Embed Size (px)
Citation preview

ORIGINAL ARTICLE Aquaculture
Use of a Noctiluca-killing bacterium Marinobacter salsuginis strainBS2 to reduce shrimp mortality caused by Noctiluca scintillans
Teeyaporn Keawtawee • Kimio Fukami •
Putth Songsangjinda
Received: 26 September 2011 / Accepted: 15 March 2012 / Published online: 13 April 2012
� The Japanese Society of Fisheries Science 2012
Abstract The ability of an algicidal bacterium Marin-
obacter salsuginis strain BS2, isolated from shrimp pond
water, to reduce shrimp mortality was investigated
under laboratory conditions. When two species of shrimp
(Penaeus monodon and Litopenaeus vannamei) (body
length 1.5–1.8 cm) were cultured together with the dino-
flagellate Noctiluca scintillans, nearly 80 % of the shrimps
died within 7 days. However, when bacterial strain BS2
was also added to the culture, N. scintillans was killed
within 48 h, and shrimp survival rates on the 7th day
improved from 23 to 87 % for both P. monodon and
L. vannamei. The bacterium BS2 alone had no effect on
shrimp condition. Under conditions of increased dissolved
oxygen, the effect of using BS2 was greater, and shrimp
survival rates improved even more dramatically, from 26 to
98 %. These studies provide the first evidence that the use
of killing bacteria, isolated from shrimp culture water, can
suppress harmful algal blooms (HABs) and thus restore the
efficiency of shrimp production. The control of HABs in
this way in shrimp culture farms would be a major benefit
for shrimp production.
Keywords Killing bacteria � Noctiluca scintillans �HABs � Shrimp culture
Introduction
Shrimp aquaculture is one of the fastest developing eco-
nomic activities in the Asia–Pacific region, where almost
80 % of the world’s production of farmed shrimp occurs
[30, 33]. In an intensive shrimp culture system, the concen-
tration of ammonia, resulting from excretion by the cultured
animals and mineralization of organic detritus, such as
unconsumed feed and feces, can reach toxic levels. The
accumulation of ammonia in pond water may cause the
deterioration of water quality, reduced growth, increased
oxygen consumption and ammonia–N excretion by shrimps,
and even high mortality [5, 32]. The lethal effects of
ammonia on juveniles of penaeid shrimps have been reported
for Penaeus chinensis, P. penicillatus [9], P. monodon [8],
P. paulensis [24], P. semisulcatus [31], P. japonicas juve-
niles [7], and Metapenaeus ensis [23].
Dinoflagellate blooms also cause disease and increased
mortality of cultured shrimp [2, 6]. Noctiluca scintillans is
a large, luminescent, heterotrophic dinoflagellate which
commonly forms red tides in temperate and subtropical
coastal regions. N. scintillans blooms have been reported in
New Zealand [4], Japan, China, Hong Kong [6, 16],
Indonesia, Malaysia, and Thailand [28, 29], providing
evidence that such bloom events are often associated with
massive fish and marine invertebrate mortality events.
Extensive blooms have been reported on the east and west
coasts of India, where they have been implicated in the
decline of fishery resources [1, 3]. The dense blooms of
green Noctiluca sp. are occasionally a stress factor for
shrimps, with a lower or even total loss of production
reported in Thailand [27] and mass mortality of shrimp
populations in China [6]. Although Noctiluca does not
produce toxins, it is catalogued as a harmful algal bloom
(HAB) species on the basis of its association with mass
T. Keawtawee (&) � K. Fukami
Graduate School of Kuroshio Science, Kochi University,
200 Monobe, Nankoku, Kochi 783-8502, Japan
e-mail: [email protected]
P. Songsangjinda
Department of Fisheries, Marine Shrimp Culture Research
Institute, Coastal Fisheries Research and Development Bureau,
Kaset-Klang, Chatuchak, Bangkok 10900, Thailand
123
Fish Sci (2012) 78:641–646
DOI 10.1007/s12562-012-0497-1

mortalities in farmed finfish due to oxygen depletion, gill
clogging, and high ammonia levels [25] resulting from the
high levels of ammonium and phosphate in N. scintillans
cells [15, 22, 25].
Previous studies have shown that various bacterial spe-
cies in the natural environment are capable of inhibiting or
controlling the growth of HAB [11, 13, 17]. Other studies
have shown that naturally occurring bacteria are also
capable of controlling the growth of phytoplankton [10, 12,
18, 20] and playing important roles during the development
and decay periods of red tide by influencing the initiation,
growth, maintenance and termination of phytoplankton
populations [14, 19, 21, 26]. In a recent study carried out
by our group, we found that some bacteria, which had been
isolated from a shrimp culture pond and identified as
Marinobacter sulsuginis, Vibrio sp., and Pseudoaltero-
monas sp. could suppress the growth of N. scintillans [20].
We also found that several strains of killing bacteria iso-
lated from shrimp culture water showed species-specific
inhibition against the growth of N. scintillans by either
direct or indirect attack [20]. However, to the best of our
knowledge, there have been no published reports on the
role of algicidal bacteria on shrimps in culture systems. It is
therefore essential to study the effect of isolated bacterial
strains that inhibit the growth of HABs, N. scintillans in
particular, in shrimp culture systems. If it is possible to use
killing bacteria to control the formation of N. scintillans
bloom, this would represent a promising approach by
which to improve the efficiency of shrimp farming man-
agement and shrimp production.
The aim of this study was to determine the ability of a
killing bacterium to suppress N. scintillans growth in a
shrimp culture system under laboratory conditions as a first
step in evaluating the potential applications of the isolated
bacterium for suppressing N. scintillans and controlling
HABs in shrimp farms.
Materials and methods
Cultures of N. scintillans and killing bacteria
Noctiluca scintillans was isolated from seawater at the
Sriracha Station (13.12�N, 100.56�E), Mueang Chon Buri,
Chonburi Province in June–July 2009 and from Don Hoy
Load Station, Samut Songkhram Province, Thailand in
January–February 2010.
The most effective killing bacterium (Marinobacter sul-
suginis strain BS2) was isolated from a pond used for the
culture of the shrimp P. monodon in June to July 2008 during
60–110 days of shrimp stocking at the Domrong shrimp farm
(6.845�N, 101.00�E), Thepha, Songkhla Province, Thailand
[20]. The ranges of salinity, temperature, and pH in the
shrimp pond were 23–26 ppt, 29–30 �C and 7.8–8.8,
respectively [20].
Shrimp culture experiments
The bacterium Marinobacter sulsuginis BS2 strain was
incubated in FeTY liquid medium at 28 ± 1 �C for 48 h in
preparation for the experiment. The cell suspension was
then centrifuged at 4500 rpm for 10–15 min, and the cells
were rinsed with sterile seawater and resuspended in fil-
tered seawater. The initial bacterial density was determined
by the DAPI method.
The first experiment was carried out to compare the
sensitivity of shrimps of different species and sizes to
N. scintillans in shrimp culture under laboratory condi-
tions. Ten individuals of each shrimp species, i.e., Penaeus
monodon (Black tiger shrimp) and Litopenaeus vannamei
(White shrimp), with a body length of about 1.5–1.8 cm
(small size) and about 3.5–4.0 cm (larger size), were reared
in 500 ml seawater in 1-l glass bottles (Fig. 1). The first
control (S) system contained only shrimps without any
addition of N. scintillans or bacterial strain BS2. The sec-
ond control (SB) system contained shrimps and BS2, but
no N. scintillans. The first treatment (SN) system contained
shrimps and N. scintillans without BS2, and the test system
contained shrimps, N. scintillans, and bacterial strain BS2
(SNB). The initial N. scintillans density was 10 cells/ml,
which had increased to 20 cells/ml with the addition of N.
scintillans cells after a 12-h incubation, followed by the
addition of BS2 (concentration approx. 103 cells/ml) in the
SB and SNB treatments. Each treatment was carried out in
Fig. 1 Shrimp rearing procedure under laboratory conditions.
S shrimp, SB shrimp ? Noctiluca-killing bacteria strain BS2, SNshrimp ? N. scintillans, SNB shrimp ? N. scintillans ? Noctiluca-
killing bacteria strain BS2
642 Fish Sci (2012) 78:641–646
123
triplicate. The supply of dissolved oxygen (DO) was con-
trolled at more than 4–5 mg O2/l by means of an air pump.
Observations were made at 12-h intervals for 168 h.
Shrimp were assumed to be dead when they were immobile
and showed no response when touched with a glass rod.
Dead shrimps were removed daily from the containers.
N. scintillans abundance, total bacteria density, and shrimp
survival were monitored every day until the 7th day
according to the procedure described in Fig. 1.
For the second experiment, the effect of DO on shrimp
survival rates was investigated. P. monodon (Black tiger
shrimp) juveniles (body length about 2.8–3.2 cm) were
used in treatments with and without DO [concentration of
DO in the treatment with DO: 4–5 mgO2/l (controlled by
an air pump)] . The DO concentration was determined
using a DO meter twice daily (7:00 a.m. and 3:00 p.m.)
The same series of treatments was used, in triplicate, as in
the first experiment. For each treatment, 30 shrimps were
placed in a 20-l aquarium tank. The initial cell density was
20 cells/ml for N. scintillans and approximately 103 cells/
ml for bacterial strain BS2. The abundance of N. scintillans
and the shrimp survival were monitored every day for
7 days. Water samples (30 ml) were collected and used for
determining ammonia concentration on days 0, 1, 3, 5, and
7 of the experiment by means of TRACCS-800.
Statistical analysis
The significance of variations in shrimp mortality among
treatments was tested by one-way analysis of variance
(ANOVA). The mean comparison was performed by
Duncan’s multiple range tests. All of the statistical tests
were considered to be significant at the 95 % significant
level of confidence.
Results
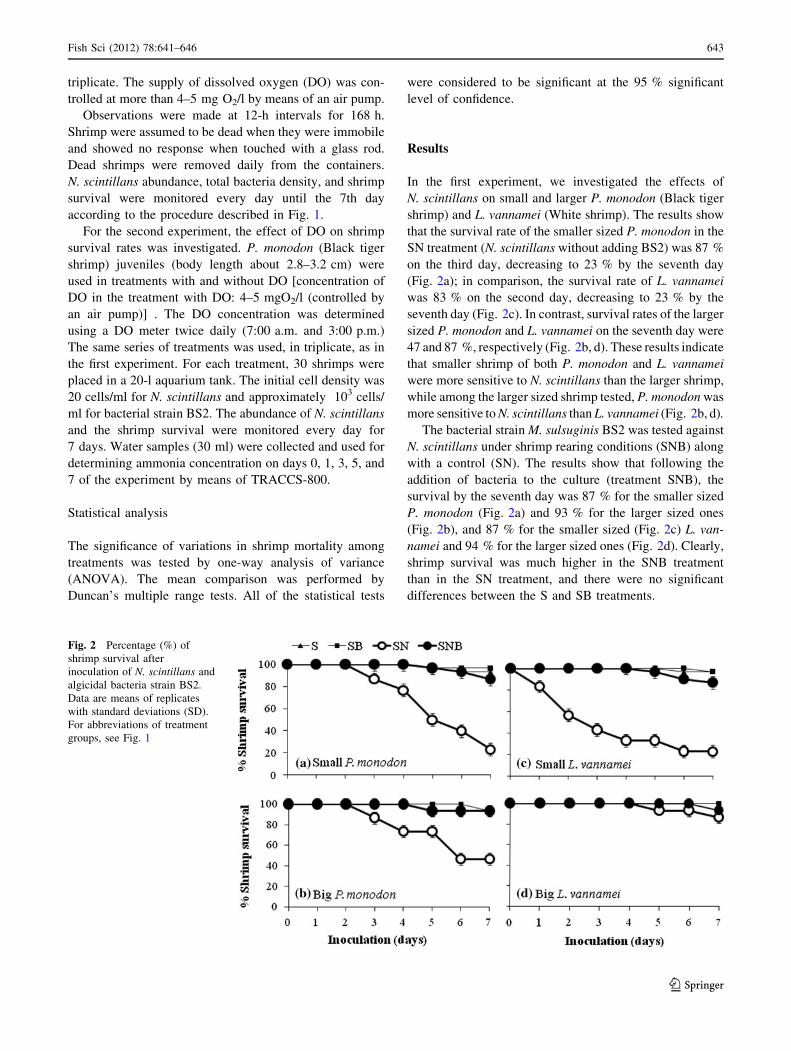
In the first experiment, we investigated the effects of
N. scintillans on small and larger P. monodon (Black tiger
shrimp) and L. vannamei (White shrimp). The results show
that the survival rate of the smaller sized P. monodon in the
SN treatment (N. scintillans without adding BS2) was 87 %
on the third day, decreasing to 23 % by the seventh day
(Fig. 2a); in comparison, the survival rate of L. vannamei
was 83 % on the second day, decreasing to 23 % by the
seventh day (Fig. 2c). In contrast, survival rates of the larger
sized P. monodon and L. vannamei on the seventh day were
47 and 87 %, respectively (Fig. 2b, d). These results indicate
that smaller shrimp of both P. monodon and L. vannamei
were more sensitive to N. scintillans than the larger shrimp,
while among the larger sized shrimp tested, P. monodon was
more sensitive to N. scintillans than L. vannamei (Fig. 2b, d).
The bacterial strain M. sulsuginis BS2 was tested against
N. scintillans under shrimp rearing conditions (SNB) along
with a control (SN). The results show that following the
addition of bacteria to the culture (treatment SNB), the
survival by the seventh day was 87 % for the smaller sized
P. monodon (Fig. 2a) and 93 % for the larger sized ones
(Fig. 2b), and 87 % for the smaller sized (Fig. 2c) L. van-
namei and 94 % for the larger sized ones (Fig. 2d). Clearly,
shrimp survival was much higher in the SNB treatment
than in the SN treatment, and there were no significant
differences between the S and SB treatments.
Fig. 2 Percentage (%) of
shrimp survival after
inoculation of N. scintillans and
algicidal bacteria strain BS2.
Data are means of replicates
with standard deviations (SD).
For abbreviations of treatment
groups, see Fig. 1
Fish Sci (2012) 78:641–646 643
123
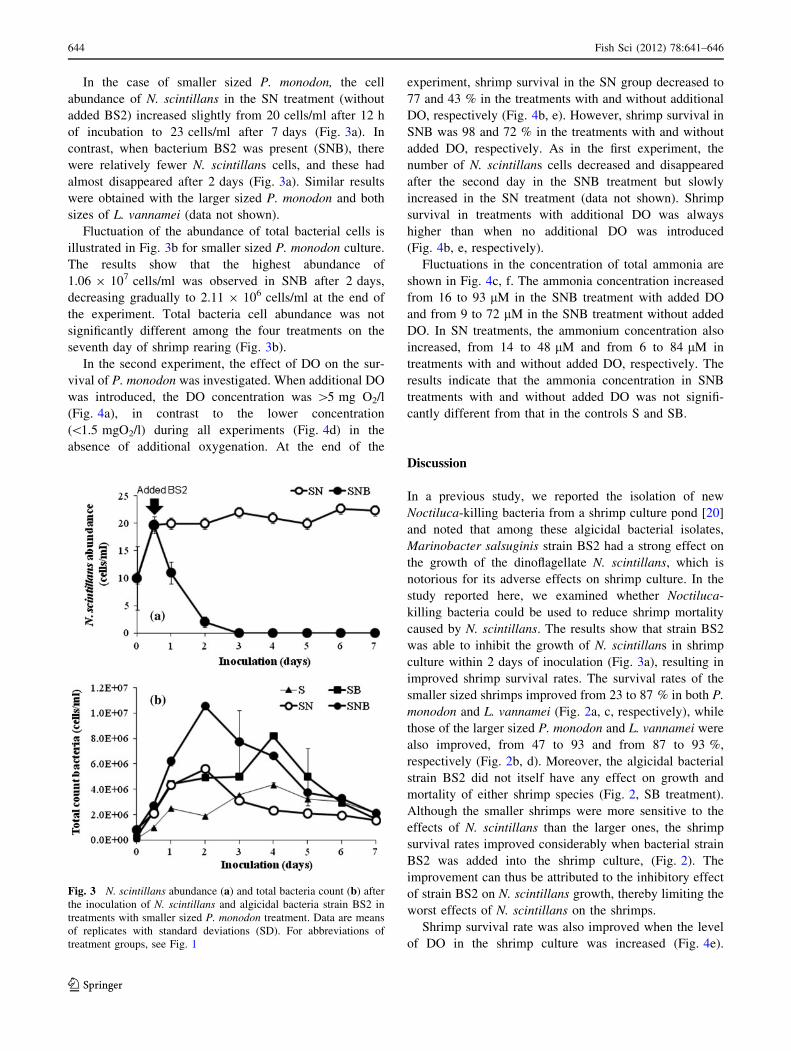
In the case of smaller sized P. monodon, the cell
abundance of N. scintillans in the SN treatment (without
added BS2) increased slightly from 20 cells/ml after 12 h
of incubation to 23 cells/ml after 7 days (Fig. 3a). In
contrast, when bacterium BS2 was present (SNB), there
were relatively fewer N. scintillans cells, and these had
almost disappeared after 2 days (Fig. 3a). Similar results
were obtained with the larger sized P. monodon and both
sizes of L. vannamei (data not shown).
Fluctuation of the abundance of total bacterial cells is
illustrated in Fig. 3b for smaller sized P. monodon culture.
The results show that the highest abundance of
1.06 9 107 cells/ml was observed in SNB after 2 days,
decreasing gradually to 2.11 9 106 cells/ml at the end of
the experiment. Total bacteria cell abundance was not
significantly different among the four treatments on the
seventh day of shrimp rearing (Fig. 3b).
In the second experiment, the effect of DO on the sur-
vival of P. monodon was investigated. When additional DO
was introduced, the DO concentration was [5 mg O2/l
(Fig. 4a), in contrast to the lower concentration
(\1.5 mgO2/l) during all experiments (Fig. 4d) in the
absence of additional oxygenation. At the end of the
experiment, shrimp survival in the SN group decreased to
77 and 43 % in the treatments with and without additional
DO, respectively (Fig. 4b, e). However, shrimp survival in
SNB was 98 and 72 % in the treatments with and without
added DO, respectively. As in the first experiment, the
number of N. scintillans cells decreased and disappeared
after the second day in the SNB treatment but slowly
increased in the SN treatment (data not shown). Shrimp
survival in treatments with additional DO was always
higher than when no additional DO was introduced
(Fig. 4b, e, respectively).
Fluctuations in the concentration of total ammonia are
shown in Fig. 4c, f. The ammonia concentration increased
from 16 to 93 lM in the SNB treatment with added DO
and from 9 to 72 lM in the SNB treatment without added
DO. In SN treatments, the ammonium concentration also
increased, from 14 to 48 lM and from 6 to 84 lM in
treatments with and without added DO, respectively. The
results indicate that the ammonia concentration in SNB
treatments with and without added DO was not signifi-
cantly different from that in the controls S and SB.
Discussion
In a previous study, we reported the isolation of new
Noctiluca-killing bacteria from a shrimp culture pond [20]
and noted that among these algicidal bacterial isolates,
Marinobacter salsuginis strain BS2 had a strong effect on
the growth of the dinoflagellate N. scintillans, which is
notorious for its adverse effects on shrimp culture. In the
study reported here, we examined whether Noctiluca-
killing bacteria could be used to reduce shrimp mortality
caused by N. scintillans. The results show that strain BS2
was able to inhibit the growth of N. scintillans in shrimp
culture within 2 days of inoculation (Fig. 3a), resulting in
improved shrimp survival rates. The survival rates of the
smaller sized shrimps improved from 23 to 87 % in both P.
monodon and L. vannamei (Fig. 2a, c, respectively), while
those of the larger sized P. monodon and L. vannamei were
also improved, from 47 to 93 and from 87 to 93 %,
respectively (Fig. 2b, d). Moreover, the algicidal bacterial
strain BS2 did not itself have any effect on growth and
mortality of either shrimp species (Fig. 2, SB treatment).
Although the smaller shrimps were more sensitive to the
effects of N. scintillans than the larger ones, the shrimp
survival rates improved considerably when bacterial strain
BS2 was added into the shrimp culture, (Fig. 2). The
improvement can thus be attributed to the inhibitory effect
of strain BS2 on N. scintillans growth, thereby limiting the
worst effects of N. scintillans on the shrimps.
Shrimp survival rate was also improved when the level
of DO in the shrimp culture was increased (Fig. 4e).
Fig. 3 N. scintillans abundance (a) and total bacteria count (b) after
the inoculation of N. scintillans and algicidal bacteria strain BS2 in
treatments with smaller sized P. monodon treatment. Data are means
of replicates with standard deviations (SD). For abbreviations of
treatment groups, see Fig. 1
644 Fish Sci (2012) 78:641–646
123
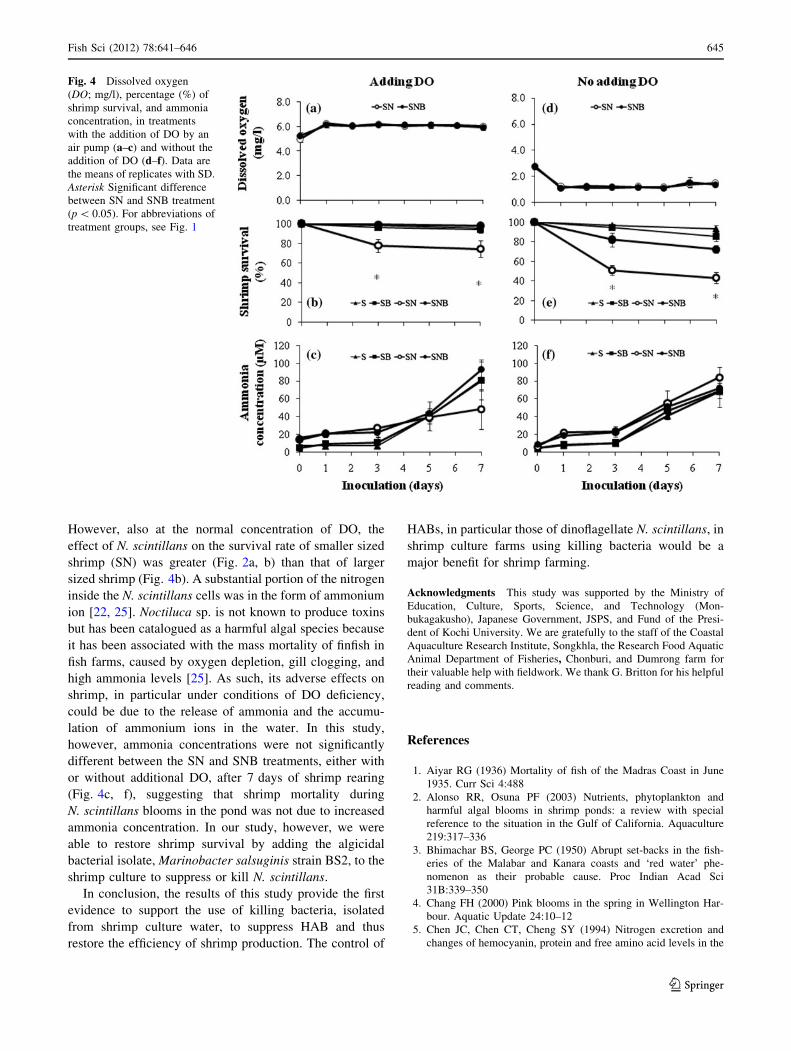
However, also at the normal concentration of DO, the
effect of N. scintillans on the survival rate of smaller sized
shrimp (SN) was greater (Fig. 2a, b) than that of larger
sized shrimp (Fig. 4b). A substantial portion of the nitrogen
inside the N. scintillans cells was in the form of ammonium
ion [22, 25]. Noctiluca sp. is not known to produce toxins
but has been catalogued as a harmful algal species because
it has been associated with the mass mortality of finfish in
fish farms, caused by oxygen depletion, gill clogging, and
high ammonia levels [25]. As such, its adverse effects on
shrimp, in particular under conditions of DO deficiency,
could be due to the release of ammonia and the accumu-
lation of ammonium ions in the water. In this study,
however, ammonia concentrations were not significantly
different between the SN and SNB treatments, either with
or without additional DO, after 7 days of shrimp rearing
(Fig. 4c, f), suggesting that shrimp mortality during
N. scintillans blooms in the pond was not due to increased
ammonia concentration. In our study, however, we were
able to restore shrimp survival by adding the algicidal
bacterial isolate, Marinobacter salsuginis strain BS2, to the
shrimp culture to suppress or kill N. scintillans.
In conclusion, the results of this study provide the first
evidence to support the use of killing bacteria, isolated
from shrimp culture water, to suppress HAB and thus
restore the efficiency of shrimp production. The control of
HABs, in particular those of dinoflagellate N. scintillans, in
shrimp culture farms using killing bacteria would be a
major benefit for shrimp farming.
Acknowledgments This study was supported by the Ministry of
Education, Culture, Sports, Science, and Technology (Mon-
bukagakusho), Japanese Government, JSPS, and Fund of the Presi-
dent of Kochi University. We are gratefully to the staff of the Coastal
Aquaculture Research Institute, Songkhla, the Research Food Aquatic
Animal Department of Fisheries, Chonburi, and Dumrong farm for
their valuable help with fieldwork. We thank G. Britton for his helpful
reading and comments.
References
1. Aiyar RG (1936) Mortality of fish of the Madras Coast in June
1935. Curr Sci 4:488
2. Alonso RR, Osuna PF (2003) Nutrients, phytoplankton and
harmful algal blooms in shrimp ponds: a review with special
reference to the situation in the Gulf of California. Aquaculture
219:317–336
3. Bhimachar BS, George PC (1950) Abrupt set-backs in the fish-
eries of the Malabar and Kanara coasts and ‘red water’ phe-
nomenon as their probable cause. Proc Indian Acad Sci
31B:339–350
4. Chang FH (2000) Pink blooms in the spring in Wellington Har-
bour. Aquatic Update 24:10–12
5. Chen JC, Chen CT, Cheng SY (1994) Nitrogen excretion and
changes of hemocyanin, protein and free amino acid levels in the
Fig. 4 Dissolved oxygen
(DO; mg/l), percentage (%) of
shrimp survival, and ammonia
concentration, in treatments
with the addition of DO by an
air pump (a–c) and without the
addition of DO (d–f). Data are
the means of replicates with SD.
Asterisk Significant difference
between SN and SNB treatment
(p \ 0.05). For abbreviations of
treatment groups, see Fig. 1
Fish Sci (2012) 78:641–646 645
123

hemolymph of Penaeus monodon exposed to different concen-
trations of ambient ammonia–N at different salinity levels. Mar
Ecol Prog Ser 110:85–94
6. Chen YQ, Gu XG (1993) An ecological study of red tides in the
East China Sea. In: Smayda TJ, Schimizu L (eds) Toxic phyto-
plankton blooms in the sea. Elsevier, Amsterdam, pp 217–221
7. Chen JC, Kuo YZ (1992) Effect of ammonia on growth and
molting of Penaeus japonicas juveniles. Aquaculture 104:
249–260
8. Chen JC, Lei SC (1990) Toxicities of ammonia and nitrite to
Penaeus monodon juveniles. J World Aquatic Soc 21:300–306
9. Chen JC, Lin CY (1992) Lethal effects of ammonia on Penaeuschinensis Osbeck juveniles at different salinity levels. J Exp Mar
Biol Ecol 156:139–148
10. Clinton EH, Elif D, Kathryn CJ, CARY SC, Kirchman DL, David
AH (2005) A bacterium that inhibits the growth of Pfiesteriapiscicida and other dinoflagellates. Harmful Algae 4:221–234
11. Doucette GJ, McGovern ER, Babinchak JA (1999) Algicidal
bacteria active against Gymnodinium breve (Dinophyceae).
I. Bacterial isolation and characterization of killing activity.
J Phycol 35:1447–1454
12. Fukami K, Nishijima T, Murata H, Doi S, Hata Y (1991) Dis-
tribution of bacteria influential on the development and the decay
of Gymnodinium nagasakiense red tide and their effect on algal
growth. Nippon Suisan Gakkaishi 57:2321–2326
13. Fukami K, Yuzawa A, Nishijima T, Hata Y (1992) Isolation and
properties of a bacterium inhibiting the growth of Gymnodiniumnagasakiense. Nippon Suisan Gakkaishi 58:1073–1077
14. Fukami K, Yuzawa A, Sakaguchi K, Nishijima T (1995) Pre-
vention of noxious red tide by algicidal bacteria. In: Proc Int Conf
Ecological System Enhancement Technology for Aquatic Envi-
ronments. Japan International Science and Technology Federa-
tion, Tokyo, pp 89–94
15. Fukuyo Y, Takano H, Chihara M, Matsuoka K (1990) Red tide
organisms in Japan. An illustrated taxonomic guide. Uchida
Rokakuho, Tokyo
16. Huang C, Qi H (1997) The abundance cycle and influence factors
on red tide phenomena of Noctiluca scintillans (Dinophyceae) in
Dapeng Bay, the South China Sea. J Plank Res 19:303–318
17. Imai I, Ishida Y, Hata Y (1993) Killing of marine phytoplankton
by a gliding bacterium Cytophaga sp., isolated from the coastal
Sea of Japan. Mar Biol 116:527–532
18. Imai I, Ishida Y, Sawayama K, Hata Y (1991) Isolation of marine
gliding bacterium that kills Chattonella antique (Raphidophy-
ceae). Nippon Suisan Gakkaishi 57:1409
19. Imai I, Kim MC, Nagasaki K, Itakura S, Ishida Y (1998) Rela-
tionships between dynamics of red tide-causing raphidophycean
flagellates and algicidal micro-organisms in the coastal sea of
Japan. Phycol Res 46:139–146
20. Keawtawee T, Fukami K, Songsangjinda P, Muangyao P (2011)
Isolation and characterization of Noctiluca-killing bacteria from a
shrimp aquaculture pond in Thailand. Fish Sci 7:657–664
21. Kodama M, Doucette GJ, Green DH (2006) Relationships
between bacteria and harmful algae. In: Graneli E, Turner JT
(eds) Ecology of harmful algae, ecological studies, vol 189.
Springer, Heidelberg, pp 244–255
22. Montani S, Pithakpol S, Tada K (1998) Nutrient regeneration in
coastal sea by Noctiluca scintillans, a red tide causing dinofla-
gellate. J Mar Biotechnol 6:224–228
23. Nan FH, Chen JC (1991) Lethal effect of ammonia to juvenile
Metapenaeus ensis. J Fish Soc Taiwan 18:41–46
24. Ostrensky A, Wasielesky JW (1995) Acute toxicity of ammonia
to various life stages of the Sao Paulo shrimp, Penaeus paulensisPerez-Farfante, 1967. Aquaculture 132:339–347
25. Okaichi T, Nishio S (1976) Identification of ammonia as the toxic
principle of red tide of Noctiluca miliaris. Bull Plank Soc Jpn
23:75–80
26. Riquelme CE, Fukami K, Ishida Y (1988) Effects of bacteria on
the growth of a marine diatom, Asterionella glacialis. Bull Jpn
Soc Microb Ecol 3:29–34
27. Songsangjinda P, Yamamoto T, Fukami K, Keawtawee T (2006)
Importance of controlling community structure of living organ-
isms in intensive shrimp culture ponds. Coast Mar Sci 30:91–99
28. Suwapeepun S (1995) Red tide and a study of red tide in Thai
waters. In: Proc Workshop Seminar on Red Tide. Eastern Marine
Fisheries Development Center, Bangkok
29. Thon-gra-ar W, Pinkaew K, Musika C, Wongsudawan W (1998)
A baseline study on the occurrence of red tide plankton in the
coastal waters from the Bang Pakong River Mouth to Sri Racha,
Chonburi province. Thai J Aquatic Sci 4:60–78
30. Trott LA, Alongi DM (2000) The impact of shrimp pond effluent
on water quality and phytoplankton biomass in a tropical man-
grove estuary. Mar Pollut Bull 40:947–951
31. Wajsbrot N, Gasith A, Krom MD, Samocha TM (1990) Effect of
dissolved oxygen and the molt stage on the acute toxicity of
ammonia to juvenile green tiger prawn Penaeus semisulcatus.
Environ Toxicol Chem 9(4):497–504
32. Wickins JF (1976) Tolerance of warm-water prawns to recircu-
lated water. Aquaculture 9:19–37
33. Wolanski E, Spagnol S, Thomas S, Moore K, Alongi DM, Trott
L, Davidson A (2000) Modelling and visualizing the fate of
shrimp pond effluent in a mangrove-fringed tidal Creek. Estuar
Coast Shelf Sci 50:85–97
646 Fish Sci (2012) 78:641–646
123





























