Embed Size (px)
Citation preview

1
Universidade Federal de Santa Catarina
Centro de Ciências Biológicas
Departamento de Biologia Celular, Embriologia e Genética
LAMAR – Laboratório de algas marinhas
Influência da radiação UVB no desenvolvimento de plântulas de Gelidium
floridanum (RHODOPHYTA)
Frederico Trevisan Santos
Florianópolis, SC
Outubro, 2009

2
Universidade Federal de Santa Catarina
Centro de Ciências Biológicas
Influência da radiação UV-B no desenvolvimento de plântulas de Gelidium
floridanum (RHODOPHYTA)
Aluno: Frederico Trevisan Santos
Orientadora: Profa. Dra. Zenilda L. Bouzon
Co-orientadora: Lidiane Â. Scariot
Laboratório de Algas Marinhas – LAMAR, CCB, UFSC
Florianópolis, SC
Outubro, 2009
Trabalho de conclusão de curso apresentado
ao curso de Graduação em Biologia, da
Universidade Federal de Santa Catarina.

3
Banca examinadora
Zenila Bouzon
Luciane Ouriques
Paulo Horta

4
Agradecimentos
À Professora Zenilda e a Lidiane pela dedicação dispensada.

5
Lista de Figuras
Fig. 1: Talo tetrasporofítico adulto de Gelidium floridanum.
Fig. 2: Ciclo de vida de G. floridanum.
Fig. 3: Processo Germinativo de G. floridanum, do esporo à plântula.
Fig. 4: Local de coleta dos talos tetrasporofíticos de G.floridanum.
Fig. 5: Sala de cultura onde as amostras foram mantidas para crescimento e exposições à radiação UVB, sob
condições controladas.
Fig. 6: Plântulas no oitavo dia de desenvolvimento e primeiro dia de exposição à UVB (“dia zero”).
Fig. 7: Crescimento em comprimento de plântulas controle e tratadas com UVB (µm).
Fig. 8: Evolução do ângulo de curvatura da plântulas de G. floridanum tratadas com UVB.
Fig. 9: Crescimento em comprimento ao longo de 23 dias de crescimento de plântulas de G. floridanum
controle e tratadas com UVB.
Fig. 10: Crescimento em largura ao longo de 23 dias de crescimento de plântulas de G. floridanum
tratadas com UVB.
Fig 11. Plântulas de G. floridanum com 13 dias de desenvolvimento, coradas com AT-O.
Fig. 12. Plântulas de G. floridanum com 13 dias, coradas com Coomassie brilliant blue (CBB).
Fig. 13. Plântulas de G. floridanum com 13 dias, coradas com PAS.
Fig. 14. Plântulas de G. floridanum com 18 dias, coradas com AT-O.
Fig. 15. Plântulas de G. floridanum com 18 dias, coradas com CBB.
Fig. 16. Plântulas de G. floridanum com 18 dias, coradas com PAS.
Fig. 17. Plântulas de G. floridanum com 23 dias, coradas com AT-O.
Fig. 18. Plântulas de G. floridanum com 23 dias, coradas com CBB.
Fig. 19. Plântulas de G. floridanum com 23 dias, coradas com PAS.

6
Sumário
Resumo 7
Introdução 8
Objetivos 13
Objetivo geral 13
Objetivo específico 13
Materiais e métodos 14
Local de coleta 14
Processamento de amostras 17
Microscopia óptica 17
Histoquímica 17
Resultados 19
Morfologia externa 19
Crescimento 22
Taxa de crescimento 23
Observações histoquímicas 25
Discussão 40
Bibliografia 45

7
Resumo
A incidência da radiação ultravioleta sobre a superfície terrestre vem
aumentando devido à redução da espessura da camada de ozônio, especialmente no
hemisfério sul. Este aumento, principalmente da radiação UVB, atinge negativamente
os diferentes organismos aquáticos, entre eles as macroalgas vermelhas presentes nas
regiões entre-marés. São organismos de grande importância ecológica e econômica.
Dentre estas, se destaca o gênero Gelidium, cuja importância econômica reside na
extração do ágar de qualidade. No presente trabalho, plântulas de G. floridanum foram
expostas à radiação UVB, sob condições de laboratório, ao longo de 15 dias. Os
resultados mostraram uma nítida diminuição na taxa de crescimento das plântulas
expostas quando comparadas com as plântulas controle. As plântulas expostas tiveram
um crescimento médio de 0,729mm ao longo do período analisado, ao passo que as
plântulas controle cresceram até 1,004mm em média. A largura, por outro lado,
apresentou diferenças mais sutis: 0,072mm para as plantas tratadas com radiação UVB
e, 0,078mm para as controle. Plântulas expostas apresentaram curvatura do talo de 59°
no quinto dia de irradiação, 55.5° no décimo dia e 42,5° no décimo quinto dia. Outro
efeito observado foi a emissão anômala de rizóides laterais, próximos ao ápice das
plântulas expostas à radiação. Os testes histoquímicos revelaram um nítido
espessamento na parede celular nas plântulas tratadas evidenciado por uma forte reação
metacromática nesta região. Também foi possível observar um padrão diferente de
distribuição dos grãos de amido. As células localizadas na região convexa do talo
curvado mostraram uma maior concentração de grãos de amido. Quando coradas com
CBB, as células corticais das plântulas tratadas apresentaram conteúdo heterogêneo e as
medulares, por sua vez, apresentaram diâmetros irregulares, ao contrário das plântulas
controle. Estes dados comprovam que a radiação UVB interfere no crescimento, na
morfologia e na fisiologia de plântulas de G. floridanum.

8
Introdução
As algas são dominantes em habitats aquáticos, onde podem ser os contribuintes mais
importantes para a produtividade destes ecossistemas. Ao longo dos costões rochosos podem
ser encontradas dispostas em faixas ou camadas bem distintas (Raven, 2001). Elas podem
crescer desde uns poucos milímetros até mais de 60 metros formando as chamadas “kelps” que
são florestas submersas. São importantes também no que se refere à fixação de carbono da
atmosfera, interagem com vários organismos aquáticos desempenhando papel de grande
importância, como provedoras de alimento e abrigo. Desta forma, proporcionam um cenário
adequado para o recrutamento e a dispersão dos animais no ecossistema (Bischof et al, 2006).
Dentre os grupos de algas do costão rochoso, as algas vermelhas diferenciam dos
demais organismos fotossintetizantes por duas características marcantes: a presença de clorofila
“a” e “d” e ficobilinas em seus cloroplastos e também a ausência de flagelos (Oliveira, 2003).
Existem mais de 6000 espécies, sendo que a grande maioria vive no ambiente marinho. São
bentônicas e macroscópicas quase que em sua totalidade. As ficobilinas e alguns pigmentos
acessórios mascaram a cor da clorofila “a” e dão às algas vermelhas sua coloração
característica (Oliveira, 2003).
Estas algas têm uma ampla importância como recurso alimentar marinho rico em
vitaminas, sais minerais, proteínas com baixo teor de gordura e, por isso, com grande valor
comercial. Da mesma forma, a produção de ficocolóides, as agaranas e as carregenanas,

9
apresenta inúmeras aplicações industriais insubstituíveis. A maioria destas aplicações, quase
90%, é na indústria alimentícia como emulsificantes, estabilizantes, gelificantes, para a
clarificação de bebidas, como elementos suspensores, entre outros usos. São também utilizadas
na indústria cosmética, de medicamentos, biotecnologia e outros (Armisén & Galatas, 1987;
Oliveira, 2003).
O ágar é um colóide hidrofílico produzido por algumas espécies de Rhodophyta. Dentre
os gêneros produtores podemos destacar os gêneros Pterocladiella, Gracilaria e Gelidium.
Originalmente o ágar de Gelidium é considerado o ágar genuíno. Embora existam outras fontes
de ágar, estes não possuem as mesmas propriedades do ágar de Gelidium, porém podem ser
usados como substitutos em certas condições (Armisén & Gálatas, 1987).
A espécie Gelidium floridanum Taylor, W.R. (Fig. 1), apresenta o ciclo de vida trifásico
assim como as demais ordens da classe Florideophyceae, constituído por uma fase gametofítica
haplóide (gametófito) e duas fases esporofíticas diplóides: o carposporófito e o tetrasporófito
(Ouriques, 2002).
Fig. 1: Talo tetrasporofítico adulto de Gelidium floridanum. Seta: localização dos ramos férteis no talo. Cabeças
de seta: “estiquídios”, ramos laterais modificados, com tetrasporângios. (Foto: Lidiane Scariot)
O gametófito masculino libera o espermácio que fica a deriva no ambiente encontrando-
se ao acaso com a tricogine do gametófito feminino. Neste momento há o reconhecimento do
espermácio ocorrendo a fecundação e desenvolvimento do carposporófito. O carposporófito

10
libera carpósporos diplóides, os quais se desenvolvem formando o tetrasporófito. Nos ramos
férteis, os tetrasporângios sofrem meiose espórica produzindo tetrásporos (Fig. 2).
Fig. 2: Ciclo de vida de G. floridanum. (Esquema: Lidiane Scariot)
Os esporos das algas vermelhas são liberados com uma mucilagem transparente que é
responsável pela adesão destes ao substrato (Avanzini, 1989). No caso de Pterocladiella
capillacea cerca de duas horas depois da liberação, os esporos já estão firmemente fixados no
substrato (Ouriques, 2002). Logo em seguida, uma parede celular é produzida e a germinação
tem início (Ouriques, 2002; Bouzon et al. 2006).
Em G. floridanum, o esporo aderido ao substrato emite rapidamente uma
protuberância lateral, denominada tubo germinativo, para a qual é transferido todo o
conteúdo protoplasmático do esporo. Em seguida, forma-se um septo isolando o tubo
germinativo do esporo inicial (Fig. 3A e 3B). No lado oposto à fixação do esporo
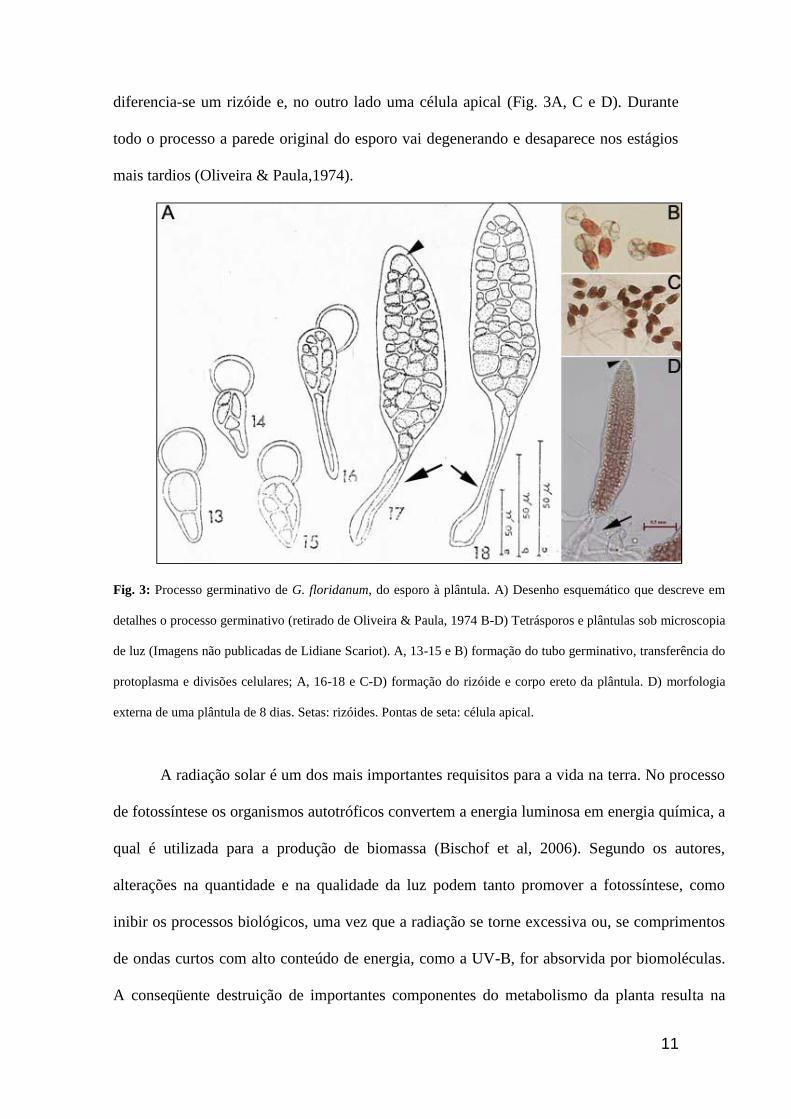
11
diferencia-se um rizóide e, no outro lado uma célula apical (Fig. 3A, C e D). Durante
todo o processo a parede original do esporo vai degenerando e desaparece nos estágios
mais tardios (Oliveira & Paula,1974).
Fig. 3: Processo germinativo de G. floridanum, do esporo à plântula. A) Desenho esquemático que descreve em
detalhes o processo germinativo (retirado de Oliveira & Paula, 1974 B-D) Tetrásporos e plântulas sob microscopia
de luz (Imagens não publicadas de Lidiane Scariot). A, 13-15 e B) formação do tubo germinativo, transferência do
protoplasma e divisões celulares; A, 16-18 e C-D) formação do rizóide e corpo ereto da plântula. D) morfologia
externa de uma plântula de 8 dias. Setas: rizóides. Pontas de seta: célula apical.
A radiação solar é um dos mais importantes requisitos para a vida na terra. No processo
de fotossíntese os organismos autotróficos convertem a energia luminosa em energia química, a
qual é utilizada para a produção de biomassa (Bischof et al, 2006). Segundo os autores,
alterações na quantidade e na qualidade da luz podem tanto promover a fotossíntese, como
inibir os processos biológicos, uma vez que a radiação se torne excessiva ou, se comprimentos
de ondas curtos com alto conteúdo de energia, como a UV-B, for absorvida por biomoléculas.
A conseqüente destruição de importantes componentes do metabolismo da planta resulta na

12
redução da atividade fotossintética e do metabolismo geral, apresentando decréscimo na
produção de biomassa (Bischof et al, 2006).
A radiação ultra-violeta é uma das frações da luz solar dividida em três faixas: UVA
315-400 nm, UVB 280-315 nm e UVC 190-280 nm (Bischof et al, 2006). Cálculos baseados
nas características de absorção do ozônio indicam que com a redução de 10% da coluna da
camada de ozônio, a irradiação aumenta em aproximadamente 5% para comprimentos de onda
em torno de 320 nm, porém aumenta em 100% para comprimentos em torno de 300 nm
(Bischof et al, 2006).
Uma série de fatores influencia no alcance da radiação UV-B na superfície da Terra:
distância entre a Terra e o sol, níveis de gases atmosféricos e aerossóis, ângulo de Zênite solar,
cobertura de nuvens, altitude, latitude, superfície de albedo, comprimento do dia e outros (Diaz
et al, 2006). A influência da radiação também depende da capacidade desta de penetrar na
coluna d´agua. A radiação UVB (RUVB) capaz de afetar negativamente a produtividade
primária das plantas alcança até 5-6 metros de profundidade (Bischof et al, 2006).
Segundo Diaz et al, 2006, depois que se descobriu a diminuição dos níveis
estratosféricos de ozônio durante a primavera na região Antártica, o estudo da variação dos
níveis de UVB tornou-se um tópico de grande interesse na comunidade científica. A
diminuição da camada de ozônio estratosférica causa um aumento na quantidade de RUVB que
alcança a superfície terrestre e efeitos desta radiação têm sido detectados, embora
primeiramente em estágios multicelulares (Altamirano et al, 2003). A América do sul é uma
região de particular relevância para este tipo de estudo por causa da proximidade com a área
afetada por esta influência. Ao final da primavera massas de ar com baixas quantidades deste
gás passam sobre a região sul da América do Sul diminuindo a proteção que é oferecida pelo
ozônio às diferentes comunidades (Diaz et al, 2006).

13
Os registros referentes à diminuição da camada de ozônio na região Antártica indicam
uma redução de 40% a 50% desse elemento na estação da primavera. Por esta redução estar
restrita àquela região se denominou buraco na camada de ozônio. O íon cloro, quando exposto
a radiação ultravioleta apresenta uma alta reatividade com as moléculas de ozônio causando a
destruição das mesmas. Existem outros fatores que também são apontados como vilões do
ozônio como os óxidos nítricos, por exemplo (Martínez, 2004)
Os efeitos da exposição à radiação ultravioleta nos sistemas biológicos são variados,
indo do nível molecular ao organismo, afetando o crescimento, a produtividade e mesmo a
estrutura e função do ecossistema (Bischof et al, 2006; Hongxia et al, 2007).
Os estudos sobre a influência da radiação UVB em macroalgas estão focados em algas
de interesse comercial e nas alterações que essa radiação pode provocar na fisiologia destes
organismos. Já se sabe que o crescimento e morfologia do talo são afetados pela radiação
(Navarro et al, 2007; Martinez et al, 2004; Altamirano et al, 2000; Mansila et al, 2006; Schimdt
et al, 2009). Os produtos extraídos de determinadas macroalgas são alvo de análises
quantitativas e qualitativas. Alterações nestes fatores podem significar colapsos econômicos em
comunidades que dependem desta produção. As comunidades de peixes que tem as macroalgas
como fonte de alimento e local para procriação também podem ser afetadas influenciando
diretamente na economia e subsistência das comunidades humanas (Bischof et al, 2006).
O presente experimento teve por finalidade observar a influência da radiação UVB no
desenvolvimento de plântulas de Gelidium floridanum.

14
Objetivos
Objetivo geral
- Avaliar os efeitos da radiação UVB no desenvolvimento de plântulas de G. floridanum.
Objetivos específicos
- Avaliar o crescimento das plântulas, ao longo do tempo, sob influência da RUVB;
- Identificar alterações morfológicas decorrentes da presença da RUVB;
- Analisar alterações na estrutura celular, através de testes histoquímicos, nas plântulas
submetidas a RUVB.
Materiais de métodos
Local de coletada
Praia da Ponta do Sambaqui, Baía Norte da Ilha de Santa Catarina (27°29’18,8”S e
48°32’12,9”W), município de Florianópolis, Ilha de Santa Catarina (Fig. 4).

15
Fig. 4. Local de coleta dos talos tetrasporofíticos de G. floridanum. A) localização da Praia do Sambaqui,
Florianópolis, Santa Catarina. B) rocha mostrando faixa composta por G. floridanum, em período de maré baixa.
C) características do costão rochoso, com rochas independentes e espaçadas.
As coletas foram realizadas durante os períodos de marés baixas, na região entremarés,
com o auxílio de uma espátula. As amostras foram acondicionadas em sacos plásticos e
transportadas para o Laboratório de Algas Marinhas – LAMAR, da Universidade Federal de
Santa Catarina (UFSC).
No laboratório, as plantas mais vigorosas e aparentemente livres de epífitas foram
selecionadas e lavadas em água do mar filtrada e esterilizada, para remoção dos detritos e
organismos contaminantes.
Ramos com tetrásporângios foram selecionados e dispostos sobre lâminas de vidro,
dentro de placas de Petri contendo água do mar esterilizada. Foram utilizadas 4 lâminas por
placa e quatro placas de Petri (duas para controle e duas para o tratamento com RUVB). As
placas foram mantidas no escuro por aproximadamente 12h, no primeiro dia, para estimular a

16
liberação dos tetrásporos. No dia seguinte, os ramos foram retirados e foi verificada a
deposição de esporos sobre as lâminas.
Os esporos foram mantidos em 100 mL de meio composto por água do mar filtrada e
esterilizada utilizando lâmpada UV-C 8W e filtro de 5 e 1 micrômetro, acrescido de meio
nutritivo von Stoch 4%, em salinidade de 35 ups. O período de desenvolvimento tetrásporo-
plântula foi de 8 dias. O meio foi renovado a cada três dias.
Os esporos e plântulas foram mantidos em sala de cultura (Fig. 5A) a temperatura de
24°C (±2°C), sob condições de luminosidade (irradiância fotossinteticamente ativa) de 80
μmol.fótons.m²/s, fornecida por lâmpadas do tipo “luz do dia” (Philips C-5 Super 84 16W/840,
Brasil), dispostas acima das placas e medida através de um medidor de quanta (Li-cor light
meter 250, United States of America, USA), sob fotoperíodo de 12h.
As amostras foram expostas a radiação ultravioleta B (RUVB) artificial fornecida por
lâmpada (Vilber Lourmat VL-6LM, Marne La Vallée, France) (Fig. 5B e C), 2 horas por dia
(13:00 as 15:00 horas), a partir do 8° dia da liberação dos tetrásporos e, ao longo de 15 dias. A
intensidade da radiação UVB foi 310 mW.cm-2
(Radiometer-model IL 1400A - International
Light, Newburyport, MA, USA).

17
Fig. 5. Sala de cultura onde as amostras foram mantidas para crescimento e exposições à radiação UVB, sob
condições controladas. A) espaço onde o experimento foi desenvolvido. As setas indicam o plástico de proteção à
radiação. B) detalhe da lâmpada de RUVB artificial (seta). C) visualização dos espaços destinados ao controle e ao
tratado.
Amostragens foram realizadas a cada 5 dias a partir do 8 dia (considerado dia zero):
no quinto dia (13 dia de desenvolvimento), no décimo dia (18 ) e no décimo quinto dia (23 ).
Parte do material foi fotografado in vivo em microscópio de luz Leica DM500 equipado
com sistema de captura de imagem (LAS EZ versão 1.6). Este foi observado visando à análise
da morfologia externa, a realização das medidas de comprimento do eixo maior da plântula e
da largura da região mediana, assim como da determinação da curvatura do talo da plântula.
As medidas dos ângulos de curvatura foram tomadas a partir da imagem capturada, traçando
duas retas tangentes à parte externa da plântula curvada e, medido o ângulo entre estas retas
com o auxílio de um transferidor. A outra parte foi processada para microscopia de luz e
histoquímica.

18
A taxa de crescimento foi calculada medindo o comprimento e a largura de 30 plântulas
do controle e do tratamento em cada período analisado. As plântulas foram selecionadas
aleatoriamente.
Processamento das amostras
Microscopia ótica
Para microscopia óptica, as amostras foram fixadas em solução de paraformaldeído
2,5% em tampão fosfato 0,1M e mantidas em câmara úmida, overnight, em temperatura de
4 C. O material foi lavado duas vezes com tampão fosfato 0,1M durante 20 minutos cada,
desidratado em série crescente de etanol (30%, 50%, 70%, 90% e 100%), durante 30 minutos
cada. Após as desidratações as amostras foram incluídas em uma mistura de partes iguais de
etanol 100% e historesina pura por 12 horas. Depois disso, foram colocadas em historesina
pura por mais 12 horas e então emblocadas em historesina com polimerizador por 24 horas.
Os blocos de resina foram seccionados em micrótomo Leica RM2135 e preparadas 15
lâminas de cada amostra: 8 dia e dias 5, 10 e 15 do material tratado e controle.
Histoquímica
O material para microscopia óptica recebeu três tipos de tratamento histoquímico: azul
de toluidina (AT-O), azul brilhante de Coomassie (CBB) e reação de ácido periódico de Schiff
(PAS). Três lâminas de cada amostra para cada tipo de tratamento histoquímico. Os diferentes
testes foram posteriormente observados e fotografados utilizando o microscópio e o sistema de
captura de imagem citado acima.

19
Azul de Toluidina - AT-O
Este corante foi usado para identificar polissacarídeos ácidos através da reação de
metacromasia. As lâminas contendo os cortes foram tratadas com solução aquosa de AT-O
0,5% acidificada, com pH 3,0, por 30 segundos, lavadas em água, secas ao ar e montadas com
bálsamo do Canadá.
Azul Brilhante de Coomassie - CBB
O CBB é utilizado para identificar a presença de proteínas totais nas amostras. Para o
procedimento de coloração, as lâminas ficaram imersas em solução (0,4g diluídas em 100 mL
de água destilada) por 45 minutos e, posteriormente lavadas em solução de Clark (ácido acético
e álcool absoluto 1:3) por 5 minutos, secas ao ar e montadas com bálsamo do Canadá.
Ácido Periódico de Schiff - PAS
O PAS é usado para verificar a presença de polissacarídeos neutros. No caso de algas
vermelhas, identifica principalmente o amido das florídeas e, também, celulose. As lâminas
foram imersas em ácido periódico 1% por 20 minutos, lavadas durante 15 minutos em água
corrente, imersas novamente em reativo de Schiff por mais 20 minutos e, finalmente, lavadas
por 15 minutos em água corrente. Posteriormente, são secas ao ar e montadas com bálsamo do
Canadá.

20
Resultados
Morfologia externa
Os experimentos de tratamento com radiação UVB tiverem início quando as plântulas
estavam no 8° dia de desenvolvimento após a liberação e subseqüente germinação. Neste
estágio, as plântulas mediam entre 0,3 e 0,4 mm de comprimento e, 0,04-0,06 mm de largura na
região mediana (Fig. 6 A, B e C). Estas plântulas são caracteristicamente cilíndricas na base e
achatadas em direção ao ápice. O talo é constituído por numerosas células fortemente
pigmentadas. O crescimento destas plântulas se dá por proliferação da célula apical (Fig. 6 D).
São fixadas ao substrato por longos rizóides, hialinos com aproximadamente 1mm de
comprimento e 75µm de largura, podendo ocorrer em número de um ou mais por plântula (Fig.
6 A e E).
Fig. 6. Plântulas de 8 dias (dia zero), no início do experimento. A e B) plântulas com sistema rizoidal
apresentando um rizóide (A) e vários rizóides (B). C) plântulas agrupadas. D) detalhe do ápice. E) detalhe do
rizóide.

21
Ao longo dos 15 dias de experimento, foi possível distinguir duas características em
especial entre as plântulas do controle e aquelas expostas à radiação UVB: a primeira
relacionada à ocorrência de um encurvamento do talo da plântula e, a segunda relacionada ao
surgimento de emissões lateralizadas de rizóides, localizados próximo ao ápice das plântulas,
ambas observadas apenas no grupo de plântulas expostas à radiação. Estas podem ser
facilmente visualizadas na figura 7.
Algumas plântulas cresciam em aglomerados podendo-se considerar que as plântulas
mais externas sofriam incidência direta da radiação enquanto as dispostas internamente ficavam
“protegidas”.

22
Fig. 7: Desenvolvimento do décimo terceiro ao vigésimo terceiro dia das plântulas controle e irradiadas com
RUVB. A-C: características gerais das plântulas do controle. D-F: plântulas irradias ao longo de 5 dias; D-E:
plântulas apresentando nítido encurvamentodo talo, a partir da região mediana do talo, em direção ao ápice; F:
detalhamento dos rizóides. G-I: morfologia geral apresentada pelas plântulas do controle após 10 dias de
experimento. Notar início de emissão de bifurcação de ramos em G; H: grupamento típico de plântulas. J-M:
plântulas expostas por 10 dias evidenciando em J e K o padrão anômalo de emissão lateral de rizóides; em L, o
encurvamento ainda presente do talo, embora em menor grau. É possível verificar a região apical mais estreita
quando comparada às plântulas controle. N-S: morfologia apresentada pelas plântulas controle após 15 dias de
experimento. Q e R: detalhe de ocorrência de bifurcação apical do talo; S: sistema rizoidal longo. T-V: plântulas
após 15 dias de exposição à RUVB. T: o encurvamento do talo diminui, mas ainda verifica-se a presença de
rizóides laterais; U: algumas plântulas apresentam desenvolvimento tardio; V: detalhe dos rizóides, menos
numerosos. As demais algas verdes que aparecem nas imagens demonstram contaminação ocorrida ao longo do
experimento.

23
Crescimento
As plântulas controle, com 13 dias de desenvolvimento atingiram um
comprimento médio de 0,671mm e largura de 0,077mm. Estas plântulas se mantiveram
eretas e apresentaram uma maior proliferação de rizóides em várias direções,
aumentando desta forma a adesão das plântulas ao substrato (Fig 7 A, B e C). Por outro
lado, as plântulas tratadas com 13 dias de desenvolvimento e, após o quinto dia de
exposição à radiação UVB, mostraram uma acentuada curvatura do talo, com ângulo de
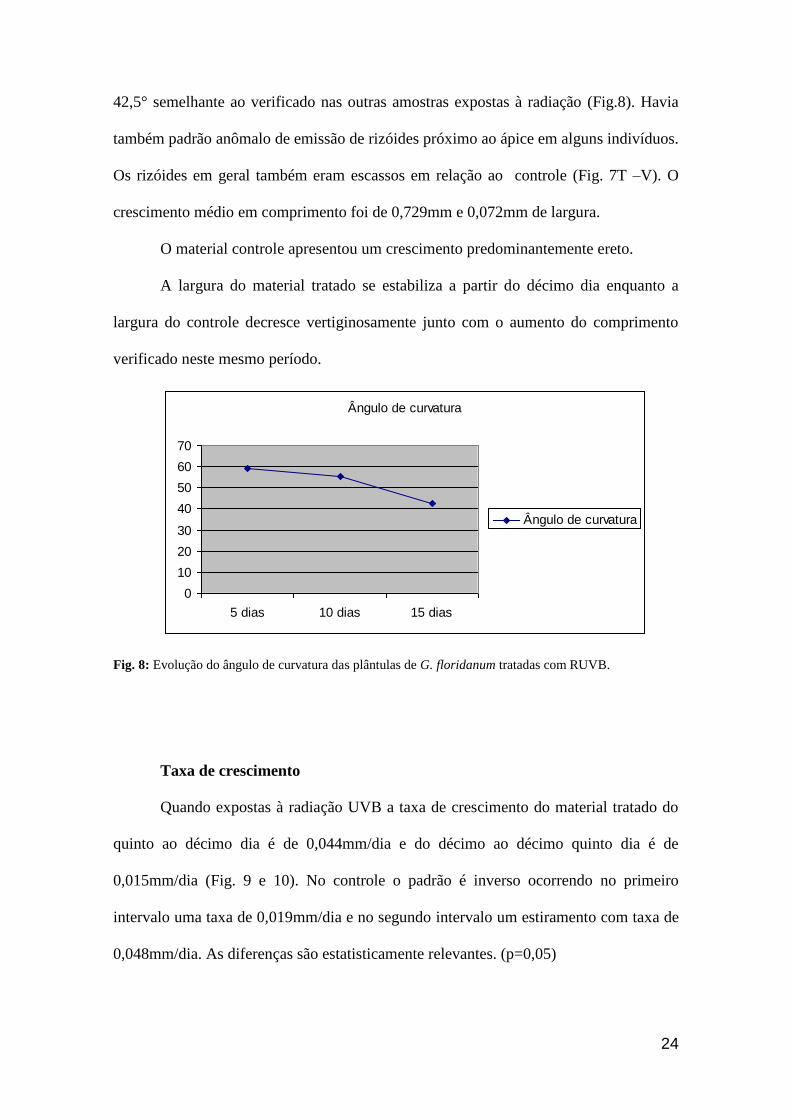
curvatura em média de 59° (Fig. 8). O material tratado apresentou um significativo
aumento na curvatura das plântulas em relação ao controle. Estas plântulas atingiram
um comprimento médio de 0,433mm e largura de 0,059mm e apresentaram um menor
número de rizóides quando comparadas ao controle (Fig. 7D – F).
As plântulas controle no 18° dia de desenvolvimento atingiram um comprimento
médio de 0,766 e largura de 0,091mm. Também apresentam um grande número de
rizóides e mantiveram-se eretas com uma tendência a afinarem próximo a base
mantendo a largura próxima ao ápice. Algumas já apresentavam as primeiras
bifurcações junto à base (Fig. 7G – I). Já as plântulas de 18° dias tratadas com UVB
(por 10 dias) apresentaram um encurvamento forte com ângulo médio de 55,5° e
diminuição no número de rizóides emitidos (Fig. 8). Algumas apresentaram um padrão
anômalo de emissão de rizóides próximo ao ápice (Fig. K – M). Estas plântulas tratadas
tiveram um crescimento médio em comprimento de 0,651mm e largura de 0,070mm.
Já as plântulas controle com 23 dias de desenvolvimento chegaram a um
comprimento de 1,004mm e largura de 0,078mm. Estas plântulas também se mostraram
eretas e com rizóides abundantes emitidos a partir da base. Em algumas delas foram
verificadas bifurcações no ápice demonstrando início de ramificação do talo (Fig. 7N –
S). O material de 23 dias tratado com UVB (por 15 dias) apresentou encurvamento de
24
42,5° semelhante ao verificado nas outras amostras expostas à radiação (Fig.8). Havia
também padrão anômalo de emissão de rizóides próximo ao ápice em alguns indivíduos.
Os rizóides em geral também eram escassos em relação ao controle (Fig. 7T –V). O
crescimento médio em comprimento foi de 0,729mm e 0,072mm de largura.
O material controle apresentou um crescimento predominantemente ereto.
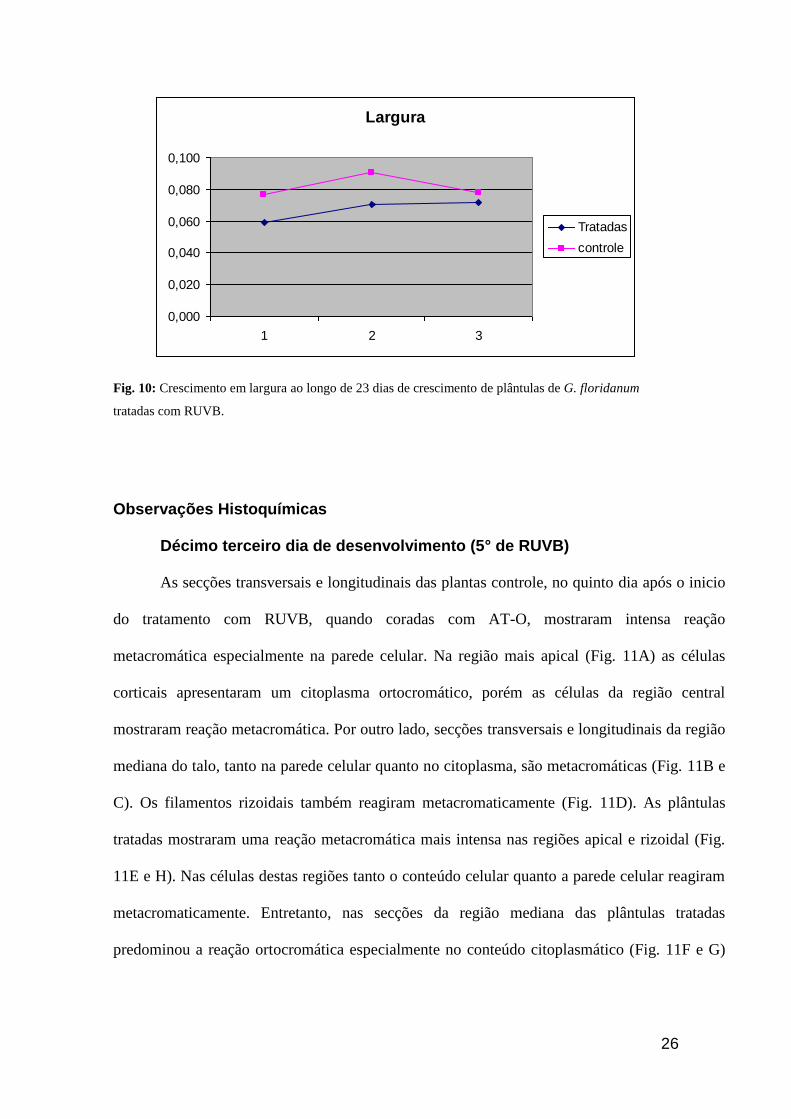
A largura do material tratado se estabiliza a partir do décimo dia enquanto a
largura do controle decresce vertiginosamente junto com o aumento do comprimento
verificado neste mesmo período.
Ângulo de curvatura
0
10
20
30
40
50
60
70
5 dias 10 dias 15 dias
Ângulo de curvatura
Fig. 8: Evolução do ângulo de curvatura das plântulas de G. floridanum tratadas com RUVB.
Taxa de crescimento
Quando expostas à radiação UVB a taxa de crescimento do material tratado do
quinto ao décimo dia é de 0,044mm/dia e do décimo ao décimo quinto dia é de
0,015mm/dia (Fig. 9 e 10). No controle o padrão é inverso ocorrendo no primeiro
intervalo uma taxa de 0,019mm/dia e no segundo intervalo um estiramento com taxa de
0,048mm/dia. As diferenças são estatisticamente relevantes. (p=0,05)
25
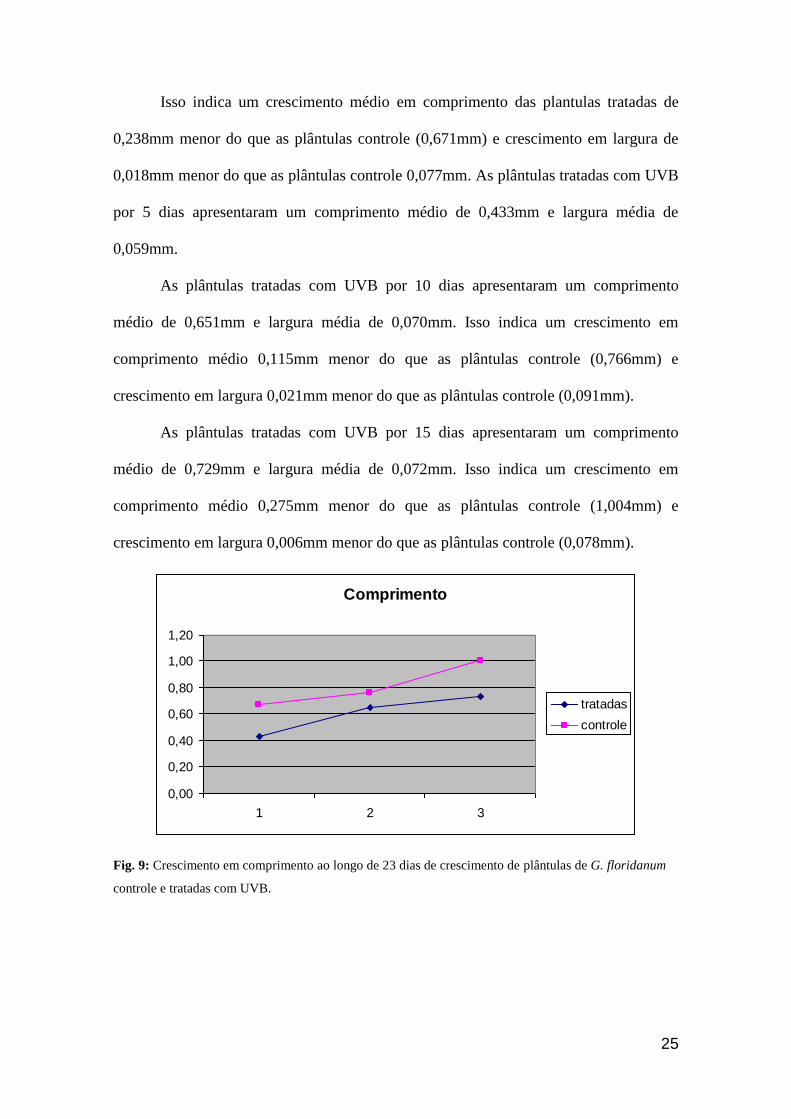
Isso indica um crescimento médio em comprimento das plantulas tratadas de
0,238mm menor do que as plântulas controle (0,671mm) e crescimento em largura de
0,018mm menor do que as plântulas controle 0,077mm. As plântulas tratadas com UVB
por 5 dias apresentaram um comprimento médio de 0,433mm e largura média de
0,059mm.
As plântulas tratadas com UVB por 10 dias apresentaram um comprimento
médio de 0,651mm e largura média de 0,070mm. Isso indica um crescimento em
comprimento médio 0,115mm menor do que as plântulas controle (0,766mm) e
crescimento em largura 0,021mm menor do que as plântulas controle (0,091mm).
As plântulas tratadas com UVB por 15 dias apresentaram um comprimento
médio de 0,729mm e largura média de 0,072mm. Isso indica um crescimento em
comprimento médio 0,275mm menor do que as plântulas controle (1,004mm) e
crescimento em largura 0,006mm menor do que as plântulas controle (0,078mm).
Comprimento
0,00
0,20
0,40
0,60
0,80
1,00
1,20
1 2 3
tratadas
controle
Fig. 9: Crescimento em comprimento ao longo de 23 dias de crescimento de plântulas de G. floridanum
controle e tratadas com UVB.
26
Largura
0,000
0,020
0,040
0,060
0,080
0,100
1 2 3
Tratadas
controle
Fig. 10: Crescimento em largura ao longo de 23 dias de crescimento de plântulas de G. floridanum
tratadas com RUVB.
Observações Histoquímicas
Décimo terceiro dia de desenvolvimento (5° de RUVB)
As secções transversais e longitudinais das plantas controle, no quinto dia após o inicio
do tratamento com RUVB, quando coradas com AT-O, mostraram intensa reação
metacromática especialmente na parede celular. Na região mais apical (Fig. 11A) as células
corticais apresentaram um citoplasma ortocromático, porém as células da região central
mostraram reação metacromática. Por outro lado, secções transversais e longitudinais da região
mediana do talo, tanto na parede celular quanto no citoplasma, são metacromáticas (Fig. 11B e
C). Os filamentos rizoidais também reagiram metacromaticamente (Fig. 11D). As plântulas
tratadas mostraram uma reação metacromática mais intensa nas regiões apical e rizoidal (Fig.
11E e H). Nas células destas regiões tanto o conteúdo celular quanto a parede celular reagiram
metacromaticamente. Entretanto, nas secções da região mediana das plântulas tratadas
predominou a reação ortocromática especialmente no conteúdo citoplasmático (Fig. 11F e G)

27
Nesta região observou-se uma curvatura do talo onde as células da camada inferior ficaram
compactadas.
Quando coradas com CBB as plântulas controles e as tratadas com RUVB mostraram
uma reação positiva no conteúdo celular de todos os tipos celulares observados (Fig. 12A-F).
Porém, nas células do controle a reação foi mais intensa nas células corticais das regiões
apicais, evidenciando maior concentração de material protéico (Fig. 12A e D). Nas secções
transversais na porção intermediária das plântulas controle foi possível observar uma
distribuição uniforme das células da medula. Estas células mostraram conteúdo citoplasmático
levemente granular diferindo das corticais que apresentaram citoplasma homogêneo (Fig. 12B).
Entretanto, em secção transversal da porção mediana das plântulas tratadas foi possível
observar distribuição e diâmetros irregulares das células medulares. As células corticais
apresentaram citoplasma com conteúdo heterogêneo (Fig. 12F). Na porção basal, tanto do
controle quanto do tratado, observou-se que os rizóides originam-se da região medular (Fig.
12C e G). Nas plântulas tratadas, na região rizoidal, inúmeras conexões intercelulares entre
célula-célula e célula-células rizoidais (Fig. 12G). As paredes celulares tanto do tratado quanto
dos controles não reagiram ao CBB.
Quando coradas com PAS as plântulas controle mostraram, em secções longitudinais,
uma distribuição desigual de grãos de amido das florídeas. De forma que nas células das
regiões intermediárias e basais o citoplasma é preenchido por numerosos grãos, entretanto, em
direção à região apical as células mostram uma redução considerável no número de grãos
mostrando um conteúdo citoplasmático levemente positivo ao PAS (Fig. 13A). Nas regiões
mais próximas a base, tanto em corte longitudinal quanto transversal, observou-se que a maior
concentração de grãos de amido ocorreu na camada de células sub-corticais (Fig. 13B e D). No
citoplasma das células rizoidais não foi observado grãos de amido, porém a parede celular
reagiu fortemente ao PAS evidenciando a presença de polissacarídeos neutros como a celulose

28
(Fig. 13C). Este padrão de reação da parede celular foi também observado nas demais células.
Diferindo do observado nas plântulas controle, as tratadas com RUVB e coradas com
PAS, mostraram em secções longitudinais células ricas em grãos amido nas regiões apicais
(Fig. 13E). Em secções transversais, da região mediana das plântulas, foi possível detectar uma
distribuição desigual de células ricas em grãos de amido, onde a maior concentração destes
grãos foi observada nas células localizadas na porção côncava do talo (Fig. 13F). As células
rizoidais apresentaram reação semelhante ao observado nas células do controle (Fig. 13H).
Com a reação de PAS foi possível observar que toda a superfície do talo tanto das
plântulas controle quanto das tratadas com RUVB é revestida por uma camada PAS-positiva
(Fig. 13).
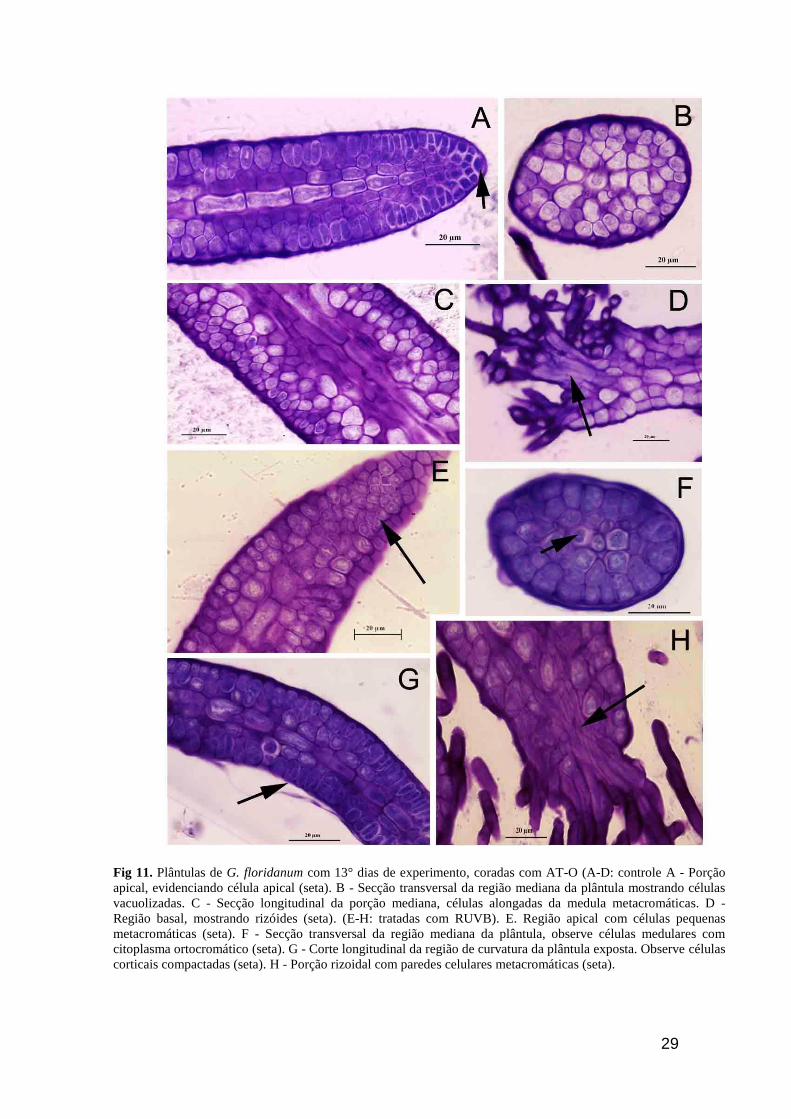
29
Fig 11. Plântulas de G. floridanum com 13° dias de experimento, coradas com AT-O (A-D: controle A - Porção
apical, evidenciando célula apical (seta). B - Secção transversal da região mediana da plântula mostrando células
vacuolizadas. C - Secção longitudinal da porção mediana, células alongadas da medula metacromáticas. D -
Região basal, mostrando rizóides (seta). (E-H: tratadas com RUVB). E. Região apical com células pequenas
metacromáticas (seta). F - Secção transversal da região mediana da plântula, observe células medulares com
citoplasma ortocromático (seta). G - Corte longitudinal da região de curvatura da plântula exposta. Observe células
corticais compactadas (seta). H - Porção rizoidal com paredes celulares metacromáticas (seta).

30
Fig 12. Plântulas de G. floridanum no 13° dia de desenvolvimento e 5° dias de exposição, coradas com coomassie
brilliant blue (CBB) (A-C: controle). A - Secção longitudinal da região apical, onde observa-se intensa reação
positiva. Conexão intercelular na região medular (seta). B - Secção transversal da região medular Células corticais
(seta). C - Porção rizoidal em secção longitudinal, rizóides (setas). (D-G: tratadas com RUVB). D - Secção
longitudinal da região apical mostrando células de tamanhos diferentes, detalhe da célula apical (seta). E - Secção
longitudinal da região mediana mostrando células medulares grandes e regulares (seta). F – Corte transversal
mostrando células corticais positivas e medulares negativas ao CBB. G - Porção rizoidal em secção longitudinal,
com numerosos rizóides (setas) e inúmeras conexões intercelulares (cabeça de seta).

31
Fig. 13. Plântulas de Gelidium floridanum com 13 dias,coradas com PAS. (A-D: controle). A - Secção
longitudinal evidenciando a diminuição da concentração de amido da base para o ápice. B - Região basal em corte
longitudinal. C – Rizóides levemente positivos ao PAS. D - Secção transversal da região basal com grânulos de
amido perifericamente e uniformemente distribuídos. (E-H: tratadas com RUVB). E - Detalhe da célula apical
(seta) sem amido. F - Distribuição desigual dos grânulos na plântula. G - Detalhe da origem medular do rizóide
(seta).

32
Décimo oitavo dia de desenvolvimento (10° de RUVB)
As plântulas controle de 18 dias (10 dias de exposição à RUVB) apresentaram reação
metacromática na parede e levemente no citoplasma quando coradas com AT-O, mostrando a
presença de polissacarídeos neutros. Porém, em secção longitudinal, as células corticais, mais
distais, da região apical apresentaram citoplasma ortocromático, bem homogêneo (Fig. 14A).
Nesta secção foi possível observar que as células medulares são cilíndricas e que reagiram
metacromaticamente ao AT-O. Em secção transversal pode-se observar uma reação
metacromática intensa da parede celular (Fig. 14C). Os rizóides das plântulas controle também
são ortocromáticos no citoplasma e mostraram paredes metacromática ao AT-O (Fig. 14D).
Nas plântulas tratadas com RUVB por 10 dias, as células da região apical também
apresentaram reação levemente ortocromática no citoplasma quando coradas com AT-O (Fig.
14E). Entretanto, as células da região mediana em secção longitudinal revelam um citoplasma
metacromático vacuolizado (Fig. 14F). Em secção transversal, desta mesma região, as células
medulares reagiram metacromaticamente e mostraram uma disposição irregular no talo (Fig.
14H). Os rizóides mostraram uma reação semelhante ao observado no controle (Fig. 14G).
As plântulas controle de 18 dias coradas com CBB, seguindo o padrão do 13 dia,
mostraram forte reação citoplasmática, mais uniforme nas células corticais e, um pouco
heterogêneo nas células medulares (Fig. 15A-D). As células das plântulas tratadas também
seguiram o padrão observado no 13° dia onde foi possível observar distribuição e diâmetros
irregulares das células medulares e células corticais com citoplasma com conteúdo heterogêneo
(Fig. 15E-H) As paredes celulares tanto do tratado quanto dos controles não reagiram ao CBB.
As plântulas controle tratadas com PAS revelam uma concentração menor de grãos de
amido da região basal para a região apical, quando comparadas ao observado no 13° dia de
desenvolvimento das plântulas controle (Fig. 16A e B). Em secção transversal pode-se

33
verificar, nas células da região mediana maior quantidade de grãos de amido na região cortical
(Fig. 16C), porém, nesta mesma figura, células mais apicais apresentaram o citoplasma PAS-
positivo sem grãos de amido. Os rizóides reagem positivamente ao PAS na parede celular (Fig.
16D). As células da região apical das plântulas tratadas com RUVB mostraram o mesmo
padrão de redução na concentração de grãos de amido observado nas células do controle (Fig.
16E). As plântulas tratadas revelaram uma concentração de células ricas em grãos de amido
lateralizados, onde o fotossintetizado se concentra na região côncava do encurvamento da
plântula (Fig. 16F e H).
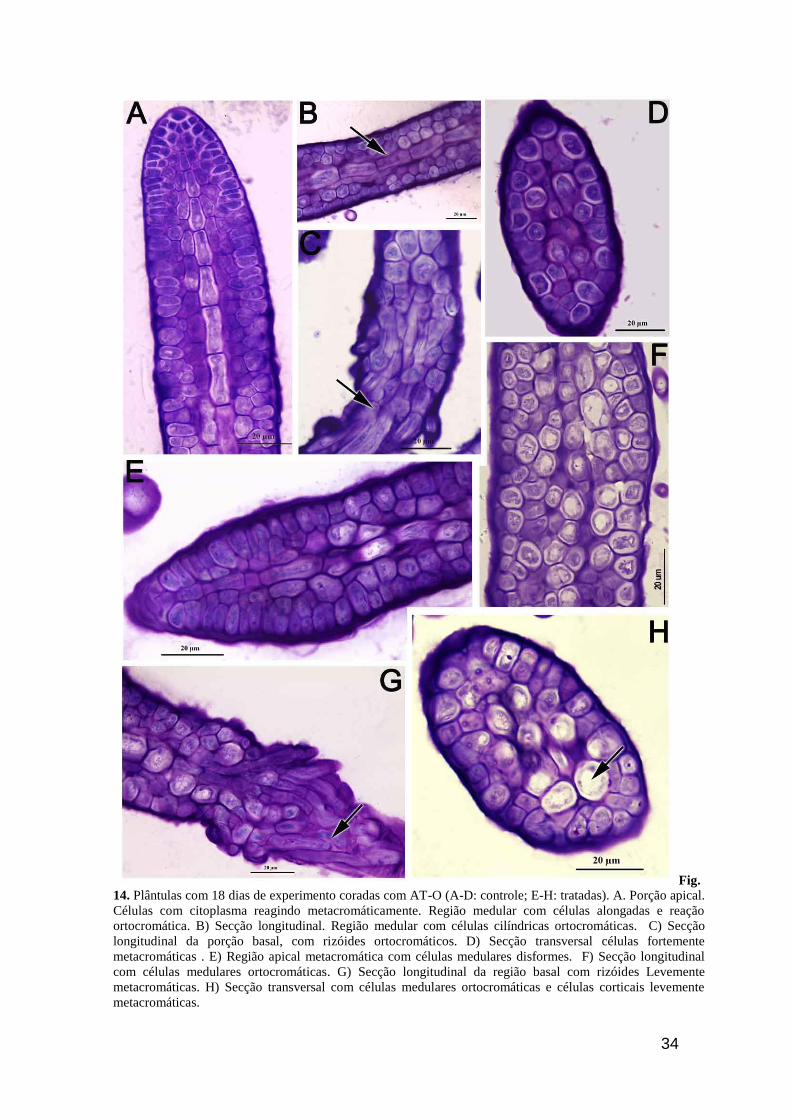
34
Fig.
14. Plântulas com 18 dias de experimento coradas com AT-O (A-D: controle; E-H: tratadas). A. Porção apical.
Células com citoplasma reagindo metacromáticamente. Região medular com células alongadas e reação
ortocromática. B) Secção longitudinal. Região medular com células cilíndricas ortocromáticas. C) Secção
longitudinal da porção basal, com rizóides ortocromáticos. D) Secção transversal células fortemente
metacromáticas . E) Região apical metacromática com células medulares disformes. F) Secção longitudinal
com células medulares ortocromáticas. G) Secção longitudinal da região basal com rizóides Levemente
metacromáticas. H) Secção transversal com células medulares ortocromáticas e células corticais levemente
metacromáticas.
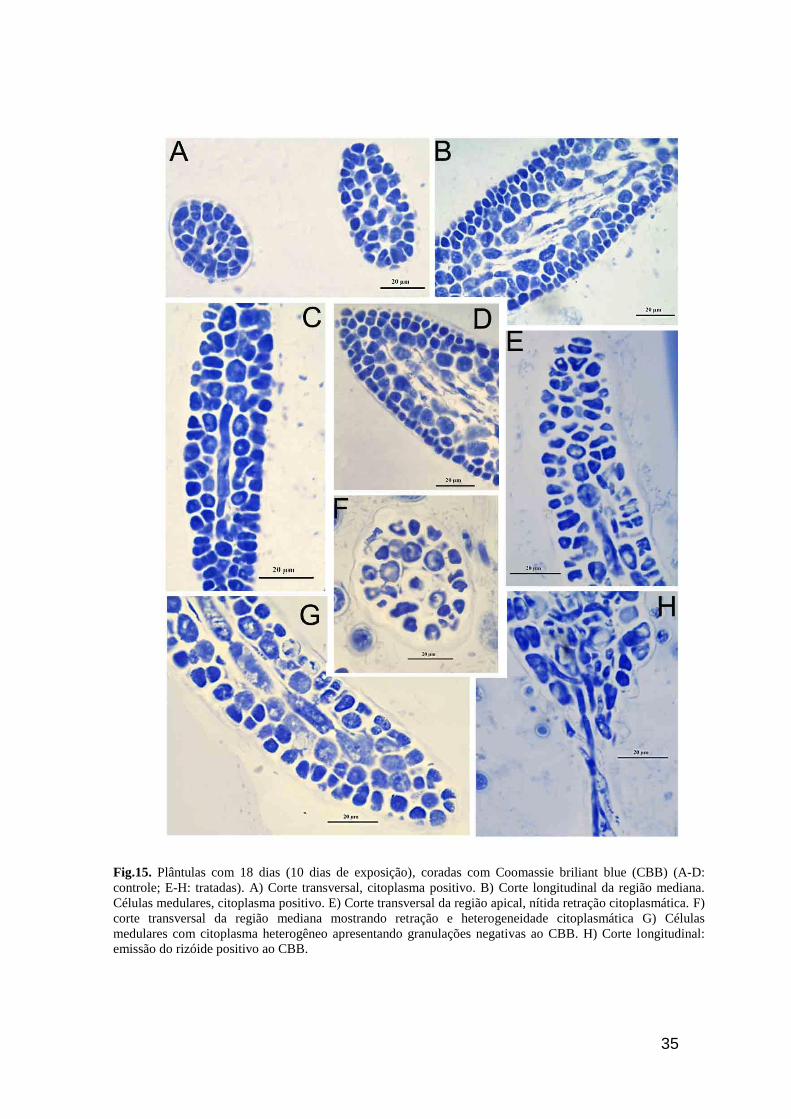
35
Fig.15. Plântulas com 18 dias (10 dias de exposição), coradas com Coomassie briliant blue (CBB) (A-D:
controle; E-H: tratadas). A) Corte transversal, citoplasma positivo. B) Corte longitudinal da região mediana.
Células medulares, citoplasma positivo. E) Corte transversal da região apical, nítida retração citoplasmática. F)
corte transversal da região mediana mostrando retração e heterogeneidade citoplasmática G) Células
medulares com citoplasma heterogêneo apresentando granulações negativas ao CBB. H) Corte longitudinal:
emissão do rizóide positivo ao CBB.

36
Fig. 16. Plântulas com 18 dias, coradas com PAS (A-D: plântulas controle; E-H: plântulas tratadas). A)
Secção longitudinal da região superior da plântula. B e C) Secção transversal da região apical, alguns grânulos
dispersos na região central. D) Detalhe dos rizóides negativos ao PAS. E) Região apical em corte longitudinal.
F) corte transversal da região mediana mostrando a distribuição desigual de grânulos de amido (setas). G)
corte longitudinal da região mediana. H) Região basal em corte transversal mostrando grânulos de amido nas
células e a ausência destes no rizóide.

37
Vigésimo terceiro dia de desenvolvimento (15° dia de RUVB)
As plântulas controle de 23 dias quando coradas com AT-O apresentaram reação
fortemente metacromática na parede celular e levemente no citoplasma dos diferentes tipos
celulares (Fig. 17A). Esta reação também foi observada na região intermediária da plântula
onde as paredes celulares e os citoplasmas reagiram metacromaticamente (Fig. 17B e C). Os
rizóides também têm comportamento metacromático em relação ao AT-O. As plântulas
tratadas com RUVB mostraram a parede celular metacromática, embora o citoplasma se mostre
levemente ortocromático (Fig. 17E e F). Os rizóides reagiram de forma semelhante ao
observado nas plântulas controle (Fig. 17E)
Em relação às plântulas coradas com CBB foi possível perceber que as células da região
apical mostraram o conteúdo citoplasmático positivo revelando uma grande concentração
protéica (Fig. 18A e C). Estas células apresentaram justaposição das células, organização
semelhante ao demonstrado nas análises anteriores. Na região mediana, as células são menos
compactadas, onde foi possível perceber várias conexões intercelulares. Assim como observado
no 13° e 18°, as células medulares são longas e apresentam o citoplasma com conteúdo
heterogêneo devido a presença de vacúolos (Fig. 18B e D). Nas plântulas tratadas com RUVB
foram observadas irregularidades na distribuição e formato das células da região apical (Fig.
18F). Estas células apresentaram-se mais separadas umas das outras do que o observado no
controle e com citoplasma com diferentes graus de concentração de grânulos de amido. Em
secção transversal das células da região mediana, verificou-se uma distribuição desigual das
células e com citoplasma fortemente positivo ao CBB, onde a concentração protéica foi maior
em um dos lados, mostrando uma atividade diferenciada (Fig. 18G e H). Os rizóides
apresentaram citoplasma corado pelo CBB tanto nas plântulas tratadas como controle (Fig. 18E
e I).

38
Quando coradas através da reação de PAS, as plântulas controle de 23 dias mostraram
o mesmo padrão observado nos estágios anteriores, onde a região apical não apresentou grãos
de amido nas células (Fig. 19 A). Entretanto, em poucas plântulas foi possível observar na
região apical células com conteúdo fortemente positivo ao PAS (Fig. 19 B). Em secções
longitudinais e transversais da região mediana das plântulas (Fig. 10C e D) verificou-se
distribuição simétrica de células ricas em dos grãos de amido. Os rizóides são PAS positivos
somente na parede celular (Fig. 19E). Nas células da região apical das plântulas tratadas
observou-se um padrão distinto do encontrado nos estágios anteriores, estas células não
apresentaram grãos de amido (Fig. 19F). Porém, na região mediana das plaântulas, onde
ocorreu o encurvamento, a distribuição de células ricas em grãos de amido foi assimétrica com
a maior concentração de grãos na parte côncava do encurvamento (Fig. 19G e H). Os rizóides
das plântulas tratadas reagiram de forma semelhante aos observados nas plântulas controle
(Fig. I). As paredes celulares de todos os tipos de células reagiram positivamente ao PAS
evidenciando a presença de polissacarídeos neutros.
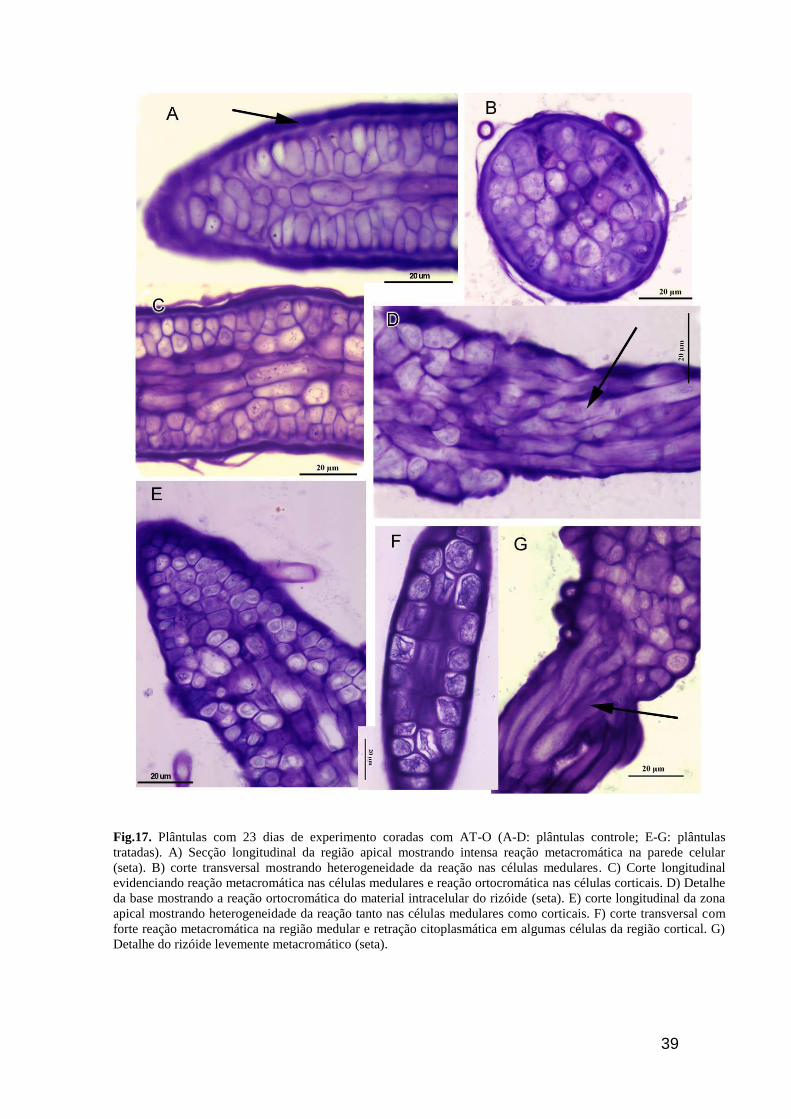
39
Fig.17. Plântulas com 23 dias de experimento coradas com AT-O (A-D: plântulas controle; E-G: plântulas
tratadas). A) Secção longitudinal da região apical mostrando intensa reação metacromática na parede celular
(seta). B) corte transversal mostrando heterogeneidade da reação nas células medulares. C) Corte longitudinal
evidenciando reação metacromática nas células medulares e reação ortocromática nas células corticais. D) Detalhe
da base mostrando a reação ortocromática do material intracelular do rizóide (seta). E) corte longitudinal da zona
apical mostrando heterogeneidade da reação tanto nas células medulares como corticais. F) corte transversal com
forte reação metacromática na região medular e retração citoplasmática em algumas células da região cortical. G)
Detalhe do rizóide levemente metacromático (seta).
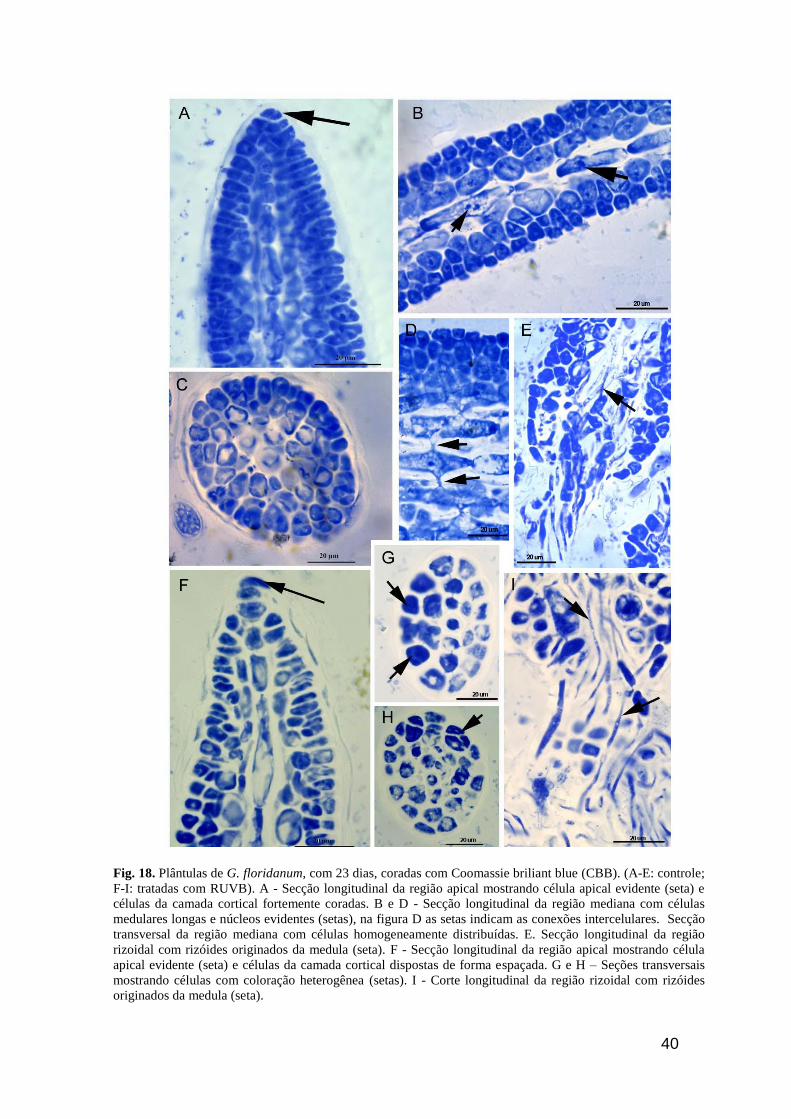
40
Fig. 18. Plântulas de G. floridanum, com 23 dias, coradas com Coomassie briliant blue (CBB). (A-E: controle;
F-I: tratadas com RUVB). A - Secção longitudinal da região apical mostrando célula apical evidente (seta) e
células da camada cortical fortemente coradas. B e D - Secção longitudinal da região mediana com células
medulares longas e núcleos evidentes (setas), na figura D as setas indicam as conexões intercelulares. Secção
transversal da região mediana com células homogeneamente distribuídas. E. Secção longitudinal da região
rizoidal com rizóides originados da medula (seta). F - Secção longitudinal da região apical mostrando célula
apical evidente (seta) e células da camada cortical dispostas de forma espaçada. G e H – Seções transversais
mostrando células com coloração heterogênea (setas). I - Corte longitudinal da região rizoidal com rizóides
originados da medula (seta).
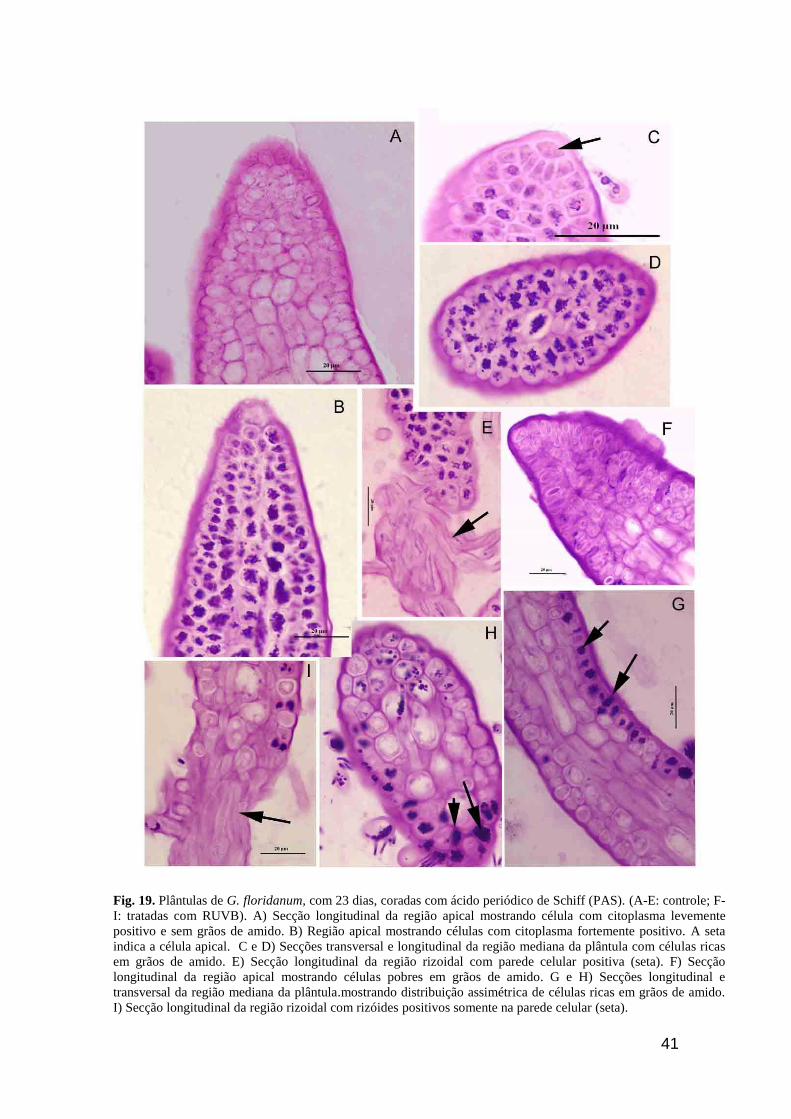
41
Fig. 19. Plântulas de G. floridanum, com 23 dias, coradas com ácido periódico de Schiff (PAS). (A-E: controle; F-
I: tratadas com RUVB). A) Secção longitudinal da região apical mostrando célula com citoplasma levemente
positivo e sem grãos de amido. B) Região apical mostrando células com citoplasma fortemente positivo. A seta
indica a célula apical. C e D) Secções transversal e longitudinal da região mediana da plântula com células ricas
em grãos de amido. E) Secção longitudinal da região rizoidal com parede celular positiva (seta). F) Secção
longitudinal da região apical mostrando células pobres em grãos de amido. G e H) Secções longitudinal e
transversal da região mediana da plântula.mostrando distribuição assimétrica de células ricas em grãos de amido.
I) Secção longitudinal da região rizoidal com rizóides positivos somente na parede celular (seta).

42
Discussão
Os dados avaliados no presente experimento indicaram efeitos negativos
produzidos pela radiação UVB sobre o crescimento e a morfologia de plântulas de G.
floridanum.
De acordo com o levantamento realizado neste trabalho são poucos os registros
da utilização da análise histoquímica para avaliar a influência da radiação ultravioleta B
sobre a estrutura morfológica de algas (Schmidt et al, 2009) e plantas.
Esta diminuição do crescimento já foi observada anteriormente em inúmeros
experimentos com radiação UVB: em tetrasporófitos jovens de Iridea cordata por
Martinez 2004, em Ulva rígida por Altamirano et al 2000, por Mansila et al 2006, por
Altamirano et al 2003. Experimentos anteriores mostram claramente que a radiação
UVB tem efeitos negativos em plântulas das macroalgas Gigartina skottsbergii
(Setchell & Gardner) e Sarcothalia crispata (Bory). Estas alterações no crescimento
podem estar relacionadas com vias de biossíntese, processos de divisão celular, danos
na membrana dos cloroplastos ou dos tilacóides e dano ao DNA das células. As taxas de
crescimento mais baixas podem representar os efeitos nocivos que esta radiação tem em
diversos mecanismos celulares como, por exemplo, enzimas responsáveis pelo
metabolismo do nitrogênio na alga (Mansilla et al, 2006; Altamirano et al, 2003,
Hongxia et al 2007).
A exposição aos comprimentos de onda curtos pode modificar o pareamento
normal das bases de DNA pela formação de lesões entre moléculas de pirimidina
adjacentes, resultando em dímeros conhecidos como fotoprodutos. A formação destes
dímeros de pirimidina vem sendo referida como uma das causas de atraso na divisão

43
celular e conseqüente redução nas taxas de crescimento. A principal conseqüência disso
é a incapacidade das enzimas de RNA e DNA polimerase de reconhecerem estes
dímeros danificados, provocando a interrupção na transcrição e replicação do DNA
(Navarro, 2004; Hongxia et al, 2007; Steinhoff et al, 2008). Esta pode ser uma causa da
diminuição do crescimento em comprimento e largura encontrado nas plântulas de
Gelidium floridanum neste experimento.
Relatos sobre as alterações morfológicas produzidas pela radiação UVB em
espécies de macroalgas são comuns na literatura. Os efeitos morfológicos resultantes
dessa radiação têm sido reportados para plantas superiores. Neste último caso,
receptores específicos da UVB estariam sendo ativados produzindo uma sutil alteração
nas características morfológicas, tais como tamanho e número de folhas, grau de
branqueamento e a elongação do caule. (Navarro, 2004). O encurvamento do talo das
plântulas de Gelidium floridanum, pode ser percebido já no quinto dia de exposição à
UVB o que também acontece com tetrasporófitos de Iridea cordata. Segundo o mesmo
autor, estas alterações morfológicas podem ser interpretadas como uma forma de defesa
à radiação.
Algumas plântulas cresciam muito próximas umas das outras, formando um
aglomerado, isto possivelmente protegia as mais internas e deixava expostas as
periféricas. Esta característica de crescerem aglomeradas pode ter influenciado na
diferença de curvatura entre as plântulas tratadas com UVB sendo que as periféricas
sofriam mais influência da radiação.
A emissão de rizóides lateralmente é um padrão encontrado exclusivamente em
plântulas tratadas com UVB provando que esta característica é influenciada pela
radiação. Não há registro na literatura deste tipo de emissão anômala.

44
As plântulas controle têm uma diminuição da concentração de amido em direção
ao ápice. No caso do encurvamento das plântulas tratadas a parte convexa não apresenta
amido ou apresenta redução significativa da concentração de amido em relação a parte
côncava.
É possível relacionar dois efeitos: a curvatura do talo e o acúmulo e
deslocamento dos grãos de amido. O lado côncavo da curvatura da plântula reage
positivamente ao Ácido periódico de Schiff definindo uma grande quantidade de
fotossintetizado, isto não acontece no lado convexo sugerindo que o material foi
esgotado na multiplicação celular. O encurvamento da plântula ocorre por causa do
aumento do número de células em apenas um dos lados. Podemos assim sugerir que a
exposição à radiação UVB induziu as células de um lado a se multiplicarem encurvando
a plântula. A inibição do fotossistema II pela UVB foi verificada em algas do gênero
Gelidium por Gómez et al, 2001. Em Laminaria hyperborea foram identificadas
dramáticas mudanças nos cloroplastos de algas expostas a UVB + UVA a 17 °C
(Steinhoff, 2008).
O espessamento da parede celular em macroalgas tratadas com RUVB pode ser
interpretado como uma aclimatação a exposição. Este efeito foi observado em duas
variantes de Kappaphycus alvarezii e também em Audouniella savina (F.S Collins)
Woekerlling expostas a radiação onde o espessamento da parede foi considerado como
uma maneira de proteção dos organismos (Schimdt, 2009; Talarico et al, 2000;
Altamirano et al 2000).
Neste grupo de rodófitas a citocinese é incompleta, ficando a membrana celular
contínua entre as células filhas. O poro formado é fechado pela deposição de natureza
protéica, a rolha da conexão intercelular (Bouzon, 1999). Em algumas células medulares

45
de plântulas tratadas pode-se perceber a continuidade do citoplasma entre duas células
conectadas.
Sendo assim fica claro que a exposição à RUVB causa efeitos diversos sobre a
morfologia, distribuição de amido e crescimento das plântulas de G. floridanum na fase
do ciclo de vida, alvo do presente experimento. Deve-se investir em estudos
complementares que corroborem estas inferências.

46
Referências
Altamirano, M., Flores-Moya, A. & Figueroa, F.L. 2000. Long-term effects of natural
sunlight under various ultraviolet conditions on growth and photosynthesis of intertidal
Ulva rigida (Chlorophyceae) cultivated in situ. Botanica Marina, 43: 119-126
Altamirano, M., Flores-Moya, A; Kuhlenkamp, R; Figueroa, F.L. 2003. Stage-dependet
sensitivity to ultraviolet radiation in zygotes of the Brown alga Fucus serratus. Zygote,
11:101-106.
Altamirano, M., Flores-Moya, A.& Figueroa, F.L. 2003. Effects of UV radiation and
temperature on growth of germlings of three species os Fucus (Phaeophycae). Aquatic
Botany, 75: 9-20.
Armisén, R. & Galatas, F. 1987. Production, properties and uses of agar. In: Production
and utilization of products from commercial seaweeds. D.J. McHugh (Ed.), pp. 1-57.
FAO fishing tech. paper.
Avanzini, A. 1989. La ultra estructura de las esporas de Rhodophyta. Insula, 19: 7-10.
Bischof, K., Gómez, I., Molis, M., Hanelt, D., Karsten, U., Lüder, U., Roleda, M. Y.,
Zacher, K. & Wiencke, C. 2006. Ultraviolet radiation shapes seaweed communities.
Reviews in Environmental Science and Biotechnology, 5: 141-166.
Bouzon, Z.L. 1993. Aspectos histoquímicos e ultra-estruturais da porção vegetativa e
reprodutiva de estiquídios de Hypnea musciformis (Gigartinales – Rhodophyta).
Dissertação apresentada à Universidade Federal do Paraná, Curitiba, PR. 66p.
Bouzon, Z.L. 1999. Histoquímica e Ultra-estrutura da espermatogênese e
carposporogênese de algas gracilarióides (Rhodophyta). Tese apresentada ao Instituto
de Biociências da USP, São Paulo, SP. 116p.

47
Bouzon, Z.L., Ouriques, L.C. & Oliveira, E.C. 2006. Spore adhesion and cell wall
formation in Gelidium Floridanum (Rhodophyta, Gelidiales). Journal of Applied
Phycology, 18: 287-294.
Diaz, S., Camilión, C., Deferrari, G., Fuenzalida, H., Armstrong, R., Booth, C.,
Paladini, A., Cabrera, S., Casiccia, C., Lovengreen, C., Pedroni, J., Rosales, A.,
Zagarese, H. & Vernet, M. 2006. Symposium-in-Print: UV effects on aquatic and
coastal ecosystems. Ozone and UV radiation over Southern South America:
Climatology and Anomalies. Photochemistry and Photobiology, 82: 834-843.
Gómez, I., Figueroa, F.L., Sousa-Pinto, I., Viñegla, B., Pérez-Rodríguez, E., Maestre,
C., Coelho, S., Felga, A., & Pereira, P. 2001. Effects of UV radiation and temperature in
hotosynthesis as mesured by PAM Fluorescence in red alga Gelidium pulchellum
(Turner). Botanica Marina, 44:9-16.
Hongxia, J., Kunshan, G. & Helbing, E.W. 2007 Effects of solar UV radiaton on
germination of conchospores and morphogenesis of sporelings in Porphyra haitanensis.
Marine Biology, 151:1751-1759
Mansilla, A; Werlingar, C; Palacios, M; Navarro, N.P & Cuadra, P. 2006. Effects of
UVB radiation on the initial stages of growth of Gigartina skottsbergii, Sarcothalia
crispata and Mazzaella laminarioides (Gigartinales, Rhodophyta), Journal of Applied
Phycology 18:451-459.
Martínez, N.N., 2004. Efeitos da radiação UVB em Irideae cordata
(Gigartinales,Rhodophyta). Dissertação apresentada ao Instituto de Biociências de São
Paulo, São Paulo, SP.87p.
Navarro, N.P., Mansilla, A. & Palacios, M. 2007. UVB effects on early development
stages of commercially important macroalgae in southern Chile. Journal of Applied
phycology, 20:897-906.
Oliveira, E.C. & Paula, E.J.1974. Estudos sobre a germinação de esporos de Rodofíceas
do litoral brasileiro. Anais de Sociedade de Botânica do Brasil, 1:125-133.

48
Oliveira, E.C. 2003. Introdução a Biologia Vegetal. 2. Ed. São Paulo: Edusp- Editora da
Universidade de São Paulo, 266p.
Ouriques, L.C., 2002. Padrões de germinação e aspectos ultra-estruturais de esporos de
Rhodophyta. Tese apresentada ao Instituto de Biociências da Universidade Estadual
Paulista “Júlio de Mesquita Filho”, Rio Claro, SP. 107pp.
Ouriques. L.C., Bouzon, Z.L. 2003. Ultrastructure of germination of tetraspores of
Hypnea musciformis (Gigartinales, Rhodophyta). Plant Biosystems (Firenze), Bologna-
Italy, v.137, n. 2, p. 193-201, 2003.
Raven, H. P., Evert, R. F. & Eichhorn, S. 2001. Biologia Vegetal. Guanabara Koogan,
Rio de Janeiro, RJ. 906pp.
Schmidt, E.C. 2009. Efeitos da radiação ultravioleta-B sobre a morfofisiologia de
Kappaphycus alvarezii (Gigartinales), variantes pigmentares verde e vermelha.
Dissertação apresentada ao curso de pós-graduação em Biologia Vegetal da
Universidade Federal de Santa Catarina, SC. 101pp.
Schmidt, E.C., Scariot, L.A., Rover, T. & Bouzon, Z.L. 2009. Changes in ultrastructure
and histochemistry of two red macroalgae strains of Kappaphycus alvarezii
(Rhodophyta, Gigartinales), as a consequence of ultraviolet B radiation exposure.
Micron, 40:860-689
Steinhoff, F.S., Wiencke, C., Muller, R. & Bischof, K. 2008. Effects of ultraviolet
radiation and temperature on the ultrastructure of zoospores of the brown macroalga
Laminaria hyperborean. Plant Biology, 10:388-397
Talarico, L & Maranzana, G. 2000. Light and adaptative responses in red macroalgae:
an overview. Journal of Photochemistry and Photobiology, 56: 1-11





























