Embed Size (px)
Citation preview

J Neurosurg Pediatrics 2:150–157, 2008
150 J. Neurosurg.: Pediatrics / Volume 2 / August 2008
YELOMENINGOCELE is a clinically significant prima-ry neurulation defect leading to significant mor-bidity and mortality rates.7,21 The incidence of
MMC has changed during the past decades according togeographic region and ethnicity.12,35 The precise worldwideincidence of MMC has been difficult to assess given thevariations in prenatal diagnosis and in policies regardingthe elective termination of pregnancy.8,21 In more techno-logically advanced countries, despite folic acid supplemen-tation and routine prenatal screening, administered as early
as the first trimester, MMC still affects nearly 1 in 2000 livebirths.1 Newborns with MMC present with various degreesof lower-extremity paralysis, sexual and sphincter dysfunc-tion, tethering of the spinal cord, skeletal deformation,brainstem dysfunction, hydrocephalus, and ventriculoperi-toneal shunt complications.19,25,26 Even with appropriateneonatal treatment and multidisciplinary medical care,MMC is still associated with a mortality rate that can rangebetween 14 and 35% in the first 5 years of age.36
Vascular and apoptotic changes in the placode ofmyelomeningocele mice during the final stages of in uterodevelopment
Laboratory investigation
JOAQUIM L. REIS, M.D., PH.D.,1,2 JORGE CORREIA-PINTO, M.D., PH.D.,3,4
MARIANA P. MONTEIRO, M.D., PH.D.,1 MADALENA COSTA, B.SC.,1
AND GROVER M. HUTCHINS, M.D.5
1Department of Anatomy, Abel Salazar Institute for the Biomedical Sciences and Unit forMultidisciplinary Biomedical Research, University of Porto; 2Department of Neurosurgery, SantoAntónio General Hospital; 4Department of Pediatric Surgery, São João Hospital, Porto; 3Life andHealth Sciences Research Institute, School of Health Sciences, University of Minho, Braga, Portugal;and 5Department of Pathology, Johns Hopkins School of Medicine, Baltimore, Maryland
Object. Myelomeningocele (MMC) is a primary neurulation defect that is associated with devastating neurologicaldisabilities in affected newborns. To better characterize the in utero neurodegenerative process of MMC, the authorsinvestigated the changes in vascular organization, apoptosis, and the presence of inflammatory cells during gestationby using a mutant mouse model of MMC.
Methods. The curly tail/loop tail (ct/lp) mutant mouse model of MMC was chosen to obtain fetuses at differentstages of gestation. Mouse fetuses harboring MMC were harvested by caesarean section at embryonic Days 14.5, 16.5,and 18.5 (complete mouse gestation at 19 days, 6 mice/group); littermate fetuses with the same gestational age butwithout an MMC were used as controls. Samples of the MMC placode or normal spinal cord were stained for immuno-cytochemical labeling with caveolin antibody (endothelium marker) and activated caspase-3 antibody (apoptosis mark-er). Samples were morphometrically analyzed with a computer-assisted image analyzer.
Results. The MMC mice presented with an increase in vascular density from embryonic Days 16.5–18.5 and anenhanced number of apoptotic cells at embryonic Day 18.5, compared with controls. There were scarce signals of aninflammatory reaction in the MMC placode, as a few infiltrating neutrophils were seen only at embryonic Day 18.5.
Conclusions. Fetal placodes in MMC mice showed evidence of increased vascular density since embryonic Day16.5 and increased apoptosis at embryonic Day 18.5. These new data support the view that in utero changes of theMMC placode, occurring during the last stages of gestation, contribute to the neuropathological manifestations in full-term newborns with MMC. (DOI: 10.3171/PED/2008/2/8/150)
KEY WORDS • apoptosis • endothelium • inflammation • immunocytochemistry •myelomeningocele • spina bifida
M
Abbreviation used in this paper: MMC = myelomeningocele.This article contains some figures that are displayed in
color online but in black and white in the print edition.

The exact origin and timing of MMC lesion formationare not fully understood. Authors of several pathophysio-logical studies have challenged the concept that an intrinsicembryonic error is the sole determinant of an MMC lesionand instead have favored the hypothesis that the intrauter-ine environment is a major cause of injury to the placode,through either trauma or chemical toxicity.5,9,13,15 Based onthe concept that neurological deficits caused by the MMClesion occur during the second half of gestation, in uterosurgical correction of MMC in humans was initiated a dec-ade ago.4 The clinical outcomes of in utero surgery havesuggested clear improvement in some neurological lesions,namely in the reversal of ectopic cerebellar tonsils of theMMC-associated Chiari malformation, decreasing it from95 to 38%, with a consequent reduction in the need for aventricular shunt from 91 to 59%.3,31–34 In contrast, deficitsin the lower limbs and in urodynamic function in patientssubmitted to MMC correction, through either fetal or clas-sic postnatal surgery, were similar to those of controls.14,30,27
Attending to these data, the potential benefits of in utero re-pair of MMC are under scrutiny in a randomized trial de-signed to compare the clinical outcomes of patients withMMC treated with either fetal or postnatal surgery.2
Recently, using a mouse model of MMC, we document-ed astrocytic proliferation during gestation together withneuronal degeneration, after embryonic Day 16.5 (full-termgestation: 19 days), predominantly in the dorsal part of theplacode.23 These findings are in accordance with clinicaland electromyography findings in newborn humans withMMC who display a loss of leg movements during the first2 postnatal weeks, suggesting that neuronal damage wasinitiated before birth.27 To better characterize the cytologicaldynamics in the MMC placode that could lead to neuronaldegeneration during gestation, we looked for other cytolog-ical alterations not clearly evaluated in previous scientificstudies.10,11,15,17,23 In this way we evaluated the changes invascular structure, frequency of apoptotic cells, and inflam-mation in the placode of MMC mutant mouse fetuses dur-ing the final stages of gestation.
Methods
Mouse Strains and Breeding
The curly tail (ct) mouse strain was initially supplied tous by the Jackson Laboratory, and the loop-tail (lp) lineagewas kindly given to us by Dr. Andrew Copp. These ct andlp mouse lineages were housed as separated colonies in acontrolled-temperature room maintained on a daily 12-hourlight/dark period. Water and food were freely availablewith no restriction. To obtain newborns with MMC, doublyheterozygous males (lp/+, ct/+) were mated overnight withhomozygous curly tail females (+/+, ct/ct). Mating waspromoted during the dark phase, and the female mice werechecked for vaginal plugs in the next daylight period. Theday of copulation plugging was defined as embryonic Day0.5. Mice harboring both curly tail and loop-tail mutations(ct/ct, lp/+) are known to express severe MMC almost in-variabily.29
Sample Collection and Experimental Protocol
Our goal was to study fetuses of ct/lp mice with MMC atdifferent gestational ages. Fetuses were harvested by cae-
sarean section at embryonic Days 14.5, 16.5, and 18.5 ofgestation (end of gestation: 19 days). Fetuses without MMCwith a straight tail and apparently normal hindlimb functionwere used as controls for each of the 3 gestational agegroups. Each of the 6 groups of fetuses (3 MMC groups and3 control groups) comprised 6 mouse fetuses. All experi-mental protocols, including animal care, were reviewed andapproved by the Animal Care and Use Committee of ourinstitution (Abel Salazar Institute for Biomedical Sciences).The directives set forth by the National Institutes of HealthGuide for the Care and Use of Laboratory Animals as wellas the guidelines from the European Union on animal pro-tection (Directive 86/609/EC) were also followed.
The pregnant mice were killed with a lethal dose of ket-amine and xylazine hydrochloride, and fetuses were har-vested through a caesarean section procedure. Newbornpups were photographed, evaluated for muscular tonus,killed, and fixed in a buffered solution of 4% paraformalde-hyde with 14% picric acid for 1 hour. After fixation, thefetuses were cut at the MMC level and further fixed for 24hours in the same fixative solution. Tissue samples werethen routinely processed and embedded in paraffin. Sec-tions were mounted on poly-L-lysine-coated glass slides,dewaxed in xylene, rehydrated with grading ethanols, andstained with H & E or immunocytochemical methods.
For immunocytochemistry labeling, antigen retrieval wasperformed by boiling the mounted section in a pressurecooker containing a 10 mM citrate buffer (pH 6.0) for 3minutes. Samples were incubated at room temperature in3% hydrogen peroxide in methanol to quench endogenousperoxidase, followed by normal serum of swine (DakoCy-tomation) for 20 minutes to block nonspecific staining. Forlabeling of apoptotic cells, samples were incubated with thecleaved caspase-3 antibody (Asp175, Cell Signaling Tech-nology) in a 1:100 dilution for 1 hour. After rinsing withtris-buffered saline, incubation with a second antibody wasperformed using swine anti–rabbit antibody (DakoCytoma-tion) for 30 minutes. Tissue sections were again washed intris-buffered saline and incubated in avidin-biotin-peroxi-dase complex (DakoCytomation) at room temperature for30 minutes. After several washes, the reaction product wasmade visible with diaminobenzidine tetrahydrochloride(DakoCytomation) treatment. Tissues sections were coun-terstained with hematoxylin.
To label endothelial cells, caveolin staining was used in amanner similar to the aforementioned method for cleavedcaspase-3 staining. Briefly, nonspecific staining wasblocked by incubation with a postprimary block (Novocas-tra Laboratories) for 5 minutes at room temperature, fol-lowed by primary antibody incubation with caveolin anti-body (caveolin, BD Biosciences) at a 1:400 dilution for 90minutes before incubation with a secondary antibody bind-ing system (Novocastra Laboratories). Before detection ofthe reaction product with diaminobenzidine tetrahydrochlo-ride, the samples were incubated with polymer from theNovoLink Polymer Detection System (Novocastra Labora-tories) for 30 minutes at room temperature. Backgroundstaining of tissue sections was again done with hematoxy-lin.
Negative controls for the specificity of immunolabelingwere performed by omitting the primary antibody. Verifica-tion of the staining of anterior spinal artery and other ab-dominal vessels of fetal samples and mouse thymus sam-
J. Neurosurg.: Pediatrics / Volume 2 / August 2008
Vascular density and apoptosis in myelomeningocele
151
J. L. Reis et al.
ples were used as internal positive controls for caveolin andcleaved caspase-3, respectively. For each antibody, stainingof the different sample sections was performed on the sameday for all tissue sections to minimize interassay variability.
Inflammatory reactions were evaluated by quantifyingthe number of neutrophil cells in the MMC or normal spi-nal cord on hematoxylin-stained sections.
Morphometric Analysis
Each spinal cord section was photographed using lightmicroscopy, and morphometric analysis was performedwith a computer-assisted imaging analyzer (Leica Qwin,Leica Microsystems). Under the same magnification, im-munolabeling was quantified in each sample of spinal cordtissue with a square area of 13,100 µm2. The number ofimmunoreactive cells for cleaved caspase-3 antibody wascounted in each sample, and the proportion of these cells perµm2 was calculated. The number of stained capillaries ineach sample was counted, and its density per µm2 was cal-culated. In each spinal cord section, high-magnificationphotographs of 6 different areas were obtained: 3 from theventral area that included the white and gray columns andfrom the area around the central channel. The procedure forthe dorsal area was similar. For the MMC groups, areas ofplacode selected for morphometric analysis were 3 ventraland 3 dorsal regions, according to the following criteria: thefirst area was located on the basal plate (medial), the secondon the alar plate (lateral), and a third intermediate areaaround the sulcus limitans.
Statistical Analysis
A comparison of the means between MMC and controlsfor each gestational age was performed using a two-tailed,unpaired t-test. A probability value < 0.05 was consideredto reflect a statistically significant difference.
Results
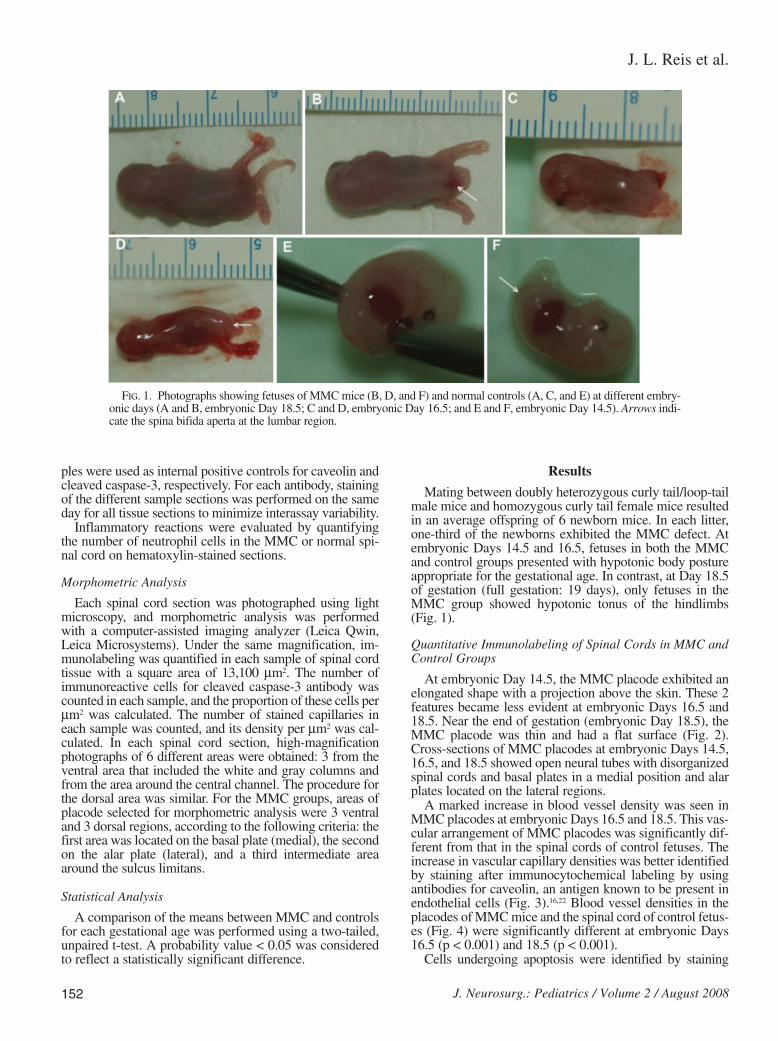
Mating between doubly heterozygous curly tail/loop-tailmale mice and homozygous curly tail female mice resultedin an average offspring of 6 newborn mice. In each litter,one-third of the newborns exhibited the MMC defect. Atembryonic Days 14.5 and 16.5, fetuses in both the MMCand control groups presented with hypotonic body postureappropriate for the gestational age. In contrast, at Day 18.5of gestation (full gestation: 19 days), only fetuses in theMMC group showed hypotonic tonus of the hindlimbs(Fig. 1).
Quantitative Immunolabeling of Spinal Cords in MMC andControl Groups
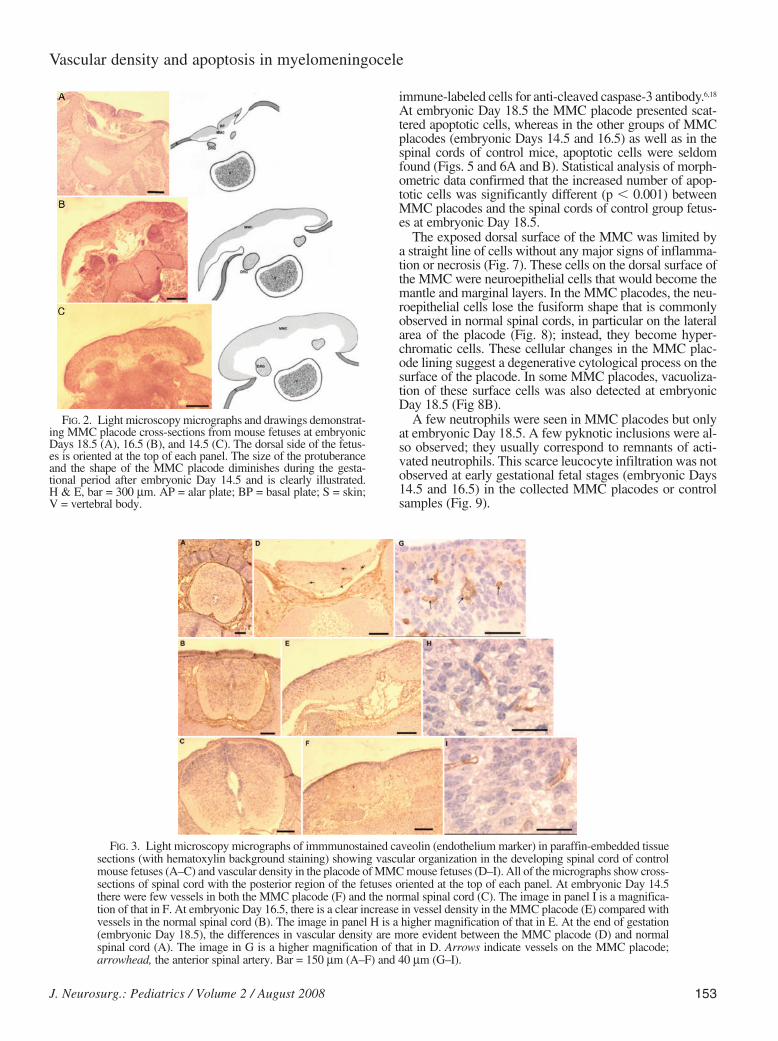
At embryonic Day 14.5, the MMC placode exhibited anelongated shape with a projection above the skin. These 2features became less evident at embryonic Days 16.5 and18.5. Near the end of gestation (embryonic Day 18.5), theMMC placode was thin and had a flat surface (Fig. 2).Cross-sections of MMC placodes at embryonic Days 14.5,16.5, and 18.5 showed open neural tubes with disorganizedspinal cords and basal plates in a medial position and alarplates located on the lateral regions.
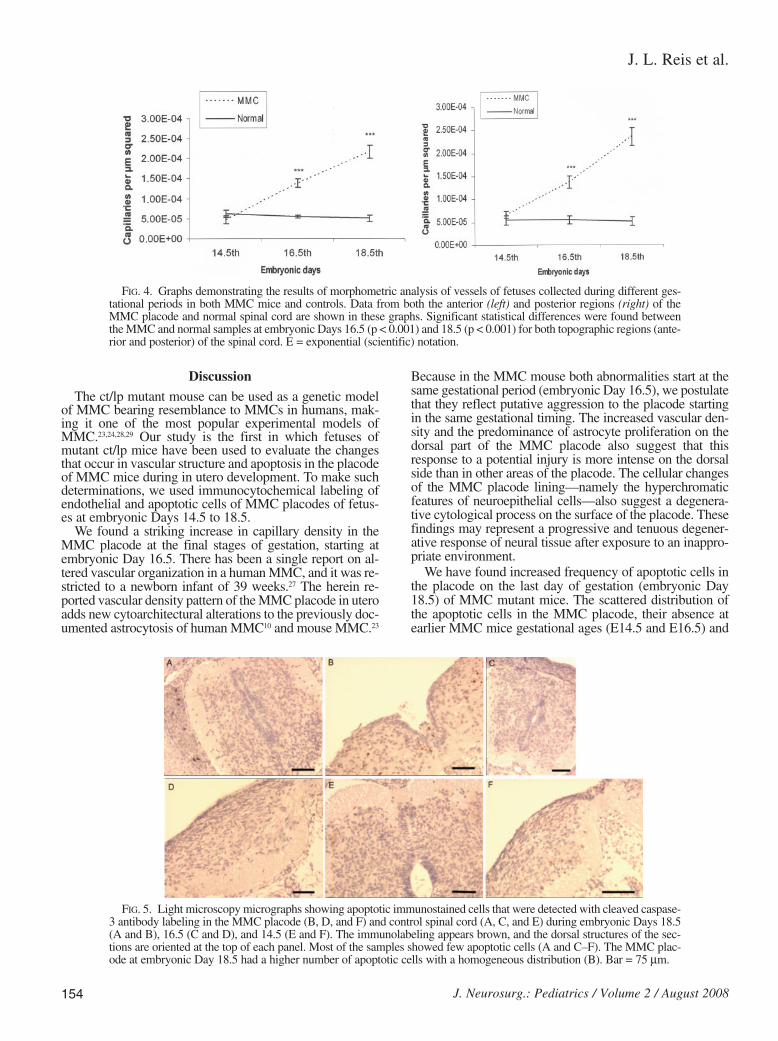
A marked increase in blood vessel density was seen inMMC placodes at embryonic Days 16.5 and 18.5. This vas-cular arrangement of MMC placodes was significantly dif-ferent from that in the spinal cords of control fetuses. Theincrease in vascular capillary densities was better identifiedby staining after immunocytochemical labeling by usingantibodies for caveolin, an antigen known to be present inendothelial cells (Fig. 3).16,22 Blood vessel densities in theplacodes of MMC mice and the spinal cord of control fetus-es (Fig. 4) were significantly different at embryonic Days16.5 (p < 0.001) and 18.5 (p < 0.001).
Cells undergoing apoptosis were identified by staining
152 J. Neurosurg.: Pediatrics / Volume 2 / August 2008
FIG. 1. Photographs showing fetuses of MMC mice (B, D, and F) and normal controls (A, C, and E) at different embry-onic days (A and B, embryonic Day 18.5; C and D, embryonic Day 16.5; and E and F, embryonic Day 14.5). Arrows indi-cate the spina bifida aperta at the lumbar region.
immune-labeled cells for anti-cleaved caspase-3 antibody.6,18
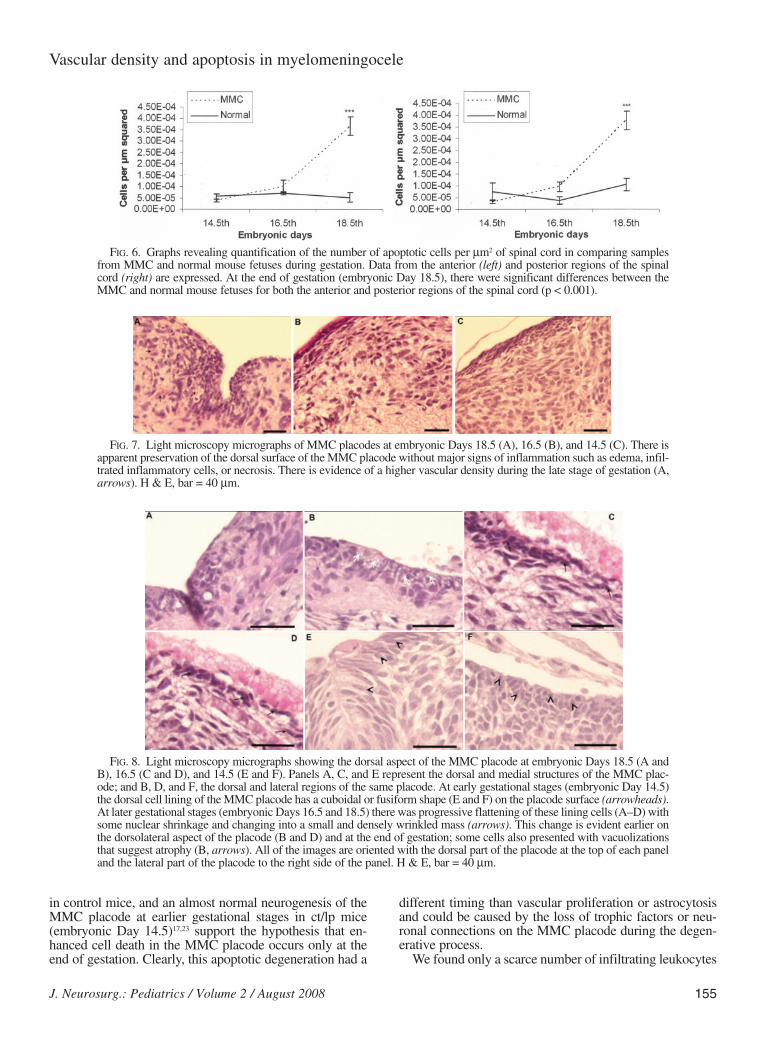
At embryonic Day 18.5 the MMC placode presented scat-tered apoptotic cells, whereas in the other groups of MMCplacodes (embryonic Days 14.5 and 16.5) as well as in thespinal cords of control mice, apoptotic cells were seldomfound (Figs. 5 and 6A and B). Statistical analysis of morph-ometric data confirmed that the increased number of apop-totic cells was significantly different (p , 0.001) betweenMMC placodes and the spinal cords of control group fetus-es at embryonic Day 18.5.
The exposed dorsal surface of the MMC was limited bya straight line of cells without any major signs of inflamma-tion or necrosis (Fig. 7). These cells on the dorsal surface ofthe MMC were neuroepithelial cells that would become themantle and marginal layers. In the MMC placodes, the neu-roepithelial cells lose the fusiform shape that is commonlyobserved in normal spinal cords, in particular on the lateralarea of the placode (Fig. 8); instead, they become hyper-chromatic cells. These cellular changes in the MMC plac-ode lining suggest a degenerative cytological process on thesurface of the placode. In some MMC placodes, vacuoliza-tion of these surface cells was also detected at embryonicDay 18.5 (Fig 8B).
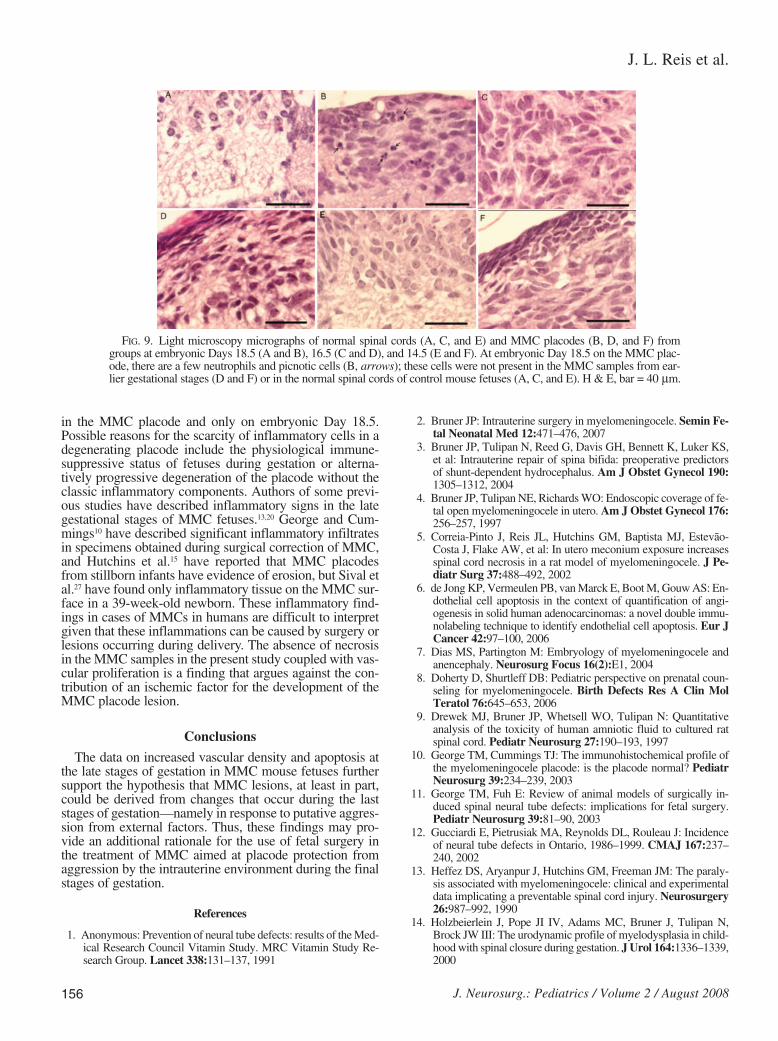
A few neutrophils were seen in MMC placodes but onlyat embryonic Day 18.5. A few pyknotic inclusions were al-so observed; they usually correspond to remnants of acti-vated neutrophils. This scarce leucocyte infiltration was notobserved at early gestational fetal stages (embryonic Days14.5 and 16.5) in the collected MMC placodes or controlsamples (Fig. 9).
FIG. 2. Light microscopy micrographs and drawings demonstrat-ing MMC placode cross-sections from mouse fetuses at embryonicDays 18.5 (A), 16.5 (B), and 14.5 (C). The dorsal side of the fetus-es is oriented at the top of each panel. The size of the protuberanceand the shape of the MMC placode diminishes during the gesta-tional period after embryonic Day 14.5 and is clearly illustrated. H & E, bar = 300 µm. AP = alar plate; BP = basal plate; S = skin;V = vertebral body.
FIG. 3. Light microscopy micrographs of immmunostained caveolin (endothelium marker) in paraffin-embedded tissuesections (with hematoxylin background staining) showing vascular organization in the developing spinal cord of controlmouse fetuses (A–C) and vascular density in the placode of MMC mouse fetuses (D–I). All of the micrographs show cross-sections of spinal cord with the posterior region of the fetuses oriented at the top of each panel. At embryonic Day 14.5there were few vessels in both the MMC placode (F) and the normal spinal cord (C). The image in panel I is a magnifica-tion of that in F. At embryonic Day 16.5, there is a clear increase in vessel density in the MMC placode (E) compared withvessels in the normal spinal cord (B). The image in panel H is a higher magnification of that in E. At the end of gestation(embryonic Day 18.5), the differences in vascular density are more evident between the MMC placode (D) and normalspinal cord (A). The image in G is a higher magnification of that in D. Arrows indicate vessels on the MMC placode;arrowhead, the anterior spinal artery. Bar = 150 µm (A–F) and 40 µm (G–I).
J. Neurosurg.: Pediatrics / Volume 2 / August 2008 153
Vascular density and apoptosis in myelomeningocele
J. L. Reis et al.
DiscussionThe ct/lp mutant mouse can be used as a genetic model
of MMC bearing resemblance to MMCs in humans, mak-ing it one of the most popular experimental models ofMMC.23,24,28,29 Our study is the first in which fetuses ofmutant ct/lp mice have been used to evaluate the changesthat occur in vascular structure and apoptosis in the placodeof MMC mice during in utero development. To make suchdeterminations, we used immunocytochemical labeling ofendothelial and apoptotic cells of MMC placodes of fetus-es at embryonic Days 14.5 to 18.5.
We found a striking increase in capillary density in theMMC placode at the final stages of gestation, starting atembryonic Day 16.5. There has been a single report on al-tered vascular organization in a human MMC, and it was re-stricted to a newborn infant of 39 weeks.27 The herein re-ported vascular density pattern of the MMC placode in uteroadds new cytoarchitectural alterations to the previously doc-umented astrocytosis of human MMC10 and mouse MMC.23
Because in the MMC mouse both abnormalities start at thesame gestational period (embryonic Day 16.5), we postulatethat they reflect putative aggression to the placode startingin the same gestational timing. The increased vascular den-sity and the predominance of astrocyte proliferation on thedorsal part of the MMC placode also suggest that thisresponse to a potential injury is more intense on the dorsalside than in other areas of the placode. The cellular changesof the MMC placode lining—namely the hyperchromaticfeatures of neuroepithelial cells—also suggest a degenera-tive cytological process on the surface of the placode. Thesefindings may represent a progressive and tenuous degener-ative response of neural tissue after exposure to an inappro-priate environment.
We have found increased frequency of apoptotic cells inthe placode on the last day of gestation (embryonic Day18.5) of MMC mutant mice. The scattered distribution ofthe apoptotic cells in the MMC placode, their absence atearlier MMC mice gestational ages (E14.5 and E16.5) and
154 J. Neurosurg.: Pediatrics / Volume 2 / August 2008
FIG. 4. Graphs demonstrating the results of morphometric analysis of vessels of fetuses collected during different ges-tational periods in both MMC mice and controls. Data from both the anterior (left) and posterior regions (right) of theMMC placode and normal spinal cord are shown in these graphs. Significant statistical differences were found betweenthe MMC and normal samples at embryonic Days 16.5 (p < 0.001) and 18.5 (p < 0.001) for both topographic regions (ante-rior and posterior) of the spinal cord. E = exponential (scientific) notation.
FIG. 5. Light microscopy micrographs showing apoptotic immunostained cells that were detected with cleaved caspase-3 antibody labeling in the MMC placode (B, D, and F) and control spinal cord (A, C, and E) during embryonic Days 18.5(A and B), 16.5 (C and D), and 14.5 (E and F). The immunolabeling appears brown, and the dorsal structures of the sec-tions are oriented at the top of each panel. Most of the samples showed few apoptotic cells (A and C–F). The MMC plac-ode at embryonic Day 18.5 had a higher number of apoptotic cells with a homogeneous distribution (B). Bar = 75 µm.
in control mice, and an almost normal neurogenesis of theMMC placode at earlier gestational stages in ct/lp mice(embryonic Day 14.5)17,23 support the hypothesis that en-hanced cell death in the MMC placode occurs only at theend of gestation. Clearly, this apoptotic degeneration had a
different timing than vascular proliferation or astrocytosisand could be caused by the loss of trophic factors or neu-ronal connections on the MMC placode during the degen-erative process.
We found only a scarce number of infiltrating leukocytes
FIG. 6. Graphs revealing quantification of the number of apoptotic cells per µm2 of spinal cord in comparing samplesfrom MMC and normal mouse fetuses during gestation. Data from the anterior (left) and posterior regions of the spinalcord (right) are expressed. At the end of gestation (embryonic Day 18.5), there were significant differences between theMMC and normal mouse fetuses for both the anterior and posterior regions of the spinal cord (p < 0.001).
FIG. 7. Light microscopy micrographs of MMC placodes at embryonic Days 18.5 (A), 16.5 (B), and 14.5 (C). There isapparent preservation of the dorsal surface of the MMC placode without major signs of inflammation such as edema, infil-trated inflammatory cells, or necrosis. There is evidence of a higher vascular density during the late stage of gestation (A,arrows). H & E, bar = 40 µm.
FIG. 8. Light microscopy micrographs showing the dorsal aspect of the MMC placode at embryonic Days 18.5 (A andB), 16.5 (C and D), and 14.5 (E and F). Panels A, C, and E represent the dorsal and medial structures of the MMC plac-ode; and B, D, and F, the dorsal and lateral regions of the same placode. At early gestational stages (embryonic Day 14.5)the dorsal cell lining of the MMC placode has a cuboidal or fusiform shape (E and F) on the placode surface (arrowheads).At later gestational stages (embryonic Days 16.5 and 18.5) there was progressive flattening of these lining cells (A–D) withsome nuclear shrinkage and changing into a small and densely wrinkled mass (arrows). This change is evident earlier onthe dorsolateral aspect of the placode (B and D) and at the end of gestation; some cells also presented with vacuolizationsthat suggest atrophy (B, arrows). All of the images are oriented with the dorsal part of the placode at the top of each paneland the lateral part of the placode to the right side of the panel. H & E, bar = 40 µm.
J. Neurosurg.: Pediatrics / Volume 2 / August 2008 155
Vascular density and apoptosis in myelomeningocele
J. L. Reis et al.
in the MMC placode and only on embryonic Day 18.5.Possible reasons for the scarcity of inflammatory cells in adegenerating placode include the physiological immune-suppressive status of fetuses during gestation or alterna-tively progressive degeneration of the placode without theclassic inflammatory components. Authors of some previ-ous studies have described inflammatory signs in the lategestational stages of MMC fetuses.13,20 George and Cum-mings10 have described significant inflammatory infiltratesin specimens obtained during surgical correction of MMC,and Hutchins et al.15 have reported that MMC placodesfrom stillborn infants have evidence of erosion, but Sival etal.27 have found only inflammatory tissue on the MMC sur-face in a 39-week-old newborn. These inflammatory find-ings in cases of MMCs in humans are difficult to interpretgiven that these inflammations can be caused by surgery orlesions occurring during delivery. The absence of necrosisin the MMC samples in the present study coupled with vas-cular proliferation is a finding that argues against the con-tribution of an ischemic factor for the development of theMMC placode lesion.
Conclusions
The data on increased vascular density and apoptosis atthe late stages of gestation in MMC mouse fetuses furthersupport the hypothesis that MMC lesions, at least in part,could be derived from changes that occur during the laststages of gestation—namely in response to putative aggres-sion from external factors. Thus, these findings may pro-vide an additional rationale for the use of fetal surgery inthe treatment of MMC aimed at placode protection fromaggression by the intrauterine environment during the finalstages of gestation.
References
1. Anonymous: Prevention of neural tube defects: results of the Med-ical Research Council Vitamin Study. MRC Vitamin Study Re-search Group. Lancet 338:131–137, 1991
2. Bruner JP: Intrauterine surgery in myelomeningocele. Semin Fe-tal Neonatal Med 12:471–476, 2007
3. Bruner JP, Tulipan N, Reed G, Davis GH, Bennett K, Luker KS,et al: Intrauterine repair of spina bifida: preoperative predictors of shunt-dependent hydrocephalus. Am J Obstet Gynecol 190:1305–1312, 2004
4. Bruner JP, Tulipan NE, Richards WO: Endoscopic coverage of fe-tal open myelomeningocele in utero. Am J Obstet Gynecol 176:256–257, 1997
5. Correia-Pinto J, Reis JL, Hutchins GM, Baptista MJ, Estevão-Costa J, Flake AW, et al: In utero meconium exposure increasesspinal cord necrosis in a rat model of myelomeningocele. J Pe-diatr Surg 37:488–492, 2002
6. de Jong KP, Vermeulen PB, van Marck E, Boot M, Gouw AS: En-dothelial cell apoptosis in the context of quantification of angi-ogenesis in solid human adenocarcinomas: a novel double immu-nolabeling technique to identify endothelial cell apoptosis. Eur JCancer 42:97–100, 2006
7. Dias MS, Partington M: Embryology of myelomeningocele andanencephaly. Neurosurg Focus 16(2):E1, 2004
8. Doherty D, Shurtleff DB: Pediatric perspective on prenatal coun-seling for myelomeningocele. Birth Defects Res A Clin MolTeratol 76:645–653, 2006
9. Drewek MJ, Bruner JP, Whetsell WO, Tulipan N: Quantitativeanalysis of the toxicity of human amniotic fluid to cultured ratspinal cord. Pediatr Neurosurg 27:190–193, 1997
10. George TM, Cummings TJ: The immunohistochemical profile ofthe myelomeningocele placode: is the placode normal? PediatrNeurosurg 39:234–239, 2003
11. George TM, Fuh E: Review of animal models of surgically in-duced spinal neural tube defects: implications for fetal surgery.Pediatr Neurosurg 39:81–90, 2003
12. Gucciardi E, Pietrusiak MA, Reynolds DL, Rouleau J: Incidenceof neural tube defects in Ontario, 1986–1999. CMAJ 167:237–240, 2002
13. Heffez DS, Aryanpur J, Hutchins GM, Freeman JM: The paraly-sis associated with myelomeningocele: clinical and experimentaldata implicating a preventable spinal cord injury. Neurosurgery26:987–992, 1990
14. Holzbeierlein J, Pope JI IV, Adams MC, Bruner J, Tulipan N,Brock JW III: The urodynamic profile of myelodysplasia in child-hood with spinal closure during gestation. J Urol 164:1336–1339,2000
156 J. Neurosurg.: Pediatrics / Volume 2 / August 2008
FIG. 9. Light microscopy micrographs of normal spinal cords (A, C, and E) and MMC placodes (B, D, and F) fromgroups at embryonic Days 18.5 (A and B), 16.5 (C and D), and 14.5 (E and F). At embryonic Day 18.5 on the MMC plac-ode, there are a few neutrophils and picnotic cells (B, arrows); these cells were not present in the MMC samples from ear-lier gestational stages (D and F) or in the normal spinal cords of control mouse fetuses (A, C, and E). H & E, bar = 40 µm.

15. Hutchins GM, Meuli M, Meuli-Simmen C, Jordan MA, HeffezDS, Blakemore KJ: Acquired spinal cord injury in human fetuseswith myelomeningocele. Pediatr Pathol Lab Med 16:701–712,1996
16. Kasper M: Phenotypic characterization of pulmonary arteries innormal and diseased lung. Chest 128 (6 Suppl):547S–552S, 2005
17. Keller-Peck CR, Mullen RJ: Patterns of neuronal differentiationin neural tube mutant mice: curly tail and Pax3 splotch-delayed.J Comp Neurol 368:516–526, 1996
18. Lossi L, Tamagno I, Merighi A: Molecular morphology of neu-ronal apoptosis: analysis of caspase 3 activation during postnataldevelopment of mouse cerebellar cortex. J Mol Histol 35:621–629, 2004
19. McDonnell GV, McCann JP: Why do adults with spina bifida andhydrocephalus die? A clinic-based study. Eur J Pediatr Surg 10(1 Suppl):31–32, 2000
20. Meuli M, Meuli-Simmen C, Hutchins GM, Yingling CD, Hoff-man KM, Harrison MR, et al: In utero surgery rescues neurolog-ical function at birth in sheep with spina bifida. Nat Med 1:342–347, 1995
21. Mitchell LE, Adzick NS, Melchionne J, Pasquariello PS, SuttonLN, Whitehead AS: Spina bifida. Lancet 364:1885–1895, 2004
22. Nag S, Venugopalan R, Stewart DJ: Increased caveolin-1 expres-sion precedes decreased expression of occludin and claudin-5during blood-brain barrier breakdown. Acta Neuropathol 114:459–469, 2007
23. Reis JL, Correia-Pinto J, Monteiro MP, Hutchins GM: In uterotopographic analysis of astrocytes and neuronal cells in the spinalcord of mutant mice with myelomeningocele. J Neurosurg 106(6 Suppl):472–479, 2007
24. Selcuki M, Manning S, Bernfield M: The curly tail mouse modelof human neural tube defects demonstrates normal spinal corddifferentiation at the level of the meningomyelocele: implicationsfor fetal surgery. Childs Nerv Syst 17:19–23, 2001
25. Shaffer J, Friedrich WN, Shurtleff DB, Wolf L: Cognitive andachievement status of children with myelomeningocele. J Pedi-atr Psychol 10:325–336, 1985
26. Singhal B, Mathew KM: Factors affecting mortality and morbid-ity in adult spina bifida. Eur J Pediatr Surg 9 (1 Suppl):31–32,1999
27. Sival DA, van Weerden TW, Vles JS, Timmer A, den Dunnen WF,Staal-Schreinemachers AL, et al: Neonatal loss of motor functionin human spina bifida aperta. Pediatrics 114:427–434, 2004
28. Stiefel D, Copp AJ, Meuli M: Fetal spina bifida in a mousemodel: loss of neural function in utero. J Neurosurg 106 (3 Sup-pl):213–221, 2007
29. Stiefel D, Shibata T, Meuli M, Duffy PG, Copp AJ: Tethering ofthe spinal cord in mouse fetuses and neonates with spina bifida.J Neurosurg 99 (2 Suppl):206–213, 2003
30. Sutton LN: Fetal surgery for neural tube defects. Best Pract ResClin Obstet Gynaecol 22:175–188, 2007
31. Sutton LN, Adzick NS, Bilaniuk LT, Johnson MP, CrombleholmeTM, Flake AW: Improvement in hindbrain herniation demon-strated by serial fetal magnetic resonance imaging following fetalsurgery for myelomeningocele. JAMA 282:1826–1831, 1999
32. Tulipan N, Hernanz-Schulman M, Bruner JP: Reduced hindbrainherniation after intrauterine myelomeningocele repair: a report offour cases. Pediatr Neurosurg 29:274–278, 1998
33. Tulipan N, Hernanz-Schulman M, Lowe LH, Bruner JP: Intraut-erine myelomeningocele repair reverses preexisting hindbrainherniation. Pediatr Neurosurg 31:137–142, 1999
34. Tulipan N, Sutton LN, Bruner JP, Cohen BM, Johnson M, AdzickNS: The effect of intrauterine myelomeningocele repair on theincidence of shunt-dependent hydrocephalus. Pediatr Neurosurg38:27–33, 2003
35. Velie EM, Shaw GM: Impact of prenatal diagnosis and elective ter-mination on prevalence and risk estimates of neural tube defects inCalifornia, 1989–1991. Am J Epidemiol 144:473–479, 1996
36. Worley G, Schuster JM, Oakes WJ: Survival at 5 years of a cohortof newborn infants with myelomeningocele. Dev Med ChildNeurol 38:816–822, 1996
Manuscript submitted January 17, 2008.Accepted April 17, 2008.Sources of support: The present research work was financially
supported by the Portuguese Foundation for Science and Technol-ogy and the Unit for Multidisciplinary Investigation in Biomedicineat the Abel Salazar Institute for Biomedical Sciences/University ofPorto.
Address correspondence to: Joaquim L. Reis, M.D., Ph.D., De-partment of Anatomy, ICBAS, Abel Salazar Institute for BiomedicalSciences, University of Porto, Largo Prof. Abel Salazar, 2, 4099-033Porto, Portugal. email: [email protected].
J. Neurosurg.: Pediatrics / Volume 2 / August 2008 157
Vascular density and apoptosis in myelomeningocele







![35. Which is the best position for operating the mouse? a) Tail away from the user c) Tail facing the right e) None of the above [IBPS Clerk 2011] b) Tail towards the user36. Which](https://img.pdfslide.us/doc/110x75/5e87c604e7966e5cc44070a9/-35-which-is-the-best-position-for-operating-the-mouse-a-tail-away-from-the.jpg)



















