Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
CENTRO DE BIOCIÊNCIAS
PROGRAMA DE PÓS-GRADUAÇÃO EM PSICOBIOLOGIA
ECOLOGIA DE COMUNIDADES E COMPORTAMENTO REPRODUTIVO DE
ANFÍBIOS ANUROS EM SAVANA AMAZÔNICA
CARLOS EDUARDO COSTA CAMPOS
NATAL/RN
2015

ii
UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
CENTRO DE BIOCIÊNCIAS
PROGRAMA DE PÓS-GRADUAÇÃO EM PSICOBIOLOGIA
ECOLOGIA DE COMUNIDADES E COMPORTAMENTO REPRODUTIVO DE
ANFÍBIOS ANUROS EM SAVANA AMAZÔNICA
CARLOS EDUARDO COSTA CAMPOS
ORIENTADORA: DRA. ELIZA MARIA XAVIER FREIRE
Tese de Doutorado apresentada ao Programa de Pós-
Graduação em Psicobiologia da Universidade Federal
do Rio Grande do Norte (Área de Concentração:
Comportamento Animal; Linha de Pesquisa:
Ecologia e Comportamento Animal), como requisito
para obtenção do título de Doutor em Psicobiologia.
NATAL/RN
2015

iii
UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
CENTRO DE BIOCIÊNCIAS
PROGRAMA DE PÓS-GRADUAÇÃO EM PSICOBIOLOGIA
A TESE: ECOLOGIA DE COMUNIDADES E COMPORTAMENTO REPRODUTIVO
DE ANFÍBIOS ANUROS EM SAVANA AMAZÔNICA
ELABORADA POR: Carlos Eduardo Costa Campos
e aprovada por todos os membros da Banca Examinadora, foi aceita pelo Programa de
Pós-Graduação em Psicobiologia e homologada pelos membros da banca, como requisito
à obtenção do título de: DOUTOR EM PSICOBIOLOGIA
DATA DA DEFESA: 23 de Fevereiro de 2015
BANCA EXAMINADORA
____________________________________________
Profa. Dra. Eliza Maria Xavier Freire (Presidente)
Universidade Federal do Rio Grande do Norte
____________________________________________
Profa. Dra. Gilda Vasconcellos de Andrade
Universidade Federal do Maranhão
____________________________________________
Prof. Dr. Marcelo Nogueira de Carvalho Kokubum
Universidade Federal de Campina Grande
____________________________________________
Prof. Dr. Wallisen Tadashi Hattori
Universidade Federal do Rio Grande do Norte
____________________________________________
Profa. Dra. Maria de Fátima Arruda de Miranda
Universidade Federal do Rio Grande do Norte
____________________________________________
Prof. Dr. Arrilton Araújo de Souza (Suplente)
Universidade Federal do Rio Grande do Norte


iv
DEDICATÓRIA
AOS 6 M’S PRESENTES NA MINHA VIDA...
MARIA SALETE COSTA (DONA BRANCA, MÃE)
MARIA SOCORRO COSTA (TIA CO)
MAYARA MELO FURTADO (ESPOSA)
MARIA EDUARDA E JOÃO EDUARDO MELO (FILHOS)
ELIZA MARIA XAVIER FREIRE (JUJU, ORIENTADORA)

v
Cobras cegas são notívagas
O orangotango é profundamente solitário
Macacos também preferem o isolamento
Certas árvores só frutificam de 25 em 25 anos
Andorinhas copulam no vôo
O mundo não é o que pensamos.
Carlos Drummond de Andrade

vi
AGRADECIMENTOS
Agradeço a Deus, não só nesta etapa, mas em toda minha vida, pois me presenteou com
vida, saúde, força, coragem e sabedoria para entender e superar as provações.
A Profa. Eliza Maria Xavier Freire pela orientação, discussões, idéias, construções,
destruições e reconstruções e, acima de tudo pelo profissionalismo, dedicação, amizade e
paciência. Agradeço imensamente por ter me aceitado como orientando e por estar sempre
disposta a me ajudar em todos os momentos.
A Universidade Federal do Rio Grande do Norte e ao Programa de Pós Graduação em
Psicobiologia pela oportunidade para realização do Doutorado.
A todos os professores do Programa de Pós Graduação em Psicobiologia/UFRN pelos
ensinamentos adicionados durante o curso, principalmente sobre comportamento animal.
A Universidade Federal do Amapá e ao Colegiado do Curso de Ciências Biológicas pela
liberação integral para cursar o Doutorado.
A Fundação de Amparo a Pesquisa do Estado do Amapá (FAPEAP, Processo N0.
250.203/059/2014), que através do Edital N0. 05/2013 – Custeio Tese, forneceu apoio
financeiro para realização deste trabalho.
A Empresa Brasileira de Pesquisa Agropecuária do Estado do Amapá, em especial, ao
Dr. Silas Mochiutti e aos funcionários do Campo Experimental da Embrapa.
Ao ICMBio pela licença concedida para coleta dos anfíbios anuros.
Ao Núcleo de Hidro-Metereologia e Energia Renováveis do Instituto de Pesquisas
Científicas e Tecnológicas do Estado do Amapá por fornecer os dados ambientais
utilizados neste trabalho.
A todos os amigos do Laboratório de Herpetologia da Universidade Federal do Rio
Grande do Norte – Jaqueiuto, Mateus, Miguel, Raul, Bruno, Luan, Jaqueline, Kallyne,
Iaponira, Thaise, Mycarla e Jadna por todo o apoio nos momentos “raros” de convivência.
Aos estagiários do Laboratório de Herpetologia da Universidade Federal do Amapá,
Auridan, Ananda, Nayara, Endrea, Telma, Keison, Yuri, Daniel, Mayara, Raimundo,
Hélio, Bruna, Lígia, Deise. Agradeço a oportunidade de trabalhar, conviver e aprender
com vocês, pelo companheirismo e pelas memoráveis noites molhados e sem dormir.
A Marinus Hoogmoed, Albertina Lima, Thiago Carvalho e André Pansonato pela
identificação e confirmação dos anuros coletados na Savana amapaense.
A Júlio Sá e Andréa Soares pelo incentivo, críticas, idéias e sugestões fornecidas para a
execução desse trabalho. Mais do que amigos... irmãos.

vii
Ao Prof. Dr. Raimundo Nonato Picanço Souto, do Laboratório de Artrópodes da
Universidade Federal do Amapá, pela ajuda e identificação dos atrópodes presentes no
conteúdo estomacal dos anuros.
A Huann Vasconcelos pela ajuda na elaboração dos mapas e formatação da Tese.
Um agradecimento especial a minha esposa e grande amor Mayara Melo, que participou
de todas as fases deste trabalho. Seu apoio foi fundamental para elaboração desta Tese,
desde sua companhia e auxílio em várias fases de campo; e cujo carinho e amor sempre
me deram forças para agüentar a ausência e a distância.
A toda minha família que sempre esteve, mesmo que de muito longe, me ajudando e me
dando forças para seguir em frente.
E por fim, a todas as pessoas que participaram diretamente ou indiretamente na realização
deste trabalho e que aqui não foram citados.
MUITO OBRIGADO!!!
EDUARDO CAMPOS

viii
SUMÁRIO
RESUMO GERAL x
ABSTRACT xi
1. INTRODUÇÃO GERAL E REVISÃO DA LITERATURA 1
2. METODOLOGIA GERAL 9
2.1. CARACTERIZAÇÃO DA ÁREA DE ESTUDO 9
2.2. PROCEDIMENTOS METODOLÓGICOS 13
CAPÍTULO 1. RIQUEZA E COMPOSIÇÃO DE UMA TAXOCENOSE DE ANUROS DE
ÁREA DE SAVANA AMAZÔNICA
16
1. ABSTRACT 16
2. RESUMO 17
3. INTRODUÇÃO 17
4. MATERIAL E MÉTODOS 18
4.1. Área de Estudo 18
4.2. Procedimentos Metodológicos 19
4.2.1. Trabalho em campo 19
4.2.2. Análise dos dados 20
5. RESULTADOS E DISCUSSÃO 22
5.1. Riqueza e Composição de espécies 22
5.2. Eficiência amostral: Curva de rarefação e Estimadores de riqueza de espécies 25
5.3. Diversidade e Equitabilidade de espécies 27
5.4. Análise de Similaridade (Cluster analysis) e Ordenamento (NMDS) 28
6. REFERÊNCIAS BIBLIOGRÁFICAS 32
CAPÍTULO 2. ECOLOGIA DE COMUNIDADES DE ANUROS EM DIFERENTES
FITOFISIONOMIAS DE UMA ÁREA DE SAVANA AMAZÔNICA
40
1. RESUMO 40
2. ABSTRACT 41
3. INTRODUÇÃO 41
4. MATERIAL E MÉTODOS 42
4.1. Área de Estudo 42
4.2. Procedimentos Metodológicos 43
4.2.1. Constância de ocorrência e Abundância de espécies 44
4.2.2. Distribuição espacial (habitat e microhabitat) e sazonal 44
4.2.3. Distribuição temporal (período de atividade) 45
4.2.4. Análise da dieta 45
4.3. Análise dos dados 46
5. RESULTADOS E DISCUSSÃO 47
5.1. Uso e partilha de habitat e microhabitat pelas espécies 47
5.2. Constância das espécies 51
5.3. Abundância das espécies por habitat e microhabitat 52
5.4. Distribuição temporal (período de atividade) 58
5.5. Distribuição sazonal das espécies 62
5.6. Abundância, riqueza e sazonalidade 65
5.7. Composição da dieta 67
6. REFERÊNCIAS BIBLIOGRÁFICAS 76

ix
CAPÍTULO 3. ECOLOGIA E COMPORTAMENTO REPRODUTIVO DE ESPÉCIES DE
ANUROS EM SAVANA AMAZÔNICA
86
1. RESUMO 86
2. ABSTRACT 87
3. INTRODUÇÃO 87
4. MATERIAL E MÉTODOS 88
4.1. Área de Estudo 88
4.2. Procedimentos Metodológicos 89
4.2.1. Padrões temporais do comportamento reprodutivo 90
4.2.2. Modos reprodutivos 90
4.2.3. Estratégias reprodutivas comportamentais 90
4.2.4. Sítios de vocalização 91
4.2.5. Comportamentos reprodutivos 91
4.3. Análises dos dados 92
5. RESULTADOS E DISCUSSÃO 93
5.1. Padrões temporais do comportamento reprodutivo 93
5.2. Modos reprodutivos e Cuidado parental 96
5.3. Estratégias reprodutivas comportamentais 99
5.4. Sítios de vocalização 100
5.5. Comportamentos reprodutivos 109
5.5.1. Leptodactylus fuscus 111
5.5.2. Phyllomedusa hypochondrialis 112
5.5.3. Hypsiboas punctatus 113
5.5.4. Rhinella margaritifera 114
6. REFERÊNCIAS BIBLIOGRÁFICAS 117
CONSIDERAÇÕES FINAIS 126
REFERÊNCIAS 127

x
RESUMO GERAL
As Savanas amazônicas ocorrem em manchas disjuntas por extensas áreas de floresta nos
Estados do Amapá, Amazonas, Pará e Roraima. Para essas áreas é reconhecida variação
considerável na composição de espécies da anurofauna entre as localidades e as
fitofisionomias das Savanas amazônicas; no entanto, em função da carência de estudos
ecológicos, especialmente sobre comportamento reprodutivo, uma amostragem
sistemática, especificamente na Savana amapaense, é relevante na Amazônia Oriental.
Nessa perspectiva, foi estudada uma área de Savana no Estado do Amapá quanto à
composição, ecologia e comportamento reprodutivo de anfíbios anuros. Para as
amostragens dos anuros, foram realizadas 24 excursões para observacões e/ou coletas, de
janeiro de 2009 a dezembro de 2010 em cada fitofisionomia (Savana gramíneo-lenhosa,
Savana gramíneo-lenhosa arbustiva, Savana parque e Savana arbórea); para as análises
do comportamento reprodutivo foram feitas observações durante o período de janeiro a
dezembro de 2013, com duração de quatro dias consecutivos. As amostragens foram
realizadas através de busca ativa e auditiva ao longo de 20 parcelas de 100x50 metros.
Foram registradas 21 espécies de anuros, incluindo quatro novos registros de ocorrência
para o Estado do Amapá: Dendropsophus walfordi, Scinax fuscomarginatus,
Pseudopaludicola boliviana e Elachistocleis helianneae. A análise de variância de
Kruskal-Wallis revelou diferenças significativas quanto a riqueza e a diversidade de
espécies entre as fitofisionomias (p < 0.05). A análise de similaridade de Bray Curtis
reconheceu as fitofisionomias em três grupos: Savana arbórea, Savana gramíneo lenhosa
e gramíneo lenhosa arbustiva, e Savana parque. Através do Ordenamento por
escalonamento não-métrico multidimensional, a comunidade de anuros resultou em
agrupamentos distintos nas três fitofisionomias amostradas, com diferenças significativas
(ANOSIM, R = 0.823; p < 0.001), indicando diferença na composição de espécies. No
estudo de ecologia de comunidades, os valores obtidos para largura dos nichos espacial,
temporal e trófico sugerem que as espécies de anuros da Savana amapaense é composta
predominantemente por espécies generalistas. As análises de modelos nulos indicaram a
ocorrência de estruturação na comunidade quanto aos nichos temporal e trófico,
indicando influência significativa de fatores ecológicos contemporâneos nesse processo.
A ausência de estrutura quanto ao nicho espacial pode ser explicada pela segregação
espacial na distribuição e ocupação dos anuros nas diferentes fitofisionomias da Savana
amapaense, resultante de especializações. As estratégias reprodutivas, 11 espécies de
anuros foram classificadas com padrão de reprodução prolongado, intrinsecamente ligado
ao período chuvoso e ao modo reprodutivo da maioria das espécies, que apresentaram
desova em corpos d’água lênticos. Foram registrados seis modos reprodutivos, com
cuidado parental para as espécies Leptodactylus macrosternum e L. podicipinus, que
apresentaram modos reprodutivos caracterizados pelas desovas em ninhos de espuma.
Quanto às estratégias reprodutivas comportamentais, a estratégia do macho vocalizador
foi constatada em todas as espécies de anuros; a estratégia do macho satélite foi registrada
apenas para as espécies D. walfordi, Hypsiboas multifasciatus, S. nebulosus e S.
fuscomarginatus; a procura ativa por fêmeas foi registrada para as espécies Phyllomedusa
hypochondrialis e L. fuscus e, a estratégia de macho deslocador ocorreu apenas em
Rhinella major e R. margaritifera. Dos comportamentos reprodutivos registrados, a
exibição da região gular e do saco vocal está relacionado ao comportamento de corte e
territorialidade exibido pelos machos. Além dos comportamentos de corte, sinais visuais
associados foram registrados para os anuros da Savana amapaense.
PALAVRAS-CHAVE: Anuros, Savana amazônica, Amapá, Composição, Riqueza,
Estrutura de comunidades, Comportamento reprodutivo.

xi
ABSTRACT
The Amazon savannas occur as isolated patches throughout extensive areas of forest in
the states of Amapá, Amazonas, Pará, and Roraima. There is a considerable variation in
the composition of anuran assemblages in the localities and phytophysiognomies of
Amazon savannas and given the absence of studies on reproductive behavior, a systematic
and geographically wide sampling has been carried out in the Amapá savanna, located in
the Eastern Amazon. The study was conducted in a savanna area in the state of Amapá to
examine the composition, ecology, and reproductive behavior of anuran amphibians. We
carried out 24 field trips in each phytophysiognomy (gramineous-woody savana,
gramineous-herbaceous-woody savana, park savana, and arboreal savanna); for analysis
of reproductive behavior observations were made during the period January to December
2013, lasting four consecutive days. Samples were collected by active and acoustic search
along 20 plots of 100x50 meters. Twenty-one anuran species were recorded, of which
four are new records for the state of Amapá: Dendropsophus walfordi, Scinax
fuscomarginatus, Pseudopaludicola boliviana e Elachistocleis helianneae. The Kruskal-
Wallis ANOVA revealed significant differences between richness and species diversity
in the phytophysiognomies (p < 0.05). The Bray-Curtis similarity coefficient divided the
phytophysiognomies into three groups: arboreal savana, gramineous-woody savanna and
gramineous-herbaceous-woody savanna, and park savanna. According to the non-metric
multidimensional scaling, the structure of the anuran community resulted in a separation
into three phytophysiognomies, with significant differences in the structure of
communities (ANOSIM, R = 0.823; p < 0.001). In the study of community ecology, the
results obtained for spatial, temporal, and trophic niche breadth suggest that the
assemblage of anurans of the Amapá savanna is not composed of predominantly
generalist species. Also, the presence of other specialist anurans may explain the
processes of speciation associated with the isolation of habitats, resulting in heterogeneity
and spatial discontinuity in the phytophysiognomies with open formations. The null
model analysis revealed that the community is structured based on temporal and trophic
niche, indicating a significant influence of contemporary ecological factors on the
assemblage. The absence of structure based on spatial niche might be explained by the
spatial segregation in the distribution and occupation of anurans in the different
phytophysiognomies of the Amapá savanna. Regarding the reproductive behavior of
anurans, 11 species were classified as having a long breeding season, intrinsically
associated with the rainy season and the reproductive mode of most species that lay egg
clutches in lentic water bodies. Six reproductive modes were recorded and parental care
was observed in Leptodactylus macrosternum and L. podicipinus, whose reproductive
mode is characterized by foam nests. Regarding behavioral reproductive strategies,
calling males were observed in all species of anurans, satellite males were recorded only
for D. walfordi, Hypsiboas multifasciatus, S. nebulosus and S. fuscomarginatus; active
search for females was observed for Phyllomedusa hypochondrialis and L. fuscus, and
male displacement was recorded only for Rhinella major and R. margaritifera. Of the
reproductive behaviors observed, throat and vocal sac display is associated with courtship
and territorial behavior exhibited by males. In addition to courtship behavior, visual
signals associated with courtship strategies were recorded for the anurans of the Amapá
savanna.
KEYWORDS: Anurans, Amazon savanna, Amapá, Composition, Richness, Community
structure, Reproductive behavior.

1
1. INTRODUÇÃO GERAL E REVISÃO DA LITERATURA
As formações abertas da América do Sul representam um papel crucial para a
conservação da biodiversidade, visto que são ambientes chaves para manutenção de
espécies tipicamente florestais em um contexto regional de paisagem savânica (Silva &
Bates 2002). Apesar da elevada riqueza de espécies e endemismos, espécies
compartilhadas com domínios vizinhos e algumas espécies amplamante distribuídas
(Colli et al. 2002; Valdujo et al. 2011), ainda existem consideráveis lacunas amostrais e
o número de estudos básicos que possam fornecer subsídios para a conservação, como
inventários faunísticos, é ainda muito pequeno.
O Cerrado brasileiro constitui um dos componentes de vegetação terrestre mais
importante do continente sul-americano, ocorrendo ao longo de mais de 200 de latitude
(Ratter et al. 1997; Silva & Bates 2002). É também o segundo maior Domínio
Fitogeográfico brasileiro (Klink & Machado 2005) e o quarto colocado, dentre os 34
hotspots de biodiversidade do planeta (Mittermeier et al. 2005); porém, nos últimos 35
anos, mais da metade da sua área foi transformada em diferentes usos e ocupações, tais
como pastagens e plantações de grãos (Klink & Machado 2005).
Neste contexto, Machado et al. (2004) reconheceu que cerca de 20% da vegetação
original de Cerrado se encontram preservadas, e apenas 1,2% está protegida em Unidades
de Conservação. Embora reduzido, o Cerrado se estende por uma grande área contínua
no Brasil Central e, de forma disjunta, em áreas descontínuas ao sul e ao norte do país
(Ribeiro & Walter 1998, 2008). As manchas disjuntas de Cerrado na floresta tropical
úmida dos Estados do Amapá, Amazonas, Pará e Roraima são reconhecidas como
Savanas amazônicas (Prance 1996, Sanaiotti et al. 1997, Barbosa et al. 2005) e fazem
parte da Ecorregião das Savanas das Guianas (Barbosa & Miranda 2005).
Pelo fato de serem constituídas por um conjunto diversificado de fitofisionomias,
formadas por mosaicos não-florestais (formações abertas) e florestais, associadas a
diferentes tipos de solo e clima, a Ecorregião das Savanas das Guianas é classificada de
forma diferente do Cerrado do Brasil Central, uma vez que possui vegetação aberta,
dominada pelo estrato herbáceo (ervas e capins), eventualmente com árvores e arbustos
em diferentes densidades (Barbosa et al. 2007, Barbosa & Campos 2011, Jati et al. 2014).
As Savanas amazônicas ocorrem por toda a bacia Amazônica, correspondendo a
aproximadamente 5% do total da Amazônia brasileira, ocupando uma área de 200.000
km2 (Cavalcante et al. 2014). O complexo Roraima-Rupununi, localizado no norte de

2
Roraima, sul da Venezuela e sudoeste da Guiana, é a maior área de Savana amazônica,
ocupando 42.706 km2 (Barbosa & Campos 2011). Outras áreas semelhantes as Savanas
são encontradas nos Estados do Pará (e.g. Alter do Chão e Serra do Cachimbo), Amazonas
(e.g. Campos de Humaitá), Rondônia (e.g. Áreas contínuas no sul do Estado), e Amapá
(e.g. Campos savânicos de Macapá, Porto Grande, Ferreira Gomes, Tartarugalzinho,
Pracuúba, Amapá, Calçoene, Oiapoque e Jari) (Ratter et al. 2006, Magnusson et al. 2008,
Rocha et al. 2014) (Figura 1).
Figura 1 – Distribuição das Savanas amazônicas no Brasil. 1. Alter do Chão; 2. Amapá; 3.
Roraima; 4. Humaitá; 5. Serra do Cachimbo; 6. Rondônia. Em hachurado: distribuição do Cerrado
do Brasil Central. Adaptado de Vieira et al. (2002).
As Savanas amazônicas do Estado do Amapá são, aparentemente, mais recentes
que outras formações que estão localizadas em antigos terrenos cristalinos do Cerrado do
Brasil Central (Rabelo et al. 2008). Embora a Savana amapaense possa conservar algumas
semelhanças com o Cerrado do Brasil Central, possui características particulares tais

3
como estrato herbáceo composto por representantes das famílias Poaceae, Cyperaceae e
Leguminosae (Sanaiotti et al. 1997, Rocha et al. 2014), atribuídas à sua história evolutiva
no âmbito dos regimes amazônicos, que definem vários tipos de mosaicos
fitofisionômicos, gerando uma grande complexidade e heterogeneidade de habitats.
Nesse contexto, as Savanas amazônicas são consideradas “refúgios” de uma
vegetação aberta que, provavelmente, se expandiu através do corredor amazônico durante
o Cenozóico (Haffer & Prance 2002). Apesar da sua importância para a biogeografia da
Amazônia, essas áreas têm sido mal amostradas em comparação com as florestas tropicais
da região amazônica, e os trabalhos sobre a anurofauna se concentraram em Savanas
Amazônicas dos estados brasileiros do Pará (Neckel-Oliveira et al. 2000, Pinheiro et al.
2012, Neckel-Oliveira et al. 2012) e de Roraima (Nascimento 2005, Azarak et al. 2011).
Apesar da extensão territorial que ocupam e da sua heterogeneidade, as Savanas
amazônicas são áreas pouco estudadas do ponto de vista de suas comunidades de anuros
e, apesar da fragmentação, nenhuma constitui Unidade de Conservação da Amazônia,
fatos que justificam estudos substanciais sobre sua anurofauna e consequente
identificação de prioridades para conservação (Azevedo-Ramos & Galatti 2002). Em
adição, o fato das Savanas amazônicas constituírem ambientes historicamente
pressionados por atividade antrópica (Sugai et al. 2014, Bitar et al. 2014), reforça a
importância da realização de inventários.
A complexidade e a heterogeneidade encontradas nas diversas fitofisionomias
abertas e fechadas das Savanas amazônicas, bem como a influência dos biomas vizinhos,
revelam a existência de padrões de distribuição geográfica, diversidade, riqueza e
abundância de espécies, particularmente de anfíbios anuros, vinculadas a essas áreas
(Souza & Eterovick 2011, Fahr & Kalko 2011). Além disso, dados da literatura sugerem
que as savanas com formações abertas tendem a apresentar riqueza e abundâncias mais
altas do que formações florestais (Colli et al. 2002, Maffei et al. 2011, Valdujo et al. 2011,
Conte et al. 2013).
O conhecimento sobre a composição da anurofauna das formações abertas da
Amazônia é também fundamental para a compreensão dos padrões de distribuição e
evolução dos anuros na América do Sul (Werneck et al. 2011, Werneck et al. 2012). Nas
Savanas amazônicas, por estarem isoladas da distribuição contínua do Cerrado, poucos
estudos foram realizados na tentativa de entender a distribuição das espécies de anuros,
bem como as características destas comunidades. A sazonalidade e a estrutura de
comunidades de anuros podem estar correlacionadas com a estrutura da vegetação e

4
disponibilidade de corpos d’água (Neckel-Oliveira et al. 2000), variações climáticas
(Azarak et al. 2011), distribuição espacial (Pinheiro et al. 2012) e período reprodutivo
(Azarak et al. 2011, Pinheiro et al. 2012, Bitar et al. 2012).
Dentre os estudos realizados em áreas de Savanas amazônicas podem ser
destacados: Neckel-Oliveira et al. (2000) sobre diversidade e distribuição de anuros em
Alter do Chão, Estado do Pará; Nascimento (2005) sobre a herpetofauna de áreas abertas
de Roraima; Azarak et al. (2011) sobre a composição, habitat, reprodução e dieta de
anuros do lavrado de Roraima; Pinheiro et al. (2012) e Neckel-Oliveira et al. (2012)
listaram as espécies de anfíbios anuros da Floresta Nacional de Carajás, Estado do Pará;
Bitar et al. (2012) estudaram a composição de espécies e os modos reprodutivos de anuros
na Amazônia Meridional; Sugai et al. (2014) sobre a composição de espécies de anuros
de uma área semelhante à Savana amazônica no município de Camapuã, Estado de Mato
Grosso do Sul e Bitar et al. (2014) que, em estudos realizados com comunidades de anuros
em área de transição entre a floresta Amazônica e a Savana, sugeriram que a conversão
de ambientes florestais em ambientes abertos pode resultar na substituição de espécies e
na variação da composição de espécies entre os habitats.
A anurofauna atualmente conhecida para as Savanas amazônicas é composta por
18 espécies, listadas a partir de inventários em Alter do Chão, Estado do Pará (Neckel-
Oliveira et al. 2000) e de 16 espécies em áreas abertas do lavrado do Estado de Roraima
(Azarak et al. 2011). Levando em consideração que grande parte das Savanas amazônicas
não foi adequadamente amostrada e que novos registros de espécies e de distribuição de
ocorrência nestas áreas são frequentes, a real riqueza de espécies de anuros para as
Savanas amazônicas pode estar subestimada.
No que se refere à ecologia de comunidades de anuros, diversos fatores são
responsáveis pela estruturação das comunidades e têm sido abordados sob diferentes
aspectos. Estudos ecológicos podem fornecer informações sobre as interações entre as
populações e/ou comunidades, bem como a forma que estas podem ser influenciadas por
processos abióticos e bióticos (competição e predação), detectando a influência relativa
de fatores históricos e contemporâneos na distribuição e regulação da riqueza e
diversidade de anuros (Toledo et al. 2007, Harrison & Cornell 2008, Thomé et al. 2010).
A coexistência de espécies em uma determinada comunidade é possibilitada, entre
outros fatores, pela partilha dos recursos disponíveis (Prado & Pombal Jr. 2005) que, por
sua vez, é determinada tanto por fatores ecológicos quanto históricos (Valdujo et al.
2013). As dimensões de nicho espacial (e.g. espécies que exploram habitats ou

5
microhabitats distintos), temporal (ativas em diferentes períodos do dia) e trófico (com
dieta diferente) são consideradas os principais descritores das relações entre as espécies,
determinando a estruturação das comunidades (Schoener 1974, Pianka 1994).
O nicho espacial é um fator determinante da estrutura de comunidades de anuros
e a coexistência entre as espécies pode resultar do uso diferenciado de habitats ou
microhabitats para vocalização, reprodução e desenvolvimento da fase larval (Kopp et al.
2010, Melo et al. 2013). A heterogeneidade dos habitats ou microhabitats exerce grande
influência na variação da riqueza e composição de espécies; em ambientes heterogêneos
que oferecem uma maior quantidade de recursos disponíveis e abrigam um maior número
de espécies (Bastazini et al. 2007, Vasconcelos et al. 2009), é esperada uma maior
segregação (Vasconcelos & Rossa-Feres 2008).
Nas comunidades de anuros, o nicho temporal é um importante componente na
segregação das espécies (Hartel et al. 2011, Maffei et al. 2011) e na sua estruturação
(Valdujo et al. 2013). A distribuição temporal é fortemente influenciada por processos
abióticos tais como a pluviometria e a temperatura, sendo esses, os principais fatores que
determinam os períodos de atividades dos anuros (Pirani et al. 2013). Por apresentarem
forte associação com a pluviosidade e a temperatura (Kopp et al. 2010), diversas espécies
de anuros apresentam respostas semelhantes a uma combinação diferente de variáveis
ambientais (Saenz et al. 2006), podendo ocorrer sobreposição no turno de vocalização
(Both et al. 2008) e na sua distribuição sazonal (Bastazini et al. 2007).
A dieta é uma dimensão importante do nicho trófico, que fornece subsídios para a
descrição de interações ecológicas entre os anuros e pode ser partilhada pelas espécies de
uma taxocenose (Menin et al. 2005). Contudo, esta dimensão de nicho também promove
grande sobreposição entre as espécies, tendo em vista que, de forma geral, os anuros são
considerados predadores oportunistas em relação aos recursos alimentares (Toft 1985).
Como a dieta é um aspecto fundamental do nicho ecológico, a estrutura da comunidade é
baseada na partilha de recursos alimentares entre as espécies coexistentes (Sabagh &
Carvalho-e-Silva 2008).
A segregação das espécies nas comunidades pode ocorrer em função das
diferenças na utilização das dimensões de nicho, possibilitando a redução de interações
interespecíficas, como a competição e a predação, permitindo assim, que uma maior
variedade de espécies consiga coexistir (Ernst & Rödel 2006, Paula 2012). No entanto, a
coexistência é possível não só porque as espécies apresentam padrões distintos quanto à
ocupação de habitats e microhabitats e à utilização dos recursos ambientais, mas porque

6
podem apresentar ocorrência sazonal diferenciada (Costa et al. 2013), com sua própria
composição de indivíduos entre as espécies e com uma história evolutiva e filogenética
distintas (Araújo et al. 2013).
Os anuros que ocorrem na região Neotropical podem ser divididos em dois grupos
ecológicos: um grande grupo de espécies restritas a florestas tropicais e um grupo menor
de espécies típicas de formações abertas, que ocorrem nos domínios fitogeográficos do
Cerrado, Chaco e Caatinga (Valdujo 2011). A anurofauna do Cerrado compreende um
grande número de espécies endêmicas (Bastos 2007, Valdujo et al. 2012), espécies
compartilhadas com domínios da diagonal de formações abertas e algumas espécies de
ampla distribuição (Colli et al. 2002, Araújo et al. 2009).
A distribuição da riqueza total de espécies e da riqueza de espécies endêmicas de
anuros no Cerrado do Brasil Central é heterogênea, e, apesar do esforço de amostragem
ser historicamente mais intenso nas regiões sul e sudoeste (Diniz-Filho et al. 2008,
Valdujo 2011), outras localidades bem amostradas em diferentes partes do Cerrado
diferem substancialmente em composição e riqueza de espécies (Valdujo et al. 2009,
2012, Araújo et al. 2013). Além da heterogeneidade espacial e ambiental do Cerrado
(Vasconcelos et al. 2009, Silva et al. 2011), essas diferenças na composição e na riqueza
podem ter sido favorecidas pelo contato com quatro grandes domínios fitogeográficos da
América do Sul: Amazônia, Mata Atlântica, Caatinga e Chaco (Joly et al. 1999).
De acordo com Souza & Eterovick (2011), a heterogeneidade ambiental registrada
nas formações florestais e abertas do Cerrado, bem como a influência dos domínios
fitogeográficos adjacentes, permite a coexistência de um maior número de espécies do
que ambientes homogêneos, por disponibilizar uma diversidade de habitats e
microhabitats, que é um fator determinante dos padrões de distribuição geográfica,
riqueza e abundância das espécies de anuros (Colli et al. 2002).
Estudos sobre ecologia de comunidade de anuros realizados no Cerrado do Brasil
Central evidenciaram que a segregação das espécies ocorre principalmente quanto ao
nicho temporal (Giaretta et al. 2008, Oda et al. 2009, Maffei et al. 2011, Paula 2012). A
pluviosidade e a temperatura são fatores promotores do início das atividades de
vocalização e reprodução das espécies, ficando as características ambientais como fatores
secundários. Adicionalmente, a ausência de estrutura na comunidade em função da
heterogeneidade ambiental (nicho espacial) pode ser atribuída a partilha de recursos,
diminuindo assim, a competição e a predação, consideradas de importância primária na
estruturação das comunidades (Crump 1982).

7
Em áreas de Savanas amazônicas, os estudos realizados com comunidades de
anuros abordaram apenas a riqueza e a composição de espécies, inexistindo estudos
ecológicos sobre a estruturação das comunidades. Quanto a riqueza e a composição
destacam-se aqueles realizados por Neckel-Oliveira et al. (2000), Nascimento (2005),
Neckel-Oliveira et al. (2012), Pinheiro et al. (2012) e Sugai et al. (2014).
Entender como as comunidades de anuros estão estruturadas e os mecanismos que
agem sobre a utilização do habitat e do microhabitat pelas espécies, os itens alimentares
que fazem parte da dieta e o período de atividade têm gerado subsídios para a formulação
de hipóteses sobre a evolução e a estrutura de comunidades (Pianka 1994). Recentemente,
fatores históricos têm sido considerados importantes para explicar a estrutura de
comunidades (Webb et al. 2002, Valdujo et al. 2012) uma vez que espécies relacionadas
filogeneticamente são associadas a fatores ecológicos semelhantes. Dessa forma, a
estruturação de comunidades não é influenciada somente pelos fatores ecológicos
contemporâneos, mas também por fatores históricos e restrições filogenéticas (Buckley
& Jetz 2007, 2008).
No que se refere ao comportamento reprodutivo das espécies de anuros, a
coexistência de diversas populações em um mesmo habitat é facilitada por divergências
ecológicas, especialmente na distribuição espacial e temporal das comunidades (Wells
1977, 2007). As taxocenoses de anuros são excelentes modelos por formarem agregados
conspícuos durante os eventos reprodutivos (Scott & Woodward 1994). A coexistência
entre diferentes espécies nas agregações reprodutivas é comumente explicada pela
partilha espacial, temporal e acústica (Silva et al. 2008, Kopp et al. 2010, Costa et al.
2013).
Embora a maioria dos anuros da região Neotropical apresentem atividade
reprodutiva sazonal, a distribuição e abundância de água, principalmente na forma de
chuva é o fator extrínseco primário no controle dos padrões, estratégias e modos
reprodutivos e comportamentos das espécies (Prado et al. 2005, Gambale et al. 2014). A
sazonalidade reprodutiva de algumas espécies de anuros também tem uma função chave
nas interações ecológicas e na estruturação das comunidades (Canavero et al. 2008, Oda
et al. 2009, Wachlevski et al. 2014).
A grande variedade de comportamentos reprodutivos exibidos pelas diferentes
espécies de anuros podem envolver complexas comunicações vocais (Hartmann et al.
2006, Costa & Toledo 2013), visuais (Hödl & Amézquita 2001, Hartmann et al. 2005,
Miranda et al. 2008), químicas e táteis (Wells 1977). A descrição de comportamentos

8
como corte (Costa et al. 2006, Muniz et al. 2008, Kokubum & Souza 2008, Delia et al.
2010, Lantyer-Silva et al. 2014) e cuidado parental (Santos & Amorim 2006, Vargas-
Salinas 2014), não são incomuns, mas ainda estão longe de serem conhecidos para todas
as espécies de anuros.
O comportamento reprodutivo dos anuros pode ser categorizado de acordo com
os padrões temporais anuais, como proposto por Wells (1977): 1) anuros com reprodução
prolongada; 2) anuros com reprodução explosiva. Esses padrões temporais influenciam
na organização social, na competição entre os machos por parceiras reprodutivas e nas
estratégias reprodutivas adotadas pelas populações de anuros. Na reprodução prolongada,
a estação reprodutiva é longa, a densidade de machos é relativamente baixa e a chegada
das fêmeas é assincrônica ao longo do tempo; a reprodução explosiva é caracterizada pela
curta duração do evento reprodutivo, alta densidade de machos resultando em estratégias
de procura ativa (Wells 1979) e competição, chegada sincrônica das fêmeas aos locais de
reprodução e baixa oportunidade de escolha de parceiros (Wells 1977, 2007).
As estratégias reprodutivas em anuros derivaram de uma combinação de atributos
morfológicos, fisiológicos e comportamentais, adaptados a condições ambientais (Toledo
et al. 2012). Quanto às estratégias reprodutivas comportamentais, dois tipos básicos são
reconhecidos: macho vocalizador e estratégias reprodutivas alternativas (Howard 1978).
Das estratégias reprodutivas, o macho vocalizador é a mais comum, na qual o macho
através da vocalização atrai fêmeas para a reprodução. Já as estratégias reprodutivas
alternativas, são divididas nas estratégias: de procura ativa por fêmeas, de macho
deslocador e de macho satélite (Pombal Jr. & Haddad 2007).
Os modos reprodutivos são uma combinação de caracteres que incluem o local da
desova, as características dos ovos e da desova, a duração dos estágios de
desenvolvimento, o estágio e o tamanho dos girinos e o tipo de cuidado parental (Salthe
& Duellman 1973, Crump 1974, Pombal Jr. & Haddad 2007). Os anuros da região
Neotropical apresentam a maior diversidade de modos reprodutivos (Haddad & Prado
2005), devido à existência de espécies que apresentam modos mais generalizados, com
os ovos depositados diretamente na água, e de espécies que apresentam modos de
reprodução mais especializados, inclusive em ambientes terrestres (Pombal Jr. & Haddad
2005, 2007, Toledo et al. 2012).
Para o Cerrado do Brasil Central, o número de modos reprodutivos é baixo,
ocorrendo apenas nove, com predomínio de modos generalizados (Kopp et al. 2010, Bitar
et al. 2012, Conte et al. 2013). De acordo com Hödl (1990), em estudo sobre a diversidade

9
dos modos reprodutivos de anuros amazônicos, espécies de formações abertas possuem
modos generalizados com desovas aquáticas ou em ninho de espuma e, espécies de
formações florestais, apresentam modos especializados. Dessa forma, uma pequena
variedade de modos é esperada em formações abertas em relação a outros domínios
fitogeográficos do país (Silva et al. 2012), uma vez que a diversificação e especialização
dos modos reprodutivos tem relação mais intrínseca com as características do ambiente
do que com as relações filogenéticas (Duellman & Trueb 1994).
Apesar da diversidade de padrões, estratégias e modos reprodutivos registrados
para a anurofauna do Cerrado do Brasil Central (Costa et al. 2006, Kopp et al. 2010,
Maffei et al. 2011, Conte et al. 2013, Gambale et al. 2014), existem poucos estudos,
principalmente sob o ponto de vista do comportamento reprodutivo, de modo particular
para as espécies de áreas de Savana amazônica. Nessas áreas, os únicos estudos que
abordam a reprodução e os modos reprodutivos estão contextualizados em Azarak et al.
(2011) e Bitar et al. (2012).
Nesse contexto, é reconhecida variação considerável na composição da
anurofauna entre as localidades e as fitosionomias das Savanas amazônicas; no entanto,
em função da carência de estudos ecológicos, especialmente sobre comportamento
reprodutivo, justifica-se uma amostragem sistemática para a Savana, localizada na
Amazônia Oriental. Especificamente, esta Tese tem como objetivos estudar a
composição, ecologia de comunidades e comportamento reprodutivo de anfíbios anuros
de área de Savana amapaense.
2. METODOLOGIA GERAL
2.1. CARACTERIZAÇÃO DA ÁREA DE ESTUDO
A área estudada é parte da Savana amapaense, que ocupa uma estreita faixa
longitudinal de 9.861,89 Km2, correspondente a 6,9% da superfície territorial do Estado
do Amapá (Rabelo et al. 2008). Ocupando uma superfície geomorfológica Terciária que
forma a transição entre os setores costeiros do Holoceno, que são dominados por
manguezais e florestas de várzea, e os setores do Pré-Cambriano, revestidos
principalmente por florestas densas de terra firme, a Savana amapaense se estende
paralelamente à costa atlântica, desde a região do município do Laranjal do Jarí até o alto
curso do rio Uaçá (Azevedo 1967, Rabelo et al. 2008).

10
O trabalho de campo foi realizado no Campo Experimental da Savana amapaense
da Empresa Brasileira de Pesquisa Agropecuária (00°023’5” N, 51°02’2” W), localizado
a 48 km ao Norte de Macapá, ocupando uma área 1.381 ha e perímetro de 15.080 m
(Melém-Júnior et al. 2003) (Figura 2).
Figura 2 – Campo Experimental da Savana amapaense da Empresa Brasileira de Pesquisa
Agropecuária. Fonte: Secretaria Estadual do Meio Ambiente do Estado do Amapá (2014).
O tipo climático predominante é Am, de acordo com a classificação Köppen
(Alvares et al. 2013) caracterizado como clima tropical chuvoso com nítido período seco.
Devido à concentração das chuvas em seis meses seguidos (janeiro a julho), o clima ao
longo do ano pode ser tipicamente reconhecido em apenas duas estações: uma estação
seca bem evidente (4.6 mm) e uma estação chuvosa com precipitação pluviométrica
elevada (4.827 mm). Com relação à temperatura média mensal, a mínima é de 24,4o C e
a máxima de 28,4o C (Figura 3).

11
Figura 3 – Médias mensais da pluviosidade (Barras) e das temperaturas mínima (linha cinza) e
máxima (linha tracejada) do Campo Experimental da Savana amapaense da EMBRAPA durante
os anos de 2009, 2010 e 2013. Fonte: Núcleo de Hidrometeorologia e Energia Renováveis do
Instituto de Pesquisas Científicas e Tecnológicas do Estado do Amapá (NHMET/ IEPA).
O relevo é plano ou suavemente ondulado, sobre solos que podem ocorrer sob
duas formas principais: os Latossolos e as Areias quartzosas. O domínio da Savana ocorre
sobre dois tipos básicos de unidades de relevo: planaltos cristalinos ou sedimentares e
depressões interplanálticas. Em geral, nas regiões de planaltos, predominam a savana
gramíneo-lenhosa e a savana gramíneo lenhosa arbustiva, enquanto que nas depressões
interplanáticas são mais comuns a savana parque e a savana arbórea (Aguiar et al. 2004,
Barbosa et al. 2005).
O domínio fitofisionômico da Savana amapaense é caracterizado por dois estratos
principais, um arbóreo e outro rasteiro, enquanto os hábitats são divididos em dois grupos
principais: hábitats ripários e hábitats de interflúvio (Ribeiro & Walter 2008). Nos
hábitats de interflúvio ocorre o predomínio de formações abertas englobando a Savana
gramíneo lenhosa, gramíneo lenhosa-arbustiva, parque e arbórea (Figura 4).

12
Figura 4 – Desenho esquemático das Fitofisionomias amostradas no Campo Experimental da
Savana amapaense da EMBRAPA: A – Savana gramíneo lenhosa; B – Savana gramíneo lenhosa-
arbustiva; C – Savana parque; D – Savana arbórea. Adaptado de Ribeiro & Walter (1998).
O estudo em campo foi efetuado em quatro fitofisionomias do Campo
Experimental da Savana amapaense da EMBRAPA (Figura 5); a definição de cada
fitofisionomia segue Miranda et al. (2003):
Savana gramíneo-lenhosa, caracterizada por extensos campos de gramíneas e
estrato herbáceo abundante, com estrato lenhoso disperso e composto por árvores anãs
esparsas (< 1,0 m);

13
Savana gramíneo-lenhosa arbustiva, caracterizada por estrato herbáceo coberto
por gramíneas em grande densidade e estrato lenhoso com a presença irregular de
pequenas árvores esparsas (< 3,8 m);
Savana parque, apresenta estrato herbáceo menos desenvolvido e cobertura
arbóreo-arbustiva relativamente baixa (< 4,3 m) e densa formando um dossel
descontínuo;
Savana arbórea, caracterizada pela ausência de estrato herbáceo e pela presença
de árvores com alturas superiores a 2 m em grande densidade, formando um dossel
contínuo.
Figura 5 – Vegetação das fitofisionomias do Campo Experimental da Savana amapaense da
EMBRAPA: A – Savana gramíneo lenhosa; B – Savana gramíneo lenhosa-arbustiva; C – Savana
parque; D – Savana arbórea.
2.2. PROCEDIMENTOS METODOLÓGICOS
Para a amostragem dos anfíbios anuros foram realizadas excursões mensais de
janeiro de 2009 a dezembro de 2010 (Capítulo 1 e 2), totalizando 24 excursões para
observações e/ou coletas em cada fitofisionomia, sendo 12 no período chuvoso e 12 no

14
período seco; para as análises do comportamento reprodutivo de espécies de anuros na
Savana amapaense (Capítulo 3) foram feitas observações durante o período de janeiro a
dezembro de 2013, com duração de quatro dias consecutivos, totalizando 288
horas.homem.
Foram estabelecidas 20 unidades amostrais em cada fitofisionomia (Savana
gramíneo lenhosa, Savana gramíneo lenhosa-arbustiva, Savana parque e Savana arbórea),
dispostas de acordo com a disponibilidade de corpos d’água, totalizando 80 unidades
amostrais. Cada unidade amostral foi representada por uma parcela de 100x50 metros
(0,5 ha), equidistantes pelo menos 500 metros, conforme metodologia descrita em Bitar
et al. (2012). Cabe destacar que o esforço amostral foi o mesmo nas quatro
fitofisionomias, possibilitando comparações. Cada fitofisionomia foi examinada por
quatro pesquisadores e, em cada noite, visitada uma parcela de 100x50 metros (0,5 ha)
(Figura 6).
Figura 6 – Delineamento amostral dos anfíbios anuros e variáveis ambientais da Savana
amapaense. (M – meses de amostragem; SGL – Savana gramíneo lenhosa; SGLA – Savana
gramíneo lenhosa-arbustiva; SP – Savana parque; SA – Savana arbórea).

15
As amostragens dos anuros foram feitas através de busca ativa e auditiva (cf.
Heyer et al. 1994). Inicialmente, os horários de atividade e amostragem dos anuros foram
ajustados através de coletas piloto, visando identificar turnos e picos de vocalização.
Como a maioria das espécies apresentou horários de atividade mais amplos, iniciando a
vocalização no começo da noite, por volta das 18:00h e com diminuição as 23:30h, e
poucas espécies apresentaram horários restritos a apenas algumas horas, estabeleceu-se o
horário de observações e/ou coletas entre 18:00h e 24:00h.
Cada parcela foi amostrada por quatro pesquisadores, caminhando lentamente
lado a lado por pelo menos 4 horas, entre 18:00h a 24:00h. A cada 10 m
aproximadamente, os observadores paravam por cerca de dois minutos e registravam o
número de indivíduos de cada espécie e machos em atividade de vocalização (cf.
Zimmerman 1994) e, ao mesmo tempo, procuravam visualmente na vegetação
circundante. Para este método, o esforço de coleta foi calculado pela multiplicação do
número de horas no campo pelo número de pesquisadores (horas.homem).
Os anuros coletados foram acondicionados separadamente em sacos plásticos. No
momento do registro, algumas informações como, fitofisionomia da Savana/habitat onde
o espécime foi primeiramente avistado, se o animal estava em atividade de vocalização,
horário, posição em relação ao sítio de vocalização (paralelo, vocalizando para cima,
vocalizando para baixo, diagonal), substrato utilizado (solo, água, estrato herbáceo,
arbóreo, arbustivo), altura em relação ao poleiro e a superfície da água e distância do sítio
de vocalização até a margem do corpo d’água.
Depois de coletados, os anuros foram mortos com anestésico (lidocaína 5%),
numerados e fixados de acordo com metodologia proposta por Heyer et al. (1994). Os
espécimes foram coletados de acordo com a Licença N0. 18243-1, expedida pelo Sistema
de Autorização e Informação em Biodiversidade – SISBIO/ICMBio. O material
testemunho está depositado na Coleção de Fauna do Instituto de Pesquisas Científicas e
Tecnológicas do Estado do Amapá e na Coleção Herpetológica da Universidade Federal
do Rio Grande do Norte (UFRN).
O detalhamento dos métodos utilizados para analisar a riqueza e composição de
espécies, ecologia de comunidades e o comportamento reprodutivo dos anuros da Savana
amapaense estão descritos nos capítulos correspondentes.

16
CAPÍTULO 1 ARTIGO SUBMETIDO AO PERIÓDICO AMPHIBIA-REPTILIA (QUALIS B1)
RIQUEZA E COMPOSIÇÃO DE UMA TAXOCENOSE DE ANUROS
EM ÁREA DE SAVANA AMAZÔNICA
Carlos Eduardo Costa-Campos1,2*; Eliza Maria Xavier Freire3
1 Universidade Federal do Amapá, Departamento de Ciências Biológicas e da Saúde,
Laboratório de Herpetologia, Rod. Juscelino Kubitschek, km 02, Jardim Marco Zero,
CEP 68.902-280, Macapá, AP, Brasil. 2 Universidade Federal do Rio Grande do Norte, Programa de Pós-Graduação em
Psicobiologia, Lagoa Nova, CEP 59.072-970, Natal, RN, Brasil.
3 Universidade Federal do Rio Grande do Norte, Departamento de Botânica, Ecologia e
Zoologia, Laboratório de Herpetologia, Lagoa Nova, CEP 59072-970, Natal, RN, Brasil.
* Corresponding author. E-mail: [email protected]
1. RESUMO
As Savanas amazônicas ocupam aproximadamente 200.000 km2 da Amazônia brasileira,
ocorrendo em manchas isoladas dispersas por extensas áreas de floresta nos Estados do
Amapá, Amazonas, Pará e Roraima. Apesar de haver considerável variação na
composição da anurofauna entre localidades e entre fitofisionomias das savanas, uma
amostragem sistemática e geograficamente ampla ainda não havia sido realizada para a
savana amapaense localizada no Norte do Brasil, Amazônia Oriental. Nessa perspectiva,
foi estudada uma área de savana no Estado do Amapá quanto à riqueza, composição,
diversidade e abundância de espécies. Este estudo foi realizado no Campo Experimental
da Empresa Brasileira de Pesquisa Agropecuária (00°023’5” N, 51°02’2” W). Para as
amostragens dos anuros, nós realizamos 24 coletas mensais em cada fitofisionomia
(savana gramíneo-lenhosa, savana gramíneo-lenhosa arbustiva, savana parque e savana
arbórea), através de busca ativa e auditiva ao longo de 20 parcelas de 100x50 metros em
cada fitofisionomia. Foram registradas 21 espécies de anuros pertencentes a 4 famílias:
Bufonidae (3 spp.); Hylidae (11); Leptodactylidae (6) e Microhylidae (1). Quanto à
composição de espécies por fitofisionomia, para a savana gramíneo-lenhosa arbustiva
registrou-se um maior número de indivíduos. A curva de rarefação de espécies para a área
total atingiu uma assintota, sugerindo que o esforço de coleta de dados foi suficiente para
amostrar satisfatoriamente a riqueza de espécies da área. A análise de variância de
Kruskal-Wallis revelou diferenças significativas entre a riqueza e a diversidade de
espécies entre as fitofisionomias (p<0,05). A análise de similaridade de Bray-Curtis
separou as fitofisionomias em três grupos principais: savana arbórea, savana gramíneo-
lenhosa e gramíneo-lenhosa arbustiva e savana parque. Através do Ordenamento por
escalonamento não-métrico multidimensional (NMDS), a estrutura de comunidade de
anuros da savana resultou em uma separação entre três fitofisionomias amostradas, com
diferenças significativas (ANOSIM, R = 0,823; p < 0,001) e com nível de stress de 0,07,
indicando diferença na estrutura de comunidades das três fitofisionomias amostradas. A
riqueza local (21 espécies) corresponde a 14 % das 150 espécies típicas que possuem
distribuição fortemente associada ao Cerrado do Brasil Central e 35,6% das 59 espécies
típicas de domínios vizinhos e que ocorrem apenas marginalmente no Cerrado,
representando uma parte considerável da riqueza de espécies de anuros registradas na
savana da porção Oriental da Amazônia brasileira.
Palavras-chave: Amapá, Amazônia Oriental, Comunidades de Anuros.

17
2. ABSTRACT
The Amazonian savannas occupy approximately 200.000 km2 of the Brazilian Amazon,
taking place in scattered isolated patches over large areas of forest in the states of Amapá,
Amazonas, Pará and Roraima. Despite having considerable variation in the composition
of Anurans between locations and between the savannas physiognomies, a systematic and
geographically wide sampling has not been performed for the savanna from Amapá yet,
located in the northern of Brazil, Eastern Amazonia. In this perspective, there were studies
made on the richness, composition, diversity and abundance of anurans species, an area
of savanna in Amapá State, specifically in the Experimental Field of the Brazilian
Agricultural Research Agency (00°023’5”N; 51°02’2”W). For sampling of anurans, we
performed 24 samples in each physiognomies (grassland savanna, scrub grassland
savanna, parkland savanna, open woodland savanna), through active and auditive search
over 20 parcels of 100x50 meters in each physiognomies. It was registered 21 species of
frogs belonging to 5 families: Bufonidae (3 spp.), Hylidae (11), Leptodactylidae (6) e
Microhylidae (1). Regarding the species richness by physiognomies, scrub grassland
savanna registered a greater number of individuals. The species rarefaction curve for the
total area has reached an asymptote, suggesting that the data collection effort was enough
to adequately sample the species richness of the area. The Kruskal-Wallis variance
analysis revealed significant differences on the species richness and diversity among the
physiognomies (p<0.05). The Bray-Curtis similarity analysis grouped the physiognomies
into three main groups: open woodland savanna, grassland savanna and scrub grassland
savanna and parkland savanna. Through ordering by non-metric multidimensional scaling
(NMDS), the species composition from the savanna anuran community resulted in a
separation among three physiognomies sampled, with significant differences (ANOSIM,
R = 0.823, p<0.001) and level of stress of 0.07, indicating differences in community
composition of the three physiognomies sampled. The local richness (21 species)
corresponds to 14% out of the15 typical species that have distribution strongly associated
with the Cerrado from Central Brazil and 35.6% out of 59 typical species of neighboring
domains and which occur only marginally in the Cerrado, representing a considerable
part of frog species richness recorded in the savanna of eastern portion of the Brazilian.
Key-Words: Amapá, Eastern Amazon, Frog Communities.
3. INTRODUÇÃO
As Savanas amazônicas ocupam normalmente áreas planas, cobertas por uma
vegetação aberta constituída por estrato herbáceo dominado por gramíneas, apresentando
árvores e arbustos dispersos em diferentes densidades de cobertura (Rabelo et al. 2008).
As savanas ocupam aproximadamente 200.000 km2 da Amazônia brasileira (Cavalcante
et al. 2014), ocorrendo em manchas isoladas dispersas por extensas áreas de floresta nos
Estados do Amapá, Amazonas, Pará e Roraima (Sanaiotti et al. 1997, Magnusson et al.
2008). O maior bloco contínuo de savanas do extremo norte da Amazônia é o complexo
Roraima-Rupununi, localizado no norte de Roraima, sul da Venezuela e no sudoeste da
Guiana, possuindo cerca de 43.000 km2 (Barbosa et al. 2007, Barbosa & Campos 2011).

18
A complexidade e a heterogeneidade encontradas nas diferentes fitofisionomias
das savanas (Eiten 1972, Coutinho 1978), bem como a influência dos biomas vizinhos,
demonstram a existência de padrões de distribuição geográfica, diversidade, riqueza e
abundância das espécies habitantes nessas áreas (Neckel-Oliveira et al. 2000). No que se
refere aos estudos faunísticos, dada a grande diversidade dos anfíbios anuros, a
quantidade de estudos sobre ecologia de comunidade vem aumentando ao longo dos anos
abordando diferentes aspectos, tais como, os mecanismos que agem sobre a seleção e o
uso dos hábitats e os fatores responsáveis pela manutenção da diversidade e a própria
estrutura de comunidades (Brasileiro et al. 2005, Prado et al. 2005, Santos et al. 2009,
Araújo et al. 2013, Sugai et al. 2014).
Nesse contexto, apesar da extensão territorial e da heterogeneidade, as savanas
amazônicas são áreas extremamente desconhecidas do ponto de vista de suas
comunidades de anuros, não constituem Unidades de Conservação e estão altamente
ameaçadas pela expansão da atividade humana; portanto é premente a realização de
inventários sobre a anurofauna (Neckel-Oliveira et al. 2000, Brasileiro et al. 2008,
Pinheiro et al. 2012, Bitar et al. 2014). A forte pressão antrópica e a presença de espécies
endêmicas de anuros em áreas de savana, são outros fatores que justificam a importância
destes inventários (Sugai et al. 2014).
Apesar da considerável variação na composição da anurofauna entre localidades
e entre as fitofisionomias da savana (Neckel-Oliveira et al. 2000, Bitar et al. 2012), uma
amostragem sistemática e geograficamente ampla ainda não havia sido realizada para a
savana da Amazônia Oriental. Nessa perspectiva, foi efetuado inventário de anuros da
área de savana do Amapá, para conhecer a composição, riqueza e diversidade ao longo
de diferentes fitofisionomias. Para averiguar a existência de padrões nas savanas, a
composição de espécies locais foi comparada a de áreas de savanas amazônicas da região
de Alter do Chão e de Carajás, Estado do Pará, além de outras localidades bem amostradas
do Cerrado no Brasil Central.
4. MATERIAL E MÉTODOS
4.1. Área de estudo
A savana amapaense ocupa uma estreita faixa longitudinal de 9.861,89 Km2, o
que corresponde a 6,9% da superfície territorial do estado do Amapá (Rabelo et al. 2008).
Situada desde a cidade de Macapá, se estende na direção norte do Estado até a cidade de
Calçoene, num percurso de 374 km. Nesta área, especificamente no Campo Experimental

19
da Empresa Brasileira de Pesquisa Agropecuária – EMBRAPA (00°023’5” N, 51°02’2”
W), foi realizado este estudo; a área total da savana é 1.381 ha e perímetro de 15.080 m
(Melém-Júnior et al. 2003).
Foram amostradas quatro fitofisionomias do Campo Experimental da
EMBRAPA, seguindo descrição proposta por Miranda et al. (2003): savana gramíneo
lenhosa, caracterizada por extensos campos de gramíneas e estrato herbáceo abundante,
com estrato lenhoso disperso e composto por árvores anãs esparsas (< 1,0 m); savana
gramíneo-lenhosa arbustiva, com estrato herbáceo coberto por gramíneas em grande
densidade e estrato lenhoso com presença irregular e pequenas árvores esparsas (< 3,8
m); savana parque, com estrato herbáceo menos desenvolvido e cobertura arbóreo-
arbustiva relativamente baixa (< 4,3 m) e densa formando um dossel descontínuo e,
savana arbórea, com ausência de estrato herbáceo e árvores com maior densidade,
formando um dossel contínuo (> 2,0 m).
O tipo climático predominante é Am, de acordo com a classificação Köppen
(Alvares et al. 2013) caracterizado como clima tropical chuvoso com nítido período seco.
Devido à concentração das chuvas em seis meses seguidos (janeiro a junho), o clima ao
longo do ano pode ser tipicamente reconhecido em apenas duas estações: uma estação
seca bem evidente (4.6 mm) e uma estação chuvosa com precipitação pluviométrica
elevada (4.827 mm). Com relação à temperatura média mensal, a mínima é de 24,4o C e
a máxima de 28,4o C.
O relevo é plano ou suavemente ondulado, sobre solos que podem ocorrer sob
duas formas principais: os Latossolos e as Areias quartzosas. O domínio da savana ocorre
sobre dois tipos básicos de unidades de relevo: planaltos cristalinos ou sedimentares e
depressões interplanálticas. Em geral, nas regiões de planaltos, predominam a savana
gramíneo-lenhosa e a savana gramíneo lenhosa arbustiva, enquanto que nas depressões
interplanáticas são mais comuns a savana parque e a savana arbórea (Aguiar et al. 2004,
Barbosa et al. 2005).
4.2. Procedimentos metodológicos
4.2.1. Trabalho em campo
Para a amostragem dos anfíbios anuros da Savana amapaense foram realizadas 24
excursões em cada fitofisionomia, sendo 12 no período chuvoso e 12 no período seco.
Foram estabelecidas 20 unidades amostrais em cada fitofisionomia (Savana gramíneo-
lenhosa, Savana gramíneo-lenhosa arbustiva, Savana parque e Savana arbórea), dispostas
de acordo com a disponibilidade de corpos d’água. Cada unidade amostral foi

20
representada por uma parcela de 100 x 50 metros (0,5ha), distantes pelo menos 500
metros entre si, conforme sugerido por Bitar et al. (2012). Nessas unidades amostrais
foram realizados os métodos de busca ativa e acústica (Crump & Scott 1994, Heyer et al.
1994, Zimmerman 1994).
A busca visual ativa e auditiva foi efetuada em cada parcela por quatro
pesquisadores, das 18:00 as 24:00h. A cada 10 m percorridos foram anotados o número
de indivíduos de cada espécie e em atividade de vocalização. Em ambos os métodos, o
esforço amostral foi calculado pela multiplicação do número de horas no campo pelo
número de pesquisadores participantes da coleta (horas.homem).
Os anuros coletados foram acondicionados separadamente em sacos plásticos. No
momento do registro, algumas informações como, fitofisionomia da savana/hábitat onde
o espécime foi primeiramente avistado, se o animal estava em atividade de vocalização,
horário, posição em relação ao sítio de vocalização (paralelo, perpendicular, vocalizando
para cima, vocalizando para baixo, diagonal), substrato utilizado (solo, água, estrato
herbáceo, arbóreo, arbustivo), altura em relação ao poleiro e a superfície da água e
distância do sítio de vocalização até a margem do corpo d’água.
As variáveis climatológicas pluviometria mensal (mm), temperatura mínima e
máxima (°C) e umidade relativa (%) foram obtidas do Núcleo de Hidro-Metereologia e
Energia Renováveis do Instituto de Pesquisas Científicas e Tecnológicas do Estado do
Amapá (NHMET/IEPA).
Depois de coletados, os animais foram mortos com anestésico (lidocaína 5%),
numerados e fixados em formol e conservados de acordo com Heyer et al. (1994). Os
espécimes foram coletados de acordo com a Licença N0. 18243-1, expedida pelo Sistema
de Autorização e Informação em Biodiversidade – SISBIO/ICMBio. O material
testemunho está depositado na Coleção de Fauna do Instituto de Pesquisas Científicas e
Tecnológicas do Estado do Amapá e na Coleção Herpetológica da Universidade Federal
do Rio Grande do Norte (UFRN).
4.2.2. Análise dos dados
A diversidade dos anuros da área total foi calculada por meio do Índice de
Diversidade de Shannon-Winner (Krebs 2014). Nós também utilizamos este índice para
averiguar a existência de diferenças de diversidade de espécies entre as quatro
fitofisionomias da Savana amapaense.

21
𝐻′ = ∑(𝑝𝑖)𝑙 = 1 (ln 𝑝𝑖)
O Índice de Equitabilidade foi calculado através da razão entre a diversidade
obtida e a diversidade máxima. Este índice mostra a homogeneidade da população ou
como as espécies estão representadas por número de indivíduos de cada espécie dentro
da comunidade (Magurran 2004). Este índice varia de 0, quando uma espécie se sobressai
em abundância, a 1, quando as espécies são igualmente abundantes no ambiente.
𝐽 =𝐻′
𝐻𝑚𝑎𝑥′
Para analisar a riqueza de espécies de anuros da Savana amapaense e das
fitofisionomias foram construídas curvas de rarefação de espécies baseadas no número de
indivíduos e no número de amostras (cf. Gotelli & Colwell 2001) utilizando-se o
programa EstimateS 9.1 (Colwell 2013) com 1.000 aleatorizações. A riqueza de espécies
foi comparada pelos estimadores de riqueza Chao 1, Chao 2, Jacknife 1, Jacknife 2 e
Bootstrap no programa EstimateS 9.1 (Colwell 2013).
As possíveis diferenças nas variações da riqueza, diversidade e equitabilidade nas
diferentes fitofisionomias foram verificadas através de Análise de variância de Kruskal-
Wallis e Teste de Dunn a posteriori (Zar 2010). Todas as análises foram realizadas através
dos Programas “BioEstat 5.0 (Ayres et al. 2007) e PAST 2.09 (Hammer et al. 2001). Os
valores foram considerados significativos para p < 0,05.
Para verificar se a composição das comunidades diferia entre as fitofisionomias,
foi realizada uma Análise de agrupamento (Cluster analysis) e de Ordenamento
Escalonamento Multidimensional Não-Métrico (NMDS). As diferenças entre a
composição das espécies e as fitofisionomias foram avaliadas através de uma Análise de
Similaridade ANOSIM (Clarke 1993). Para detectar quais as espécies de anuros são
responsáveis pelas diferenças entre os grupos foi utilizada a Análise de Quebra de
Porcentagem da Similaridade (SIMPER), considerando o Índice de Similaridade de Bray-
Curtis.
Onde j = equitabilidade; H’ = diversidade obtida; Hmax’ =
diversidade máxima.
Onde H' = índice de diversidade de espécies; pi = Ni/N = probabilidade
que um indivíduo pertença à espécie i de um total de "S" espécies; Ni =
número total de indivíduos da espécie i; N = número total de espécies.

22
5. RESULTADOS E DISCUSSÃO
5.1. Riqueza e Composição de espécies
Em 1.920 horas.homem de esforço amostral foram encontradas 21 espécies de
anfíbios anuros pertencentes às famílias: Bufonidae (3 espécies); Hylidae (11);
Leptodactylidae (6) e Microhylidae (1) (Tabela 1, Figura 1). Das espécies registradas na
savana amapaense, dois hilídeos (Dendropsophus walfordi e Scinax fuscomarginatus),
um leptodactilídeo (Pseudopaludicola boliviana) e um microhilídeo (Elachistocleis
helianneae) representam novos registros de ocorrência para o Estado do Amapá.
Tabela 1 – Espécies de anfíbios anuros registradas para uma área de Savana amapaense estudada
durante o período de Janeiro de 2009 a Dezembro de 2010.
Família /Espécies
Bufonidae
Rhinella major (Müller & Hellmich, 1936)
Rhinella margaritifera (Laurenti, 1768)
Rhinella marina (Linnaeus, 1758)
Hylidae
Dendropsophus walfordi (Bokermann, 1962)
Hypsiboas multifasciatus (Günther, 1859)
Hypsiboas punctatus (Schneider, 1799)
Hypsiboas raniceps Cope, 1862
Osteocephalus taurinus Steindachner, 1862
Scinax fuscomarginatus (A. Lutz, 1925)
Scinax nebulosus (Spix, 1824)
Scinax ruber (Laurenti, 1768)
Scinax x-signatus (Spix, 1824)
Trachycephalus typhonius (Linnaeus, 1758)
Phyllomedusa hypochondrialis (Daudin, 1800)
Leptodactylidae
Pseudopaludicola boliviana Parker, 1927
Adenomera hylaedactyla (Cope, 1868)
Leptodactylus fuscus (Schneider, 1799)
Leptodactylus macrosternum Miranda-Ribeiro, 1926
Leptodactylus pentadactylus (Laurenti, 1768)
Leptodactylus podicipinus (Cope, 1862)
Microhylidae
Elachistocleis helianneae Caramaschi, 2010

23
Figura 1 – Anfíbios anuros registrados na Savana amapaense durante o período de Janeiro de
2009 a Dezembro de 2010. A = R. major, B = R. margaritifera, C = R. marina, D = D. walfordi,
E = H. multifasciatus, F = H. punctatus, G = H. raniceps, H = O. taurinus, I = S. fuscomarginatus,
J = S. nebulosus, K = S. ruber, L = S. x-signatus, M = T. typhonius, N = P. hypochondrialis, O =
P. boliviana, P = A. hylaedactyla, Q = L. fuscus, R = L. macrosternum, S = L. pentadactylus, T
= L. podicipinus, U = E.helianneae.

24
A riqueza registrada para a área estudada está aquém da riqueza de 43 espécies
conhecidas para localidades bem amostradas no Cerrado (Brasileiro et al. 2005,
Vasconcelos & Rossa-Feres 2005, Vitt et al. 2005, Santos et al. 2007, Brasileiro et al.
2008, Ribeiro-Júnior & Bertoluci 2009, Araújo & Almeida-Santos 2011) e de 26 espécies
em Savanas amazônicas (Neckel-Oliveira et al. 2000, Azarak et al. 2011, Pinheiro et al.
2012, Bitar et al. 2012, Neckel-Oliveira et al. 2012, Sugai et al. 2014).
A riqueza da área de Savana estudada na porção Oriental local corresponde a 14%
das 150 espécies que possuem distribuição fortemente associada ao Cerrado do Brasil
Central e 35,6% das 59 espécies típicas de domínios vizinhos e que ocorrem apenas
marginalmente no Cerrado (Valdujo et al. 2012) representando 21 espécies de anuros
registradas na savana da porção Oriental da Amazônia brasileira.
A anurofauna da Savana amapaense é caracterizada principalmente pelas famílias
Hylidae e Leptodactylidae, um padrão comumente encontrado na região Neotropical
(Brasileiro et al. 2005, Kopp & Eterovick 2006, Bertoluci et al. 2007, Canelas & Bertoluci
2007, Vaz-Silva et al. 2007, Ribeiro-Júnior & Bertoluci 2009, Sugai et al. 2014). A
família Hylidae representa mais da metade das espécies registradas neste estudo (52,4%)
apesar de Brasileiro et al. (2005), destacar que a riqueza de hilídeos em áreas abertas do
Cerrado representar 46,4% da comunidade.
A riqueza de espécies de anuros da Savana amapaense variou nas diferentes
fitofisionomias, ou seja, as áreas abertas (e.g. Savana gramíneo lenhosa) apresentaram
maior riqueza, seguidas pelas áreas florestais (e.g. Savana parque e arbórea). Estes dados
concordam com os da literatura, os quais sugerem que as áreas abertas tendem a
apresentar riquezas mais altas quando comparadas as áreas florestadas do Domínio do
Cerrado (Maffei et al. 2011, Conte et al. 2013). Trabalhos realizados por Valdujo et al.
(2011) encontraram riquezas de anuros maiores nas áreas abertas do cerrado do que nas
matas de galeria, matas ciliares e cerradões. Isso se deve ao fato de que a reprodução da
maioria das espécies de Cerrado ocorre em áreas abertas, e, como a disponibilidade de
sítios reprodutivos é um dos fatores mais importantes na seleção do hábitat para anuros,
é esperado maior riqueza de espécies e abundância de indivíduos em fitofisionomias
abertas (Colli et al. 2002, Bastazini et al. 2007).
Por outro lado, em trabalhos realizados por Neckel-Oliveira et al. (2000) em uma
área de Savana amazônica no município de Santarém, e por Pinheiro et al. (2012) na
região de Carajás, ambos no Estado do Pará, foram constatadas maior riqueza e
abundância de espécies de anuros em ambientes florestados. Diversos estudos relatam

25
que espécies de ambientes abertos podem utilizar áreas florestadas como locais de
forrageio, migração ou refúgio (Silva & Rossa-Feres 2007, Brassaloti et al. 2010).
Nesse sentido, padrões na estrutura da vegetação das fitofisionomias exercem
grande influência na distribuição dos anuros, limitando ou alterando sua riqueza (Ernst et
al. 2006, Enst & Rödel 2008). Werner et al. (2007) observou que comunidades de anuros
de áreas abertas podem ser fortemente influenciadas pela cobertura do dossel, onde áreas
com maior cobertura apresentaram menor riqueza de espécies. Outro fator que pode
interferir na riqueza de espécies de anuros são os modos reprodutivos, pois impõem
restrições à utilização das fitofisionomias (Haddad & Prado 2005).
5.2. Eficiência amostral: Curva de rarefação e Estimadores de riqueza de
espécies
A curva de rarefação de espécies para área total atingiu a assíntota, sugerindo que
o esforço de coleta de dados foi suficiente para amostrar satisfatoriamente a riqueza de
espécies da área de estudo (Figura 2); o mesmo ocorreu para análise efetuada para cada
uma das fitofisionomias estudadas (Figura 3).
Figura 2 – Curva de rarefação de espécies baseada no número de indivíduos por espécie para a
área total da Savana amapaense durante os anos de 2009 e 2010.

26
Figura 3 – Curvas de rarefação de espécies para cada fitofisionomia amostrada: A) savana
gramíneo lenhosa; B) savana gramíneo lenhosa-arbustiva; C) savana parque e D) savana arbórea.
Segundo Magurran (2004) e Santos (2006), as curvas de rarefação são excelentes
métodos para avaliar a eficiência de inventários, por estimar todas as espécies de
determinados locais ou habitats. Semelhante à rarefação para área total, as curvas de
rarefação de espécies para todas as fitofisionomias também se estabilizaram. No entanto,
para a savana arbórea, estabilizou em uma riqueza de espécies relativamente baixa, o que
indica que o esforço de amostra adicional seria necessário nos ambientes florestais.
Neste contexto, três fatores não mutuamente exclusivos e associados à estrutura
da cobertura vegetal podem ser responsáveis pelos diferentes padrões riqueza e
composição de espécies de anuros observados nas fitofisionomias estudadas: 1) a
capacidade de algumas espécies em colonizar ambientes caracterizados por vegetação
mais esparsa; 2) tolerâncias fisiológicas distintas das espécies em relação à temperatura
da água, a intensidade luminosa e a umidade próxima à superfície do solo; e 3)
dependência de microhabitats específicos para a reprodução.
As características particulares de cada fitofisionomia em relação a estes fatores,
possivelmente determinam e limitam a distribuição das espécies presentes nas
fitofisionomias da Savana amapaense, uma vez que diferentes espécies de anuros
respondem diferencialmente às condições ambientais, apresentando diferentes taxas de
crescimento e desenvolvimento, além de exibirem graus distintos de resistência e
tolerância à perda de água (Halverson et al. 2003). A seleção de habitats reprodutivos

27
associada às limitações impostas por modos reprodutivos específicos pode restringir a
utilização de um determinado habitat (Haddad & Prado 2005, Pombal Jr. & Haddad
2007), como é o caso de R. margaritifera, registrada apenas na Savana arbórea e P.
hypochondrialis e L. podicipinus, registrados na Savana gramíneo lenhosa arbustiva.
Apesar da curva acumulativa de espécies apresentar tendência a estabilização, não
é descartada a possibilidade de incremento na riqueza local, mas com a continuidade do
estudo, o aumento de esforço contribuiria muito lentamente para a adição do aumento da
riqueza de espécies, conforme evidenciado pelos estimadores de riqueza. Isso evidencia
a importância da realização de inventários com a associação de diferentes métodos de
amostragem (e.g. armadilhas de intercepção e queda) para um conhecimento mais
completo da anurofauna (Greenberg et al. 1994, Brown 1997, Maritz et al. 2007, Ribeiro-
Júnior et al. 2008), pois a riqueza de especies é estreitamente relacionada com o esforço
da amostragem (Ugland & Gray 2004, Silveira et al. 2010).
5.3. Diversidade e Equitabilidade de espécies
A diversidade de espécies obtida para a área estudada foi de H’=2,728. Por outro
lado, os índices de diversidade entre as fitofisionomias amostradas variaram de H’=0,814
a H’=2,459 e de equitabilidade variaram de J = 0,587 a J = 1,887 (Tabela 2). A análise
de variância de Kruskal-Wallis revelou diferenças significativas entre a riqueza de
espécies (H = 38,38; gl = 3; p = 0,001), diversidade (H = 35,98; gl = 3; p = 0,001) e
equitabilidade (H = 15,92; gl = 3; p = 0,001).
Tabela 2 – Riqueza, Diversidade (H’) e Equitabilidade (J) de anuros amostrados nas
fitofisionomias da Savana amapaense durante os anos de 2009 e 2010.
Fitofisionomias Riqueza H’ J
Savana gramíneo lenhosa 13 2,276 0,8874
Savana gramíneo lenhosa-arbustiva 19 2,459 0,8353
Savana parque 8 1,000 1,8874
Savana arbórea 4 0,814 0,5873
Área total 21 2,728 0,8824
Quanto à diversidade por fitofisionomia, constatou-se a maior diversidade na
savana gramíneo lenhosa arbustiva. A alta diversidade registrada nesta fitofisionomia
corrobora com trabalhos realizados por Brasileiro et al. (2005), Araujo et al. (2009),
Araujo & Almeida-Santos (2011), Valdujo et al. (2011) em estudos comparativos de

28
diversidade de espécies de áreas abertas com formações campestres, savânicas e florestais
em regiões do Cerrado do Brasil Central.
Os índices de riqueza de espécies, diversidade e equitabilidade apresentaram
diferenças significativas entre as fitofisionomias amostradas (Tabela 3). Este resultado
era esperado, em função das fitofisionomias apresentarem diferenças na heterogeneidade
ambiental, conforme defendido por Vasconcelos et al. (2009, 2010). A heterogeneidade
ambiental possui um papel importante na determinação da riqueza de espécies e na
estruturação das comunidades de anuros (Huston 1994, Buckley & Jetz 2007), oferecendo
ambientes e condições diversas para espécies com diferentes requisitos ecofisiológicos
(Valdujo et al. 2011).
Tabela 3 – Análise de variância de Kruskal-Wallis e Teste de Dunn a posteriori entre a riqueza
de espécies, diversidade e equitabilidade das fitofisionomias da Savana amapaense. SGL =
Savana gramíneo lenhosa; SGLA = Savana gramíneo lenhosa-arbustiva; SP = Savana parque; SA
= Savana arbórea. Resultados significativos estão em negrito (p < 0,05).
Fitofisionomias Riqueza H’ J
SGL vs. SGLA H = 19,17; p = 0,001 H = 16,25; p = 0,001 H = 4,20; p = 0,040
SGLA vs. SP H = 21,59; p = 0,001 H = 20,76; p = 0,001 H = 4,25; p = 0,039
SGLA vs. SA H = 30,24; p = 0,001 H = 29,41; p = 0,001 H = 18,57; p = 0,001
Adicionalmente, vários estudos têm demonstrado que ambientes heterogêneos e
complexos favorecem a coexistência de um maior número de espécies quando
comparados a ambientes homogêneos (Conte & Rossa-Feres 2007, Santos et al. 2007,
Vasconcelos et al. 2009, Silva et al. 2011). Além da heterogeneidade ambiental registrada
no Cerrado (Silva et al. 2011), diferenças em sua composição e riqueza de espécies entre
várias localidades amostradas devem ter sido favorecidas pelo contato com quatro
grandes domínios fitogeográficos da América do Sul: Amazônia, Mata Atlântica,
Caatinga e Chaco (Joly et al. 1999).
5.4. Análise de Similaridade (Cluster analysis) e Ordenamento (NMDS)
A análise de similaridade a partir do índice de Bray-Curtis separou as
fitofisionomias em três grupos principais: (A) Savana arbórea, (B) Savana parque e (C)
Savana gramíneo lenhosa e Savana gramíneo lenhosa-arbustiva. O grupo (A) é
caracterizado pela ocorrência de R. margaritifera. O grupo (B) é caracterizado pela alta
frequência de ocorrência de H. punctatus. E o grupo (C), as espécies mais frequentes
foram P. hypochondrialis e L. fuscus (Figura 4).

29
Figura 4 – Dendrograma de similaridade de Bray-Curtis A linha pontilhada corta o demdrograma
em três clusters, representando as três fitofisionomias da Savana amapaense. A (Savana arbórea
– SA); B (Savana parque – SP) e C (Savana gramíneo lenhosa – SGL e Savana gramíneo lenhosa-
arbustiva – SGLA).
A análise de similaridade (SIMPER) mostrou que a similaridade média dentro das
fitofisionomias da Savana amapaense foi de 27,5% para a Savana gramíneo lenhosa,
48,8% para a Savana gramíneo lenhosa-arbustiva, 58,7% Savana arbórea e 47,0% para a
Savana parque. As espécies que mais contribuíram para a similiaridade média dentro das
fitofisionomias foram: Leptodactylus fuscus (53,6%, Savana gramíneo lenhosa);
Phyllomedusa hypochondrialis (26,7%, Savana gramíneo lenhosa-arbustiva); Hypsiboas
punctatus (81,8%, Savana parque) e, R. margaritifera (88,4%, Savana arbórea).
A dissimilaridade média das comunidades entre as fitofisionomias foi maior entre:
Savana arbórea e Savana parque (99,5 %) e, menor entre a Savana gramíneo lenhosa e a
Savana gramíneo lenhosa-arbustiva (75,2 %). As espécies que mais contribuíram para a
dissimilaridade média entre as fitofisionomias foram: Rhinella margaritifera (32,0%,
Savana arbórea e Savana parque); Hypsiboas punctatus (21,7%, Savana gramíneo lenhosa
e Savana parque) e, Phyllomedusa hypochondrialis (17,4%, Savana gramíneo lenhosa e
Savana gramíneo lenhosa-arbustiva).
A contribuição expressiva das espécies R. margaritifera, H. punctatus e P.
hypochondrialis na dissimilaridade média entre as fitofisionomias pode ser explicada pela
existência de variáveis ambientais na estruturação das comunidades de anuros estudada,

30
determinando uma distribuição não aleatória ao longo dos gradientes formados pelas
fitofisionomias da Savana amapaense. Um fator importante que pode explicar a influência
da estrutura do ambiente nas comunidades de anuros é a disponibilidade de nichos,
influenciando, desta forma, nos padrões de coexistência e diversidade das espécies da
comunidade estudada (Werner et al. 2007, Oliveira & Eterovick 2010).
Através do NMDS construído a partir da matriz de similaridade de Bray-Curtis,
ficou demonstrado que a comunidades de anfíbios anuros da área estudada está distribuída
entre as três fitofisionomias amostradas, com diferenças significativas (ANOSIM, R =
0,823; p < 0,001) e com nível de stress de 0,07. Desta forma, as comunidades formaram
grupos distintos no NMDS, indicando diferença na composição das comunidades das três
fitofisionomias (Figura 5).
Figura 5 – Ordenação por Escalonamento Multidimensional Não-Métrico (NMDS) das
fitofisionomias da área estudada, gerado a partir da matriz de similaridade de Bray-Curtis. SGL
= Savana gramíneo lenhosa; SGLA = Savana gramíneo lenhosa-arbustiva; SA = Savana arbórea;
SP = Savana parque.
O índice de Bray-Curtis obtido apresentou agrupamentos que refletem diferenças
encontradas entre as principais fitofisionomias amostradas e revelou uma maior
similaridade entre a Savana gramíneo lenhosa e a Savana gramíneo lenhosa-arbustiva.
Ambas as fitofisionomias diferem quanto à composição florística das espécies arbóreas

31
dominantes, substrato herbáceo e arbustivo, e topografia (Batalha 2011); provavelmente,
estas diferenças tiveram influência nos resultados obtidos. Além disso, áreas
geograficamente próximas, geralmente apresentam características estruturais
semelhantes, as quais dão suporte a comunidades semelhantes em sua riqueza e
composição de espécies (Dixo & Verdade 2006). Mesmo não favorecendo espécies mais
exigentes quanto ao habitat, as fitofisionomias Savana parque e Savana arbórea, com
menor complexidade estrutural, são relativamente favoráveis na estruturação das
comunidades de anuros, contribuindo para a preservação da diversidade regional em áreas
de mosaicos vegetacionais (Gardner et al. 2006).
A composição da comunidade de anuros parece influenciada por fatores
ecológicos históricos e contemporâneos, incluindo heterogeneidade ambiental (Buckley
& Jetz 2007), história evolutiva (Seymour et al. 2001, Smith et al. 2007) e disponibilidade
de água (Oliveira & Diniz-Filho 2010). Estes exemplos ilustram a importância dos fatores
históricos e comtemporâneos em escala local, determinados pela dinâmica espacial e
temporal das fitofisionomias da Savana amapaense. Deste modo, a composição e a
riqueza local dos anuros é condicionada tanto pelas interações ecológicas das espécies
entre os diferentes tipos de fitofisionomias, como pelas interações históricas,
determinadas pelas barreiras e corredores utilizados para a dispersão das espécies.
Os padrões de distribuição geográfica das espécies ocorrentes na Savana
amapaense mostram a importância do intercâmbio faunístico com regiões adjacentes para
o incremento da riqueza local. O predomínio de espécies características do domínio
Cerrado e a existência de espécies de domínios adjacentes, principalmente
compartilhadas com o domínio Amazônico, revelam que a proximidade geográfica e a
dinâmica espacial histórica entre os dois domínios favorecem a troca faunística, sobretudo
nas áreas abertas e florestais da Savana amapaense.
A presença de populações compartilhadas destaca a urgência em se considerar a
singularidade da Savana amapaense nos esforços conservacionistas. Igualmente, a
possibilidade de ocorrência prévia de extinções locais e a acelerada destruição dos
habitats, subsidiam a realização de inventários da anurofauna em áreas pouco amostradas
e o estabelecimento de corredores que conectem os remanescentes de áreas de Savana
amazônica. Neste contexto, devemos ressaltar a importância da realização de inventários
de longa duração, gerando bancos de dados confiáveis que sirvam de subsídios para
estudos que procuram explicar os padrões de distribuição de espécies, além de auxiliar na
tomada de decisões para políticas de conservação.

32
Adicionalmente, o conhecimento da composição e riqueza atual de anuros das
diferentes fitofisionomias da Savana amapaense, além do preenchimento de lacunas
amostrais, é de grande importância para a detecção e monitoramento de espécies sensíveis
ou tolerantes à antropização do ambiente, bem como para estudos biogeográficos que
possibilitem avaliar a existência de diferentes padrões de distribuição de espécies. Estas
informações são de grande importância pois, conforme os resultados obtidos no presente
estudo, são relevantes para planejamento da conservação, considerando que os anfíbios
estão entre os vertebrados mais ameaçados do planeta.
6. REFERÊNCIAS BIBLIOGRÁFICAS
Aguiar, L.M.S., Machado, R.B. & Marinho-Filho, J. 2004. A diversidade biológica do
Cerrado. In: Cerrado: ecologia e caracterização (Org. por L.M. Aguiar & A.J.A.
Camargo), pp.17-40. Brasília: Embrapa Cerrados.
Alvares, C.A., Stape, J.L., Sentelhas, P.C., Gonçalves, J.L.M. & Sparovek, G. 2013.
Köppen’s climate classification map for Brazil. Meteorologische Zeitschrift, 22 (6),
711-728.
Araújo, C.O. & Almeida-Santos, S.M. 2011. Herpetofauna de um remanescente de
Cerrado no estado de São Paulo, sudeste do Brasil. Biota Neotropica, 11 (3), 47-62.
Araújo, C.O., Condez, T.H. & Sawaya, R.J. 2009. Anfíbios Anuros do Parque Estadual
das Furnas do Bom Jesus, sudeste do Brasil, e suas relações com outras taxocenoses
no Brasil. Biota Neotropica, 9 (2), 77-98.
Araújo, C.O., Corrêa, D.T. & Almeida-Santos, S.M. 2013. Anuros da Estação
Ecológica de Santa Bárbara, um remanescente de formações abertas de Cerrado no
estado de São Paulo. Biota Neotropica, 13 (3), 230-240.
Ayres, M., Ayres Jr., M.A., Ayres, D.L. & Dos Santos, A.S. 2007. BioEstat: aplicações
estatísticas nas áreas das ciências biológicas e médicas. Belém: Publicações Avulsas
Mamirauá.
Azarak, P.A., Nascimento, S.P. &Carvalho, C.M. 2011. Anfíbios do Lavrado de
Roraima. Biologia Geral e Experimental, 11 (1), 4-14.
Barbosa, R.I., Nascimento, S.P., Amorim, P.A.F. & Silva, R.F. 2005. Notas sobre a
composição arbóreo-arbustiva de uma fisionomia das savanas de Roraima, Amazônia
Brasileira. Acta Botanica Brasilica, 19 (2), 323-329.

33
Barbosa, R.I., Campos, C., Pinto, F. & Fearnside, P.M. 2007. The “Lavrados” of
Roraima: Biodiversity and Conservation of Brazil’s Amazonian Savannas. Functional
Ecosystems and Communities, 1, 29-41.
Barbosa, R.I. & Campos, C. 2011. Detection and geographical distribution of clearing
areas in the savannas (‘lavrado’) of Roraima using Google Earth web tool. Journal of
Geography and Regional Planning, 4, 122-136.
Bastazini, C.V., Munduruca, J.V., Rocha, P.L.B. & Napoli, M.F. 2007. Which
environmental variables better explain changes in anuran community composition? A
case study in the Restinga of Mata de Sao Joao, Bahia, Brazil. Herpetologica, 63, 459-
471.
Batalha, M.A. 2011. O Cerrado não é um bioma. Biota Neotropica, 11 (1), 21-24.
Bertoluci, J., Brassaloti, R.A., Ribeiro-Júnior, J.W., Vilela, V.M.F.N. & Sawakuchi,
H.O. 2007. Species composition and similarities among anuran assemblages of forest
sites in southeastern Brazil. Scientia Agricola, 64 (4), 364-374.
Bitar, Y.O.C., Pinheiro, L.P.C., Abe, P.S. & Santos-Costa, M.C. 2012. Species
composition and reproductive modes of anurans from a transitional Amazonian forest,
Brazil. Zoologia, 29 (1), 19-26.
Bitar, Y.O.C., Juen, L., Pinheiro, L.P.C. & Santos-Costa, M.C. 2014. Anuran Beta
Diversity in a Mosaic Anthropogenic Landscape in Transitional Amazon. Journal of
Herpetology, 48 (4), 000-000.
Brasileiro, C.A., Sawaya, R.J., Kiefer, M.C &, Martins, M. 2005. Amphibians of a
cerrado fragment in Southeastern Brazil. Biota Neotropica, 5 (2), 93-109.
Brasileiro, C.A., Lucas, L.M., Oyamaguchi, H.M., Thomé, M.T.C. & Dixo, M. 2008.
Anurans, Northern Tocantins River Basin, states of Tocantins and Maranhão, Brazil.
Check List, 4 (2), 185-197.
Brassaloti, R.A., Rossa-Feres, D.C. & Bertoluci, J. 2010. Anurofauna da Floresta
Estacional Semidecidual da Estação Ecológica dos Caetetus, sudeste do Brasil. Biota
Neotropica, 10 (1), 275-291
Brown, L.J. 1997. An Evaluation of Some Marking and Trapping Techniques Currently
Used in the Study of Anuran Population Dynamics. Journal of Herpetology, 31, 410-
419.
Buckley, L. & Jetz, B.W. 2007. Environmental and historical constraints on global
patterns of amphibian richness. Proceedings of the Royal Society, 274, 1167-1173.

34
Canelas, M.A.S. & Bertoluci, J. 2007. Anurans of the Serra do Caraça, southeastern
Brazil: Species composition and fenological patterns of calling activity. Iheringia,
Série Zoologia, 97 (1), 21-26.
Cavalcante, C.O., Flores, A.S. & Barbosa, R.I. 2014. Fatores edáficos determinando a
ocorrência de leguminosas herbáceas em savanas amazônicas. Acta Amazônica, 44 (3),
379-386.
Clarke, K.R. 1993. Non-parametric multivariate analysis of changes in community
structure. Austral Ecology, 18, 117-143.
Colli, G.R., Bastos, R.P. & Araújo, A.F.B. 2002. The character and dynamics of the
Cerrado herpetofauna. In: The Cerrados of Brazil: Ecology and Natural History of a
Neotropical Savanna. (Org. por P.S.Olivera & R.J.Marquis), pp. 223-241. New York:
Columbia University Press.
Colwell, R.K. 2013. EstimateS: Statistical estimation of species richness and shared
species from samples. Version 9.1. Encontrado em 10 de Julho de 2013,
http://purl.oclc.org/estimates.
Conte, C.E. & Rossa-Feres, D.C. 2007. Riqueza e distribuição espaço-temporal de
anuros em um remanescente de Floresta de Araucária no sudeste do Paraná. Revista
Brasileira de Zoologia, 24 (4), 1025-1037.
Conte, C.E., Silva, D.R. & Rodrigues, A.P. 2013. Anurofauna da bacia do Rio Tijuco,
Minas Gerais, Brasil e sua relação com taxocenoses de anfíbios do Cerrado e suas
transições. Iheringia, 103 (3), 280-288.
Coutinho, L.M. 1978. O conceito de cerrado. Revista Brasileira de Botânica, 1: 17-23.
Crump, M.L. & Scott, N.J.J. 1994. Visual encounter surveys. In: Measuring and
monitoring biological diversity: standard methods for amphibians (Org. por W.R.
Heyer., M.A. Donnelly., R.W. McDiarmid., L.A.C. Hayek. & M.S. Foster), pp. 84-92.
Washington, USA: Smithsonian Institution Press.
Dixo, M. & Verdade, V.K. 2006. Herpetofauna de serrapilheira da Reserva Biológica
de Morro Grande, Cotia (SP). Biota Neotropica, 6 (2), 1-20.
Eiten, G. 1972. The Cerrado vegetation of Brazil. The Botanical Review, 38, 201-341.
Ernst, R., Linsenmair, E. & Rödel, M. 2006. Diversity erosion beyond the species level:
dramatic loss of functional diversity after selective logging in two tropical amphibians
communities. Biological Conservation, 133: 143-155.

35
Ernst, R. & Rödel, M. 2008. Patterns of community in two tropical tree frog
assemblages: Separating spatial structure and environmental effects in disturbed and
undisturbed forest. Journal of Tropical Ecology, 24, 111-120.
Gardner, T.A., Ribeiro-Júnior, M.A., Barlow, J., Ávila-Pires, T.C.S., Hoogmoed,
M.S. & Peres, C.A. 2006. The Value of Primary, Secundary, and Plantation Forest
for a Neotropical Herpetofauna. Conservation Biology, 21 (3), 775-787.
Gotelli, N. & Colwell, R.K. 2001. Quantifying biodiversity: Procedures and pitfalls in
the measurement and comparison of species richness. Ecology Letters, 4, 379-391.
Greenberg, C.H., Neary, D.G. & Harris, L.D. 1994. A comparison of herpetofaunal
sampling effectiveness of pitfall, single-ended, and double-ended funnel traps with
drift fences. Journal of Herpetology, 28, 319-324.
Haddad, C.F.B. & Prado, C.P.A. 2005. Reproductive modes in frogs and their
unexpected diversity in the Atlantic Forest of Brazil. BioScience, 55, 207-217.
Halverson M.A., Skelly D.K., Kiesecker J.M. & Freidenburg L.K. 2003. Forest
mediated light regime linked to amphibian distribution and performance. Oecologia,
134, 360-364.
Hammer, O., Harper, D.A.T. & Ryan, P.D. 2001. PAST: Paleontological Statistics
software package for education and data analysis. Version 2.01. Paleontologia
Electronica 4. Encontrado em 3 de Setembro de 2010, http://palaeo-
electronica.org/2001_1/past/issue1_01.htm.
Heyer, W.R., Donnelly, M.A., Mcdiarmid, R.W., Hayek, L.C. & Foster, M.S. 1994.
Measuring and monitoring biological diversity: standard methods for amphibians.
Washington: Smithsonian Institution.
Houston, M.A. 1994. Biological Diversity. The coexistence of species changing
landscape. Cambridge: Cambridge University Press.
Joly, C.A., Aidar, M.P.M., Klink, C.A., Mcgrath, D.G., Moreira, A.G., Moutinho,
P., Nepstad, D.C., Oliveira, A.A., Pott, A., Rodal, M.J.N. & Sampaio, E.V.S.B.
1999. Evolution of the Brazilian phytogeography classification systems: implications
for biodiversity conservation. Ciência e Cultura, 51, 331-348.
Kopp, K. & Eterovick, P.C. 2006. Factors influencing spatial and temporal structure of
frog assemblages at ponds in southeastern Brazil. Journal of Natural History, 40 (29-
31), 1813-1830.
Krebs, C.J. 2014. Ecological methodology. 3a ed. Vancouver: University of British
Columbia.

36
Maffei, F., Ubaid, F.K. & Jim, J. 2011. Anurans in an open cerrado area in the
municipality of Borebi, Sao Paulo state, Southeastern Brazil: habitat use, abundance
and seasonal variation. Biota Neotropica, 11, 221-233.
Magnusson, W.E., Lima, P.A., Albernaz, A.L.K.M., Sanaiotti, T.M. & Guillaumet,
J.L. 2008. Composição florística e cobertura vegetal das savanas na região de Alter
do Chão, Santarém – PA. Revista Brasileira de Botânica, 31, 165-177.
Magurran, A.E. 2004. Measuring biological diversity. Oxford: Blackwell Science.
Maritz, B., Gavin, M., Mackay, D. & Alexander, G. 2007. The effect of funnel trap
type and size of pitfall trap on trap success: implications for ecological field studies.
Amphibia-Reptilia, 28: 321-328.
Melém-Júnior, N.J., Farias-Neto, J.T. & Yokomizo, G.K.I. 2003. Caracterização dos
Cerrados do Amapá. Comunicado Técnico Embrapa, 105: 1-5.
Miranda, I.S., Absy, M.L. & Rebêlo, G.H. 2003. Community Structure of Woody
Plants of Roraima Savannahs, Brazil. Plant Ecology, 164: 109-123.
Neckel-Oliveira, S., Magnusson, W.E. & Lima, A.P. 2000. Diversity and distribution
of frogs in an Amazonian savanna in Brazil. Amphibia-Reptilia, 21 (3), 317-326.
Neckel-Oliveira, S., Galatti, U., Gordo, M., Pinheiro, L.P.C. & Maschio, G.F. 2012.
Anfíbios. In: Fauna da Floresta Nacional de Carajás: Estudos Sobre Vertebrados
Terrestres (Org. por F.D. Martins., A.F. Castilho., J. Campos., F.M. Hatano &, S.
Rolim), pp. 67-79. São Paulo: Editora Rona.
Oliveira, F.F.R. & Eterovick, P.C. 2010. Patterns of Spatial Distribution and
Microhabitat Use by Syntopic Anuran Species along Permanent Lotic Ecosystems in
the Cerrado of Southeastern Brazil. Herpetologica, 66 (2), 159-171.
Oliveira, G. & Diniz-Filho, J.A.F. 2010. Spatial patterns of terrestrial vertebrates
richness in Brazilian semiarid, Northeastern Brazil: Selecting hypotheses and
revealing constraints. Journal of Arid Environments, 74, 1418-1426.
Pinheiro, L.C., Bitar, Y.O.C., Galatti, U., Neckel-Oliveira, S. & Santos-Costa, M.C.
2012. Amphibians from southeastern state of Pará: Carajás Region, northern Brazil.
Check List, 8 (4), 693-702.
Pombal Jr., J.P. & Haddad, C.F.B. 2007. Estratégias e modos reprodutivos em anuros.
In: Herpetologia no Brasil II (Org. por L.B. Nascimento., P.M.E. Oliveira), pp. 101-
116. Belo Horizonte: Sociedade Brasileira de Herpetologia.

37
Prado, C.P.A., Uetanabaro, M. & Haddad, C.F.B. 2005. Breeding activity patterns,
reproductive modes, and habitat use by anurans (Amphibia) in a seasonal environment
in the Pantanal, Brasil. Amphibia-Reptilia. 26, 211-221.
Rabelo, B.V., Pinto, A.C.; Simas, A.P.S.C., Tardin, A.T., Fernandes, A.V., Souza,
C.B., Monteiro, E.M.P.B., Facundes, F.S., Ávila, J.E.S., Souza, J.S.A., Costa-
Guedes, L.A., Penha, O.A.A., Melo, R.M.S. & Gibson, V.M. 2008.
Macrodiagnóstico do Estado do Amapá: primeira aproximação do ZEE. Macapá:
Instituto de Pesquisas Científicas e Tecnológicas do Estado do Amapá (IEPA).
Ribeiro-Júnior, M.A., Gardner T.A. & Ávila-Pires, T.C.S. 2008. Evaluating the
Effectiveness of Herpetofaunal Sampling Techniques across a Gradient of Habitat
Change in a Tropical Forest Landscape. Journal of Herpetology, 42 (4), 733-749.
Ribeiro-Júnior, J.W. & Bertoluci, J. 2009. Anurans of the cerrado of the Estação
Ecológica and the Floresta Estadual de Assis, southeastern Brazil. Biota Neotropica,
9 (1), 207-206.
Sanaiotti, T., Bridgewater, S. & Ratter, J. 1997. A floristic study of the savanna
vegetation of the state of Amapá, Brazil, and suggestions for its conservation. Boletim
do Museu Paraense Emílio Goeldi, Série Botânica, 13, 3-29.
Santos, A.J. 2006. Estimativas de riqueza em espécies. In: Métodos de estudos em
biologia da conservação e manejo da vida silvestre (Org. por L. Cullen Júnior., R.
Rudran & C. Valladares-Pádua), pp. 19-41. Curitiba: Editora UFPR.
Santos, P., Siqueira, C.C., Goyannes-Araújo, P., Fernandes, C.G.A., Rubião, E.C.N.
& Van Sluys, M. 2007. A Survey of the Leaf-Litter Frog Assembly from an Atlantic
Forest Area (Reserva Ecológica de Guapiaçu) in Rio de Janeiro State, Brazil, with an
estimate of frogs densities. Tropical Zoology, 20, 99-108.
Santos, T.G., Rossa-Feres, D.C. & Casatti, L. 2007. Diversidade e distribuição espaço-
temporal de anuros em região com pronunciada estação seca no sudeste do Brasil.
Iheringia, Série Zoologia, 97 (1), 37-49.
Santos, T.G., Vasconcelos, T.S., Rossa-Feres, D.C. & Haddad, C.F.B. 2009. Anurans
of a seasonally dry tropical forest: Morro do Diabo State Park, São Paulo State, Brazil.
Journal of Natural History, 43 (15-16), 973-993.
Seymour, C.L., De Klerk, H.M., Channing, A. & Crowe, T.M. 2001. The
biogeography of the Anura of sub-equatorial Africa and the prioritization of areas for
their conservation. Biodiversity and Conservation, 10, 2045-2076.

38
Silva, F.R. & Rossa-Feres, D.C. 2007. Uso de fragmentos florestais por anuros
(Amphibia) de área aberta na região noroeste do Estado de São Paulo. Biota
Neotropica, 7 (2), 141-148.
Silva, R.A., Martins, I.A. & Rossa-Feres, D.C. 2011. Environmental heterogeneity:
Anuran diversity in homogeneous environments. Zoologia, 28 (5), 610-618.
Silveira, L.F., Beisiegel, B.M., Curcio, F.F., Valdujo, P.H., Dixo, M., Verdade, V.K.,
Mattox, G.M.T. & Cunningham, P.T.M. 2010. Para que servem os inventários de
fauna? Estudos avançados, 68, 173-207.
Smith, S.A., Oca, A.N.M., Reeder, T.W. & Wiens, J.J. 2007. A phylogenetic
perspective on elevational species richness patterns in Middle American treefrogs.
Why so few species in lowland tropical rainforests? Evolution, 61, 1188-1207.
Sugai, J.L.M.M., Terra, J.S. & Ferreira, V.L. 2014. Anurans of a threatened savanna
area in western Brazil. Biota Neotropica, 14 (1), 1-9.
Ugland, K.I. & Gray, J.S. 2004. Estimation of species richness: analysis of the methods
developed by Chao and Karakassis. Marine Ecology Progress Series, 284, 1-8.
Valdujo, P.H., Camacho, A., Recoder, R.S., Júnior, M.T., Ghellere, J.M.B., Mott,
T., Nunes, P.M.S., Nogueira, C. & Rodrigues, M.T. 2011. Anfíbios da Estação
Ecológica Serra Geral do Tocantins, região do Jalapão, Estados do Tocantins e
Bahia. Biota Neotropica, 11 (1), 251-262.
Valdujo, P.H., Silvano, D.L., Colli, G.R. & Martins, M. 2012. Anuran Species
Composition and Distribution Patterns in Brazilian Cerrado, a Neotropical Hotspot.
South American Journal of Herpetology, 7 (2), 63-78.
Vasconcelos, T.S. & Rossa-Feres, D.C. 2005. Diversidade, distribuição espacial e
temporal de anfíbios anuros (Amphibia, Anura) na região Noroeste do Estado de São
Paulo, Brasil. Biota Neotropica, 5 (2), 1-14.
Vasconcelos, T.S., Santos, T.G., Rossa-Feres, D.C. & Haddad, C.F.B. 2009. Influence
of the environmental heterogeneity of breeding ponds on anuran assemblages from
southeastern Brazil. Canadian Journal Zoology, 87, 699-707.
Vasconcelos, T.S., Santos, T.G., Haddad, C.F.B. & Rossa-Feres, D.C. 2010. Climatic
variables and altitude as predictors of anuran species richness and number of
reproductive modes in Brazil. Journal of Tropical Ecology, 26, 423-432.
Vaz-Silva, W., Guedes, A.G., Azevedo-Silva, P.L., Gontijo, F.F., Barbosa, R.S.,
Aloísio G.R. & Almeida, F.C.G. 2007. Herpetofauna, Espora Hydroelectric Power
Plant, state of Goiás, Brazil. Check List, 3 (4), 338-345.

39
Vitt, L.J., Caldwell, J.P., Colli, G.R., Garda, A.A., Mesquita, D.O., França, F.G.R.,
Shepard, D.B., Costa, G.C., Vasconcellos, M.M. & Novaes e Silva. V. 2005. Uma
atualização do guia fotográfico dos répteis e anfíbios da região do Jalapão no Cerrado
brasileiro. Sam Noble Oklahoma Museum of Natural History, 2, 1-24.
Werner, E.E., Skelly, D.K., Relyea, R.A. & Yurewicz, K.L. 2007. Amphibian species
richness across environmental gradients. Oikos, 116, 1697-1712.
Zar, J. H. 2010. Biostatistical Analysis. 5a ed. New Jersey: Prentice Hall.
Zimmerman, B.L. 1994. Audio Strip Transects. In: Measuring and monitoring
biological diversity: standard methods for amphibians (Org. por W.R. Heyer., M.A.
Donnelly., R.W. McDiarmid., L.A.C. Hayek & M.S. Foster), pp. 92-97. Washington:
Smithsonian Institution Press.

40
CAPÍTULO 2
ARTIGO A SER SUBMETIDO AO PERIÓDICO ZOOLOGIA (QUALIS B1)
ECOLOGIA DE COMUNIDADES DE ANUROS EM DIFERENTES FITOFISIONOMIAS
DE UMA ÁREA DE SAVANA AMAZÔNICA
Carlos Eduardo Costa-Campos1,2*; Eliza Maria Xavier Freire3
1 Universidade Federal do Amapá, Departamento de Ciências Biológicas e da Saúde,
Laboratório de Herpetologia, Rod. Juscelino Kubitschek, km 02, Jardim Marco Zero,
CEP 68.902-280, Macapá, AP, Brasil. 2 Universidade Federal do Rio Grande do Norte, Programa de Pós-Graduação em
Psicobiologia, Lagoa Nova, CEP 59.072-970, Natal, RN, Brasil.
3 Universidade Federal do Rio Grande do Norte, Departamento de Botânica, Ecologia e
Zoologia, Laboratório de Herpetologia, Lagoa Nova, CEP 59072-970, Natal, RN, Brasil.
* Corresponding author. E-mail: [email protected]
1. RESUMO
No estudo de ecologia de comunidades, diversos fatores estão envolvidos na estruturação
das comunidades e têm sido abordados sob diferentes aspectos. As dimensões de nicho
espacial, temporal e trófico podem fornecer informações sobre as interações entre as
populações, bem como a influencia de processos abióticos e bióticos sobre essas
populações. Objetivando analisar a utilização, largura e sobreposição de nicho espacial,
temporal e trófico na estruturação da comunidade de anuros, este estudo foi realizado em
área de Savana amazônica localizada no Campo Experimental da Empresa Brasileira de
Pesquisa Agropecuária, Estado do Amapá, Amazônia Oriental. Para avaliar os valores de
sobreposição de nicho entre os pares de espécies foram construídos modelos nulos a partir
da aleatorização dos dados da comunidade. Os valores obtidos pela largura de nicho
espacial, temporal e trófico sugerem que as espécies de anuros da Savana amapaense é
composta predominantemente por espécies generalistas. A presença também de anuros
especialistas pode ajudar a explicar os processos de especiação decorrentes do isolamento
dos hábitats como resultado da heterogeneidade e descontinuidade espacial encontrada
no domínio fitogeográfico das formações abertas. As análises de modelos nulos indicaram
a ocorrência de estruturação de comunidade quanto aos nichos temporal e trófico,
indicando influência significativa dos fatores ecológicos contemporâneos sobre a
comunidade. A ausência de estrutura quanto ao nicho espacial pode ser explicada pela
segregação espacial na distribuição e ocupação dos anuros nas diferentes fitofisionomias
da Savana amapaense. A especificidade da distribuição no nicho espacial, apresentada
por algumas espécies, também pode ser atribuída à limitação imposta pelos modos
reprodutivos específicos que podem restringir a utilização de um determinado habitat e
microhabitat.
PALAVRAS-CHAVE: Taxocenose de anuros; Nicho ecológico; Modelos nulos.

41
2. ABSTRACT
In the study of community ecology, several factors are responsible for structuring
communities and have been examined from different perspectives. The dimensions of
spatial, temporal, and trophic niche may provide information about the interactions
among populations and/or communities, as well as how they may be affected by abiotic
and biotic processes. This study was aimed at examining the use, breadth, and overlap of
spatial, temporal, and trophic niche in the structuring of anuran assemblages in an area of
Amazon savanna located in the Experimental Station of the Brazilian Agricultural Research Agency, state of Amapá, Western Amazon. To assess niche overlap between
pairs of species, null models were constructed from the randomization of data obtained
for the community. The values obtained based on the breadth of spatial, temporal, and
trophic niche suggest that the anuran species of the Amapá savanna is not composed
predominantly of generalist species. The presence of specialist anurans may help explain
the speciation processes associated with the isolation of habitats as result of the
heterogeneity and spatial discontinuity found in the phytophysiognomies of open
formations. The analysis of the null models suggest a community structure based on
temporal and trophic niche, and a significant influence of contemporary ecological factors
on the assemblage. The absence of structure based on spatial niche may be explained by
the spatial segregation in the distribution and occupation of anurans in the different
physiognomies of the Amapá savanna. The specificity in the distribution of the spatial
niche observed for some species may be also due to the restrictions of specific
reproductive modes that can prevent the use of a given habitat and microhabitat.
KEY-WORDS: Anuran community; Ecological niche; Null models.
3. INTRODUÇÃO
O estudo de comunidade de anfíbios anuros pode apresentar diversas abordagens,
dentre eles a distribuição espacial e temporal das espécies (Oda et al. 2009, Maffei et al.
2011, Campos et al. 2013), os mecanismos pelos que a distribuição das espécies pode ser
influenciada por fatores abióticos e bióticos (Kopp et al. 2010) e a disponibilidade de
recursos alimentares entre as espécies coexistentes (Lima & Magnusson, 2000). Neste
contexto, vários fatores influenciam a distribuição dos anuros dentro de uma comunidade
(Pirani et al. 2012, Conte et al. 2013, Melo et al. 2013, Steen et al. 2013).
A variação sazonal na estrutura da comunidade tem relação direta com a
pluviometria, propiciando a formação de agregados reprodutivos conspícuos durante os
eventos reprodutivos para diversas espécies de anuros (Giaretta et al. 2008, Nomura et al.
2012). Uma sucessão de espécies pode ocorrer no mesmo ambiente conforme a dinâmica
climática sazonal dos habitats (Silva et al. 2011), provavelmente relacionada ao modo
reprodutivo e tolerância fisiológica (Abrunhosa et al. 2006). A chuva é um componente
fundamental nas atividades de vocalização, reprodução e desenvolvimento larval dos
anuros (Hartel et al. 2011, Vasconcelos et al. 2011).

42
A temperatura também tem um importante papel na distribuição espaço-temporal
(Cassemiro et al. 2012) e na atividade de vocalização e comportamental dos anuros, sendo
que algumas espécies iniciam sua atividade de acordo com a variação térmica (Santos et
al. 2008, Canavero et al. 2009). Entretanto, a temperatura, na maioria das vezes, pode
estar relacionada com a umidade do ar; locais com altas temperaturas associadas a altas
taxas de umidade favorecem a formação de sítios reprodutivos para os anuros (Costa et
al. 2013). Outros fatores como a claridade natural (luz lunar) também interferem no
comportamento das espécies de anuros (Prado & Pombal Jr. 2005).
Outro fator relacionado com a distribuição espacial das espécies de anuros em
determinada área é a ocupação dos habitats e microhabitats disponíveis, que difere tanto
no plano horizontal quanto no vertical (Tews et al. 2004). Neste contexto, a coexistência
das espécies varia de acordo com a capacidade de exploração dos microhabitats com
características distintas (Abrunhosa et al. 2006). Dentro do mesmo habitat e microhabitat,
as espécies podem apresentar segregação quanto ao sítio de vocalização e quando há
sobreposição, esta é diretamente relacionada com a riqueza e a composição de espécies
(Zina et al. 2007).
A dieta é outro fator relevante na estrutura de comunidade, e pode ser partilhada
para o conhecimento das interações entre as espécies de anuros coexistentes de uma
taxocenose, fornecendo informações sobre as condições e os recursos alimentares
existentes no habitat (Andrew & Christensen 2001, Menin et al. 2005). Neste contexto,
estudos sobre os hábitos alimentares podem servir muitas vezes como informação básica
para o desenvolvimento de estratégias de conservação de populações naturais (Wiens &
Graham 2005).
A importância da descrição desses processos vai além da determinação de uma
possível estrutura de comunidade do grupo, fornecendo também dados essenciais para
estudos e elaborações de modelos ecológicos, de especial importância para as Savanas
amazônicas. Neste contexto, este trabalho tem como objetivo analisar a utilização, largura
e sobreposição de nicho espacial, temporal e trófico na estruturação da taxocenose de
anuros em área de Savana amazônica no Estado do Amapá.
4. MATERIAL E MÉTODOS
4.1. Área de estudo
As Savanas do Amapá ocorrem sobre uma estreita faixa longitudinal de 9.861,89
Km2, estendendo-se paralelamente à costa atlântica, desde a região do Jarí até o alto curso

43
do rio Uaçá (extremo norte do Amapá) e correspondendo a 6,9% da superfície territorial
do Estado (Rabelo et al. 2008). Este estudo foi realizado especificamente no Campo
Experimental da Empresa Brasileira de Pesquisa Agropecuária – EMBRAPA (00°023’5”
N, 51°02’2” W), uma área de Savana amazônica com 1.381 ha e perímetro de 15.080 m
(Melém-Júnior et al. 2003), localizada a 48 km ao norte da cidade de Macapá e com
limites a oeste com a BR-156 e limites norte, sul e leste com propriedades particulares.
O Campo Experimental da EMBRAPA possui quatro fitofisionomicas distintas
de vegetação nativa de Savana amazônica (cf. Miranda et al. 2003), a saber: savana
gramíneo lenhosa, caracterizada por extensos campos de gramíneas e estrato herbáceo
abundante, com estrato lenhoso disperso e composto por árvores anãs esparsas (< 1,0 m);
savana gramíneo lenhosa-arbustiva, com estrato herbáceo coberto por gramíneas em
grande densidade e estrato lenhoso com presença irregular e pequenas árvores esparsas
(< 3,8 m); savana parque, com estrato herbáceo menos desenvolvido e cobertura arbóreo-
arbustiva relativamente baixa (< 4,3 m) e densa formando um dossel descontínuo e,
savana arbórea, com estrato herbáceo ausente e árvores com maior densidade, formando
um dossel contínuo (> 2,0 m).
De acordo com a classificação Köppen, o clima predominante é Am, caracterizado
como tropical chuvoso com nítida estação seca (Alvares et al. 2013). A precipitação anual
varia entre 4,6 mm (estação seca) a 4,827 mm (estação chuvosa) e o clima ao longo do
ano pode ser tipicamente reconhecido em apenas duas estações: uma estação seca bem
evidente e uma estação chuvosa com precipitação pluviométrica elevada (janeiro a
junho). A temperatura média mensal varia de 24,4oC a 28,4oC, com umidade relativa
média de 84,7%.
O domínio da Savana amapaense ocorre sobre dois tipos de unidades de relevo:
planaltos cristalinos ou sedimentares e depressões interplanálticas. Em geral, nas regiões
dos planaltos, predominam a savana gramíneo lenhosa e a savana gramíneo lenhosa-
arbustiva, enquanto que nas depressões interplanáticas a savana parque e a savana arbórea
(Aguiar et al. 2004, Barbosa et al. 2005). O relevo é plano ou suavemente ondulado,
ocorrendo em duas formas principais: os Latossolos e as Areias quartzosas.
4.2. Procedimentos metodológicos
Para a amostragem dos anfíbios anuros da Savana amapaense foram realizadas 24
incursões em cada fitofisionomia, sendo 12 no período chuvoso e 12 no período seco.
Foram estabelecidas 20 unidades amostrais em cada fitofisionomia (Savana gramíneo

44
lenhosa, Savana gramíneo lenhosa-arbustiva, Savana parque e Savana arbórea), dispostas
de acordo com a disponibilidade de corpos d’água. Cada unidade amostral foi
representada por uma parcela de 100 x 50 metros (0,5ha), distantes pelo menos 500
metros entre si, conforme sugerido por Bitar et al. (2012). Nessas unidades amostrais
foram realizados os métodos de busca ativa visual e acústica (Crump & Scott 1994, Heyer
et al. 1994, Zimmerman, 1994).
4.2.1. Constância de ocorrência e Abundância de espécies
As espécies observadas e/ou coletadas nas quatro fitofisionomias da Savana
amapaense foram classificadas segundo o Índice de Constância de Ocorrência (C),
calculado conforme Krebs (2014).
𝑪 =𝒑 (𝟏𝟎𝟎)
𝑷
De acordo com os valores de C, as espécies foram categorizadas em: a) espécies
comuns, presentes em mais de 50% da amostragem; b) relativamente comuns, entre 25 a
50% e, c) esporádicas, espécies registradas em menos de 25% das amostragens.
A abundância de cada espécie foi estimada através da contagem do número de
vocalizações e/ou avistamento dos anuros. Para evitar superestimativas em decorrência
de recontagens de indivíduos em amostragens subsequentes, a abundância total de cada
espécie foi considerada igual à máxima abundância registrada em cada fitofisionomia.
Em cada fase de campo, a sequência dos horários das amostragens nas fitofisionomias foi
alterada, a fim de evitar erros no registro da abundância em função da variação no turno
de vocalização e/ou avistamento das espécies (SANTOS ET AL. 2007; RIBEIRO ET AL. 2012).
4.2.2. Distribuição espacial (habitat e microhabitat) e sazonal
A distribuição espacial das espécies de anuros foi analisada através de dois níveis:
tipos de habitats (Savana gramíneo lenhosa, gramíneo lenhosa-arbustiva, parque e
arbórea) e tipos de microhabitats. Os microhabitats utilizados como sítios de vocalização
e reprodução foram caracterizados pelas seguintes variáveis: (i) solo nu, espécies que
vocalizam no solo; (ii) água, espécies que vocalizam no interior de corpos d’água, com
parte ou todo corpo submerso; (iii) estrato herbáceo, espécies empoleiradas até 50 cm de
altura; (iv) estrato arbustivo, espécies empoleiradas entre 51 e 150 cm; (v) estrato arbóreo,
espécies empoleiradas acima de 151 cm de altura (Rossa-Feres & Jim 2001).
Onde p = número de espécies registradas, e P = número total de espécies

45
A distribuição sazonal foi feita com base nos dados mensais de abundância das
espécies de anuros registradas durante os dois anos de amostragem (2009 a 2010) e das
variáveis climatológicas (pluviometria, temperatura e umidade relativa). Para avaliar a
distribuição sazonal das espécies, foram considerados como picos de atividade os
períodos de maior abundância de machos vocalizantes (Kopp et al. 2010).
4.2.3. Distribuição temporal (período de atividade)
Para verificar a distribuição temporal ao longo da noite, de hora em hora, foi
estimado o número de machos em atividade de vocalização, através de classes
estabelecidas: (1) de 1 a 2 machos, (2) de 3 a 5 machos, (3) de 6 a 10 machos, (4) de 11
a 20 machos e (5) de 21 a 50 machos, de cada espécie (Bertoluci & Rodrigues 2002, Kopp
& Eterovick 2006). Os anuros foram classificados em, crepusculares (com vocalização
entre as 18:00 e 19:59 h), noturnos (20:00 e 23:59 h) e contínuos (18:00 e 23:59 h).
4.2.4. Análise da dieta
Para análise da dieta, os espécimes coletados foram mortos após a coleta com
anestésico (lidocaína 5%), fixados com formol a 10% e conservados em álcool a 70%.
Em laboratório, foram dissecados e através de incisão ventral, os estômagos foram
retirados, abertos e os seus conteúdos analisados através de microscópio estereoscópio.
As presas foram identificadas a nível de ordem e, quando possível, em categorias
inferiores. As presas registradas inteiras tiveram seu comprimento e largura medidos com
paquímetro digital Mitutoyo® (precisão de 0,01 mm) e seu volume estimado através da
fórmula do volume do elipsóide (cf. Magnusson et al. 2003).
𝑽 =𝟒
𝟑 𝝅 (
𝒍
𝟐) 𝒙 (
𝒘
𝟐)
𝟐
As porcentagens numéricas e volumétricas dos itens alimentares foram
quantificadas utilizando os métodos de frequência de ocorrência e volumétrico. Na
frequência de ocorrência foram registrados o número de estômagos em que cada item
ocorre, em relação à somatória de todos os itens alimentares. No método volumétrico
foram obtidos o volume de cada item alimentar, em relação ao total de itens. O índice de
importância de cada categoria de presa na dieta, foi calculado com a utilização da fórmula,
conforme descrito em Werneck et al. (2009) e Kolodiuk et al. (2010).
𝑰𝑹 = (𝑭% + 𝑵% + 𝑽%
𝟑)
Onde l= comprimento da presa e w= largura da presa
Onde IR = índice de importância; F% = porcentagem de ocorrência;
N% = porcentagem numérica; V% = porcentagem volumétrica.

46
4.3. Análise dos dados
As sobreposições no uso do nicho espacial (habitat e micro-habitat), temporal e
trófico entre os pares de espécies de anuros foram calculadas através da equação de
sobreposição (cf. Pianka 1973).
∅𝒊𝒋 =∑ 𝒑𝒊𝒋 𝒑𝒊𝒌
𝒏𝒊=𝟏
√∑ 𝒑𝒊𝒋𝟐 ∑ 𝒑𝒊𝒌𝟐𝒏𝒊=𝟏
𝒏𝒊=𝟏
A sobreposição varia de zero (nenhuma sobreposição) a um (sobreposição total).
Os valores de sobreposição foram considerados altos (>0,6) ou baixos (<0,4). Esses
índices foram obtidos e aleatorizados 1.000 vezes com o auxílio do programa Ecosim
7.71 (Gotelli & Entsminger 2005).
Para avaliar os valores de sobreposição de nicho espacial e trófico entre os pares
de espécies foram construídos modelos nulos a partir da aleatorização dos dados da
comunidade (Gotelli & Colwell 2001). A presença de padrões não aleatórios na
sobreposição de nicho foi obtida com a utilização do módulo de sobreposição de nicho
“Niche Overlap Module” do programa EcoSim 7.71 (Gotelli & Entsminger 2005),
utilizando as opções: Índice de sobreposição de nicho de Pianka e Algoritmo de
randomização para substituir as categorias de nicho da matriz original por números
uniformes e aleatórios entre 0 e 1 (Crawford et al. 2009).
Para cada espécie foram calculadas a amplitude do nicho espacial e trófico através
do índice de Levins (B) como descrito em Pianka (1986) e Vitt et al. (2001).
𝑩 = 𝟏 ∑ 𝒑𝒊𝟐
𝒏
𝒊=𝟏
⁄
Os valores obtidos de B foram utilizados para calcular o índice padronizado de
Levins (Bp). Para calcular os índices de largura de nicho foi utilizado o software
Ecological Metodology, Version 5.2 (Krebs 2014). Por produzir valores entre um
intervalo conhecido (0 - 1), este índice foi utilizado para comparar as larguras de nicho
das espécies de anuros.
𝑩𝒑 =(𝜷−𝟏)
(𝒏−𝟏)
Onde B = índice de Levins; i = categoria de habitat, microhabitat ou dieta; n = número de
categorias de habitat, micro-habitat ou dieta; pi = proporção de indivíduos associados à
categoria i de habitat, microhabitat ou dieta.
Onde Bp = índice padronizado de Levins; B = índice de Levins; n = número de
categorias utilizadas.
Onde Øij = índice de similaridade de nicho; p = a proporção da
categoria de habitat, microhabitat ou dieta; j e k = representam o par de
espécies comparadas.

47
As possíveis diferenças nas variações espaço-temporais da riqueza e abundância
das espécies e variáveis climatológicas foram verificadas através da Análise de Variância
de Kruskal-Wallis e Dunn a posteriori (Zar 2010). Para testarmos diferenças
significativas entre a sazonalidade (período chuvoso e seco) com as variações espaço-
temporais citadas, utilizamos o Teste de Mann-Whitney (com correção de Bonferroni).
Para testar a relação entre as variáveis ambientais (pluviometria, temperatura e
umidade relativa) e a riqueza e a abundância das espécies de anuros foi utilizado o método
de regressão linear simples (Zar 2010). Todas as análises foram realizadas através dos
programas “BioEstat 5.0 (Ayres et al. 2007) e PAST 2.09 (Hammer et al. 2001). Os
valores foram considerados significativos para p < 0,05.
5. RESULTADOS E DISCUSSÃO
5.1. Uso e partilha de habitat e microhabitat pelas espécies
Quanto à composição das espécies por habitat, o maior número de indivíduos foi
registrado na Savana gramíneo lenhosa-arbustiva (1.039), seguido pela Savana gramíneo
lenhosa (184), ambas áreas abertas da Savana estudada. Phyllomedusa hypochondrialis e
L. podicipinus ocorreram apenas na Savana gramíneo lenhosa-arbustiva e R.
margaritifera na Savana arbórea (Tabela 1).
Tabela 1 – Famílias/Espécies, número de indivíduos (N) e porcentagem de anuros (%) registrados
para a Savana amapaense, por habitat, durante o período de Janeiro de 2009 a Dezembro de 2010.
SGL = savana gramíneo lenhosa; SGLA = savana gramíneo lenhosa-arbustiva; SP = savana
parque e SA = savana arbórea.
Famílias/Espécies Habitats
SGL SGLA SP SA
N % N % N % N %
Bufonidae
Rhinella major --- --- 1 0,1 10 7,8 1 1,3
Rhinella margaritifera --- --- --- --- --- --- 54 68,4
Rhinella marina 5 2,7 10 1,0 --- --- --- ---
Hylidae
Dendropsophus walfordi 2 1,1 35 3,4 --- --- --- ---
Hypsiboas multifasciatus 20 10,9 96 9,2 16 12,4 --- ---
Hypsiboas punctatus --- --- 23 2,2 47 36,4 --- ---
Hypsiboas raniceps 12 6,5 19 1,8 20 15,5 --- ---
Osteocephalus taurinus 8 4,3 28 2,7 9 7,0 --- ---
Scinax fuscomarginatus 14 7,6 5 0,5 --- --- --- ---
Scinax nebulosus 3 1,6 167 16,1 --- --- --- ---

48
Tabela 1 (Continuação) – Famílias/Espécies, número de indivíduos (N) e porcentagem de
anuros (%) registrados para a Savana amapaense, por habitat, durante o período de Janeiro de
2009 a Dezembro de 2010. SGL = savana gramíneo lenhosa; SGLA = savana gramíneo lenhosa-
arbustiva; SP = savana parque e SA = savana arbórea.
Famílias/Espécies Habitats
SGL SGLA SP SA
N % N % N % N %
Hylidae
Scinax ruber 16 8,7 123 11,8 --- --- --- ---
Scinax x-signatus 3 1,6 55 5,3 --- --- --- ---
Trachycephalus typhonius 17 9,2 121 11,6 --- --- --- ---
Phyllomedusa hypochondrialis --- --- 174 16,7 --- --- --- ---
Leptodactylidae
Pseudopaludicola boliviana 15 8,2 4 0,4 --- --- --- ---
Adenomera hylaedactyla 23 12,5 44 4,2 --- --- 20 25,3
Leptodactylus fuscus 46 25,0 91 8,8 --- --- --- ---
Leptodactylus macrosternum --- --- --- --- 10 7,8 4 5,1
Leptodactylus pentadactylus --- --- 16 1,5 3 2,3 --- ---
Leptodactylus podicipinus --- --- 6 0,6 --- --- --- ---
Microhylidae
Elachistocleis helianneae --- --- 21 2,0 14 10,9 --- ---
Total 184 1039 129 79
Uma das diferenças na composição de anuros entre os habitats amostrados da
Savana amapaense podem ser explicados com relação a estrutura da vegetação. Diversos
estudos demonstram que a vegetação influencia a taxocenose de anuros, seja limitando a
sua presença, ou alterando sua abundância (Enst & Rödel 2005, Ernst et al. 2006). Em
estudo realizado por Neckel-Oliveira et al. (2000) em uma área de Savana amazônica no
município de Santarém, Pará, a maior riqueza e abundância de anuros foi observada em
formações florestais, indicando que algumas espécies são generalistas, enquanto outras
estão restritas a habitats específicos.
Além da vegetação, outro fator que pode explicar a diferença na composição de
espécies entre os habitats pode estar relacionado à distribuição não homogênea e a
disponibilidade de sítios aquáticos adequados a reprodução (Nomura et al. 2012, Araújo
et al. 2013). Adicionalmente, características morfofisiológicas ou comportamentais
(Conte et al. 2013), e consequentemente, os modos reprodutivos com estágio larval
aquático na maioria das espécies (Becker et al. 2010) são um dos principais fatores que
influenciam na composição e na distribuição de anuros.

49
Na Savana amapaense, dois fatores não mutuamente exclusivos podem explicar a
maior riqueza e composição de espécies na Savana gramíneo lenhosa-arbustiva: 1) a
heterogeneidade do habitat, com vegetação estruturada por estrato herbáceo coberto por
gramíneas e estrato lenhoso com presença irregular e pequenas árvores esparsas,
possibilita a ocupação vertical por hilídeos e por espécies terrestres, principalmente
bufonídeos e leptodactilídeos, e 2) a plasticidade no desenvolvimento larval das espécies
de anuros, com período larval curto como forma de minimizar o risco acumulado de
mortalidade por predação ou dessecação do habitat (Campos et al. 2013).
A menor riqueza e composição de espécies na savana arbórea pode estar
relacionada ao menor número de microhabitats disponíveis e aos modos reprodutivos das
espécies. A quantidade de microhabitats com substratos para vocalização pode ser um
fator determinante para diferenciar o habitat com maior riqueza e composição de espécies
de anuros (Afonso & Eterovick 2007, Pirani et al. 2013). Outro fator que pode contribuir
para uma diminuição na riqueza e composição de anuros é a existência de espécies que
apresentam modos reprodutivos mais generalizados, caracterizado pela deposição de ovos
em corpos de água lênticos e em ninhos de espuma com girinos aquáticos (Pombal Jr. &
Haddad 2007).
Neste contexto, como explicar as diferentes composições de espécies de anuros
entres os habitats da Savana amapaense? Alguns estudos verificam que apesar dos
habitats serem próximos entre si, existem diferenças quanto à composição da taxocenose
de anuros (Pinheiro et al. 2012). Isto se deve ao fato de que algumas espécies ocupam
uma variedade de habitats, enquanto outras possuem requerimentos de habitat mais
especializados (Valdujo et al. 2012), como observado no presente estudo.
Consequentemente, diferenças na estrutura da vegetação e disponibilidade de corpos
d’água entre os habitats, podem explicar as diferenças entre as taxocenoses de anfíbios,
limitando sua composição e abundância (Toft 1985, Oliveira & Ratter 2002).
A elevada riqueza de anuros com as áreas abertas do Cerrado registradas por
diversos autores (Colli et al. 2002, Brasileiro et al. 2005, Araújo et al. 2009, Valdujo et
al. 2011, Campos et al. 2013, Araújo et al. 2013, Gambale et al. 2014) também foi
observada em áreas abertas (Savana gramíneo lenhosa e Savana gramíneo lenhosa-
arbustiva) presentes na Savana amapaense. Apesar da diminuição da riqueza de espécies
nas formações florestais (Savana parque e Savana arbórea), vale ressaltar a importância
destes habitats mésicos como locais de refúgio, forrageio, hibernação e migração
utilizados por anuros adultos e juvenis (Silva & Rossa-Feres 2007, Silva et al. 2011).

50
No que se refere à composição das espécies por microhabitat, no estrato herbáceo
foi registrado um maior número de indivíduos (523), seguido pelo estrato arbutivo (437)
e pelo solo (283) Das espécies registradas, R. margaritifera e R. marina ocorreram apenas
no microhábitat de solo (Tabela 2).
Tabela 2 – Famílias/Espécies, número de indivíduos (N) e porcentagem de anuros (%) registrados
para a Savana amapaense, por microhabitat, durante o período de Janeiro de 2009 a Dezembro de
2010. S = Solo; A = Água; Eherb = Estrato herbáceo; Earbu = Estrato arbustivo; Earbo = Estrato
arbóreo.
Famílias/Espécies
Microhabitats
S A Eherb Earbu Earbo
N % N % N % N % N %
Bufonidae
Rhinella major 8 2,8 4 4,9 --- --- --- --- --- ---
Rhinella margaritifera 54 19,1 --- --- --- --- --- --- --- ---
Rhinella marina 15 5,3 --- --- --- --- --- --- --- ---
Hylidae
Dendropsophus walfordi --- --- --- --- 25 4,8 12 2,7 --- ---
Hypsiboas multifasciatus 14 4,9 --- --- 97 18,5 21 4,8 --- ---
Hypsiboas punctatus --- --- --- --- 28 5,4 42 9,6 --- ---
Hypsiboas raniceps --- --- --- --- 27 5,2 24 5,5 --- ---
Osteocephalus taurinus --- --- --- --- 5 1,0 33 7,6 7 6,6
Scinax fuscomarginatus --- --- --- --- 6 1,1 13 3,0 --- ---
Scinax nebulosus --- --- --- --- 108 20,7 60 14,2 --- ---
Scinax ruber --- --- --- --- 74 14,1 57 13,0 8 7,5
Scinax x-signatus --- --- --- --- 20 3,8 22 5,0 16 15,1
Trachycephalus typhonius 14 4,9 --- --- 14 2,7 62 14,2 48 45,8
Phyllomedusa hypochondrialis 12 4,2 --- --- 46 8,8 89 20,4 27 25,5
Leptodactylidae
Pseudopaludicola boliviana 9 3,2 10 12,2 --- --- --- --- --- ---
Adenomera hylaedactyla 14 4,9 --- --- 73 14,0 --- --- --- ---
Leptodactylus fuscus 112 39,6 25 30,5 --- --- --- --- --- ---
Leptodactylus macrosternum 4 1,4 10 12,2 --- --- --- --- --- ---
Leptodactylus pentadactylus 17 6,0 2 2,4 --- --- --- --- --- ---
Leptodactylus podicipinus 4 1,4 2 2,4 --- --- --- --- --- ---
Microhylidae
Elachistocleis helianneae 6 2,1 29 35,4 --- --- --- --- --- ---
Total 283 82 523 437 106
A composição ou a coexistência de espécies está diretamente ligada com a
capacidade de exploração dos microhabitats (Vasconcelos & Rossa-Feres 2008). De
acordo com Haddad & Prado (2005), quanto maior o número de microhabitats maior é a
riqueza de anuros. De modo geral, o padrão de distribuição espacial das espécies da
Savana amapaense não difere de estudos em outras localidades do Cerrado (Brasileiro et

51
al. 2005, Araújo et al. 2009; Araújo & Almeida-Santos 2011, Valdujo et al. 2011), pois
os membros da família Hylidae, em sua maioria arborícolas, geralmente estão associados
a microhabitats para vocalização acima do chão. Já membros de outras famílias, como
Bufonidae, Leptodactylidae e Microhylidae estão restritos ao solo ou às margens de
corpos d‘água (Toledo et al., 2003; Morais et al., 2011).
5.2. Constância das espécies
O índice de constância para as espécies de anuros amostradas na Savana
amapaense apresentou 13 espécies comuns, cinco espécies relativamente comuns e três
espécies esporádicas (Tabela 3).
Tabela 3 – Índice de constância dos anuros amostrados durante o período de Janeiro de 2009 a
Dezembro de 2010 nas fitofisionomias da Savana amapaense: SGL – savana gramíneo lenhosa;
SGLA – savana gramíneo lenhosa-arbustiva; SP – savana parque e SA – savana arbórea. C =
espécie comum; RC = espécie relativamente comum; E – espécie esporádica.
Famílias/Espécies Fitofisionomias
SGL SGLA SP SA
% Status % Status % Status % Status
Bufonidae
Rhinella major --- --- 4,2 E 17,6 E 7,7 E
Rhinella margaritifera --- --- --- --- --- --- 100,0 C
Rhinella marina 18,7 E 20,8 E --- --- --- ---
Hylidae
Dendropsophus walfordi 12,5 E 33,3 RC --- --- --- ---
Hypsiboas multifasciatus 50,0 RC 58,3 C 23,5 E --- ---
Hypsiboas punctatus --- --- 16,7 E 100,0 C --- ---
Hypsiboas raniceps 12,5 E 33,3 RC 47,1 RC --- ---
Osteocephalus taurinus 12,5 E 29,2 RC 29,4 RC --- ---
Scinax fuscomarginatus 25,0 RC 8,3 E --- --- --- ---
Scinax nebulosus 6,2 E 83,3 C --- --- --- ---
Scinax ruber 31,2 RC 62,5 C --- --- --- ---
Scinax x-signatus 37,5 RC 41,7 RC --- --- --- ---
Trachycephalus typhonius 37,5 RC 87,5 C --- --- --- ---
Phyllomedusa hypochondrialis --- --- 95,8 C --- --- --- ---
Leptodactylidae
Pseudopaludicola boliviana 37,5 RC 4,2 E --- --- --- ---
Adenomera hylaedactyla 37,5 RC 50,0 RC 46,3 RC --- ---
Leptodactylus fuscus 68,7 C 91,7 C --- --- --- ---
Leptodactylus macrosternum --- --- --- --- 23,5 E 7,7 E
Leptodactylus pentadactylus --- --- 25,0 RC 5,9 E --- ---
Leptodactylus podicipinus --- --- 8,3 E --- --- --- ---
Microhylidae
Elachistocleis helianneae --- --- 16,7 E 23,5 E --- ---

52
O índice de constância para as espécies amostradas na Savana amapaense destaca
a presença de espécies ecologicamente generalistas de áreas abertas, como: D. walfordi,
S. fuscomarginatus, S. nebulosus, S. ruber, S. x-signatus, T. typhonius, P. boliviana e L.
fuscus (Santos et al. 2009). Aspectos como a capacidade das espécies de colonizar áreas
abertas, tolerâncias fisiológicas distintas e a necessidade de microhabitats específicos
para a reprodução são possíveis fatores responsáveis pelos distintos padrões de frequência
de ocorrência da taxocenose de anuros (Melo et al. 2007, Moraes et al. 2007).
5.3. Abundância das espécies por habitat e microhabitat
As espécies mais abundantes foram P. hypochondrialis, S. nebulosus e S. ruber,
correspondendo a 34,1% do total de indivíduos registrados para a Savana amapaense. Nos
habitats amostrados, P. boliviana foi a espécie mais abundante na Savana gramíneo
lenhosa, com 78,9% dos indivíduos amostrados. Na Savana parque as espécies mais
abundantes foram R. major (83,4%) e L. macrosternum (71,4%). As espécies registradas
em apenas um habitat foram: P. hypochondrialis e L. podicipinus, na Savana gramíneo
lenhosa-arbustiva e R. margaritifera, na Savana arbórea (Figura 1).
Figura 1 – Abundância relativa das espécies de anfíbios anuros registradas durante o período de
Janeiro de 2009 a Dezembro de 2010 nos habitats da Savana amapaense (SGL = savana gramíneo
lenhosa; SGLA = savana gramíneo lenhosa-arbustiva; SP = savana parque; SA = savana arbórea).

53
As maiores abundâncias de anuros foram registradas em áreas abertas (Savana
gramíneo lenhosa e Savana gramíneo lenhosa-arbustiva) associadas a corpos d’água
temporários utilizados como sítios reprodutivos, corroborando com os resultados obtidos
em regiões com sazonalidade climática (Santos et al. 2007, Vasconcelos et al. 2009,
Campos et al. 2013). As áreas abertas com corpos d’água temporários, permite que os
girinos explorem fontes de alimento antes da colonização de insetos e peixes, evitando ao
mesmo tempo as pressões de predação e competição (Höld 1990, Prado et al. 2005).
Em contraste, a redução na abundância de anuros registrados em áreas florestais
(Savana parque e Savana arbórea) com corpos d’água permanentes está associada
principalmente com a maior estratificação vegetal e com os modos reprodutivos
especializados. A mudança nas comunidades de plantas gera variações na disponibilidade
de microhabitats, que pode afetar padrões demográficos em taxocenoses de anuros
(Vasconcelos et al. 2009).
As espécies que utilizaram um maior número de microhabitats foram P.
hypochondrialis (Solo = 6,9%; Estrato herbáceo = 26,4%; Estrato arbustivo = 51,2% e
Estrato arbóreo = 15,5%) e T. typhonius (Solo = 10,1%; Estrato herbáceo = 10,1%; Estrato
arbustivo = 44,9% e Estrato arbóreo = 34,8%). O menor número de microhabitats foi
registrado para as espécies R. margaritifera e R. marina, observadas no solo (Figura 2).
Figura 2 – Abundância relativa das espécies de anfíbios anuros registradas durante o período de
Janeiro de 2009 a Dezembro de 2010 nos microhabitats da Savana amapaense (S = Solo; A =
Água; Eherb = Estrato herbáceo; Earbu = Estrato arbustivo e Earbo = Estrato arbóreo).

54
A abundância das espécies registradas na Savana amapaense diferiu
significativamente (H = 11,77; gl = 3; p = 0,0082). Nos testes à posteriori de Dunn, houve
diferença significativa apenas entre a Savana gramíneo lenhosa-arbustiva e Savana
arbórea (p < 0,05).
Das espécies registradas, os bufonídeos, leptodactlídeos e microhilídeos são
terrestres (solo e água) e os hilídeos são trepadores. As espécies R. margaritifera e R.
marina foram as únicas espécies registradas no microhabitat solo. Todos os
leptodactilídeos foram registrados no solo ou em poças temporárias, fato atribuído por
serem espécies terrícolas e com desenvolvimento indireto com girinos em ninhos de
espuma (Pombal Jr. & Haddad 2007, Araújo et al. 2009).
O solo também foi utilizado como substrato por três espécies trepadoras, H.
multifasciatus, P. hypochondrialis e T. typhonius. Este fato pode estar relacionado a
ausência de vegetação sobre a água e a disponibilidade de poleiros emergentes, fazendo
com estas espécies utilizem o solo como sítio reprodutivo (Melo et al. 2007). As espécies
P. hypochondrialis e T. typhonius utilizaram o solo para vocalização e os poleiros acima
dos corpos d’água para formação de casais em amplexo.
Os valores de largura do nicho espacial para o habitat variaram de 0,04 a 0,94 para
a Savana gramíneo lenhosa e Savana gramíneo lenhosa-arbustiva, de 0,21 a 0,82 para a
Savana parque e de 0,21 a 0,94 para a Savana arbórea. A largura de nicho espacial para o
microhabitat variou de 0,23 a 0,99, com 61,9% de espécies generalistas (Tabela 4).
Os valores obtidos pela largura de nicho espacial e o padrão de distribuição local,
sugerem que as espécies de anuros da Savana amapaense não é composta predominan-
temente por espécies generalistas. A presença também de anuros especialistas pode ajudar
a explicar os processos de especiação ligados com o isolamento dos habitats como
resultado da heterogeneidade e descontinuidade espacial encontrada no domínio
fitogeográfico das formações abertas (Cavalcanti & Joly 2002, Oliveira-Filho & Ratter
2002, Giaretta et al. 2008, Oda et al. 2009).
Neste contexto, a variação da largura de nicho nos habitats e microhabitats com a
presença de espécies especialistas e generalistas tem relação direta com as características
reprodutivas e com a heterogeneidade estrutural dentro da taxocenose (Amèzquita &
Hodl 2004). Estes resultados vão de acordo com o postulado de que, taxocenoses de
anuros que habitam áreas com alta heterogeneidade espacial podem acomodar um grande
número de espécies, por apresentarem uma grande quantidade de microhabitats
disponíveis (Campos & Vaz-Silva 2010).

55
Tabela 4 – Largura de nicho espacial para os habitats e microhabitats utilizados pela comunidade
de anuros da Savana amapaense, durante o período de Janeiro de 2009 a Dezembro de 2010.
O maior valor dos índices de sobreposição de nicho espacial quanto ao uso do
habitat ocorreu nos pares de espécies P. hypochondrialis vs. L. podicipinus, registrados
apenas na Savana gramíneo lenhosa-arbustiva. Quanto ao uso de microhabitats, a maior
sobreposição ocorreu nos pares de espécies R. marina vs. R. margaritifera e L.
podicipinus vs. R. major (Tabela 5).
De acordo com Oda et al. (2009) e Melo et al. (2013), o padrão de variação
fitofisionômica e de heterogeneidade local dos habitats e microhabitats utilizados pelos
anuros cria um gradiente de condições ambientais e variações na utilização de recursos
que podem ser exploradas por um grande número de espécies, contribuindo assim para a
alta sobreposição. A associação desses fatores constitui para a determinação do número
de espécies que podem explorar um determinado nicho espacial (Campos et al. 2013).
Espécies Habitats Microhabitats
Rhinella major 0,21 0,80
Rhinella margaritifera 0 0
Rhinella marina 0,80 0
Dendropsophus walfordi 0,11 0,78
Hypsiboas multifasciatus 0,38 0,37
Hypsiboas punctatus 0,79 0,92
Hypsiboas raniceps 0,94 0,89
Osteocephalus taurinus 0,59 0,37
Scinax fuscomarginatus 0,63 0,76
Scinax nebulosus 0,04 0,86
Scinax ruber 0,26 0,60
Scinax x-signatus 0,11 0,97
Trachycephalus typhonius 0,28 0,62
Phyllomedusa hypochondrialis 0 0,59
Pseudopaludicola boliviana 0,50 0,99
Adenomera hylaedactyla 0,82 0,37
Leptodactylus fuscus 0,80 0,42
Leptodactylus macrosternum 0,69 0,69
Leptodactylus pentadactylus 0,36 0,23
Leptodactylus podicipinus 0 0,80
Elachistocleis helianneae 0,92 0,40

56
Tabela 5 – Sobreposição de nicho baseado no uso dos habitats (acima) e microhabitats (abaixo) utilizados pelos anuros da Savana amapaense durante o período
de Janeiro de 2009 a Dezembro de 2010. Rmj = R. major, Rmarg = R. margaritifera, Rm = R. marina, Dw = D. walfordi, Hm = H. multifasciatus, Hp = H.
punctatus, Hr = H. raniceps, Ot = O. taurinus, Sf = S. fuscomarginatus, Sn = S. nebulosus, Sr = S. ruber, Sx-s = S. x-signatus, Tt = T. typhonius, Ph = P.
hypochondrialis, Pb = P. boliviana, Ah = A. hylaedactyla, Lf = L. fuscus, Lm = L. macrosternum, Lpenta = L. pentadactylus, Lpod = L. podicipinus, Eh =
E.helianneae. Os números em negrito indicam valores de sobreposição de nicho ≥ 95 %.
Espécies Rmj Rmarg Rm Dw Hm Hp Hr Ot Sf Sn Sr Sx-s Tt Ph Pb Ah Lf Lm Lpent Lpod Eh
Rhinella major 0,10 0,09 0,10 0,26 0,93 0,72 0,38 0,03 0,10 0,10 0,10 0,10 0,10 0,03 0,12 0,09 0,96 0,28 0,10 0,63
Rhinella margaritifera 0,89 0 0 0 0 0 0 0 0 0 0 0 0 0 0,37 0 0,37 0 0 0
Rhinella marina 0,89 1 0,92 0,95 0,39 0,74 0,94 0,72 0,90 0,95 0,92 0,95 0.89 0,66 0,93 1 0 0,88 0,89 0,74
Dendropsophus walfordi 0 0 0 0,98 0,44 0,65 0,93 0,39 1 1 1 1 1 0,31 0,85 0,92 0 0,98 1 0,83
Hypsiboas multifasciatus 0,13 0,14 0,14 0,96 0,57 0,80 0,99 0,52 0,97 0,98 0,98 0,99 0,97 0,44 0,88 0,95 0,15 0,98 0,97 0,89
Hypsiboas punctatus 0 0 0 0,86 0,71 0,87 0,67 0,15 0,44 0,44 0,44 0,68 0,44 0,11 0,36 0,39 0,83 0,60 0,44 0,86
Hypsiboas raniceps 0 0 0 0,96 0,86 0,97 0,88 0,59 0,64 0,68 0,65 0,68 0,63 0,55 0,69 0,74 0,62 0,74 0,63 0,89
Osteocephalus taurinus 0 0 0 0,55 0,35 0,89 0,75 0,56 0,92 0,95 0,93 0,95 0,92 0,49 0,87 0,94 0,27 0,96 0,92 0,93
Scinax fuscomarginatus 0 0 0 0,77 0,60 1 0,92 0,94 0,35 0,46 0,39 0,46 0,96 1 0,68 0,73 0 0,33 0,34 0,28
Scinax nebulosus 0 0 0 1 0,94 0,90 0,98 0,61 0,82 0,99 1 0,99 0,81 0,28 0,83 0,90 0 0,98 1 0,83
Scinax ruber 0 0 0 0,98 0,89 0,94 1 0,72 0,88 0,99 1 1 0,89 0,38 0,87 0,94 0 0,98 0,99 0,83
Scinax x-signatus 0 0 0 0,82 0,71 0,87 0,88 0,82 0,84 0,84 0,90 1 0,94 0,31 0,84 0,92 0 0,98 1 0,83
Trachycephalus typhonius 0,16 0,17 0,17 0,49 0,35 0,73 0,64 0,89 0,77 0,53 0,65 0,88 0,90 0,39 0,87 0,95 0 0,97 0.99 0,82
Phyllomedusa hypochondrialis 0,10 0,12 0,12 0,77 0,62 0,95 0,90 0,94 0,34 1 0,99 1 0,99 0,26 0,82 0,89 0 0,98 1 0,83
Pseudopaludicola boliviana 0,93 0,67 0,67 0 0,09 0 0 0 0 0 0 0 0,12 0,08 0,13 0,82 0,94 0,75 0,93 0,86
Adenomera hylaedactyla 0,17 0,19 0,19 0,89 0,98 0,55 0,73 0,14 0,41 0,85 0,78 0,58 0,20 0,45 0,63 0,18 0,14 0,81 0,82 0,68
Leptodactylus fuscus 0,97 0,98 0,98 0 0,14 0 0 0 0 0 0 0 0,17 0,11 0,67 0,93 0 0,88 0,89 0,74
Leptodactylus macrosternum 0,75 0,37 0,37 0 0,05 0 0 0 0 0 0 0 0,06 0,04 0 0,70 0,57 0,17 0 0,52
Leptodactylus pentadactylus 0,94 1 1 0 0,14 0 0 0 0 0 0 0 0,17 0,11 0,25 0,19 1 0,48 0,98 0,92
Leptodactylus podicipinus 1 0,89 0,89 0 0,13 0 0 0 0 0 0 0 0,16 0,10 0,26 0,17 0,97 0,75 0,94 0,83
Elachistocleis helianneae 0,62 0,20 0,20 0 0,03 0 0 0 0 0 0 0 0,04 0,02 0,21 0,04 0,41 0,98 0,32 0,62

57
A média de sobreposição de nicho na utilização dos habitats pelas espécies de
anuros não foi significativamente diferente entre os valores observado e esperado (média
observada = 0,60; média esperada = 0,54; p = 0,069). A média de sobreposição do uso do
microhabitat também não mostrou diferença significativa entre os valores observado e
esperado pelo modelo nulo (média observada = 0,39; média esperada = 0,40; p = 0,480),
indicando ausência de estrutura com relação às dimensões de nicho espacial (Figura 3).
Figura 3 – Simulações da sobreposição de uso de habitats (a) e microhabitats (b) da comunidade
de anuros da Savana amapaense durante o período de Janeiro de 2009 a Dezembro de 2010. As
setas indicam as médias observadas de sobreposição de nicho espacial.
No presente estudo, a ausência de estrutura na taxocenose de anuros da Savana
amapaense quanto ao nicho espacial (habitat e microhabitat) pode ser explicada pela
segregação espacial na distribuição e ocupação dos anuros, sendo os padrões de
sobreposição e largura das espécies similares ao esperado em distribuições aleatórias nos
diferentes nichos. A especificidade na distribuição de um determinado nicho espacial
apresentada por algumas espécies também pode ser atribuída à limitação imposta pelos

58
modos reprodutivos específicos que podem restringir a utilização de um determinado
habitat e microhabitat (Haddad & Prado 2005, Pombal Jr. & Haddad 2007).
Assim, R. margaritifera, P. hypochondrialis e L. podicipinus podem ser
consideradas totalmente dependentes de seus habitats. Rhinella margaritifera somente foi
registrada na Savana arbórea, demonstrando forte dependência a esse tipo de habitat.
Phyllomedusa hypochondrialis e L. podicipinus foram apenas registradas na Savana
gramíneo lenhosa-arbustiva. As duas espécies, apesar de terem sido registradas no mesmo
habitat, apresentam modos reprodutivos distintos, com ovos arborícolas e ovos em ninho
de espuma flutuante em água acumulada em piscinas construídas, respectivamente.
5.4. Distribuição temporal (período de atividade)
A partir da análise de distribuição temporal de machos em atividade de
vocalização, as espécies R. margaritifera, L. macrosternum e L. pentadactylus
apresentaram período de atividade crepuscular (14,3%), sete espécies (33,3%)
apresentaram atividade noturna e 11 espécies (52,4%) apresentaram atividade contínua,
sendo este o período de atividade predominante da taxocenose estudada (Tabela 6).
Tabela 6 – Período de atividade de vocalização das espécies de anuros registradas na Savana
amapaense durante o período de Janeiro de 2009 a Dezembro de 2010. vocalização contínua,
vocalização crepuscular, vocalização noturna.
Espécies 18:00 - 18:59 19:00 - 19:59 20:00 - 20:59 21:00 - 21:59 22:00 - 22:59 23:00 -23:59
Rhinella major
Rhinella margaritifera
Rhinella marina
Dendropsophus walfordi
Hypsiboas multifasciatus
Hypsiboas punctatus
Hypsiboas raniceps
Osteocephalus taurinus
Scinax fuscomarginatus
Scinax nebulosus
Scinax ruber
Scinax x-signatus
Trachycephalus typhonius
Phyllomedusa hypochondrialis
Pseudopaludicola boliviana
Adenomera hylaedactyla
Leptodactylus fuscus
Leptodactylus macrosternum
Leptodactylus pentadactylus
Leptodactylus podicipinus
Elachistocleis helianneae
Horário/Turno de vocalização

59
Os hilídeos e leptodactilídeos apresentaram um período de atividade mais extenso,
iniciando as atividades mais cedo às 18 h e estendendo até às 24 h. Os valores de largura
do nicho temporal para o período de atividade variaram de 0,26 a 0,93, com 19,1% das
espécies especialistas e 80,9% generalistas (Tabela 7). A maior amplitude de nicho
temporal ocorreu entre as espécies R. major, S. ruber e T. typhonius.
Tabela 7 – Número de espécies, período de atividade de vocalização e amplitude de nicho
temporal dos anuros registrados na Savana amapaense durante o período de Janeiro de 2009 a
Dezembro de 2010.
Os maiores valores dos índices de sobreposição do período de atividade das
espécies de anuros ocorreu nos pares de espécies S. x-signatus vs. E. helianneae, H.
punctatus vs. P. boliviana e S. fuscomarginatus vs. P. boliviana, com índice de
sobreposição de 0,98. A sobreposição total ocorreu nos pares de espécies H.
multifasciatus vs. L. fuscus e R. margaritifera vs. L. pentadactylus (Tabela 8).
Os modelos nulos mostraram que a sobreposição média do período de atividade
das espécies de anuros da Savana amapaense foi significativamente diferente entre os
valores observado e esperado pelo modelo nulo (média observada = 0,60; média esperada
= 0,47; p < 0,001), indicando a existência de estrutura com relação ao período de atividade
(Figura 4).
Espécies 18:00 - 18:59 19:00 - 19:59 20:00 - 20:59 21:00 - 21:59 22:00 - 22:59 23:00 -23:59 N Amplitude
Rhinella major 9 14 18 15 0 0 56 0,93
Rhinella margaritifera 28 4 0 0 0 0 32 0,28
Rhinella marina 0 0 8 14 0 0 22 0,86
Dendropsophus walfordi 0 0 28 9 4 0 41 0,45
Hypsiboas multifasciatus 6 23 21 12 15 8 85 0,80
Hypsiboas punctatus 8 17 20 3 0 0 48 0,67
Hypsiboas raniceps 0 4 18 6 0 0 28 0,54
Osteocephalus taurinus 0 0 15 7 8 0 30 0,88
Scinax fuscomarginatus 0 18 26 4 0 0 48 0,63
Scinax nebulosus 6 17 24 26 16 18 107 0,86
Scinax ruber 8 12 14 14 17 0 65 0,93
Scinax x-signatus 4 28 18 5 0 0 55 0,54
Trachycephalus typhonius 0 0 17 12 9 14 52 0,93
Phyllomedusa hypochondrialis 0 3 32 24 5 0 64 0,50
Pseudopaludicola boliviana 4 18 25 0 0 0 47 0,64
Adenomera hylaedactyla 18 26 10 17 4 0 75 0,75
Leptodactylus fuscus 12 25 28 14 16 10 105 0,84
Leptodactylus macrosternum 2 15 0 0 0 0 17 0,26
Leptodactylus pentadactylus 18 3 0 0 0 0 21 0,32
Leptodactylus podicipinus 18 24 6 0 0 0 48 0,73
Elachistocleis helianneae 0 22 19 4 0 0 45 0,67
Horário/Turno de vocalização

60
Tabela 8 – Sobreposição de nicho temporal utilizado pelos anuros da Savana amapaense durante o período de Janeiro de 2009 a Dezembro de 2010. Rmj = R.
major, Rmarg = R. margaritifera, Rm = R. marina, Dw = D. walfordi, Hm = H. multifasciatus, Hp = H. punctatus, Hr = H. raniceps, Ot = O. taurinus, Sf = S.
fuscomarginatus, Sn = S. nebulosus Sr = S. ruber, Sx-s = S. x-signatus, Tt = T. typhonius, Ph = P. hypochondrialis, Pb = P. boliviana, A. hylaedactyla, Lf = L.
fuscus, Lm = L. macrosternum, Lpenta = L. pentadactylus, Lpod = L. podicipinus, Eh = E.helianneae. Os números em negrito indicam valores de sobreposição
de nicho ≥ 95 %
Espécies Rmj Rmarg Rm Dw Hm Hp Hr Ot Sf Sn Sr Sx-s Tt Ph Pb Ah Lf Lm Lpent Lpod Eh
Rhinella major 0,38 0,76 0,75 0,86 0,90 0,84 0,71 0,75 0,83 0,82 0,85 0,63 0,84 0,83 0,89 0,89 0,52 0,39 0,69 0,84
Rhinella margaritifera 0 0 0,24 0,37 0,03 0 0,08 0,18 0,32 0,23 0 0,01 0,21 0,57 0,34 0,27 1 0,69 0,11
Rhinella marina 0,73 0,55 0,45 0,77 0,73 0,51 0,74 0,64 0,39 0,71 0,91 0,40 0,53 0,57 0 0 0,10 0,44
Dendropsophus walfordi 0,67 0,72 0,97 0,94 0,81 0,70 0,66 0,55 0,78 0,94 0,76 0,40 0,72 0 0 0,19 0,65
Hypsiboas multifasciatus 0,85 0,74 0,74 0,83 0,92 0,92 0,86 0,74 0,72 0,82 0,83 1 0,62 0,26 0,68 0,86
Hypsiboas punctatus 0,83 0,63 0,95 0,70 0,72 0,94 0,51 0,68 0,98 0,81 0,89 0,65 0,39 0,80 0,95
Hypsiboas raniceps 0,88 0,91 0,73 0,66 0,71 0,73 0,93 0,87 0,53 0,77 0,20 0,03 0,34 0,80
Osteocephalus taurinus 0,71 0,78 0,81 0,49 0,84 0,92 0,66 0,44 0,77 0 0 0,16 0,58
Scinax fuscomarginatus 0,70 0,67 0,92 0,58 0,76 0,98 0,67 0,84 0,56 0,09 0,60 0,97
Scinax nebulosus 0,88 0,67 0,90 0,81 0,65 0,74 0,92 0,38 0,19 0,46 0,69
Scinax ruber 0,68 0,70 0,75 0,66 0,81 0,92 0,43 0,33 0,57 0,67
Scinax x-signatus 0,41 0,57 0,92 0,84 0,85 0,83 0,25 0,82 0,98
Trachycephalus typhonius 0,81 0,51 0,41 0,76 0 0 0,13 0,47
Phyllomedusa hypochondrialis 0,68 0,55 0,75 0,07 0,01 0,21 0,65
Pseudopaludicola boliviana 0,68 0,84 0,59 0,22 0,69 0,96
Adenomera hylaedactyla 0,84 0,75 0,59 0,88 0,75
Leptodactylus fuscus 0,57 0,35 0,70 0,85
Leptodactylus macrosternum 0,29 0,86 0,74
Leptodactylus pentadactylus 0,71 0,12
Leptodactylus podicipinus 0,72
Elachistocleis helianneae
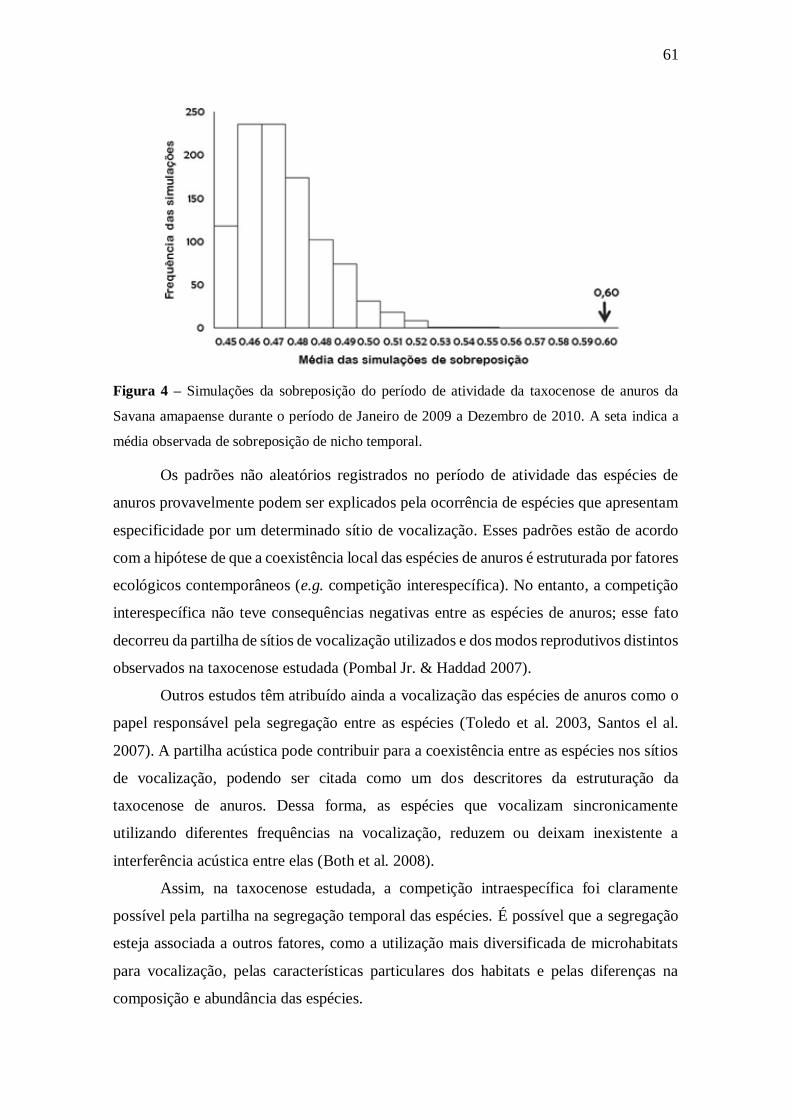
61
Figura 4 – Simulações da sobreposição do período de atividade da taxocenose de anuros da
Savana amapaense durante o período de Janeiro de 2009 a Dezembro de 2010. A seta indica a
média observada de sobreposição de nicho temporal.
Os padrões não aleatórios registrados no período de atividade das espécies de
anuros provavelmente podem ser explicados pela ocorrência de espécies que apresentam
especificidade por um determinado sítio de vocalização. Esses padrões estão de acordo
com a hipótese de que a coexistência local das espécies de anuros é estruturada por fatores
ecológicos contemporâneos (e.g. competição interespecífica). No entanto, a competição
interespecífica não teve consequências negativas entre as espécies de anuros; esse fato
decorreu da partilha de sítios de vocalização utilizados e dos modos reprodutivos distintos
observados na taxocenose estudada (Pombal Jr. & Haddad 2007).
Outros estudos têm atribuído ainda a vocalização das espécies de anuros como o
papel responsável pela segregação entre as espécies (Toledo et al. 2003, Santos el al.
2007). A partilha acústica pode contribuir para a coexistência entre as espécies nos sítios
de vocalização, podendo ser citada como um dos descritores da estruturação da
taxocenose de anuros. Dessa forma, as espécies que vocalizam sincronicamente
utilizando diferentes frequências na vocalização, reduzem ou deixam inexistente a
interferência acústica entre elas (Both et al. 2008).
Assim, na taxocenose estudada, a competição intraespecífica foi claramente
possível pela partilha na segregação temporal das espécies. É possível que a segregação
esteja associada a outros fatores, como a utilização mais diversificada de microhabitats
para vocalização, pelas características particulares dos habitats e pelas diferenças na
composição e abundância das espécies.

62
5.5. Distribuição sazonal das espécies
Durante o estudo foram registrados 1.203 espécimes no período chuvoso (janeiro
a junho) e 228 no período seco (julho a dezembro). De acordo com a Figura 5, as espécies
são mais frequentes no início da estação chuvosa, apresentando um padrão nitidamente
sazonal, como observado por Menin et al. (2005), Oda et al. (2009) e Maffei et al. (2011).
Figura 5 – Distribuição sazonal das espécies de anuros da Savana amapaense registradas durante
o período de Janeiro de 2009 a Dezembro de 2010.

63
Figura 5 (Continuação) – Distribuição sazonal das espécies de anuros da Savana amapaense
registradas durante o período de Janeiro de 2009 a Dezembro de 2010.

64
Figura 5 (Continuação) – Distribuição sazonal das espécies de anuros da Savana amapaense
registradas durante o período de Janeiro de 2009 a Dezembro de 2010.
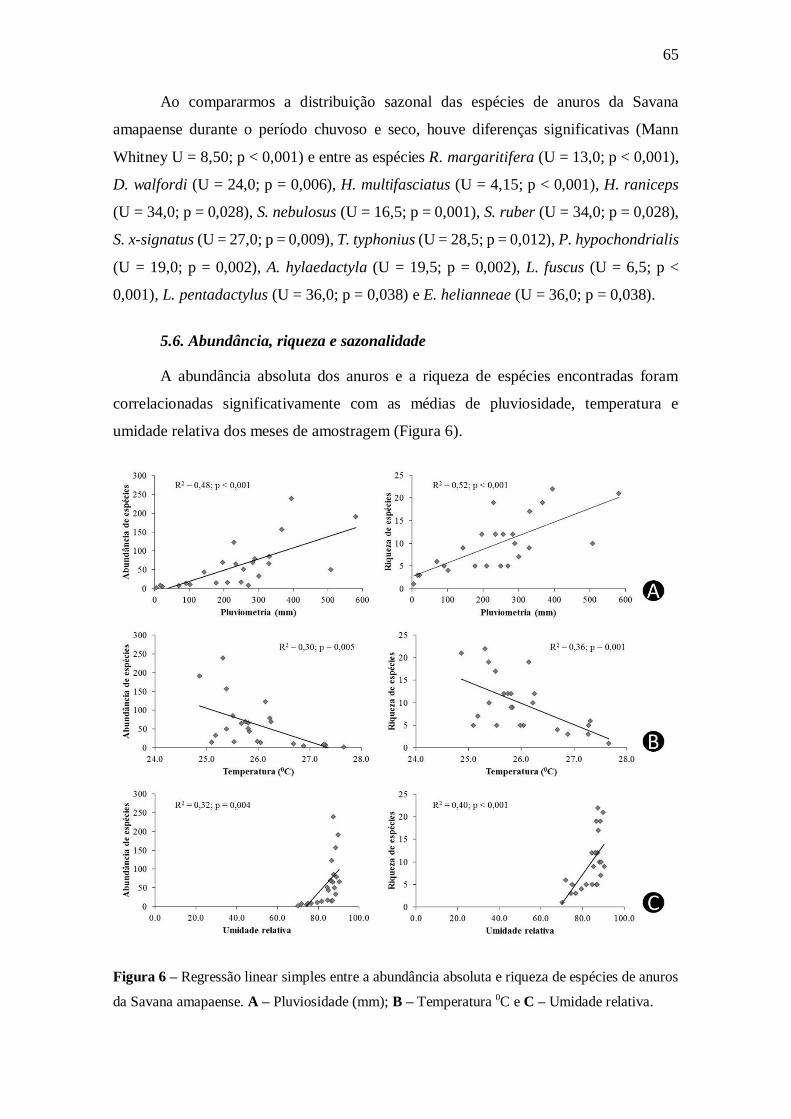
65
Ao compararmos a distribuição sazonal das espécies de anuros da Savana
amapaense durante o período chuvoso e seco, houve diferenças significativas (Mann
Whitney U = 8,50; p < 0,001) e entre as espécies R. margaritifera (U = 13,0; p < 0,001),
D. walfordi (U = 24,0; p = 0,006), H. multifasciatus (U = 4,15; p < 0,001), H. raniceps
(U = 34,0; p = 0,028), S. nebulosus (U = 16,5; p = 0,001), S. ruber (U = 34,0; p = 0,028),
S. x-signatus (U = 27,0; p = 0,009), T. typhonius (U = 28,5; p = 0,012), P. hypochondrialis
(U = 19,0; p = 0,002), A. hylaedactyla (U = 19,5; p = 0,002), L. fuscus (U = 6,5; p <
0,001), L. pentadactylus (U = 36,0; p = 0,038) e E. helianneae (U = 36,0; p = 0,038).
5.6. Abundância, riqueza e sazonalidade
A abundância absoluta dos anuros e a riqueza de espécies encontradas foram
correlacionadas significativamente com as médias de pluviosidade, temperatura e
umidade relativa dos meses de amostragem (Figura 6).
Figura 6 – Regressão linear simples entre a abundância absoluta e riqueza de espécies de anuros
da Savana amapaense. A – Pluviosidade (mm); B – Temperatura 0C e C – Umidade relativa.

66
Dentre as espécies, H. multifasciatus, H. punctatus, S. nebulosus, P.
hypochondrialis, A. hylaedactyla e L. fuscus tiveram sua abundância significativamente
correlacionada com a pluviosidade, temperatura e umidade relativa. As abundâncias de
R. major, R. marina, D. walfordi, S. fuscomarginatus, L. macrosternum e L. pentadactylus
não apresentaram correlação significativa com temperatura e umidade relativa (Tabela 9).
Tabela 9 – Valores da regressão linear simples e significância entre as abundâncias das espécies
de anuros e as variáveis climatológicas registradas na Savana amapaense. Os valores
significativos estão em negrito.
Espécies Pluviosidade Temperatura Umidade Relativa
R2
p R2 p R
2 P
Bufonidae
Rhinella major 0,28 0,008 0,09 0,088 0,86 0,163 Rhinella margaritifera 0,33 0,004 0,09 0,133 0,30 0,006
Rhinella marina 0,19 0,033 0,11 0,122 0,09 0,162
Hylidae Dendropsophus walfordi 0,37 0,002 0,16 0,054 0,11 0,122
Hypsiboas multifasciatus 0,42 0,001 0,30 0,006 0,48 < 0,001 Hypsiboas punctatus 0,30 0,005 0,28 0,008 0,18 0,038
Hypsiboas raniceps 0,27 0,009 0,22 0,021 0,15 0,061 Osteocephalus taurinus 0,30 0,006 0,19 0,033 0,11 0,107
Scinax fuscomarginatus 0,28 0,008 0,15 0,060 0,08 0,187
Scinax nebulosus 0,36 0,002 0,25 0,013 0,28 0,006 Scinax ruber 0,23 0,017 0,15 0,062 0,20 0,030
Scinax x-signatus 0,16 0,052 0,11 0,111 0,11 0,113
Trachycephalus thyphonius 0,23 0,018 0,15 0,062 0,21 0,023
Phyllomedusa hypochondrialis 0,51 < 0,001 0,37 0,002 0,39 0,001
Leptodactylidae
Pseudopaludicola boliviana 0,34 0,003 0,21 0,023 0,11 0,107
Adenomera hylaedactyla 0,27 0,009 0,18 0,038 0,21 0,024 Leptodactylus fuscus 0,42 0,001 0,26 0,010 0,38 0,001 Leptodactylus macrosternum 0,20 0,027 0,11 0,109 0,08 0,187
Leptodactylus pentadactylus 0,29 0,007 0,16 0,055 0,12 0,100 Leptodactylus podicipinus 0,09 0,147 0,07 0,202 0,04 0,320
Microhylidae
Elachistocleis helianneae 0,33 0,003 0,17 0,046 0,12 0,106
O período de atividade dos anuros esteve associado aos meses chuvosos, seguindo
a mesma variação sazonal encontrada em vários estudos com anuros da região
Neotropical (Toledo et al. 2003, Melo et al. 2007, Oda et al. 2009, Maffei et al. 2011). A
chuva tem relação direta com a umidade do ar, que é fator determinante na atividade dos
anuros, devido à pele permeável (Stebbins & Cohen 1994). Além de aumentar a umidade
do ar, a chuva disponibiliza ou renova os sítios reprodutivos, influenciando positivamente
a atividade de vocalização e reprodução das espécies de anuros (Goottsberger & Gruber
2004, Vasconcelos & Rossa-Feres 2005).

67
As taxocenoses de anuros respondem diretamente as modificações do hábitat,
podendo ter sua riqueza e abundância influenciados por diversas variáveis ambientais
(Ernst & Rödel 2008, Vasconcelos et al. 2009). No presente estudo, a pluviosidade foi a
variável que mais influenciou na variação sazonal dos anuros. Esses dados concordam
com Giaretta et al. (2008) e Kopp et al. (2010) que a riqueza e abundância das espécies
de anuros registradas no Cerrado são fortemente influenciadas pela pluviosidade e de que
a vocalização e reprodução ocorre principalmente durante as chuvas.
Alguns estudos têm destacado uma forte associação da pluviosidade com os
padrões de distribuição e estruturação da riqueza e abundância de espécies de anuros
(Vasconcelos et al. 2010, Hartel et al. 2011) sendo esta considerada o principal fator
extrínseco que controla a atividade reprodutiva (Prado et al. 2005).
5.7. Composição da dieta
Foram analisados um total de 796 estômagos, (Savana gramíneo lenhosa, N = 178;
Savana gramíneo lenhosa-arbustiva, N = 494; Savana parque, N = 124; Savana arbórea,
N = 72). Os estômagos das espécies T. typhonius, S. fuscomarginatus, P. boliviana e E.
helianneae não foram analisados por estarem vazios ou com digestão avançada.
Para as espécies de anuros da Savana gramíneo lenhosa foram registradas 12
categorias de presas, com valores de importância variando de 1,84 (Chilopoda para H.
multifasciatus) a 132,47 (Coleoptera para H. raniceps; Tabela 10). Para a Savana
gramíneo lenhosa-arbustiva, 15 categorias de presas foram identificadas, com o maior
valor de importância de 299,96 (Coleoptera para L. podicipinus; Tabela 11).
Tabela 10 – Índice de Importância de presas na dieta de anuros da Savana gramíneo lenhosa. Rm
= R. marina, Dw = D. walfordi, Hm = H. multifasciatus, Hr = H. raniceps, Ot = O. taurinus, Sf
= S. fuscomarginatus, Sn = S. nebulosus, Sr = S. ruber, Sx-s = S. x-signatus, Tt = T. typhonius,
Pb = P. boliviana, Ah = A. hylaedactyla, Lf = L. fuscus.
Espécies de anuros Presas Rm Dw Hm Hr Ot Sf Sn Sr Sx-s Tt Pb Ah LfAraneae 26,17 48,32 37,69 4,53 5,47Acari 51,73 3,08 4,76 9,97
Opilliones 32,69 9,24 78,28 3,11 Chilopoda 1,84 Odonata 5,80 10,04 Orthoptera 21,76 64,38 38,56 13,18 20,04 5,14Hemiptera 4,88 31,55 68,54 57,47 4,51 62,59Coleoptera 58,60 6,32 2,32 132,47 32,10 6,32 14,19Hymenoptera 32,53 6,12 13,25 Mollusca 17,35 1,87Nematoda 17,45 40,01LNI 9,43

68
Tabela 11 – Índice de Importância de presas na dieta de anuros da Savana gramíneo lenhosa-arbustiva. Rmj = R. major, Rm = R. marina, Dw = D. walfordi,
Hm = H. multifasciatus, Hp = H. punctatus, Hr = H. raniceps, Ot = O. taurinus, Sf = S. fuscomarginatus, Sn = S. nebulosus, Sr = S. ruber, Sx-s = S. x-signatus,
Tt = T. typhonius, Ph = P. hypochondrialis, Pb = P. boliviana, Ah = A. hylaedactyla, Lf = L. fuscus, Lpenta = L. pentadactylus, Lpod = L. podicipinus, Eh = E.
helianneae.
Presas Rmj Rm Dw Hm Hp Hr Ot Sf Sn Sr Sx-s Tt Ph Pb Ah Lf Lpenta Lpo EhAraneae 9,33 8,97 50,47 33,11 37,31 6,28 16,43 Acari 112,17 8,73 14,97 5,27 66,76 68,80 31,57 Opilliones 44,58 10,54 42,63 10,37 6,17 30,00 Chilopoda 9,17 Odonata 11,58 74,89 Orthoptera 8,67 95,64 22,37 10,82 42,37 10,21 10,28
Blattaria 11,36 Hemiptera 16,40 13,47 7,51 15,83 117,68 104,97 82,65 11,38 31,30 Coleoptera 1,36 8,73 129,87 5,45 47,26 19,27 11,50 28,36 28,38 46,26 299,96 Hymenoptera 12,08 84,53 10,98 61,18 17,94 Larva de Lepidoptera 10,41
Diptera 3,24 15,67 12,58 Mollusca 20,99 9,98 Nematoda 4,98 18,56 30,87 63,32 LNI 21,76 1,73
Espécies de anuros

69
Na Savana parque, foram identificadas 15 categorias de presas, com valor de
importância variando de 0,54 (Acari para H. multifasciatus) a 157,48 (Hemiptera para O.
taurinus). Para as espécies de anuros da Savana arbórea foram registradas oito categorias
de presas, com o menor valor de importância de 1,47 (Araneae para A. hylaedactyla) e o
maior de 152,47 (Hymenoptera para R. margaritifera; Tabela 12).
Tabela 12 – Índice de Importância de presas na dieta de anuros da Savana parque e da Savana
arbórea. Rmj = R. major, Rmarg = R. margaritifera, Hm = H. multifasciatus, Hp = H. punctatus,
Hr = H. raniceps, Ot = O. taurinus, Ah = A. hylaedactyla, Lm = L. macrosternum, Lpenta = L.
pentadactylus, Eh = E. helianneae.
Constataram-se diferenças na composição e número de presas na dieta dos anuros.
Isto certamente reflete a variedade de presas registradas e as estratégias de alimentação
utilizadas, ou seja, forrageadores do tipo senta e espera e forrageadores oportunistas (Solé
& Rödder 2010). Além disso, cada espécie de anuro apresentou um grupo de presa
específico e com maior representatividade em sua dieta. Estas diferenças nas estratégias
de forrageamento e na utilização dos recursos reduzem a competição, pois permite as
espécies capturar uma grande variedade de presas, o que pode funcionar como uma
estratégia de coexistência, refletindo o grau de diferenciação no uso do nicho trófico entre
as espécies (Duré & Kehr 2004, Araújo et al. 2009).
Representantes da família Bufonidae são considerados de hábito alimentar
generalista, com sua dieta variando de acordo com a disponibilidade de presas no habitat
(Batista et al. 2011, Maragno & Souza 2011). Entretanto, estudos que avaliaram a
disponibilidade de presas no habitat sugerem que algumas espécies do gênero Rhinella
Presas Rmj Hm Hp Hr Ot Lm Lpenta Eh Rmj Rmarg Ah Lm
Oligochaeta 155,70
Araneae 10,67 12,18 38,46 1,47
Acari 0,54 1,84 64,45 43,78 28,43 45,67
Opilliones 31,73 85,26 55,72 18,34
Chilopoda 3,69
Collembola 16,67
Odonata 5,67
Orthoptera 20,16 132,99 6,63
Blattaria 18,76
Hemiptera 22,18 92,15 157,48 78,16
Coleoptera 6,38 1,55 21,07 63,29 12,32 2,72 37,49 8,67 126,58
Hymenoptera 47,89 3,26 7,48 152,47 13,48
Diptera 2,36 89,00 4,27
Nematoda 4,37 16,50 19,13
LNI 27,72
Savana parque Savana arbórea

70
são capazes de selecionar suas presas, consumindo itens em maior ou menor proporção
do que a esperada em função da disponibilidade, sendo classificados como especialistas
em formigas e forrageadores ativos (Toft 1980, 1981, 1995, Sabagh et al. 2012). No
presente estudo, os bufonídeos apresentaram dieta constituída principalmente por
formigas e coleópteros, confirmando o hábito alimentar especialista.
Os hilídeos são predadores generalistas com estratégia de forrageamento
oportunista. Toft (1981) estudando as estratégias de forrageamento de anuros verificou
que espécies especialistas quanto a sua estratégia alimentar possuem preferência
alimentar por formigas e ácaros, e espécies generalistas evitam formigas e ácaros. No
presente estudo, a preferência alimentar dos hilídeos por Orthoptera e Hemiptera está de
acordo com estudo realizado por Toft (1981). Esse resultado pode estar relacionado com
a influência do habitat sobre a disponibilidade de presas. Esse tipo de dieta está
relacionado com o forrageio oportunista, onde suas presas são caracterizadas por
artrópodes mais ativos e de corpo mole (Simon & Toft 1991).
Os leptodactilídeos apresentam dieta generalista com estratégia de forrageamento
considerada intermediária entre ativa e senta e espera (Duré & Kehr 2004, Wells 2007).
No presente estudo, a dieta do gênero Leptodactylus foi constituída por Coleoptera e
Hemiptera corroborando com a estratégia senta e espera descrita para outros trabalhos
com o gênero (Solé et al. 2009, Sugai et al. 2012 Camera et al. 2014). No entanto, a
presença de outros artrópodes no conteúdo alimentar (Araneae, Acari e Opiliones) sugere
um comportamento alimentar oportunista e diferenças na disponibilidade de presas no
habitat ocupado pela população de leptodactilídeos (Solé & Rödder 2010).
Os anuros são tradicionalmente descritos como predadores generalistas, com
comportamento de forrageamento oportunista. Estudos sugerem que muitas espécies
apresentam algum grau de especialização na dieta (Simon & Toft 1991, Toft 1995). Toft
(1980, 1981) identificou dois principais padrões de dieta em anuros: os "especialistas" em
formigas que consomem mais formigas e ácaros e os "especialistas em não-formigas” que
comem uma maior quantidade de ortópteros e hemípteros. Essas especializações na dieta
estão intrinsecamente ligadas às estratégias de forrageamento (senta e espera ou busca
ativa), padrões de atividade noturno ou diurno, mecanismos de defesa, habitat ocupado e
a variabilidade e abundância sazonal dos recursos (Toft 1980, 1981, 1985).
Neste contexto, os bufonídeos são classificados como especialista em formigas,
Hylidae e Leptodactylidade como especialistas em não formigas. Seguindo essa linha, os
anuros registrados na Savana amapaense podem ser classificados em forrageadores

71
ativos, que consumiram presas pequenas e móveis em grande quantidade, forrageadores
passivos (senta e espera) que consumiram presas grandes e móveis em pouca quantidade
e predadores especialistas ou generalistas (Toft 1980, 1981, López et al. 2005, 2009).
Apesar desses dois tipos básicos de estratégias de forrageamento registrado para os
anuros, uma mesma espécie pode utilizar diferentes estratégias de acordo com a
disponibilidade de presas no ambiente. Isto permite a partilha de recursos pelas espécies
presentes na taxocenose, reduzindo a competição (Menin et al. 2005).
Para a Savana gramíneo lenhosa houve uma alta sobreposição de nicho alimentar
entre os pares de espécies S. nebulosus vs. S. x-signatus (0,89) e H. multifasciatus vs. S.
nebulosus (0,88). Os altos valores de sobreposição foram registrados entre as espécies
com dieta constituída preferencialmente por Araneae e Hemiptera. A ausência de
sobreposição ocorreu nos pares de espécies: R. marina vs. O. taurinus; R. marina vs. S.
x-signatus; D. walfordi vs. O. taurinus; D. walfordi vs. S. x-signatus; H. raniceps vs. S. x-
signatus; O. taurinus vs. S. x-signatus e O. taurinus vs. A. hylaedactyla (Tabela 13).
Tabela 13 – Sobreposição de nicho alimentar registrado pelos anuros da Savana gramíneo
lenhosa, durante o período de Janeiro de 2009 a Dezembro de 2010. Rm = R. marina, Dw = D.
walfordi, Hm = H. multifasciatus, Hr = H. raniceps, Ot = O. taurinus, Sn = S. nebulosus, Sr = S.
ruber, Sx-s = S. x-signatus, Ah = A. hylaedactyla, Lf = L. fuscus.
O alto valor de sobreposição de nicho registrado entre os hilídeos pode ser
resultado não apenas da proximidade taxonômica, mas também da distribuição vertical
dos seus indivíduos, cuja variação é muito pequena nos estratos verticais que ocupam
(Menin et al. 2005) e do hábito alimentar generalista (Toft 1981). De acordo com
Zimmerman & Simberloff (1996), a sobreposição da dieta é maior entre espécies
taxonomicamente semelhantes, devido estas compartilharem traços comportamentais,
Rmj Dw Hm Hr Ot Sn Sr Sx-s Ah Lf
Rhinella major 0,10 0,31 0,72 0 0,03 0,65 0 0,40 0,45
Dendropsophus walfordi 0,19 0,11 0 0,12 0,13 0 0,36 0,08
Hypsiboas multifasciatus 0,36 0,06 0,88 0,32 0,71 0,39 0,27
Hypsiboas raniceps 0,12 0,11 0,74 0 0,13 0,49
Osteocephalus taurinus 0,04 0,05 0 0 0,69
Scinax nebulosus 0,24 0,89 0,34 0,23
Scinax ruber 0,31 0,30 0,69
Scinax x-signatus 0,36 0,29
Adenomera hylaedactyla 0,23
Leptodactylus fuscus

72
fisiológicos e morfológicos. Nossos resultados corroboram essa teoria mostrando que os
maiores valores de sobreposição foram registrados entre espécies da mesma família
(Hylidae) e menores valores entre espécies de famílias diferentes.
A sobreposição de nicho alimentar para a Savana gramíneo lenhosa-arbustiva
variou entre 0 (R. marina vs. H. punctatus; R. major vs. O. taurinus; D. walfordi vs. O.
taurinus; H. punctatus vs. H. raniceps; H. raniceps vs. S. x-signatus; O. taurinus vs. L.
pentadactylus; S. nebulosus vs. L. podicipinus e S. x-signatus vs. L. pentadactylus) e 0,94
(D. walfordi vs. L. podicipinus) (Tabela 14).
Tabela 14 – Sobreposição de nicho alimentar registrado pelos anuros da Savana gramíneo
lenhosa-arbustiva, durante o período de Janeiro de 2009 a Dezembro de 2010. Rmj = R. major,
Rm = R. marina, Dw = D. walfordi, Hm = H. multifasciatus, Hp = H. punctatus, Hr = H. raniceps,
Ot = O. taurinus, Ph = P. hypochondrialis, Sn = S. nebulosus, Sr = S. ruber, Sx-s = S. x-signatus,
Ah = A. hylaedactyla, Lf = L. fuscus, Lpenta = L. pentadactylus, Lpod = L. podicipinus.
As espécies D. walfordi vs. L. podicipinus apresentaram um alto valor de
sobreposição mesmo sendo espécies de famílias distintas e possuindo tamanhos do corpo
diferentes. Isto pode estar associado ao fato dessas duas espécies terem apresentado
elevado consumo de indivíduos da ordem Coleoptera. É importante ressaltar que altos
valores nas sobreposições de nichos nem sempre indicam competição, já que a
abundância do recurso alimentar no habitat resulta em partição do mesmo sem prejuízos
ou perdas para as espécies consumidoras (Jiménez & Bolaños 2012).
Em taxocenoses de anuros, os padrões de utilização dos recursos alimentares estão
relacionados, principalmente, às habilidades plesiomórficas que são herdadas de suas
Rmj Rm Dw Hm Hp Hr Ot Sn Sr Sx-s Ph Ah Lf Lpenta Lpo
Rhinella major 0,92 0,25 0,12 0,09 0,12 0 0,42 0,05 0 0,51 0,75 0,06 0,14 0,27
Rhinella marina 0,28 0,24 0 0,19 0 0,47 0,05 0 0,39 0,77 0,08 0,41 0,30
Dendropsophus walfordi 0,56 0,18 0,41 0 0,02 0,16 0,15 0,52 0,30 0,35 0,48 0,94
Hypsiboas multifasciatus 0,57 0,66 0,31 0,45 0,56 0,59 0,61 0,49 0,63 0,54 0,45
Hypsiboas punctatus 0 0,26 0,26 0,54 0,84 0,70 0,46 0,60 0 0
Hypsiboas raniceps 0,39 0,15 0,38 0 0,38 0,09 0,33 0,41 0,44
Osteocephalus taurinus 0,06 0,26 0,09 0,14 0,02 0,80 0 0
Scinax nebulosus 0,14 0,26 0,30 0,59 0,22 0,19 0
Scinax ruber 0,78 0,49 0,17 0,28 0,09 0,17
Scinax x-signatus 0,49 0,44 0,42 0 0
Phyllomedusa hypochondrialis 0,57 0,40 0,23 0,44
Adenomera hylaedactyla 0,37 0,21 0,15
Leptodactylus fuscus 0,11 0,21
Leptodactylus pentadactylus 0,51
Leptodactylus podicipinus

73
respectivas linhagens e que surgiram concomitantemente com a divergência das famílias
(Toft 1980) ou pelas interações ecológicas contemporâneas.
Na sobreposição de nicho quanto a composição da dieta para a Savana parque, os
maiores valores foram entre os pares de espécies H. multifasciatus vs. H. punctatus (0,75)
e H. punctatus vs. O. taurinus (0,78). Os maiores valores de sobreposição da dieta foram
registrados entre as espécies com dieta predominantemente de hemípteros. A ausência de
sobreposição ocorreu nos pares de espécies: R. major vs. O. taurinus; H. punctatus vs. H.
raniceps e O. taurinus vs. L. pentadactylus (Tabela 15).
Tabela 15 – Sobreposição de nicho alimentar registrado pelos anuros da Savana parque, durante
o período de Janeiro de 2009 a Dezembro de 2010. Rmj = R. major, Hm = H. multifasciatus, Hp
= H. punctatus, Hr = H. raniceps, Ot = O. taurinus, Lm = L. macrosternum, Lpenta = L.
pentadactylus.
Para a Savana arbórea não houve ausência de sobreposição de nicho alimentar
para as espécies de anuros. Os maiores valores foram registrados para os pares de espécies
R. margaritifera vs. A. hylaedactyla (0,76) que apresentaram dieta constituída por
coleópteros. Os pares de espécies R. major vs. R. margaritifera, R. major vs. A.
hylaedactyla e R. major vs. L. macrosternum apresentaram valores de sobreposição
baixos, variando de 0,14 a 0,29 (Tabela 16).
Tabela 16 – Sobreposição de nicho alimentar registrado pelas espécies de anuros da Savana
arbórea, Savana amapaense, durante o período de Janeiro de 2009 a Dezembro de 2010. Rmj =
R. major, Rmarg. = R. margaritifera, Ah = A. hylaedactyla, Lm = L. macrosternum.
Rmj Hm Hp Hr Ot Lm Lpenta
Rhinella major 0,08 0,10 0,05 0 0,11 0,03
Hypsiboas multifasciatus 0,75 0,45 0,73 0,19 0,27
Hypsiboas punctatus 0 0,78 0,05 0
Hypsiboas raniceps 0,06 0,49 0,67
Osteocephalus taurinus 0 0
Leptodactylus macrosternum 0,29
Leptodactylus pentadactylus
Rmj Rmarg Ah Lm
Rhinella major 0,29 0,24 0,14
Rhinella margaritifera 0,76 0,43
Adenomera hylaedactyla 0,48
Leptodactylus macrosternum

74
O grau de especialização para amplitude de nicho no uso de recursos da dieta
variou entre as espécies (0 < Bp ≤ 1). As espécies com maiores amplitudes de nicho
trófico, em ordem decrescente, foram: S. x-signatus (Savana gramíneo lenhosa); D.
walfordi (Savana gramíneo lenhosa-arbustiva); L. macrosternum (Savana parque) e R.
major (Savana arbórea). Das espécies de anuros registradas, 67,5% foram generalistas e
32,5% especialistas (Tabela 17).
Tabela 17 – Amplitude de nicho alimentar para as espécies de anuros registradas Savana
amapaense, durante o período de Janeiro de 2009 a Dezembro de 2010. SGL = Savana gramíneo
lenhosa; SGLA = Savana gramíneo lenhosa-arbustiva; SP = Savana parque; SA = Savana arbórea.
Bp = Índice padronizado de Levins.
Os modelos nulos mostraram que a sobreposição média da dieta entre as espécies
de anuros da Savana amapaense foi significativamente diferente entre os valores
observado e esperado pelo modelo nulo (média observada = 0,25; p < 0,001), indicando
a existência de estrutura com relação a dieta (Figura 7).
Espécies N Bp N Bp N Bp N Bp
Rhinella major 21 0,38 13 0,18 9 0,78
Rhinella marina 20 0,80 44 0,54
Rhinella margaritifera 32 0,53
Dendropsophus walfordi 16 0,24 52 0,99
Hypsiboas multifasciatus 16 0,48 63 0,90 38 0,51
Hypsiboas punctatus 29 0,94 20 0,41
Hypsiboas raniceps 25 0,86 14 0,80 17 0,63
Osteocephalus taurinus 11 0,75 14 0,72 20 0,17
Scinax nebulosus 16 0,48 41 0,43
Scinax ruber 20 0,69 25 0,50
Scinax x-signatus 10 0,92 30 0,77
Phyllomedusa hypochondrialis 72 0,62
Adenomera hylaedactyla 13 0,84 28 0,60 17 0,39
Leptodactylus fuscus 31 0,38 42 0,58
Leptodactylus macrosternum 5 1 14 0,64
Leptodactylus pentadactylus 8 0,91 11 0,41
Leptodactylus podicipinus 12 0
Fitofisionomias
SGL SGLA SP SA
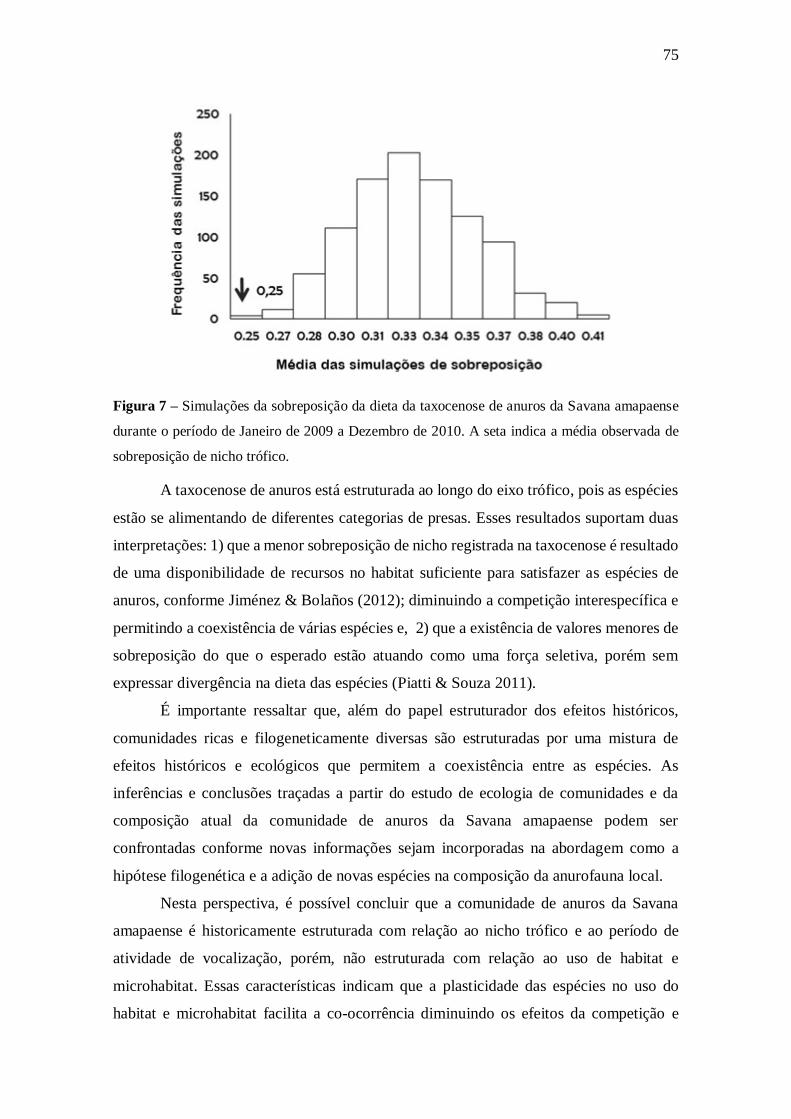
75
Figura 7 – Simulações da sobreposição da dieta da taxocenose de anuros da Savana amapaense
durante o período de Janeiro de 2009 a Dezembro de 2010. A seta indica a média observada de
sobreposição de nicho trófico.
A taxocenose de anuros está estruturada ao longo do eixo trófico, pois as espécies
estão se alimentando de diferentes categorias de presas. Esses resultados suportam duas
interpretações: 1) que a menor sobreposição de nicho registrada na taxocenose é resultado
de uma disponibilidade de recursos no habitat suficiente para satisfazer as espécies de
anuros, conforme Jiménez & Bolaños (2012); diminuindo a competição interespecífica e
permitindo a coexistência de várias espécies e, 2) que a existência de valores menores de
sobreposição do que o esperado estão atuando como uma força seletiva, porém sem
expressar divergência na dieta das espécies (Piatti & Souza 2011).
É importante ressaltar que, além do papel estruturador dos efeitos históricos,
comunidades ricas e filogeneticamente diversas são estruturadas por uma mistura de
efeitos históricos e ecológicos que permitem a coexistência entre as espécies. As
inferências e conclusões traçadas a partir do estudo de ecologia de comunidades e da
composição atual da comunidade de anuros da Savana amapaense podem ser
confrontadas conforme novas informações sejam incorporadas na abordagem como a
hipótese filogenética e a adição de novas espécies na composição da anurofauna local.
Nesta perspectiva, é possível concluir que a comunidade de anuros da Savana
amapaense é historicamente estruturada com relação ao nicho trófico e ao período de
atividade de vocalização, porém, não estruturada com relação ao uso de habitat e
microhabitat. Essas características indicam que a plasticidade das espécies no uso do
habitat e microhabitat facilita a co-ocorrência diminuindo os efeitos da competição e

76
permitindo a coexistência local de espécies com distintas exigências ecológicas,
sobretudo no uso dos recursos alimentares e período de atividade, e foram determinantes
na gênese da comunidade de anuros da Savana amapaense.
Estes exemplos ilustram como as variações existentes na estruturação das
comunidades podem resultar de diferenças nas condições ambientais ecológicas e,
também, de diferenças históricas entre as linhagens evolutivas que as compõem.
Adicionalmente, a heterogeneidade ambiental em escala regional, determinada pela
dinâmica espacial e temporal das fitofisionomias registradas na Savana amapaense,
exerce grande influência na variação da riqueza e diversidade de espécies da região.
Além disso, os resultados aqui registrados indicam que o ambiente está agindo
como filtro sobre as espécies, que além de exibirem claramente relação entre o uso do
ambiente e as características históricas, não estão aptas a utilizar as mesmas categorias de
habitats e microhabitats disponíveis. Assim, a ausência de estrutura na comunidade pode
ser atribuída à grande disponibilidade de recursos partilhados pelas espécies, o que
diminui a pressão de competição entre elas.
Os padrões de estruturação das comunidades de anuros em áreas de Savana
amazônica ainda não foram estudados e poucos são os estudos com estrutura de
comunidade de anuros que ocorrem nas diferentes fitosionomias desse bioma. Assim
sendo, a ampliação de estudos sobre a anurofauna nas diversas Ecorregiões buscando
entender como as comunidades estão interagindo no tempo e espaço, ajudarão a traçar
planos de manejo para a conservação de Áreas Prioritárias das Savanas amazônicas.
6. REFERÊNCIAS BIBLIOGRÁFICAS
Abrunhosa, P.A., Wogel, H. & Pombal Jr., J.P. 2006. Anuran temporal occupancy in
a temporary pond from the Atlantic Rain Forest, South-Eastern Brazil. Herpetological
Journal, 16, 115-122.
Afonso, L.G. & Eterovick, P.C. 2007. Spatial and temporal distribution of breeding
anurans in streams in Southeastern Brazil. Journal of Natural History, 41 (13-16), 949-
963.
Aguiar, L.M.S., Machado, R.B. & Marinho-Filho, J. 2004. A diversidade biológica do
Cerrado. In: Cerrado: ecologia e caracterização (Org. por L.M. Aguiar & A.J.A.
Camargo), pp. 17-40. Brasília: Embrapa Cerrados.

77
Alvares, C.A., Stape, J.L., Sentelhas, P.C., Gonçalves, J.L.M. & Sparovek, G. 2013.
Köppen’s climate classification map for Brazil. Meteorologische Zeitschrift, 22 (6),
711-728.
Amézquita, A. & Hödl, W. 2004. How, when, and where to perform visual displays:
The case of the Amazonian frog Hyla parviceps. Herpetologica, 60, 420-429.
Andrew, S. & Christensen, B. 2001. Optimal diet theory: when does it work, and when
and why does it fail? Animal Behavior, 61, 379-390.
Araújo, C.O., Condez, T.H. & Sawaya, R.J. 2009. Anfíbios Anuros do Parque Estadual
das Furnas do Bom Jesus, sudeste do Brasil, e suas relações com outras taxocenoses
no Brasil. Biota Neotropica, 9 (2), 77-98.
Araújo, C.O. & Almeida-Santos, S.M. 2011. Herpetofauna de um remanescente de
cerrado no estado de São Paulo, sudeste do Brasil. Biota Neotropica, 11 (3), 47-62.
Araújo, C.O., Corrêa, D.T. & Almeida-Santos, S.M. 2013. Anuros da Estação
Ecológica de Santa Bárbara, um remanescente de formações abertas de Cerrado no
estado de São Paulo. Biota Neotropica, 13 (3), 230-240.
Ayres, M., Ayres Jr., M.A., Ayres, D.L. & Dos Santos, A.S. 2007. BioEstat: aplicações
estatísticas nas áreas das ciências biológicas e médicas. Belém: Publicações Avulsas
Mamirauá.
Barbosa, R.I., Nascimento, S.P., Amorim, P.A.F. & Silva, R.F. 2005. Notas sobre a
composição arbóreo-arbustiva de uma fisionomia das savanas de Roraima, Amazônia
Brasileira. Acta Botanica Brasilica, 19 (2), 323-329.
Batista, R.C., Brito-de-Carvalho, C., Freitas, E.B., Franco, S.C., Batista, C.C.,
Coelho, W.A. & Faria, R.G. 2011. Diet of Rhinella schneideri (Werner, 1894)
(Anura: Bufonidae) in the Cerrado, Central Brazil. Herpetological Notes, 4, 17-21.
Becker, C.G., Fonseca, C.R., Haddad, C.F.B. & Prado, P.I. 2010. Habitat split as a
cause of local population declines of amphibians with aquatic larvae. Conservation
Biology, 24, 287-294.
Bertoluci, J. & Rodrigues, J.T. 2002. Seasonal patterns of breeding activity of Atlantic
Rainforest anurans at Boracéia, Southeastern Brazil. Amphibia-Reptilia, 23: 161-167.
Bitar, Y.O.C., Pinheiro, L.P.C., Abe, P.S. & Santos-Costa, M.C. 2012. Species
composition and reproductive modes of anurans from a transitional Amazonian forest,
Brazil. Zoologia, 29 (1), 19-26.

78
Both, C., Kaefer, I.L., Santos, T.G. & Cechin, S.T.Z. 2008. An austral assemblage in
the Neotropics: seasonal occurrence correlated with photoperiod. Journal of Natural
History, 42 (3-4), 205-222.
Brasileiro C.A., Sawaya R. J., Kiefer M.C. & Martins M. 2005. Amphibians of an
open Cerrado fragment in Southeastern Brazil. Biota Neotropica, 5 (2), 93-109.
Camera, B.F., Krinski, D. & Calvo, I.A. 2014. Diet of the Neotropical frog
Leptodactylus mystaceus (Anura: Leptodactylidae). Herpetology Notes, 7, 31-36.
Campos, F.S. & Vaz-Silva, W. 2010. Distribuição espacial e temporal da anurofauna em
diferentes ambientes no município de Hidrolândia, GO, Brasil Central. Neotropical
Biology and Conservation, 5 (3), 179-187.
Campos, V.A., Oda, F.H., Juen, L., Barth, A. & Dartora, A. 2013. Composição e
riqueza de espécies de anfíbios anuros em três diferentes habitat em um agrossistema
no Cerrado do Brasil central. Biota Neotropica, 13 (1), 124-132.
Canavero, A., Arim M. & Brazeiro, A. 2009. Geographic variations of seasonality and
coexistence in communities: The role of diversity and climate. Austral Ecology, 34:
741-750.
Cassemiro, F.A.S., Gouveia, S.F. & Diniz-Filho, J.A.F. 2012. Distribuição de Rhinella
granulosa: integrando envelopes bioclimáticos e respostas ecofisiológicas. Revista da
Biologia, 8, 38-44.
Cavalcanti, R.B. & Joly, C.A. 2002. Biodiversity and conservation priorities in the
Cerrado Region. In: The Cerrados of Brazil: Ecology and Natural History of a
Neotropical Savanna (Org. por P.S. Oliveira & R.J.Marquis), pp. 351-367. Nova York:
Columbia University Press.
Colli, G.R., Bastos, R.P. & Araújo, A.F.B. 2002. The character and dynamics of the
Cerrado herpetofauna. In: The Cerrados of Brazil: Ecology and Natural History of a
Neotropical Savanna. (Org. por P.S.Olivera & R.J.Marquis), pp. 223-241. New York:
Columbia University Press.
Conte, C.E., Silva, D.R. & Rodrigues, A.P. 2013. Anurofauna da bacia do Rio Tijuco,
Minas Gerais, Brasil e sua relação com taxocenoses de anfíbios do Cerrado e suas
transições. Iheringia, 103 (3), 280-288.
Costa, W.P., Almeida, S.C. & Jim, J. 2013. Anurofauna em uma área na Depressão
Periférica, no centro-oeste do estado de São Paulo, Brasil. Biota Neotropica, 13 (2),
163-174.

79
Crawford, J.A., Shepard, D.B. & Conner, C.A. 2009. Diet composition and overlap
between recently metamorphosed Rana areolata and Rana sphenocephala:
implications for a frog of conservation concern. Copeia, 2009 (4), 642-646.
Crump, M.L. & Scott, N.J.J. 1994. Visual encounter surveys. In: Measuring and
monitoring biological diversity: standard methods for amphibians, (Org. por, W.R.
Heyer., M.A. Donnelly., R.W. McDiarmid., L.A.C. Hayek & M.S. Foster), pp. 84-92.
Washington: Smithsonian Institution Press.
Duré, M.I. & Kehr, A.I. 2004. Influence of microhabitat on the trophic rcology of two
Leptodactylids from Northeastern Argentina. Herpetologica, 60 (3), 295-303.
Ernst, R. & Rödel, M. 2005. Anthropogenically induced changes of predictability in
tropical anuran assemblages. Ecology, 86, 3111-3118.
Ernst, R. & Rödel, M. 2008. Patterns of community composition in two tropical tree
frog assemblages: separating spatial structure and environmental effects in disturbed
and undisturbed forests. Journal of Tropical Ecology, 24 (2), 111-120.
Ernst, R., Linsenmair, E. & Rödel, M. 2006. Diversity erosion beyond the species level:
dramatic loss of functional diversity after selective logging in two tropical amphibians
communities. Biological Conservation, 133, 143-155.
Gambale, P.G., Woitovicz-Cardoso, M., Vieira, R.R., Batista, V.G., Ramos, J. &
Bastos, R.P. 2014. Composição e riqueza de anfíbios anuros em remanescentes de
Cerrado do Brasil Central. Iheringia, 104 (1), 50-58.
Giaretta, A.A., Menin, M., Facure, K.G., Kokubum, M.N.C. & De Oliveira Filho,
J.C. 2008. Species richness, relative abundance, and habitat of reproduction of
terrestrial frogs in the Triangulo Mineiro region, Cerrado biome, southeastern Brazil.
Iheringia, 98 (2), 181-188.
Goottsberger, B. & Gruber, E. 2004. Temporal partitioning of reproductive activity in
a neotropical anuran Community. Journal of Tropical Ecology, 20, 271-280.
Gotelli, N. & Colwell, R.K. 2001. Quantifying biodiversity: Procedures and pitfalls in
the measurement and comparison of species richness. Ecology Letters, 4: 379-391.
Gotelli N.J. & Entsminger, G.L. 2005. EcoSim: Null Models Software for Ecology.
Version 7.72. Encontrado em 10 de setembro de de 2013.
http://together.net/~gentsmin/ecosim.htm
Haddad, C.F.B. & Prado, C.P.A. 2005. Reproductive modes in frogs and their
unexpected diversity in the Atlantic forest of Brazil. BioScience, 55 (3), 207-217.

80
Hammer, O., Harper, D.A.T. & Rian, P.D. 2001. Past: Palaeonthological statistics
software package for education and data analysis. Version. 1.37. Encontrado em 12 de
novembro de 2012. http://palaeo-electronica.org/2001_1/past/issue1_01.htm
Hartel, T., Bancila, R. & Cogalniceanu, D. 2011. Spatial and temporal variability of
aquatic habitat use by amphibians in a hydrologically modified landscape. Freshwater
Biology, 56, 2288-2298.
Heyer, W.R., Donnelly, M.A., Mcdiarmid, R.W., Hayek, L.C. & Foster, M.S. 1994.
Measuring and monitoring biological diversity: standard methods for amphibians.
Washington: Smithsonian Institution.
Hödl, W. 1990. Reproductive diversity in Amazonian lowland frogs. Fortschritte der
Zoologie, 38, 41-60.
Jiménez, R. & Bolaños, F. 2012. Use of food and spatial resources by two frogs of the
genus Dendropsophus (Anura: Hylidae) from La Selva, Costa Rica. Phyllomedusa, 11
(1), 51-62.
Kolodiuk, M.F., Ribeiro, L.B. & Freire, E.M.X. 2010. Diet and foraging behavior of
two species of Tropidurus (Squamata, Tropiduridae) in the Caatinga of northeastern
Brazil. South American Journal of Herpetology, 5 (1), 35-44.
Kopp, K. & Eterovick P.C. 2006. Factors influencing spatial and temporal structure of
frog assemblages at ponds in southeastern Brazil. Journal of Natural History, 40 (29-
31): 1813-1830.
Kopp, K., Signorelli, L. & Bastos, R.P. 2010. Distribuição temporal e diversidade de
modos reprodutivos de anfíbios anuros no Parque Nacional das Emas e entorno, estado
de Goiás, Brasil. Iheringia, 100 (3), 192-200.
Krebs, C.J. 2014. Ecological methodology. 3a ed. Vancouver: University of British
Columbia.
Lima, A.P. & Magnusson, W.E. 2000. Does foraging activity change with the
ontogeny? An assessment for six sympatric species of postmetamorphic litter anurans
in Central Amazonia. Journal of Herpetology, 34 (2), 192-200.
López, J.A., Peltzer, M.P. & Lajmanovich, R.C. 2005. Dieta y solapamiento del
subnicho trófico de nueve espécies de leptodactílidos en el Parque General San Martín
(Argentina). Revista Española de Herpetología, 19, 19-31.
López, J. A., Scarabotti, P.A., Medrano, M.C. & Ghirardi, R. 2009. Is the red spotted
Green frog Hypsiboas punctatus (Anura: Hylidae) selecting it’s prey? The importance
of prey availability. Revista de Biología Tropical, 57, 847-857.

81
Maffei, F., Ubaid, F.K. & Jim, J. 2011. Anurans in an open cerrado area in the
municipality of Borebi, Sao Paulo state, Southeastern Brazil: habitat use, abundance
and seasonal variation. Biota Neotropica, 11, 221-233.
Magnusson, W.E., Lima, A.P., Silva, W.A. & Araújo, M.C. 2003. Use of geometric
forms to estimate volume of invertebrates in ecological studies of dietary overlap.
Copeia, 2003, 13‑19.
Maragno, F.P. & Souza, F.L. 2011. Diet of Rhinella scitula (Anura, Bufonidae) in the
Cerrado, Brazil: the importance of seasons and body size. Revista Mexicana de
Biodiversidad, 82, 879-886.
Melém-Júnior, N.J., Farias-Neto, J.T. & Yokomizo, G.K.I. 2003. Caracterização dos
Cerrados do Amapá. Comunicado Técnico Embrapa, 105, 1-5.
Melo, G.V., Rossa-Feres, D.C. & Jim, J. 2007. Variação temporal no sítio de
vocalização em uma comunidade de anuros de Botucatu, Estado de São Paulo, Brasil.
Biota Neotropica, 7 (2), 93-102.
Melo, M., Fava, F., Pinto, H.B.A., Bastos, R.P. & Nomura, F. 2013. Anuran diversity
(Amphibia) in the Extractivist Reserve Lado do Cedro, Goiás. Biota Neotropica, 13
(2), 205-217.
Menin, M., Rossa-Feres, D.C. & Giaretta, A. 2005. Resource use and coexistence of
two syntopic hylid frogs (Anura: Hylidae). Revista Brasileira de Zoologia, 22 (1), 61-
72.
Miranda, I.S., Absy, M.L. & Rebêlo, G.H. 2003. Community Structure of Woody
Plants of Roraima Savannahs, Brazil. Plant Ecology, 164, 109-123.
Moraes, R.A., Sawaya, R.J. & Barrella, W. 2007. Composição e diversidade de
anfíbios anuros em dois ambientes de Mata Atlântica no Parque Estadual Carlos
Botelho, São Paulo, sudeste do Brasil. Biota Neotropica, 7 (2), 27-36.
Morais, A.R., Signorelli, L., Gambale, P.G., Kopp, K., Nomura, F., Guimarães, L.D.,
Vaz-Silva, W., Ramos, J. & Bastos, R.P. 2011. Anfíbios anuros associados a corpos
d’água do sudoeste do estado de Goiás, Brasil. Biota Neotropica, 11 (3), 355-363.
Neckel-Oliveira, S., Magnusson, W.E. & Lima, A.P. 2000. Diversity and distribution
of frogs in an Amazonian savanna in Brazil. Amphibia-Reptilia, 21 (3), 317-326.
Nomura, F., Maciel, N.M., Pereira, E.B. & Bastos, R.P. 2012. Diversidade de anuros
(Amphibia) em áreas recuperadas de atividade mineradora e de plantio de Eucalyptus
urophyla, no Brasil Central. Bioscience Journal, 28 (2), 312-324.

82
Oda, F.H., Bastos, R.P. & Sá Lima, M.A.D.C. 2009. Taxocenose de anfíbios anuros no
Cerrado do Alto Tocantins, Niquelândia, Estado de Goiás: diversidade, distribuição
local e sazonalidade. Biota Neotropica, 9 (4): 219-232.
Oliveira Filho, A.T. & Ratter, J.A. 2002. Vegetation physiognomies and woody flora
of the cerrado biome. In: Ecology and Natural History of a Neotropical Savanna (Org.
por P.S. Oliveira. & R.J. Marquis), pp. 91-120. Nova York: Columbia University
Press.
Pianka, E.R. 1973. The structure of lizard communities. Annual Review of Ecology and
Systematics, 4, 53-74.
Pianka, E.R. 1986. Ecology and natural History of Desert Lizards – Analysis of the
Ecological Niche and Community Structure. New Jersey: Princeton University Press.
Piatti, L. & Souza F.L. 2011. Diet and resource partitioning among anurans in irrigated
rice fields in Pantanal, Brazil. Brazilian Journal of Biology, 71, 653-661.
Pinheiro, L.C., Bitar, Y.O.C., Galatti, U., Neckel-Oliveira, S. & Santos-Costa, M.C.
2012. Amphibians from southeastern state of Pará: Carajás Region, northern Brazil.
Check List, 8 (4), 693-702.
Pirani, R.M., Nascimento, L.B. & Feio, R.N. 2013. Anurans in a forest remnant in the
transition zone between cerrado and atlantic rain forest domains in southeastern Brazil.
Anais da Academia Brasileira de Ciências, 85 (3), 1093-1104.
Pombal Jr., J.P. & Haddad, C.F.B. 2007. Estratégias e modos reprodutivos em anuros.
In: Herpetologia no Brasil II (Org. por L.B. Nascimento & P.M.E. Oliveira), pp. 101-
116. Belo Horizonte: Sociedade Brasileira de Herpetologia.
Prado, C.P.A., Uetanabaro, M. & Haddad, C.F.B. 2005. Breeding activity patterns,
reproductive modes, and habitat use by anurans (Amphibia) in a seasonal environment
in the Pantanal, Brasil. Amphibia-Reptilia, 26: 211-221.
Prado, G.M. & Pombal Jr. J.P. 2005. Distribuição espacial e temporal dos anuros em
um brejo da Reserva Biológica de Duas Bocas, sudeste do Brasil. Arquivos do Museu
Nacional, 63 (4), 685-705.
Rabelo, B.V., Pinto, A.C.; Simas, A.P.S.C., Tardin, A.T., Fernandes, A.V., Souza,
C.B., Monteiro, E.M.P.B., Facundes, F.S., Ávila, J.E.S., Souza, J.S.A., Costa-
Guedes, L.A., Penha, O.A.A., Melo, R.M.S. & Gibson, V.M. 2008.
Macrodiagnóstico do Estado do Amapá: primeira aproximação do ZEE. Macapá:
Instituto de Pesquisas Científicas e Tecnológicas do Estado do Amapá (IEPA).

83
Ribeiro-Júnior, J.W., Lima, A.P. & Magnusson, W.E. 2012. The Effect of Riparian
Zones on Species Diversity of Frogs in Amazonian Forests. Copeia, 2012 (3), 375-
381.
Rossa-Feres, D.C. & Jim, J. 2001. Similaridade no sítio de vocalização em uma
comunidade de anfíbios anuros na região noroeste do Estado de São Paulo, Brasil.
Revista Brasileira de Zoologia, 18 (2), 439-454.
Sabagh, L.T., Mello, R.S. & Rocha, C.F.D. 2012. Food niche overlap between two
sympatric leaf-litter frog species from Central Amazonia. Zoologia, 29 (1), 95-98.
Santos, T.G., Rossa-Feres, D.C. & Casatti, L. 2007. Diversidade e distribuição espaço-
temporal de anuros em região com pronunciada estação seca no sudeste do Brasil.
Iheringia, 97 (1), 37-49.
Santos, T.G., Kopp, K., Spies, M. R., Trevisan, R. & Cechin, S. Z. 2008. Temporal
and spatial distribution of anurans in the Pampa Region (Santa Maria, RS). Iheringia,
98, 244-253.
Silva, F.R. & Rossa-Feres, D.C. 2007. Uso de fragmentos florestais por anuros
(Amphibia) de área aberta na região noroeste do Estado de São Paulo. Biota
Neotropica, 7 (2), 141-147
Silva, F.R., Gibbs, J.P. & Rossa-Feres, D.C. 2011. Breeding Habitat and Landscape
Correlates of Frog Diversity and Abundance in a Tropical Agricultural Landscape.
Wetlands, 31, 1079-1087.
Silva, R.A., Martins, I.A. & Rossa-Feres, D.C. 2011. Environmental heterogeneity:
Anuran diversity in homogeneous environments. Zoologia, 28 (5), 610-618.
Simon, M.P. & Toft, C.A. 1991. Diet specialization in small vertebrates: mite-eating in
frogs. Oikos, 61, 263-278.
Solé. M., Dias, I.R., Rodrigues, E.A.S., Marciano-JR, E., Branco, S.M.J.,
Cavalcante, K.P. & Rödder, D. 2009. Diet of Leptodactylus ocellatus (Anura:
Leptodactylidae) from a cacao plantation in southern Bahia, Brazil. Herpetology
Notes, 2, 9-15.
Solé, M. & Rödder, D. 2010. Dietary Assessments of adult amphibians. In: Amphibian
Ecology and Conservation: a handbook of techniques (Org. por C.K. Dodd), pp. 167-
184. Oxford: Oxford University Press.
Stebbins, R.C. & Cohen, N.W. 1997. A Natural History of Amphibians. New Jersey:
Princeton University Press.

84
Steen, D.A., Mcclure, C.J.W. & Graham, S.P. 2013. Relative influence of weather and
season on anuran calling activity. Canadian Journal of Zoology, 91 (7), 462-467.
Sugai, J.L.M.M., Terra, J.S. & Ferreira, V.L. 2012. Diet of Leptodactylus fuscus
(Amphibia: Anura: Leptodactylidae) in the Pantanal of Miranda river, Brazil. Biota
Neotropica, 12 (1), 99-104.
Tews, J., Brose, U., Grimm, V., Tielborger, K., Wichmann, M.C., Schwager, M. &
Jeltsch, F. 2004. Animal species diversity driven by habitat heterogeneity/diversity:
the importance of keystone structures. Journal of Biogeography, 31, 79-92.
Toft, C.A. 1980. Feeding ecology of thirteen syntopic species of anurans in a seasonal
tropical environment. Oecologia, 45, 131-141.
Toft, C.A. 1981. Feeding ecology of Panamanian litter anurans: patterns in diet and
foraging mode. Journal of Herpetology, 15, 139-144.
Toft, C.A. 1985. Resource Partitioning in Amphibians and Reptiles. Copeia, 1985, 1-20.
Toft, C.A. 1995. Evolution of diet specialization in poison-dart frogs (Dendrobatidae).
Herpetologica, 51, 202-216.
Toledo, L.F., Zina, J. & Haddad, C.F.B. 2003. Distribuição espacial e temporal de uma
comunidade de anfíbios anuros do Município de Rio Claro, São Paulo, Brasil. Holos
Environment, 3 (2), 136-149.
Valdujo, P.H., Camacho, A., Recoder, R.S., Júnior, M.T., Ghellere, J.M.B., Mott,
T., Nunes, P.M.S., Nogueira, C. & Rodrigues, M.T. 2011. Anfíbios da Estação
Ecológica Serra Geral do Tocantins, região do Jalapão, Estados do Tocantins e
Bahia. Biota Neotropica, 11 (1), 251-262.
Valdujo, P.H., Silvano, D.L., Colli, G.R. & Martins, M. 2012. Anuran Species
Composition and Distribution Patterns in Brazilian Cerrado, a Neotropical Hotspot.
South American Journal of Herpetology, 7 (2), 63-78.
Vasconcelos, T.S. & Rossa-Feres, D.C. 2008. Habitat heterogeneity and use of physical
and acoustic space in anuran communities in southeastern Brazil. Phyllomedusa, 7,
125-140.
Vasconcelos, T.S., Santos, T.G., Rossa-Feres, D.C. & Haddad, C.F.B. 2009. Influence
of the environmental heterogeneity of breeding ponds on anuran assemblages from
southeastern Brazil. Canadian Journal Zoology, 87, 699-707.
Vasconcelos, T.S., Santos T.G., Rossa-Feres, D.C. & Haddad, C.F.B. 2011. Spatial
and temporal distribution of tadpole assemblages (Amphibia, Anura) in a seasonal dry
tropical forest of southeastern Brazil. Hydrobiologia, 673, 93-104.

85
Vasconcelos, T.S., Santos, T.G., Haddad, C.F.B. & Rossa-Feres, D.C. 2010. Climatic
variables and altitude as predictors of anuran species richness and number of
reproductive modes in Brazil. Journal of Tropical Ecology, 26, 423-432.
Vitt, L.J., Sartorius, S.S., Ávila-Pires, T.C.S. & Espósito, M.C. 2001. Life on the leaf
litter: the ecology of Anolis nitens tandai in the Brazilian Amazon. Copeia, 2001 (2),
401-412.
Wells, K.D. 2007. The Ecology and Behavior of Amphibians. Chicago and London: The
University of Chicago Press.
Werneck, F.P., Colli, G.R. & Vitt, L.J. 2009. Determinants of assemblage structure in
Neotropical dry forest lizards. Austral Ecology, 34, 97-115.
Wiens, J.J. & Graham, C.H. 2005. Niche conservatism: Integrating Evolution, Ecology
and Conservation Biology. Annual Review of Ecology, Evolution and Systematics, 36,
519-539.
Zar J.H. 2010. Bioestatistical Analysis. 5a ed. New Jersey: Prentice-Hall.
Zimmerman, B.L. & Simberloff, D. 1996. An historical interpretation of habitat use by
frogs in a Central Amazonian Forest. Journal of Biogeography, 23, 27-46.
Zimmerman, B.L. 1994. Audio Strip Transects. In: Measuring and monitoring
biological diversity: standard methods for amphibians (Org. por W.R. Heyer., M.A.
Donnelly., R.W. McDiarmid., L.A.C. Hayek. & M.S. Foster), pp. 92-97. Washington:
Smithsonian Institution Press.
Zina, J., Ennser, J., Pinheiro, S.C.P., Haddad, C.F.B. & Toledo, L.F. 2007.
Taxocenose de anuros de uma mata semidecídua do interior do Estado de São Paulo e
comparações com outras taxocenoses do Estado, sudeste do Brasil. Biota Neotropica,
7 (2): 49-58.

86
CAPÍTULO 3
ARTIGO A SER SUBMETIDO AO PERIÓDICO HERPETOLOGICAL JOURNAL (QUALIS B1)
ECOLOGIA E COMPORTAMENTO REPRODUTIVO DE ESPÉCIES DE ANUROS
EM SAVANA AMAZÔNICA
Carlos Eduardo Costa-Campos1,2*; Eliza Maria Xavier Freire3
1 Universidade Federal do Amapá, Departamento de Ciências Biológicas e da Saúde,
Laboratório de Herpetologia, Rod. Juscelino Kubitschek, km 02, Jardim Marco Zero,
CEP 68.902-280, Macapá, AP, Brasil. 2 Universidade Federal do Rio Grande do Norte, Programa de Pós-Graduação em
Psicobiologia, Lagoa Nova, CEP 59.072-970, Natal, RN, Brasil.
3 Universidade Federal do Rio Grande do Norte, Departamento de Botânica, Ecologia e
Zoologia, Laboratório de Herpetologia, Lagoa Nova, CEP 59072-970, Natal, RN, Brasil.
* Corresponding author. E-mail: [email protected]
1. RESUMO
As taxocenoses de anuros são compostas por um grande número de espécies que têm uma
grande variedade de padrões, modos, estratégias e comportamentos reprodutivos. A
grande variedade de comportamentos reprodutivos exibidos pelos anuros podem envolver
complexas comunicações vocais, visuais, químicas e táteis. A existência de
comportamentos e estratégias alternativas de reprodução podem ter evoluído para
maximizar o sucesso reprodutivo, evitar a predação e a competição ou podem estar
relacionadas às condições ambientais específicas, indicando a importância de processos
históricos na composição das taxocenoses. Nesse contexto, este estudo teve como
objetivos estudar os padrões temporais, modos, estratégias e comportamento reprodutivo
de anuros. Das 21 espécies de anuros registradas na Savana amapaense, 11 foram
classificadas como de padrão de reprodução prolongado, com reprodução no mínimo de
três meses consecutivos, o que representa 52,4% das espécies registradas. Esse padrão
reprodutivo está intrinsecamente ligado ao período chuvoso e ao modo reprodutivo da
maioria das espécies, que apresentaram desova em corpos d’água lênticos. Foram
registrados seis modos reprodutivos, sendo o modo 1 o mais representativo, presente em
15 espécies (71,4%). O cuidado parental foi registrado nas espécies Leptodactylus
macrosternum e L. podicipinus, que apresentaram modos reprodutivos caracterizados
pelas desovas em ninhos de espuma. Quanto às estratégias reprodutivas comportamentais,
a estratégia do macho vocalizador foi observada em todas as 21 espécies de anuros; a
estratégia do macho satélite foi registrada apenas para as espécies Dendropsophus
walfordi, Hypsiboas multifasciatus, Scinax nebulosus e S. fuscomarginatus; a procura
ativa por fêmeas foi observada para as espécies Phyllomedusa hypochondrialis e L. fuscus
e, a estratégia de macho deslocador foi registrada apenas para as espécies Rhinella major
e R. margaritifera. Dos comportamentos reprodutivos registrados, a exibição da região
gular e do saco vocal está relacionada ao comportamento de corte e territorialidade
exibida pelos machos. Além dos comportamentos de corte, sinais visuais associados a
estratégias de corte foram registrados para as espécies em agregações reprodutivas.
PALAVRAS-CHAVE: Comportamento reprodutivo, estratégias reprodutivas, Anuros.

87
2. ABSTRACT
Assemblages of anurans are composed of a large number of species with a wide variety
of reproductive patterns, modes, strategies, and behaviors. The broad array of
reproductive behaviors by anurans may involve complex vocal, visual, chemical and
tactile communication. The existence of alternative reproductive behaviors and strategies
might have evolved to maximize reproductive success, avoid predation and competition,
or might be associated with specific environmental conditions, indicating the importance
of historical processes in assemblage composition. In this context, this study was aimed
at examining the temporal patterns, modes, and strategies, and reproductive behavior of
anurans. Of the 21 anuran species recorded in the Amapá savanna, 11 were classified as
prolonged reproductive pattern, with a breeding season of at least three consecutive
months, representing 52.4% of the recorded species. This reproductive pattern is closely
associated with the rainy season and the reproductive mode of most species, whose egg
clutches were found in lentic water bodies. Six reproductive modes were observed, with
mode 1 as the most representative, observed in 15 species (71.4%). Parental care was
recorded in the species Leptodactylus macrosternum and L. podicipinus, which exhibited
reproductive modes characterized by egg clutches in foam nests. Regarding male mating
strategies, male calling was observed in all 21 anuran species; satellite behavior was
recorded only for the species Dendropsophus walfordi, Hypsiboas multifasciatus, Scinax
nebulosus, and S. fuscomarginatus; active search for females was observed in the species
Phyllomedusa hypochondrialis and L. fuscus; displacement behavior was recorded only
for the species Rhinella major and R. margaritifera. Of the reproductive behaviors
observed, throat and vocal sac display is associated with courtship behavior and
territoriality by males. In addition to courtship behavior, visual signals associated with
strategies of courtship were recorded for the species observed in breeding aggregations.
KEY-WORDS: Breeding behavior, Reproductive strategies, Anurans.
3. INTRODUÇÃO
As taxocenoses de anuros neotropicais são compostas por um grande número de
espécies que têm uma grande variedade de modos de reprodução (Hödl 1990, Haddad &
Prado 2005, Pombal Jr. & Haddad 2007, Toledo et al. 2012). A existência de
comportamentos e estratégias alternativas de reprodução podem ter evoluído para
maximizar o sucesso reprodutivo ou essa evolução de estratégias reprodutivas podem
estar relacionadas às condições ambientais específicas (Taborsky et al. 2008, Taborsky &
Brockmann 2010, Toledo et al. 2012), indicando a importância de processos históricos na
composição das taxocenoses.
O comportamento reprodutivo dos anuros pode ser categorizado de acordo com
dois padrões temporais de reprodução: explosivo e prolongado, conforme descrição de
Wells (1977a, 2007). Espécies com padrão explosivo apresentam reprodução em curto
espaço de tempo, com grande quantidade de machos e fêmeas sexualmente maduros no
local de desova; é comum que os machos exibam procura ativa por fêmeas e não

88
defendam território. No padrão prolongado, as estações reprodutivas ocorrem em vários
meses, na qual a chegada de fêmeas é assincrônica; nesse padrão, o sucesso reprodutivo
do macho depende da sua capacidade de atrair fêmeas ao sítio de vocalização e de manter
distância segura de outros machos.
Apesar da utilidade de classificação proposta por Wells (1977a), ela não se adapta
a todos os anfíbios anuros, podendo diferir localmente de acordo com fatores ambientais.
Crump (1974) encontrou quatro padrões temporais de reprodução em espécies de anfíbios
anuros: (1) contínuo, reprodução em todas as noites, exceto naquelas secas e com luar;
(2) oportunista, regular após chuvas fortes; (3) esporádico úmido, reprodução esporádica
após chuvas fortes; (4) esporádico seco, esporádica durante infrequentes períodos de seca.
Essa classificação reflete, em parte, a complexidade do comportamento reprodutivo de
espécies de anfíbios anuros neotropicais.
O modo reprodutivo dos anuros é uma combinação de caracteres que incluem o
local onde os ovos são depositados, as características dos ovos e da desova, o ritmo e a
duração dos estágios de desenvolvimento, o estágio e o tamanho dos girinos e, se houver,
o tipo de cuidado parental (Haddad & Prado 2005). A diversidade de modos reprodutivos
é uma das maiores observadas para os vertebrados e isso se deve à existência de espécies
que apresentam modos de reprodução mais generalizados, onde os ovos são depositados
diretamente na água, e de espécies que apresentam modos de reprodução mais
especializados, inclusive em ambientes terrestres (Pombal Jr. & Haddad 2007).
Diante do exposto e considerando a ausência de estudos sobre os comportamentos
reprodutivos de anuros em área de Savana localizada na Amazônia Oriental, este estudo
teve como objetivos, (1) verificar os padrões temporais do comportamento reprodutivo,
(2) identificar os modos reprodutivos e o cuidado parental, (3) descrever as estratégias
reprodutivas comportamentais, (4) caracterizar os sítios de vocalização e (5) analisar os
comportamentos reprodutivos de espécies nas fitofisionomias da Savana amapaense.
4. MATERIAL E MÉTODOS
4.1. Área de estudo
A Savana amapaense se estende na direção norte do Estado do Amapá até o
município de Calçoene, ocupando uma estreita faixa longitudinal de 9.861,89 Km2, o que
corresponde a 6,9% da superfície territorial do Estado do Amapá (Rabelo et al. 2008).
Este estudo foi realizado em uma área de 1.381 ha e perímetro de 15.080 m (Melém-
Júnior et al. 2003), especificamente no Campo Experimental da Empresa Brasileira de

89
Pesquisa Agropecuária – EMBRAPA (00°023’5” N, 51°02’2” W), uma área de vegetação
nativa de Savana no Estado do Amapá.
Para tanto, foram amostradas quatro fitofisionomias distintas de vegetação nativa
da Savana amapaense (cf. Miranda et al. 2003), a saber: savana gramíneo lenhosa,
caracterizada por extensos campos de gramíneas e estrato herbáceo abundante, com
estrato lenhoso disperso e composto por árvores anãs esparsas (< 1,0 m); savana gramíneo
lenhosa-arbustiva, com estrato herbáceo coberto por gramíneas em grande densidade e
estrato lenhoso com presença irregular e pequenas árvores esparsas (< 3,8 m); savana
parque, com estrato herbáceo menos desenvolvido e cobertura arbóreo-arbustiva
relativamente baixa (< 4,3 m) e densa formando um dossel descontínuo e, savana arbórea,
com estrato herbáceo ausente e árvores com maior densidade, formando um dossel
contínuo (> 2,0 m).
O clima predominante na Savana amapaense é Am, de acordo com a classificação
Köppen, caracterizado como tropical chuvoso com nítida estação seca (Alvares et al.
2013). O clima ao longo do ano pode ser tipicamente reconhecido em apenas duas
estações: uma estação seca bem evidente e uma estação chuvosa, com precipitação anual
variando de 4.6 mm (estação seca) a 4.827 mm (estação chuvosa). A temperatura média
mensal varia de 24.4oC a 28.4oC, com umidade relativa média de 84.7%.
O relevo é plano ou suavemente ondulado, ocorrendo em duas formas principais:
os Latossolos e as Areias quartzosas e, o domínio da Savana amapaense ocorre sobre dois
tipos de unidades de relevo: planaltos cristalinos ou sedimentares e depressões
interplanálticas. Nas regiões dos planaltos predominam as fitofisionomias abertas: savana
gramíneo lenhosa e savana gramíneo lenhosa-arbustiva; enquanto que nas depressões
interplanáticas predominam fitofisionomias florestais: savana parque e a savana arbórea
(Aguiar et al. 2004, Barbosa et al. 2005).
4.2. Procedimentos metodológicos
Para a amostragem dos anfíbios anuros da Savana amapaense foram realizadas 12
excursões em cada fitofisionomia durante o período de janeiro a dezembro de 2013. Para
tanto, foram estabelecidas 20 unidades amostrais em cada fitofisionomia (Savana
gramíneo lenhosa, Savana gramíneo lenhosa-arbustiva, Savana parque e Savana arbórea),
dispostas de acordo com a disponibilidade de corpos d’água. Cada unidade amostral foi
representada por uma parcela de 100 x 50 metros (0,5ha), distantes pelo menos 500
metros entre si, conforme sugerido por Bitar et al. (2012). Nessas unidades amostrais

90
foram realizados os métodos de busca ativa visual e acústica (Crump & Scott 1994, Heyer
et al. 1994, Zimmerman 1994).
4.2.1. Padrões temporais do comportamento reprodutivo
O padrão reprodutivo de cada espécie foi determinado com base em machos em
atividade de vocalização, casais em amplexo e girinos, conforme descrição proposta por
Crump (1974) e Wells (1977a). Os padrões temporais de comportamento reprodutivo
exibidos pelas espécies foram classificados considerando as seguintes categorias: (1)
contínuo, onde se tem fêmeas com ovos, desovas e girinos o ano todo; (2) prolongado,
quando se estende por várias semanas; e (3) explosivo, quando se estende por poucos
dias, e em geral possui características comportamentais associadas com um grande
número de casais simultaneamente em amplexo, machos em procura ativa por fêmeas e
vocalizações sobrepostas (Pombal Jr & Haddad 2005, Prado et al. 2005).
4.2.2. Modos reprodutivos
Para classificação dos modos reprodutivos foi utilizado o método proposto por
Haddad & Prado (2005) e Pombal Jr. & Haddad (2007). A determinação dos modos
reprodutivos foi realizada a partir da observação do sítio de oviposição, características
dos ovos e da desova, característica e local de desenvolvimento dos girinos e cuidado
parental, se presente (cf. Haddad & Prado 2005, Pombal Jr & Haddad 2005, Pombal Jr &
Haddad 2007, Toledo et al. 2012).
4.2.3. Estratégias reprodutivas comportamentais
Quanto às estratégias reprodutivas comportamentais exibidas pelos machos,
adotamos o método proposto por Pombal Jr & Haddad (2007). Estas estratégias foram
divididas em dois tipos básicos: (1) estratégia do macho vocalizador, na qual os machos
através da vocalização, atrai fêmeas reprodutivas e, (2) estratégias reprodutivas
alternativas (cf. Howard 1978), que são divididas em três categorias: (2a) estratégia do
macho satélite, onde machos menores se associam a machos dominantes (vocalmente
ativos) para interceptar fêmeas que se aproximem do macho vocalizante ou simplesmente
para esperar vacância dos sítios reprodutivos; (2b) estratégia de procura ativa por fêmeas,
onde o macho percorre o ambiente a procura de fêmeas reprodutivas que são interceptadas
para o acasalamento e (2c) estratégia do macho deslocador, onde o macho tenta deslocar
machos em amplexo para acasalar com as fêmeas.

91
4.2.4. Sítios de vocalização
O sítio de vocalização de cada macho foi caracterizado pelos seguintes
descritores: 1) posição dos anuros em relação ao sítio de vocalização; 2) altura em relação
à superfície da água ou do solo, para as espécies cujos machos vocalizam empoleirados;
3) distância do sítio de vocalização até a margem do corpo d’água.
4.2.5. Comportamentos reprodutivos
Para estudo dos comportamentos reprodutivos foram analisadas quatro espécies
de anuros com maior abundância na taxocenose estudada. As observações em campo
foram feitas mensalmente com duração de quatro dias consecutivos, totalizando 107 horas
para cada espécie. Dentre as quatro espécies registradas, 36 indivíduos de Leptodactylus
fuscus (Savana gramíneo-lenhosa), 40 de Phyllomedusa hypochondrialis (Savana
gramíneo-lenhosa-arbustiva), 38 de Hypsiboas punctatus (Savana parque) e 45 de
Rhinella margaritifera (Savana arbórea).
Para as observações dos comportamentos reprodutivos utilizamos os métodos de
amostragem ad libitum e animal focal (Lehner 2003). Os comportamentos reprodutivos
das espécies foram identificados com base na ocorrência de machos em atividade de
vocalização, comportamentos de corte e sinais visuais (Wells 2007). As descrições dos
comportamentos foram baseadas em Hodl & Amézquita (2001) e Hartmann et al. (2005).
Os períodos de observação foram iniciados as 17:30 h e terminaram somente
depois de cessada a atividade de vocalização, aproximadamente às 24:00 h. Para cada
indivíduo foi destinado um período de amostragem de 15 minutos. Os registros dos
comportamentos reprodutivos foram transcritos em caderno de campo e os
comportamentos filmados utilizando Filmadora Digital Full HD. As filmagens tiveram
duração de cinco minutos, totalizando 284 horas de registro.
Os indivíduos foram identificados individualmente pela presença de marcas
naturais e pelo padrão de coloração dorsal (presença ou ausência de linhas em forma de
ampulheta, marcas circulares e linhas no dorso e entre os olhos, marcas triangulares entre
as narinas e barras transversais na região da coxa) através de fotografias (Doody 1995,
Del Lama et al. 2011).

92
4.3. Análise dos dados
A sobreposição de nicho foi calculada pela equação descrita por Pianka (1974). O
resultado desta equação varia entre zero (0 = nenhuma sobreposição e um (1 =
sobreposição total). Os valores de sobreposição foram considerados altos (>0,6) ou baixos
(<0,4). Esses índices foram obtidos e aleatorizados 1000 vezes com o auxílio do programa
Ecosim 7.71 (Gotelli & Entsminger 2005).
∅𝒊𝒋 =∑ 𝒑𝒊𝒋 𝒑𝒊𝒌
𝒏𝒊=𝟏
√∑ 𝒑𝒊𝒋𝟐 ∑ 𝒑𝒊𝒌𝟐𝒏𝒊=𝟏
𝒏𝒊=𝟏
Para avaliar os valores de sobreposição para o sítio de vocalização entre os pares
de espécies foram construídos modelos nulos a partir da aleatorização dos dados da
comunidade (Gotelli & Colwell 2001, Gotelli & Entsminger 2005). A presença de
padrões não aleatórios na sobreposição de nicho foi obtida com a utilização do módulo
de sobreposição de nicho “Niche Overlap Module” do programa EcoSim 7.71 (Gotelli &
Entsminger 2005), utilizando as opções: Índice de sobreposição de nicho de Pianka e
Algoritmo de randomização para substituir as categorias de nicho da matriz original por
números uniformes e aleatórios entre 0 e 1 (Crawford et al. 2009).
A amplitude de nicho para o sítio de vocalização foi determinada pelo índice de
Levins padronizado (Krebs 2014), para as dimensões distância da margem (para todas as
espécies registradas) e altura de empoleiramento (para os hilídeos). As espécies foram
consideradas generalista para os recursos que apresentaram valores B ≥ 0,50.
𝑩 = 𝟏 ∑ 𝒑𝒊𝟐
𝒏
𝒊=𝟏
⁄
Os valores obtidos de B foram utilizados para calcular o índice padronizado de
Levins (Bp), que produz valores entre zero e um (0 = largura mínima, 1 = largura
máxima). Por produzir valores entre um intervalo conhecido (0 - 1), este índice foi
utilizado para comparar as larguras de nicho das espécies de anuros. Para calcular os
índices de largura de nicho foi utilizado o software Ecological Metodology, Version 5.2
(Krebs 2014).
Onde B = índice de Levins; i = categoria de habitat, microhabitat ou dieta; n = número
de categorias de habitat, micro-habitat ou dieta; pi = proporção de indivíduos
associados à categoria i de habitat, microhabitat ou dieta.
Onde Øij = índice de similaridade de nicho; p = a proporção da
categoria de habitat, microhabitat ou dieta; j e k = representam o par de
espécies comparadas.

93
𝑩𝒑 =(𝜷−𝟏)
(𝒏−𝟏)
Para testar a relação entre as variáveis ambientais (pluviometria, temperatura e
umidade relativa) e as espécies de anuros em reprodução foi utilizado o método de
regressão linear simples (Zar 2010).
Para verificar diferenças entre os comportamentos reprodutivos exibidos pelas
espécies de anuros foi realizada uma Análise de agrupamento (Cluster analysis)
utilizando o índice de similaridade de Bray-Curtis.
Todas as análises foram realizadas através dos programas “BioEstat 5.0 (Ayres et
al. 2007) e PAST 2.09 (Hammer et al. 2001). Os valores foram considerados
significativos para p < 0,05.
5. RESULTADOS E DISCUSSÃO
5.1. Padrões temporais do comportamento reprodutivo
Do total de espécies de anuros registradas na Savana amapaense, 11 foram
classificadas como de padrão de reprodução prolongado, com reprodução no mínimo de
três meses consecutivos, o que representa 52,4% das espécies registradas. Sete espécies
(33,3%) apresentaram padrão de reprodução explosiva e três espécies (14,3%)
apresentaram padrão de reprodução contínuo (Tabela 1).
Estes resultados estão de acordo com Wells (1977b, 2007), no qual o período
reprodutivo prolongado é o mais frequente para a maioria das espécies da região
Neotropical. Nas espécies que apresentam esse padrão reprodutivo, que ocorre por
semanas ou meses, algumas características da vocalização e da qualidade dos sítios
reprodutivos e de oviposição tem papel fundamental na atração das fêmeas e escolha do
parceiro pelas fêmeas (Wells 1977a). Entretanto, uma desvantagem da reprodução
prolongada é que os ovos, girinos e imagos ficam mais tempo expostos a predação.
Esse padrão reprodutivo está intrinsecamente ligado ao período chuvoso e ao
modo reprodutivo da maioria das espécies (desova em corpos d’água lênticos). No caso
de P. hypochondrialis registramos atividade de vocalização e casais em amplexo no solo
em pequenas poças isoladas com vegetação de gramíneas rasteiras. De acordo com Wogel
et al. (2005) e García et al. (2013), esse comportamento é um dos últimos recursos
utilizados, já que esse gênero tem uma desova especializada onde os ovos são liberados
em folhas dobradas para retenção de umidade próximas a poças temporárias.
Onde Bp = índice padronizado de Levins; B = índice de Levins; n = número de
categorias utilizadas.

94
As espécies R. major, R. margaritifera, R. marina, S. x-signatus, T. typhonius, L.
macrosternum e E. helianneae apresentaram reprodução explosiva, utilizando poças
temporárias formadas após curtos períodos de chuva. Esses dados estão de acordo com
estudos realizados com anuros que apresentam reprodução explosiva (Toledo et al. 2003,
Prado & Pombal Jr. 2005), que são caracterizados pela presença frequente de machos em
poças temporárias após a chuva e pela presença de coros reprodutivos que definem os
locais de desova e atração das fêmeas.
O período chuvoso coincidiu também com o encontro de casais em amplexo,
girinos e imagos dos anuros registrados. A chuva parece ser o principal fator regulador
das atividades reprodutivas dos anuros, não apenas pela vulnerabilidade desses animais à
dessecação, mas também porque a chuva determina a viabilidade e duração dos sítios
reprodutivos (Gottsberger & Gruber 2004).
Tabela 1 – Espécies de anuros registrados na Savana amapaense durante o período de Janeiro a
Dezembro de 2013 com seus respectivos padrões reprodutivos (E = explosivo, P = prolongado, C
= contínuo), fitofisionomias (SGL = savana gramíneo-lenhosa, SGLA = savana gramíneo-lenhosa
arbustiva, SP = savana parque, SA = savana arbórea) e períodos reprodutivos (barras cinzas).
Espécies Padrão Fitofisionomia Período Reprodutivo
Rhinella major E SGLA, SP, SA
Rhinella margaritifera E SA
Rhinella marina E SGL, SGLA
Dendropsophus walfordi P SGL, SGLA
Hypsiboas multifasciatus P SGL, SGLA, SP
Hypsiboas punctatus P SGLA, SP
Hypsiboas raniceps P SGL, SGLA, SP
Osteocephalus taurinus P SGL, SGLA, SP
Scinax fuscomarginatus P SGL, SGLA
Scinax nebulosus C SGL, SGLA
Scinax ruber P SGL, SGLA
Scinax x-signatus E SGL, SGLA
Trachycephalus typhonius E SGL, SGLA
Pseudopaludicola boliviana P SGL, SGLA
Adenomera hylaedactyla P SGL, SGLA, SA
Phyllomedusa hypochondrialis P SGLA
Leptodactylus fuscus C SGL, SGLA
Leptodactylus macrosternum E SP, SA
Leptodactylus pentadactylus P SGLA, SP
Leptodactylus podicipinus C SGLA
Elachistocleis helianneae E SGLA, SP
Jan Fev Mar Abr Mai Jun Jul Ago Set Out Nov Dez
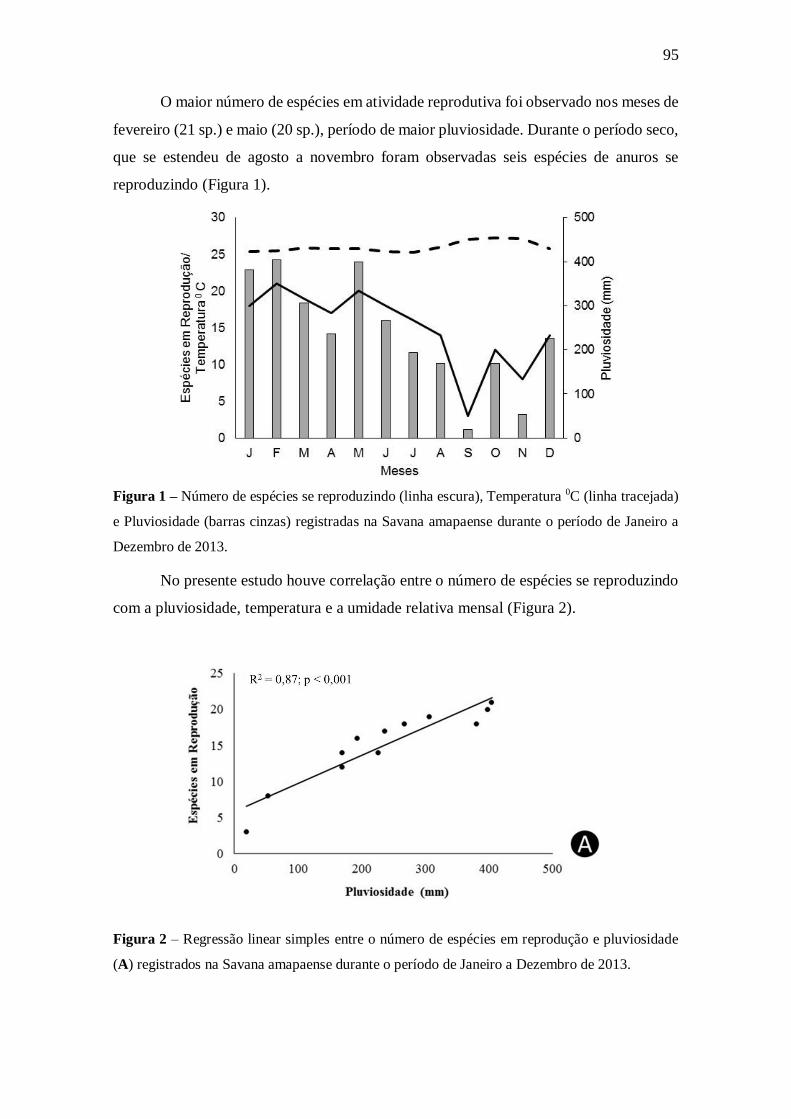
95
O maior número de espécies em atividade reprodutiva foi observado nos meses de
fevereiro (21 sp.) e maio (20 sp.), período de maior pluviosidade. Durante o período seco,
que se estendeu de agosto a novembro foram observadas seis espécies de anuros se
reproduzindo (Figura 1).
Figura 1 – Número de espécies se reproduzindo (linha escura), Temperatura 0C (linha tracejada)
e Pluviosidade (barras cinzas) registradas na Savana amapaense durante o período de Janeiro a
Dezembro de 2013.
No presente estudo houve correlação entre o número de espécies se reproduzindo
com a pluviosidade, temperatura e a umidade relativa mensal (Figura 2).
Figura 2 – Regressão linear simples entre o número de espécies em reprodução e pluviosidade
(A) registrados na Savana amapaense durante o período de Janeiro a Dezembro de 2013.

96
Figura 2 (Continuação) – Regressão linear simples entre o número de espécies em reprodução
e Temperatura 0 C (B) e umidade relativa (C) registrados na Savana amapaense durante o período
de Janeiro a Dezembro de 2013.
5.2. Modos reprodutivos e Cuidado parental
Foram registrados seis modos reprodutivos nas espécies de anuros presentes na
Savana amapaense. O modo reprodutivo 1, caracterizado por ovos e girinos exotróficos
em corpos d’água lênticos, foi o mais representativo, presente em 15 espécies (71,4%)
das famílias Bufonidae, Hylidae, Leptodactylidae e Microhylidae, seguido pelo modo
reprodutivo 30, presente em duas espécies (9,5%) da família Leptodactylidae (Tabela 2,
Figura 3). Esse modo é caracterizado por ninhos de espuma com ovos e estágios larvais
iniciais em ninhos subterrâneos construídos, que após a inundação, girinos exotróficos
em corpos d’água lênticos.

97
Tabela 2 – Espécies de anuros registradas na Savana amapaense durante o período de Janeiro a
Dezembro de 2013 com descrição dos modos reprodutivos, desova e girinos.
Figura 3 – Alguns modos reprodutivos das espécies de anuros registrados na Savana amapaense.
A) Desova aquática em cordões gelatinosos de R. margaritifera; B) Ovos aquáticos em corpos
d’água lênticos de D. walfordi; C) Desova em ninhos de espuma na superfície da água de L.
macrosternum; D) Desova em ninhos de espuma flutuante em água acumulada em panelas
construídas de L. podicipinus; E) Ovos arborícolas de P. hypochondrialis (Modo 24); F) Desova
em ninhos de espuma de L. pentadactylus.
Espécies Modos Desova Girinos
Rhinella major 1 Corpos d’água lênticos Exotróficos
Rhinella margaritifera 1 Corpos d’água lênticos Exotróficos
Rhinella marina 1 Corpos d’água lênticos Exotróficos
Dendropsophus walfordi 1 Corpos d’água lênticos Exotróficos
Hypsiboas multifasciatus 1 Corpos d’água lênticos Exotróficos
Hypsiboas punctatus 1 Corpos d’água lênticos Exotróficos
Hypsiboas raniceps 1 Corpos d’água lênticos Exotróficos
Osteocephalus taurinus 1 Corpos d’água lênticos Exotróficos
Scinax fuscomarginatus 1 Corpos d’água lênticos Exotróficos
Scinax nebulosus 1 Corpos d’água lênticos Exotróficos
Scinax ruber 1 Corpos d’água lênticos Exotróficos
Scinax x-signatus 1 Corpos d’água lênticos Exotróficos
Trachycephalus typhonius 1 Corpos d’água lênticos Exotróficos
Pseudopaludicola boliviana 1 Corpos d’água lênticos Exotróficos
Adenomera hylaedactyla 32 Ninhos de espuma Endotróficos
Phyllomedusa hypochondrialis 24 Arborícolas Exotróficos
Leptodactylus fuscus 30 Ninhos de espuma Exotróficos
Leptodactylus macrosternum 11 Ninhos de espuma Exotróficos
Leptodactylus pentadactylus 30 Ninhos de espuma Exotróficos
Leptodactylus podicipinus 13 Ninhos de espuma Exotróficos
Elachistocleis helianneae 1 Corpos d’água lênticos Exotróficos

98
Os modos reprodutivos observados na Savana amapaense representam 15,4% dos
39 modos reprodutivos conhecidos para o Brasil (Pombal Jr. & Haddad 2007). A
predominância de desovas aquáticas e em ninhos de espuma estão de acordo com Hödl
(1990), em estudo sobre a diversidade de modos reprodutivos de espécies de anuros
amazônicos de formações abertas.
A menor diversificação de modos reprodutivos e a predominância de modos
generalizados não terrestres (Modo 1) bem como de modos que apresentam resistência a
dessecação (Modo 11, 13, 30 e 32) é esperada em áreas de Savana, pois a diversidade de
modos reprodutivos é mais um reflexo das características ambientais e físicas do habitat
do que das relações filogenéticas (Vasconcelos & Rossa-Feres 2005, Kopp et al. 2010,
Vasconcelos et al. 2010, Valdujo et al. 2013). Nesse caso, essas características podem
estar relacionadas a fatores históricos ligados a sazonalidade climática da Savana
amapaense que talvez tenham historicamente limitado a diversificação dos modos
reprodutivos dos anuros na área de estudo.
O cuidado parental foi registrado em duas espécies da família Leptodactylidae
registradas na área de estudo: L. macrosternum e L. podicipinus, caracterizadas pelas
desovas em ninhos de espuma. Leptodactylus podicipinus exibiu cuidado maternal na
proteção dos ovos fertilizados e girinos nos estágios de desenvolvimento inicial (23-30),
intermediário (31-40) e final (41-46), segundo caracteres morfológicos contidos em
Gosner (1960). Leptodactylus macrosternum apresentou cuidado maternal com a desova
e girinos em estágio inicial de desenvolvimento (23-30).
Em anuros, a forma mais comum de cuidado parental está associada a proteção da
desova, o cuidado com os girinos, que se estende do início do desenvolvimento até a
metamorfose, e o transporte de ovos e girinos (Crump 1995, Wells 2007, Delia et al. 2010,
Castro et al. 2013, Vargas-Salinas et al. 2014). O comportamento parental também pode
ser associado com a defesa da prole contra predadores, e pode ser executado pelos machos
ou pelas fêmeas e, em alguns casos, por ambos os sexos (Santos & Amorim 2006, De Sá
et al. 2007).
No presente estudo, a presença de fêmeas no cuidado parental com a desova e
girinos em diferentes estágios de desenvolvimento, tal como observado em ambas as
espécies, pode ser uma adaptação das circunstâncias em que ocorre a desova nos sítios
reprodutivos mantidos pelos machos (Martins 2001, Rodrigues et al. 2011). O registro de
machos de L. podicipinus construindo bacias pode representar uma especialização

99
evolutiva paternal, podendo ser caracterizado como um investimento parental (Martins,
2001, Prado et al. 2002).
5.3. Estratégias reprodutivas comportamentais
Na comunidade de anuros da Savana amapaense foram observadas três diferentes
estratégias comportamentais utilizadas pelos machos durante a atividade reprodutiva. A
estratégia do macho vocalizador foi observada em todas as 21 espécies presentes na área
de estudo. A estratégia do macho satélite foi registrada apenas para as espécies da família
Hylidae: D. walfordi, H. multifasciatus, S. fuscomarginatus e S. nebulosus. A procura
ativa por fêmeas foi observada para as espécies P. hypochondrialis e L. macrosternum e,
a estratégia de macho deslocador foi registrada apenas para duas espécies da família
Bufonidae: R. major e R. margaritifera.
A estratégia do macho satélite foi observada em 12 indivíduos de D. walfordi,
com registro de dois machos satélites próximos a um único macho vocalizador. Em oito
observações, foram registrados machos satélites de H. multifasciatus. Em S.
fuscomarginatus, foi observado apenas um macho satélite próximo ao macho vocalizador.
Em cinco registros foram observados machos satélites de S. nebulosus. Das espécies
descritas, apenas dois machos satélites de S. nebulosus interceptaram fêmeas reprodutivas
e casais já amplectados. Os machos apresentaram comportamento satélite como estratégia
alternativa para aquisição de fêmeas ovadas que se aproximem de machos vocalizantes
(Wells 1977b, Pombal Jr. & Haddad 2007) e, em todas as ocasiões, os machos satélites
começaram a vocalizar quando o macho vocalizador deixava o sítio reprodutivo ou
simplesmente deixava de vocalizar.
A estratégia reprodutiva de procura ativa por fêmeas foi registrada nas espécies P.
hypochondrialis (n = 14) e L. macrosternum (n = 8), que foram observadas se deslocando
lentamente pela vegetação procurando fêmeas aptas a reprodução. Nesta estratégia
observamos os machos de P. hypochondrialis (n = 6) e L. macrosternum (n = 2)
interceptando as fêmeas ovadas. Em três observações, os machos de L. macrosternum
foram registrados interceptando casais em amplexo. Apenas P. hypochondrialis foi
observada se deslocando pela vegetação enquanto estavam em atividade de vocalização,
comportamento registrado por Pombal Jr & Haddad (2005). Esta estratégia está
relacionada às altas densidades de indivíduos nas agregações reprodutivas (Wells 1977a),
sendo comum observar tentativas de amplexo entre machos, machos solitários ou
pareados (Arak 1983, Wells 2007).

100
A estratégia do macho deslocador, observada 23 vezes, foi registrada nas espécies
R. major (n = 17) e R. margaritifera (n = 6). Durante as estratégias do macho deslocador
foram utilizadas três estratégias: (1) em todos os casos o macho deslocador agarrou a
região abdominal do macho em amplexo; (2) em três observações o macho deslocador
empurrou o macho em amplexo com as pernas e palmas do pé para a lateral do corpo da
fêmea; (3) em duas observações o macho deslocador entrou em baixo da fêmea e virou o
casal em amplexo de cabeça para baixo. Nos machos em amplexo foram observados
chutes com as pernas e vibração do corpo em resposta as estratégias do macho deslocador.
Para evitar o deslocamento dos machos em amplexo, as fêmeas também exibiam
pequenos chutes e caminhavam lentamente.
5.4. Sítios de vocalização
O maior número de espécies em atividade de vocalização ocorreu durante o
período de janeiro a julho de 2013, coincidindo com os meses de maiores índices
pluviométricos (Figura 4). Algumas espécies como R. major, H. multifasciatus, H.
punctatus, P. hypochondrialis, S. nebulosus, S. ruber, S. x-signatus e L. fuscus foram
registradas vocalizando também nos meses mais secos. De acordo com Höld (1990), a
pluviosidade e a disponibilidade de corpos d’água são provavelmente os fatores
ambientais que mais influenciam a atividade de vocalização e reprodução dos anuros.
Os machos iniciaram as atividades de vocalização e estabeleceram território antes
da chegada das fêmeas e apresentaram fidelidade em relação a determinados
microhabitats como sítios de vocalização (Pombal Jr. 1997; Prado et al. 2005). Isto está
relacionado à competição por sítios de vocalização, pois a obtenção de “bons sítios” para
vocalizar permite maiores chances de sucesso reprodutivo (Crump 1974).
Casais em amplexo foram registrados nos meses com maior concentração de
chuvas, coincidindo com a atividade de vocalização das espécies de anuros da Savana
amapaense. Para as espécies R. major, R. margaritifera, D. walfordi, H. multifascitus, H.
punctatus, S. nebulosus, S. ruber e E. helianneae, a formação de casais em amplexo foi
também registrada durante o período seco, sendo comum a formação de agrupamentos de
até oito indivíduos, dependendo da densidade de machos ao longo das margens dos corpos
d’água. Todas as espécies de anuros registradas na Savana amapaense apresentaram
amplexo do tipo axilar, considerado o mais comum entre os anuros (Wells 2007).

101
Figura 4 – Atividade de vocalização e casais em amplexo de anuros registrados na Savana
amapaense durante o período de janeiro a dezembro de 2013. As barras escuras representam
machos em vocalização e as barras cinzas casais em amplexo.

102
Nos sítios de vocalização utilizados pelos anuros na Savana amapaense, 13
espécies (57,1%) vocalizaram empoleiradas e 12 (47,6%) sobre o solo. As espécies O.
taurinus, T. typhonius e A. hylaedactyla vocalizaram empoleiradas e sobre o solo,
ocupando os mesmos sítios de vocalização (Tabela 3). Entretanto, considerando
separadamente as espécies que vocalizaram empoleiradas (Hylidae) e as que vocalizaram
no solo (Bufonidae e Leptodactylidae) ou flutuando na superfície da água
(Leptodactylidae e Microhylidae) registramos dois padrões: partilha completa em todos
os corpos d’água para os hilídeos e sobreposição entre todas as espécies que vocalizaram
no solo ou flutuando na superfície da água.
De acordo com Toledo & Haddad (2005) o tipo do sítio de vocalização é
determinado principalmente por suas características morfológicas de cada espécie (e.g.
tamanho corpóreo, altura ocupada no estrato vertical e sítios para deposição de ovos).
Assim, a ocupação dos hilídeos verticalmente está relacionada à presença de discos
adesivos nas terminações digitais (Bertoluci & Rodrigues 2002). Complementarmente,
espécies de maior tamanho corpóreo, como os bufonídeos e leptodactilídeos, ocupam
preferencialmente microhabitats terrestres e corpos d’água durante o seu ciclo de vida
(Prado & Pombal Jr. 2005, Conte & Rossa-Feres 2007).
As larguras de nicho do sítio de vocalização variaram de 0,34 a 0,99. A altura em
relação ao poleiro variou de 0,55 a 0,98, com todas as espécies generalistas; a altura do
poleiro em relação a superfície da água variou de 0,36 a 0,98, com 80% das espécies
generalistas e, a largura de nicho do sítio de vocalização para distância do sítio de
vocalização até a margem mais próxima ao corpo d’água variou de 0,34 a 0,99, com 85%
das espécies generalistas e 15% especialistas (Tabela 4).
Os valores totais dos índices de sobreposição de nicho do sítio de vocalização para
distância do sítio de vocalização até a margem mais próxima ao corpo d’água ocorreram
entre os pares de espécies H. punctatus vs. H. raniceps, H. punctatus vs. S. ruber, H.
raniceps vs. S. fuscomarginatus e S. fuscomarginatus vs. S. nebulosus. A ausência de
sobreposição ocorreu nos pares de espécies R. major vs. L. fuscus, H. raniceps vs. A.
hylaedactyla e L. fuscus vs. E. helianneae (Tabela 5).

103
Tabela 3 – Sítios de vocalização utilizados pelos anuros na Savana amapaense, durante o período de Janeiro a Dezembro de 2013. Posição dos anuros em
relação ao sítio de vocalização: P = paralelo, VC = vocalizando para cima, VP = vocalizando para baixo, D = diagonal. Altura do poleiro. Altura do poleiro em
relação a superfície do corpo d’água. Distância do sítio de vocalização até a margem mais próxima ao corpo d’água.
Espécies
P VC VP D 1-30 31-60 61-90 91-120 > 120 1-30 31-60 61-90 91-120 > 120 0-30 31-60 61-90 91-120 > 120
Rhinella major
Rhinella margaritifera
Rhinella marina
Dendropsophus walfordi
Hypsiboas multifasciatus
Hypsiboas punctatus
Hypsiboas raniceps
Osteocephalus taurinus
Scinax fuscomarginatus
Scinax nebulosus
Scinax ruber
Scinax x-signatus
Trachycephalus typhonius
Phyllomedusa hypochondrialis
Pseudopaludicola boliviana
Adenomera hylaedactyla
Leptodactylus fuscus
Leptodactylus macrosternum
Leptodactylus pentadactylus
Leptodactylus podicipinus
Elachistocleis helianneae
Posição Altura do poleiro (cm) Distância da margemSuperfície do corpo d'água

104
Tabela 4 – Largura de nicho para as variáveis do sítio de vocalização das espécies registradas na
Savana amapaense, durante o período de Janeiro a Dezembro de 2013. Altura do poleiro. Altura
do poleiro em relação a superfície do corpo d’água. Distância do sítio de vocalização até a margem
mais próxima ao corpo d’água
Espécies Altura do poleiro Superfície do corpo d'água Distância da margem
Rhinella major - - - - - - 0,97
Rhinella margaritifera - - - - - - 0,97
Rhinella marina - - - - - - 0,60
Dendropsophus walfordi 0,87 0,62 0,71
Hypsiboas multifasciatus 0,69 0,39 0,64
Hypsiboas punctatus 0,98 0,56 0,99
Hypsiboas raniceps 0,78 0,98 0,98
Osteocephalus taurinus 0,77 0,36 0,49
Scinax fuscomarginatus 0,55 0,74 0,89
Scinax nebulosus 0,71 - - - 0,75
Scinax ruber 0,98 0,63 0,92
Scinax x-signatus 0,98 0,81 0,64
Trachycephalus typhonius 0,93 0,70 0,94
Phyllomedusa hypochondrialis 0,76 0,90 0,67
Pseudopaludicola boliviana - - - - - - 0,89
Adenomera hylaedactyla - - - - - - 0,34
Leptodactylus fuscus - - - - - - 0,85
Leptodactylus macrosternum - - - - - - 0,47
Leptodactylus pentadactylus - - - - - - 0,80
Leptodactylus podicipinus - - - - - - 0,86
Elachistocleis helianneae - - - - - - - - -

105
Tabela 5 – Sobreposição de nicho para distância do sítio de vocalização até a margem mais próxima ao corpo d’água registrado pelas espécies de anuros da
Savana amapaense, durante o período de Janeiro a Dezembro de 2013. Rmj = R. major, Rmarg = R. margaritifera, Rm = R. marina, Dw = D. walfordi, Hm =
H. multifasciatus, Hp = H. punctatus, Hr = H. raniceps, Ot = O. taurinus, Sf = S. fuscomarginatus, Sn = S. nebulosus Sr = S. ruber, Sx-s = S. x-signatus, Tt =
T. typhonius, Ph = P. hypochondrialis, Pb = P. boliviana, A. hylaedactyla, Lf = L. fuscus, Lm = L. macrosternum, Lpenta = L. pentadactylus, Lpod = L.
podicipinus, Eh = E.helianneae. Os números em negrito indicam valores de sobreposição de nicho ≥ 95 %
Rmj Rmarg Rm Dw Hm Hp Hr Ot Sf Sn Sr Sx-s Tt Ph Pb Ah Lf Lm Lpent Lpod Eh
Rhinella major 0,74 0,20 0,96 0,91 0,52 0,60 0 0,66 0,71 0,44 0,27 0,26 0,32 0,94 0,61 0,68 0,91 0,56 0,64 0,62
Rhinella margaritifera 0,79 0,70 0,85 0,87 0,85 0,65 0,82 0,78 0,87 0,82 0,59 0,71 0,71 0,48 0,48 0,64 0,74 0,70 0,49
Rhinella marina 0,12 0,37 0,70 0,61 0,92 0,50 0,39 0,78 0,89 0,68 0,69 0,27 0,31 0 0,07 0,71 0,50 0,32
Dendropsophus walfordi 0,96 0,62 0,71 0 0,78 0,84 0,52 0,32 0,16 0,35 0,82 0,38 0,80 0,99 0,35 0,54 0,39
Hypsiboas multifasciatus 0,81 0,87 0,26 0,91 0,93 0,73 0,57 0,30 0,54 0,76 0,33 0,79 0,95 0,42 0,58 0,34
Hypsiboas punctatus 1 0,72 0,96 0,92 1 0,93 0,43 0,76 0,35 0 0,58 0,65 0,33 0,45 0
Hypsiboas raniceps 0,62 1 0,96 0,96 0,86 0,37 0,73 0,41 0 0,67 0,74 0,29 0,44 0
Osteocephalus taurinus 0,51 0,40 0,80 0,91 0,74 0,83 0 0,04 0,12 0 0,43 0,47 0
Scinax fuscomarginatus 1 0,91 0,79 0,31 0,68 0,45 0 0,74 0,82 0,24 0,43 0
Scinax nebulosus 0,85 0,70 0,24 0,61 0,48 0 0,79 0,88 0,18 0,42 0
Scinax ruber 0,97 0,48 0,78 0,30 0 0,49 0,54 0,37 0,44 0
Scinax x-signatus 0,54 0,79 0,18 0 0,30 0,33 0,42 0,40 0
Trachycephalus typhonius 0,84 0,36 0,53 0,34 0,10 0,63 0,89 0,42
Phyllomedusa hypochondrialis 0,25 0,16 0,59 0,36 0,38 0,74 0,07
Pseudopaludicola boliviana 0,83 0,46 0,72 0,76 0,70 0,85
Adenomera hylaedactyla 0,08 0,24 0,88 0,73 0,98
Leptodactylus fuscus 0,85 0 0,58 0
Leptodactylus macrosternum 0,22 0,46 0,24
Leptodactylus pentadactylus 0,71 0,89
Leptodactylus podicipinus 0,63
Elachistocleis helianneae

106
Os maiores valores dos índices de sobreposição de nicho do sítio de vocalização
para altura em relação ao poleiro ocorreram entre os pares de espécies D. walfordi vs. H.
punctatus (0,98) e S. nebulosus vs. S. x-signatus (0,97). Quanto ao sítio de vocalização
para altura do poleiro em relação a superfície da água, houve sobreposição total nos pares
de espécies H. punctatus vs. H. multifasciatus; S. fuscomarginatus vs. H. punctatus; S.
ruber vs. D. walfordi; S. x-signatus vs. D. walfordi e S. x-signatus vs. S. ruber (Tabela 6).
Tabela 6 – Sobreposição de nicho do sítio de vocalização para altura em relação ao
poleiro (acima) e para altura do poleiro em relação a superfície da água (abaixo)
registrado pelas espécies de anuros da Savana amapaense, durante o período de janeiro a
dezembro de 2013. Os números em negrito indicam valores de sobreposição de nicho ≥ 95 %
As espécies que vocalizaram nas margens corpos d’água exibiram uma maior
sobreposição no sítio de vocalização do que as espécies que vocalizaram empoleiradas.
Estes resultados estão de acordo com Santos & Rossa-Feres (2007) e Maffei et al. (2011),
provavelmente como consequência da menor possibilidade de partilha espacial
decorrente do uso do espaço bidimensional. Adicionalmente, variações no conjunto dos
descritores do sítio de vocalização, incluindo a distância da margem, vegetação
predominantemente herbácea dos corpos d'água amostrados e a partilha vertical, geraram
uma baixa sobreposição entre os anuros que vocalizaram empoleirados.
Essa pequena sobreposição registrada no presente estudo está de acordo com a
segregação verificada em outras comunidades de anuros da região Neotropical (Conte et
al. 2013, Costa et al. 2013, Gambale et al. 2014). Alguns estudos (Santos & Rossa-Feres
2007, Silva et al. 2008) parecem indicar que a sobreposição no sítio de vocalização é
diretamente relacionada com a riqueza de espécies, com baixa sobreposição em
Dw Hm Hp Hr Ot Sf Sn Sr Sx-s Tt Ph
Dendropsophus walfordi 0,19 0,98 0,50 0,66 0,73 0,59 0,81 0,38 0,52 0,85
Hypsiboas multifasciatus 0,51 0,24 0,94 0,18 0,36 0,81 0,71 0,89 0 0,46
Hypsiboas punctatus 0,59 1 0,56 0,58 0,84 0,68 0,85 0,49 0,46 0,89
Hypsiboas raniceps 0,94 0,78 0,84 0,33 0,63 0,94 0,91 0,95 0,14 0,71
Osteocephalus taurinus 0,06 0,18 0,17 0,12 0,22 0,26 0,52 0,12 0,91 0,64
Scinax fuscomarginatus 0,68 0,98 1 0,90 0,17 0,84 0,79 0,72 0,17 0,82
Scinax nebulosus 0,33 0,98 0,95 0,64 0,18 0,91 0,92 0,97 0,12 0,89
Scinax ruber 1 0,52 0,60 0,94 0,06 0,69 0,33 0,83 0,33 0,30
Scinax x-signatus 1 0,63 0,70 0,98 0,08 0,78 0,46 1 0 0,41
Trachycephalus typhonius 0,19 0,58 0,57 0,38 0,88 0,54 0,59 0,20 0,27 0,88
Phyllomedusa hypochondrialis 0,29 0,88 0,85 0,57 0,60 0,82 0,79 0,89 0,65 0,59

107
comunidades com pequeno número de espécies e alta sobreposição em comunidades com
grande riqueza de espécies.
Apesar da grande amplitude de nicho para os sítios de vocalização, a sobreposição
entre as espécies foi pequena. Este resultado evidencia uma homogeneidade estrutural
presente nos corpos d'água que limitam as possibilidades de partilha espacial e temporal
entre as espécies de anuros (Toledo et al. 2003, Santos & Rossa-Feres 2007, Vasconcelos
& Rossa-Feres 2008, Vasconcelos et al. 2009, Silva et al. 2011).
Os modelos nulos mostraram que a sobreposição média para a distância do sítio
de vocalização até a margem mais próxima ao corpo d’água (média observada = 0,54;
média esperada = 0,49; p = 0,013) e altura do poleiro (média observada = 0,59; média
esperada = 0,52; p = 0,047) entre as espécies de anuros da Savana amapaense foi
significativamente diferente entre os valores observado e esperado pelo modelo nulo,
indicando existência de estrutura quanto aos sítios de vocalização (Figura 5).
Figura 5 – Simulações da sobreposição dos sítios de vocalização até a margem mais próxima ao
corpo d’água (a) e altura do poleiro (b) da taxocenose de anuros da Savana amapaense, durante o
período de janeiro a dezembro de 2013. As setas indicam as médias observadas de sobreposição
de nicho quanto aos sítios de vocalização.

108
Isto pode ter sido originado pelo fato das espécies de anuros utilizarem os sítios
de vocalização de forma semelhante, co-ocorrendo para reduzir a competição, permitindo
a coexistência entre as espécies (Griffin & Silliman 2011) e mudanças nos modos
reprodutivos devido a condições adversas (Toledo et al. 2011). Como o estabelecimento
dos sítios de vocalização é ocupado durante pouco tempo (uma noite, ou pouco menos do
que isso, dependendo da espécie) não há tempo para a competição atuar, gerando uma
ocupação aleatória.
Segundo Melo et al. (2007), em áreas abertas a heterogeneidade estrutural é muito
baixa e existe uma grande semelhança estrutural entre os corpos d’água. Mesmo
utilizando áreas, aparentemente, pouco heterogêneas, as espécies possuem diferentes
modos do ciclo de vida, incluindo os modos reprodutivos, permitindo a ocupação do
mesmo local.
Os modelos nulos mostraram que a sobreposição média para altura em relação ao
poleiro e a superfície da água, para as espécies cujos machos vocalizam empoleirados
(média observada = 0,60; média esperada = 0,62; p = 0,430) não foi significativamente
diferente entre os valores observado e esperado pelo modelo nulo, indicando ausência de
estrutura (Figura 6).
Figura 6 – Simulações da sobreposição dos sítios de vocalização em relação à altura do poleiro
em relação a superfície da água da taxocenose de anuros da Savana amapaense, durante o período
de janeiro a dezembro de 2013. A seta indica a média observada de sobreposição de nicho quanto
aos sítios de vocalização.

109
De modo geral, a presença de anuros em poleiros acima dos corpos d’água
depende da disponibilidade de poleiros emergentes. A diferenciação no modo em que as
espécies exploram o microhabitat resulta em uma partilha de recursos. Espécies com
sobreposição no sítio de vocalização apresentam segregação nos parâmetros acústicos e
espécies com sobreposição nos parâmetros acústicos tendem a compartilhar o sítio de
vocalização (Silva et al. 2008).
5.5. Comportamentos reprodutivos
Durante as observações de campo foram registrados 10 comportamentos
reprodutivos (Tabela 7). Todos os sinais visuais foram exibidos em fração de segundos,
e, quase sempre foram repetidos várias vezes e acompanhados, ou alternados, pela
emissão do canto de anúncio.
Tabela 7 – Repertório do comportamento reprodutivo exibido pelas quatro espécies de anuros da
Savana amapaense: Leptodactylus fuscus; Phyllomedusa hypochondrialis; Hypsiboas punctatus
e Rhinella margaritifera. As descrições dos comportamentos foram baseadas em Hodl &
Amézquita (2001)1 e Hartmann et al. (2005)2. O asterisco (*) indica que a descrição foi adaptada.
Comportamentos Descrição
Postura ereta 1, 2 Extensão dos braços e elevação da parte anterior do corpo
Postura imóvel Os membros são retraídos junto ao corpo, mantendo a
superfície ventral do corpo em contato com o substrato.
Andando e Saltando 1, 2 Pequenos passos para qualquer direção ou saltos rápidos
para os lados, com movimentos descontínuos, elevando os
membros rapidamente do substrato.
Movimentos circulares 1 Movimentação ao redor de outro indivíduo ou simplesmente
girando em torno de seu próprio eixo
Exibição da região gular 1, 2 Pulsação da região gular sem produção de som
Exibição do saco vocal 2 O saco vocal infla, exibindo a coloração, sem vocalização
Exibição dos membros 2 Movimentos de extensão dos membros
Elevação do corpo 2 Elevação do corpo se estendendo pelos quatro membros
Elevação dos braços 1 * Extensão dos braços em ângulo de 900, provocando
vibrações
Vibração do corpo 1 * Movimentos de contração da região abdominal do corpo
provocando vibração e sons

110
Os repertórios comportamentais exibidos pelos machos das espécies de anuros da
Savana amapaense permitiu a delimitação de alguns comportamentos relacionados com
movimentos do corpo e exibição de sinais visuais. A análise de agrupamento (Coeficiente
de correlação cofenético = 0,9538) permitiu visualizar três agrupamentos para os
comportamentos reprodutivos das espécies de anuros da Savana amapaense (Figura 7).
Figura 7 – Análise de agrupamento (Cluster) dos comportamentos repodutivos exibido pelas
quatro espécies de anuros da Savana amapaense: Grupo 1 – Leptodactylus fuscus e Phyllomedusa
hypochondrialis; Grupo 2 – Hypsiboas punctatus; Grupo 3 – Rhinella margaritifera, durante o
período de janeiro a dezembro de 2013.
No presente estudo, os comportamentos mais frequentes das espécies de anuros
da Savana amapaense foram exibição da região gular, exibição do saco vocal e postura
imóvel. Esses comportamentos ocorreram pelo acesso direto a fêmeas reprodutivas nos
sítios de vocalização, e consequentemente, aumento do sucesso reprodutivo (Wells 1977,
Sullivan & Kwiatkowski 2007).
Entre as quatro espécies registradas, apenas H. punctatus apresentou
características que concordam com o sistema em lek de reprodução (Emlen & Oring
1977), onde: (1) ocorrem agrupamentos de machos em sítios de vocalização; (2) não
apresentam cuidado parental; (3) possuem padrão prolongado de reprodução. De acordo
com Toledo et al. (2005), o sistema de lek é amplamente difundido entre os hilídeos e em
espécies que vocalizam em coros reprodutivos.

111
5.5.1. Leptodactylus fuscus
Para L. fuscus, a Tabela 8 sumariza os tipos de comportamentos reprodutivos,
número de observações e a frequência de ocorrência para os machos de L. fuscus
registrados na Savana gramíneo lenhosa.
Tabela 8 – Comportamento reprodutivo, número de observações e frequência de ocorrência para
os machos de L. fuscus registrado na Savana gramíneo lenhosa, durante o período de janeiro a
dezembro de 2013.
Comportamento reprodutivo Número de observações Frequência (%)
Exibição do saco vocal 65 17.5
Postura imóvel 68 18.3
Exibição da região gular 69 18.5
Elevação do corpo 79 21.2
Postura ereta 91 24.5
A análise de agrupamento (Coeficiente de correlação cofenético = 0,9592)
permitiu visualizar dois agrupamentos para o comportamento reprodutivo de L. fuscus
(Figura 8).
Figura 8 – Análise de agrupamento (Cluster) do comportamento repodutivo de Leptodactylus
fuscus (Savana gramíneo lenhosa), durante o período de janeiro a dezembro de 2013, registrado
na Savana amapaense.

112
5.5.2. Phyllomedusa hypochondrialis
Para a espécie P. hypochondrialis foram descritos quatro tipos de comportamento
reprodutivo. A Tabela 9 sumariza o comportamento reprodutivo, número de observações
e a frequência de ocorrência para os machos de P. hypochondrialis registrado na Savana
gramíneo lenhosa-arbustiva.
Tabela 9 – Comportamento reprodutivo, número de observações e frequência de ocorrência para
os machos de Phyllomedusa hypochondrialis registrado na Savana gramíneo lenhosa-arbustiva,
durante o período de janeiro a dezembro de 2013.
Comportamento reprodutivo Número de observações Frequência (%)
Postura imóvel 62 23.0
Exibição dos membros 64 23.8
Exibição da região gular 70 26.0
Exibição do saco vocal 73 27.1
A análise de agrupamento (Coeficiente de correlação cofenético = 0,9131)
permitiu visualizar dois agrupamentos para o comportamento reprodutivo de P.
hypochondrialis (Figura 9).
Figura 9 – Análise de agrupamento (Cluster) do comportamento reprodutivo de Phyllomedusa
hypochondrialis (Savana gramíneo lenhosa-arbustiva), durante o período de janeiro a dezembro
de 2013, registrado na Savana amapaense.
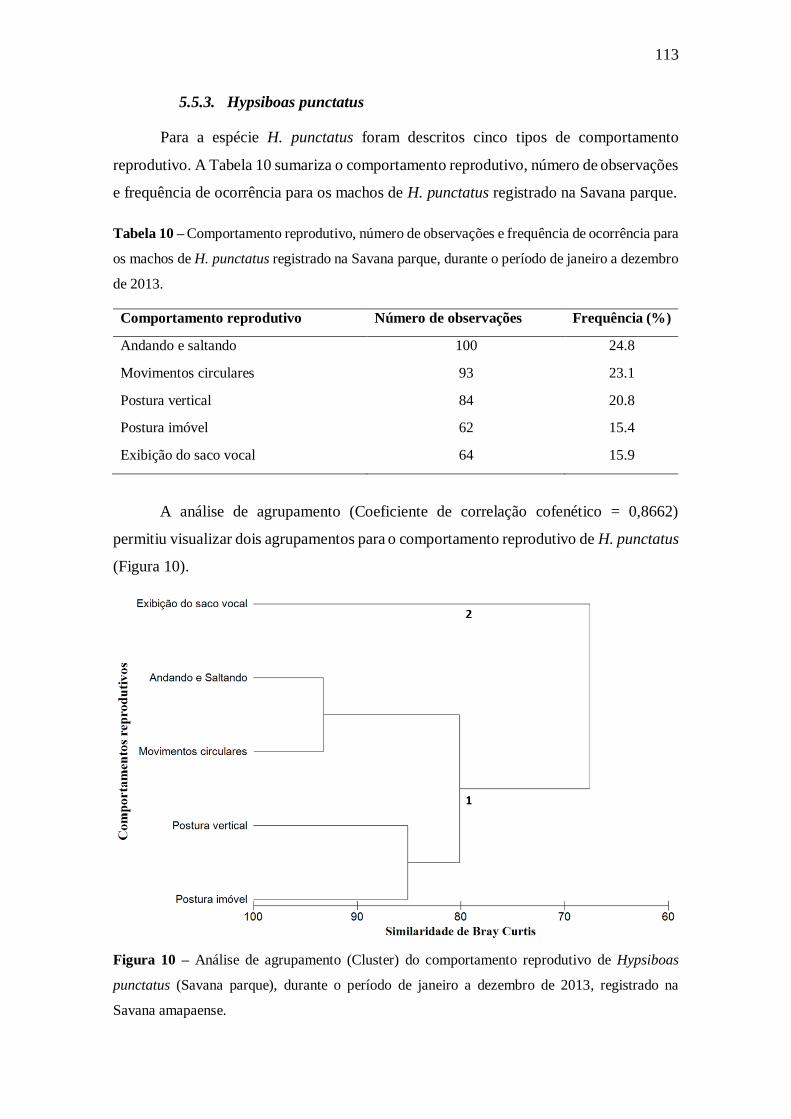
113
5.5.3. Hypsiboas punctatus
Para a espécie H. punctatus foram descritos cinco tipos de comportamento
reprodutivo. A Tabela 10 sumariza o comportamento reprodutivo, número de observações
e frequência de ocorrência para os machos de H. punctatus registrado na Savana parque.
Tabela 10 – Comportamento reprodutivo, número de observações e frequência de ocorrência para
os machos de H. punctatus registrado na Savana parque, durante o período de janeiro a dezembro
de 2013.
Comportamento reprodutivo Número de observações Frequência (%)
Andando e saltando 100 24.8
Movimentos circulares 93 23.1
Postura vertical 84 20.8
Postura imóvel 62 15.4
Exibição do saco vocal 64 15.9
A análise de agrupamento (Coeficiente de correlação cofenético = 0,8662)
permitiu visualizar dois agrupamentos para o comportamento reprodutivo de H. punctatus
(Figura 10).
Figura 10 – Análise de agrupamento (Cluster) do comportamento reprodutivo de Hypsiboas
punctatus (Savana parque), durante o período de janeiro a dezembro de 2013, registrado na
Savana amapaense.

114
5.5.4. Rhinella margaritifera
Para R. margaritifera, a Tabela 11 sumariza o comportamento reprodutivo, número
de observações e frequência de ocorrência para os machos de R. margaritifera registrado
na Savana arbórea.
Tabela 11 – Comportamento reprodutivo, número de observações e frequência de ocorrência para
os machos de R. margaritifera registrado na Savana parque, durante o período de janeiro a
dezembro de 2013.
Comportamento reprodutivo Número de observações Frequência (%)
Movimentos circulares 67 23.1
Vibração do corpo 65 22.4
Postura ereta 63 21.7
Vibração dos braços 58 20.0
Exibição da região gular 37 12.8
A análise de agrupamento (Coeficiente de correlação cofenético = 0,9355)
permitiu visualizar quatro agrupamentos para o comportamento reprodutivo de R.
margaritifera (Figura 10).
Figura 10 – Análise de agrupamento (Cluster) do comportamento reprodutivo de Rhinella
margaritifera (Savana arbórea), durante o período de janeiro a dezembro de 2013, registrado na
Savana amapaense.

115
Os sinais visuais foram exibidos principalmente em contextos reprodutivos,
aumentando a probabilidade dos machos de atrair fêmeas reprodutivas, e em encontros
agonísticos (Preininger et al. 2009). Para Hödl & Amézquita (2001), um sinal é
considerado visual quando o evento comportamental (1) gera um efeito visual durante
uma interação intra ou interespecífica, (2) é repetitivo, conspícuo e estereotipado e (3)
provavelmente provoca uma resposta imediata. Muitos anuros podem empregar posturas
estereotipadas e exibições visuais, além da comunicação acústica como sinais de ameaça
(Pombal Jr. et al. 1994). Neste sentido, muitos comportamentos complexos têm sido
descritos e cada vez mais se busca compreender o papel das interações sociais neste grupo
(Hartmann et al. 2004, Giasson & Haddad 2007).
Em alguns casos, a evolução dos sinais visuais pode ser favorecida pela pressão
das condições ambientais (Hödl & Amézquita 2001) e, alguns estudos demonstraram que
os sinais visuais são usados em diferentes contextos sociais (Barros & Feio 2011, Lipinski
et al. 2012), indicando que a sinalização visual é diversificada e tem evoluído de forma
independente em diversas famílias de anuros (Hödl & Amézquita 2001).
No caso dos anuros noturnos, que participam de coros durante a atividade
reprodutiva, o intenso barulho da vocalização simultânea de um grande número de
machos pode estar relacionado com o aparecimento de sinais visuais. O uso de sinais
visuais também pode estar diretamente relacionado aos modos reprodutivos (Wogel &
Pombal Jr. 2007).
Para L. fuscus, os comportamentos de postura ereta e imóvel, a elevação do corpo
e as características dos ninhos de espuma construídos em câmaras subterrâneas nos
momentos que antecedem a corte podem representar um critério de seleção pelas fêmeas.
O comportamento reprodutivo sugere que a escolha dos machos pelas fêmeas de L. fuscus
pode ser feito de três maneiras não excludentes: (1) através da vocalização; (2)
características das câmaras subterrâneas. Essas características foram registradas por
Martins (1988) em lagoa temporária e por Lucas et al. (2008) e De-Carvalho et al. (2008)
para L. fuscus em área de Cerrado, e parecem ser comuns para as espécies de
leptodactlídeos (e.g. Leptodactylus furnarius, L. troglodytes e L. mystacinus) que
constroem ninhos de espuma em câmaras subterrâneas próximas a corpos d’água
(Kokubum & Sousa 2008, Silva & Giaretta 2008, Azarak 2012).
Para P. hypochondrialis, o comportamento de exibição dos membros está
relacionado primariamente com a localização dos machos pelas fêmeas (Hartmann et al.
2005). Os sinais visuais exibibidos garantem a localização dos machos e o sucesso

116
reprodutivo, especialmente quando o amplexo ocorre em poleiros acima dos corpos
d’água. Como desova ocorre sobre folhas pendentes em corpos d’água, esses sítios
reprodutivos devem ser guardados pelos machos, já que folhas mais bem posicionadas
podem significar garantia de que a desova caia integralmente dentro da água, assegurando
a sobrevivência dos girinos. Nesse caso, os sinais visuais podem ser importantes fatores
de manutenção de espaço entre os machos, deixando-os visíveis e facilitando a sua
localização pelas fêmeas reprodutivas (Hartmann et al. 2005).
Os movimentos circulares e andando e saltando exibidos por H. punctatus foi
realizado nos momentos em que o macho se aproxima de fêmeas reprodutivas e foi
observado sempre acompanhado do canto de anúncio. A associação entre o canto de
anúncio e a ocorrência de sinais visuais foi também registrada por Caldart et al. (2014)
sugerindo que, além do comportamento reprodutivo, pode ser exibido em interações
agonísticas. Em H. punctatus, o macho pode dar pequenos saltos para qualquer direção,
sempre voltando ao mesmo lugar, ou pequenos passos em direção a fêmea. De acordo
com Wells (1980) e Juncá et al. (1994) estes movimentos são extremamente visíveis e
realizados durante o início da corte.
As vibrações corporais e elevação dos braços foram observadas apenas para R.
margaritifera. De acordo com Narins (1990), as vibracões do corpo e os movimentos dos
membros podem estar relacionadas com a comunicação sísmica, podendo ocorrer
simultaneamente com os sinais visuais. O comportamento de produzir sinais sísmicos
descritos aqui, representa um potencial modo de sinalização visual que pode ser
importante para os anuros terrestres que se comunicam sobre a superfície da água (Forti
& Encarnação 2012).
No caso dos machos de R. margaritifera que formam agregações reprodutivas em
poças temporárias, o intenso barulho da vocalização simultânea de um grande número de
machos está relacionado com o aparecimento de sinais visuais. Essa hipótese é sustentada,
pois o barulho do coro pode reduzir a habilidade da fêmea em discriminar o canto de
machos coespecíficos (Hödl & Amézquita, 2001; Hartmann et al., 2005). Dessa maneira,
os sinais visuais facilitam a localização dos indivíduos da mesma espécie a curtas
distâncias, diminuindo o risco de predação (Hartmann et al. 2005).
A exibição da região gular e do saco vocal presente no comportamento
reprodutivo das três espécies de anuros registradas na Savana amapaense está relacionado
ao comportamento de corte e territorialidade exibida pelos machos (Hartmann et al. 2005,
Hirschmann & Hödl 2006). Estes comportamentos estão de acordo com o postulado por

117
Wells (1978, 1980) que verificou que a alteração de coloração da região gular e exibição
do saco vocal apresenta relação com à territorialidade exibida pelos machos e na
diferenciação de machos residentes e de machos satélites. As fêmeas por sua vez, têm
poucas oportunidades de escolha de machos em agregações explosivas, pois são
interceptadas logo que chegam aos sítios reprodutivos (Wells 2007).
Considerando que a maioria dos estudos sobre os sinais visuais são realizados nas
interações entre os machos, outros estudos, incluindo informações sobre os sinais visuais
utilizados pelas fêmeas são necessários. A combinação de atributos ecológicos
relacionados com a diversidade de sinais visuais exibidos pelos anfíbios anuros fornece
dados essenciais para compreensão das estratégias reprodutivas e interações sociais,
subsidiando estratégias de conservação, de modo a satisfazer os requisitos necessários
para a manutenção da diversidade do grupo.
Cabe então enfatizar que, devido a essas características comportamentais e
reprodutivas peculiares, os anuros são organismos-modelo para estudos de diversos
aspectos de história natural, principalmente sinais visuais relacionados ao comportamento
reprodutivo (Prado et al. 2002). Apesar das espécies registradas na Savana amapaense
apresentarem um repertório de sinais visuais semelhantes, é plausível a hipótese de que o
sistema de comunicação destas espécies seja limitado por pressões seletivas e ambientais
registradas nas fitofisionomias.
Acreditamos que nossos resultados sobre a riqueza e composição de espécies,
ecologia de comunidades e comportamento reprodutivo aplicados a história natural,
possam ser úteis em ações conservacionistas, além de encorajar futuros estudos com as
espécies de anuros de áreas de Savana
6. REFERÊNCIAS BIBLIOGRÁFICAS
Aguiar, L.M.S., Machado, R.B. & Marinho-Filho, J. 2004. A diversidade biológica do
Cerrado. In: Cerrado: ecologia e caracterização (Org. por L.M. Aguiar & A.J.A.
Camargo), pp.17-40. Brasília: Embrapa Cerrados.
Alvares, C.A., Stape, J.L., Sentelhas, P.C., Gonçalves, J.L.M. & Sparovek, G. 2013.
Köppen’s climate classification map for Brazil. Meteorologische Zeitschrift, 22 (6),
711-728.
Arak, A. 1983. Male-male competition and mate choice in anuran amphibians. In: Mate
Choice (Org. por P. Batenson), pp. 181-210. Cambridge: Cambridge Universitu Press.

118
Ayres, M., Ayres Jr., M.A., Ayres, D.L. & Dos Santos, A.S. 2007. BioEstat: aplicações
estatísticas nas áreas das ciências biológicas e médicas. Belém: Publicações Avulsas
Mamirauá.
Barbosa, R.I., Nascimento, S.P., Amorim, P.A.F. & Silva, R.F. 2005. Notas sobre a
composição arbóreo-arbustiva de uma fisionomia das savanas de Roraima, Amazônia
Brasileira. Acta Botanica Brasilica, 19 (2), 323-329.
Barros, A.B. & Feio, R.N. 2011. Visual communication in Scinax maracaya (Cardoso
& Sazima, 1980) (Anura: Hylidae) at the Serra da Canastra National Park, southeastern
Brazil. Herpetology Notes, 4: 103-104.
Bertoluci, J. & Rodrigues, J.T. 2002. Seasonal patterns of breeding activity of Atlantic
Rainforest anurans at Boracéia, Southeastern Brazil. Amphibia-Reptilia, 23 (2), 161-
167.
Bitar, Y.O.C., Pinheiro, L.P.C., Abe, P.S., Santos-Costa, M.C. 2012. Species
composition and reproductive modes of anurans from a transitional Amazonian forest,
Brazil. Zoologia, 29 (1), 19-26.
Caldart, V.M., Iop, S., Cechin, S.Z. 2014. Social interactions in a neotropical stream
frog reveal a complex repertoire of visual signals and the use of multimodal
communication. Behaviour, 151: 719-739.
Castro, D.P., Borges-Leite, M.J., Lima, D.C. & Borges-Nojosa, D.M. 2013. Parental
care in two species of Leptodactylus Fitzinger, 1826 (Anura, Leptodactylidae) in
north-eastern Brazil. Herpetology Notes, 6, 267-269.
Conte, C.E. & Rossa-Feres, D.C. 2007. Riqueza e distribuição espaço-temporal de
anuros em um remanescente de Floresta com Araucária no sudeste do Paraná. Revista
Brasileira de Zoologia, 24, 1025-1037.
Conte, C.E., Silva, D.R., Rodrigues, A.P. 2013. Anurofauna da bacia do Rio Tijuco,
Minas Gerais, Brasil e sua relação com taxocenoses de anfíbios do Cerrado e suas
transições. Iheringia, 103 (3), 280-288.
Costa, W.P., Almeida, S.C. & Jim, J. 2013. Anurofauna em uma área na Depressão
Periférica, no centro-oeste do estado de São Paulo, Brasil. Biota Neotropica, 13 (2),
163-174.
Crawford, J.A., Shepard, D.B. & Conner, C.A. 2009. Diet composition and overlap
between recently metamorphosed Rana areolata and Rana sphenocephala:
implications for a frog of conservation concern. Copeia, 2009 (4), 642-646.

119
Crump, M.L. 1974. Reproductive strategies in a tropical anuran community. University
of Kansas, Museum of Natural History, Miscellaneous Publications, 61, 1-68.
Crump, M.L. & Scott, N.J.J. 1994. Visual encounter surveys. In: Measuring and
monitoring biological diversity: standard methods for amphibians (Org. por W.R.
Heyer., M.A. Donnelly., R.W. McDiarmid., L.A.C. Hayek & M.S. Foster), pp. 84-92.
Washington: Smithsonian Institution Press.
Crump, M.L. 1995. Parental Care. In: Amphibian Biology, Volume 2: Social Behavior
(Org. por H. Heatwole & B.K. Sullivan), pp. 519-522. Australia: Surrey Beatty &
Sons.
De Sá, R.O., Brandão, R.A. & Guimarães, L.D. 2007. Description of the tadpole of
Leptodactylus pustulatus Peters 1870 (Anura: Leptodactylidae). Zootaxa, 1523, 49-58.
Del Lama, F., Rocha, M.D., Andrade, M.A. & Nascimento, L.B. 2011. The use of
photography to identify individual tree frogs by their natural marks. South American
Journal of Herpetology, 6 (3), 198-204.
Delia, J., Cisneros-Heredia, D.F., Whitney, J, & Murrieta-Galindo, R. 2010.
Observations on the Reproductive Behavior of a Neotropical Glassfrog,
Hyalinobatrachium fleischmanni (Anura: Centrolenidae). South American Journal of
Herpetology, 5 (1), 1-12.
Doody, J. S. 1995. A photographic mark-recapture method for patterned amphibians.
Herpetological Review, 26 (1), 19-21.
Emlen, S.T. & Oring, L.W. 1977. Ecology, sexual selection and evolution of mating
sistems. Science, 197, 215-223.
Forti, L.R. & Encarnação, L.C. 2012. Water-wave production in the Neotropical frogs
Physalaemus albonotatus and Pseudopaludicola mystacalis: a seismic signal?
Salamandra, 48 (3), 181-184.
Gambale, P.G., Woitovicz-Cardoso, M., Vieira, R.R., Batista, V.G., Ramos, J. &
Bastos, R.P. 2014. Composição e riqueza de anfíbios anuros em remanescentes de
Cerrado do Brasil Central. Iheringia, 104 (1), 50-58.
García, C.G., Lescano, J.N. & Leynaud, G.C. 2013. Oviposition-site selection by
Phyllomedusa sauvagii (Anura: Hylidae): An arboreal nester inhabiting arid
environments. Acta Oecologica, 51, 62-65.
Giasson, L.O.M. & Haddad, C.F.B. 2007. Mate choice and reproductive biology of
Hypsiboas albomarginatus (Anura: Hylidae) in the Atlantic Forest, southeastern
Brazil. South American Journal of Herpetology, 2: 157-164.

120
Goottsberger, B. & Gruber, E. 2004. Temporal partitioning of reproductive activity in
a Neotropical anuran community. Journal of Tropical Ecology, 20, 271-280.
Gosner, K.L. 1960. A simplified table for staging anuran embryos and larvae with notes
on identification. Herpetologica, 16, 183-190.
Gotelli, N. & Colwell, R.K. 2001. Quantifying biodiversity: Procedures and pitfalls in
the measurement and comparison of species richness. Ecology Letters, 4, 379-391.
Gotelli N.J. & Entsminger, G.L. 2005. EcoSim: Null Models Software for Ecology.
Version 7.72. Encontrado em 10 de setembro de de 2013.
http://together.net/~gentsmin/ecosim.htm
Griffin, J.N. & Silliman, B.R. 2011. Resource partitioning and why it matters. Nature
Education Knowledge, 2, 8.
Haddad, C.F.B. & Prado, C.P.A. 2005. Reproductive modes in frogs and their
unexpected diversity in the Atlantic forest of Brazil. BioScience, 55 (3), 207-217.
Hammer, O., Harper, D.A.T. & Rian, P.D. 2001. Past: Palaeonthological statistics
software package for education and data analysis. Version. 1.37. Encontrado em 12 de
novembro de 2012. http://palaeo-electronica.org/2001_1/past/issue1_01.htm
Hartmann, M.T., Giasson, L.O.M., Hartmann, P.A. & Haddad, C.F.B. 2005. Visual
communication in Brasilian species of anurans from the Atlantic forest. Journal of
Natural History, 39, 1675-1685.
Heyer, W.R., Donnelly, M.A., Mcdiarmid, R.W., Hayek, L.C. & Foster, M.S. 1994.
Measuring and monitoring biological diversity: standard methods for amphibians.
Washington: Smithsonian Institution.
Hirschmann, W. & Hödl, W. 2006. Visual signaling in Phrynobatrachus krefftii
Boulenger, 1909 (Anura: Ranidae). Herpetologica, 62, 18-27.
Hödl, W. 1990. Reproductive diversity in Amazonian lowland frogs. Fortschritte der
Zoologie, 38, 41-60.
Hödl, W. & Amézquita, A. 2001. Visual signaling in anuran amphibians. In: Anuran
communication (Org. por M.J. Ryan), pp. 121-141. Washington: Smithsonian
Institution Press.
Howard, R.D. 1978. The evolution of mating strategies in bullfrogs, Rana catesbeiana.
Evolution, 32 (4), 850-871.
Juncá, F.A., Altig, R. & Gascon, C. 1994. Breeding biology of Colostethus stepheni, a
dendrobatid frog with a nontransported nidicolous tadpole. Copeia, 1994: 747-750.

121
Kokubum, M.N.C. & Souza, M.B. 2008. Reproductive ecology of Leptodactylus aff.
hylaedactylus (Anura, Leptodactylidae) from an open area in Northern Brazil. South
American Journal of Herpetology, 3 (1), 15-21.
Kopp, K., Signorelli, L. & Bastos, R.P. 2010. Distribuição temporal e diversidade de
modos reprodutivos de anfíbios anuros no Parque Nacional das Emas e entorno, estado
de Goiás, Brasil. Iheringia, 100 (3), 192-200.
Krebs, C.J. 2014. Ecological methodology. 3a ed. Vancouver: University of British
Columbia.
Lehner, P. 2003. Handbook of ethological methods. 2a ed. Cambridge: University of
Cambridge.
Lipinski, V.M., Caldart, V.M. & Iop, S. 2012. Visual communication in Hypsiboas
curupi (Anura: Hylidae) at Parque Estadual do Turvo, southern Brazil. Phyllomedusa,
11 (1): 71-74.
Lucas, E.M., Brasileiro, C.A., Oyamaguchi, H.M. & Martins, M. 2008. The
reproductive ecology of Leptodactylus fuscus (Anura, Leptodactylidae): new data
from natural temporary ponds in the Brazilian Cerrado and a review throughout its
distribution. Journal of Natural History, 42, 2305-2320.
Maffei, F., Ubaid, F.K. & Jim, J. 2011. Anurans in an open cerrado area in the
municipality of Borebi, Sao Paulo state, Southeastern Brazil: habitat use, abundance
and seasonal variation. Biota Neotroprica, 11, 221-233.
Martins, I.A. 2001. Parental care behaviour in Leptodactyulus podicipinus (Cope, 1982)
(Anura, Leptodactylidae). Herpetological Journal, 11, 29-32.
Melém-Júnior, N.J., Farias-Neto, J.T. & Yokomizo, G.K.I. 2003. Caracterização dos
Cerrados do Amapá. Comunicado Técnico Embrapa, 105, 1-5.
Melo, G.V., Rossa-Feres, D.C. & Jim, J. 2007. Variação temporal no sítio de
vocalização em uma comunidade de anuros de Botucatu, Estado de São Paulo, Brasil.
Biota Neotropica, 7 (2), 93-102.
Miranda, I.S., Absy, M.L. & Rebêlo, G.H. 2003. Community Structure of Woody
Plants of Roraima Savannahs, Brazil. Plant Ecology, 164, 109-123.
Narins, P.M. 1990. Seismic communication in anuran amphibians. Bioscience, 40, 268-
274.
Pianka, E. R. 1974. Evolutionary Ecology. New York: Harper and Row.

122
Pombal Jr., J.P., Sazima, I. & Haddad, C.F.B. 1994. Breeding behavior of the pumpkin
toadlet, Brachycephalus ephippium (Brachycephalydae). Journal of Herpetology, 28,
526-519.
Pombal Jr., J.P. 1997. Distribuição espacial e temporal de anuros (Amphibia) em uma
poça permanente na Serra de Paranapiacaba, sudeste do Brasil. Revista Brasileira de
Biologia, 57, 583-594.
Pombal Jr., J.P. & Haddad, C.F.B. 2005. Estratégias e modos reprodutivos de anuros
(Amphibia) em uma poça permanente na Serra de Paranapiacaba, Sudeste do Brasil.
Papéis avulsos de zoologia, 45 (15), 201-213.
Pombal Jr., J.P. & Haddad, C.F.B. 2007. Estratégias e modos reprodutivos em anuros.
In: Herpetologia no Brasil II (Org. por L.B. Nascimento., P.M.E. Oliveira), pp. 101-
116. Belo Horizonte: Sociedade Brasileira de Herpetologia.
Prado, C.P.A., Uentanabaro, M. & Haddad, C.F.B. 2002. Description of a new
reproductive mode in Leptodactylus (Anura, Leptodactylidae), with a review of the
reproductive specialization toward terrestriality in the genus. Copeia, 2002 (4), 1128-
1133.
Prado, C.P.A.; Uetanabaro, M. & Haddad, C.F.B. 2005. Breeding activity patterns,
reproductive modes, and habitat use by anurans (Amphibia) in a seasonal environment
in the Pantanal, Brasil. Amphibia-Reptilia, 26, 211-221.
Prado, G.M. & Pombal Jr. J.P. 2005. Distribuição espacial e temporal dos anuros em
um brejo da Reserva Biológica de Duas Bocas, sudeste do Brasil. Arquivos do Museu
Nacional, 63 (4), 685-705.
Preininger, D., Boeckle, M., & Hodl, W. 2009. Communication in noide enviroments
II: Visual signaling behavior of male foot flagging frogs Staurois latopalmatus.
Herpetologica, 65 (2), 166-173.
Rabelo, B.V., Pinto, A.C., Simas, A.P.S.C., Tardin, A.T., Fernandes, A.V., Souza,
C.B., Monteiro, E.M.P.B., Facundes, F.S., Ávila, J.E.S., Souza, J.S.A., Costa-
Guedes, L.A., Penha, O.A.A., Melo, R.M.S. & Gibson, V.M. 2008.
Macrodiagnóstico do Estado do Amapá: primeira aproximação do ZEE. Macapá:
Instituto de Pesquisas Científicas e Tecnológicas do Estado do Amapá (IEPA).
Rodrigues, A.P., Giaretta, A.A., Silva, D.R. & Facure, K.G. 2011. Reproductive
features of three maternal-caring species of Leptodactylus (Anura: Leptodactylidae)
with a report on alloparental care in frogs. Journal of Natural History, 45, 33-34.

123
Santos, E.M. & Amorim, F.O. 2006. Cuidado parental em Leptodactylus natalensis
(Amphibia, Anura, Leptodactylidae). Iheringia, 96, 491-494.
Santos, T.G. & Rossa-Feres, D.C. 2007. Similarities in calling site and advertisement
call among anuran amphibians in southeastern Brazil. South American Journal of
Herpetology, 2 (1), 17-30.
Silva, W.R. & Giaretta, A.A. 2008. Seleção de sítios de oviposição em anuros
(Lissamphibia). Biota Neotropica, 8 (3): 243-248.
Silva, R.A., Martins, I.A. & Rossa-Feres, D.C. 2008. Bioacústica e sítio de vocalização
em taxocenoses de anuros de área aberta no noroeste paulista. Biota Neotropica, 8 (3),
123-134.
Silva, R.A., Martins, I.A. & Rossa-Feres, D.C. 2011. Environmental heterogeneity:
Anuran diversity in homogeneous environments. Zoologia, 28 (5), 610-618.
Sullivan, B.K. & Kwiatkowski, M.A. 2007. Courtship displays in anurans and lizards:
theoretical and empirical contributions to our understanding of costs and selection on
males due to female choice. Functional Ecology, 21, 666-675.
Taborsky, M.; Oliveira, R.F. & Brockmann, H.J. 2008. The evolution of alternative
reproductive tactics: concepts and questions. In: Alternative reproductive tactics: an
integrative approach (Org. por R.F. Oliveira., M. Taborsky & H.J. Brockmann), pp.
1-21. Cambridge: Cambridge University Press.
Taborsky, M. & Brockmann, H.J. 2010. Alternative reproductive tactics and life
history phenotypes. In: Animal behaviour: Evolution and Mechanisms (Org. por P.
Kappeler), pp. 537-586. Berlin: Springer.
Toledo, L.F., Zina, J. & Haddad, C.F.B. 2003. Distribuição espacial e temporal de uma
comunidade de anfíbios anuros do município de Rio Claro, São Paulo, Brasil. Holos
Environment, 3 (2), 136-149.
Toledo, L.F. & Haddad, C.F.B. 2005. Acoustic Repertoire and Calling Behavior of
Scinax fuscomarginatus (Anura, Hylidae). Journal of Herpetology, 39 (3), 455-464.
Toledo, L.F., Sazima, I. & Haddad, C.F.B. 2011. Behavioural defences of anurans: an
overview. Ethology Ecology & Evolution, 23 (1), 1-25.
Toledo, L.F., Garey, M.V., Costa, T.R.N., Lourenço-de-Moraes, R., Hartmann, M.T.
& Haddad, C.F.B. 2012. Alternative reproductive modes of Atlantic forest frogs.
Journal of Ethology, 30, 331-336.
Valdujo, P.H., Carnaval, A.C. & Graham, C.H. 2013. Environmental correlates of
anuran beta diversity in the Brazilian Cerrado. Ecography, 36, 708-717.

124
Vargas-Salinas, F., Quintero-Ángel, A., Osorio-Domínguez, D., Rojas-Morales, J.A.,
Escobar-Lasso, S., Gutiérrez-Cárdenas, P.D.A., Rivera-Correa, M. &
Amézquita, A. 2014. Breeding and parental behaviour in the glass frog Centrolene
savagei (Anura: Centrolenidae). Journal of Natural History, 48 (27-18), 1689-1705.
Vasconcelos, T.S. & Rossa-Feres, D.C. 2005. Diversidade, distribuição espacial e
temporal de anfíbios anuros (Amphibia, Anura) na região Noroeste do Estado de São
Paulo, Brasil. Biota Neotropica, 5 (2), 1-14.
Vasconcelos, T.S. & Rossa-Feres, D.C. 2008. Habitat heterogeneity and use of physical
and acoustic space in anuran communities in southeastern Brazil. Phyllomedusa, 7,
125-140.
Vasconcelos, T.S., Santos, T.G., Rossa-Feres, D.C. & Haddad, C.F.B. 2009. Influence
of the environmental heterogeneity of breeding ponds on anuran Assemblages from
southeastern Brazil. Canadian Journal of Zoology, 87, 699-707.
Vasconcelos, T.S., Santos, T.G., Haddad, C.F.B. & Rossa-Feres, D. C. 2010. Climatic
variables and altitude as predictors of anuran species richness and number of
reproductive modes in Brazil. Journal of Tropical Ecology, 26, 423-432.
Wells, K.D. 1977a. The social behavior of anuran amphibians. Animal Behaviour, 25,
666-693.
Wells, K.D. 1977b. Territoriality and male mating success in the green frog (Rana
clamitans). Ecology, Cambridge, 58, 750-762.
Wells, K.D. 1978. Territoriality in the green frog (Rana clamitans): vocalizations and
agonistic interactions. Animal Behaviour, 26, 1051-1063.
Wells, K.D. 1980. Intra and interspecific comunication in the neotropical frog Hyla
ebraccata. American Zoologist, 20 (4), 724.
Wells, K.D. 2007. The ecology and behavior of amphibians. Chicago: University of
Chicago Press.
Wogel, H., Abrunhosa, P. A. & Pombal Jr., J. P. 2005. Breeding behaviour and mating
success of Phyllomedusa rohdei (Anura, Hylidae) in south-eastern Brazil. Journal of
Natural History, 39, 2035-2045.
Wogel, H. & Pombal Jr. J.P. 2007. Comportamento reprodutivo e seleção sexual em
Dendropsophus bipunctatus (Spix, 1824) (Anura, Hylidae). Papéis Avulsos de
Zoologia, 47 (13), 165-174.
Zar J.H. 2010. Bioestatistical Analysis. 5a ed. New Jersey: Prentice-Hall.

125
Zimmerman, B.L. 1994. Audio Strip Transects. In: Measuring and monitoring
biological diversity: standard methods for amphibians (Org. por W.R. Heyer., M.A.
Donnelly., R.W. McDiarmid., L.A.C. Hayek & M.S. Foster), pp. 92-97. Washington:
Smithsonian Institution Press.

126
CONSIDERAÇÕES FINAIS
A composição e a riqueza de espécies, associada a ecologia de comunidades e ao
comportamento reprodutivo, ressaltam a importância da comunidade de anuros da Savana
amapaense como uma área de interesse conservacionista. O contato com outros domínios
fitogeográficos, em especial o Amazônico, determina um alto potencial de trocas de
espécies ao longo da história biogeográfica, favorecendo a alta riqueza e diversidade de
espécies em escala local e regional.
A ocorrência de novos registros de espécies de anfíbios anuros para a Savana
amapaense e a possibilidade de extinções locais, confrontada com a acelerada e intensa
destruição dos blocos vegetacionais remanescentes de formações abertas e florestais do
Estado do Amapá, apontam para uma necessidade urgente de ações de conservação e
inventários em áreas não amostradas.
A existência de estrutura de comunidades com relação a dieta e ao período de
atividade de vocalização e a ausência de estrutura com relação ao uso de habitat e
microhabitat, indicam que a plasticidade das espécies no uso do habitat e microhabitat
facilita a co-ocorrência diminuindo os efeitos da competição e permitindo a coexistência
local de espécies de anuros da Savana amapaense.
Os repertórios comportamentais exibidos pelas espécies de anuros da Savana
amapaense estão relacionados pelo acesso direto a fêmeas reprodutivas nos sítios de
vocalização e em encontros agonísticos. Esses comportamentos são descritos pela
primeira vez para as espécies Leptodactylus fuscus, Phyllomedusa hypochondrialis,
Hypsiboas punctatus e Rhinella margaritifera registrados em área de Savana amazônica
no Estado do Amapá. Desta forma, a realização de estudos futuros que permitam um
maior detalhamento dos comportamentos exibidos pelas espécies, podem auxiliar na
compreensão de padrões reprodutivos pouco explorados em áreas de Savana amazônica.
Adicionalmente, estudos biogeográficos, de filogeografia e de genética de
populações podem ajudar a elucidar as conexões históricas e o fluxo gênico entre as
populações de anuros das formações abertas e florestais da Savana amapaense,
proporcionando o aumento do conhecimento sobre a história de vida de espécies em
vastas áreas com ambientes naturais ainda pouco perturbados e, fornecendo subsídios
teóricos para medidas adequadas de conservação em outras áreas impactadas de Savana
amazônica.

127
REFERÊNCIAS
Aguiar, L.M.S., Machado, R.B. & Marinho-Filho, J. 2004. A diversidade biológica do
Cerrado. In: Cerrado: ecologia e caracterização (Org. por L.M. Aguiar & A.J.A.
Camargo), pp.17-40. Brasília: Embrapa Cerrados.
Alvares, C.A., Stape, J.L., Sentelhas, P.C., Gonçalves, J.L.M. & Sparovek, G. 2013.
Köppen’s climate classification map for Brazil. Meteorologische Zeitschrift, 22 (6),
711-728.
Araújo, C.O., Condez, T.H. & Sawaya, R.J. 2009. Anfíbios Anuros do Parque Estadual
das Furnas do Bom Jesus, sudeste do Brasil, e suas relações com outras taxocenoses
no Brasil. Biota Neotropica, 9 (2), 77-98.
Araújo, C.O., Corrêa, D.T. & Almeida-Santos, S.M. 2013. Anuros da Estação
Ecológica de Santa Bárbara, um remanescente de formações abertas de Cerrado no
estado de São Paulo. Biota Neotropica, 13 (3), 230-240.
Azarak, P.A., Nascimento, S.P. & Carvalho, C.M. 2011. Anfíbios do Lavrado de
Roraima. Biologia Geral e Experimental, 11 (1), 4-14.
Azarak, P.A. 2012. História natural de Leptodactylus fuscus (Schneider,1799) no
lavrado de Roraima (Amphibia: Anura: Leptodactylidae). Dissertação (Mestrado em
Recursos Naturais), Universidade Federal de Roraima, Boa Vista. 52 p.
Azevedo, L.G. 1967. Tipos eco-fisionômicos de vegetação do território federal do
Amapá. Revista Brasileira de Geografia, 29 (2), 25-51.
Azevedo-Ramos, C. & Galatti, U. 2002. Patterns of amphibian diversity in Brazilian
Amazonia: conservation implications. Biological Conservation, 103, 103-111.
Barbosa, R.I. & Miranda, I.S. 2005. Fitofisionomias e diversidade vegetal nas savanas
de Roraima. In: Savanas de Roraima: etnoecologia, biodiversidade e potencialidades
agrossilvipastoris (Org. por R.I Barbosa; H.A.M. Xaud & J.M. Costa e Souza), pp.
61-78. Boa Vista: FEMACT-RR.
Barbosa, R.I., Nascimento, S.P., Amorim, P.A.F. & Silva, R.F. 2005. Notas sobre a
composição arbóreo-arbustiva de uma fisionomia das savanas de Roraima, Amazônia
Brasileira. Acta Botanica Brasilica, 19 (2), 323-329.
Barbosa, R.I., Campos, C., Pinto, F. & Fearnside, P.M. 2007. The “Lavrados” of
Roraima: Biodiversity and Conservation of Brazil’s Amazonian Savannas. Functional
Ecosystems and Communities, 1 (1), 29-41.

128
Barbosa, R.I. & Campos, C. 2011. Detection and geographical distribution of clearing
areas in the savannas (‘lavrado’) of Roraima using Google Earth web tool. Journal of
Geography and Regional Planning, 4, 122-136.
Bastazini, C.V., Munduruca, J.F.V., Rocha, P.L.B. & Napoli, M.F. 2007. Which
environmental variables better explain changes in anuran community composition? A
case study in the restinga of Mata de São João, Bahia, Brazil. Herpetologica, 63, 459-
471.
Bastos, R.P. 2007. Anfíbios do Cerrado. In: Herpetologia no Brasil II. (Org. por L.B.
Nascimento & P.M.E. Oliveira), p. 87-100. Belo Horizonte: Sociedade Brasileira de
Herpetologia.
Bitar, Y.O.C., Pinheiro, L.P.C., Abe, P.S. & Santos-Costa, M.C. 2012. Species
composition and reproductive modes of anurans from a transitional Amazonian forest,
Brazil. Zoologia, 29 (1), 19-26.
Bitar, Y.O.C., Juen, L., Pinheiro, L.P.C. & Santos-Costa, M.C. 2014. Anuran Beta
Diversity in a Mosaic Anthropogenic Landscape in Transitional Amazon. Journal of
Herpetology, 48 (4), 000-000.
Both, C.; Kaefer, I. L., Santos, T.G. & Cechin, S.T.Z. 2008. An austral assemblage in
the Neotropics: seasonal occurrence correlated with photoperiod. Journal of Natural
History, 42 (3,4), 205-222.
Buckley, L.B. & Jetz, W. 2007. Environmental and historical constraints on global
patterns of amphibian richness. Proceedings of the Royal Society B: Biological
Sciences, 274, 1167-1173.
Buckley, L.B. & Jetz, W. 2008. Linking global turnover of species and environments.
Proceedings of the National Academy of Sciences, 105, 17863-17841.
Canavero, A., Arim, M., Naya, D.E., Camargo, A., Rosa, I. & Maneyro, R. 2008.
Calling activity patterns in an anuran assemblage: the role of seasonal trends and
weather determinants. North Western Journal of Zoology, 4 (1), 29-41.
Cavalcante, C.O., Flores, A.S. & Barbosa, R.I. 2014. Fatores edáficos determinando a
ocorrência de leguminosas herbáceas em savanas amazônicas. Acta Amazônica, 44 (3),
379-386.
Colli, G.R., Bastos, R.P. & Araújo, A.F.B. 2002. The character and dynamics of the
Cerrado herpetofauna. In: The Cerrados of Brazil: Ecology and Natural History of a
Neotropical Savanna. (Org. por P.S.Olivera & R.J. Marquis), pp. 223-241. New York:
Columbia University.

129
Conte, C.E., Silva, D.R. & Rodrigues, A.P. 2013. Anurofauna da bacia do Rio Tijuco,
Minas Gerais, Brasil e sua relação com taxocenoses de anfíbios do Cerrado e suas
transições. Iheringia, 103 (3), 280-288.
Costa, R.C., Facure, K.G. & Giaretta, A.A. 2006. Courtship, vocalization, and tadpole
description of Epipedobates flavopictus (Anura: Dendrobatidae) in southern Goiás,
Brazil. Biota Neotropica, 6 (1), 1-9.
Costa, T.R.N. & Toledo, L.F. 2013. Physalaemus spiniger (Anura: Leiuperinae): A Frog
with an Extensive Vocal Repertoire. Journal of Herpetology, 47 (4), 530-538.
Costa, W.P., Almeida, S.C. & Jim, J. 2013. Anurofauna em uma área na Depressão
Periférica, no centro-oeste do estado de São Paulo, Brasil. Biota Neotropica, 13 (2),
163-174.
Crump, M.L. 1974. Reproductive strategies in a tropical anuran community.
Miscellaneous Publications of the Museum of Natural History of the University of
Kansas, 61, 1-68.
Crump, M.L. 1982. Amphibian reproductive ecology on the community level. In:
Herpetological communities (Ogr. por N.J. Scott Jr), pp. 39-47. Washington: USA.
De-Carvalho, C.B., Freitas, E.B., Faria, R.G., Batista, R.C., Batista, C.C., Coelho,
W.A. & Bocchiglieri, A. 2008. História natural de Leptodactylus mystacinus e
Leptodactylus fuscus (Anura: Leptodactylidae) no Cerrado do Brasil Central. Biota
Neotropica, 8 (3), 105-115.
Delia, J., Cisneros-Heredia, D.F., Whitney, J. & Murrieta-Galindo, R. 2010.
Observations on the Reproductive Behavior of a Neotropical Glassfrog,
Hyalinobatrachium fleischmanni (Anura: Centrolenidae). South American Journal of
Herpetology, 5 (1), 1-12.
Diniz-Filho, J.A.F., Bini, L.M., Vieira, C.M., Blamires, D., Terribile, L., Bastos, R.,
Oliveira, G. & Barreto, B. 2008. Spatial patterns of terrestrial vertebrate species
richness in the Brazilian Cerrado. Zoological Studies, 47 (2), 146-157.
Duellman, W.E. & Trueb, L. 1994. Biology of Amphibians. Baltimore and London:
McGraw-Hill.
Ernst, R. & Rödel, M.O. 2006. Community assembly and structure of tropical leaf-litter
anurans. Ecotropica, 12, 113-129.
Fahr, J. & Kalko, E.K.V. 2011. Biome transitions as centres of diversity: habitat
heterogeneity and diversity patterns of West African bat assemblages across spatial
scales. Ecography, 34, 177-195.

130
Gambale, P.G., Woitovicz-Cardoso, M., Vieira, R.R., Batista, V.G., Ramos, J. &
Bastos, R.P. 2014. Composição e riqueza de anfíbios anuros em remanescentes de
Cerrado do Brasil Central. Iheringia, 104 (1), 50-58.
Giaretta, A.A., Menin, M., Facure, K.G., Kokubum, M.N.C. & De Oliveira Filho,
J.C. 2008. Species richness, relative abundance, and habitat of reproduction of
terrestrial frogs in the Triangulo Mineiro region, Cerrado biome, southeastern Brazil.
Iheringia, 98 (2), 181-188.
Haddad, C.F.B. & Prado, C.P.A. 2005. Reproductive modes in frogs and their
unexpected diversity in the Atlantic forest of Brazil. BioScience, 55 (3), 207-217.
Haffer, J. & Prance, G.T. 2002. Impulsos climáticos da evolução na Amazônia durante
o Cenozóico: sobre a teoria dos Refúgios da diferenciação biótica. Estudos Avançados,
16 (46), 175-206.
Harrison, S. & Cornell, H. 2008. Toward a better understanding of the regional causes
of local community richness. Ecology letters, 11, 969-979.
Hartel, T., Bancila, R. & Cogalniceanu, D. 2011. Spatial and temporal variability of
aquatic habitat use by amphibians in a hydrologically modified landscape. Freshwater
Biology, 56, 2288-2298.
Hartmann, M.T., Giasson, L.O.M., Hartmann, P.A. & Haddad, C.F.B. 2005. Visual
communication in Brasilian species of anurans from the Atlantic forest. Journal of
Natural History, 39, 1675-1685.
Hartmann, M.T., Hartmann, P.A. & Haddad, C.F.B. 2006. Repertório vocal de
Hylodes phyllodes (Amphibia, Anura, Hylodidae). Papéis Avulsos de Zoologia, 46
(17), 203-209.
Heyer, W.R., Donnelly, M.A., Mcdiarmid, R.W., Hayek, L.C. & Foster, M.S. 1994.
Measuring and monitoring biological diversity: standard methods for amphibians.
Washington: Smithsonian Institution.
Hödl, W. 1990. Reproductive diversity in Amazonian lowland frogs. Fortschritte der
Zoologie, 38, 41-60.
Hödl, W. & Amézquita, A. 2001. Visual signaling in anuran amphibians. In: Anuran
communication (Org. por M.J. Ryan), pp. 121-141. Washington: Smithsonian
Institution Press.
Howard, R.D. 1978. The evolution of mating strategies in bullfrogs, Rana catesbeiana.
Evolution, 32 (4), 850-871.

131
Jati, S.R., Fearnside, S.R. & Barbosa, R.I. 2014. Densidade da madeira de árvores em
savanas do norte da Amazônia brasileira. Acta Amazônica, 44 (1), 79-86.
Joly, C.A., Aidar, M.P.M., Klink, C.A., McGrath, D.G., Moreira, A.G., Moutinho,
P., Nepstad, D.C., Oliveira, A.A., Pott, A., Rodal, M.J.N. & Sampaio, E.V.S.B.
1999. Evolution of the Brazilian phytogeography classification systems: implications
for biodiversity conservation. Ciência e Cultura, 51, 331-348.
Klink, C.A. & Machado, R.B. 2005. Conservation of the Brazilian cerrado.
Conservation Biology, 19 (3), 707-713.
Kokubum, M.N.C. & Souza, M.B. 2008. Reproductive ecology of Leptodactylus aff.
hylaedactylus (Anura, Leptodactylidae) from an open area in Northern Brazil. South
American Journal of Herpetology, 3 (1), 15-21.
Kopp, K., Signorelli, L. & Bastos, R.P. 2010. Distribuição temporal e diversidade de
modos reprodutivos de anfíbios anuros no Parque Nacional das Emas e entorno, estado
de Goiás, Brasil. Iheringia, 100 (3), 192-200.
Lantyer-Silva, A.S.F., Solé, M. & Zina, J. 2014. Reproductive biology of a
bromeligenous frog endemic to the Atlantic Forest: Aparasphenodon arapapa
Pimenta, Napoli and Haddad, 2009 (Anura: Hylidae). Anais da Academia Brasileira
de Ciências, 86 (2), 867-880.
Machado, R.B., Ramos Neto, M.B., Pereira, P.G.P., Caldas, E.F., Gonçalves, D.A.,
Santos, N.S., Tabor, K. & Steininger, M. 2004. Estimativas de perda da área do
Cerrado brasileiro. Brasília: Conservation International do Brasil.
Maffei, F., Ubaid, F.K. & Jim, J. 2011. Anurans in an open cerrado area in the
municipality of Borebi, Sao Paulo state, Southeastern Brazil: habitat use, abundance
and seasonal variation. Biota Neotroprica, 11: 221-233.
Magnusson, W.E., Lima, P.A., Albernaz, A.L.K.M., Sanaiotti, T.M. & Guillaumet,
J.L. 2008. Composição florística e cobertura vegetal das savanas na região de Alter
do Chão, Santarém – PA. Revista Brasileira de Botânica, 31, 165-177.
Martins, M. 1988. Biologia reprodutiva de Leptodactylus fuscus em Boa Vista, Roraima
(Amphibia: Anura). Revista Brasileira de Biologia, 48 (4), 969-977.
Melém-Júnior, N.J., Farias-Neto, J.T. & Yokomizo, G.K.I. 2003. Caracterização dos
Cerrados do Amapá. Comunicado Técnico Embrapa, 105, 1-5.
Melo, M., Fava, F., Pinto, H.B.A., Bastos, R.P. & Nomura, F. 2013. Anuran diversity
(Amphibia) in the Extractivist Reserve Lado do Cedro, Goiás. Biota Neotropica, 13
(2), 205-217.

132
Menin, M., Rossa-Feres, D.C. & Giaretta, A.A. 2005. Resource use and coexistence of
two syntopic hylid frogs (Anura, Hylidae). Revista Brasileira de Zoologia, 22 (1), 61-
72.
Miranda, I.S., Absy, M.L. & Rebêlo, G.H. 2003. Community Structure of Woody
Plants of Roraima Savannahs, Brazil. Plant Ecology, 164, 109-123.
Miranda, D.B., Garey, M.V., Monteiro-Filho, E.L.A. & Hartmann, M.T.
2008. Sinalização visual e Biologia Reprodutiva de Dendropsophus werneri (Anura:
Hylidae) em área de Mata Atlântica no Estado do Paraná, Brasil. Papéis Avulsos de
Zoologia, 48 (29), 335-343.
Mittermeier, R.A., Fonseca, G.A.B., Rylands, A.B. & Brandon, K. 2005. A brief
history of biodiversity conservation in Brazil. Conservation Biology, 19 (3), 601-611.
Muniz, K.P.R., Giaretta, A.A., Silva, W.R. & Facure, K.G. 2008. Auto-ecologia de
Hypsiboas albopunctatus (Anura, Hylidae) em área de Cerrado no sudeste do
Brasil. Iheringia, 98 (2), 254-259.
Nascimento, S.P. 2005. Herpetofauna de Roraima: ênfase nas áreas abertas (Lavrado).
In: Savanas de Roraima: etnoecologia, biodiversidade e potencialidades
agrossilvipastoris (Org. por R.I. Barbosa., H.A.M. Xaud., J.M. Costa e Souza), pp.
123-134. Boa Vista: FEMACT-RR.
Neckel-Oliveira, S., Magnusson, W.E. & Lima, A.P. 2000. Diversity and distribution
of frogs in an Amazonian savanna in Brazil. Amphibia-Reptilia, 21 (3), 317-326.
Neckel-Oliveira, S., Galatti, U., Gordo, M., Pinheiro, L.P.C. & Maschio, G.F. 2012.
Anfíbios. In: Fauna da Floresta Nacional de Carajás: Estudos Sobre Vertebrados
Terrestres (Org. por F.D. Martins., A.F. Castilho., J. Campos., F.M. Hatano &, S.
Rolim), pp. 67-79. São Paulo: Editora Rona.
Oda, F.H., Bastos, R.P. & Lima, M.A.C.S. 2009. Taxocenose de anfíbios anuros no
Cerrado do Alto Tocantins, Niquelândia, Estado de Goiás: diversidade, distribuição
local e sazonalidade. Biota Neotropica, 9 (4), 219-232.
Paula, A. 2012. Estrutura e dinâmica de uma comunidade de anuros no hotspot de
biodiversidade do Cerrado. Dissertação (Mestrado em Biologia Animal),
Universidade de Brasília, Brasília. 71 p.
Pianka, E.R. 1994. Evolutionary Ecology. 5a ed. New York: Harper Collins.
Pinheiro, L.C., Bitar, Y.O.C., Galatti, U., Neckel-Oliveira, S. & Santos-Costa, M.C.
2012. Amphibians from southeastern state of Pará: Carajás Region, northern Brazil.
Check List, 8 (4), 693-702.

133
Pirani, R.M., Nascimento, L.B. & Feio, R.N. 2013. Anurans in a forest remnant in the
transition zone between cerrado and atlantic rain forest domains in southeastern Brazil.
Anais da Academia Brasileira de Ciências, 85 (3), 1093-1104.
Pombal Jr., J.P. & Haddad, C.F.B. 2005. Estratégias e modos reprodutivos de anuros
(Amphibia) em uma poça permanente na Serra de Paranapiacaba, Sudeste do Brasil.
Papéis avulsos de zoologia, 45 (15), 201-213.
Pombal Jr., J.P. & Haddad, C.F.B. 2007. Estratégias e modos reprodutivos em anuros.
In: Herpetologia no Brasil II (Org. por L.B. Nascimento & P.M.E. Oliveira), pp. 101-
116. Belo Horizonte: Sociedade Brasileira de Herpetologia.
Prado, C.P.A., Uetanabaro, M. & Haddad, C.F.B. 2005. Breeding activity patterns,
reproductive modes, and habitat use by anurans (Amphibia) in a seasonal environment
in the Pantanal, Brasil. Amphibia-Reptilia, 26, 211-221.
Prado, G.M. & Pombal Jr. J.P. 2005. Distribuição espacial e temporal dos anuros em
um brejo da Reserva Biológica de Duas Bocas, sudeste do Brasil. Arquivos do Museu
Nacional, 63 (4), 685-705.
Prance, G.T. 1996. Islands in Amazonia. Philosophical Transactions of the Royal
Society of London, 351, 823-833.
Rabelo, B.V., Pinto, A.C.; Simas, A.P.S.C., Tardin, A.T., Fernandes, A.V., Souza,
C.B., Monteiro, E.M.P.B., Facundes, F.S., Ávila, J.E.S., Souza, J.S.A., Costa-
Guedes, L.A., Penha, O.A.A., Melo, R.M.S. & Gibson, V.M. 2008.
Macrodiagnóstico do Estado do Amapá: primeira aproximação do ZEE. Macapá:
Instituto de Pesquisas Científicas e Tecnológicas do Estado do Amapá (IEPA).
Ratter, J.A., Ribeiro, J.F. & Bridgewater, S. 1997. The Brazilian cerrado vegetation
and threats to its biodiversity. Annals of Botany, 80 (3), 223-230.
Ratter, J.A, Bridgewater, S. & Ribeiro, J.F. 2006. Biodiversity patterns of the woody
vegetation of the Brazilian Cerrado. In: Neotropical savannas and dry forests: plant
diversity, biogeography, and conservation (Org. por R.T. Pennington., G.P. Lewis &
J.A. Ratter), pp. 31-66. Oxford: Taylor & Francis Press.
Ribeiro, J.F. & Walter, B.M.T. 1998. Fitofisionomias do Bioma Cerrado. In: Cerrado:
ambiente e flora (Org. por S.M. Sano & S.P. Almeida), pp. 89-166. Brasília:
Embrapa/CPAC.
Ribeiro, J.F.; Walter, B.M.T. 2008. As principais fitofisionomias do Cerrado. In:
Cerrado: ecologia e flora (Org. por S.M. Sano., S.P. Almeida & J.F. Ribeiro, J.F),
pp.153-212. Brasília: Embrapa Cerrados.

134
Rocha, A.E.S., Miranda, I.S. & Costa Neto, S.V. 2014. Composição florística e chave
de identificação das Poaceae ocorrentes nas savanas costeiras amazônicas, Brasil. Acta
Amazônica, 44 (3), 301-314.
Sabagh, L.T. & Carvalho-e-Silva, A.M.P.T. 2008. Feeding overlap in two sympatric
species of Rhinella (Anura: Bufonidae) of the Atlantic Rain Forest. Revista Brasileira
de Zoologia, 25 (2), 247-253.
Saenz, D., Fitzgerald, L.A., Baum, K.A. & Conner, R.N. 2006. Abiotic correlates of
anuran calling phenology: the importance of rain, temperature, and season.
Herpetological Monographs, 20, 64-82.
Sanaiotti, T., Bridgewater, S. & Ratter, J. 1997. A floristic study of the savanna
vegetation of the state of Amapá, Brazil, and suggestions for its conservation. Boletim
do Museu Paraense Emílio Goeldi, 13, 3-29.
Santos, E.M. & Amorim, F.O. 2006. Cuidado parental em Leptodactylus natalensis
(Amphibia, Anura, Leptodactylidae). Iheringia, 96 (4), 491-494.
Salthe, S.N. & Duellman, W.E. 1973. Quantitative constrains associated with
reproductive mode in anurans. In: Evolutionary biology of the anurans: contemporary
research on major problems (Org. por J.L. Vial), pp. 229-249. Columbia: University
of Missouri Press.
Schoener, T.W. 1974. Resource partitioning in ecological communities. Science, 185,
27-38.
Scott Jr., N.J. & Woodward, B.D. 1994. Surveys at breeding. In: Measuring and
Monitoring Biological Diversity - Standard Methods for Amphibians (Org. por W.R.
Heyer., M.A. Donnelly., R.W. Mcdiarmid., L.A.C. Hayek & M.S. Foster), pp. 118-
125. Washington: Smithsonian Institution Press.
Silva, J.M.C. & Bates, J.M. 2002. Biogeographic patterns and conservation in the South
American Cerrado: a tropical savanna hotspot. BioScience, 52: 225-234.
Silva, R.A., Martins, I.A. & Rossa-Feres, D.C. 2008. Bioacústica e sítio de vocalização
em taxocenoses de anuros de área aberta no noroeste paulista. Biota Neotropica, 8 (3),
123-134.
Silva, R.A., Martins, I.A. & Rossa-Feres, D.C. 2011. Environmental heterogeneity:
Anuran diversity in homogeneous environments. Zoologia, 28 (5), 610-618.

135
Silva, F.R., Almeida-Neto, M., Prado, V.H.M., Haddad, C.F.B. & Rossa-Feres, D.C.
2012. Humidity levels drive reproductive modes and phylogenetic diversity of
amphibians in the Brazilian Atlantic Forest. Journal of Biogeography, 39 (9), 1720-
1732.
Souza, A. & Eterovick, P. 2011. Environmental factors related to anuran assemblage
composition, richness and distribution at four large rivers under varied impact levels
in southeastern Brazil. River Research and Applications, 27, 1023-1036.
Sugai, J.L.M.M., Terra, J.S. & Ferreira, V.L. 2014. Anurans of a threatened savanna
area in western Brazil. Biota Neotropica, 14 (1), 1-9.
Thomé, M.T.C., Zamudio, K.R., Giovanelli, J.G.R., Haddad, C.F.B., Baldissera,
F.A. & Alexandrino, J. 2010. Phylogeography of endemic toads and post-Pliocene
persistence of the Brazilian Atlantic Forest. Molecular Phylogenetics and Evolution,
55 (3), 1018-1031.
Toft, C.A. 1985. Resource partitioning in amphibians and reptiles. Copeia, 1985, 1-21
Toledo, L.F., Silva, R.R. & Haddad, C.F.B. 2007. Anurans as prey: an exploratory
analysis and size relationships between predators and their prey. Journal of Zoology,
271, 170-177.
Toledo, L.F., Garey, M.V.; Costa, T.R.N., Lourenço-de-Moraes, R., Hartmann,
M.T. & Haddad, C.F.B. 2012. Alternative reproductive modes of Atlantic forest
frogs. Journal of Ethology, 30, 331-336.
Valdujo P.H., Recoder, R.S., Vasconcelos, M.M. & Portella, A.S. 2009. Amphibia,
Anura, São Desidério, western Bahia uplands, northeastern Brazil. Check List, 5 (4),
903-911.
Valdujo, P.H. 2011. Diversidade e distribuição de anfíbios no Cerrado: o papel dos
fatores históricos e dos gradientes ambientais. Tese (Doutorado em Ciências),
Universidade de São Paulo, São Paulo. 125 p.
Valdujo, P.H., Camacho, A., Recoder, R.S., Júnior, M.T., Ghellere, J.M.B., Mott,
T., Nunes, P.M.S., Nogueira, C. & Rodrigues, M.T. 2011. Anfíbios da Estação
Ecológica Serra Geral do Tocantins, região do Jalapão, Estados do Tocantins e
Bahia. Biota Neotropica, 11 (1), 251-262.
Valdujo, P.H., Silvano, D.L., Colli, G. & Martins, M. 2012. Anuran species
composition and distribution patterns in Brazilian Cerrado, a neotropical hotspot.
South American Journal Herpetology, 7 (2), 63-78.

136
Valdujo, P.H., Carnaval, A.C. & Graham, C.H. 2013. Environmental correlates of
anuran beta diversity in the Brazilian Cerrado. Ecography, 36: 708-717.
Vargas-Salinas, F., Quintero-Ángel, A., Osorio-Domínguez, D., Rojas-Morales, J.A.,
Escobar-Lasso, S., Gutiérrez-Cárdenas, P.D.A., Rivera-Correa, M. &
Amézquita, A. 2014. Breeding and parental behaviour in the glass frog Centrolene
savagei (Anura: Centrolenidae). Journal of Natural History, 48 (27-18), 1689-1705.
Vasconcelos, T.S. & Rossa-Feres, D.C. 2008. Habitat heterogeneity and use of physical
and acoustic space in anuran communities in southeastern Brazil. Phyllomedusa, 7,
125-140.
Vasconcelos, T.S., Santos, T.G., Rossa-Feres, D.C. & Haddad, C.F.B. 2009. Influence
of the environmental heterogeneity of breeding ponds on anuran Assemblages from
southeastern Brazil. Canadian Journal of Zoology, 87, 699-707.
Vieira, D.L.M., Aquino, F.G., Brito, M.A., Fenades-Bulhão, C. & Henriques, R.P.B.
2002. Síndromes de dispersão de espécies arbustivo-arbóreas em cerrado sensu stricto
do Brasil Central e savanas amazônicas. Revista Brasileira de Botânica, 25 (2), 215-
220.
Wachlevski, M., Erdtmann, L.K. & Garcia, P.C.A. 2014. Anfíbios anuros em uma área
de Mata Atlântica da Serra do Tabuleiro, Santa Catarina. Biotemas, 27 (2), 97-107.
Webb, C.O., Ackerly, D.D., Mcpeek, M.A. & Donoghue, M.J. 2002. Phylogenies and
community ecology. Annual Review of Ecology and Systematics, 33, 475-505.
Wells, K.D. 1977. The social behavior of anuran amphibians. Animal Behaviour, 25: 666-
693.
Wells, K.D. 1979. Reproductive behavior and male mating success in a neotropical toad,
Bufo typhonius. Biotropica, 11 (4), 301-307.
Wells, K.D. 2007. The ecology and behavior of amphibians. Chicago: University of
Chicago Press.
Werneck, F.P., Costa, G.C., Colli, G.R., Prado, D.E. & Sites Jr., J.W. 2011.
Revisiting the Seasonally Dry Tropical Forests historical distribution: new insights
based on palaeodistribution modelling and palynological evidence. Global Ecology
and Biogeography, 20, 272-288.
Werneck, F.P., Nogueira, C., Colli, G.R., Sites Jr. J.W & Costa, G.C. 2012. Climatic
stability in the Brazilian Cerrado: implications for biogeographical connections of
South American savannas, species richness and conservation in a biodiversity hotspot.
Journal of Biogeography, 39, 1695-1706.

137
Zimmerman, B.L. 1994. Audio Strip Transects. In: Measuring and monitoring
biological diversity: standard methods for amphibians (Org. por W.R. Heyer., M.A.
Donnelly., R.W. McDiarmid., L.A.C. Hayek., M.S. Foster), pp. 92-97. Washington:
Smithsonian Institution Press.





























