Embed Size (px)
Citation preview

Vagal Preganglionic Projections to theEnteric Nervous System Characterized
With Phaseolus Vulgaris-Leucoagglutinin
MARY-CLARE HOLST, JOSHUA B. KELLY, AND TERRY L. POWLEY*
Department of Psychological Sciences, Purdue University, West Lafayette, Indiana 47907
ABSTRACTThe patterns and extent of vagal preganglionic divergence and convergence within the
gastrointestinal tract of the rat were characterized with the anterograde tracer Phaseolusvulgaris-leucoagglutinin (PHA-L). Three weeks after tracer was iontophoretically injectedinto two to four sites within the dorsal motor nucleus of the vagus, wholemounts of perfusedgut organs (stomach, duodenum, cecum) were prepared, counterstained with Cuprolinic blue,and processed for PHA-L using the avidin biotin complex with diaminobenzidine. Controlsincluded animals injected with PHA-L after intracranial deafferentations. Well-positionedinjections labeled an extremely dense and intricate network of varicose efferent axonsthroughout the gastric myenteric plexus (including that of the fundus). Individual fiberscollateralized extensively, forming a variety of pericellular arborizations and terminalcomplexes made up of both en passant and end swellings. Single axons frequently innervatedsubsets of neurons within ganglia. Most enteric neurons were contacted by varicosities ofmore than one vagal fiber. The patterns of vagal preganglionic fibers in the duodenal and cecalmyenteric plexuses resembled the organization in the stomach in many aspects, but theprojections in each organ had distinctive characteristics, and label was less dense in theintestines than in the stomach. Vagal preganglionic fibers directly innervated submucosalganglia, although sparsely. Brainstem injections of PHA-L retrogradely labeled a fewmyenteric neurons in the corpus, fundus, and duodenum: These ‘‘gastrobulbar’’ and ‘‘duode-nobulbar’’ neurons received reciprocal vagal preganglionic innervation. Finally, the PHA-Lthat spread to the nucleus of the solitary tract occasionally produced transganglionic labelingof afferent intramuscular arrays (gastric fundus). The results of this paper provide strongevidence that the traditional ‘‘command neuron’’ or ‘‘mother cell’’ hypotheses of vagal-entericorganization should be abandoned for an integrative neural network model. J. Comp. Neurol.381:81–100, 1997. r 1997 Wiley-Liss, Inc.
Indexing terms: autonomic; visceral efferents; myenteric ganglia; gastrointestinal innervation
The connections between the central nervous system(CNS) and the enteric nervous system (ENS) have beenextensively studied, beginning with the earliest descrip-tions of the autonomic nervous system (see, e.g., Meissner,1857;Auerbach, 1862; Gaskell, 1886; Ramon y Cajal, 1893;Langley and Anderson, 1894, 1895, Langley, 1899; Dogiel,1895, 1899) and continuing throughout this century (forreview, see Davenport, 1989). The vagus nerve, the sourceof parasympathetic projections to the upper gastrointesti-nal (GI) tract, has been the focus of much of this work, andit is well established that vagal preganglionic neurons inthe brainstem are organized viscerotopically (Fox andPowley, 1985; Altschuler et al., 1992), project primarily tothe myenteric plexus (Schofield, 1962; Kirchgessner andGershon, 1989; Berthoud et al., 1990), and control numer-ous motor and secretomotor responses (Laughton and
Powley, 1987; Davenport, 1989; Zhang et al., 1992). How-ever, little is known about the terminal architecture ofpreganglionics within the myenteric plexus. Neither thepattern nor the extent of divergence of individual pregan-glionic fibers has been systematically described. Similarly,potential convergence of preganglionic fibers has not beenexamined. Furthermore, the regional patterns of such
Contract grant sponsor: NIH; Contract grant number: DK27627; Con-tract grant sponsor: NIMH; Contract grant number: MH01023.*Correspondence to: Dr. Terry L. Powley, Department of Psychological
Sciences, 165 Peirce Hall, Purdue University, West Lafayette, IN 47907.E-mail: [email protected] 6 February 1996; Revised 31 October 1996; Accepted 5 Decem-
ber 1996
THE JOURNAL OF COMPARATIVE NEUROLOGY 381:81–100 (1997)
r 1997 WILEY-LISS, INC.

divergence and convergence in different parts of the GItract have not been compared.The prevailing view concerning vagal projections to the
GI tract was established by Langley (1922). To reconcilethe disparity between the few thousand preganglionicaxons of the vagus and the millions of enteric neurons,Langley hypothesized that vagal preganglionic neuronsproject to a few specialized postganglionic ‘‘vagal neurons’’or ‘‘mother cells,’’which, in turn, project divergently withinthe plexuses to coordinate autonomic control of the gut.This model assumed low pre- to postganglionic ratiossimilar to those presumed for parasympathetic pathwaysin general (cf. Wang et al., 1995). More recently, in asimilar argument, Wood (1987) proposed that vagal para-sympathetic preganglionics project to select ‘‘commandneurons’’ located within the enteric nervous system.Although the mother cell/command neuron hypothesis
has wide acceptance, it is worth stressing that the support-ing evidence for the model is largely indirect and that it isbased primarily on the discrepancy between the number ofpreganglionic neurons (or their axons) and the number ofenteric neurons. Recent attempts to determine the extentand nature of vagal preganglionic terminal arbors haveyielded conflicting results. In a survey using brainstemPhaseolus vulgaris-leucoagglutinin (PHA-L) injections totrace vagal preganglionic terminals in the gut, Kirch-gessner and Gershon (1989) observed sparse label consis-tent with the mother cell/command neuron concept. Incontrast, an experiment employing the anterograde tracerDiI to label vagal preganglionics (Berthoud et al., 1990)labeled dense networks of preganglionic endings withinganglia, with varicosities contacting large numbers ofmyenteric neurons, a pattern consistent with more exten-sive preganglionic divergence in the myenteric plexus (seealso similar patterns in Powley et al., 1994, Fig. 1;Berthoud, 1995). An electrophysiological analysis of myen-teric neurons during vagal stimulation (Schemann andGrundy, 1992) also suggested widespread parasympa-thetic projections. Neither these tracer experiments norelectrophysiological studies, however, directly examinedthe pattern or amount of divergence of individual vagalpreganglionic neurons or attempted to compare systemati-cally the patterns from different regions of the GI tract.The present experiment was designed to examine the
terminal patterns of these vagal preganglionic fibers.Iontophoretic injections of the tracer PHA-L into thedorsal motor nucleus of the vagus (DMNX) were used tolabel and, thus, to identify vagal efferent axons in theenteric plexuses, and wholemount techniques were used toassess the full compass of individual preganglionic arborsin the GI tract. The results indicate that vagal pregangli-onic fibers projecting to the gut have extensive, highlydivergent arbors with endings throughout the myentericplexus of the stomach, small intestine (duodenum), andlarge intestine (cecum). The pattern is at variance with themother cell/command neuron model of autonomic controlof the GI tract. Some of the results have been described inabstract (Boyd et al., 1993; Kelly et al., 1994).
MATERIALS AND METHODS
One hundred and two adult male Sprague-Dawley rats(Harlan Industries) weighing 200–360 g at the time ofsurgery weremaintained under standard laboratory condi-tions, including a 12:12 hour light:dark schedule, 23°C,
40–60% humidity, and ad libitum access to water andpellets. All procedures were conducted in accordance withAAALAC guidelines and were approved by the PurdueUniversity Animal Care and Use Committee.
Injection of PHA-L and perfusion
For tracer injections, the rats were anesthetized withsodium pentobarbital (60 mg/kg i.p.) and treated withatropine sulfate (1 mg/kg s.c.). For each animal, the floor ofthe fourth ventricle, vagal trigone, and surface of themedulla were exposed surgically, and PHA-L (Vector Labo-ratories, Burlingame, CA) dissolved in 10 mM sodiumphosphate-buffered saline, pH 8.0 (2.5% w/vol), was in-jected iontophoretically through a glassmicropipette (WorldPrecision Instruments; OD 5 20 µm, ID 5 15 µm) with aMidgard Precision Current Source unit (model CS-3).Individual injections consisted of 5 µA anodal current, 7seconds on/7 seconds off, and were typically for 20 minutes(the time was 15 minutes or less for 12 injections).The number of DMNX injections varied, with different
animals receiving one (n 5 5), two (both left: n 5 8, sevencontrol rats and one experimental rats; one left and oneright: n 5 46), three (one left, two right; n 5 5), or four (tworight, two left; n 5 38) injections. The rostrocaudal andmediolateral coordinates of the individual injections in agiven rat were spaced so that, in cases with two PHA-Linjections, one site was 100–200 µm rostral and 50–150 µmlateral to the other. In rats with four injections, two (oneright and one left) were near the caudal level of the areapostrema, and two were approximately 750 µm rostral tothis, although the right and left sites were at slightlydifferent levels. The pipette tip occasionally broke duringan individual series of injections, wherein the injectionswere usually completed with the larger tip (up to 50 µm).Alternatively, after a fourth injection site, the internalpipette diameter could be as small as 3 µm (from blockage).Following the PHA-L injections, the incision was su-
tured, and Demerol hydrochloride (20 mg/kg, i.m.) wasadministered as an analgesic. Following a postinjectionsurvival time of 12–41 days (20 days gave optimum label,and this survival time was used after trials), the animalwas perfused. The evening before the animal was sched-uled for perfusion, a timed fast/refeed schedule was imple-mented to increase the contents (i.e., the amount ofdistension) of the GI tract, and particularly the stomach,at perfusion. Animals were deprived overnight (total of 16hours) and then given 15 g of food 1 hour before perfusion.The rats were deeply anesthetized with a lethal dose ofsodium pentobarbital administered intraperitoneally be-fore they received a 0.1 ml injection of heparin in the leftventricle, followed by transcardial perfusion with 200 ml0.9% NaCl solution at 40°C, followed by 750 ml of 4%paraformaldehyde in 0.1 M sodium potassium phosphatebuffer, pH 7.4, at 4°C.
Preparation of tissue, counterstaining,and immunocytochemistry
Brains were postfixed in 4% paraformaldehyde with15% sucrose for 24 hours at 4°C. The medulla was thencryostat sectioned at 56 µm in the transverse plane, andsections were collected in 0.02 M potassium phosphate-buffered saline (KPBS), pH 7.4. The guts were openedalong themesenteric border, rinsed clean with running tapwater, and postfixed for 24 hours in 4% paraformaldehyde.Separate complementarymuscular (longitudinal and circu-
82 M.-C. HOLST ET AL.

lar layers with enclosedmyenteric plexus) and submucosal(mucosa removed) wholemounts were prepared for thefirst 4-5 cm of small intestine and for the complete ventraland dorsal surfaces of the cecum. For the stomach, only thefull dorsal and ventral muscular wholemounts were regu-larly prepared.All gut peels had a 10 minute endogenous peroxidase
block (1:4 solution of 3% hydrogen peroxidase and metha-nol) followed by five 5-minute KPBS rinses. Wholemountswere then stained with 0.5% w/v Cuprolinic blue (Quino-linic phthalocyanine; cat. no. 17052; Polysciences,Warring-ton, PA) dissolved in 0.05 M sodium acetate buffer, pH 5.6,to which 1.0 M MgCl2 was added (pH becomes 4.9),according to the protocol in Holst and Powley (1995). Everyfourth section of the medulla was also counterstainedexcept with 0.05–0.1% Cuprolinic blue.Gut peels and medulla sections were blocked for nonspe-
cific protein in 10% normal rabbit serum (NRS), 1% bovineserum albumin (BSA), and 0.5% Triton X-100 in KPBSovernight at 4°C and were then incubated with goatanti-PHA-L agglutinin diluted 1:1,500 in 2% NRS, 1%BSA, and 0.3% Triton X-100 in KPBS for 48 hours at 4°C(initial trials) or for 16–24 hours on a shaking table (inlater runs, because no difference between the refrigeratedand room-temperature steps was detected).All subsequentincubations and rinses were at room temperature. Tissueswere rinsed twice for 10 minutes in the above dilutent;incubated for 2 hours in biotinylated anti-goat IgG (1:200)in the same dilutent; rinsed three times for 10 minutes inKPBS; incubated in avidin biotinylated horseradish peroxi-dase complex (Vectastain ABC Elite Kit used with the Aand B solutions reduced to 15 µl/ml to lower backgroundstaining) for 1.5 hours; rinsed twice for 5 minutes in KPBSand three times for 5 minutes in 0.05 M Tris buffer, pH 7.6;and reacted in 0.06% diaminobenzidine (DAB) with 0.015%hydrogen peroxide in Tris buffer for 2-4 minutes. Allantibodies and theABC kit were from Vector Laboratories.Wholemounts and medulla sections were mounted onsubbed slides, gut peels were flattened with weights for 1hour, and both were air dried overnight, dehydrated withan alcohol series, cleared in xylenes, and coverslipped inDPX (Aldrich, Milwaukee, WI) or Pro-Texx (Baxter Diag-nostics, McGaw Park, IL).
Controls
The five rats with the two left PHA-L injections receivedin addition a complete bilateral deafferentation of theabdominal GI tract. Only the stomach and a series ofmedulla sections (see below) were processed for immunocy-tochemistry. The bilateral deafferentation and its verifica-tion were performed according to the methods of Walls etal. (1995). The deafferentation consisted of a completeposterior truncal vagotomy (i.e., cauterization of the poste-rior vagal trunk in the abdomen; Powley et al., 1987) and,6–13 days later, a left intracranial afferent (dorsal) vagalrhizotomy (i.e., a lesion of the sensory vagal rootletsadjacent to the medulla). The left DMNX was exposed inthe same surgery as the dorsal vagal rhizotomy for twoPHA-L injections, as outlined above. One animal diedfollowing the PHA-L injection. For verification of thelesions, the rats were injected intraperitoneally with Fluo-rogold (Fluorochrome, Englewood, CO) 5–8 days prior tokilling and were injected in the left nodose ganglion withwheat germ agglutinin-horseradish peroxidase (WGA-HRP; Sigma, St. Louis, MO) 11–24 hours prior to perfu-
sion. The perfusion and handling of the stomach and brainfollowed the procedure used with all experimental rats,except that the stomach was distended with approxi-mately 10 ml saline during the perfusion rather than withthe fasting/feeding regime (because these rats had beenmaintained on a liquid diet). Furthermore, one-third of thebrain sections were processed for fluorescence microscopy(for Fluorogold verification of the truncal vagotomy; Foxand Powley, 1985), one-third for WGA-HRP using tetra-methyl benzidine (for verification of the vagal afferentrhizotomy), and one-third for PHA-L (for screening theinjection site).
Analysis of data
Gut peels and sections through the DMNX were exam-ined systematically for PHA-L label with a Leitz Ortho-plan II microscope. Individual PHA-L-labeled fibers orensembles of a few fibers were reconstructed by tracingthem at 3250–400 with a digitizing system consisting ofthe Eutectic Neuron Tracing System (formerly EutecticElectronics Inc., Raleigh, NC) and a Nikon Optiphot IImicroscope. For the tracings of individual fibers, we usedthe following criteria: the axon (or its collaterals) 1) couldbe followed from near the mesenteric attachment, 2) wasnot cut, broken, or otherwise incomplete, and 3) did notconverge with other labeled fibers on common sites, mak-ing the individual identity of the axon ambiguous. Adrawing tube attached to the Leitz was used for cameralucida records of submucosal fibers and injection sites.Colored photographs were taken with Kodak Royal Gold25 film in the Leitz automatic camera, transferred to aKodak Photo CD, and computer rendered (version 3.0;Adobe Photoshop, Mountain View, CA) to enhance sharp-ness and contrast.
RESULTS
PHA-L labeling pattern in the gastricmyenteric plexus
Dense pattern. Animals with injection sites focused inthe DMNX (see below) had dense axonal label in thegastric myenteric plexus. This pattern is described firstand is then compared with cases with less extensiveinjections and with fewer labeled fibers in the next section.Throughout the myenteric plexus of the stomach, la-
beled vagal preganglionic axons, collaterals, and varicosi-ties were so densely distributed that individual gangliawere demarcated by, and filled with, a complicated mesh-work of processes (Figs. 1, 2A). Large numbers of labeledvagal fibers outlined the interganglionic connectives (Figs.2B, 3), including the smaller secondary and tertiary inter-ganglionic connectives of the main plexus, thereby estab-lishing an intricate lattice or network throughout theentire plexus of the organ. Sometimes, a single axoncollateral or a few axon collaterals (Fig. 3, arrow) appearedto ‘‘short cut’’ by projecting directly from one ganglion toanother without following the circuitous paths of theconnectives. Throughout the densely stained pregangli-onic labyrinth, the individual axons were of fine caliberand were studded with many varicosities (Figs. 1, 2A-B,4A-C). These axonal swellings were smaller within theconnectives and were more prominent close to and withinthe ganglia (e.g., Figs. 1, 2A-B, 4D-F). Many myentericneurons were contacted by varicosities frommore than one
VAGAL PREGANGLIONIC PROJECTIONS TO ENS 83
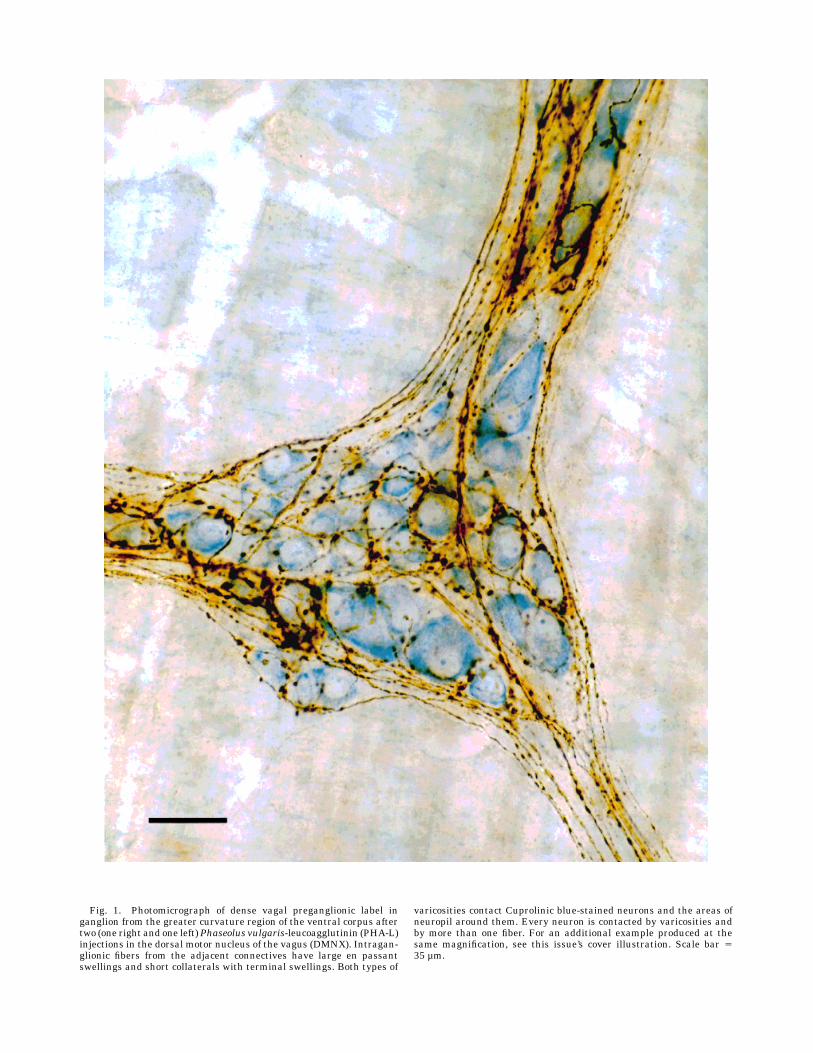
Fig. 1. Photomicrograph of dense vagal preganglionic label inganglion from the greater curvature region of the ventral corpus aftertwo (one right and one left)Phaseolus vulgaris-leucoagglutinin (PHA-L)injections in the dorsal motor nucleus of the vagus (DMNX). Intragan-glionic fibers from the adjacent connectives have large en passantswellings and short collaterals with terminal swellings. Both types of
varicosities contact Cuprolinic blue-stained neurons and the areas ofneuropil around them. Every neuron is contacted by varicosities andby more than one fiber. For an additional example produced at thesame magnification, see this issue’s cover illustration. Scale bar 535 µm.
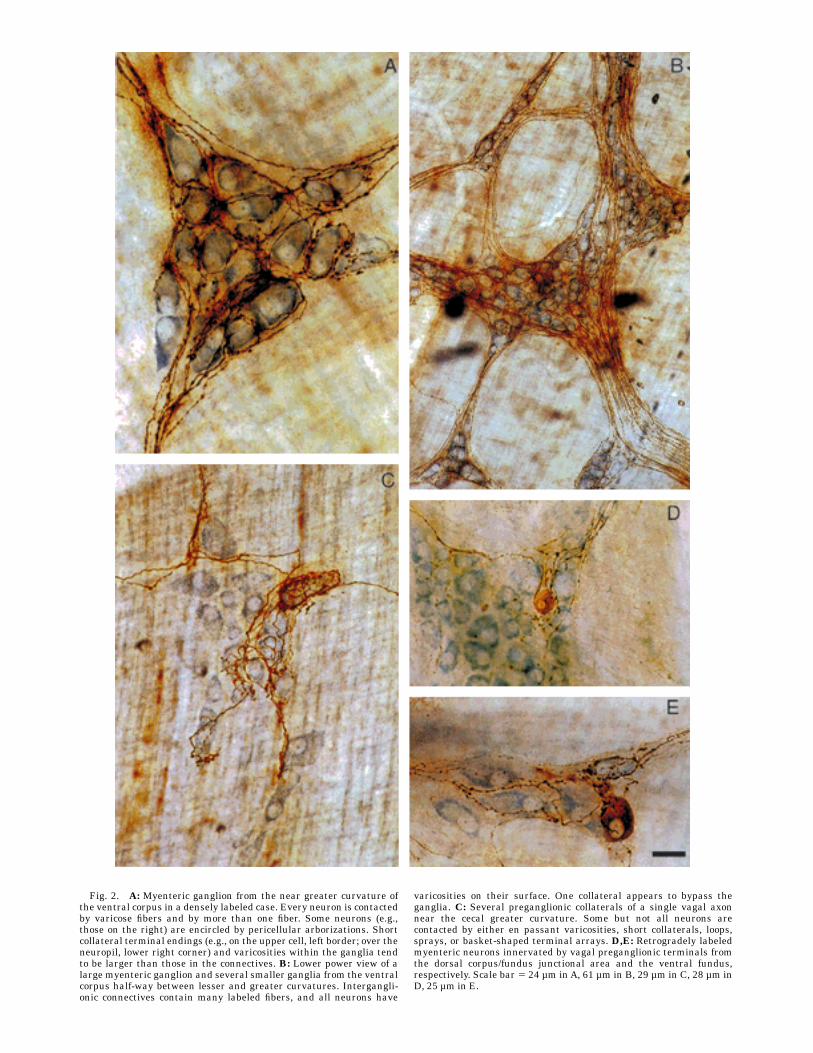
Fig. 2. A: Myenteric ganglion from the near greater curvature ofthe ventral corpus in a densely labeled case. Every neuron is contactedby varicose fibers and by more than one fiber. Some neurons (e.g.,those on the right) are encircled by pericellular arborizations. Shortcollateral terminal endings (e.g., on the upper cell, left border; over theneuropil, lower right corner) and varicosities within the ganglia tendto be larger than those in the connectives. B: Lower power view of alarge myenteric ganglion and several smaller ganglia from the ventralcorpus half-way between lesser and greater curvatures. Intergangli-onic connectives contain many labeled fibers, and all neurons have
varicosities on their surface. One collateral appears to bypass theganglia. C: Several preganglionic collaterals of a single vagal axonnear the cecal greater curvature. Some but not all neurons arecontacted by either en passant varicosities, short collaterals, loops,sprays, or basket-shaped terminal arrays. D,E: Retrogradely labeledmyenteric neurons innervated by vagal preganglionic terminals fromthe dorsal corpus/fundus junctional area and the ventral fundus,respectively. Scale bar 5 24 µm inA, 61 µm in B, 29 µm in C, 28 µm inD, 25 µm in E.
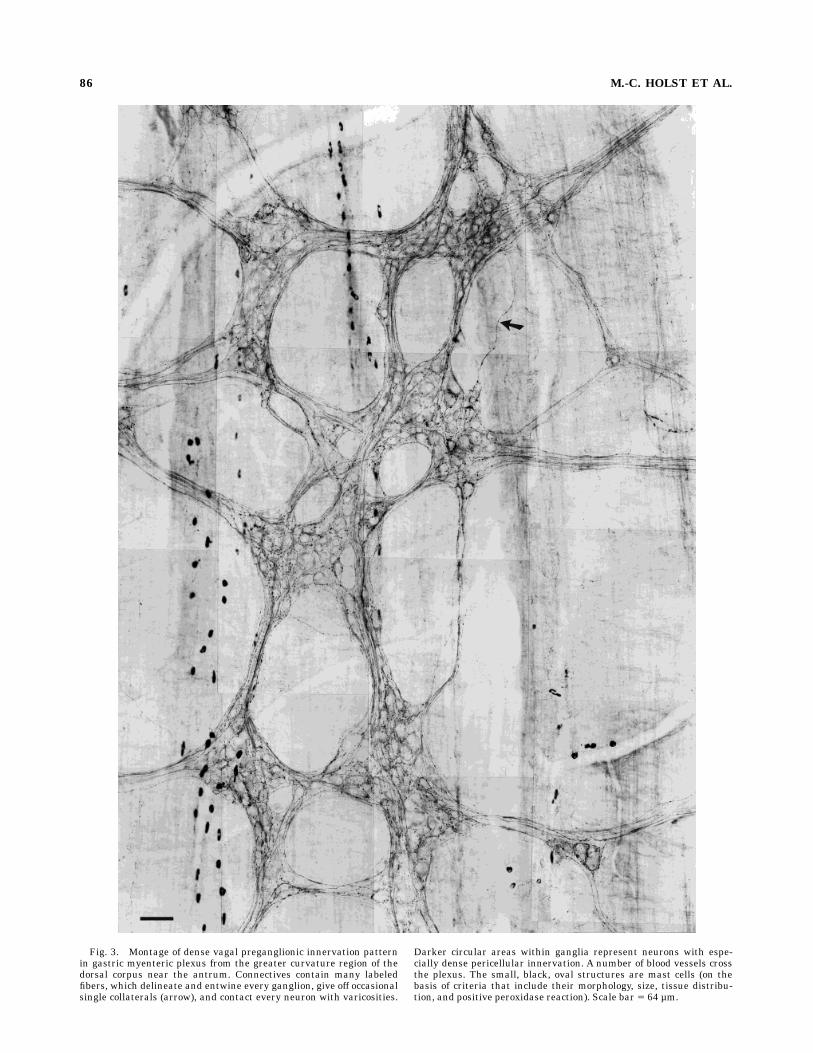
Fig. 3. Montage of dense vagal preganglionic innervation patternin gastric myenteric plexus from the greater curvature region of thedorsal corpus near the antrum. Connectives contain many labeledfibers, which delineate and entwine every ganglion, give off occasionalsingle collaterals (arrow), and contact every neuron with varicosities.
Darker circular areas within ganglia represent neurons with espe-cially dense pericellular innervation. A number of blood vessels crossthe plexus. The small, black, oval structures are mast cells (on thebasis of criteria that include their morphology, size, tissue distribu-tion, and positive peroxidase reaction). Scale bar 5 64 µm.
86 M.-C. HOLST ET AL.
converging axon (e.g., Figs. 1, 2A-B). However, someneurons were contacted with an especially heavy ring orhalo of varicosities (e.g., Figs. 2A-B, 3, 4A-C; indicated by
the darker areas scattered throughout the ganglia in lowerpower photographs). Some axons bypassed a ganglionwithout making apparent contacts, although they issued
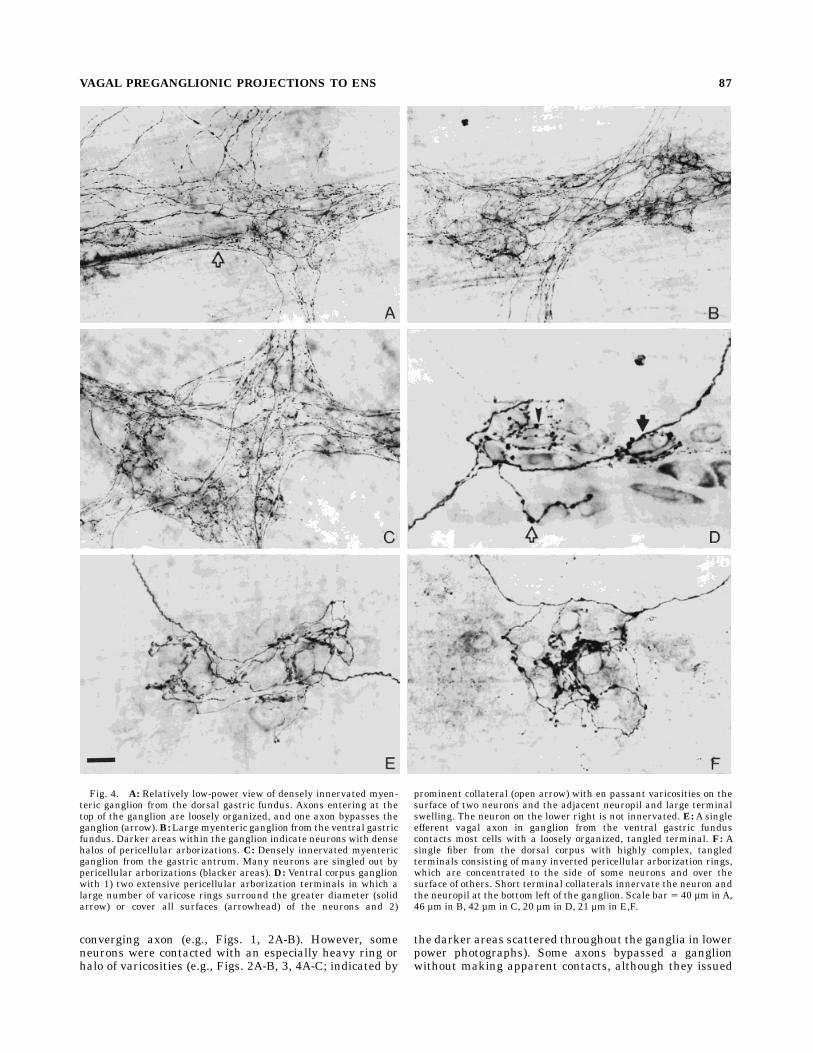
Fig. 4. A: Relatively low-power view of densely innervated myen-teric ganglion from the dorsal gastric fundus. Axons entering at thetop of the ganglion are loosely organized, and one axon bypasses theganglion (arrow).B: Largemyenteric ganglion from the ventral gastricfundus. Darker areas within the ganglion indicate neurons with densehalos of pericellular arborizations. C: Densely innervated myentericganglion from the gastric antrum. Many neurons are singled out bypericellular arborizations (blacker areas). D: Ventral corpus ganglionwith 1) two extensive pericellular arborization terminals in which alarge number of varicose rings surround the greater diameter (solidarrow) or cover all surfaces (arrowhead) of the neurons and 2)
prominent collateral (open arrow) with en passant varicosities on thesurface of two neurons and the adjacent neuropil and large terminalswelling. The neuron on the lower right is not innervated. E: A singleefferent vagal axon in ganglion from the ventral gastric funduscontacts most cells with a loosely organized, tangled terminal. F: Asingle fiber from the dorsal corpus with highly complex, tangledterminals consisting of many inverted pericellular arborization rings,which are concentrated to the side of some neurons and over thesurface of others. Short terminal collaterals innervate the neuron andthe neuropil at the bottom left of the ganglion. Scale bar 5 40 µm inA,46 µm in B, 42 µm in C, 20 µm in D, 21 µm in E,F.
VAGAL PREGANGLIONIC PROJECTIONS TO ENS 87

varicose endings in other ganglia of the plexus (e.g., Figs.2B, 3, arrow in Fig. 4A).Although the profuse preganglionic network occurred
throughout the stomach (ventral corpus, greater curvaturearea: Figs. 1, 2A; dorsal corpus, greater curvature area:Fig. 3; midventral corpus: Fig. 2B; ventral fundus: Fig. 2E;dorsal fundus: Fig. 4A; ventral fundus: Fig. 4B; dorsalantrum: Fig. 4C), sparsely labeled patches were foundeven in the densely labeled cases: Some areas had areduced number of fibers, some had only one or two axons,and, in some, it was difficult to find the poorly stainedremnants of an axon (an artifact associated with mechani-cal damage during wholemount preparation). These pat-terns of sparse labeling varied unsystematically fromanimal to animal and appeared to reflect the position of theinjection sites within the DMNX rather than an absence ofinnervation in a particular region of the stomach. Thesesparsely labeled regions were surrounded by areas withsuch dense and intricate networks that it was impossibleto trace single fibers for more than a short distance.Examination of the lesser curvature region revealed
bundles of fibers dividing from the parent gastric vagalbranches and entering the myenteric plexus. Within ashort distance, smaller bundles and single fibers split offfrom the main nerves to pass ventrally or laterally into thegeneral plexus. Once within the plexus, their courseappeared to be dictated by the position of the ganglia. Anumber of smaller branches returned to ganglia closer tothe esophagus, and a few of these fascicles passed to thecut edge of the lesser curvature (presumably to the otherside of the stomach). At the greater curvature, a number offibers turned and coursed for short distances (i.e., aboutone-quarter of the way) toward the lesser curvature beforeturning and running parallel to the longitudinal muscle.At this point, these fibers issued a number of collaterals ina series of ganglia and then ended. Along the greatercurvature, there were also small bundles coursing towardsthe lesser curvature from the greater. Some of theseappeared damaged (probably during the initial peel prepa-ration), and others were very short segments. Neverthe-less, these bundles appeared to arise both as a minorcontinuation of the innervation from one side of thestomach to the other around the greater curvature and asinnervation from a small number of vagal preganglionicfibers entering the stomach wall in neurovascular fasciclesat the greater curvature.Sparse pattern. The paths of single vagal pregangli-
onic fibers through the myenteric plexus (Figs. 4D–F,5A,C; distal parts of Fig. 5B,D) could be followed morereadily where sparse labeling occurred as a result of lesswell-positioned (outside the DMNX—see below) or lessextensive PHA-L injections sites. Individual axons rami-fied through the myenteric plexus. Collaterals circledthrough a number of ganglia, often returning to parallelthe parent original fiber (Fig. 5B–D) and looping fromganglion to ganglion, generally following a course from thelesser curvature toward the greater (see above). Theprominent varicosities on a fiber as it passed adjacent to orthrough a ganglion were most often seen in contact withneuronal somata or the surrounding neuropil. These puta-tive contacts occurred in two basic types: fiber swellingsresembling en passant endings and terminal swellings.Short terminal intraganglionic branches with one or no enpassant swellings and single terminal varicosities fre-quently arose from an axon as it passed through the
ganglion (e.g., Fig. 2A). Other intraganglionic terminalbranches were longer, with en passant boutons contactingseveral neurons as well as the adjacent neuropil beforeforming a final terminal swelling on another neuron (e.g.,Fig. 4D, open arrow).Within a ganglion, a single vagal motor axon could have
many short terminal branches passing to only the mostproximal neurons, longer branches to several myentericneurons, or an elaborate network of collaterals entwinedamong numerous somata.Anetwork of rings could encircleindividual neurons or small groups of neurons in the‘‘pericellular arborizations’’ (e.g., Figs. 2A, 4D) describedby Dogiel (1899). In addition to forming tightly apposedstrings of varicosities around myenteric neurons, theserings often extended into areas of neuropil between neu-rons, where, presumably, the varicosities contacted den-dritic processes. The pericellular arborizations had severalmodifications. One consisted of a varying number of vari-cose rings around a neuron, often with an additionalshowering of short terminal twigs, each of which had oneor two swellings (Fig. 4D). Another consisted of a ‘‘basket’’or ‘‘flask-like’’ enclosure of one or two isolated neurons (Fig.10F–H). A third common type was a complex tangle ofoverlapping loops or rings, one side of which could beinverted, so that the arborization was especially concen-trated to one side of several neurons and the adjacentneuropil (Fig. 4F). A somewhat less dense arrangement ofthis third form is seen on the single fiber illustrated inFigure 4E.From ganglion to ganglion, the same fiber might gener-
ate different patterns of selectivity. In some instances, thefiber would send a fine collateral or several branches into aganglion. In other cases, the axon would pass directlythrough a ganglion with only a few simple en passantappositions or none at all. Yet, within other ganglia, thefiber collaterals would encircle some neurons and notothers or would contact many or all of the neurons. Astriking feature of axons that innervated a smaller subsetof myenteric neurons in a given ganglion was that, fre-quently, the preganglionic endings appeared to be distrib-uted by some selective principle and not by a stochasticprocess. Such fibers singled out and formed a dense field ofvaricosities around one neuron, produced a singular enpassant varicosity apposed to other neurons, and had noapparent contact with others (e.g., Fig. 4D–F).Reconstructions of individual fiber were attempted for
the gastric myenteric plexus by using the criteria listed inMaterials and Methods. However, tracing the trajectory ofsingle axons was impractical in cases where the injectionsite was focused in the DMNX because of the extensiveoverlap and interdigitation of labeled processes. In experi-mental cases with less centrally placed injections andsparser gastric PHA-L, the complexity of the labeled axonswas such that tracing a single fiber with confidence wasnearly impossible due to the repeated divisions, extensiveramifications, and widely divergent projection fields. Thisdivergence was not restricted to the one area; it occurred inthe fundus (Fig. 5B), corpus (Fig. 5C), and antrum (Fig.5D). In contrast, many intestinal fibers could be moreeasily reconstructed (see below).Additional observations. Injections of PHA-L in the
dorsal vagal complex occasionally retrogradely labeledsomata in the myenteric ganglia. These neurons occurredin the gastric myenteric plexus of both the corpus and thefundus. Such retrogradely labeled neurons, in some in-
88 M.-C. HOLST ET AL.

Fig. 5. A:Montage of a small area of the single vagal efferent axontraced in B. The three traces are from the ventral myenteric plexus inthe gastric fundus (B), corpus (C,) and pyloric (D) regions, as indicatedin the respective sketches (top left in B–D). For these three traces,myenteric ganglia contacted by fibers in C andD and in the first part of
B are indicated by small, white-filled outlines. Part of traces B and D(from the lesser curvature and the pylorus, respectively) to the arrowsrepresent two or three fibers; trace C and distal parts of B and D aresingle fibers. Scale bar5 84 µm inA, 3.4 mm in B, 4.1 mm in C, 3.1 mmin D.

stances, were contacted reciprocally by vagal efferentvaricosities labeled with PHA-L (Fig. 2D,E).In a minority (seven) of the gastric wholemounts, we
observed a sparse number of axons with a different termi-nal pattern. These fibers (Fig. 10E) were only found in themuscle of the fundus. They formed intramuscular arrayswith short, straight collaterals (in the muscle) parallel tothe muscle fibers and have therefore been identified assensory axons. Similar fibers were not found in otherregions of the stomach or other GI tract peels, and nolabeled axons were found in the mucosa (which only has avagal afferent innervation) of otherwisewell-labeled PHA-Lcases sectioned through the pylorus (Powley et al., 1994).
PHA-L labeling pattern in the duodenalmyenteric plexus
The preganglionic vagal axons in the duodenum (Figs. 6,7) were similar to those seen in the stomach; however,some differences in overall pattern were noted. The fibersentered the duodenum from the mesenteric attachmentand branched repeatedly to form extensive projectionfields in the myenteric plexus. For example, the three orfour intertwined duodenal fibers traced in Figure 7E beganat the mesenteric border, completely traversed the organradially, and continued anally for approximately 2.3 cm. Incontrast to the pattern of the gastric myenteric plexus, theduodenal ganglia tended to be narrowly elongated andwere orientated in parallel rows running in the direction ofthe circular muscle (e.g., Fig. 7D). This organizationimposed a rectilinear and angular pattern on the vagalpreganglionic fibers. Individual axons coursed through theganglia, parallel to the circular muscle, for varying dis-tances before either collateralizing or bending at rightangles to pass to another row of ganglia (e.g., Figs. 6, 7D).Preganglionic fibers could extend for considerable dis-tances (sometimes several cm) through many ganglia withsimple en passant varicosities (Fig. 7A), issuing few if anycollaterals until reaching a point where they branched ex-tensively and formed abundant ganglionic terminals (Fig.7B,C).Although vagal preganglionic axons typically coursedcentrifugally from ganglion to ganglion, these fibers occa-sionally left a ganglion and bent within the plexus toreturn to the same or an adjacent ganglion (Fig. 6).In the duodenal peels, the vagal preganglionic innerva-
tion was never quite as dense as that observed in thecorresponding stomach wholemount, although the exten-sive projection fields of individual fibers still provided wideinnervation of the myenteric ganglia, and, in well-labeledcases, many ganglia received input from one or more fibers(Figs. 6, 7B,C). Many preganglionic axon arbors actuallyspanned much of the duodenal circumference, entering atthemesenteric border and traveling around the antimesen-tery surface to return almost to the mesentery on thecontralateral side (Fig. 7E; see also Fig. 3 in Powley et al.,1994). Individual duodenal fibers collateralized and di-verged within the plexus, fanning out asymmetrically,with an aboral bias (Fig. 7D).Althoughmost of these intestinal fibers entered from the
neurovascular bundles of the mesentery, the proximalduodenum also received projections of individual vagalpreganglionic axons that coursed from the gastric antrum,across the pylorus, to the duodenal bulb. The gastric antralfiber illustrated in Figure 5D was one of numerous fiberstraced from the junction with the pylorus rather than thelesser curvature. Similarly, fibers appeared to continueacross the pyloric end of the duodenal peels.
The form and distribution of varicosities and terminalspecializations of duodenal preganglionic axons resembledthose in the stomach. Within a ganglion, an axon couldeither contact a few of the neurons with simple en passantvaricosities (Fig. 7A), or it could contact withmore complex-ity many of the neurons (Fig. 7A–D). Pericellular arboriza-tions surrounding specific neurons within a ganglion couldbe either simple or complex (Fig. 7B,C). Loops, tangle-typeterminals (not shown—see below), and short terminalcollaterals with en passant varicosities and terminal bou-tons were common (Fig. 7B,C).Similar to the retrograde labeling that was observed in
the stomach (cf. Fig. 2 D,E), retrogradely labeled neuronswere also observed in the myenteric plexus of the proximalduodenum (not illustrated). These labeled enteric duodeno-bulbar neurons, like their counterparts in the stomachwall, were contacted by PHA-L-labeled varicose vagalefferent fibers.
PHA-L labeling pattern in the cecalmyenteric plexus
Preganglionic vagal fibers were sparser and displayed adifferent pattern in the cecum (e.g., Fig. 8H) than in theduodenum. They entered the cecum near the ileocecaljunction and fanned out parallel to the circular muscletoward the greater curvature. The shape of cecal myen-teric ganglia varied. Diamond-shaped clusters of neuronspredominated close to the lesser curvature, and elongated,radial forms resembling those in the duodenum prevailedin the rest of the plexus. Cecal preganglionic axons rami-fied extensively through the initial ganglia. The moredistal collaterals from these proximal divisions, however,could then pass for some distance through a number ofganglia with only a few simple varicosities or en passanttype swellings (similar to duodenal fiber; Fig. 7A) beforebranching or turning to contact an adjacent row of ganglia.Close to the mesenteric border of the lesser curvature, thepattern consisted of a dense meshwork with collateralsramifying from ganglion to ganglion, but the more distalcollaterals produced several separate radial elements par-allel to the circular muscle fibers, giving the overallprojection an appearance of a hand with splayed fingers.The terminals of individual cecal vagal preganglionic
fibers tended to be less complicated than those in theproximal gut, but they had some of the same characteris-tics as those in the stomach and duodenum (Fig. 8A–C,G).The simplest was the series of en passant endings men-tioned above. Terminal collaterals of varying lengths werefrequently issued within a ganglion. Simple arborizationswere common, with the collaterals frequently thinner thanthe parent fiber, and many ganglia were crossed by axonsissuing short collateral terminals (Fig. 8A,E,G). Sprays ofshort collaterals with prominent varicose endings (Fig. 8B)and single pericellular loops (Fig. 8A,B,G) were often atthe end of long intraganglionic collaterals. A less complextangled ending, consisting of loops and short collaterals,was a predominate form throughout the cecum (Fig. 8C,F). Encircling pericellular arborizations (Fig. 8D) and thebasket type of ending (Fig. 2C) were relatively rare.
PHA-L labeling pattern in the duodenal andcecal submucosal plexus
Vagal preganglionic projections to the submucosal plexusof the duodenum and cecum were sparse but exhibitedmany of the same characteristics as those in the myenteric
90 M.-C. HOLST ET AL.

Fig. 6. Montage of PHA-L-labeled vagal preganglionic axons in theduodenal myenteric plexus after two injections (one right and one left)in the DMNX. Fibers cross and make many angular turns within thegeneral plexus as well as between individual ganglia. Interganglionic
connectives are less pronounced than in the gastric myenteric plexus.Many neurons in this field are contacted by vagal motor fibers. Scalebar 5 62 µm.
VAGAL PREGANGLIONIC PROJECTIONS TO ENS 91

Fig. 7. Photomicrographs and traces of duodenalmyenteric pregan-glionic fibers. A: A single vagal fiber with simple en passant varicosi-ties on the surface of a number of neurons. This axon traveled througha number of ganglia before branching. B,C: Duodenal ganglia con-tacted by several fibers: Some neurons are contacted by more than onefiber (e.g., thin arrow in C), and several neurons were contacted withspecific pericellular arborizations (thick arrows in B and C) of varyingdegrees of complexity. The angularity of duodenal collaterals isparticularly evident. D: A single fiber forming part of the complex of
preganglionic fibers shown in E. The axon contacts a number ofelongated ganglia (gray-filled outlines) running in the direction of thecircularmuscle.E:Network of three or four fibers that extended for 2.3cm in the oral (left) to anal direction and covered most of theduodenum from one cut mesenteric border to the other. Many neuronsin this densely innervated area were contacted by fibers and theircollateral terminals. Arrows indicate the extent of the fiber shown inE. Scale bar 5 26 µm inA, 33 µm in B, 40 µm in C, 3.9 mm in D, 2 mmin E.
92 M.-C. HOLST ET AL.
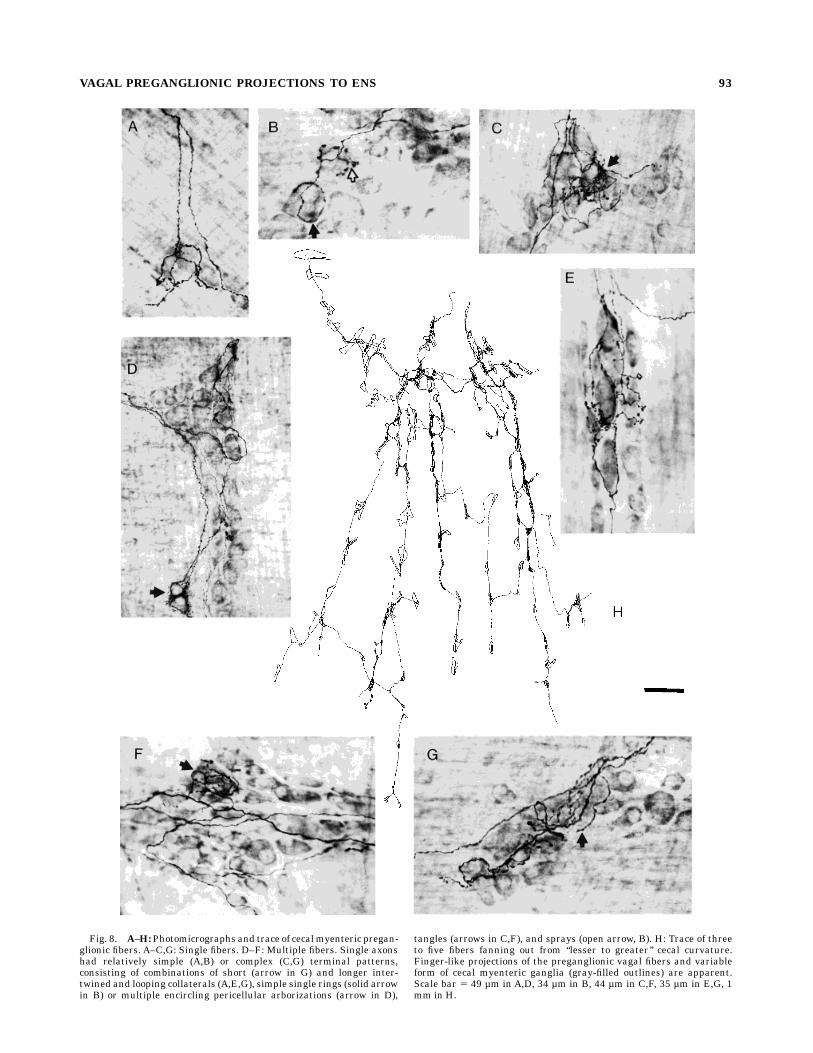
Fig. 8. A–H:Photomicrographs and trace of cecalmyenteric pregan-glionic fibers. A–C,G: Single fibers. D–F: Multiple fibers. Single axonshad relatively simple (A,B) or complex (C,G) terminal patterns,consisting of combinations of short (arrow in G) and longer inter-twined and looping collaterals (A,E,G), simple single rings (solid arrowin B) or multiple encircling pericellular arborizations (arrow in D),
tangles (arrows in C,F), and sprays (open arrow, B). H: Trace of threeto five fibers fanning out from ‘‘lesser to greater’’ cecal curvature.Finger-like projections of the preganglionic vagal fibers and variableform of cecal myenteric ganglia (gray-filled outlines) are apparent.Scale bar 5 49 µm in A,D, 34 µm in B, 44 µm in C,F, 35 µm in E,G, 1mm in H.
VAGAL PREGANGLIONIC PROJECTIONS TO ENS 93
plexus. Preganglionic fibers had numerous collaterals withen passant and terminal varicosities in proximity tosubmucosal neurons (Fig. 9A,C). These vagal efferents
contacted the neurons in an individual ganglion (Fig. 9B),forming short terminals, loops, and profusions of termi-nals resembling the tangled type (Fig. 9D,E). These submu-
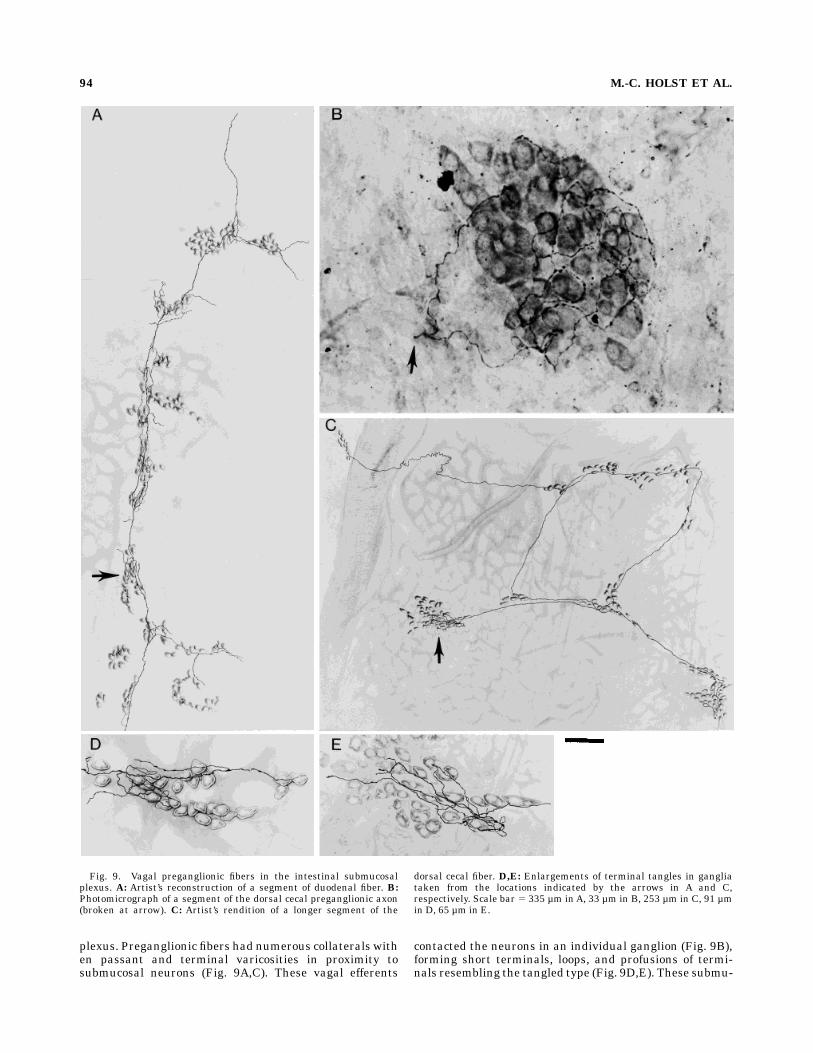
Fig. 9. Vagal preganglionic fibers in the intestinal submucosalplexus. A: Artist’s reconstruction of a segment of duodenal fiber. B:Photomicrograph of a segment of the dorsal cecal preganglionic axon(broken at arrow). C: Artist’s rendition of a longer segment of the
dorsal cecal fiber. D,E: Enlargements of terminal tangles in gangliataken from the locations indicated by the arrows in A and C,respectively. Scale bar 5 335 µm inA, 33 µm in B, 253 µm in C, 91 µmin D, 65 µm in E.
94 M.-C. HOLST ET AL.
cosal axons could only be traced for relatively shortdistances and had the appearance of being broken (e.g.,Fig. 9B, arrow) at one or both ends, as if they were eithercollaterals of a vagal efferent axon in the myenteric plexusor broken segments of longer submucosal fibers. Becausemost of these fibers were observed in rather denselystained areas with much connective tissue (i.e., in the lesswell cleaned areas of a peel) and were rarely found in the
more lightly stained areas, they could have been brokenduring the scraping required to remove the mucosa.
PHA-L injection sites in the DMNX
The tracer injections, through systematic variation ofplacement as well as normal variability, sampled most ofthe DMNX. In different animals, the PHA-L injection
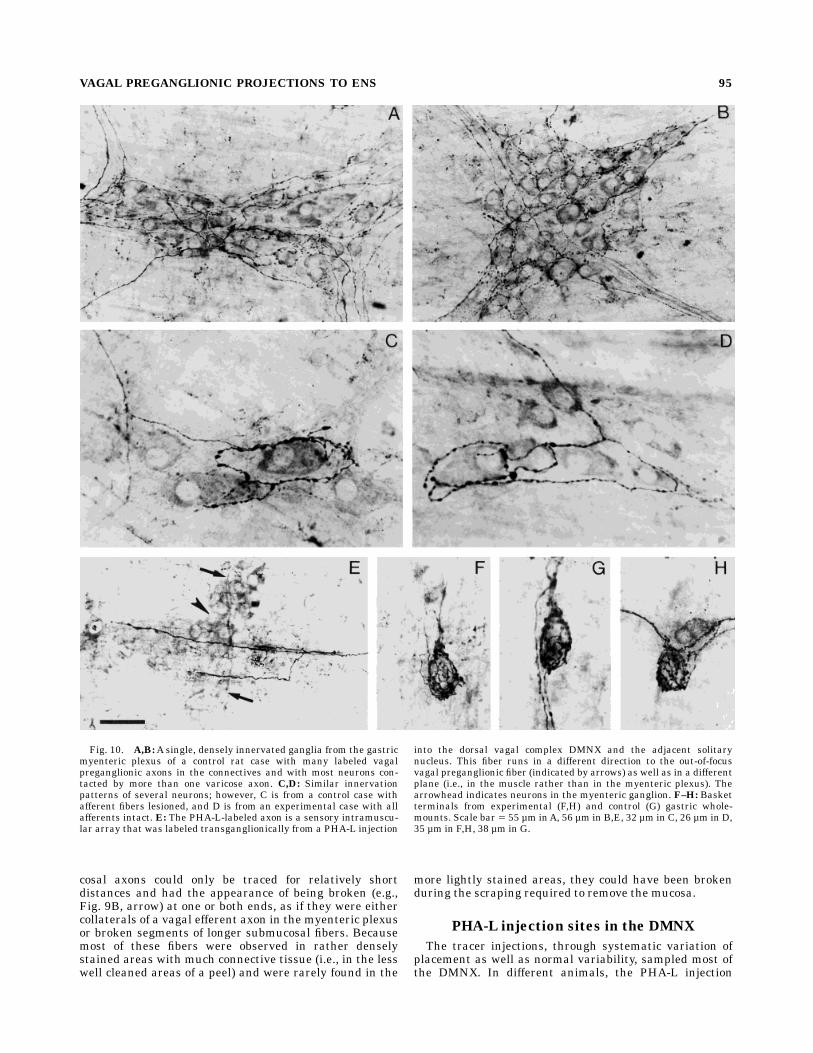
Fig. 10. A,B:A single, densely innervated ganglia from the gastricmyenteric plexus of a control rat case with many labeled vagalpreganglionic axons in the connectives and with most neurons con-tacted by more than one varicose axon. C,D: Similar innervationpatterns of several neurons; however, C is from a control case withafferent fibers lesioned, and D is from an experimental case with allafferents intact. E: The PHA-L-labeled axon is a sensory intramuscu-lar array that was labeled transganglionically from a PHA-L injection
into the dorsal vagal complex DMNX and the adjacent solitarynucleus. This fiber runs in a different direction to the out-of-focusvagal preganglionic fiber (indicated by arrows) as well as in a differentplane (i.e., in the muscle rather than in the myenteric plexus). Thearrowhead indicates neurons in the myenteric ganglion. F–H: Basketterminals from experimental (F,H) and control (G) gastric whole-mounts. Scale bar 5 55 µm inA, 56 µm in B,E, 32 µm in C, 26 µm in D,35 µm in F,H, 38 µm in G.
VAGAL PREGANGLIONIC PROJECTIONS TO ENS 95

series commenced either slightly rostral to, level with, orup to 400 µm caudal to the area postrema. Themost rostralset resulted in the densest gastric labeling of vagal pregan-glionic axons. This injection site continued to the rostralend of the area postrema or to more rostral levels of openmedulla. The center of an injection usually extended from60 µm to 300 µm in the horizontal plane, although thesurrounding labeled neurons, processes, and terminalscontinued in the rostrocaudal direction for some sections.Frequently, the dorsal part of the hypoglossal nucleus alsocontained some labeling. Concomitant axonal labeling inthe brainstem provided further evidence of the extent ofthe DMNX neuronal label (i.e., axons exited the lateralpole of the DMNX and projected ventrolaterally in fas-cicles to the lateral surface of the medulla) and also of theincidental involvement of the nucleus of the solitary tract(particularly farther rostrally in the dorsally situatedvagal afferent fascicles and the tract) and the hypoglossalnucleus (especially more caudally in the 12th nerve fas-cicles).The present PHA-L protocol was designed to maximize
labeling of DMNX efferents; hence, several variables lim-ited the topographic information obtained from the indi-vidual experimental cases. These procedural factors in-cluded 1) the use of multiple injections in individualanimals, 2) the employment of large injections that tendedto spread to several sites, 3) the use of long survival timesthat appeared to increase the diffusion and spread of thelectin within the medulla, and 4) the utilization of a highlysensitive processing protocol that may have overestimatedthe effective sphere of the PHA-L injection in the brain-stem. Overall, only a few general topographic patternsseemed to hold unequivocally: Cases with the dense pat-tern of gastric label described above always had well-focused injection sites (irrespective of the number of suchsites or the size of the pipette) with accompanying heavyneuronal staining of the DMNX. The location of the labeledDMNX neurons seemed to influence the sites of theoccasional sparsely labeled patches in the gastric myen-teric plexus as well as the number of labeled fibers presentin the duodenum and cecum.Axonal label in the intestineswas particularly sparse if the DMNX injections werefocused medially with little lateral spread.
Validation of preganglionic labeling pattern
The dorsal rhizotomy control experiments were de-signed to eliminate all vagal afferents to the GI tract and toavoid the infrequent concomitant transganglionic labelingof the intramuscular afferent fiber in the gastric fundus(see above). Three of the control cases produced verifiablelesions of all vagal afferents. Although the PHA-L labelingwas reduced to the dorsal surface of the stomach (becauseof the posterior trunk vagotomy), terminal patterns weresimilar to those described above. Labeled preganglionicfibers filled the connectives and ganglia in all parts of thestomach, and these efferents exhibited all of the character-istics observed in cases in which the vagal afferents wereintact (Fig. 10A,B). PHA-L-filled axons were varicose, thevaricosities were larger in the vicinity of the myentericganglia, and both en passant and terminal swellings ofendings were observed (Fig. 10C). Pericellular arboriza-tions, short collaterals, longer collaterals, loops (Fig. 10C),and basket (Fig. 10G) terminals resembling those inexperimental gastric wholemounts (Fig. 10D,F,H) occurred.
DISCUSSION
The present results describe an extensive network ofvagal preganglionic endings contacting themyenteric plex-uses of the GI tract, specifically, the stomach (fundus aswell as corpus and antrum), small intestine (duodenum),and large intestine (cecum). The network is comprised ofindividual efferent axons ramifying widely to producenumerous collaterals projecting to multiple ganglia andterminating in extensive, varicosity-rich telodendria in theganglia. In the stomach, for example, the axons aresufficiently divergent that they establish a network ofvagal efferent terminals, which encircle and make puta-tive contacts with virtually all myenteric neurons. Al-though this gastric preganglionic network appears to beinclusive, that is, all myenteric neurons receive contacts,examination of the arbors of individual axons indicate thatthe innervation pattern is not random: Their collateralsoften enter a myenteric ganglion and extensively contact asubset of one or more ENS neurons while making minimalor no contact with immediately neighboring neurons.Although preganglionic axons throughout the GI tractramify and diverge dramatically, the projections to theintestinal myenteric plexus appear less dense than thoseto the gastric myenteric plexus (see, however, discussion ofsampling issues below), and those to the submucosalplexus may be less dense than either myenteric projection.In contrast to the efferents projecting to the gastricmyenteric plexus, the intestinal myenteric and submuco-sal vagal preganglionic fibers appear to converge less onindividual ganglia or neurons. Before discussing this vagal-enteric network and its implications for processing in theautonomic nervous system, several control issues pertain-ing to our observations and the PHA-L technique should beconsidered.
Specificity of efferent label
An issue that is pivotal to all interpretations of thepresent results is whether the PHA-L-labeled axons andterminals we have described are efferent. The evidencesupporting this conclusion is extensive: 1) The patterns ofPHA-L label we have described were observed in thecontrol cases we prepared with verified vagal deafferenta-tion of the abdominal viscera. 2) Conversely, in complemen-tary experiments where vagal afferents were labeled bynodose injections of WGA-HRP after intracranial vagalmotor rhizotomies to eliminate efferents, the types ofprofiles we have described here in the myenteric plexushave not been observed, although distinctively differentpatterns characteristic of afferents (i.e., intramusculararrays, intraganglionic laminar endings) have been ob-tained (Wang and Powley, 1994; Powley and Wang, 1995;Wang, 1995). 3) Similarly, our earlier experiment employ-ing supranodose vagotomy to eliminate vagal efferents andlabel gastric fundic afferents with nodose ganglion injec-tions of DiI (Berthoud and Powley, 1992) found the sensorypattern of profiles mentioned above (cf. item 2), but not theprofile types described in the present paper. 4) In anadditional experiment, when DiI was injected into thenodose ganglion, and DiAwas injected into the DMNX, thecontrasting and very different sensory and motor profilepatterns were shown (Berthoud and Powley, 1992). 5)Even the cases with the heaviest axonal labeling in thepyloric region in the present experiment did not exhibitfibers in the pyloric mucosa (which is heavily innervated
96 M.-C. HOLST ET AL.

by vagal afferents, but not efferents; Powley et al., 1994). 6)In the present experiment, the densest labeling of theputatively preganglionic processes occurred with injec-tions that were centered in the DMNX and that labeled thegreatest numbers of DMNX neurons. In contrast, injec-tions situated peripherally with respect to the DMNXproduced much less extensive GI tract label. 7) Theendings described here, including their patterns of enpassant and terminal varicosities, are similar to thoseestablished as vagal efferent by different criteria (e.g.,silver: Schofield, 1962; zinc iodide and methylene blue:Roper and Ko, 1978; electron microscopy: Baluk andGabella, 1987; DiI: Berthoud et al., 1990). 8) Some of themotor profiles we have described are qualitatively similarto those identified by Kirchgessner and Gershon (1989) asefferent. However, the density, variety, and extent of theprofiles described here were not realized in this earlierPHA-L analysis. In summary, then, multiple converginglines of evidence substantiate that the axons described inthe present analysis are vagal efferents.
Comparison of present observationsand earlier experiments
In a recent electrophysiological experiment, Schemannand Grundy (1992) used an in vitro preparation of a 2 cm2
patch of guinea pig ventral gastric corpus, which preservedthe myenteric plexus and a branch of the vagus coursinginto the tissue. They reported that 98% of the ganglionicneurons near the lesser curvature received vagal efferentprojections. Also consistent with the present results, theseinvestigators observed that, when one neuron in a gan-glion was innervated, all neurons in the ganglion weretypically innervated. Furthermore, in another observationconsonant with the present data, Schemann and Grundyestimated from the latencies, amplitudes, and thresholdsof fast EPSPs that individual neurons were often inner-vated by multiple convergent vagal inputs. In anotherpotential parallel—specifically to the distinct pericellulararborization terminal types of single fibers on myentericneurons—Schemann and Grundy occasionally recordedfrom myenteric neurons, which received such a stronginput from one vagal fiber that fast EPSPs produced an‘‘action potential in an all-or-none fashion at thresholdstimulation currents.’’Given the highly complementary nature of these electro-
physiological observations and the present PHA-L results,one apparent discrepancy should be noted. Schemann andGrundy indicated that the percentage of myenteric neu-rons activated by vagal stimulation dropped progressivelyto as low as approximately 30% midway from the lesser tothe greater curvature of the corpus. They interpreted thisas a reduction in vagal innervation toward the greatercurvature. In contrast, we did not see a reduction in vagalefferent projections to myenteric ganglia near the ratgreater curvature (e.g., Figs. 1, 2A) nor at the midpoint ofeither ventral or dorsal corpus. Two sources of samplingbias (or a species difference) may have produced thereduction observed by Schemann and Grundy: Becausevagal fascicles fan out and execute a variety of divergentturns as they travel radially from the lesser to the greatercurvature, Schemann and Grundy’s patch of gastric corpuswith one vagal branch attached would have been partiallydenervated. The specimen would have lost axons thatentered the tissue obliquely or that wrapped around thegreater curvature and extended from the other side as well
as those entering the organ from neurovascular bundles atthe greater curvature. Furthermore, disruption, axonaldamage, and denervation caused by their dissection tech-nique could have biased the sample as they recordedfurther from the source of the spared vagal branch.Overall, if one allows for the discrepancy just discussed,
the earlier carbocyanine dye labeling results (Berthoud etal., 1990), the electrophysiological analysis of gastric vagal-myenteric interactions (Schemann and Grundy, 1992), andthe present observations with PHA-L describe compatibleand complementary patterns of vagal preganglionic projec-tions. At odds with this picture is essentially only theinitial PHA-L study of vagal efferents by Kirchgessner andGershon (1989). In their pioneering application of PHA-L,Kirchgessner and Gershon described much more limitedvagal efferent projections to the GI tract. They illustrateda few simple fibers and endings in the myenteric gangliaand concluded that the vagus did not project directly to thefundus or to the submucosal ganglia. Their results wereapparently limited by the undersampling that the PHA-Ltechnique can produce.
Sampling issues and limitationsof the PHA-L technique
Anterograde tracing strategies generally produce conser-vative estimates (or underestimates) of a projection, be-cause complete labeling is difficult to obtain. In the case ofthe present experiment, three sources of underestimationcan to be identified: 1) PHA-L labels only a fraction of theneurons within an injection sphere (Gerfen and Saw-chenko, 1984; Gerfen et al., 1989). 2) Both the shape of theDMNX and the orientation of its neurons make it particu-larly difficult to successfully iontophoretically inject theentire pool of GI motor neurons. The DMNX is a highlyelongated and fusiform structure extending longitudinallymore than 5 mm in the rat. The somata and dendrites ofthe ‘‘isodendritic’’ preganglionic neurons are also elon-gated in the longitudinal axis (Fox and Powley, 1992;Zhang et al., 1992). In the present experiments, the PHA-Ldid not label the entire population of DMNX neuronsbilaterally (even with four injection sites). Parenthetically,injection placement may also have resulted in a dispropor-tionate reduction in our intestinal preganglionic labeling.Most of our injection coordinates were centered in thegastric columns or subnuclei, which are situated medially(Fox and Powley, 1985), thus failing to optimally label theceliac columns that project to the distal duodenum andcecum but that lie lateral to the gastric subnuclei. 3) Therequirements of optimal PHA-L immunocytochemistry aredifficult to satisfy in the GI tract because of the tissuecomposition and organization of the gut. Processing strate-gies risk incomplete penetration of the reagents andexcessive disruption of the tissue, both of which reducelabeling (Powley et al., 1994; Holst and Powley, 1995). Ourearly trials suggested several details that could criticallyaffect the density and/or extent of the preganglionic label.When specimens were divided by separating the longitudi-nal and circular muscle layers (i.e., a common dissectionmethod), the myenteric plexus was often disrupted andfragmented, was attached variously to one muscle layer orthe other, or was completely lost. This problem wasavoided by leaving the external muscle layer as a singlewholemount and optimizing the processing to achievemaximal penetration. Other factors that appeared criticalincluded 1) sufficient survival times for PHA-L to accumu-
VAGAL PREGANGLIONIC PROJECTIONS TO ENS 97

late in terminal areas (less than 20 days could yield sparselabel), 2) adequate stretching of the tissue prior to fixationto minimize its thickness (the fast-refeed regimen was a‘‘physiological’’ means to assure the wall was thin atperfusion), and 3) good fixation and postfixation for stabi-lizing the PHA-L in the tissue.Such undersampling issues underscore the point that
even the extensive terminal network observed in thepresent analyses must represent a conservative estimateof the parasympathetic innervation. Sampling issueswouldalso seem to explain whyKirchgessner andGershon (1989)observed far less PHA-L labeling than reported here. Inaddition to technical problems relating to the tissue prepa-ration (cf. point 3 above), these authors typically employeda single, unilateral injection that was positioned morecaudally than the DMNX sites that we found to yieldoptimal labeling. Similarly, false negatives associated withincomplete label presumably explain a recent report oflabeled vagal efferents contacting roughly 47% of myen-teric neurons in a region of the fundus (Berthoud, 1995;the figure was not proffered as an absolute estimate).Rather than serving as a source for an unbiased calcula-tion, this latter figure should be questioned, because it wasbased on the notoriously problematic tracer DiA andbecause the completeness of the DMNX injections were notverified.Finally, for the present experiment, one potential source
of underestimation of the extent of divergence of indi-vidual fibers (in contradistinction to themeasure of conver-gence) should also be recognized. With our whole-organwholemounts, we were able to find fibers as they enteredthe organ from the mesenteric attachments and to tracethem through their extensive fields in the myentericplexus. This technique cannot exclude the possibility thatthese fibers were only collaterals of even more widelydivergent axons that had bifurcated before they enteredthe organ. The present observations, therefore, providelower bound estimates of the patterns of divergence ofsingle vagal preganglionics, but they would be systematicunderestimations if axonal divisions occur in the nervebranches or trunks proximal to the target organ.
Retrograde and transganglionictransport of PHA-L
Two somewhat anomalous aspects of our PHA-L labelshould also be considered. Although the occurrences wererelatively rare, we did observe both retrograde (see Fig.2D,E) and transganglionic (see Fig. 10E) label with injec-tions into the brainstem. The retrogradely labeled neuronspresumably resulted from PHA-L uptake by the centraltelodendria (i.e., in the dorsal vagal complex) of axonsoriginating from afferent ‘‘gastrobulbar’’ and ‘‘duodenobul-bar’’ neurons located in the myenteric ganglia. Similarly,neurons projecting from the myenteric plexus to the CNS(‘‘rectospinal’’) have been shown in distal parts of the GItract (Doerffler-Melly and Neuhuber, 1988; Neuhuber etal., 1993), and centripetal projections are found from theGI tract to the celiac ganglia (Messenger and Furness,1991; Cheng and Powley, 1994) as well as from the cardiacganglia to the nodose ganglion or brainstem (Cheng et al.,1997). Infrequent retrograde labeling with PHA-L hasbeen previously noted (see, e.g., Gerfen et al., 1989), andits determining variables have been discussed (Schofield,1990), although we were not able to correlate its occur-rence with any specific aspect of the PHA-L delivery. The
present observations, in addition, show that the novelgastrobulbar and duodenobulbar neurons, and, thus, notonly ENS postganglionic efferents, are innervated by vagalpreganglionic endings [cf. also the implication of Sche-mann and Grundy’s (1992) conclusion and our finding thatall neurons in a ganglion are innervated].The occasional well-labeled afferent endings (intramus-
cular arrays) in the circular and longitudinal muscle of theforestomach were easily distinguishable from the varicoseefferent endings that occurred in the plexus between themuscle sheets. Apparently, the afferent endings werelabeled transganglionically by PHA-L at the central termi-nals of nodose neurons. To our knowledge, transganglioniclabeling with PHA-L has not been previously reported. Forthe present analysis of preganglionic neurons and their GIprojections, the critical points are 1) that we observedneither transganglionic labeling of afferent endings in themuscle coats nor retrogradely labeled neurons in themyenteric plexus in experimental control animals thatreceived afferent rhizotomies before PHA-L injections and2) that we could readily distinguish each of these infre-quently occurring afferent patterns frommotor profiles.
Vagal-enteric organization:Mother cell vs. neural network pattern
The present results have major implications for tradi-tional views of the parasympathetic control of the GI tract.They suggest that the conventional view, namely, thatvagal efferents selectively contact a limited number of‘‘mother cells’’ or ‘‘command neurons’’ in the enteric ner-vous system, is incorrect. The present observations of thehighly collateralized, extensively ramifying vagal pregan-glionic telodendria and the webs of varicose pericellulararborizations enveloping most myenteric neurons (includ-ing gastrobulbar afferents as well as local afferents andinterneurons) are direct observations that challenge themother cell/command neuron explanation of vagal-entericinteractions.This revision in thinking about the articulation of vagal
efferents and neurons of the ENS has implications for ourunderstanding of the role of the CNS in the control of theENS. If the command neuron view, which has dominatedthe field since Langley, was correct and the vagal ‘‘finalcommon path’’ linking the CNS to the enteric nervoussystem had direct influence over only a few specializedenteric neurons, then one might expect central effects to bereduced to permissive go/no-go operations, with most ofthe graded effects that influence autonomic integrationoccurring locally in the gut wall. In contrast, however, ifthe vagus welds the brain and the ENS into a neuralnetwork, as the present results indicate, then one mightexpect the CNS to be an integrative focus capable ofgraded and varied responses for more of the physiologicalfunctions that influence gut function. This latter conclu-sion is reinforced by the present observations of neuronswithin the myenteric ganglia that were retrogradely la-beled by PHA-L injections in the vagal trigone. Suchobservations suggest that centripetal linkages betweenthe brain and the ENS are not restricted to the classicallysituated nodose ganglion neurons but may also includeafferent projections from neurons located in the ENS. Thenew observation that the somata of these centripetallyprojecting gastrobulbar and duodenobulbar afferents re-ceive contacts from preganglionic collaterals (see Fig.2D,E) further reinforces the concept of extensive and
98 M.-C. HOLST ET AL.

reciprocal interconnections between the brainstem andthe ENS. Thus, in addition to the extensive efferentprojections from the CNS to the ENS, afferent linkagesbetween gut and brain may be more extensive than hasbeen realized.Although the present observations suggest that a neural
network model is a better description of connectivity in theCNS-vagal-enteric motor axis than a command neuronmodel, vagal projections are not organized in a random orstochastic manner. An individual axon may ‘‘select’’ andpreferentially encircle or formbaskets of varicosities around(presumably to innervate) some neurons in a ganglion,although it does not form any varicosities in the vicinity ofother enteric neurons; or single neurons will receivepericellular arborizations (cf. Fig. 4D), whereas neighbor-ing enteric neurons are ‘‘ignored.’’ These patterns clearlysuggest that vagal projections in the ENS are determinedby organizing principles other than a random-ingrowthmodel. This same conclusion is suggested by the differentpatterns of divergence that characterize preganglionicneurons projecting to the stomach, duodenum, and cecum.The reconstructions of individual or small numbers of
vagal axons (Figs. 5, 7, 8) suggest another inference aboutvagal control of the GI tract. The majority, if not all, ofthese fibers are small, unmyelinated c-fibers with corre-spondingly slow conduction velocities. Given the morphol-ogy of the preganglionics and the biophysics of suchc-fibers, it may be that their motor capacity involvesgraded recruitment and the elicitation of different auto-nomic scores under different local conditions. It is possiblethat some collaterals or telodendria are of sufficiently finecaliber that action potentials do not invade themwith highfidelity under all conditions. These extensive axonal termi-nals could act as selective filters of action potentials as afunction of firing frequency or pattern. Local conditions inthe enteric ganglia may influence the security of theprojection from vagal preganglionic to postganglionic neu-ron. And, of course, different ending specializations, fromthe single, small varicosity to the elaborate arborizations,will influence the strength and pattern of signal expressedon postganglionic neurons. Overall, the structural detailsof the vagal-enteric network are rich and varied enough togenerate a complex and differentiated set of autonomicprograms.
ACKNOWLEDGMENTS
We thank E. Baronowsky for her considerable contribu-tions in the laboratory; D. Boyd for tissue preparation; W.Wollmann and T. Tsaousides for duodenal traces; Dr.David Kiernan, University of Western Ontario, for helpfulsuggestions about Cuprolinic blue; J. Mitchell for hisartistic skills; H. Urgell for diligently performing a multi-tude of services; M. Jarvinen, R. Phillips, and E. Walls fortheir comments on an earlier draft of the text; and RichardMyers-Walls, Center for Instructional Services, PurdueUniversity, for computer renditions of Figures 1 and 2.
LITERATURE CITED
Altschuler, S.M., L. Rinaman, andR.R.Miselis (1992) Viscerotopic represen-tation of the alimentary tract in the dorsal and ventral vagal complexesin the rat. In S. Ritter, R.C. Ritter, and C.D. Barnes (eds): Neuro-anatomy and Physiology of Abdominal VagalAfferents. Boca Raton, FL:CRC Press, Inc., pp. 21–54.
Auerbach, L. (1862) Uber einen Plexus myentericus. Breslau: Aug. Schulzand Company.
Baluk, P., and G. Gabella (1987) Fine structure of the autonomic ganglia ofthe mouse pulmonary vein. J. Neurocytol. 16:65–79.
Berthoud, H.-R. (1995) Anatomical demonstration of vagal input to nicotin-amide acetamide dinucleotide phosphate diaphorase-positive (nitrer-gic) neurons in rat fundic stomach. J. Comp. Neurol. 358:428–439.
Berthoud, H.-R., and T.L. Powley (1992) Vagal afferent innervation of therat fundic stomach: Morphological characterization of the gastrictension receptor. J. Comp. Neurol. 319:261–276.
Berthoud, H.-R., A. Jedrzejewska, and T.L. Powley (1990) Simultaneouslabeling of vagal innervation of the gut and afferent projections from thevisceral forebrain with DiI injected into the dorsal vagal complex in therat. J. Comp. Neurol. 301:65–79.
Boyd, D.B., J.B. Kelly, D.L. Kim, M.-C. Holst, and T.L. Powley (1993) Vagalefferent projections to the enteric nervous system: Command neuronhypothesis reconsidered. Soc. Neurosci. Abstr. 19:962.
Cheng, Z., and T.L. Powley (1994) Celiac ganglionic and vagal projections tothe rat duodenum compared using DiI/DiA dual labeling and confocalmicroscopy. Soc. Neurosci. Abstr. 20:1375.
Cheng, Z., T.L. Powley, J.S. Schwaber, and F.J. Doyle (1997) Vagal afferentinnervation of the atria of the rat heart reconstructed with confocalmicroscopy. J. Comp. Neurol. 381:1–17.
Davenport, H.W. (1989) Gastrointestinal physiology, 1895–1975: Motility.In S.G. Schultz, J.D. Wood, and B.B. Rauner (eds): Handbook ofPhysiology: The Gastrointestinal System. Bethesda, MD: AmericanPhysiological Society, pp. 1–101.
Doerffler-Melly, J., and W.L. Neuhuber (1988) Rectospinal neurons: Evi-dence for a direct projection from the enteric to the central nervoussystem in the rat. Neurosci. Lett. 92:121–125.
Dogiel, A.S. (1895) Zur Frage uber die Ganglien der Darmgeflechte bei denSaugethieren. Anat. Anz. 10:517–528.
Dogiel, A.S. (1899) Uber den Bau der Ganglien in den Geflechten desDarmes und der Gallenblase des Menschen und der Saugethiere. Arch.Anat. Physiol. Anat. Abstr. pp. 130–158.
Fox, E.A., and T.L. Powley (1985) Longitudinal columnar organizationwithin the dorsal motor nucleus represents separate branches of theabdominal vagus. Brain Res. 341:269–282.
Fox, E.A., and T.L. Powley (1992) Morphology of identified preganglionicneurons in the dorsal motor nucleus of the vagus. J. Comp. Neurol.322:79–98.
Gaskell, W.H. (1886) On the structure, distribution and function of thenerves which innervate the visceral and vascular systems. J. Physiol.(London) 7:2–80.
Gerfen, C.R., and P.E. Sawchenko (1984) An anterograde neuroanatomicaltracing method that shows the detailed morphology of neurons, theiraxons and terminals: Immunohistochemical localization of an axonallytransported plant lectin, Phaseolus vulgaris-leucoagglutinin (PHA-L).Brain Res. 290:219–238.
Gerfen, C.R., P.E. Sawchenko, and J. Carlsen (1989) The PHA-L antero-grade axonal tracing method. In L. Heimer and L. Zaborszky (eds):Neuroanatomical Tract-Tracing Methods 2: Recent Progress. NewYork:Plenum Press, pp. 19–47.
Holst, M.-C., and T.L. Powley (1995) Cuprolinic blue (quinolinic phthalocya-nine) counterstaining of enteric neurons for peroxidase immunocyto-chemistry. J. Neurosci. Methods 62:121–127.
Kelly, J.B., M.-C. Holst, and T.L. Powley (1994) Different vagal efferentprojection patterns in stomach, duodenum and cecum. Soc. Neurosci.Abstr. 20:1375.
Kirchgessner,A.L., andM.D. Gershon (1989) Identification of vagal efferentfibers and putative target neurons in the enteric nervous system of therat. J. Comp. Neurol. 285:38–53.
Langley, J.N. (1899) On inhibitory fibres in the vagus for the end of theesophagus and the stomach. J. Physiol. (London) 23:407–414.
Langley, J.N. (1922) Connexions of the enteric nerve cells. J. Physiol.(London) 56:39.
Langley, J.N., and H.KAnderson (1894) The constituents of the hypogastricnerves. J. Physiol. (London) 17:177–191.
Langley, J.N., and H.K. Anderson (1895) On the innervation of the pelvicand adjoining viscera. Part I. The lower portion of the intestine. J.Physiol. (London) 18:67–105.
Laughton, W.B., and T.L. Powley (1987) Localization of efferent function inthe dorsal motor nucleus of the vagus. Am. J. Physiol. 252:R13–R25.
Meissner, G. (1857) Uber die Nerven der Darmwand. Z. Ration. Med.8:364–366.
VAGAL PREGANGLIONIC PROJECTIONS TO ENS 99

Messenger, J.P,. and J.B. Furness (1991) Calbindin-immunoreactive nerveterminals in the guinea pig coeliac ganglion originate from colonic nervecells. J. Autonom. Nerv. Syst. 35:133–142.
Neuhuber, W.L., M. Appelt, J.M. Polak, W. Baier-Kustermann, L. Abelli,and G.-L. Ferri (1993) Rectospinal neurons: Cell bodies, pathways,immunocytochemistry and ultrastructure. Neuroscience 56:367–378.
Powley, T.L., and F.B. Wang (1995) Mapping regional distributions of vagalafferent projections to the gastrointestinal tract. Gastroenterology108:A670.
Powley, T.L., E.A. Fox, and H.-R. Berthoud (1987) Retrograde tracertechnique for assessment of selective and total subdiaphragmaticvagotomies. Am. J. Physiol. 253:R361–R370.
Powley, T.L., M.-C. Holst, D.B. Boyd, and J.B. Kelly (1994) Three-dimensional reconstructions of autonomic projections to the gastrointes-tinal tract. Microsc. Res. Technol. 29:297–309.
Ramon y Cajal, S. (1893) Sur les ganglions et plexus nerveux d’intestin.C.R. Soc. Biol. Paris 45:217–223.
Roper, S., and C.-P. Ko (1978) Synaptic remodeling in the partiallydenervated parasympathetic ganglion in the heart of the frog. In C.W.Cotman (ed): Neuronal Plasticity. NewYork: Raven Press, pp. 1–25.
Schemann, M., and D. Grundy (1992) Electrophysiological identification ofvagally innervated enteric neurons in guinea pig stomach. Am. J.Physiol. 263:G709–G718.
Schofield, B.R. (1990)Uptake ofPhaseolus vulgaris-leucoagglutinin (PHA-L)by fibers of passage. J. Neurosci. Methods 35:47–56.
Schofield, G.C. (1962) Experimental studies on the myenteric plexus inmammals. J. Comp. Neurol. 119:159–185.
Walls, E.K., R.J. Phillips, M.-C. Holst, J.S. Voreis, A.R. Perkins, L.E.Pollard, and T.L. Powley (1995) Selective vagal rhizotomies: A newdorsal surgical approach used for intestinal deafferentations. Am. J.Physiol. 269:R1279–R1288.
Wang, F.B. (1995) Inventory and Distribution of Vagal Afferent Projectionsin the Muscular Wall of the Rat Gastrointestinal Tract. PurdueUniversity, Ph.D. Thesis, 1995.
Wang, F.B., and T.L. Powley (1994) Taxonomy of vagal afferent projectionsto the gastrointestinal tract. Soc. Neurosci. Abstr. 20:1375.
Wang, F.B., M.-C. Holst, and T.L. Powley (1995) The ratio of pre- topostganglionic neurons and related issues in the autonomic nervoussystem. Brain Res. Rev. 21:93–115.
Wood, J.D. (1987) Physiology of the enteric nervous system. In L.R. Johnson(ed): Physiology of the Gastrointestinal Tract, 2nd Ed. NewYork: RavenPress, pp. 67–109.
Zhang, X., R. Fogel, and W.E. Renehan (1992) Physiology and morphologyof neurons in the dorsal motor nucleus of the vagus and the nucleus ofthe solitary tract that are sensitive to distension of the small intestine.J. Comp. Neurol. 323:432–448.
100 M.-C. HOLST ET AL.