Embed Size (px)
Citation preview

UvA-DARE is a service provided by the library of the University of Amsterdam (http://dare.uva.nl)
UvA-DARE (Digital Academic Repository)
The molecular mechanism of Atp8b1-deficiency
Groen, A.
Link to publication
Citation for published version (APA):Groen, A. (2009). The molecular mechanism of Atp8b1-deficiency.
General rightsIt is not permitted to download or to forward/distribute the text or part of it without the consent of the author(s) and/or copyright holder(s),other than for strictly personal, individual use, unless the work is under an open content license (like Creative Commons).
Disclaimer/Complaints regulationsIf you believe that digital publication of certain material infringes any of your rights or (privacy) interests, please let the Library know, statingyour reasons. In case of a legitimate complaint, the Library will make the material inaccessible and/or remove it from the website. Please Askthe Library: https://uba.uva.nl/en/contact, or a letter to: Library of the University of Amsterdam, Secretariat, Singel 425, 1012 WP Amsterdam,The Netherlands. You will be contacted as soon as possible.
Download date: 19 Aug 2019

Chapter 7
125
Chapter 7
Bile formation in mice lacking both Atp8b1 and Abcb4
Annemiek Groen, Cindy Kunne,
Karin van den Oever, Kam S. Ho-Mok,
Coen C. Paulusma and Ronald P.J. Oude Elferink
AMC Liver Center, Academic Medical Center, Amsterdam, The Netherlands
Manuscript

Bile formation in mice lacking both Atp8b1 and Abcb4
126
Abstract
Introduction Progressive Familial Intrahepatic Cholestasis (PFIC) can be caused by mutations
in different genes. ATP8B1 is associated with PFIC1 and ABCB4 is associated with PFIC3. Both
gene products mediate phospholipid translocation in the canalicular membrane in the liver, albeit
in opposite directions. ATP8B1 flips phosphatidylserine from the outer to the inner leaflet,
whereas ABCB4 flops phosphatidylcholine (PC) from the inner to the outer leaflet. Recent
results in Atp8b1-deficient mice show that the canalicular membrane is vulnerable to the
detergent effect of bile salts as witnessed by increased excretion of cholesterol,
phosphatidylserine (PS), sphingomyelin (SM) and canalicular ectoenzymes in bile. We created a
double knockout mouse lacking both Atp8b1 and Abcb4 (further designated as MF mice) to
investigate the importance of PS flipping in the presence and absence of PC flopping. Methods
Wild type, single and double transgenic mice of the same genetic background were infused for 2
hours with increasing amounts of taurocholate, after a depletion period of 90 minutes. Bile was
sampled and biliary phospholipid, cholesterol, bile salt and ecto-enzyme activity were measured.
Livers of untreated mice were used for histology, RT-PCR and western blotting. We also fed
these mice a diet containing 0.5% cholate to study chronic liver damage.
Results The Abcb4-/- phenotype, i.e. absence of biliary PC excretion, was also observed in
double transgenic mice. Surprisingly, the absence of both Abcb4 and Atp8b1 completely
abrogated the enhanced excretion of ectoenzymes APN and ALP that is observed in Atp8b1
single mutant mice. In addition, chronic administration of cholate led to less damage in double
transgenic mice as compared to Abcb4-/- mice. Conclusions These data demonstrate that the
function of Atp8b1 is particularly critical in the presence of Abcb4. These observations can be
interpreted by assuming that Abcb4-mediated flopping of PC makes the canalicular membrane
more fluid, which is counteracted by PS flipping by Atp8b1. In the absence of Abcb4 the latter
function is less relevant.

Chapter 7
127
Introduction
Apical plasma membranes of epithelial cells have a relative high content of sphingolipids in
the outer leaflet1. Together with cholesterol, sphingolipids (i.e. sphingomyelin and
glycosphingolipids) form rafts in which the lipids are in a rigid, liquid-ordered, structure that
confers detergent resistance to the membrane2. In the canalicular membrane of the hepatocyte
this detergent resistance is physiologically important for the protection against high bile salt
concentrations in the canalicular lumen. We have hypothesized that, under normal conditions,
the outer leaflet is largely in the liquid-ordered state because it consists for a large part of
cholesterol and sphingomyelin3.
In two inherited liver diseases the resistance towards bile salts is compromised, albeit in quite
different ways. Mutations in the ATP8B1 gene cause Progressive Familial Intrahepatic
Cholestasis (PFIC type 1). PFIC1 is characterized by pruritus, jaundice and growth retardation
caused by an impaired bile formation. This causes progressive liver damage, which can be
solved by liver transplantation4. ATP8B1 is a flippase that translocates phosphatidylserine (PS)
from the outer to the inner leaflet of the canalicular membrane5. Absence of ATP8B1 activity
will lead to an increase in PS content of the canalicular outer leaflet which in turn causes loss of
membrane rigidity due to the relative decrease in sphingolipid content.
Progressive Familial Intrahepatic Cholestasis Type 3 is characterized by the absence of
phosphatidylcholine (PC) from bile. This disease can be distinguished from PFIC type 1 on the
basis of histopathology and (high) serum GGT activity. PFIC3 is caused by mutations in the
ABCB4 gene, encoding a floppase for PC. ABCB4 translocates PC from the inner to the outer
leaflet of the canalicular membrane and thereby makes the phospholipid available for extraction
by bile salts. Excreted PC associates with bile salt micelles, reduces their detergent activity and
thereby indirectly protects the cells lining the biliary tree from further extraction of
phospholipids by bile salt micelles3.
These mechanisms create the paradoxical situation that two systems, involved in protection of
hepatocytes against bile salt-mediated lipid extraction, translocate phospholipids in opposite
directions within the canalicular membrane. This raises the question how the canalicular
membrane will behave in the absence of both systems. On the one hand one might expect that
bile salt toxicity is enhanced, because two systems of protection are absent; on the other hand
ATP8B1-mediated PS flipping might only be relevant in the presence of ABCB4-mediated PC
flopping because the first activity would counteract the fluidization of the membrane caused by

Bile formation in mice lacking both Atp8b1 and Abcb4
128
the second activity. We tested these possibilities in mice lacking both Abcb4 and Atp8b1, further
described as MF mice.
Methods
Animals. All experiments were performed with age-matched male mice at 2-6 months of age. Wild type,
Atp8b1G308V/G308V, Abcb4-/- and Atp8b1G308V/G308V,Abcb4-/- (MF) mice used in the experiments were of FVB/N
background and were bred in a pathogen free environment on a 12h light-dark cycle at the animal facility of the
Academic Medical Center. Mice were fed standard rodent chow or, when indicated, a commercial purified diet
(K4068.02, Arie Blok Diervoeders, Woerden, The Netherlands). Food and water were supplied ad libitum. All
animal experiments were approved by the institutional animal care and use committee (IUCAC) of the Academic
Medical Center.
In vivo bile salt infusions. The gallbladder and jugular vein were cannulated. Mice were placed on a heating pad
and body temperature was kept at 37°C. Prior to bile salt infusion, mice were depleted of endogenous bile salts by
bile collection without infusion. At t = 90 min, taurocholate (TC) was infused via the jugular vein. Infusion rates
were increased every 30 min with 400 nmol/min*100g (400-800-1200-1600 nmol/min*100g). Bile samples
(collected every 10 minutes) were analyzed for bile salt, choline-containing phospholipid, cholesterol, alkaline
phosphatase (ALP) and aminopeptidase N (APN or Cd13) activity,.
Bile salt, phospholipid, and cholesterol assays. Bile salt, choline-containing phospholipids, and cholesterol were
determined enzymatically. AP and Cd13/APN activities were determined using para-nitrophenylphosphate and L-
alanine-ß-naphtylamide-HBr as substrates, respectively6. All measurements were done on a Novostar analyzer
(BMG Labtech GmbH, Offenburg, Germany).
Serum BS levels were determined by HPLC/MS 7. Phospholipid species were determined by high performance
thinlayer chromatography (HP-TLC) as described previously8
Histology.
Livers were harvested and fixed in 4% paraformaldehyde. The tissue was paraffin-embedded and sectioned. The
sections were stained with 0.1% Sirius Red in saturated picric acid pH 2.0 and counterstained with haematoxylin9.
Fibrosis was scored in a blinded fashion as previously described10.
Results
The Atp8b1G308V/G308V mice that we have previously reported on11 were bred against
129/SvImJ background. Because the Abcb4-/- mice were bred against the FVB/N background we
have backcrossed Atp8b1G308V/G308V mice against the FVB/N background. Subsequently, the two
mutants were crossed to produce double transgenic mice. Thus, all experimental animals in this
report are against the same FVB/N background. Table 1 shows the basic parameters of the three
experimental groups. The table shows that the Abcb4-/- phenotype is also penetrant in
combination with the absence of Atp8b1; MF mice have similarly elevated plasma ALAT and
ASAT activities as well as increased bilirubin and bile salt levels as the Abcb4-/- mice. The bile

Chapter 7
129
parameters display a similar picture: bile flow is increased in Abcb4-/- single knockout as well as
in the MF mice. We have previously established that this is caused by an increased bile salt-
independent bile flow and we have hypothesized that this is caused by the observed bile duct
proliferation12. Finally, phospholipid excretion is similarly impaired in Abcb4-/- and MF mice.
We subsequently analyzed bile formation in these different genotypes during bile depletion and
subsequent infusion of taurocholate at increasing rates.
Table 1. Serum and biliary parameters in different genotypes
Parameter Wild type Atp8b1G308V/G308V Abcb4-/- Abcb4-/-Atp8b1G308V/G308V Plasma ASAT (U/l) ALAT (U/l) Bilirubin (µM) Bile salt (µM)
86.0 ± 45.3 53.0 ± 13.6 2.0 ± 2.3
0.6 ± 0.06
93.0 ± 53.8 44.0 ± 8.60 2.0 ± 2.3
1.9 ± 1.23
289.0 ± 84.40** 457.0 ± 103.8**
5.0 ± 2.0 10.7 ± 7.53*
345.0 ± 267.5 226.0 ± 179.0
4.0 ± 0.0 12.8 ± 16.9
Bile Flow(µl/min.100g) Bile salt (nmol/min.100g) Phospholipids (nmol/min.100g) Cholesterol (nmol/min.100g)
8.2 ± 0.6 197 ± 110 17.4 ± 13.2 0.9 ± 0.5
9.0 ±1.7 344 ± 150 37.4 ± 13.2 2.6 ± 0.5*
13.3 ± 1.8** 279.3 ± 94.12 2.6 ± 2.1** 0.8 ± 0.6
9.0 ± 1.8 324 ± 115 1.0 ± 0.5** 2.1 ± 1.0
Body weight 29.0 ± 2.00 33.4 ± 1.10 30.7 ± 0.30 29.3 ± 2.40 Liver weight (% of body weight)
4.4 ± 0.2 4.5 ± 0.2 7.0 ± 0.3** 5.9 ± 0.6**
Data are from animals that had been fed lab chow ad libitum. Biliary parameters were measured in samples collected during the first 10 minutes after gallbladder cannulation. Data are from 4 to 7 animals of each genotype. * p<0.05, **p<0.01
Fig. 1 shows the main parameters of biliary excretion in all four genotypes. Bile salt and
phospholipid excretion were not different in wild types and Atp8b1G308V/G308V mice. The
increased cholesterol excretion that we have reported before in Atp8b1G308V/G308V mice on a
C57Bl/6 background6 was only significant during the depletion phase in mice on a FVB/N
background. This is probably related to the fact that during maximal infusion of taurocholate
cholesterol excretion is much higher in FVB/N mice (>8nmol/min.100gr) than in C57Bl/6 mice
(<4nmol/min.100gr), whereas maximal bile salt excretion in FVB/N mice is about 2-fold lower
than in C57Bl/6 mice. Hence, bile salt-driven cholesterol excretion is about 4-fold higher in
FVB/N mice compared to C57Bl/6 mice. We assume that this is caused by substantially higher
Abcg5/8 expression in FVB/N mice compared to C57Bl/6. In MF mice biliary bile salt excretion
started to fall during maximal taurocholate infusion, which may suggest that bile salt-induced
cholestasis is setting in. Strikingly, phospholipid excretion in MF mice was as low as in Abcb4-/-
mice. This shows that neither in the presence nor in the absence of Atp8b1 extraction of PC from
the canalicular membrane is significant when Abcb4 is lacking.

Bile formation in mice lacking both Atp8b1 and Abcb4
130
Figure 1. Bile formation in mice of various genotypes. Mice were cannulated and bile was collected for 90 minutes. Subsequently, taurocholate was intravenously infused at infusion rates that were increased every 30 min with 400 nmol/min*100g (400-800-1200-1600 nmol/min*100g). Top panels: bile salt excretion. Middle panels phospholipid excretion. Lower panels cholesterol excretion. Left panels: Closed squares: wild type mice; open squares: Atp8b1G308V/G308V mice. Right panels: closed circles: Abcb4-/- mice; open circles: MF mice. The data represent averages from 5-7 animals per group with standard deviations.

Chapter 7
131
We have previously reported that excretion of ectoenzymes into bile is strongly enhanced in
Atp8b1G308V/G308V mice. Figure 2 shows that this is also the case in Atp8b1G308V/G308V mice on a
FVB/N background; the excretion of both aminopeptidase N and alkaline phosphatase were
significantly increased in mice lacking Atp8b1. However, the strong excretion of both enzymes
into bile was completely abrogated in MF mice. Hence, the canalicular membrane of MF mice is
apparently much more bile salt resistant that that in the Atp8b1G308V/G308V mice.
To further investigate this finding we also analyzed the phospholipid composition of bile
during taurocholate induced bile formation (i.e. in the last sample during infusion). Because
phospholipid excretion is a bile salt-driven process, we normalized the data for the amount of
bile salt in the same samples. Fig. 3 shows that similar differences can be observed between MF
and Abcb4-/- as between wild type and Atp8b1G308V/G308V mice: in both cases of Atp8b1 deficiency
a higher biliary content of PS and SM is detected. In the case of the MF mice also significant
increases in PI and PC were observed. In conclusion, Atp8b1 deficiency causes a similar relative
increase in phospholipid extraction in the absence of Abcb4 as in its presence, but the absolute
level of phospholipid extraction is much lower than in the presence of Abcb4.
To analyze chronic liver damage in this model of double deficiency, we also studied the
effects of chronic bile salt administration via a 0.5% cholate-supplemented diet. Mice from the
various genotypes were fed with this diet for 7 days. In these animals bile was collected and liver
specimens were analyzed for fibrosis. Fig. 4A shows the excretion of phospholipids during the
first 15 min of bile duct cannulation. The data in the figure were normalized for bile salts, which
provides the best reflection of the excretion capacity. These data largely confirm those from the
acute infusion experiments in that total phospholipid excretion is strongly reduced in both the
Abcb4-/- and the MF mice. However, there was nevertheless a significantly higher phospholipid
excretion in MF mice compared to Abcb4-/- mice. In addition, there was a tendency towards
higher cholesterol excretion in the Atp8b1G308V/G308V and the MF mice compared to wild type and
Abcb4-/-, but this was not significant (Fig. 4B). The excretion of the ectoenzymes aminopeptidase
N (Fig. 4C) and alkaline phosphatase (Fig. 4D) into bile also provided a similar picture as the
acute infusion experiment. In single Atp8b1G308V/G308V mice a high activity of both enzymes was
observed which was completely (APN) or partly (ALP) abrogated in the double knockout (MF)
mice.

Bile formation in mice lacking both Atp8b1 and Abcb4
132
Figure 2. Biliary excretion of ectoenzymes Mice were cannulated and bile was collected for 90 minutes. Subsequently, taurocholate was intravenously infused at infusion rates that were increased every 30 min with 400 nmol/min*100g (400-800-1200-1600 nmol/min*100g). Top panels Aminopeptidase N (APN). Lower panels: alkaline phosphatase (ALP). Left panels: Closed squares: wild type mice; open squares: Atp8b1G308V/G308V mice. Right panels: closed circles: Abcb4-/- mice; open circles: MF mice. The data represent averages from 5-7 animals per group with standard deviations.
Figure 3. Biliary phospholipid species in mice of various genotypes. Samples from the experiment of Figure 1 (taken at 160 min) were analyzed for phospholipid species by HPTLC. The data were normalized for the amount of total bile salt in the same sample. Please note that the ordinate has a logarithmic scale. The data represent mean values from 5 animals in each group with standard deviations. Asterisks indicate a significant difference (p<0.05) with the corresponding group lacking Atp8b1. The data represent averages from 5-7 animals per group with standard deviations.

Chapter 7
133
Quite strikingly, serum aminotransferase (ALAT) activity learned that hepatic damage was
considerably lower in the MF mice compared with both single knockout models (because of the
strong variability of serum ALAT in Atp8b1G308V/G308V the difference between these and MF mice
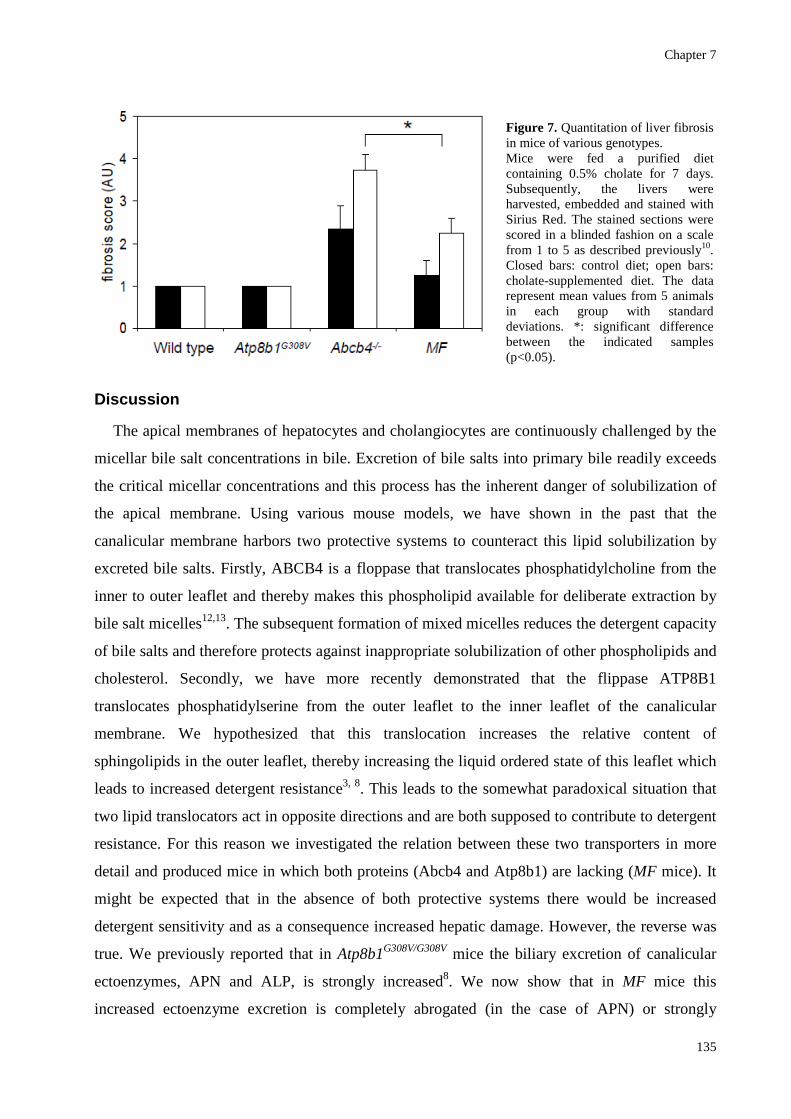
was not significant (Fig. 5). We therefore also analyzed fibrosis in all genotypes by picro-sirius
red staining and we found a very strong difference in fibrosis between the single Abcb4-/- mice
and the double deficient MF mice (Fig. 6). To substantiate this finding, we scored section from
all animals in a blinded fashion (Fig. 7). This confirmed that the fibrosis score was significantly
lower in the double deficient MF mice as compared to single Abcb4-/- mice.
Figure 4. Bile composition in mice of various genotypes after feeding a cholate-supplemented diet. Mice were fed a purified diet containing 0.5% cholate for 7 days. Subsequently, bile was collected for 15 minutes and analyzed for its composition. Panel A: phospholipid/bile salt ratio. Panel B: cholesterol/bile salt ratio. Panel C: aminopeptidase N (APN) activity. Panel D: alkaline phosphatase (ALP) activity. The data represent mean values from 5 animals in each group with standard deviations. #: significant difference between the indicated samples (p<0.05). *: significant difference with wild type (p<0.05).
A. B.
C. D.

Bile formation in mice lacking both Atp8b1 and Abcb4
134
Figure 5. Serum aminotransferase activity in mice of various genotypes. Mice were fed a purified diet containing 0.5% cholate for 7 days. The data represent mean values from 5 animals in each group with standard deviations. #: significant difference between the indicated samples (p<0.05). *: significant difference with wild type (p<0.05).
Figure 6. Liver fibrosis in mice of various genotypes. Mice were fed a purified diet containing 0.5% cholate for 7 days. Subsequently, the livers were harvested, embedded and stained with picro sirius red. While the single Abcb4-/- mice display extensive fibrosis, this is nearly absent in double deficient MF mice.
Chapter 7
135
Discussion
The apical membranes of hepatocytes and cholangiocytes are continuously challenged by the
micellar bile salt concentrations in bile. Excretion of bile salts into primary bile readily exceeds
the critical micellar concentrations and this process has the inherent danger of solubilization of
the apical membrane. Using various mouse models, we have shown in the past that the
canalicular membrane harbors two protective systems to counteract this lipid solubilization by
excreted bile salts. Firstly, ABCB4 is a floppase that translocates phosphatidylcholine from the
inner to outer leaflet and thereby makes this phospholipid available for deliberate extraction by
bile salt micelles12,13. The subsequent formation of mixed micelles reduces the detergent capacity
of bile salts and therefore protects against inappropriate solubilization of other phospholipids and
cholesterol. Secondly, we have more recently demonstrated that the flippase ATP8B1
translocates phosphatidylserine from the outer leaflet to the inner leaflet of the canalicular
membrane. We hypothesized that this translocation increases the relative content of
sphingolipids in the outer leaflet, thereby increasing the liquid ordered state of this leaflet which
leads to increased detergent resistance3, 8. This leads to the somewhat paradoxical situation that
two lipid translocators act in opposite directions and are both supposed to contribute to detergent
resistance. For this reason we investigated the relation between these two transporters in more
detail and produced mice in which both proteins (Abcb4 and Atp8b1) are lacking (MF mice). It
might be expected that in the absence of both protective systems there would be increased
detergent sensitivity and as a consequence increased hepatic damage. However, the reverse was
true. We previously reported that in Atp8b1G308V/G308V mice the biliary excretion of canalicular
ectoenzymes, APN and ALP, is strongly increased8. We now show that in MF mice this
increased ectoenzyme excretion is completely abrogated (in the case of APN) or strongly
Figure 7. Quantitation of liver fibrosis in mice of various genotypes. Mice were fed a purified diet containing 0.5% cholate for 7 days. Subsequently, the livers were harvested, embedded and stained with Sirius Red. The stained sections were scored in a blinded fashion on a scale from 1 to 5 as described previously10. Closed bars: control diet; open bars: cholate-supplemented diet. The data represent mean values from 5 animals in each group with standard deviations. *: significant difference between the indicated samples (p<0.05).

Bile formation in mice lacking both Atp8b1 and Abcb4
136
reduced (in the case of ALP). Apparently, the canalicular membrane of Atp8b1G308V/G308V mice is
more sensitive to bile salt-mediated extraction than the membrane in the double deficient mice.
This can be understood if it is assumed that the canalicular membrane of Atp8b1G308V/G308V mice
is more in a liquid disordered state than that of MF mice. The simplest hypothesis to explain this
difference is that Abcb4 is the most important determinant for creating this liquid disordered
state by flopping PC into the outer leaflet. In line with this hypothesis we observed that the
abrogation of PC excretion in the absence of Abcb4 is a dominant phenotype that is fully
penetrant in the MF mice as well. Thus, although deficiency of Atp8b1 increases the relative
amount of glycerophospholipids in the outer leaflet and thereby brings this leaflet in a more
liquid disordered state, this does not lead to major phospholipid and cholesterol extraction as
witnessed by our data from Fig. 1 and 3A. The MF mice do excrete significantly more
phospholipid into bile than the Abcb4-/- mice but this is still more than 20-fold lower than in wild
type and Atp8b1G308V/G308V mice. These data would suggest that Atp8b1 is particularly important
in the presence of Abcb4 (i.e. when PC is flopped to the outer leaflet). On the basis of these data
we propose the following model: Abcb4 flops PC for excretion into bile, but the inherent
consequence of this is an increased liquid disordered state of the outer leaflet with increased
detergent sensitivity. Atp8b1, by flipping PS to the inner leaflet, tries to counteract this effect as
much as possible and brings the outer leaflet back into the liquid ordered state. Hence, Atp8b1 is
particularly important as a defensive mechanism in the presence of PC flopping by Abcb4.
Analysis of our present and previous experiments6 shows that there is a major difference in
cholesterol excretion between FVB/N mice and C57Bl/6 mice on the other hand. If one
calculates the cholesterol excretion normalized for bile salt excretion (because it is a bile salt
driven process), it becomes clear that cholesterol excretion in FVB/N mice is about 4-fold lower
than in C57Bl/6. As a consequence the increased cholesterol excretion in Atp8b1 deficient mice
is much more prominent on a C57Bl/6 background than on a FVB/N background and for this
reason the difference did not reach significance in the present study, whereas it was quite
prominent in our previously reported study using C57Bl/6 mice6.
As discussed above, the biliary phenotype of MF mice was less pronounced than that of
Atp8b1G308V/G308V mice. It was quite striking, however, that hepatic damage in the MF mice was
also less pronounced than in Abcb4-/- mice. Upon administration of a cholate-supplemented diet,
serum ALAT activity was significantly less increased in MF mice than in Abcb4-/- mice (Fig. 5)
and the characteristic induction of fibrosis in Abcb4-/- mice was hardly observed in MF mice. It
must be concluded that the absence of Atp8b1 in some way reduces hepatic damage caused by
the absence of Abcb4. We attribute this to the higher phospholipid excretion in MF mice

Chapter 7
137
compared to Abcb4-/- mice. After bile salt feeding for 7 days the phospholipid over bile salt ratio
(PL/BS) in bile of MF mice is 0.029 while that in Abcb4-/- mice is 0.012. Hence, although
phospholipid excretion is low in both genotypes, it is 2-fold higher in MF mice compared to
Abcb4-/- mice. In mice with low transgenic expression of the human ABCB4 homolog against
Abcb4-/- background, we have previously shown that as little as 8% of normal phospholipid
excretion is sufficient to prevent bile salt-induced hepatic toxicity14. The phospholipid excretion
in MF mice is 11% of that in wild type mice which, according to our old data is indeed sufficient
to prevent hepatic damage caused by cholate-feeding.
In conclusion, our data show that Atp8b1-mediated flipping of PS is particularly important in
the setting of Abcb4-mediated PC flopping. In the absence of PC-flopping the canalicular
membrane appears to be sufficiently rigid to withstand bile salt-mediated extraction without PS-
flopping by Atp8b1.
Acknowledgement
This study was supported by a grant from The Netherlands Organization for Scientific
Research (NWO; program grant 912-02–73).
References
1. Van Meer, G. & Simons, K. The function of tight junctions in maintaining differences in lipid composition
between the apical and the basolateral cell surface domains of MDCK cells. EMBO. J. 1986; 5: 1455-1464.
2. Simons, K. & Vaz, W.L. Model systems, lipid rafts, and cell membranes. Annu. Rev. Biophys. Biomol. Struct.
2004; 33: 269-295.
3. Oude Elferink, R.P., Paulusma, C.C., & Groen, A.K. Hepatocanalicular transport defects: pathophysiologic
mechanisms of rare diseases. Gastroenterology 2006; 130: 908-925.
4. Lykavieris, P. et al. Progressive familial intrahepatic cholestasis type 1 and extrahepatic features: no catch-up of
stature growth, exacerbation of diarrhea, and appearance of liver steatosis after liver transplantation. J. Hepatol.
2003; 39: 447-452.
5. Paulusma, C.C. et al. ATP8B1 requires an accessory protein for endoplasmic reticulum exit and plasma
membrane lipid flippase activity. Hepatology 2008; 47: 268-278.
6. Groen, A. et al. Abcg5/8 independent biliary cholesterol excretion in Atp8b1-deficient mice. Gastroenterology
2008; 134: 2091-2100.
7. Bootsma, A.H. et al. Rapid analysis of conjugated bile acids in plasma using electrospray tandem mass
spectrometry: application for selective screening of peroxisomal disorders. J. Inherit. Metab Dis. 1999; 22: 307-
310.
8. Paulusma, C.C. et al. Atp8b1 deficiency in mice reduces resistance of the canalicular membrane to hydrophobic
bile salts and impairs bile salt transport. Hepatology 2006; 44: 195-204.

Bile formation in mice lacking both Atp8b1 and Abcb4
138
9. James, J., Bosch, K.S., Aronson, D.C., & Houtkooper, J.M. Sirius red histophotometry and spectrophotometry of
sections in the assessment of the collagen content of liver tissue and its application in growing rat liver. Liver
1990; 10: 1-5.
10. van Nieuwkerk, C.M. et al. Effects of Ursodeoxycholate and cholate feeding on liver disease in FVB mice with a
disrupted mdr2 P-glycoprotein gene. Gastroenterology 1996; 111: 165-171.
11. Pawlikowska, L. et al. A mouse genetic model for familial cholestasis caused by ATP8B1 mutations reveals
perturbed bile salt homeostasis but no impairment in bile secretion. Hum. Mol. Genet. 2004; 13: 881-892.
12. Oude Elferink, R.P. et al. Regulation of biliary lipid secretion by mdr2 P-glycoprotein in the mouse. J. Clin.
Invest 1995; 95: 31-38.
13. Smit,J.J. et al. Homozygous disruption of the murine mdr2 P-glycoprotein gene leads to a complete absence of
phospholipid from bile and to liver disease. Cell 1993; 75: 451-462.
14. Smith,A.J. et al. Hepatocyte-specific expression of the human MDR3 P-glycoprotein gene restores the biliary
phosphatidylcholine excretion absent in Mdr2 (-/-) mice. Hepatology 1998; 28: 530-536.





























