Embed Size (px)
Citation preview

ARNO and Apical Endocytosis
1
ARNO THROUGH ITS COILED-COIL DOMAIN REGULATES ENDOCYTOSIS AT THE APICAL SURFACE OF POLARIZED EPITHELIAL CELLS
Miriam Shmuel, Lorraine C. Santy#, Scott Frank+, Dana Avrahami, James E.
Casanova*, and Yoram Altschuler
Department of Pharmacology, School of Pharmacy, Faculty of Medicine, Hebrew University of
Jerusalem, Israel 91120. *Department of Cell Biology, University of Virginia Health Sciences
Center, Charlottesville, VA 22908. #Current address: Dept. of Biochemistry and Molecular Biology,
Pennsylvania State University, +Current address: Children’s Hospital, Boston MA.
Running title: ARNO and Apical Endocytosis
Address correspondence to: Yoram Altschuler, Department of Pharmacology, School of
Pharmacy, Faculty of Medicine, Hebrew University of Jerusalem, Israel 91120. Tel: +972-2-
6757578; fax +972-2-6758927; E mail: [email protected]
ARNO is a guanine nucleotide exchange protein for the ARF family of GTPases. Here we show that in polarized epithel ial cells, ARNO is localized exclusively to the apical plasma membrane, where it regulates endocytosis. Expression of ARNO stimulates apical endocytosis of the polymeric immunoglobulin receptor, and co-expression of ARF6 with ARNO leads to a synergistic stimulation of apical endocytosis. Expression of a dominant negative ARF6 mutant, ARF6-T27N, antagonizes this stimulatory effect. Deletion of the N-terminal coiled-coil (CC) domain of ARNO causes the mutant ARNO to localize to both the apical and basolateral plasma membranes. Expression of the CC domain alone abolishes ARNO-induced apical endocytosis as wel l as colocalization of IgA/receptor complexes with ARNO and clathrin. These results suggest that the CC domain contributes to the specificity of apical localization of ARNO through association with components of the apical plasma membrane. We conclude that ARNO acts together with ARF6 to regulate apical endocytosis. The ADP-ribosylation factors (ARFs) are a family of Ras-like small GTPases that are ubiquitously expressed and involved in membrane trafficking as well as cytoskeleton rearrangement by the recruitment of cytosolic
coat proteins, cytoskeletal proteins and activation of lipid-modifying enzymes (1-4). ARFs can be subdivided into three classes based on sequence homology. Class I is comprised of ARFs 1, 2 and 3, which function primarily in the Golgi and endosomes. Class II contains ARFs 4 and 5, whose functions are poorly understood. ARF6, the sole representative of Class III, has been found to be involved in endocytosis and postendocytic recycling (5-11), exocytosis, specifically with secretory granules in chromaffin cells (12), Glut4 trafficking in adipocytes (13,14) and phagocytosis in macrophages (15,16). Additionally, ARF6 influences the cortical actin cytoskeleton (17-21).
Clathrin-mediated endocytosis (CME) at the apical plasma membrane (PM) of epithelial cells controls the concentration of diverse apical PM transport proteins, which mediate the exchange of materials between the organism and the outside world (22). Apical CME is thus highly regulated by numerous extracellular signals and intracellular signaling pathways. Previously, we reported that in polarized Madin-Darby canine kidney (MDCK) epithelial cells, ARF6 is localized exclusively to the apical PM, where it controls CME (23). When a constitutively active ARF6 mutant (ARF6-Q67L) is expressed in MDCK cells, it is found exclusively at the apical membrane and strongly stimulates CME. A dominant inhibitory mutant ARF6-T27N also stimulates apical endocytosis, though far less potently than ARF6-Q67L.
http://www.jbc.org/cgi/doi/10.1074/jbc.M513723200The latest version is at JBC Papers in Press. Published on February 16, 2006 as Manuscript M513723200
Copyright 2006 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on May 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from

ARNO and Apical Endocytosis
2
Like other small GTPases, ARF6 cycles between an active GTP-bound form and an inactive GDP-bound form. However, ARF6 is unusual in that it remains associated with membranes in its inactive form (24,25). Activation of ARFs relies on a large and diverse family of guanine-nucleotide exchange factors (GEFs), referred to as the Sec7 family (1). Diversity within the Sec7 family derives from differences both within and outside of the 200-aa catalytic domain (26). Among the 15 ARF GEFs in the human genome, the ARNO/cytohesin, EFA6 and BRAG subfamilies contain pleckstrin homology (PH) domains that regulate their association with the plasma membrane and thus are candidates to catalyze ARF6 nucleotide exchange in vivo.
Based on in-vitro studies, ARNO/cytohesins, EFA6A and GEP100/BRAG2 have all been suggested to activate ARF6 (27,28). It is likely that each of these GEFs mediates ARF6 activation at different subcellular sites, or in different cell types. Localization of each GEF within cells is presumably determined by structural elements outside the catalytic domain. Apart from the C-terminal PH domain, members of the ARNO/cytohesin subfamily contain a coiled-coil (CC) domain at their N-terminus that mediates homodimerization, and also participates in other protein/protein interactions. At least three proteins have been identified that interact with the CC domain of ARNO family members, GRASP, CASP and IPCEF (29-32). Both CASP and IPCEF have been shown to co-migrate to the PM together with ARNO in response to EGF stimulation (31,32).
ARNO, ARF6 and ARF1 have been localized to within the apical domain of kidney proximal tubule epithelial cells (33). ARF1 was localized to both the Golgi and apical domain, whereas ARF6 was found exclusively apically in these cells. In addition, both ARNO and ARF6 in proximal tubule cells appear to translocate from the cytosol to endosomes in a manner that is dependent on endosome acidification. In polarized MDCK cells, EFA6 localizes to the apical domain and participates in tight-junction formation by regulating the association of the tight junction with the apical actin ring (34). Recently, within the subapical domain of MDCK as well as in renal proximal tubules, EFA6 was shown to associate with TWIK1, which belongs to the family of K+ channels. This association was stabilized in BHK cells by
expression of an ARF6-GDP mutant (35). Given the diversity of localizations and
actions reported for ARF6, ARNO and EFA6, it is vital to determine which GEF is responsible for activation of ARF6 at specific sites in individual cell types. In particular, the unique localization of ARF6 to the apical PM and its function in the regulation of apical CME make the question of how ARF6 is activated a central issue in epithelial cell biology and physiology. Here we report that in polarized epithelial cells, ARNO is localized, through its CC domain, exclusively to the apical PM, where it acts to regulate apical endocytosis by activating ARF6.
Experimental Procedures
Most of the methods are described in
detail in our recent papers on ARF6 and ARNO (23,36). Antibodies - The polyclonal rabbit antiserum against human ARNO has been described previously (36). For tissue immunolabeling, this antibody was affinity-purified using a GST- ARNO fusion protein lacking the CC domain of ARNO (residues 2-60). This was necessary as antibodies against the CC epitope cross-reacted with vimentin in both tissues and fixed cells. 9E10 antibody was purchased from Covance (Berkeley CA), flag antibody was from Sigma, sheep anti-human secretory IgA was from Cappel Laboratories (Cochranville, PA), X22 antibody against clathrin heavy chain and TD1 against clathrin light chain were kindly provided by Frances Brodsky (UCSF, San Francisco, CA). Secondary antibodies, anti-mouse and anti-rabbit conjugated to either Alexa 488 or Alexa 594 (minimal reactivity) were from Molecular Probes (Eugene, OR). Localization of endogenous ARNO - Tissues from Sprague-Dawley rats were fixed in paraformaldehyde-lysine-periodate by intravascular perfusion. They were then coarsely chopped with a razor blade, further fixed overnight at 4°C then cryoprotected by incubation in 30% sucrose/PBS for 3 h. Tissue slices were then snap-frozen in Tissue-Tec embedding medium and 5-µ cryosections prepared using a Reichert cryostat. For immunolabeling, tissues were preblocked in PBS containing 10% normal goat serum and 0.25% saponin, then incubated with primary antibody (10 µg/ml) in blocking buffer for 30 min at room
by guest on May 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from

ARNO and Apical Endocytosis
3
temperature. After washing, samples were incubated for 30 min with Cy2-conjugated donkey anti-rabbit antibody (Jackson Immunoresearch, West Grove, PA). Caco-2 monolayers were fixed in 4% paraformaldehyde, then blocked and labeled as described for tissues. Brush-border (BB) preparation - Confluent monolayers of Caco-2 cells were scraped from two 10-cm plates in a volume of 2 ml/plate Tris-mannitol buffer (2 mM Tris, pH 7.1, 50 mM mannitol, 1 mM PMSF). The suspended cells were then sonicated using a Misonix probe sonicator at a setting of 5 for 30 s. CaCl2 was added to a final concentration of 18 mM and the sample incubated on ice for 10 min. Non-BB membranes were then pelleted by centrifugation for 10 min at 950 x g, and BB membranes recovered from the supernatant by centrifugation for 40 min at 150,000 x g, 4°C. Enzymatic assays were conducted on each fraction as described previously (35). For Na, K-ATPase, activity was quantitated as ouabain-inhibitable phosphate release (37). Exogenous expression of ARF6 and ARNO - Recombinant adenoviruses expressing ARF6, ARNO and their mutants were prepared and used to infect filter-grown MDCK cells as previously described (23,36). The ARNO-∆coil mutation was constructed by deleting residues 2-53 in frame. The CC domain was constructed by PCR amplification of a DNA sequence corresponding to the flag tag followed by 62 N-terminal amino acids of ARNO. The level of expression of the exogenous proteins was under the control of the tetracycline-repressible system. We estimate that the level of expression of exogenous ARNO-WT and ARNO-∆coil was approximately 10- to 20-fold higher than that of endogenous ARNO. A principal source of uncertainty in this measurement is that we do not know precisely how well our antibody against ARNO cross-reacts with the canine ARNO endogenous to MDCK cells. However, we used this antibody to confirm that the expression levels of exogenous ARNO and the mutants used in this report were all reasonably constant, within ~20%. Immunofluorescence of MDCK cells - These procedures were performed as described previously (23). As a convenient and reproducible marker for the apical PM, we used either MDCK cells transfected with Concentrative Nucleoside Transporter 1 fused to
GFP (CNT1-GFP), which we have recently shown to be localized to the apical PM (38), or cells labeled at 4oC with IgA by incubating the cells with cold IgA for 60 min at 4oC, followed by extensive washes with PBS containing 1 mM MgCl2 and 1 mM CaCl2 and fixation. Images were taken using a BioRad confocal or NIKON TE-2000S (Nikon, Melville, NY) inverted fluorescence microscope with a plan Apo 60X objective lens (Nikon), equipped with a Z stepper and a Hammamatzo CCD ORCAII camera (Hammamtzo, Tucson, AZ). Images were all deconvolved with SimplePCI software (Improvision, Coventry, UK). All images were compiled using Adobe Photoshop, and/or Canvas software (ACD Systems International Inc., Saanichton, British Columbia, Canada), and are representative of the original data.
Assays of membrane traffic - Assays of apical and basolateral endocytosis, basolateral to apical transcytosis, and apical recycling, utilized radio-iodinated dimeric IgA, bound to the rabbit polymeric immunoglobulin receptor (pIgR), which had been stably transfected into the cells. Methods for these assays have been described previously (39). Biosynthetic delivery of proteins to the PM was measured as described (40). Fractionation of MDCK cells - Subcellular fractionation was modified from a previously described protocol to maintain actin in polymerized form (41). Briefly, MDCK cells were grown in 10-cm dishes for 3 days, infected with appropriate recombinant adenoviruses and incubated for 18 h. Cells were rinsed with cold PBS (containing 1 mM CaCl2 and 1 mM MgCl2), scraped and resuspended in ice-cold homogenization buffer (100 mM NaCl, 3 mM imidazole, 300 mM sucrose, 20 mM Tris, 1 mM DTT protease inhibitor cocktail pH 8.0). Cells were homogenized by 80 to 90 passages through a 27G syringe. Subsequently, the homogenate was centrifuged at 1000 x g, 4oC for 5 min to remove nuclei and large debris. The supernatant was then centrifuged at 10,000 x g, 4oC for 30 min to obtain a crude PM fraction. The resulting supernatant was centrifuged at 165,000 x g, 4oC for 2 h to obtain a light membrane fraction (endoplasmic reticulum and Golgi) and the supernatant (Cyt) containing the cell cytosol. After each centrifugation, the resulting pellet was rinsed briefly with ice-cold PBS before subsequent fractionations to avoid possible crossover contamination.
by guest on May 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from

ARNO and Apical Endocytosis
4
Results ARNO is localized to the apical surface of epithelial cells
We examined the subcellular localization of endogenous ARNO in both rat epithelial sections and the Caco-2 epithelial cell line, as well as by recombinant expression of ARNO in MDCK cells. In rat epithelial sections, ARNO was found primarily at the apical PM, as shown by immunofluorescent staining of sections of rat small intestine (Fig. 1A), kidney proximal tubule (Fig. 1B), submandibular gland (Fig. 1C) and parotid gland (Fig. 1D). This staining is characteristic of the apical PM facing the inner lumen of the tubules. Fig. 3A shows co-staining of endogenous ARNO with the apical marker Ezrin in cultured Caco-2 human intestinal cells. This latter figure illustrates the fine punctate pattern characteristic of apical microvilli in a section taken through the apical PM.
To confirm the apical localization of endogenous ARNO in epithelial cells, we took a biochemical approach in which we isolated a BB membrane fraction from the Caco-2 intestinal cell line. As shown in Fig. 2A, this BB fraction was significantly enriched in the apical marker protein alkaline phosphatase, relative to non-BB membranes, indicating that it is enriched in apical proteins. Immunoblotting of the different fractions revealed that most of the endogenous ARNO is present in the cytosolic fraction, as described previously (36), but the membrane-bound ARNO is detected only in BB membrane fractions, and not in the non-BB membrane fractions. Endogenous ARF6 is also enriched in the BB membrane, confirming our previous results that exogenously expressed ARF6 is localized to the apical PM (23). Ezrin, a well-characterized apical protein, also shows a distribution similar to that of ARNO, confirming correct separation of the membrane into its BB (apical) and basolateral parts (Fig. 2B).
Next, we determined the localization of recombinant ARNO in MDCK cells. Since our polyclonal antibody fails to detect endogenous ARNO by immunofluorescence in MDCK cells, we expressed exogenous myc-tagged human ARNO (36) in MDCK cells using a recombinant adenovirus system that incorporates a tetracycline-regulated promoter.
In Fig. 3, representative X-Y confocal sections of apical and basolateral focal planes demonstrate that ARNO localizes exclusively to the apical PM and is not detected below the tight junctions on the lateral PM (Fig. 3B). To show that ARNO was precisely at the apical PM, and not in the cytoplasm immediately below it, we performed colocalization with an integral apical PM protein. As an apical marker, we expressed CNT1-GFP, which is an integral protein of the apical PM (38). This gave results identical to colocalization with the apical marker GP135 (localized by indirect immunofluorescence), but was more reproducible and convenient. As shown in Fig. 3C, ARNO and CNT1-GFP colocalized precisely. Moreover, we did not detect either endogenous or exogenous ARNO in a perinuclear or supranuclear cytoplasmic location, characteristic of the Golgi in MDCK (Figs. 1, 3).
ARNO in concert with ARF6 induces CME at the apical PM
To examine the function of ARNO in CME at the apical PM, we performed a biochemical endocytosis assay tracing the rate of IgA endocytosis by the polymeric immunoglobulin receptor (pIgR) which was stably expressed in these cells. In the first experiment, we measured the effect of exogenous expression of ARNO on apical and basolateral CME. For this purpose, MDCK cells were infected with adenovirus encoding wild-type ARNO (Fig. 4A) and incubated for 18 h to induce expression. As shown in Fig. 4B, expression of wild-type ARNO induced apical endocytosis of IgA approximately sixfold, but had no significant effect on basolateral endocytosis of IgA (Fig. 4C). We observed no effect on the delivery of newly made pIgR to either apical or basolateral PM, transcytosis of IgA by pIgR from the basolateral to apical PM, or recycling of apical or basolateral internalized IgA back to the PM (data not shown).
We have previously reported that ARF6 localizes to the apical PM and regulates CME there. Although overexpression of dominant active ARF6-Q67L, and to a lesser extent of dominant negative ARF6-T27N, stimulates apical CME, the wild-type form of ARF6 did not induce any stimulation of apical CME (23). We hypothesized that in epithelia, ARNO may be the exchange factor for ARF6.
by guest on May 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from

ARNO and Apical Endocytosis
5
To test this hypothesis, we co-expressed a submaximal level of ARNO that induced approximately half the effect on apical endocytosis (~15% of bound IgA endocytosed in a 5-min warm-up) with overexpressed ARF6-WT. Co-expression of these two proteins produced synergistic stimulation of apical endocytosis (Fig. 4D).
Typically, dominant negative GTPases function by sequestering endogenous exchange factors in a complex with the mutant GTPase. As shown in Fig. 4E, we found that co-expression of ARF6-T27N antagonizes the stimulation of endocytosis by ARNO. Taken together, the biochemical and morphological data provide evidence for localization of ARNO to the apical PM where it participates in apical endocytosis, and are consistent with a model in which ARNO activates ARF6, which in turn stimulates apical CME.
Functional role for the CC and PH domains of ARNO
Although much progress has been made in understanding how integral PM proteins are correctly localized to the apical or basolateral PM (22), far less is known about the equally important question of how peripheral PM proteins, such as ARNO, achieve a polarized localization in epithelial or other polarized cells (42,43).
As shown in Fig. 4A, ARNO contains a catalytic Sec7 homology domain, flanked by an N-terminal CC domain known to participate in homodimerization as well as protein-protein interactions, and a C-terminal PH domain that regulates binding to phosphatidylinositol-containing membranes. Therefore, we decided to test the role of both N-terminal and C-terminal domains in targeting ARNO to the apical PM in MDCK cells. Cells expressing a mutant ARNO lacking the PH domain (Fig. 4A) showed exclusive apical localization (Fig. 5A) with no staining on the basolateral side, identical to the apical localization of wild-type (WT) ARNO (Fig. 3A). Despite the exclusive apical localization of the ARNO-∆PH mutant, its expression provoked significantly reduced stimulation of apical endocytosis (Fig. 4B) compared to the effect observed with ARNO-WT, suggesting that the PH domain is important in regulating endocytic function. In keeping with its apical localization, no effect was observed on basolateral endocytosis (Fig.
4C). In parallel, we expressed a mutant ARNO lacking the 52-aa CC domain (ARNO-∆coil) (Fig. 4A). In contrast to the ∆PH mutant, ARNO-∆coil lost its exclusive apical localization and was observed on both the apical and basolateral PM (Fig. 5A). Utilizing the adenovirus delivery system together with the tetracycline-regulated expression system, the level of expression of the mutant ARNO was comparable to that of the ARNO-WT in other experiments, indicating that the loss of exclusive apical localization is not due to overexpression. We then wanted to confirm whether ARNO-∆coil is localized to the PM, the endosomal compartment or both. For that purpose, we colocalized ARNO-∆coil with IgA bound to the pIgR on the basolateral PM at 4°C to ensure that we differentiate between the PM and early endosomal localization. As shown in Fig. 5B, ARNO-∆coil is localized exclusively to the basolateral PM and does not appear in endosomal membranes, indicating that ARNO-∆coil was truly at the basolateral PM and not associated with sub-PM vesicles or organelles (Fig. 5B). We also subjected the samples to mild digitonin treatment to extract all cytoplasmic ARNO-∆coil. We found that IgA decorates the PM in a punctate fashion. In contrast, the ARNO-∆coil appears evenly distributed, indicating that the CC domain not only causes apical targeting but also specific targeting within the PM. We did not detect ARNO-∆coil associated with intracellular structures, such as endosomes, Golgi or the endoplasmic reticulum (data not shown). In addition, mutations within the Sec7 domain that render ARNO catalytically inactive (ARNO-E156K) or mutations that abolish the function of the PH domain (ARNO-R280D), do not alter the apical localization of the protein (Fig. 3E). These results strongly suggest that the remaining domain, i.e. the CC domain, mediates apical PM localization.
To better characterize the role of the CC domain, we fractionated MDCK cells into heavy membranes (PM), light membranes (endoplasmic reticulum, Golgi and endosomes) and cytosol (Fig. 6B). We quantified the ratio between the PM and cytosol fractions and found that the ARNO-∆coil mutant appears four- to fivefold more prominently in the PM fraction compared to ARNO-WT. This ratio is similar to that between the apical and basolateral membranes, indicating that the PH domain does not
by guest on May 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from

ARNO and Apical Endocytosis
6
distinguish between the two membranes, but rather recognizes a membrane motif that appears in both (44-48).
To establish the role of the ARNO CC domain in targeting ARNO to the apical PM, we constructed a mutant harboring only the CC domain preceded by a flag tag. As shown in Fig. 5C, expression of this construct creates a pool of CC peptides which are distributed in a punctate staining pattern within the apical domain in a region that is just above the tight junction (Fig. 5C).
We hypothesized that pIgR associates either directly or indirectly with ARNO through the latter's CC domain to recruit the endocytic machinery. Therefore, ARNO would be expected to localize transiently to the PM in association with the IgA-pIgR complex. To detect this transient state, we co-expressed a dynamin I K44A mutant that arrests endocytosis at a late stage and thus causes accumulation of clathrin-coated pits and their cargo on the PM. In the presence of this dynamin mutant, IgA was observed decorating the apical PM (Fig. 5D). When co-expressed with ARNO-WT, a high degree of colocalization was observed between ARNO and IgA, as well as a significant increase in the colocalization of IgA with clathrin (second row of panels). In contrast, the isolated CC domain exhibits significant colocalization with IgA, but little colocalization with clathrin (compare left and right panels). This result suggests that ARNO associates through its CC domain with the cargo, i.e. IgA-pIgR, and not with the endocytic machinery. To confirm that the effects of ARNO on endocytosis require the CC domain, we performed an endocytosis assay (Fig. 6A). In this experiment, expression of the CC domain alone had no effect on apical endocytosis, raising the possibility that the basal level of endocytosis may be ARNO-independent. However, when co-expressed with ARNO-WT, the CC domain almost completely abolished the ARNO-induced stimulation of apical pIgR-IgA complex endocytosis. This inhibitory effect suggests that the CC domain can compete with ARNO for binding to cargo, or to components of the endocytic machinery, or both. We did not detect CC domain at either the basolateral PM or in other internal vesicles or organelles within the basolateral domain. The basolateral localization of ARNO-∆coil raises the question of whether ARNO is
responsible for the apical localization of ARF6. In cells that express both ARNO-∆coil and ARF6-WT, apical localization of ARF6 was not disrupted, and the rate of basolateral endocytosis of IgA was unaffected (data not shown). Therefore, apical ARF6 localization is independent of the localization of the exchange factor ARNO. Taken together, we find that ARF6 and ARNO independently localize to the apical PM but operate in concert to recruit the endocytic machinery to the IgA-pIgR complex. More precisely, ARNO localization to the apical domain, and specifically to the IgA-pIgR complex, is dependent on its CC domain.
Discussion To establish the epithelial apical localization of the exchange factor ARNO, we looked at endogenous ARNO in both rat epithelial tissues and human intestinal epithelial cells (Caco-2), as well as its recombinantly expressed counterpart in canine kidney (MDCK) cells. In all cases, ARNO was localized to the apical domain and apical PM. Specifically, both the nucleoside transporter CNT1, which localizes exclusively to the apical PM, and Ezrin, a member of the ezrin-radixin-moesin family, which is a conspicuous component of the apical F-actin-based scaffold
in simple epithelial cells, colocalized extensively with ARNO. These results agree with our recently published ones linking the actin cytoskeleton to the apical CME machinery, as well as with published results on apical localization of ARNO (33,49-51). An in-depth structure-function study revealed that deletion of the PH domain or a mutation that renders it inactive does not alter the apical localization of ARNO. In addition, a mutation that also abolishes the catalytic activity of ARNO did not interfere with its apical localization (Fig. 3E and data not shown). In contrast, a mutant ARNO lacking the CC domain localized to both the apical and basolateral PMs. These results indicate that the PH domain targets nonspecifically to the PM and not to other membranes in the cell, and most likely to PM domains containing PIP2- or PIP3-modified membranes (52,53). Previous studies have attributed several possible functions to the CC domain. One well-characterized function is its mediation of ARNO homodimerization (54,55).
by guest on May 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from

ARNO and Apical Endocytosis
7
Additionally, the CC domain of a different ARNO family member has been shown to interact with Munc13-1, and has therefore been proposed to be involved in targeting the exchange factor to the presynaptic active zone in neurons (57). ARNO has also been shown to interact with other molecules such as CASK or IPCEF (31,32), which appear to act as adaptors, and could serve to link ARNO to endocytic cargo. GRP1, which is closely related to ARNO, binds through its CC domain to a similar adaptor protein, Tamalin/GRASP, which in turn binds metabotropic glutamate receptors. In addition, ARNO has been shown to directly bind ß-arrestin, and to promote the internalization of G-protein-coupled receptors (58). It therefore seems plausible that ARNO could be linked to pIgR or other cargo by a similar bridging mechanism. Downstream activation of ARF6 would then lead to recruitment of the clathrin endocytic machinery, presumably through the activation of PIP5 kinase (Fig. 7). We show that ARNO expression stimulates apical endocytosis exclusively, without altering other membrane transport events or specifically altering basolateral endocytosis. Even mislocalization to the basolateral PM, which occurs upon deletion of the CC domain, did not alter endocytosis at the basolateral surface. The failure of ARNO-∆coil to recruit ARF6 to the basolateral domain and influence endocytosis there may indicate that the apical ARF6 targeting mechanism is independent of that of ARNO. Given the paucity of information on the mechanisms of localization of peripheral PM proteins, our findings on the effects of the CC domain represent a significant advance. We have previously shown that ARF6 is localized to the apical PM and that it stimulates endocytosis which is actin-clathrin- and dynamin I-dependent (23,49). In the present study, we link this finding to the GEF ARNO. We show that ARF6-WT, when expressed alone, does not stimulate apical endocytosis. When co-expressed with ARNO-
WT, a synergistic stimulation of apical endocytosis takes place. Conversely, when ARNO-WT is co-expressed with ARF6-T27N, an inhibitory effect on ARNO-mediated apical endocytosis occurs. Taking the morphological and functional results on endocytosis together, and given that endogenous ARNO is localized to the apical PM in a variety of epithelial tissues, it is likely that ARNO is indeed an authentic GEF for ARF6 in epithelial cells in vivo. However, this does not rule out the possibility that other ARNO family members also act on ARF6 at this PM. In particular, GRP-1/ARNO3, which like ARNO is ubiquitously expressed, may serve a similar function. EFA6 is a more distantly related member of the ARNO family, which shows a preference for ARF6 in vitro (27). EFA6 in MDCK cells regulates tight-junction assembly. The exogenous protein appears to be primarily apical, but is concentrated in functional regions. Upon calcium-dependent cell adhesion, EFA6 is recruited to a Triton-insoluble fraction and is mainly concentrated in the forming tight junction. This suggests that ARF6 functions in tight-junction assembly as well as apical endocytosis (34). In conclusion, our data indicate that ARF6 and its activator, ARNO, are both localized to the apical PM of polarized epithelial cells, where they function together to regulate apical endocytosis. Although ARNO concentrates with endocytic cargo (pIgR) at clathrin-coated structures, it remains to be determined whether it associates with cargo directly or through an associated adaptor molecule. In any case, association of ARNO with clathrin-coated pits requires the N-terminal CC domain. The apical PM of epithelial cells is a specialized cellular structure with unique vital and fundamental physiological functions that are regulated in part by endocytosis. Thus evidence is provided for ARNO and ARF6 as principal players in epithelial physiology.
.
REFERENCES
1. Chavrier, P., and Goud, B. (1999) Curr. Opin. Cell Biol. 11(4), 466-475
by guest on May 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from

ARNO and Apical Endocytosis
8
2. Donaldson, J. G., and Jackson, C. L. (2000) Curr Opin Cell Biol 12(4), 475-482 3. Austin, C., Boehm, M., and Tooze, S. A. (2002) Biochemistry 41(14), 4669-4677 4. Takatsu, H., Yoshino, K., Toda, K., and Nakayama, K. (2002) Biochem J 365(Pt 2),
369-378 5. Claing, A. (2004) Biochem Cell Biol 82(6), 610-617 6. Donaldson, J. G. (2003) J Biol Chem 278(43), 41573-41576 7. Cavenaugh, M. M., Whitney, J. A., Carroll, K., Zhang, C.-J., Boman, A. L.,
Rosenwald, A. G., Mellman, I., and Kahn, R. A. (1996) J. Biol. Chem. 271, 21767-21774
8. D'Souza-Schorey, C., Li, G., Colombo, M. I., and Stahl, P. D. (1995) Science 267, 1175-1178
9. D'Souza-Schorey, C., van Donselaar, E., Hsu, V. W., Yang, C., Stahl, P. D., and Peters, P. J. (1998) J. Cell Biol. 140, 603-616
10. Peters, P. J., Hsu, V. W., Ooi, C. E., Finazzi, D., Teal, S. B., Oorschot, V., Donaldson, J. G., and Klausner, R. D. (1995) J. Cell Biol. 128, 1003-1017
11. Radhakrishna, H., and Donaldson, J. G. (1997) J. Cell Biol. 139, 49-61 12. Vitale, N., Chasserot-Golaz, S., and Bader, M. F. (2002) Ann N Y Acad Sci 971, 193-
200 13. Yang, C. Z., and Mueckler, M. (1999) J Biol Chem 274(36), 25297-25300 14. Park, J. G., Bose, A., Leszyk, J., and Czech, M. P. (2001) J Biol Chem 276(51),
47751-47754 15. Zhang, Q., Cox, D., Tseng, C. C., Donaldson, J. G., and Greenberg, S. (1998) J. Biol.
Chem. 273, 19977-19981 16. Niedergang, F., Colucci-Guyon, E., Dubois, T., Raposo, G., and Chavrier, P. (2003) J
Cell Biol 161(6), 1143-1150 17. Boshans, R. L., Szanto, S., van Aelst, L., and D'Souza-Schorey, C. (2000) Mol. Cell
Biol. 20(10), 3685-3694 18. D'Souza-Schorey, C., Boshans, R. L., McDonough, M., Stahl, P. D., and Van Aelst,
L. (1997) EMBO J. 16, 5445-5454 19. Radhakrishna, H., Al-Awar, O., Khachikian, Z., and Donaldson, J. G. (1999) J. Cell
Sci. 112(Pt 6), 855-866 20. Radhakrishna, H., Klausner, R. D., and Donaldson, J. G. (1996) J. Cell Biol. 134,
935-947 21. Song, J., Khachikian, Z., Radhakrishna, H., and Donaldson, J. G. (1998) J. Cell Sci.
111, 2257-2267 22. Mostov, K. E., Verges, M., and Altschuler, Y. (2000) Curr Opin Cell Biol 12(4), 483-
490 23. Altschuler, Y., Liu, S.-H., Katz, L., Tang, K., Hardy, S., Brodsky, F., and Mostov, K.
(1999) J. Cell Biol. 147, 7-12 24. Gaschet, J., and Hsu, V. W. (1999) J Biol Chem 274(28), 20040-20045 25. Macia, E., Luton, F., Partisani, M., Cherfils, J., Chardin, P., and Franco, M. (2004) J
Cell Sci 117(Pt 11), 2389-2398 26. Jackson, C. L., and Casanova, J. E. (2000) Trends Cell Biol. 10, 60-67 27. Franco, M., Peters, P. J., Boretto, J., van Donselaar, E., Neri, A., D'Souza-Schorey,
C., and Chavrier, P. (1999) EMBO J. 18(6), 1480-1491 28. Someya, A., Sata, M., Takeda, K., Pacheco-Rodriguez, G., Ferrans, V. J., Moss, J.,
and Vaughan, M. (2001) Proc Natl Acad Sci U S A 98(5), 2413-2418 29. Nevrivy, D. J., Peterson, V. J., Avram, D., Ishmael, J. E., Hansen, S. G., Dowell, P.,
Hruby, D. E., Dawson, M. I., and Leid, M. (2000) J Biol Chem 275(22), 16827-16836.
by guest on May 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from

ARNO and Apical Endocytosis
9
30. Neeb, A., Koch, H., Schurmann, A., and Brose, N. (1999) Eur J Cell Biol 78(8), 533-538
31. Mansour, M., Lee, S. Y., and Pohajdak, B. (2002) J Biol Chem 277(35), 32302-32309 32. Venkateswarlu, K. (2003) J Biol Chem 278(44), 43460-43469 33. Maranda, B., Brown, D., Bourgoin, S., Casanova, J. E., Vinay, P., Ausiello, D. A.,
and Marshansky, V. (2001) J Biol Chem 276(21), 18540-18550 34. Luton, F., Klein, S., Chauvin, J. P., Le Bivic, A., Bourgoin, S., Franco, M., and
Chardin, P. (2004) Mol Biol Cell 15(3), 1134-1145 35. Decressac, S., Franco, M., Bendahhou, S., Warth, R., Knauer, S., Barhanin, J.,
Lazdunski, M., and Lesage, F. (2004) EMBO Rep 5(12), 1171-1175 36. Frank, S., Upender, S., Hansen, S. H., and Casanova, J. E. (1998) J. Biol. Chem. 273,
23-27 37. Coppi, M. V., and Guidotti, G. (1997) Arch. Biochem. Biophys. 346(2), 312-321 38. Mangravite, L. M., Lipschutz, J. H., Mostov, K. E., and Giacomini, K. M. (2001) Am
J Physiol Renal Physiol 280(5), F879-885 39. Breitfeld, P., Casanova, J. E., Harris, J. M., Simister, N. E., and Mostov, K. E. (1989)
Meth. Cell Biol. 32, 329-337 40. Reich, V., Mostov, K., and Aroeti, B. (1996) J. Cell Sci. 109, 2133-2139 41. Huttner, W. B., Schiebler, W., Greengard, P., and De Camilli, P. (1983) J Cell Biol
96(5), 1374-1388 42. Chuang, J. Z., and Sung, C. H. (1998) J. Cell Biol. 143, 1245-1256 43. Lecat, S., Verkade, P., Thiele, C., Fiedler, K., Simons, K., and Lafont, F. (2000) J
Cell Sci 113 (Pt 14), 2607-2618 44. Santy, L. C., Frank, S. R., Hatfield, J. C., and Casanova, J. E. (1999) Curr. Biol.
9(20), 1173-1176 45. Paris, S., Beraud-Dufour, S., Robineau, S., Bigay, J., Antonny, B., Chabre, M., and
Chardin, P. (1997) J. Biol. Chem. 272(35), 22221-22226 46. Langille, S. E., Patki, V., Klarlund, J. K., Buxton, J. M., Holik, J. J., Chawla, A.,
Corvera, S., and Czech, M. P. (1999) J. Biol. Chem. 274(38), 27099-27104 47. Klarlund, J. K., Guilherme, A., Holik, J. J., Virbasius, J. V., Chawla, A., and Czech,
M. P. (1997) Science 275(5308), 1927-1930 48. Hernandez-Deviez, D. J., Roth, M. G., Casanova, J. E., and Wilson, J. M. (2004) Mol
Biol Cell 15(1), 111-120 49. Hyman, T., Shmuel, M., and Altschuler, Y. (2006) Mol Biol Cell 17(1), 427-437 50. Louvet-Vallee, S. (2000) Biol Cell 92(5), 305-316 51. Polesello, C., and Payre, F. (2004) Trends Cell Biol 14(6), 294-302 52. Macia, E., Paris, S., and Chabre, M. (2000) Biochemistry 39(19), 5893-5901 53. Venkateswarlu, K., Oatey, P. B., Tavare, J. M., and Cullen, P. J. (1998) Curr Biol
8(8), 463-466 54. Chardin, P., Paris, S., Antonny, B., Robineau, S., Beraud-Dufour, S., Jackson, C. L.,
and Chabre, M. (1996) Nature 384(6608), 481-484 55. Cherfils, J., Menetrey, J., Mathieu, M., Le Bras, G., Robineau, S., Beraud-Dufour, S.,
Antonny, B., and Chardin, P. (1998) Nature 392(6671), 101-105 56. Lee, S. Y., and Pohajdak, B. (2000) J Cell Sci 113 (Pt 11), 1883-1889 57. Neeb, A., Koch, H., Schurmann, A., and Brose, N. (1999) Eur J Cell Biol 78(8), 533-
538. 58. Claing, A., Chen, W., Miller, W. E., Vitale, N., Moss, J., Premont, R. T., and
Lefkowitz, R. J. (2001) J Biol Chem 276(45), 42509-42513
by guest on May 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from

ARNO and Apical Endocytosis
10
The abbreviations used are: ARF, ADP-ribosylation factor; BB, brush border; CC, coiled-coil; CME, clathrin-mediated endocytosis; CNT1, Concentrative Nucleoside Transporter 1; MDCK, Madin-Darby canine kidney; PH, pleckstrin homology; pIgR, polymeric immunoglobulin receptor; PM, plasma membrane.
FIGURE LEGENDS
Fig. 1. Localization of endogenous ARNO in epithel ial tissues . Frozen 5-µ sections of rat
proximal small intestine (A), kidney proximal tubules (B), submandibular gland (C), or parotid
gland (D) were labeled with affinity-purified ARNO antibody. All sections reveal epithelial apical
localization of endogenous ARNO.
Fig. 2. ARF6 and ARNO are enriched in brush-border (BB) membranes from Caco-2
cells.
A. BB membranes were prepared as described in Methods. Aliquots of total cell lysates, non-BB
membranes, BB membranes and cytosol were analyzed for the presence of the endoplasmic
reticulum marker α-glucosidase II, the Golgi marker α-mannosidase, the apical PM marker
alkaline phosphatase, or the basolateral PM marker Na, K-ATPase. Note that BB membranes are
significantly enriched in alkaline phosphatase activity. B. Equivalent amounts of protein from
each fraction were immunoblotted with antibodies to Ezrin, ARF6 and ARNO. Fractionation
reveals that both ARF6 and ARNO are enriched in the BB fraction, indicating apical PM
localization.
Fig. 3. ARNO is found at the apical surface of polarized epithel ial MDCK and Caco-2
cells.
A. A monolayer of Caco-2 cells was fixed in 4% paraformaldehyde and stained with affinity-
purified anti-ARNO antibody (green) and anti-Ezrin, a brush-border marker. ARNO is shown to
completely localize with the apical marker, Ezrin. B-E. MDCK tet-off cells were uninfected
(control), or infected with ARNO-WT (B-D), or ARNO-E157A or ARNO-R280D (E)
adenoviruses, incubated for 18 h for expression and subsequently processed for
immunofluorescence. B. Series of X-Y confocal sections of cells stained for myc-tagged
exogenous ARNO (green) and anti-tight junction marker ZO-1 (red). The distance (in microns)
of the section above or below the center of the ZO-1 staining is indicated on the top left. C-E.
Section through the apical domain showing colocalization of the apical PM marker CNT1-GFP
(green) and either ARNO-WT (C, D) or ARNO mutants (E) (red). D. X-Z confocal section
showing colocalization between CNT1-GFP (green) and ARNO (red). E. Apical section of MDCK
tet-off cells expressing the catalytically inactive mutant ARNO-E157A and PH-domain-inactive
mutant ARNO-R280D (red) stained with the apical marker CNT1-GFP (green). Note that ARNO
is localized exclusively to the apical PM, independent of the activities of the Sec7 or PH
by guest on May 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from

ARNO and Apical Endocytosis
11
domains. Bar 2 µM.
Fig. 4. Effects of ARNO and ARF6 on apical endocytosis.
A. Schematic structural diagram of ARNO as well as the deletion mutants. B-C. MDCK tet-off
cells were grown for 3 days on Corning transwells. Control cells (uninfected) and cells expressing
ARNO-WT, ARNO-∆coil or ARNO-∆PH for 18 h were used in a 5-min assay of radio-iodinated
IgA endocytosis from either the apical (B) or basolateral (C) surface. ARNO-WT strongly stimulates
apical endocytosis. Mutants show reduced levels of stimulated apical endocytosis of IgA (B); no
effect is observed on endocytosis of IgA from the basolateral PM (C). D-E. MDCK tet-off cells were
grown for 3 days on Corning transwells. Control cells (uninfected) and cells expressing ARNO-WT,
ARF6-WT or ARF6-T27N, or co-expressing ARNO-WT with ARF6-WT (D) or ARF6-T27N
(E) for 18 h were used in a 5-min assay of radio-iodinated IgA endocytosis from the apical surface.
The level of ARNO expression was approximately 50% of that used in B and C to allow
monitoring of both stimulatory and inhibitory effects. Cells co-expressing ARNO-WT and ARF6-
WT show a synergistic stimulatory effect on apical endocytosis (D). Note that cells co-expressing
ARNO-WT with dominant negative ARF6-T27N show no effect on apical endocytosis, indicating
that ARF6-T27N abolished ARNO's stimulatory effect on endocytosis (E).
Fig. 5. The CC domain of ARNO regulates apical targeting.
MDCK tet-off cells were infected with recombinant adenoviruses to express myc-tagged ARNO-
∆coil and ARNO-∆PH or flag-tagged ARNO-CC domain, as indicated, for 18 h and subsequently
processed for immunofluorescence. A. X-Y confocal sections through apical and basolateral
domains show that while ARNO-∆PH maintained apical PM localization, the ARNO-∆coil lost
exclusive apical localization and in addition stained the basolateral domain. B. Following 18-h
expression of ARNO-∆coil (red), cells were cooled down and IgA (green) was bound to pIgR at the
basolateral surface for 60 min at 4oC. Unbound IgA was washed away, and one set was treated
with digitonin (bottom panels) to remove cytoplasmic ARNO-∆coil. Cells were fixed and
processed for immunofluorescence. ARNO-∆coil is shown to decorate the basolateral PM in a
punctate fashion. C. MDCK tet-off cells were infected with flag-tagged ARNO-CC domain (red)
and co-stained with either the tight-junction marker ZO-1 (green) or apical PM-bound IgA
(green), as indicated. The ARNO-CC domain is shown to localize primarily to the apical domain,
not on the apical PM. D. MDCK tet-off cells were single- or double-infected to express dominant
negative dynamin I K44A alone or with either myc-tagged ARNO-WT or flag-tagged ARNO-CC
domain as indicated, for 18 h of expression, and subsequently processed for immunofluorescence.
Expression of dynamin I K44A inhibits endocytosis at a late stage and results in trapping the
endocytic machinery in the clathrin-coated pit. Left panels show co-staining with IgA bound to
the apical PM (green) and either ARNO-WT or ARNO-CC domain (red). Right panels show co-
staining with IgA bound to the apical PM (green) and clathrin (red). Left panels show that when
by guest on May 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from

ARNO and Apical Endocytosis
12
endocytosis is inhibited by dominant negative dynamin, the ARNO-WT or ARNO-CC domain
colocalizes strongly with IgA, indicative of transient colocalization between ARNO through its
CC domain and the IgA cargo. Right panels show that ARNO-WT expression increases
colocalization of the cargo (IgA) with clathrin. Expression of the CC domain alone reduces cargo
(IgA) colocalization with clathrin.
Fig. 6. Expression of the CC domain of ARNO abolishes apical IgA endocytosis.
A. MDCK tet-off cells were grown for 3 days on Corning transwells. Control cells (uninfected) and
cells expressing either ARNO-WT or ARNO-CC domain, or co-expressing both for 18 h, were used
in a 5-min assay of radio-iodinated IgA endocytosis from the apical surface. The level of ARNO
expression was that used in Fig. 4B and C to allow monitoring of the inhibitory effect. Expression
of the CC domain of ARNO abolished the stimulatory effect of ARNO-WT on apical endocytosis. B.
Subcellular fractionation of MDCK cells expressing ARNO-WT and ARNO-∆coil was performed as
described in Materials. Crude PM and cytosolic fractions were resolved by SDS-PAGE, and
membranes were probed with anti-myc antibodies to detect ARNO-WT and ARNO-∆coil (B). To
verify accurate fractionation, membranes were probed with anti-pIgR (SC166) antibodies (not
shown). Western blots of three different experiments were scanned and densitometry performed using
NIH Image software. The histogram illustrates the ratio between ARNO-WT and ARNO-∆coil
appearing in the crude PM compared to the cytosolic fraction (mean ± SD, n = 3; p < 0.001 indicates
significantly different from control by t test). ARNO-∆coil expression results in increased distribution
to the PM fraction, most likely due to additional binding to the basolateral PM.
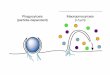
Fig. 7. The CC domain of ARNO links the receptor-ligand complex to the endocytic
system at the apical PM of epithel ia.
ARNO is positioned in the cytosol (A). Following phosphatase activity, ARNO is translocated to
the apical PM, to PIP2- and PIP3-containing membranes (B) where it colocalizes with the cargo
and associates with it (either directly or indirectly through an adaptor, X) via the ARNO-CC
domain. Subsequently, ARNO activates ARF6 by performing an exchange of GTP for GDP on
the latter that results in recruitment of the clathrin endocytic system (C). B’. When mutant
containing only CC domain is expressed, it competes with ARNO for binding the cargo, which
results in a lack of recruitment of the endocytic system and inhibition of CME.
by guest on May 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from

B.
0
5
10
15
20
25
30
Control ARNO-WT
Api
calI
gAup
take
(%of
tota
lIgA
boun
d)
ARNO-D coil
ARNO-D PH
C.
0
10
20
30
40
50
control
Bas
olat
eral
IgA
upta
ke(%
ofto
talI
gAbo
und)
ARNO-WT
ARNO- Dcoil
ARNO-D PH
Figure 4
CC Sec7 PH
CC Sec7
Sec7 PH
CC100 200 300 4000
ARNO-WT
ARNO-D PH
ARNO-D coil coiled
Coil coiled
A.
by guest on May 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from

D.
0
5
10
15
20
25
30
control ARF6-WT
Ap
ical
IgA
up
take
(%o
fto
talI
gA
bo
un
d)
ARNO-WT +
ARF6-WT
E.
Figure 4
0
5
10
15
20
25
30
control ARF6-T27N
Ap
ical
IgA
up
take
(%o
fto
talI
gA
bo
un
d)
ARNO-WT +
ARF6-T27N
ARNO-WT
ARNO-WT
by guest on May 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from

CB
ARNO
X CC
PH
Sec7
ARF6
ARF6
GTP
GTP
Apical plasma membrane
X
CC
PH
Sec7
Apical plasma membrane
CC
B
X CC
PH
Sec7
CC
PH
Sec7X
Receptor
Adaptor
A
Figure 7 by guest on M
ay 18, 2020http://w
ww
.jbc.org/D
ownloaded from

and Yoram AltschulerShmuel Miriam, Lorraine C. Santy, Scott Frank, Dana Avrahami, James E. Casanova
polarized epithelial cellsArno through its coiled-coil domain regulates endocytosis at the apical surface of
published online February 16, 2006J. Biol. Chem.
10.1074/jbc.M513723200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on May 18, 2020
http://ww
w.jbc.org/
Dow
nloaded from