Embed Size (px)
Citation preview

Journal of NeurochemistryRaven Press, Ltd., New York
1995 international Society for Neurochemistry
Vasoactive Intestinal Peptide and Pituitary AdenylateCyclase-Activating Polypeptide Potentiate cfos Expression
Induced by Glutamate in Cultured Cortical Neurons
Jean-Luc Martin, Didier Gasser, and Pierre J . Magistretti
Laboratoire de Recherche Neurologique, Institut de Physiologie et Service de Neurologie du CHUV, Faculté de Médecine,
Abstract : Previous reports have demonstrated that gluta-mate stimulates c-fos mRNA expression in primary cul-tures of mouse cerebral cortical neurons. We show herethat vasoactive intestinal peptide (VIP) induces c-fosmRNA expression ; however, this effect of VIP is com-pletely inhibited by the noncompetitive NMDA receptorantagonist MK-801, therefore indicating that VIP stimu-lates c-fos expression in a glutamate-dependent manner .A similar effect was observed with pituitary adenylate cy-clase-activating polypeptide27 (PACAP27) . At the intra-cellular level, coactivation of protein kinases A and C me-diates the glutamate-dependent stimulation of c-fos ex-pression evoked by VIP, because either H-89 orstaurosporin inhibits the effect of VIP as well as that ofglutamate. These results point to a "biochemical ANDgate" mechanism, which implies the obligatory activationof both protein kinases Aand C in the transduction of c-fos expression . In summary, this article provides evidencethat VIP and PACAP27 potentiate the effect of glutamate,the principal effector on c-fos expression, suggesting thatboth peptides can increase the "throughput" or"strength" of glutamate-containing circuits in the cere-bral cortex . Key Words: c-fos-Glutamate-Vasoactiveintestinal peptide-Pituitary adenylate cyclase-activat-ing polypeptide27-Modulation -Cerebral cortex .J. Neurochem. 65, 1-9 (1995) .
Vasoactive intestinal peptide (VIP) is a 28 aminoacid polypeptide originally isolated from porcine duo-denum by Said and Mutt (1970) . VIP immunoreactiv-ity has subsequently been demonstrated outside thegastrointestinal tract, including the PNS and CNS (Lo-ran et al ., 1979) . In the rodent CNS, VIP is highlyconcentrated in the cerebral cortex, where it is con-tained in a homogeneous population of intracorticaland radially oriented bipolar neurons (Morrison et al .,1984) . VIP is released in a Ca`+ -dependent manner byappropriate stimuli (Martin and Magistretti, 1989a,b) .Specific VIP receptors have been demonstrated byquantitative autoradiography with the highest levelsobserved in the cerebral cortex, olfactory bulb, dentate
Université de Lausanne, Lausanne, Switzerland
gyrus, and locus ceruleus (Martin et al ., 1987) . In thecerebral cortex, three subtypes of VIP receptors havebeen recently characterized and are differentially local-ized in astrocytes, microvessels, and synaptosomalmembranes (Martin et al ., 1992) . In addition to affect-ing neuronal excitability of identified neurons (Ferronet al ., 1985 ; Haas and Gähwiler, 1992), activation ofVIP receptors stimulates glycogenolysis in astrocytes(Magistretti et al ., 1983 ; Sorg and Magistretti, 1991)as well as cyclic AMP (cAMP) formation in corticalneurons, astrocytes, and intraparenchymal microves-sels (Chneiweiss et al ., 1985 ; Martin et al ., 1992) . Anelevation in cytoplasmic levels of cAMP triggers theactivation of the catalytic subunit of the protein kinaseA . Among possible protein kinase A-phosphorylatedproteins, the transcription factor cAMP-response ele-ment binding protein (CREB) is a likely target . Phos-phorylation of the transcription factor CREB stimu-lates its activity, therefore inducing transcription ofgenes with a CRE element in their promoter region(Yamamoto et al ., 1988) . Among the many geneswhose promoter bears a CRE element such as proen-kephalin and VIP, the cf.s immediate early gene(IEG) is the best characterized . Many IEGs, includingcfos, have been found to encode transcription factors
Received September 22, 1994 ; revised manuscript received De-cember 23, 1994 ; accepted December 23, 1994 .
Address correspondence and reprint requests to Dr. J .-L. Martinat Institut de Physiologie, Faculté de Médecine, Université de Lau-sanne, rue du Bugnon 7, CH 1005 Lausanne, Switzerland .Abbreviations used : AP3, 2-amino-3-phosphonopropionic acid ;
cAMP, cyclic AMP; CNQX, 6-cyano-7-nitroquinoxaline-2,3-dione ;CREB, cAMP-response element binding protein ; dBcAMP, dibu-tyryl-cAMP; DMEM, Dulbecco's modified Eagle's medium ; FCS,fetal calf serum ; GFAP, filial fibrillary acidic protein ; H-89,N-[2-(p-bromocinnamylamino)-ethyll-5-isoquinolinesulfonamide ;IBMX, isobutylmethylxanthine; IEG, immediate early gene ; LDH,lactate dehydrogenase ; MK-801, (5R, l OS)-(+)-5-methyl-10,1 I -di-hydro-5H-dibenzo[a,d]cyclohepten-5,10-imine ; PACAP, pituitaryadenylate cyclase-activating polypeptide ; PBSglc, phosphate-buf-fered saline supplemented with glucose ; SDS, sodium dodecyl sul-fate ; SSC, saline-sodium citrate ; VIP, vasoactive intestinal peptide .

2
that are believed to regulate the subsequent transcrip-tion of other genes termed late response genes (Shengand Greenberg, 1990) . To stimulate the expression oflate response genes, the Fos protein forms a heterodi-meric transcription factor with another protein of theJun family of DNA-binding proteins (Sheng andGreenberg, 1990) . Fos/Jun complexes recognize aspecific DNA sequence element known as AP-1 andthereby regulate transcription of late response genescontaining this sequence within their regulatory region(Sheng and Greenberg, 1990) . In summary, by actingas transcription factors, IEGs such as c-fbs translateshort-term stimuli into relatively long-term changes inneuronal function .
In view of the foregoing and because VIP is a potentstimulator of cAMP formation in cultured cortical neu-rons (Chneiweiss et al ., 1985), we have investigatedthe effect of VIP on the expression of the IEG cfbsin primary cultures of neurons originating from mousecerebral cortex .VIP shares a high degree of sequence identity with
pituitary adenylate cyclise-activating polypeptide(PACAP) (Miyata et al ., 1989) . PACAP, which wasoriginally isolated from ovine hypothalamus, exists intwo amidated forms, PACAP38 and PACAP27, shar-ing the same N-terminal 27 amino acids (Arimura,1992) . PACAP is involved in various functions suchas the synthesis and secretion of catecholamines fromthe adrenal medulla (Arimura, 1992), the stimulationof neuritogenesis and survival of cultured rat sym-pathetic neuroblasts (DiCicco-Bloom and Deutsch,1992), and the promotion of neurite outgrowth inPC 12 cells (Deutsch and Sun, 1992) . In the CNS, thepresence of PACAP-like immunoreactive neurons hasbeen demonstrated in the ovine posterior pituitary andthe septum (K6ves et al ., 1990), as well as in thethalamus, amygdaloid complex, hippocampus, and ce-rebral cortex of the rat (K6ves et al ., 1991) . Further-more, the presence of PACAP receptors has been ob-served in various regions of the CNS including thecerebral cortex, the striatum, the hippocampus, thethalamus, the hypothalamus, and the olfactory bulb(Masuo et al ., 1992) . In view of these observationsand to gain further insight into the role played by thisneuropeptide in the CNS, we have investigated theregulation of c-fôs expression by PACAP27 in corticalneurons.
In this article we report that VIP or PACAP27 appli-cation results indeed in c-fs induction ; however, thisaction of VIP or PACAP27 is indirect, being mediatedby glutamate acting on NMDA receptors.
MATERIALS AND METHODS
MaterialsPoly-t,-ornithine (molecular weight, 30,000-70,000),
Dulbecco's modified Eagle's medium (D-7777; DMEM),bovine pancreas insulin, human apo-transferrin, putrescine,progesterone, and L-glutamate were all obtained from Sigma;
l . Nr"urorhem., Vol. 65, No . /, 1995
J.-L. MARTIN ET AL.
VIP and PACAP27 were from Bachem (Bubendorf, Switzer-land) ; calmidazolium, staurosporin, and H-89 were fromBIOMOL (Hamburg, Germany) ; 6-cyano-7-nitroquinoxa-line-2,3-dione (CNQX) and 2-amino-3-phosphonopropionicacid (AP3) were from Tocris Neuramin (Bristol, U.K .) ;(5R, I0S)-(+)-5-methyl-l0, I I-dihydro-5H-dibenzola,dl-cyclohepten-5,10-imine (MK-801 ) was from Research Bio-chemicals Incorporated (Natick, MA, U .S.A .) .
Preparation of primary cultures of mousecerebral cortical neurons
Primary cultures of cerebral cortical neurons were pre-pared from 16-17-day-old Swiss mice embryos accordingto the method described by DiPorzio and colleagues withsubstantial modifications (DiPorzio et al ., 1980) . The cere-bral cortex was dissected and the meninges were carefullyremoved. Cells were then dissociated mechanically througha flame-narrowed Pasteur pipette by two series of 15-20 gentle flushes, first in 5 ml and then in 12 ml of phos-phate-buffered saline supplemented with 33 mM glucose(PBSglc) . Cell debris was discarded by three centrifugationsat 100 g for 4 min. Cells (800,000/ml) were plated into 100x 15-mm plates previously coated for at least 2 h with 15p,g/nnl polyornithine, rinsed once with water, and then oncewith PBSglc . To avoid contaminations by glial cells no fetalcall serum (FCS) was added either in the culture mediumor for coating the culture plates (Stella et al ., 1995) . Becausethe effect of glutamate was examined, glutamate as well asaspartate were omitted from the culture medium . The serum-free medium was composed of DMEM (lacking glutamateand aspartate) supplemented with 2 mM glutamine, 7.5 mMsodium bicarbonate, 5 mM HEPESbuffer (pH 7.4), 0.1 mg/ml streptomycin, and 0.06 mg/ml penicillin . A mixture ofhormones and salt containing 25 p.g/ml insulin, 100 Vg/niltransterrin, 60 p.M putrescine, 20 nM progesterone, and 30nM sodium selenate was added to the culture medium . Cellswere maintained for 6 days at 37°C in a humidified atmo-sphere of 95% air and 5% CO, .
Immunostaining of cortical cultures with cell-specific anti-bodies yields 94%, neuron-specific enolase-immunoreactivecells and 99% neurofilament-immunoreactive cells. No de-tectable immunostaining was observed with antibodiesagainst glial fibrillary acidic protein (GFAP), galactocere-broside, or MAC 1, which are specific markers for astrocytes,oligodendrocytes, and microglia, respectively (Stella et al .,1995) .
Exposure of cortical neurons to agonistsAfter 6 days in culture without changing the medium,
cortical neurons were exposed to agonists as follows: One-third (i .e ., 5 ml) of the medium was removed and agonistsdissolved therein . The remaining two-thirds ofthe incubationvolume was removed, placed in the incubator, and replacedby the 5 ml containing the agonist or agonists . At the endof the agonist application (10 min), the medium was re-placed by the 10 ml (which was maintained in the incubator)and the incubation continued for 50 min . This complex ma-nipulation was necessary to avoid nonspecific increases incfo .s expression due to cellular stress . Glutamate receptorantagonists and protein kinase inhibitors were added 15 minand 30 min before the agonists, respectively .
Northern blottingElectrophoresis . At the end of the stimulation, total RNA
was extracted from cultured cortical neurons using CsCl

VIP-GLU7AMA7E INTERACTION ON c-fos EXPRESSION
centrifugation procedure according to Chirgwin and collabo-rators ( 1979) . Ten micrograms of total RNA was electro-phoresed on a 1 .2% agarosc/2 M formaldehyde gel andtransferred onto Gene-Screen nylon membrane . Hybridiza-tion was performed overnight at 65°C in 50% formamide,5X saline-sodium citrate (SSC), IX PE 1250 mM Tris-HCI, pH 7.5, 0.5% sodium pyrophosphate, 5% sodium dode-cyl sulfate (SDS), 1 % polyvinylpyrrolidone, 1% , Ficoll, 25mM EDTA, 1 % bovine serum albumin 1 150 pg/ml salmonsperm DNA with a'ZP-anti-sense v- .fbs riboprobe (SP6 RNApolymerase) . Filters were washed under high stringencyconditions, first with 2X SSC/0. I % SDS at 65°C for 15 minfollowed by two washes of 15 min each in 0. I X SSC/0. I (YcSDS at 65°C . Filters were dried and apposed to Kodak ARfilm for 5-15 h with an intensifying screen .The levels of cfos mRNA were determined by scanning
densitometry (Bio Image System, Millipore, Bedford, MA,U .S.A .) and normalized for differences in gel loading relativeto the levels of actin mRNA that were determined by rehy-bridization after stripping of the filters . Hybridization andwashing conditions for actin were identical to those forc-fi)s .
Determination of glutamate concentrationsConcentrations of glutamate, present in the culture me-
dium under control or stimulated (with VIP or PACAP27)conditions, were determined by reversed-phase HPLC aftero-phthalaldehyde precolumn derivatization as described indetail in Klancnik and collaborators ( 1992) .
Determination of lactate dehydrogenase (LDH )activityMeasurement of LDH activity in the culture medium is
a simple and quantitative method for assessing glutamate-mediated toxicity in cortical cell cultures (Koh and Choi,1987) . After 6 days in culture, cortical neurons were incu-bated for 10 min in the presence or not of 100 pM glutamate;I, 3, 5, or 9 h later, LDH activity in the culture medium wasassessed according to the method of Wrobleski and LaDue( 1955) as modified by Koh and Choi (1987) .
Determination of cAMP levelsCAMP levels were measured by radioimmunoassay (Am-
ersham) . After 6 days in culture, cortical neurons plated on35-mm-diameter Petri dishes (50-100 pg of protein) werestimulated by increasing concentrations of VIP or PACAP27directly in the culture medium in which 1 mM isobutylmeth-ylxanthine (IBMX) had been previously dissolved . The reac-tion was stopped by a brief sonication ; the suspension wasthen boiled for 10 min at 95°C and centrifuged for 2 min at9,980 g. An aliquot of the supernatant was taken to assessCAMP levels by radioimmunoassay using ' -ç1-cAMP astracer (Amersham) . Protein content was measured using themethod of Bradford (1976) .
RESULTS
The effect of VIP on c-os mRNA expression wasanalyzed by northern blot in primary cultures of mousecerebral cortical neurons. In preliminary experiments,aimed at determining suitable conditions to assess c-fo .s expression, nonspecific induction of c-t- mRNAdue to medium changes was observed . To circumventthis drawback, stimulations were performed directly inthe culture medium according to the protocol described
FIG . 1 . Involvement of glutamate in the induction of c-fos mRNAexpression by VIP in cultured cortical neurons . Cultures of corti-cal neurons were stimulated for 10 min by VIP 1 pM in the pres-ence or not of various glutamate receptor antagonists. MK-8011 NM, CNQX 100 pM, or AP3 1 mM was added 15 min beforeVIP . After 1-h incubation, total RNA was isolated according tothe procedure described in Materials and Methods . In each lane,10 pg was loaded and electrophoresed . Autoradiogram showsresults from one experiment, repeated twice with similar results .
in Materials and Methods. Using this procedure, basalc-f~)s expression was barely detectable (see Figs . 1-7) . It is interesting that basal expression of c-f)s wasinhibited totally in the presence of the noncompetitiveNMDA receptor antagonist MK-801 (not shown),therefore suggesting that basal cfos expression wasdue to glutamate present in the medium . Indeed, theconcentration of glutamate in the extracellular fluidwas 1 .2 ± 0.22 pM (n = 6) after 6 days in culturewithout renewal of the medium .
Stimulation of cultured cortical neurons with VIP 1pM induced c-f)s mRNA expression by almost fivefold(Fig . I ) . VIPinduction of c-fls mRNA expression wasinhibited completely by the noncompetitive NMDAreceptor antagonist MK-801 (Fig . I) . The cx-amino-3-hydroxy-5-methyl-4-isoxazolepropionate (AMPA)/kainate receptor antagonist CNQX was only margin-ally inhibitory and the metabotropic receptor antago-nist AP3 was devoid of any effect (Fig . 1 ) . It is im-portant that MK-801 did not affect the VIP receptor-mediated signal transduction, because the stimulationof cAMP evoked by VIP was not inhibited by MK-801 JcAMP levels (pmol/mg of protein) : control,241 .7 --±- 16 ; I pM VIP, 1,669.2 ± 223; I pM VIP + lpM MK-801, 1,741 .6 - 182. n = 3 J . These observa-tions point to a mediation by glutamate, acting onNMDA receptors, in the induction by VIP of cf)sexpression . Although being mediated by glutamate, theeffect of VIP on c,fi)s expression is not due to thestimulation of glutamate release by the peptide becauseapplication of VIP or PACAP27 (see below), in condi-tions identical to those in which c-ft)s expression wasdetermined (see Materials and Methods), did not in-
.1. Ncmrochcin- Vol . 05, No . 1, 1995

FIG . 2 . Induction of c-fos mRNA expression by glutamate incultured cortical neurons . Cultures of cortical neurons were stim-ulated for 10 min by glutamate 100 p.M in the presence or notof various glutamate receptor antagonists (for concentrations,see legend to Fig . 1) added 15 min before glutamate . After 1-hincubation, total RNA was isolated according to the proceduredescribed in Materials and Methods . In each lane, 10 yg wasloaded and electrophoresed . Autoradiogram shows results fromone experiment, repeated twice with similar results .
crease glutamate concentration in the incubation me-dium . Thus, glutamate concentrations were (hM) :control, 1 .2 -±- 0.22 ; 1 pM VIP, 1 .69 ± 0 .31 ; 0.01 pMPACAP27, 1 .27 -!- 0.12 ; n = 6 .The effect of exogenously added glutamate on c-fos
expression was also examined . As previously reportedin detail (Hisanaga et al ., 1992 ; Vaccarino et al ., 1992)and as indicated in Fig . 2, glutamate induced c-f)smRNA expression . To rule out the possibility that thestimulation by glutamate of c-fos mRNA expressioncould be ascribed to a potential excitotoxic effect oncortical neurons, LDH released into the culture me-dium was determined . As indicated in Table 1, expo-sure of cortical neurons to 100 pM glutamate during10 min did not affect LDH activity released into the
TABLE 1 . Absence of glularnate toxicity in culturedcortical neurons
J. Neuroehe711., Vol. 65, No. 1, 1995
LDH release (U/ml)
Control
24.7 ± 2.2
26.7 ± 0.8
26.7 ± 0.8
29.3 ± 0 .8Glutamate,
100 PM
24.7-0.8
26.7
0.8
27.3+0.8
30.7+0.8
Sister cultures of cortical neurons were exposed or not (control)to 100 N,M glutamate for 10 min . Samples of bathing media weretaken I, 3, 5, and 9 h after the pulse stimulation and released LDHwas measured according to the method of Wrobleski and LaDue(1955) as modified by Koh and Choi (1987) . Results, which areexpressed in conventional units (U) per milliliter, are mean - SEMvalues of triplicate determinations from one experiment .
J .-L . MARTIN ET AL.
FIG . 3. Additive effect of VIP and glutamate on c-fos mRNAexpression in cultured cortical neurons . Cultures of cortical neu-rons were stimulated for 10 min by VIP 1 NM, by glutamate 100NM, or by VIP 1 pM + glutamate 100 NM. The effect of variousglutamate receptor antagonists (for concentrations, see legendto Fig . 1) added 15 min before the agonists was examined onthe stimulation of c-fos expression evoked by VIP + glutamate .After 1-h incubation, total RNA was isolated according to theprocedure described in Materials and Methods . In each lane,10 mg was loaded and electrophoresed . Autoradiogram showsresults from one experiment, repeated twice with similar results .
culture medium even 9 h after the end of the pulsestimulation . These results, which were confirmed bytrypan blue exclusion staining (not shown), are ingood agreement with those of Choi and colleagues,who have previously reported that young cultures ofcortical neurons (5-7 days in vitro) are not vulnerableto glutamate excitotoxicity (Choi et al ., 1987) . Induc-tion of c-os expression by glutamate was inhibitedcompletely by the noncompetitive NMDA receptor an-tagonist MK-801, whereas CNQX was only marginallyeffective (Fig . 2) . It is interesting that the metabotropicreceptor antagonist AP3 potentiated glutamate-evokedc-fbs expression, possibly by removing the inhibitoryeffect exerted by glutamate on cAMP formation, whichis mediated by metabotropic receptors (Fig . 2) .When added together, VIP and glutamate exhibited
additive effects on cfos mRNA levels (Fig . 3) . Thecombined effect of VIP and glutamate on c-fos mRNAexpression was also inhibited by MK-801, whereas c-fxp.s expression was not detestably altered by CNQX orAP3 (Fig . 3) . These results support an involvement ofNMDA receptors in the induction of cfos expressionevoked by VIP and glutamate added together .
As a second step toward the characterization of c-,fos induction evoked by VIP, we set out to determinethe intracellular pathway(s) likely to mediate c-jbsexpression evoked by VIP . To this aim, different pro-tein kinase inhibitors were tested including N-[2-(p-bromocinnamylamino)-ethyl]-5-isoquinolinesulfon-amide (H-89), an isoquinolinesulfonamide, which isa specific protein kinase A inhibitor (Chijiwa et al .,1990), calmidazolium, which inhibits calmodulin and
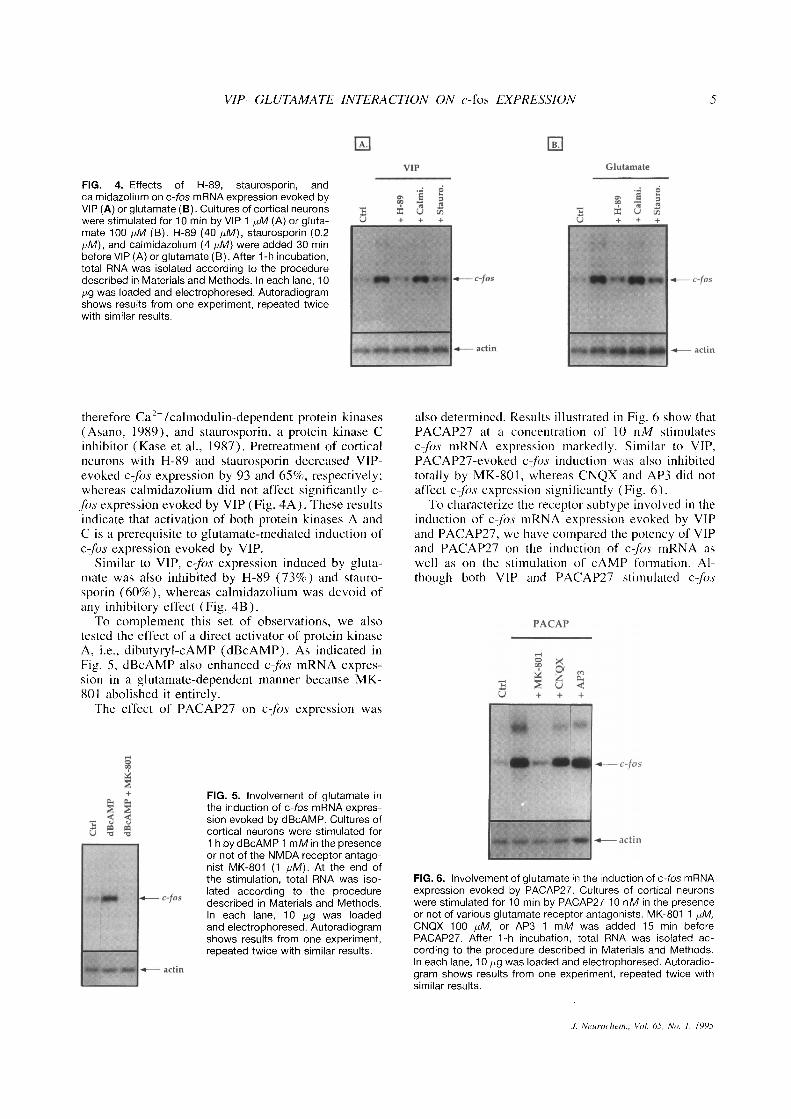
FIG . 4 . Effects of H-89, staurosporin, andcalmidazolium on c-fos mRNA expression evoked byVIP (A) or glutamate (B) . Cultures of cortical neuronswere stimulated for 10 min by VIP 1 pM (A) or gluta-mate 100 p,M (B) . H-89 (40 p,M), staurosporin (0 .2pM), and calmidazolium (4 pM) were added 30 minbefore VIP (A) or glutamate (B) . After 1-h incubation,total RNA was isolated according to the proceduredescribed in Materials and Methods . In each lane, 10pg was loaded and electrophoresed . Autoradiogramshows results from one experiment, repeated twicewith similar results .
VIP-GLUTAMATE INTERACTION ON c-fos EXPRESSION
therefore Caz+/calmodulin-dependent protein kinases(Asano, 1989), and staurosporin, a protein kinase Cinhibitor (Kase et al ., 1987) . Pretreatment of corticalneurons with H-89 and staurosporin decreased VIP-evoked c-fos expression by 93 and 65%, respectively ;whereas calmidazolium did not affect significantly c-fos expression evoked by VIP (Fig. 4A) . These resultsindicate that activation of both protein kinases A andC is a prerequisite to glutamate-mediated induction ofc-fôs expression evoked by VIP .
Similar to VIP, cfôs expression induced by gluta-mate was also inhibited by H-89 (73%) and stauro-sporin (60%), whereas calmidazolium was devoid ofany inhibitory effect (Fig . 4B) .To complement this set of observations, we also
tested the effect of a direct activator of protein kinaseA, i .e ., dibutyryl-CAMP (dBcAMP) . As indicated inFig . _5, dBcAMP also enhanced c-fos mRNA expres-sion in a glutamate-dependent manner because MK-801 abolished it entirely .The effect of PACAP27 on cfos expression was
FIG . 5 . Involvement of glutamate inthe induction of c-fos mRNA expres-sion evoked by dBcAMP . Cultures ofcortical neurons were stimulated for1 h by dBcAMP 1 mM in the presenceor not of the NMDA receptor antago-nist MK-801 (1 pM) . At the end ofthe stimulation, total RNA was iso-lated according to the proceduredescribed in Materials and Methods .In each lane, 10 pg was loadedand electrophoresed . Autoradiogramshows results from one experiment,repeated twice with similar results .
also determined . Results illustrated in Fig . 6 show thatPACAP27 at a concentration of 10 nM stimulatesc -fibs mRNA expression markedly . Similar to VIP,PACAP27-evoked c-f)s induction was also inhibitedtotally by MK-801, whereas CNQX and AP3 did notaffect cfos expression significantly (Fig . 6) .To characterize the receptor subtype involved in the
induction of c-fos mRNA expression evoked by VIPand PACAP27, we have compared the potency of VIPand PACAP27 on the induction of cfôs mRNA aswell as on the stimulation of cAMP formation . Al-though both VIP and PACAP27 stimulated cfôs
FIG. 6 . Involvement of glutamate in the induction of c-fos mRNAexpression evoked by PACAP27 . Cultures of cortical neuronswere stimulated for 10 min by PACAP27 10 nM in the presenceor not of various glutamate receptor antagonists . MK-801 1 pM,CNQX 100 pM, or AP3 1 mM was added 15 min beforePACAP27 . After 1-h incubation, total RNA was isolated ac-cording to the procedure described in Materials and Methods .In each lane, 10 pg was loaded and electrophoresed . Autoradio-gram shows results from one experiment, repeated twice withsimilar results .
J. Neuroc'kem ., Vol. 6 .5, No . 1, 1995
6
FIG . 7 . Induction of c-fos mRNA expression by increasing con-centrations ofVIP or PACAP27 . Cultures of cortical neurons werestimulated for 10 min by increasing concentrations of VIP orPACAP27 . After 1-h incubation, total RNA was isolated ac-cording to the procedure described in Materials and Methods .In each lane, 5 Ng was loaded and electrophoresed .
expression in a concentration-dependent manner,PACAP27 was more potent (- 100 times) and efficientthan VIP to stimulate c- o .s expression (Fig . 7) . Asimilar rank order of potency and efficacy was alsoobserved on the stimulation of adenylate cyclise, withEC, () values of 0.7 and 200 nM for PACAP27 andVIP, respectively (Fig . 8) .
DISCUSSION
In the cerebral cortex, c-f~)s expression is enhancedby various manipulations such as metrazole-inducedseizures (Morgan et al ., 1987), spreading depression(Herrera et al ., 1993), and focal ischernia (Uemura etal ., 1991) . In addition to these stimuli and similar toother neurotransmitters such as the monoamines dopa-mine (Vaccarino et al ., 1993) and noradrenaline (Binget al ., 1991 ), as well as the excitatory amino acidglutamate (Hisanaga et al ., 1992), VIP also stimulatesc-fos mRNA expression in the cerebral cortex (Hisa-naga et al ., 1992 ; Vaccarino et al ., 1993) . However,the present article shows clearly that VIP does notstimulate cfb.s expression directly but rather by inter-acting with glutamate present in the incubation me-dium and acting on NMDA receptors (Fig . 1) . In otherwords, VIP potentiates the effect of otherwise margin-ally effective concentrations of glutamate (-I pM),which acts as the real effector . These results agreewith those of Hisanaga and collaborators (1992), whopreviously reported that c-f)s expression stimulated byVIP was inhibited by the NMDA receptor antagonistsMK-801, CPP (4-(3-phosphonopropyl)piperazin-2-carboxylic acid), and APH (2-amino-7-phophonohep-tanoate) . However, in the report of Hisanaga and col-laborators, the blockade of VIP-evoked c-f~)s expres-sion by MK-801 was partial, due probably to the ratherlow concentration of MK-801 used, i .e ., 0 .1 pM insteadof I pM in our experiments . On the other hand, these
1. Neuro( 'lic" nn . . Vo/. 65, No. / . 1995
J.-L. MARTIN ET AL.
results contrast with those of Vaccarino and collabora-tors, who did not observe any blockade of VIP-evokedc-f~)s expression with the glutamate receptor antago-nists MK-801 and CNQX (Vaccarino et al ., 1993) .This discrepancy may be attributed to their paradigmof stimulation, involving the replacement of the me-dium (in which cultures were bathed for several days)by a Locke's solution, therefore removing accumulatedglutamate (Vaccarino et al ., 1993) . In addition, 7-10% of the cells present in their cultures were GFAP-positive astrocytes due probably to the presence ofFCS in their culture medium (Vaccarino et al ., 1993),whereas no GFAP-positive astrocytes were observedin our cultures ( Stel la et al ., 1995 ) . Nevertheless, ourresults and those of Vaccarino and collaborators( 1993) demonstrate that VIP can induce c-fs mRNAexpression either by potentiating the effect of endoge-nous glutamate or by acting directly . In this context,the complete blockade of VIP-evoked c-fOs mRNAexpression observed with MK-801 rules out the exis-tence in our study of a direct induction of c-fC)s expres-sion by VIP .
Inhibition of VIP-evoked cfbs expression by H-89,staurosporin, but not calmidazolium provides clues tounderstanding the mechanism or mechanisms by whichthe combined activation of' VIP and glutamate recep-tors is transduced to the nucleus to stimulate c-fpsexpression . The inhibitory effect of H-89 on the induc-tion of cfos expression evoked by VIP (Fig . 4A) cor-relates with the stimulatory effect of VIP on cAMPformation observed in this preparation (Fig . 8) . Inhibi-
FIG . 8 . Stimulation of CAMP formation evoked by increasingconcentrations of VIP or PACAP27 . Cultures of cortical neuronswere stimulated by increasing concentrations of PACAP27 (0)or VIP (0) in the presence of 1 mM IBMX. The reaction wasstopped by a brief sonication ; the suspension was then boiledfor 10 min at 95°C and centrifuged for 2 min at 9,980 g . Analiquot of the supernatant was taken to assess CAMP levelsby radioimmunoassay using ' 751-cAMP as tracer (Amersham) .Protein content was measured using the method of Bradford(1976) .
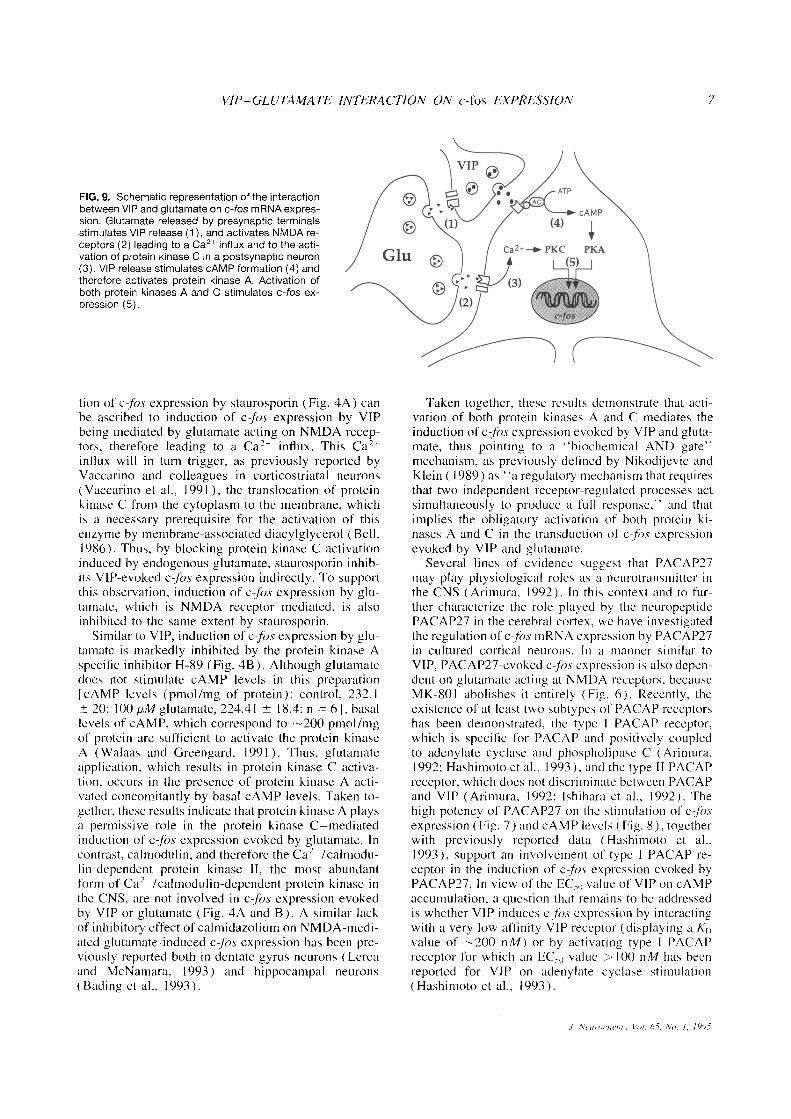
FIG. 9 . Schematic representation of the interactionbetween VIP and glutamate on c-fos mRNA expres-sion . Glutamate released by presynaptic terminalsstimulates VIP release (1), and activates NMDA re-ceptors (2) leading to a Ca21 influx and to the acti-vation of protein kinase C in a postsynaptic neuron(3) . VIP release stimulates cAMP formation (4) andtherefore activates protein kinase A . Activation ofboth protein kinases A and C stimulates c-fos ex-pression (5) .
VIP-GLUTAMATE INTERACTION ON c-fos EXPRESSION
tion of cfos expression by staurosporin (Fig . 4A) canbe ascribed to induction of c-fbs expression by VIPbeing mediated by glutamate acting on NMDA recep-tors, therefore leading to a Ca" influx . This Caz+influx will in turn trigger, as previously reported byVaccarino and colleagues in corticostriatal neurons(Vaccarino et al ., 1991), the translocation of proteinkinase C from the cytoplasm to the membrane, whichis a necessary prerequisite for the activation of thisenzyme by membrane-associated diacylglycerol (Bell,1986) . Thus, by blocking protein kinase C activationinduced by endogenous glutamate, staurosporin inhib-its VIP-evoked c-fos expression indirectly . To supportthis observation, induction of c-fos expression by glu-tamate, which is NMDA receptor mediated, is alsoinhibited to the same extent by staurosporin .
Similar to VIP, induction of c-ft)s expression by glu-tamate is markedly inhibited by the protein kinase Aspecific inhibitor H-89 (Fig . 4B) . Although glutamatedoes not stimulate cAMP levels in this preparation[cAMP levels (pmol/mg of protein) : control, 232.1± 20 ; 100 pM glutamate, 224.41 ± 18 .4 ; n = 6], basallevels of cAMP, which correspond to -200 pmol/mgof protein are sufficient to activate the protein kinaseA (Walaas and Greengard, 1991) . Thus, glutamateapplication, which results in protein kinase C activa-tion, occurs in the presence of protein kinase A acti-vated concomitantly by basal CAMP levels . Taken to-gether, these results indicate that protein kinase A playsa permissive role in the protein kinase C-mediatedinduction of c-ft)s expression evoked by glutamate . Incontrast, calmodulin, and therefore the Ca 2 '/calmodu-lin-dependent protein kinase II, the most abundantform of Ca'+/calmodulin-dependent protein kinase inthe CNS, are not involved in cfns expression evokedby VIP or glutamate (Fig . 4A and B) . A similar lackof inhibitory effect of calmidazolium on NMDA-medi-ated glutamate-induced c-fs expression has been pre-viously reported both in dentate gyrus neurons (Lereaand McNamara, 1993) and hippocampal neurons(Biding et al ., 1993) .
Taken together, these results demonstrate that acti-vation of both protein kinases A and C mediates theinduction of c-fbs expression evoked by VIP and gluta-mate, thus pointing to a "biochemical AND gate"mechanism, as previously defined by Nikodijevic andKlein ( 1989) as "a regulatory mechanism that requiresthat two independent receptor-regulated processes actsimultaneously to produce a full response," and thatimplies the obligatory activation of both protein ki-nases A and C in the transduction of c-ps expressionevoked by VIP and glutamate .
Several lines of evidence suggest that PACAP27may play physiological roles as a neurotransmitter inthe CNS (Arimura, 1992) . In this context and to fur-ther characterize the role played by the neuropeptidePACAP27 in the cerebral cortex, we have investigatedthe regulation of c-f)s mRNA expression by PACAP27in cultured cortical neurons . In a manner similar toVIP, PACAP27-evoked c-ft)s expression is also depen-dent on glutamate acting at NMDA receptors, becauseMK-801 abolishes it entirely (Fig . 6) . Recently, theexistence of at least two subtypes of PACAP receptorshas been demonstrated, the type I PACAP receptor,which is specific for PACAP and positively coupledto adenylate cyclase and phospholipase C (Arimura,1992 ; Hashimoto et al ., 1993), and the type II PACAPreceptor, which does not discriminate between PACAPand VIP (Arimura, 1992; Ishihara et al ., 1992) . Thehigh potency of PACAP27 on the stimulation of c-ft)sexpression (Fig . 7) and cAMP levels (Fig . 8), togetherwith previously reported data (Hashimoto et al .,1993), support an involvement of type I PACAP re-ceptor in the induction of c-f)s expression evoked byPACAP27 . In view of the EC,,, ) value of VIP on cAMPaccumulation, a question that remains to be addressedis whether VIP induces c-f)s expression by interactingwith a very low affinity VIP receptor (displaying a K�value of' -200 nM) or by activating type I PACAPreceptor for which an EC�, value > 100 nM has beenreported for VIP on adenylate cyclase stimulation(Hashimoto et al ., 1993) .
J . Ncurnrhcm ., Vo/. 0 5, No. 1, 1995

In summary, results reported in this article provideevidence that VIP and PACAP27 can potentiate theeffect of otherwise marginally effective concentrationsof glutamate (see Figs . I, 3, 4, 6, and 7) . This effectis postsynaptic in nature because VIP and PACAP27do not stimulate glutamate release. Conversely, previ-ous reports have demonstrated that VIP release in thecerebral cortex is enhanced by glutamate (Wang et al .,1986) . Taken together, these results suggest that VIPand glutamate can interact synergistically to increasethe throughput or strength of glutamate-containing cir-cuits in the cerebral cortex (Fig . 9) . Furthermore, be-cause VIP and PACAP27 potentiate the effect of gluta-mate on c-fos expression and because cfos can affectthe expression of late genes, these results suggest thatVIP and PACAP27 could modulate indirectly synapticplasticity in the cerebral cortex .
Acknowledgment : We are grateful to Dr . Kim Do andMs . Beatrix Benz of the Brain Research Institute in Zurich(Switzerland) for HPLC determinations of glutamate . Weare also grateful to Nephi Stella and Luc Pellerin for expertadvice on neuronal culture preparation and to MauricetteMaillard for cyclic AMP measurements . This work was sup-ported by Fonds National de la Recherche Scientifique grantsNo . 31-39'468 .93 to J .-L.M . and No . 31-26'427 .89 to P .J.M .
REFERENCESArimura A . (1992) Pituitary adenylate cyclase activating polypep-
tide (PACAP) : discovery and current status of research . Regal .Pept. 37, 287-303 .
Asano M . (1989) Divergent pharmacological effects of three cal-modulin antagonists, N-(6-aminohexyl)-5-chloro-l-naphtha-lenesulfonamide (W-7), chlorpromazine and calmidazolium, onisometric tension development and myosin light chain phos-phorylation in intact bovine tracheal smooth muscle. J. Pharma-col . Exp. Ther. 251, 764-773 .
Bading H ., Ginty D . D ., and Greenberg M . E . (1993) Regulationof gene expression in hippocampal neurons by distinct calciumsignaling pathways . Science 260, 181-186 .
Bell R . M . ( 1986) Protein kinase C activation by diacylglycerolsecond messengers . Cell 45, 631-632 .
Bing G ., Filer D ., Miller J . C ., and Stone E . A . (1991 ) Noradrenergicactivation of immediate early genes in rat cerebral cortex . Mal.Brain Res. 11, 43-46 .
Bradford M . M . (1976) A rapid and sensitive method for the quanti-tation of microgram quantities of protein using the principle ofprotein-dye binding . Anal. Biochem. 72, 248-254 .
Chijiwa T ., Mishima A ., Hagiwara M., Sano M ., Hayashi K ., InoueT ., Naito K ., Toshioka T ., and Hidaka H . (1990) Inhibition offorskolin-induced neurite outgrowth and protein phosphorylation by a newly synthesized selective inhibitor of cyclic AMP-dependent protein kinase, N-[2-(p-bromocinnamylamino)-ethyl 1 -5-i soquinolinesulfonamide (H-89), of PC 12D pheochro-mocytoma cells . J . Biol. Chem . 265, 5267-5272 .
Chirgwin J . M ., Przybyla A . E., MacDonald R . J ., and Rutter W . J .( 1979) Isolation of biologically active ribonucleic acid fromsources enriched in ribonuclease . Biochemistry 18, 5294-5299 .
Chneiweiss H ., Glowinski J., and Premont J . (1985) Vasoactiveintestinal polypeptide receptors linked to an adenylate cyclase,and their relationship with biogenic amine- and somatostatin-sensitive adenylate cyclases on central neuronal and glial cellsin primary cultures . J . Neurochem. 44, 779-786 .
Choi D . W ., Maulucci-Gelde M., and Kriegstein A . R . (1987) Gluta-mate neurotoxicity in cortical cell culture . J. Neurosci. 7, 357-368 .
1. Neurochem. . Vol. 65, No. 1 . 1995
J.-L. MARTIN ET AL.
Deutsch P . J . and Sun Y . ( 1992) The 38-amino acid form of pituitaryadenylate cyclase-activating polypeptide stimulates dual signal-ing cascades in PC12 cells and promotes neurite outgrowth . J.Biol. Chem . 267, 5108-5113 .
DiCicco-Bloom E . and Deutsch P . J . (1992) Pituitary adenylatecyclase activating polypeptide (PACAP) potently stimulatesmitosis, neuritogenesis and survival in cultured rat sympatheticneuroblasts . Regul. Pept. 37, 319 .
DiPorzio U ., Daguet M-C ., Glowinski J ., and Prochiantz A . (1980)Effect of striatal cells on in vitro maturation of mesencephalicdopaminergic neurones grown in serum-free conditions . Nature288, 370-373 .
Ferron A ., Siggins G . R ., and Bloom F . E . (1985) Vasoactive intesti-nal polypeptide acts synergistically with norepinephrine to de-press spontaneous discharge rate in cerebral cortical neurons .Proc. Nad. Acad. Sci. USA 82, 8810-8812 .
Haas H . L . and Gähwiler B . H . (1992) Vasoactive intestinal poly-peptide modulates neuronal excitability in hippocampal slicesof the rat . Neuroscience 47, 273-277 .
Hashimoto H ., Ishihara T ., Shigemoto R ., Mori K ., and Nagata S .( 1993) Molecular cloning and tissue distribution of a receptorfor pituitary adenylate cyclase-activating polypeptide . Neuron11, 333-342 .
Herrera D . G ., Maysinger D ., Gadient R., Boeckh C ., Otten U ., andCuello A . C . (1993) Spreading depression induces c-ft) .s-likeimmunoreactivity and NGF mRNA in the rat cerebral cortex .Brain Res. 602, 99- 103 .
Hisanaga K ., Sagar S . M ., and Sharp F . R . (1992) N-Methyl-D-aspartate antagonists block Fos-like protein expression inducedvia multiple signaling pathways in cultured cortical neurons . .l.Neurochem. 58, 1836-1844 .
Ishihara T ., Shigemoto R ., Mori K ., Takahashi K ., and Nagata S .( 1992) Functional expression and tissue distribution of a novelreceptor for vasoactive intestinal polypeptide . Neuron 8, 811-819 .
Kase H ., Iwahashi K ., Nakanishi S ., Matsuda Y ., Yamada K ., Taka-hashi M., Murakata C ., Sato A ., and Kaneko M . (1987) K-252compounds, novel and potent inhibitors of protein kinase C andcyclic nucleotide-dependent protein kinases . Biochem. Biophys .Res. Commun. 142, 436-440 .
Klancnik J . M ., Cu6nod M., Gähwiler B . H ., Jiang Z . P ., and DoK . Q . (1992) Release of endogenous amino acids, includinghomocysteic acid and cysteine sulphinic acid, from rat hippo-campal slices evoked by electrical stimulation of Schaffer col-lateral -commissural fibres . Neuroscience 49, 557-570 .
Koh J . Y . and Choi D . W . ( 1987) Quantitative determination ofglutamate mediated cortical neuronal injury in cell culture bylactate dehydrogenase efflux assay . J. Neurosci. Methods 20,83-90 .
K6ves K ., Arimura A ., Somogyvari-Vigh A ., Vigh S ., and Miller J .( 1990) Immunohistochernical demonstration of a novel hypo-thalamic peptide, pituitary adenylate cyclase activating polypep-tide, in the ovine hypothalamus . Endocrinology 127, 264-271 .
K6ves K ., Arimura A ., G6res T. G ., and Vigh S . ( 1991) Comparativedistribution of immunoreactive pituitary adenylate cyclase-acti-vating polypeptide and vasoactive intestinal polypeptide in ratforebrain . Neuroendocrinology 54, 159-169 .
Lerea L. S . and McNamara J . O . ( 1993) lonotropic glutamate recep-tor subtypes activate c-fos transcription by distinct calcium-requiring intracellular signaling pathways . Neuron 10, 31-41 .
Lor6n I ., Emson P . C ., Fahrenkrug J ., Bj6rklund A ., Alumets J .,Hakanson R ., and Sundler F . (1979) Distribution of vasoactiveintestinal polypeptide in the rat and mouse brain . Neuroscience4, 1953-1976 .
Magistretti P . J ., Manthorpe M ., Bloom F. E ., and Varon S . (1983 )Functional receptors for vasoactive intestinal polypeptide in cul-tured astroglia from neonatal rat brain . Regul . Pept. 6, 71-80.
Martin J . L . and Magistretti P . J . (1989x) Release of vasoactiveintestinal peptide in mouse cerebral cortex : evidence for a roleof arachidonic acid metabolites . J . Neurosci. 9, 2536-2542 .
Martin J . L . and Magistretti P. J . ( 1989b) Pharmacological evidencefor a role of voltage-sensitive Ca - '-channels of the T type in

VIP-GLUTAMATE INTERACTION ON c-fos EXPRESSION
the release of vasoactive intestinal peptide evoked by K' inmouse cerebral cortical slices . Neuroscience 30, 423-431 .
Martin .I . L ., Diet] M . M ., Hof P. R ., Palacios J . M ., and MagistrettiP. J . (1987) Autoradiographic mapping of [mono[' -5 11iodo-Tyr' °,Met0 "]-vasoactive intestinal peptide binding sites in therat brain . Neuroscience 23, 539-565 .
Martin J .-L., Feinstein D . L ., Yu N ., Sorg O ., Rossier C ., and Magis-tretti P . J . (1992) VIP receptor subtypes in mouse cerebralcortex : evidence for a differential localization in astrocytes,microvessels and synaptosomal membranes . Brain Res . 587, 1-12 .
Masuo Y ., Ohtaki T ., Masuda Y ., Tsuda M., and Fujino M . (1992)Binding sites for pituitary adenylate cyclase activating polypep-tide (PACAP) : comparison with vasoactive intestinal polypeptide (VIP) binding site localization in rat brain sections . BrainRes . 575, 113-123 .
Miyata A ., Arimura A ., Dahl R . R ., Minamino N ., Uehara A ., JiangL., Culler M . D ., and Coy D . H . (1989) Isolation of'a novel 38residue-hypothalamic polypeptide which stimulates adenylatecyclase in pituitary cells . Biochem. Biophys. Res . Commun . 164,567--574.
Morgan J . I ., Cohen D . R ., Hempstead J . L ., and Curran T . (1987)Mapping patterns of cfbs expression in the central nervoussystem after seizure . Science 237, 192-197 .
Morrison J . H ., Magistretti P . J ., Benoit R ., and Bloom F . E . ( 1984)The distribution and morphological characteristics of the intra-cortical VTP-positive cell : an immunohistochemical analysis.Brain Res . 292, 269-282 .
Nikodijevic O . and Klein D . C. ( 1989) Adenosine stimulates adeno-sine 3',5'-monophosphate and guanosine 3',5'-monophosphateaccumulation in rat pinealocytes: evidence for a role for adenosine in pineal neurotransmission . Endocrinology 125, 2150-2157 .
Said S . 1 . and Mutt V . (1970) Polypeptide with broad biologicalactivity : isolation from small intestine. Science 169, 1217-1218 .
Sheng M . and Greenberg M. E . (1990) The regulation and function
of c-fbs and other immediate early genes in the nervous system .Neuron 4, 477-485 .
Sorg O. and Magistretti P. J . ( 1991 ) Characterization of the glyco-genolysis elicited by vasoactive intestinal peptide, noradrenalineand adenosine in primary cultures of mouse cerebral corticalastrocytes . Brain Res. 563, 227-233 .
Stella N ., Pellerin L ., and Magistretti P . J . (1995) Modulation ofthe glutamate-evoked release of arachidonic acid from mousecortical neurons : involvement of a pH-sensitive membranephospholipase A Z . J. Neurosci. (in press) .
Uemura Y ., Kowall N . W ., and Moskowitz M . A . ( 1991) Focalischemia in rats causes time-dependent expression of c-fos pro-tein immunoreactivity in widespread regions of ipsilateral cor-tex . Brain Res. 552, 99- 105 .
Vaccarino F. M ., Liljequist S ., and Tallman J . F . ( 1991 ) Modulationof protein kinase C translocation by excitatory and inhibitoryamino acids in primary cultures of neurons . J. Neurochern. 57,391-396 .
Vaccarino F . M ., Hayward M . D ., Nestler E. J ., Duman R . S ., andTallman J . F . ( 1992) Differential induction of immediate earlygenes by excitatory amino acid receptor types in primary cultures of cortical and striatal neurons . Mot . Brain Res. 12, 233-241 .
Vaccarino F. M ., Hayward M . D ., Le H . N ., Hartigan D . J ., DumanR . S ., and Nestler F: . J . ( 1993 ) Induction of immediate earlygenes by cyclic AMP in primary cultures of neurons from ratcerebral cortex . Mot. Brain Re.s . 19, 76-82 .
Walaas S . 1 . and Greengard P . ( 1991 ) Protein phosphorylation andneuronal function . Pharmacol. Res . 43, 299-349 .
Wang J.-Y ., Yaksh T. L., Harty G . J ., and Go V . L. W . (1986)Neurotransmitter modulation of VIP release from cat cerebralcortex . Am. J . Phvsiol . 250, R104-RI 11 .
Wrobleski F. and LaDue J . S . ( 1955) Lactic dehydrogenase activityin blood . Proc . Soc . E'.vp. Biol. Med. 90, 210-213 .
Yamamoto K . K ., Gonzalez G . A ., Biggs W . H . III ., and MontminyM . R . (1988) Phosphorylation-induced binding and transcrip-tional efficacy of nuclear factor CREB . Nature 334, 494-498 .
1. Neurorhem ., Vo/ . 65, No . 1, 1995
![Terlipressin versus other vasoactive drugs for hepatorenal ...[Intervention Review] Terlipressin versus other vasoactive drugs for hepatorenal syndrome Mads Israelsen 1, Aleksander](https://img.pdfslide.us/doc/110x75/60a7d279a5295c2888291ae4/terlipressin-versus-other-vasoactive-drugs-for-hepatorenal-intervention-review.jpg)




























