Embed Size (px)
Citation preview

“ U
DEPARTAMENTO DE INGENIERIA AGROINDUSTRXAL
EVALUACI~N DE ARROZ ENTERO Y RESIDUAL COMO SUSTRATOS
PARA LA PRODUCCIÓN DE ESPORAS DE Beauveria bassiana
Chapingo, México, Febrero de 1999
TESIS PROFESIONAL
Que como requisito parcial para obtener el título de
INGENIERO AGROINDUSTRIAL
PRESENTA:
PATRICIA ELIZABETH BAÑOS GUEVARA


* pí 'PrOfesor-Inwtig tesis,quien me apoyó
como se me
* ,NcProfeJor-Investig
aéCimprescindi61é anáíiris estadirtico.
* Aím. c. ~üaviá83~6w Xmánaéz, porsu disponi6Uuiády t k p o áe-stmadb en Ió revirwn aé esta tesiry sus vabsas aportaciones.

* jl mis compaiiems IE. inwstriqnción: @

P.4G CONTENIDO
ICE GENERAL .................................................................................................................... m I C E DE FIGURAS ............................................................................................ m I C E DE CUADROS .................................. ...................................................................... m I C E DE GRÁFIC As .............................................................................................................
I . RESUMEN ............................................................................................................................. SUMMARY .............................. .........................................................................................
n . INTRODUCCI~N ................................................................................................................
m . JUJIIFICACI~N .................................................................................................................
iV . OBJETIVOS . . 4.1. Objetivo general ........................................................................................................... . . 4.2. Objetivos particulares .................................................................................................
V . REVISIÓN DE LITERATURA 5.1. Broca del grano de café (Hpthenemus hampei, Ferrari) .....................................
5.1.2. importancia económica de la plaga .................................................................... 5.1.3. Distnbución de la broca ....................................................................................... 5.1.3.1. Formas de dispersion ...................................................................................... 5.1.3.2. Distribución en México ......................................................................
5.1.4. Taxonomía y sinoruma ........................................................................................ 5.1.5. Biología de la broca del grano de café ................................................................ 5.1.5.1. Morfología y ciclo biológico .... .................................................................
5 . I . 6. Ecología de la broca del grano de café ................................................................ 5.1.6.1. Habitos ................................................................................................ 5.1.6.2. Daños ...............................................................................................................
5.1.7. Métodos de control de la broca del grano de café .............................................
5.1.1. Origen y antecedentes de la broca del grano de café ..................... .........
. . ..
. .
. .
....................................................................... 5.1.7.2. Control 1 ...............................................................
....................................................................... .......................................................................
............................................................ 5.1.7.5.1. Hongos ...................................................................................................... 5.1.7.5.2. Bacterias ............................................. .................................................. 5.1.7.5.3. Protozoarios .............................................................................................. 5.1.7.5.4.Vinis .......................................................................................................... 5.1.7.5.5. Nemátodos ................................................................................................
5.1.8. Enemigos naturales de la b m del grano de café .............................................
5.2. El hongo entomopatógeno Beauveria bassima (Balsamo) Vuiile min ..................... 5.2.1. Antecedentes históricos y origen ......................................................................... 5.2.2. Taxonomía y sinonutua ........................................................................................ 5.2.2.1. Clasificación taxonómica ............................................................................... 5.2.2.2. Sinonunia ............................................................................................ 5.2.2.3. Clave para identificación ..........................................................................
5.2.3. Morfología .............................................................................................................
5.1.8.1. Uso del hongo entomopatógeno Beauveria .....................................
. .
. .
i iv
vi
1 2
3
4
V
5 5
6 6 6 7 7 8 8 9 9 10 10 11 12 12 12 13 14 15 17 18 18 18 18 19 19 22 22 23 24 25 25 26
.- Patricia Elizabeth Bañm Gumma i

� ICE GENERAL ~
5.2.4. propágulos infectivos de B . bassiana .................................................................. ......................................................... n la mortalidad por Beauveria
bmiana ............................................................................................................ 5.2.5. Mecanismos de acclon ..........................................................................................
5.2.5.1. Modos de infección a insectos ....................................................................... 5.2.5.2. Micosis .............................................................................................................
5.2.5.2.2. Desarrullo de la nuwsis ........................................................................... 5.2.5.3. Período de infección letal ...............................................................................
5.2.4.1. Fases de genninación a wnidiación ................................................ ... 5.2.4.2. Formas de dispersión .................... 5.2.4.3. Influencia de la cantidad de e
..
5.2.5.2.1. Condiciones favorables para el desarrullo de una micosis ..................
5.2.5.3.1. Defensas del hospedero después de la infección .................................. 5.2.5.3.2. Cambios de comportamiento de hospederos infectados letalmente ....
. .
5.2.5.3.3. Desarrollo saprufítico del hongo ............................................................ 5.2.6. Producción de rnetabolitos ......................................... ..............................
5.2.6.1. Producción de e-as .... 5.2.6.2. Produccion de toxinas .................................................................................... 5.2.6.3. Producción de ácidos metabóliws ................................................................ 5.2.6.4. Producción de bactericidas y pigmentos ......................................................
.................................................... 5.2.7.1. Ferhhzantes ..................................................................................................... 5.2.7.2. Herbicidas .............................................................................. 5.2.7.3. insecticidas ...................................................................................................... 5.2.7.4. Fungicidas .......................................................... .......................................... 5.2.7.5. Reguladores de crecimiento y wadyuvantes ................................................
5.2.8. Producción de hongos entomopatógenos ............................................................ 5.2.8.1. Caracterización y selección de cepas ............................................................
5.2.8.1.1. Caracterización de cepas ......................................................................... 5.2.8.1.2. Seleccion de cepas .................................................................................... 5.2.8.1.3. Técnicas de aislamiento ........................................................................... 5.2.8.1.4. Conservación de aislamientos ........................................................... 5.2.8.1.5. Seguridad ..................................................................................................
5.2.8.2.1. Producción en cultivo sólido ................................................................... 5.2.8.2.2. Producción en cultivo liquido ......................................................... 5.2.8.2.3. Producción en cultivo difásiw ................................................................
5.2.9. Producción de B . bassiana en cultivo sólido ............................................................. 5.2.9.1. Proceso de producción .............................................................................................
5.2.9.2. Tipos de sustratos sólidos ............................................................................... 5.2.9.2.1. Uso del almidón corno sustrato para procesos de cultivo sólido ......... 5.2.9.2.2. Estructura química del almidón .............................................................
5.2.9.3. Pretratamiento de sustratos de almidón ....................................................... 5.2.9.4. Condiciones de desamllo .......................................................................
5.2.9.4. I . Temperatura ......................................................................................... 5.2.9.4.2. Humedad relativa ..................................................................................... 5.2.9.4.3. Lununosidad ............................................................................................. 5.2.9.4.4. Agua .......................................................................................................... 5.2.9.4.5. Odgeno ................................................ ................................................. 5.2.9.4.6. pH .............................................................................................................. 5.2.9.4.7. Nutrientes ................................................................................... 5.2.9.4.8. Control de contaminantes .......................................................................
...................................................... ..
5.2.7. Efecto de agroquímiws sobre B . bassiana .. . . . .
. .
..
5.2.8.2. Técnicas de producción de hongos ............................
. .
FÁG . 26 27 27
2 8 29 29 30 30 30 33 33 35 35 36 36 38 39 40 40 41 41 41 4 3 44 44 44 44 45 45 46 48 48 48 4 9 50 52 52 52 53 53 54 54 55 56 57 59 60 61 61 63
. Paíricia Elizabeth Baños Guevara U

. . . 5.2.9.4.9. Viabilidad .................................................................................................. 5.2.9.5. Patrones de crecuniento .................................................................................
5.2.9.5.1. Degradación del almidón por microorganismos .................................. 5.2.9.5.2. Productos metabóliws de la fermentación sólida .................................
. .
5.2.10. Exalamiento d o t o de producción de 8 . bassima en cultivo sólido ............ 5.2.10.1. Formulaciones comerciales de B . bassima .............................................. 5.2.10.1.1. Formulacion y wnservacion ................................................................. 5.2.10.1.2. Dosis recomendadas de algunos productos comerciales ...................
.. ..
PAC . 63 64 65 65 66 69 70 71
Vi . MATERIALES Y IMÉTODOS . . . 6.1. Lugar y tiempo de la investigation ............................................................................ 6.2. Fuentes de financuuniento .......................................................................................... 6.3. Origen y mantenimiento de la cepa del hongo ........................................................
. .
6.4. Selección de sustratos y análisis químico proximal ................................................. 6.5. Pruebas preliminares de acondicionamiento del hongo y del sustrato ..................
6.5.3. Curvas de ajuste de contenido de agua ...............................................................
6.5.1. Perfil de esporulación de 8 . bassima .................................................................. 6.5.2.AjustedepH ................................................................ ..............................
6.5.4. Desarrollo del método colorimétriw para determinación de almidón soluble ....................................................................................................................
6.5.4.1. Preparación de la muestra ............................................................................. 6.5.4.2. Reacción colorimétrica y lectura de datos .................................................... 6.5.4.3.Analisisdedatos ........................................................................................
6.5.5. Afinación del método para determinación de viabilidad .................................. 6.6. Primera fase del experimento .....................................................................................
6.6.1. Diseno experimental .............................................................................................
6.6.3. Cosecha .................................................................................................................. 6.6.4. Medición de las variables de respuesta
6.6.4.1. Conteo de esporas ........................................................................................... 6.7. Segunda fase del experimento ....................................................................................
6.7.1. Selección de los mejores tratamientos ................................................................. 6.7.2.1. Contw de esporas .......................................................................... 6.7.2.2. Determinación de o residual ............................................................. 6.7.2.3. Pruebas de viabilidad ............................................... 6.7.2.4. Cambio de pH .................................................................................................
6.8. Análisis estadístico .......................... ........................................................ 6.8.1. Primera fase ........................................................................................................... 6.8.2. Segunda fase ..........................................................................................................
. . .
. - 6.6.2. Preparación de las unidades experimentales .....................
............................
W . RESULTADOS Y DISCUSIÓN . . . . 7.1. Analisls qumco pmximal .......................................................................................... 7.2. Pruebas preliminares de acondicionamiento del hongo y del sustrato ..................
7.2.1. Perfil de esponilación de 8 . bassima .................................................................. 7.2.2. Ajuste de pH ........................................................................................................... 7.2.3. Curvas de ajuste de contenido de agua ............................................................... 7.2.4. Desarrollo de un método colorimétriw para determinación de almidón
soluble .................................................................................................................... 7.2.5. Método para determinación de viabilidad ..........................................................
7.3.1. Selección de los mejores tratamientos .................................................................
. . . 7.3. Primera fase del experimento .................................
73 73 73 73 74 74 74 74
74 75 75 75 76 76 76 76 77 77 77 78 78 78 78 78 78 79 79 79
81 82 82 83 84
85 89 89 89
... Patricia Elizabeth Baños Guevara u

p.ic . 92 92
7.4.2. Determinación de almidón soluble residual ...................................................... 95 7.4.3. Pruebas de viabilidad ....................................................................................... 98 7.4.4. Cambio de pH .......................... ........................................................................ 99
WI . CONCLUSIONES ............................................................................................................. 100
M . RECOMENDACIONES ....................................................................................................... 101
7.4. Segunda fase del experimento .................................................................................... 7.4.1. Perfil de esporulacion ........................................................................................... ..
. . .
x . BIBLIOG RAFÍA. ................................................................................................................... 102
Xi . ANEXOS ............................................................................................................................. 1.09
ÍNDICE DE FIGURAS NO . TiTULO PAC .
Ciclo biológico general de un hongo entomopatógeno sobre un insecto ................ 31 Ciclo infedivo de Beauveria bassiana sobre la broca del café ................................. 34 Diagrama de bloques de la metodología seguida en la investigación ..................... 80
1 2 3
. Patricia Elizabeth Baiím Guevara iv

ÍNDICE GENFRAL
ÍNDICE DE CUADROS
No. 1 2 3 4 5 6 7 8
9
10
1 1
12
13
14 15 16
17
I8
19
r n L 0 Ventajas y desventajas del control biológico .............................................................. Enemigos naturales de la broca del café (Hs.pofhenemus hmpei, Ferran) ........... Diferencias entre cultivo en sustrato sólido y cultivo líquido sumergido ............... Arreglo del diseño experimental y diseño de tratamientos ...................................... Análisis químico proximal ....................................... .............................................. Perfil de esporulación de B. bassima en cultivo cional.. .................................. Resultados del Ajuste de pH ................................................................................ Datos de ajuste de contenido de agua para a m z entero y residual, antes y después de esterilizar .................................................................................................... Datos del método colorimétrico para determinación de almidón soluble probando entrifugación y filtración a vacio .............................................................. Datos de absorúancia y datos ajustados por regresión lineal de las curvas tipo de glucosa y almidón soluble ....................................................................................... Datos de absorbancia y datos ajustados por regresión lineal para las curvas tipo de arroz entero y residual empleadas en el método colorimétrico para
determinación de almidón soluble ..................................... Influencia del contenido de agua y temperatura en la e
B. bassiana en arroz entero ................................................. influencia del contenido de agua y temperatura en la esporulación de B. bassima en a m z residual ....................................................................................... Datos reales y datos ajustados del perfil de esporulación en arroz entero ............. Datos reales y datos ajustados del perfil de esponilación en arruz residual .......... Datos reales y ajustados de la determinación de almidón soluble residual del perfil de esporulacion en arroz entero ...................................................................... Datos reales y ajustados de la determinación de almidón soluble residual del perfil de esporulación en arroz residual ................................................................... Resultados de las pruebas de viabilidad en arruz entero y residual durante el perfil de esporulación ....... ..................................................................................... Comportamiento del pH al
. .
.,
cio y final del perfil de esponilación ..............
PAC.
16 20 51 76 81 82 83
84
86
87
88
90 92 94
96
98 99
Pafricia Elizabeth Baños Guevarn Y

ÍNDICE GENERAL.
MICE DE GRAFICAS No.
1 2
3 4 5 6 7 8
9
10 1 1 12
13
14
TirVLO PAC. 82
Comportamiento de la esponilación de Beauveria bassima a diferentes valores de pH .............................................................................................................................. 83 Curva de ajuste de contenido de agua en ~ f i o z entero ............................................. 84
8 5 86 87 88
90
Perfil de esporulación de Beauveria bassima en cultivo tradicional ......................
Curva de ajuste de contenido de agua en a m z residual .......................................... Curvas del método colorimétrico para determinación de almidón soluble ............ Comparación de curvas tipo ajustadas, de almidón soluble y glucosa .................... Curvas tipo ajustadas de arxuz entero y residual (método colorimétrico) .............. Influencia de la temperatura y contenido de agua durante la esporulación de Beauveria bassima en a m z entero ............................................................................ Influencia de la temperatura y contenido de agua durante la esporulación de Beauveria bassima en arroz residual ............................................................... Curva de esporulación de Beauveria bassima en a m z entero.. Curva de esporulación de Beauveria bassimaen arroz residual ............................ Curva de ajuste de almidón soluble residual del perfil de esporulación de Beauveria bassima en arroz entero ................................................................ Curva de ajuste de almidón soluble residual del perfil de esporulación de Beauveria bassima en a m z residual .......................................................................... Comportamiento de la viabilidad durante la fase final del perfil de esporulacion de B. bassima ........................................................................................
35
!37
!38 .,
- Patricia -~ Elizabeth Baños Guevara

I. RESUMEN
EXI esta investigación se evaluaron el uso del arroz entero y residual como sustratos para obtener la mayor producción de esporas del hongo entomopatógeno Beauverja bassiana. Eí contenido de agua del medio de cultivo y la temperabra de incubación fueron las variables principales, mientras los perfiles de esporulación, el contenido de almidón residual, la viabilidad y la influencia del pH fueron los indicadores considerados como respuestas para cada tratamiento.
Los indicadores obtenidos de las pruebas preliminares fueron: un análisis químico proximal de ambos sustratos, el perfil de esponilación del hongo en a m z entero, la selección de un pH óptimo para la esponilación y las cullras de ajuste de contenido de agua.
El experimento se llevó a cabo en dos fases:
(1) La primera fase consistió en el diseño experimental. Se aplicó un experimento completamente al azar con arreglo factorial, se realizaron 16 tratamientos y 3 repeticiones; el experimento duró 15 días a días a diferentes temperaturas y contenidos de agua. Se seleccionaron los mejores tratamientos.
(2) La segunda fase, fue el análisis de resultados, del cual se observó que el mejor tratamiento, en caso de arroz entero, fue a 25OC de temperatura de incubación y a 65% de contenido de agua; para arroz residual fue a 2O0C de temperatura de incubación y 70% de contenido de agua. Para ambos sustratos se realizó un perfil de esporulación por ajuste estadístico; se obtuvieron dos curvas de tipo sigmoidal con una producción de: 12.7 x 101oy de 7.3 x 10'0 esporas/g sustrato a los 8 y 10 días para arroz entero y arroz residual, respectivamente. Las muestras de sustrato residual fueron congeladas y cuando se terminó el perfil de esporulación se determinó el contenido de almidón soluble residual, los datos fueron ajustados estadísticamente y proporcioaron una tendencia decreciente.
Otra variable de respuesta fue el porcentaje de viabilidad, éste fue de 75% para a m z entero y 69% para arroz residual después de 15 días de incubación, estos resultados no fueron satisfactorios debido al manejo previo de la cepa de trabajo. El pH final idel cultivo no íuvo variación significativa, de un pH inicial de 6.0 descendió a 5.7 en a m z entero y a 5.5 en arroz residual, esto puede ser explicado como un producto de la actividad metabólica del hongo.
Palabras clave: Hongo entomopatógeno, perfil de esporulación, viabilidad, almidón residual.
- - I Pairicia Elizabeth Baños Guevara

SUMMARY
SUMMARY
in this investigation it was evaluated the use of rice and residual rice as sustrates for the major production of spores of Beauveria bassima spores of the entomopathogeneus fungus. The water content of the culture medioum and the incubation temperature were the principal variables; whereas the esporulation profiles, the residual starch content, the viability and the pH influence where the indicators considered as responses for each treatment.
An proximal chemical analysis of both sustrates, the sporulation profile of the fungus on rice, the selection of an optimum pH for the sporulation and the curves of water content adjust; were indicators obtained form the preliminar tests.
The experiment was carried out in two phases:
(1) The first one consisted of an experimental design. A randomise experiment with a factorial arrangement was applied, 16 treatments and 3 repetitions were made; the experimental lasted for 15 days at different temperatures and moisture contents. The best treatments were selected.
( 2 ) The second one, was the results analysis, of which we observed that the best treatment, in the case of rice, was a 25OC of incubation temperature and a 65% of water content; for residual rice it was 20°C of incubation temperature and 70% of water content. For both sustrates a sporulation profile by a statistic adjust was made; there were obtained two sigmoidal curves with a 12.7 x 10'O and 7.3 x 1 0 ' O spores per gram of sustrate after 8 and 10 days for rice and residual rice, respectively. The residual sustrate samples were frozen and when the profile sporulation determination was finished the residual content of soluble starch was measured, the data were statistically adjusted and provide a decreased tendency.
Another response varible was the viability porcent, it was about a 75% for rice and 69% for residual rice after 15 days of incubation; these results were not satisfactory due the previous handle of the work strain. The final pH of culture had no significative variation, from a 6.0 initial pH it descend to a 5.7 in rice and to 5.5. for residual rice; this can be explained as a product of the metabolical activity of the fungus.
Key words: Entomopathogeneus fungus, sporulation profile, viability, residual starch
7 - Papicia Elizabeth Baños Guevara

INTRODUCCI~N
II. INTRODUCCI~N
El control biológico es el sistema de control de plagas m á s antiguo que existe, debido a que es de forma natural y está asociado con la agricultura; sin embargo, puede definll.se, desde el punto de vista ecológico, como la acción de parásitos, predadores o patógenos que mantiene la población de un insecto plaga en un promedio más bajo que aquella que existiria en su ausencia. Dicho combate se efectúa mediante la utilización deliberada y sistemática de sus enemigos naturales, a través de la liberación y protección de insectos benéficos y el empleo de microorganismos entomopatógenos tales como virus, bacterias, hongos y otros como protowarios y nemátodos.
Actualmente, a nivel mundial, se está tratando de luchar contra la inminente contaminación y residualidad de los productos químicos elaborados por el hombre, empleados en el ataque de los perjudiciales insectos plaga; los estudios más reciente:; nos revelan que el control biológico es la mejor opción para evitar el daño a la fauna benéfica y prevenir que se alcancen niveles de desequilibrio ecológico.
Uno de estos insectos plaga de alto índice infedivo en el ámbito cafetalero es la broca del grano de café (Hypohenemus hampi, Ferrari), el cual es un problema fitosanitario que se encuentra en un acelerado proceso de expansión por las regiones más importantes de América Latina y Oceania donde se ha transformado en la plaga número uno por su creciente repercución económica.
Ejerciendo un control biológico, la broca del café puede ser atacada por el hongo Beauveria bassima (Balsamo) hillemin, el cual se considera que puede jugar un papel importante en la regulación y disminución de poblaciones de la broca bajo las condiciones en que se desarrollan las plantaciones de café.
Diversas cepas de este hongo presentan diferente grado de patogenicidad sobre sus hospederos, por esta radn es posible y necesario llevar a cabo una selección permanente de cepas para afinar su espectro de acción, centrándolo sobre la broca y haciéndolo menos importante sobre insectos benéficos (Morales et al., 1991). Las esporas de este entomopatógeno se han encontrado infectando la broca en forma natural prácticamente en todos los paises en donde ha llegado esta plaga (Bustillos et al., 1991).
La técnica de producción que se ha establecido para el cultivo de este hongo es "in vitro" y se realiza en medios de cultivo o sustratos sólidos que contengan fuentes de carbono, sin embargo, la producción de conidias es pequeña y limitada (Gam, 1995). El sustrato sólido m á s estudiado para el cultivo de Beauveria bassima es el a m z pulido entero, con el que se han obtenido rendimientos aceptables de esporulación.
Por lo anterior, el presente trabajo de investigación evalúa el a m z entero y residual, subproducto de la producción comercial de Beauveria bassima, como susiratos sólido^ potenciales para la producción de esporas del hongo entomopatógeno haciendo interactuar condiciones diferentes de temperatura y contenido de humedad.
P
Patricia Elizabeth Baños Guevara 3

JLJSI'IFICACI~N
III. JUSTIFICACI~N
El café es el cultivo que más divisas genera para México, ya que una cantidad importante de éste es comercializado internacionalmente por su excelente calidad, sin embargo, enfrenta el problema del ataque continuo de plagas y enfermedades económicamente importantes que tienden a disminuir la producción del fruto, afectando con ello los volúmenes potenciales de exportación. Esto r e p e n t e directa e indirectamente en la economía de los productores y afecta a todos los agentes que intervienen de alguna forma en el proceso productivo de este cultivo.
Los actuales problemas de contaminación ambiental de los diferentes ecosistemas naturales y las consecuencias que estos provocan inciden en la reducción del uso de pesticidas químicos para el control de plagas y enfermedades en los cultivos.
Por lo anteriormente expuesto, es imperante e inminente el desarrollo de tecnologías alternativas que conlleven a la obtención de productos biológicos que coadyuven eficientemente al control de estas plagas y conjuntamente protejan el entorno físico evitando el impacto ambiental que provocan los productos sintéticos.
Las mayores dificultades para el empleo del hongo B. bassiam como insecticida de uso corriente han sido: encontrar métodos de producción eficientes y rentables de grandes volúmenes de propágulos, poder proteger este material contra la tensión ambiental y contra antagonistas químicos y biológicos y así obtener un producto final que permita un manejo tan eficiente como el de cualquier insecticida comercial (Morales et al., 1991).
Actualmente, en el Estado de Veracruz se está produciendo en pequeña escala a nivel comercial el bioinsecticida Bio-Zentla Bb en el Laboratorio Reproductor de Hongos Entomopatógenos de Zentla (LARHEZ) ubicado en la Colonia Manuel González Zentla de Huatusco, Veracruz. Los problemas a los que se han enfrentado durante el cultivo del hongo es la heterogeneidad genética del mismo, el fluctuante nivel de esporulación, la variación de viabilidad e infectividad de las esporas en campo.
Por lo cual, la presente tesis plantea una alternativa biotecnológica para la producción de un micopesticida biológico cuyo ingrediente activo serán las esporas del hongo entomopatógeno B. bsssiana obtenidas a partir de sustratos naturales y económicos corno el grano de arroz.
Patricia Elizabeth Baños Guevara - 4
I

OBJ€i?VOS
N. OBJETNOS
4.1. Objetivo general
Evaluar el uso del a m z entero y residual como sustratos sólidos para obtener la máxima producción de esporas del hongo entomopatógeno Beauveria bassima.
4.2. Objetivos particulares
* Determinar los efectos del contenido de agua del medio de cultivo y de latemperahira de incubación del hongo Beauveria bassima en la producción de esporas.
* Realizar perfiles de esponilación y de viabilidad a nivel laboratorio en los medios de cultivo bajo condiciones que resulten óptimas de contenido de agua del medio de cultivo y de la temperatura de incubación, a valor constante de pH.
Paíricia Elizabeth Baños Guevara d5
1

REVISION DE LITERATURA
V. REVISIÓN DE LITERATURA
5.1. B m a del grano de café (Hypafhenemus hampi, Fernad
5.1.1. Origen y antecedentes de la broca del pano de café
La broca del grano de café, Hqpothenemus hampei, es un insecto originario de &rica Ecuatorial, se encontró por primera vez en Francia en frutos importados de África !i fue reportada como plaga en este continente en 1901 en Gabón; de 1902 a 1904 se diseminó en el Congo, en 1908 se enwntfÚ en Uganda y Kenia de donde se expandió entre 1910 y 1965 por toda África: Guinea, Sierra Leona, Liberia, Ghana, Nigeria, Costa de Marfil, Fernando Po, Camenin, República Centroafricana, Ruanda, Burundi, Angola, Tanganica, Nysalandia, Rhodesia, Mozanbique, Santo Tomás y Príncipe. En Java apareció en 1909 (Koch, 1989). Por Asia se dispersó entre 1904 y 1963 y actualmente se encuentra distribuida en Tailandia, Vietnam del Sur, Malasia, Indonesia, Filipinas, Timor, Yemen, Laos y Taiwan (Budlo, 1992; Jiménez, 1995).
Actualmente, casi todos los países productores de café padecen de esta plaga. En América fue introducida accidentalmente en 1913 en Brasil, de donde se distribuyó al resto de los países de América; de acuerdo a un orden cmnológico en Perú se descubrió en 1959, en Guatemala en 1971, en Honduras en 1977, en Bolivia en 1978, en Jamaica y en México en 1978, en Ecuador y en El Salvador en 1981, en Nicaragua en 1988 y en Colombia en 1988; en Oceania ya invadió Nueva Caledonia, Papua y Nueva Guinea (Le PeUey, 1973; Jiménez, 1995).
5.1.2. Importancia económica de la plaga
La broca del café es un problema para los productores porque afecta directamente el fruto, que es la parte comercial de este cultivo. De hecho las normas de calidad están enfocadas a la cereza y se paga según se cumplan dichas normas (Rocha, 1996).
Según Koch (1989), la broca del café es una de las plagas m á s dañinas que existe en diversos países de África, Indonesia y América. Las pérdidas ocasionadas por la broca del café están estimadas en función del porcentaje de frutos infestados al momento de la cosecha y estas pueden ser variables, ya sea de un 25% de cerezas (fruto con 2 granos) infestadas hasta un 90% (Monterroso, 1981) y el deterioro en la calidad comercial alcanza algunas veces hasta el 50% (ANACAFE, 198 1). Tal aseveración radica en que la broca entra a una cereza para hacer su cámara de oviposición y las larvas generalmente se alimentan de uno de los granos y el otro queda sin ser atacado. Entonces se tiene una infestación de 5%, es decir que de cada 100 frutos hay un daño de 1.25 granos y son los que se van a perder, únicamente al inicio del ciclo y conforme avanza la temporada de cosecha, la broca puede completar 4-6 ciclos de vida, lo que incrementa el % de infestación. En México hay reportes de daños de hasta un 80%, pero esto es en caso de no controlar o dejar abandonado el predio. Este problema se presenta cuando baja el precio del café y los agricultores abandonan totalmente los cafetales, lo cual sigrufica que no hacen control alguno de malezas, plagas y enfermedades, por lo tanto, el daño se agudiza @echa, 1996).
- - - Pahicia Elizabeth Baños Guevara 6

REVISIÓN DE ISIZRATURA
El café es un cultivo de importancia económica para las regiones tropicales de México; especialmente en algunas de ellas, en donde es la principal fuente de ingresos. Los cafeticultores individuales, así como los países productores de café, siempre han estado expuestos a un grado de inseguridad económica debido a las fuertes fluctuaciones del precio de su producto. Al respecto, estos países han cambiado sus métodos tradicionales de producción para aumentar la calidad y rendimiento de sus cafetales al menor costo posible @ayer de México, 1980).
5.1.3. Distribución de la broca
5.1.3.1. Formas de dispersión
La broca del grano del café es un insecto capaz de volar sólo pequeñas distancias, de modo que en su diseminación de una región a otra, de un país a otro y de un continente a otro, principalmente hacia América, evidentemente el responsable ha sido el hombre (SARH, 1980).
Aunque algunas brucas llegan a volar hasta 375 metros, la mayoría vuelan menos de 50; su mayor actividad ocurre entre las cuatro y seis de la tarde. En su hábitat original el café crece bajo sombra, a la cual la broca está adaptada, lo que expiica las relativamente menores poblaciones de la plaga observadas en cafetales a pleno sol., aunque hay situaciones contradictorias como es el caso de Colombia, donde también hay plantaciones a pleno sol y el manejo agronómico es diferente. Se incrementa considerablemente el número de plantas por hectárea y provoca que entre sí se proporcionen sombra de la parte media a la parte baja de las plantas. Además, con un mayor número de plantas se dificultan las prácticas culturales de recolección de frutos brocados, las pepenas, repelas y arojes, lo que incrementa el porcentaje de infestación Uiménez, 1998, comunicación directa).
La dispersión ocurre por varias causas (Velas.Cn, 1995):
* Las hembras fecundadas buscan donde ovipositar. * Las hembras buscan otra cereza por exceso de humedad o calor. * Por inundación de l as cerezas infestadas. Las hembras de intercosecha son las que
propician nuevas infestaciones, ellas permanecen en gran número en los frutos caídos al suelo y en los dejados en las plantas. De ahí que la recolección de estos granos es la mejor medida para reducir las poblaciones iniciales del insecto.
Se ha comprobado que además de la capacidad de vuelo de la broca, la causa más importante de diseminación de la plaga ocurre por actividades humanas relacionadas con el cultivo del café entre las que se tienen (Velasco, 1995):
* La movilización del grano de café es la principal causa de dispersión. a
* Por los medios de transporte animal y mecánico. El movimiento de personal, utensilios y materiales de campo.
- - Patricia Elizabeth Baños Guevara 7

REVISIÓN DE LTERA'iWR.4
5.1.3.2. Distribución en Mexico
La broca del grano de café fue observada en México a fines de agosto de 1978 en el Sureste del Estado de Chiapas en el ejido Mixcum, municipio de Cacahotán en un beneficio húmedo, por lo que el 27 de octubre del mismo año se estableció la Cuarentena interior No. 12 quedando bajo nigimen cuarentenano los municipios de Cacahotán, Unión de Juarez (región del Soconusco) y Tuxtla Gutiérrez. Según el INMECAFÉ, para los go's, aproximadamente 80 mil ha de las 89 mil existentes en la región del Soconusco, se encontraban infestadas por esta plaga. Otra región del Estado de Chiapas invadida por la broca es el Centro-Norte, sin embargo se desconoce la superficie afectada (INmCAFÉ, 1983). Además de Chiapas, la broca se ha detectado en otros estados de la República Mexicana como son Oaxaca, Veracmz, Puebla y Guerrero. Desafortunadamente, ya re ha encontrado en el estado de Nayant, siendo la región m á s alejada del centro y del Sureste del país en donde se siembra café (SARH, 1980; Baker, 1985).
5.1.4. Taxonomía y sinonimia
La broca del grano de café Hflothenernus hampei Ferr. ha tenido cambios en su nombre científico. Por su tamaño pequeño se ha confundido con otras especies de la familia Scolytidae y su clasificación taxonómica actual de acuerdo con Wood (1982, citado por Villanueva, 1986 y por De la Rosa, 1993) es:
Reino Phylum Subphylum Clase Subclase División Orden Sub or den Superfamilia Familia Subfamilia Tribu Género Especie Nombre científico:
Animal Arthropoda Uniramia Insecta Pterygota Endopterygota Coleoptera Polyphaga Curnilinoidea Scolytidae Scolytidae o ipinae Cryphahi Hpthenernus (Westwood, 1836) hampei (Ferrari, 1867) Hypothenernus hampei Ferrari, (Ferrari, 1867)
Dentro del género Hpthenernus se han encontrado 27 especies en México, H. hampei es la única especie considerada que ataca a los frutos del género Coffeea spp. (Wood, I982 citado por De la Rosa, 1993). En relación a la sinonimia empleada en la literatura, se citan las siguientes en orden cronológico:
Cwhalus hampei Ferrari, 1887 Stephanodem coffeae Hargerdon, 1910 Xyleboms coffeivoms Stephanoderes cmki Hopkins, 19 15 Xyleboms coffeicola Campos Novaes, 1922 Stephanaderes punctatus Eggers, 1924
Van der Weele, 19 1 O
Paíricia Elizabeth Baños Guevara 8

REVISION DE LiTFXATURA P
5.1.5. Biología de la broca del grano de café
5.1.5.1. Morfología y ciclo biológico
La broca del grano del café, H hampei, tiene metamorfosis completa (Holometábola), pasando durante su ciclo de vida por cuatro estados: huevecillo, larva, pupa y adulto. La biología de este insecto ha sido estudiada intensamente en Java, Uganda, Malaya, S n Lanka, El Congo, Brasil, México y Honduras (Méndez, 1990).
Cada hembra pone un total de 30 a 74 huevecillos, que se incuban en temperaturas de entre 22 y 27°C durante un lapso de 10 a 12 días y vive alrededor de 250 días; dichos huevecillos son globosos ligeramente elípticos, de aspecto lechoso, muy pequeños, difícil de apreciarlos a simple vista; se les encuentra individual o en grupo dentro del fruto y su tamaño es de alrededor de 0.44 mm. Recién puestos son blancos, cambiando después a color amadento.
Después de 5 a 13 días de eclosionados los huevecillos, nacen las larvas que son gusanos blanquecinos, del tamaño semejante a la cabeza de un alfiler; sin patas, ligeramente curvados y adelgazándose hacia la cola. La cabeza está bien desamllada y es de color a m a d o grisáceo. La larva es de color blanco lechoso y tiene aspecto de un a m z b u t 0 en forma de “C” sin patas y con cabeza negra endurecida. Ai final de su desamllo miden de 1.88 a 2.23 mm de largo.
El estado de pupa dura aproximadamente de 4 a 9 días al cabo de los cuales se transforma en pupa, la cual es similar a la larva, pero carece de movimiento, o sea, es un estado de reposo antes de convertirse en adulto. Su color vana de blanco lechoso a pardo amarillento; toda la fase de larva a adulto se lleva a cabo dentro del fruto y dura alrededor de 20 a 36 días transformándose entonces en adulto y completando así su ciclo de vida (Le Pelley, 1973).
El adulto es un pequeño coleóptero que recién emergido es de color café claro y con el tiempo se toma de café obscuro hasta negro, de forma cilíndrica, ligeramente oblongo en su parte dorsal, del tamaño de la cabeza de un aifiler, cubierto de pelos que crecen hacia atrás, la cabeza es globular, antenas en forma de mazo y con diminutas puntuaciones finamente reticuladas, élitros o primer par de alas con rayas paralelas deprimidas longitudinaimente; en las hembras el segundo par de alas son membranosas adaptadas para el vuelo, en los machos estas están atrofiadas, por lo que no pueden volar. El tamaño del macho es de 1 a 1.25 nun de largo y la hembra de 1.25 a 1.82 mm (Velasco, 1995).
La duración del ciclo biológico de la broca del café, varía de acuerdo con las condiciones de desarrollo; en laboratorio el ciclo varia de 27.5 a 35.8 días a 24.6OC. Para las condiciones de Brasil y Guatemala, el tiempo promedio para que la broca complete su ciclo biológico de huevecillo a adulto está entre 21 y hasta 80 días, dependiendo de las condiciones bióticas y abióticas. La proporción entre hembras y machos es en promedio de 10: 1,10 cual puede deberse a que la disponibilidad de alimento es limitada y la especie tiene más potencial de aumento con una mayor cantidad de hembras que de machos. Esta proporción vana con la temporada y es en la de secas cuando hay más hembras por su mayor capacidad de sobrevivencia. De igual manera las hembras son más longevas que los machos, pueden vivir mínimo 135 días y los machos 46 días en promedio.
- 5- Pairicia Elizabeth Baños Guevara 9

REVISIÓN DE LlTERATuRA
El macho emerge uno o dos días antes que la hembra y permanece en la galería debido a su incapacidad de vuelo. La reproducción puede ser consanguínea dentro de la misma cereza, lo que d u c e la variabilidad genética de la población. Esto se debe a que el ambiente dentro de la cereza es relativamente estable. La cópula ocum cuando la hembra alcanza la madurez sexual ( t res a cinco d í a s después de la emergencia), la mayoría de las hembras abandonan el fruto después de ser fecundadas.
Con respecto al número de generaciones por año, Baker (1984, citado por Méndez, 1990) realizó conteos biológicos de broca en frutos de la planta y en aquellos que se encontraban en el suelo y consideró que bajo estas condiciones muchos insectos pueden completar de 3 a 4 generaciones por año y que esto depende de la disponibilidad de frutos maduros durante la emergencia de adultos. Baker y Barrera (1985) mencionan que en condiciones de campo en el Soconusco, Chiapas es muy probable que la broca no pueda completar más de d o s ciclos entre la maduración del fruto y la cosecha.
5.1.6. Fkología de la broca del grano de café
5.1.6.1. Hábitos
Villanueva (1986) menciona que una característica muy importante de la broca, es el hábito de reproducirse únicamente dentro de los granos, por ello es una plaga casi exclusiva del género Coffea Las hembras ovipositan sólo cuando el peso seco del fruto es del veinte por ciento o más, sino, permanecen algún tiempo en el canal de perforación hasta que la condición del fruto se adecua ovipositando luego en la galería un promedio de 30 huevecillos. Estudios recientes wiizados en Honduras indican que la hembra es capaz de reproducirse sin el macho, o sea, partenogenéticamente (Velasw, 1995).
La broca es atraída al fruto principalmente por su olor, aunque también influyen el color y la forma, los desechos y las heces de las brocas en frutos dañados. La broca ataca con mayor intensidad a los frutos de las primeras floraciones y la ocurrencia de muchas floraciones en los cafetos propicia el incremento de la población, ya que siempre habrá frutos adecuados por más tiempo. A los 57 y 97 días después de la floración (estado acuoso y semilechoso) la broca perfora y abandona el fruto; a los 107 días después de la floración (estado lechoso) la broca perfora y permanece en el canal de penetración; a los 137 días (estado semiconsistente) la broca empieza a preparar sus cámaras de oviposición y a los 147 días (estado consistente) ya se observaron los estadios de huevo y larva; entre los 207 y 227 días (estado duro y duro maduro) se aprecian todos los estadios biológicos y los endospermos destruidos. Por tanto, para controlar la bnxa es importante tomar en cuenta la etapa vegetativa del a h s t o , porque el insecto va a aparecer en el cultivo cuando los frutos tienen cierta consistencia, y no en la etapa acuosa por no ser atractivos para oviposita. Quizá perfore la cereza y al verla inmadura no prosiga su camino, en ésta etapa no hay tanto problema; lo fuerte empieza cuando el fruto se encuentra en estado masoso. Es entonces cuando el fruto capta la atención del insecto para ser perforado y hacer una especie de mina para llegar al centro de la cereza y ovipositar.
Debido a las bajas temperaturas que Ocurren en cafetales de altura (más de 1000 msnm), los niveles de infestación de la bruca han sido menores y su efecto no ha constituido problema económico en estos casos, aunque faltan evaluaciones a diferentes latitudes, ya que el comportamiento puede ser diferente en el Soconusco, Chis., más al sur, que en la Sierra Norte de Puebla o Hidalgo.
- Patricia Elizabeih Bañar Guevara I O

Las brocas son más activas en la oscuridad y con alta humedad relativa, con 55% de humedad su actividad se reduce aún en la oscuridad y en estas condiciones la mortalidad aumenta. Por esto, se recomienda la regulación de la sombra y la poda del cafeto, ya que estas prácticas reducen la humedad relativa en el cafetal. Probablemente la humedad del fruto sea un indicador para la bruca sobre las condiciones ambientales existentes. La lluvia parece ser más importante que la humedad relativa para la emergencia. El corte de la maleza antes y después de la cosecha propicia el secado de los frutos caídos en el suelo reduciendo la actividad y supervivencia de la plaga (Velasco, 1995).
Galán y Bodegas (1984) mencionan que las variedades de café comúnmente explotadas en el Soconusco, Chiapas, tienen diferencias en el desarrullo fenológico del fruto, la variedad Bourbon fue de maduración más temprana seguida de Catuai y la más tarda fue la variedad Robusta (Coffeea canephora), que en ocasiones llega a madurar hasta el mes de febrero, además de ser preferida por el ataque de broca, mientras que Bourbon y Catuai son cosechadas uno o dos meses antes. Estos investigadores observaron que en la asociación Bourbon-Robusta al final del ciclo y en ausencia de frutos de Bourbon, las poblaciones se incrementan en Robusta, mientras que en la asociación Catuai-Bourbon, notaron la preferencia de la broca por la variedad Bourbon, probablemente debido a los frutos más desarrollados para la oviposición.
5.1.6.2. Daños
Tal como lo señala Velasco (1995) los daños y pérdidas por ataque de broca son de diferente magnitud de acuerdo con la madurez fisiológica del fruto infestado. Los daños son causados en primer término por las hembras que penetran en la corona del fruto, tardando aproximadamente cuatro horas para llegar a la s e d a en donde forma su galería y oviposita.
Entre los principales daños causados por el ataque de la broca se pueden mencionar:
* Fruto joven, sirve de alimentación a las hembras que provienen de los frutos secos de la planta y del suelo, al ser más o menos perforado, se pudre y cae al suelo. Esta caída depende de la maduración del grano y puede variar del 5% al 10% de los frutos.
* El fruto verde ya formado semi-consistente o el fruto maduro atacado que no cae, pierde peso en proporción al grado que es parasitado y consumido por el insecto.
* El fruto dañado que no flota, como vano, también pesa menos y puede provocar castigos en las partidas de pergamino u oro por su apariencia, lo que disminuye el valor comercial del café.
* Las pérdidas totales de frutos dañados pueden afectar signrficativamente el rendimiento desde 6 a 60%, cuando el nivel de infestación vana del 7 al 100%.
* Un lote de café pergamino u oro con más del 14% de humedad en el lugar de almacenamiento, permite la supervivencia de numerosas brocas que pueden constituir una fuente de infestación durante cerca de 4 meses.
Patricia Elizabeth Baños Guevwa I 1

5.1.7. Métodos de control de la broca del grano de cafe
5.1.7.1. Muestre0
Al penetrar la broca en el grano del café no sale en semanas, hasta después de haber ovipositado, abandonándolo sólo bajo presión de algún parasitoide o bien para escapar de la acción de un pmducto químico. Al no salir del fruto, su control químico es difícil. Lo conveniente es aplicar un control antes de que entre a la cereza y para elio es necesario hacer muestreos continuos a partir de la etapa fenológica, aproximadamente a los 130 dias después de la floración y en adelante.
Por elio se recomienda hacer un programa de control en el que es necesario saber el momento adecuado para considerar a la broca en cantidad suficiente para ser combatida por diversos métodos. Por un lado, se hace un muestre0 donde al aparecer 5% de infestación es la indicación para iniciar medidas de control contra la plaga. Es muy recomendable monitorear las huertas al menos cada 30 días y llevar un historial de la plaga (Rocha, 1996).
5.1.7.2. Control legal
Cuando la broca del fruto del café llega a un nuevo país o área lo hace para establecerse, si las condiciones ecológicas lo permiten. Todos los países invadidos por la broca sin excepción, se han visto en la necesidad de poner en práctica las medidas de control legal tendientes al control y/o erradicación de la plaga. Estos instrumentos son la expedición de decretos que declaran en cuarentena l as áreas afectadas para impedir la diseminación de la plaga hacia las áreas libres y la creación de campañas tendientes a lograr la erradicación total de la plaga. Estas medidas se han aplicado en Perú, Guatemala, El Salvador, Honduras (Campos, 1985 y Muñoz, 1985, citados por Méndez, 1990). En México, la Dirección General de Sanidad Vegetal expidió la cuarentena interior No. 12 contra la broca del grano de café, misma que fue publicada en el Diario Oficial de la Federación el 27 de Octubre de 1978, en la cual se especificaban las medidas a tomar para evitar la diseminación de la plaga (SARH, 1980), esta cuarentena fue deroga& y actualmente hay otras normas más recientes.
No obstante la aplicación de campañas y cuarentenas, la broca ha continuado su avance a partir de sus focos de infestación; el fracaso de las campatias se debe a varios factores: al tamaño diminuto de la broca, su gran capacidad de colonización; la continuidad de las zonas cafetaleras de muchos países vecinos; el movimiento inescrupulosos de semillas infestadas, el iraslado de cortadores de un país vecino a otro durante la época de cosecha y por mera casualidad, debido al transporte de la plaga en cualquier utensilio o medio de transporte (Alonzo, 1985). Este mismo autor indica que es ilusono pensar en la erradicación de la plaga, debido a factores tales como: el tamaño extenso de las fincas, la topografía accidentada de las mismas, el elevado númem de frutos por inspeccionar, tamaño relativamente pequeño del fruto, la presencia de hospederos alternos y la movilización de jornaleros y equipo a lo largo del año.
Existen Nonnas Oficiales Mexicanas relacionadas con el problema de la brow del café; tal es el caso de:
Patricio Elizabeth Baños Guevora 12

REVISIÓN DE LITERATURA P
* La Norma Oficial Mexicana (con carácter de emergencia) NOM-EM-019-FiTO-1994. Cuarentena exterior para prevenir la introducción y diseminación de las plagas del café, publicada en el Diario Oficial de la Federación el día 16 de noviembre de 994.
* La Norma Oficial Mexicana (con carácter de emergencia) NOM-EM-122-FiTO-1994. Aviso de inicio de funcionamiento y certificación de personas físicas o morales interesadas en prestar los servicios de tratamientos fitosanitarios a vegetales, sus productos y subproductos de importación, exportación o de movilización nacional, publicada en el Diario Oficial de la Federación el día 2 7 de diciembre de 1994.
El 22 de agosto de 1995, se publicó en el Diario Oficial de la Federación el Proyecto de Norma Oficial Mexicana NOM-002-FITO-1995, en la que se establece la campaña contra la broca del café y en donde la Ley Federal de Sanidad Vegetal establece t res tipos de zonas o áreas geográficas para permitir la movilización por el territorio nacional de los vegetales, sus productos y subproductos, que son: zonas bajo control, zonas de baja prevalencia y zonas libres. Esta Norma Oficial es aplicable en los siguientes casos: productos y subproductos vegetales del café (cereza de café, café pergamino, café om, pulpa de café, cascada de café); industrializadoras (beneficios secos y beneficios húmedos); centros de almacenamiento; áreas de producción (cafetales); medios de transporte (vehícdos, contenedores, canastos, costalera) (Diario Oficial de la Federación, 1997).
5.1.7.3. Control químico
El control químico es, a la vez, la mejor y la peor de las soluciones al problema de la broca. Mejor por su efecto inmediato y su confiabilidad, peor por sus efectos secundarios indeseables después de la aplicación; tales como: contaminación ambiental, residuos 'en la cosecha, resistencia del insecto a los insecticidas y desequilibrio ecológico.
El uso de productos químicos constituye quizás una de las áreas m á s exploradas para el control de H. h m p i Además del daño directo por ataque de la broca, el control químico no apropiado puede ocasionar disturbios ecológicos perjudiciales tales como (Velasco, 1995):
* Resistencia del insecto a los químicos. Resultados obtenidos en Nueva Caledonia, Oceania, indican que poblaciones de broca han adquirido resistencia al Endosulfán, estos insectos soportan mil veces la cantidad del producto para alcanzar una mortalidad similar a las brocas susceptibles. En Centroamérica no se ha observado resistencia de la broca, a pesar de que en algunas zonas de Guatemala se ha aplicado Endosuifán por 17 años.
* Desequilibrio ecológico. Como resultado de la muerte de enemigos naturales y el surgimiento de nuevas plagas perjudiciales que antes eran potenciales.
* Residuos tóxicos. Estos pueden rebasar los limites de tolerancia y contaminar el grano para
* intoxicaciones.
consumo, la pulpa, el suelo y el agua.
- - Patricia Elizabeth Baños Gumma 13

REVISIÓN DE LITERATURA
5.1.7.4. Control cultural
La aplicación de las prácticas culturales pretenden reducir las poblaciones de broca y propiciar la formación de un mirroclima desfavorable para el desarrollo de la plaga. Para ello es necesario realizar toda una serie de labores como son: poda de árboles de sombra, cosechar d o n n e vayan madurando los frutos, poda de cafetos, aumentar la distancia de plantación de los cafetos, control de malezas y la aplicación adecuada de fertilizantes (ANACAFÉ, 1981; Rocha, 1996).
* Pepena y repela: Posiblemente lo que más sobresale del control cultural es el control manual que consiste en la recolección de frutos después de la cosecha. Esta actividad se recomienda realizar en todas aquellas áreas en donde la broca se encuentra establecida; para lo cual es necesario recoger todos los frutos que se encuentren en el árbol o en el suelo;.ya que las hembras de interperíodo que son las formas de supervivencia de la plaga se alojan en los frutos secos dejados en la planta o en el suelo y en los frutos verdes de floraciones fuera de petiodo, esta es la “cosecha sanitaria”. Los frutos recogidos se hierven por 5 minutos o se fumigan con pastillas de Fostoxin en la proporción que señala la Norma Oficial (Velasco, 1995; Rocha, 1996).
El costo de la pepena y la repela puede ser compensado por el valor del cafe recolectado, pero aunque éste no sea el caso, la eliminación minuciosa de las hembras de intercosecha puede evitar la necesidad del control químico en la próxima fructificación (Rocha, 1996).
* Repase: Es otra labor no menos importante, consiste en la recolección de los frutos tempranos, producto de las floraciones “locas”; estos frutos representan una mínima parte de la cosecha total, pero son los primeros en ser atacados por la broca; a la vez que se colectan los frutos perforados durante esta fructificación para evitar que la broca tenga progenie en su interior ya que se sabe que el insecto va a invernar dentro de ellos y esperar ahí la siguiente floración. Se utiliza bastante mano de obra para revisar todas las plantas. Esto lo puede hacer el pequeño y mediano productor por el tamaño de la superficie cultivada (Velasco, 1995; Rocha, 1996).
Los frutos recolectados de la pepena, la repela y el repase pueden tener varios destinos dependiendo del grado de infestación: si están muy deteriorados y existe abundancia de estados biológicos, lo más recomendable es enterrarlos, previo tratamento con Endosulfan polvo al 3%. En el caso de que aún puedan ser aprovechados, deben sumergirse en agua hirviendo durante cinco minutos para matar cualquier fase de desarrollo de la plaga (ANACAFÉ, 198 I).
* Regulación de sombra: Debe evitarse el exceso regulando el número de árboles, siendo de 100,90 y 80 en alturas de 600,900 y 1200 msnm, respectivamente; y su copa debe estar en forma de paraguas para que permita el paso de luz. La poda de los árboles de sombra debe realizarse al inicio de las üuvias, que es también el inicio de la fructificación (Velasco, 1995).
Específicamente lo que respecta al manejo de sombra se sabe que el insecto es afectado por la cantidad de ésta, de manera que, cuando encontramos cafetales muy sombreados, la broca se presenta en grandes cantidades, hecho que no sucede en los cafetales donde hay sombraje regular.
Pnhicin Eliznbeth Baños Gumma 14

han logrado éxitos en el sureste con árboles del género inga, y hablando en números se dice que una sombra regular se logra con 100 árboles por hectárea. Por otro lado, se sugiere la poda de los cafetos y de los árboles de sombra para reducir el problema de broca y, además, eliminar microclimas favorables para el desarrollo de enfermedades (Rccha, 1996).
* Poda de los cafetos: Debe adoptarse una poda agresiva con ramas mal formadas, viejas y enfermas para permitir mayor ventilación y luz en la mayoría de las ramas, lo cual d u c e la humedad relativa en perjuicio de la broca, ya que la supervivencia y actividad son mayores con alta humedad y obscuridad. La poda por planta puede ser más adecuada para cafeticultores pequeños, mientras que la poda por calles será mejor para extensiones mayores del cultivo (Velasco, 1995).
Para evitar los daños que ocasiona la broca, se emplean medidas preventivas (cuarentenas), de erradicación (campañas), de control integrado de la plaga a base de medidas culturales (pepena, repase, podas, etc.) y las prácticas agrunómicas tales como (velasco, 1995; Rocha, 1996):
* Control de malezas. Para facilitar la recolección manual de los frutos del suelo (pepena) se recomienda una limpia con machete antes y después de la cosecha. En caso de que queden algunos frutos en el suelo, el sol y el aire los secan aumentando la mortalidad dt: los diferentes estados biológicos de la broca.
* Fertiiización. Una fertilización adecuada puede resultar en menos floraciones discontinuas (“locas”) que permitan la supervivencia de la plaga y las floraciones principales serán las más abundantes y uniformes con un período de fructificación más corto.
* Uniformidad varietal. El uso de variedades de floración temprana y tardía permiten a la broca disponer de frutos por un mayor período de tiempo antes y después de la cosecha; por lo tanto habrá mayor posibilidad de que la broca sobreviva en cantidad importante durante y hasta la siguiente cosecha.
5.1.7.5. Control biológico
Debido a que la broca es una plaga exótica, no nativa del país, ésta se encuentra libre de la mayoría de sus enemigos naturales que la atacan en su lugar de origen. Con base en lo anterior, el control biológico clásico puede ser una estrategia de éxito como uno de los componentes del manejo integrado de la plaga (Velasco, 1995).
El uso del control biológico se viene realizando desde hace algunos siglos. La National Academic of Sciences en 1980, menciona que la idea de utilizar los agentes micrubianos en el control de insectos se concibió en el siglo Wiü. Constantemente se aislan nuevos organismos, hasta ahora han sido encontrados 1165 microorganismos relacionados con los insectos, casi todos son patógenos. Estos comprenden 90 especies de variedades bacterianas, 260 especies de virus y rickettsias, 460 especies de hongos, 255 especies de protozoarios y 100 especies de nemátodos (Hernández, 1995; Berlanga, 1995).
z Patricia Elizabeth Bañar Guama 1 5

REVISION DE mm
* Er un método muy económico. * El control biológico a diferencia del control químico no acarrea amenazas de contaminación y daños si medio ambiente; también e b la posible aparición de razas resistentes entre la población de la plaga, lo que requiere del desarrollo y uso de nuevos pesticidas con consecuentes nuevos gastos. * Ertablece un sistema regulador permanente que mantiene a la plaga bajo control, sin tener que realizar trabajos ni gastos posteriores. * Tiene mayores ventajas en comparación con el control quimim que requiere repeticiones regulare y caras por periodos prolongados. * El control biológico virtualmente no usa energía, en comparación de muchos productos químicos que se basan en combustibles fbiies (carhjn, petróleo y gas) y su elaboración
El control biológico es u11 método de combatir las plagas ya sean de plantas, de animales o del hombre, exponiétidolas al ataque de sus enemigos naturales. Virtualmente todo organismo vivo (incluidas l a s plagas) tienen enemigos naturales que pueden usarse para ese propósito. El método clásico de control biológico es introducir en un área, en la que tcdavia no existe, un enemigo natural adecuado para una plaga, que con frecuencia llegó al área accidentalmente. El control biológico es el método m á s económico de combate. El concepto de control biológico se ha ampliado para incluir todos los métodos que emplean organismos vivos para combatir plagas Esos métodos comprenden el mejoramiento de plantas resistentes a las enfermedades, la liberación de machos estériles de una plaga para interferir en la reproducción, el uso de sustancias que atraen sexualmente a los machos y permiten atraparlos y eliminarlos antes de que puedan unirse con las hembras y el manejo genético de poblaciones de plagas a fin de producir individuos estériles (Agricultura de las Américas, 1978). En el cuadro 1 se exponen las ventajas y desventajas del control biológico.
* La certeza y grado de control a1 alcamrse varían en diferentes circunstancias. * La investigación para el desarrollo de medios biológicos de control puede requerir mucho tiempo, mientras tmtc la plaga se disemina más rápidamente. Después de la aplicación de iales métodos pueden tardar a h n o s mior para a l c a m r su plena eficiencia. * L o s proyectos de control biológiu, de cada phga deben tratarse separadamente, es decir hay alta especificidad en los bioinsecticidas.
ZUADRO I. VENTAJAS Y DESVENTAJAS DEL CONTROL BIOLÓGICO vENT.qA.5 I DESVENT"4.5
I requiere s a n cantidad de enexyja. RJENT!2 Elaboración propia
I
Hay dos tipos de control biológico, inundación y aumentación, ambos pueden ser practicados con microbios y otros organismos; el control por inundación involucra el uso de un gran número de organismos por un corto tiempo para suprimir o destruir una población; el control por aumentación inicia supliendo una población natural existente la cual tiene el poder de controlar una plaga o enfermedad (Lisansky, 1985).
En 1944, en la Universidad de California, se inició el Servicio de Diagnóstico como una ayuda para los entomólogos de la misma universidad y se desarrolló rápidamente hasta convertirse en un servicio mundial. Agruparon a los patógenos de insectos en cinco categorías principales: hongos, bacterias, pmtozoarios, virus y nemátodos (Agricultura de las Américas, 1978).
La comercialización de pxvductos orgánicos se incrementa y diversifica en México, es destacable que el mercado internacional está asegurado ya que contribuye al ingreso de divisas al país, mejora los sistemas de producción agrícola y prácticamente se nulifica el impacto ecológico de esta actividad. Debido a esto en las últimas décadas el control biológico ha tenido mayor auge, además de ser más económico y no hay residualidad ya que es específico para el tipo de plaga que se pretenda combatir.
Patricia Elizabeth Bañm Guevma 16

5.1.7.5.1. Hongos
IA búsqueda de hongos entomopatógenos como enemigos naturales de insectos plaga es la primera acción en la implementación del control microbiano, la finalidad de esia exploración es la de detedar y de ser posible aislar los organismos asociados a plagas de importancia económica (Khachatourians, 1986, citado por Thomas et al., 1987).
Lisansky (1985) menciona que se conocen alrededor de 400 especies de hongos que atacan insectos y gorgojos. Los hongos usualmente infectan a sus hospederos por invasión directa lo que difiere de la mayoría de bacterias y viruses, que atacan a las plagas s i n ser ingeridos.
El uso de agentes entomopatógenos como altemativa ante los agroquímicos es un hecho que en nuestro país ha venido cobrando fuem, sobre todo en lo que va del presente decenio. Desde 1989, se inicia de manera sistematizada y masiva la producción de estos agentes en el antiguo Centro Reproductor de Organismos Benéficos. Actualmente existen en México 8 laboratorios autorizados para el aislamiento, identificación y evaluación de cepas y especies de hongos: (I) la Escuela de la Frontera Sur (ECOSUR), ubicada en Tapachula Chiapas; (2) el Instituto Tecnológico de Oaxaca (ITAO), con sede en Oaxaca, Oax.; (3) Laboratorio Rural Reproductor de Hongos Entomopatógenos de Zentla, ubicado en la Col. Manuel González Zentla de Huatusco, Ver.; (4) Colegio de Postgraduados (CP); (5) Centro Nacional de Referencia de Control Biológico, ubicado en Tecomán, Colima; (6) el Programa de Café del instituto Nacional de investigaciones Agrícolas y Forestales (INIFAP), que reside en Rosario izapa, Chiapas; (7) la Unidad de Estudios de Postgrado de Acapulco, Guerrero; y (8) el Centro Regional Universitario Oriente (CRUO-UACh) (Ruales, 1996). Los agentes multiplicados son principalmente Metarhizium anisopliae, Beauveria bassiana, Aschemonia alepodis, Paecilomyces fumosoroseus, Verticillium lecanii, Hirsutella thompsorrii y Nomuraea rileyi y otras especies que están en evaluación.
Ciertamente los hongos entomopatógenos no ocupan un lugar preponderante, pero están distribuidos entre todos los grupos de los hongos imperfectos (Phycomicetes) pasando a través de Ascomycetes y Basidiomycetes. Además, esta característica les concede un potencial particular de acción debido a la variedad de su modo de reproducción, el cual es particular para cada grupo sistemático y su capacidad a desarrollarse bajo diferentes condiciones ecológicas (Ferrun, 1978).
Los hongos son particularmente importantes para el control de Coleópteros, porque las infecciones virales y bacterianas son raras entre los escarabajos. Los hongos entomopatógenos están asociados con insectos que viven en diversos habitats, incluyendo agua dulce, suelo, superficies del suelo y locaciones aéreas (Hajek y Leger, 1994). Además se presentan como una alternativa real para el control de insectos, dado que su vía de infección es cuticular y no requieren ser ingeridos para actuar (Fuxa, 1987, Carruthers y Hural, 1990, Dolling, 1991 y Alatorre, 1990, citados por Hemández, 1993).
No obstante, de las 700 especies de hongos entomopatógenos conocidos actualmente, sólo 10 especies han sido, o se están desarrollando para control, y el alto potencial de los hongos entomopatógenos no ha sido aprovechado. Para desarrollar hongos con propósitos de control, es n d o entender los requerimientos para inducir altos niveles de transmisión de la infección en el campo que son características de las epizootias (Hemández, 1993).
I 7 PaMcia Elizabeth Baños Guevnra

5.1.7.5.2. Bacterias
Se han descrito alrededor de 90 especies de bacterias que infectan insectos; actualmente hay productos comerciales basados en al menos dos de ellas y existen otros que se están produciendo experimentalmente. Los primeros ensayos biológicos se realizaron con la bacteria Bacillus thuringiensis en 1958 y 1959, salieron al mercado wmo producto comercial hasta 1984; s l l i embargo no existe sino hasta 1993 la primera presentación de producción nacional; los formulados que se vendían para finales de 1995 eran importados, por lo cual se depende del exterior. Las personas que realizan trabajo sobre esta bacteria no han llegado a sensibilizar a los empresarios para desarrollar productos más adecuados a las necesidades de los mismos insectos, clima, etc.. Las especies de Bacillus thuringiensis son usadas para el control de plagas de orugas, mosquitos y moscas negras; Bacillus ppilli8e para el control de escarabajos Japoneses, Bacillus sphaericus es considerados un patógeno altamente promisono para el control del mosquito pero todavía no está es producción comercial.
Las bacterias producen cristales proteicos, los cuales se vuelven insecticidas después de la digestión en medio alcalino encontrándose en el intestino medio de especies susceptibles. Los cristales varían entre los muchos serotipos y aislamientos de B. thuringiensis produciendo un espectro variado de actividad contra diferentes insectos:. L a s dos cepas comerciales son “Kurstaki”, usadas contra orugas, e “Israelensis” (H-14), efectiva contra mosquitos y moscas. Algunas cepas de B, thuringiensis producen una toxina soluble llamada beta-exotoxina la cual es altamente tóxica para las moscas caseras y no necesita ser ingerida (Lisansb, 1985).
5.1.7.5.3. Protozoarios
Los protomanos patógenos se han estudiado poco porque en general no pueden desamiiarse en medios artificiales y deben identificarse en la naturaleza a través del tamaño, forma, estructura y formación de las esporas (Apicultura de las América, 1978).
5.1.7.5.4. Virus
En Estados Unidos el Dr. Entwistle tiene un laboratorio en el Instituto de Vimlogía en Oxford, y ha estado caracterizando por muchos años la acción del Virus de la Polihedrosis Nuclear (VF’N o NPV) de Neodiprion sefifex Hay muchos m á s viruses que son activamente conocidos contra insectos que pueden ser producidos y comercializados wmo agentes de control de plagas con un costo accesible para el productor (Lisansly, 1985). En México, en el Centro de Investigaciones de Estudios Avanzados del instituto Politécnico Nacional (CINVESTAV) se ha Wciado un pmgrama de trabajo que involucra aspectos de biología molecular y de ingeniería genética sobre este virus.
5.1.7.5.5. Nemátodos
Los nemátodos son un grupo sobre el cual se ha empezado a trabajar en México, con grandes posibilidades de producción y comercialización. Son agentes que por su naturaleza son los únicos w n capacidad para “buscar” a su presa, pero son, al mismo tiempo, menos específicos y no distinguen un insecto “blanco” a diferencia de los dos anteriores.
Paíricio Elizabeth Bañas Guama 18

REMSION DE LiTERA'IzIRA
5.1.8. Enemigos naturales de la broca del grano del cafe
Siendo la bruca originaria del centro africano, es en este continente donde se han encontrado los más importantes enemigos naturales. El control biotecnológico de esta plaga se ha encaminado al estudio de los enemigos naturales que parasiten la broca (Bustillo et al., 1997), los que se han ampliado notablemente como se puede observar en el Cuadro 2.
El control biológico utilizando parasitoides, podría ser la alternativa que permita frenar el avance de la broca del café a lugares no infestados, basándose en el hecho de que el cafeto es un cultivo tropical perenne considerado como un buen hábitat para la proliferación de enemigos naturales, además de los grandes beneficios ecológicos derivados del control biológico incluyendo algunos parasitoides como: Sclerdermus cadavericus y Aphanogmus úictpna (Quezada, 1985).
5.1.8.1. Uso del hongo Beauveria bassiana Los hongos entomopatógenos usados para el control de la broca del café,
Hypothenemus hampei (Ferrari), son una opción fundamental en el desarrollo de un programa de manejo integrado, cuya finalidad sea la preservación del ambiente y el uso racional de insecticidas químicos. Entre los enemigos naturales que atacan a la broca del café se consideran a los hongos entomopatógenos B. bassima (Balsamo) Vuillemin, Spicaria javanica y Metarhizium anisopliae, bajo las condiciones de los ecosistemas cafetaleros mexicanos. Se sugiere, además, la alternancia de las diferentes estrategias de control, ya que si se tiene pensado liberar algunos parasitoides, habría que planear la aplicación de un hongo para no afectar al organismo benéfico, de ahí la importancia de B. bassiana.
Fernández et al. (1985, citados por Méndez, 1990) mencionan que B. bassiana en el hongo entomopatógeno que más daños causa a la broca del café Hypothenernus hampei. Al probar el efecto de B. bmiana inoculado sobre granos, hojas e insectos, observaron que el tratamiento a los insectos dio el peor resultado (52% de mortalidad), mientras que la aplicación sobre las hojas y granos presentó una mortalidad del 92% y 8!3.1% respectivamente.
En el Congo Belga se probó B. bassiana para controlar la broca del café llamada entonces Stephandem hampei, indicando que el material fungoso debe aplicarse antes de que el insecto invada la cereza, de este experimento se obtuvo que la máxima mortalidad de la broca por efecto del hongo fue de 63% en los meses de julio y agosto, mientras la menor incidencia se presentó en m a m y abril; considerando el grado de maduración de los frutos, se encontró un 48.2% de mortalidad en frutos verdes y un 3.28% en los maduros, concluyendo que la infección del insecto se efectúa en la superficie del fruto probablemente cuando los insectos están volando de un fruto a otro para ovipositar.
Algunos investigadores señalan que B. bassima forma una capa algodonosa de color blanco-rosado en la entrada del orificio perforado por la broca bajo condiciones ambientales favorables de alta humedad relativa (80%) y temperatura de 20 a 3OoC, además de altas poblaciones de la plaga (Aregger, 1992).
L
Pairicia Elizabeth Baños Guevara 19

I
REVISIÓN DE IslzRATuRA
CUADRO 2. ENEMlGOS NATURALES DE LA BROCA DEL CAFÉ (Hpthenemus hampel, Ferran)
TIPO Y ESPECIE IMENóYr€uos unilia: Bethviidae ztoparásitoi y predatores) Cephaionomia stephanoderis letrem, 1961)
Cephaionomia spp.
proxlpsnasuta (Waterston, 1923:
amilia: Eulophidae Phymastichus coffa
amilia: Braconidae Hetemspilus coffeicoia Schmiedeknecht, 1923)
amilia: Tetrastichinae (No LaSifiUido) Sclemderma cadavericus
amilia: Fonnicidae (predator) CIimaiogmter cuwhphosus
May.)
EMtPImos amilia: Pyrrhocorida (predator) Dhdmusmb&kzosus (F).
PAIS
Casta de Marfil, Togo, África
Oriental, México y Ecuador
Brasil
Brasil, Camenin, Uganda,Tanzania, Costa de Marfil,
Kenia,Togo
Togo, Benin, brnenin y ,asta de
Marfil, Africa
Uganda,Tanzania, Cox30
Togo,Kenia, Uganda
Brasil
Java y Brasil
OBSERVACIONES Y REFERENCIAS
b En México fue criado en laboratorio y el 31 de octubre de 1988 se realizó la primera liberación en campo. En los primeros meses después de la liberación se registraron los valores máximos de parasitismo, hasta 80%. Después de un año de la liberación el parasitismo requerido fluctuó de 4 a :!3%. introducida en México y Ecuador en 1988. En Indonesia en 1989. (ie Pelley, 1968, La Saiie, 1990, y Barrera et al., 1991, .citados porKoch, 1989) En Espíritu Santo en “coniion” (Benassi, 1989, citado por Koch, 1989). Bautizado como la “avispita de Uganda”. Descubierta en Brasil por el Instituto Biológico de Sa0 Paulo, en 1929 y se adaptó desde 1993. introducida en Mexim y Ecuador en 1988, en Perú en 1962, sin éxito. Reintroducida en indonesia en 1’389. Citado por Bergamin, 1944; Le Peiiey (1968, citado por Koch, 1989) y La ;Salle (1990,citado por Koch, 1989). Parasita adultos de Hpthenemus hamPr,; fue colectada entre 1991 y 1992 y cuarentenada en los laboratorios del instituto internacional de Control Biológico (IIBC) de ingiaterra e introducido a México, de los se is envios que se hicieron emergieron un total de 1268 parasitoides adultos. Citado por Le Peiiey (1968, citado por Koch, 1989); La Salle (1990, citado por Koch, 1989).
Parasita larvas mediante largo ovipositor.
Nuevo género y especie parasita adultos. Causa dermatitis en el hombre.
Hormiga que destruye un número considerable de formas inmaduras de broca que se encuentran en el interior del fruto infestado. Reportado por Pinto de Fonseca y Araujo (1999,citado por Koch, 1989). -
Picorrdrido que se alimenta del adulto de la broca. Reportado por Klein-Koch et al. (1988, citado por Koch, 1989).
- - Patricia Elizabeth Baños Guevara 20

I
TíPO Y ESPECIE PAIS -TODOS
Hetemrhaáditis spp. Noreportados
HONGOS * ikauveria bassima CongoBelga,Ken%
C a ~ ~ , Uganda, Togo,Zak,CoS~
de MarfilJava, Indonesia, Brasil,
Peni, Ecuador, Colombia,
Guatemala y Méxicn
Java> Indonesia, Asia y África
* hfeiarhuium mimpfiae
* Nomuraea riieyi Phaecilmyces (=Spicaria)
javanicus
* Hirsuteiia eleufhrratorum ’ Iuwiwn omspnun
* p: tenujpes
OBSEKVACIONES Y m C I A S
Citado por CAB-International institute of B i o l ~ c a l Control (1 989). - Enemigo natural, cosmopolita. Existen diversas razas. Monterroso (1981) seríala que en 1978 se reportó en Guatemala. Villacorta ( 1 984 , citado por Mendez, 1990) ruilizó estudios en Brasil, encontrándolc hasta en café pergamino.
Reportado por CAB-international Institute o1 Biological Control (1989). . Reportado p CAB-international Institute ol Biological Control (1989).
Reportado por (Koch, 1989).
Méndez (1990), establece que el crecimiento micelial del hongo emerge de los costados de la broca y de las partes membranosas de la región bucal; los ramilletes de conidióforos alcanzan a cubrir todo el cuerpo, excepto las áreas quitinkadas como son los élitros y la parte superior del pronoto; generalmente bajo condiciones de laboratorio del micelio de B. bmima emerge y esporula de cualquier parte del insecto; cuando la inferdm ocurre en el interior del fruto del café, el micelio envuelve totalmente al cuerpo de la broca y la esponilación ocurre con mayor abundancia en la parte exterior del orificio de entrada de la cereza sin llegar a invadir el tejido vegetal.
Existen pocos estudios que permitan conocer la abundancia del hongo sobre la plaga, en el Ecuador, se ha registrado su presencia principalmente en cerezas verdes y rojas, y la incidencia de B. bassima está dada de acuerdo a las condiciones favorables del medio ambiente, como alta humedad relativa, alta nubosidad y temperatura óptima, pero sobre todo ,abundante población de la plaga.
El hongo entomopatógeno B. bmima se encuentra presente en el Soconusco, Chispas, afectando a las poblaciones en estado adulto de broca, Hpfhenemus hampei, en proporciones que no superan el 1%. El hongo se localiza tanto en frutos de C o f a artibica corno de C. canephora variedad robusta, y al realizar bioensayos con cepas del hongo observó que las cepas “Ecuador” y “Caen” resultaron las mejores para el control de H. hampei. 8. bmima es considerado como el mejor microorganismo entomopatógeno que puede utilizarse contra la broca del café, ya que se le ha encontrado en cualquier lugar donde se presenta la plaga, independientemente de la variedad de café de que se trate.
* 21 Patricia Elizabeth Bañar Guevara
~

Bustillos et al. (1991) reporta que se ha encontrado este hongo en forma nativa en muchas de las regiones de Colombia, a donde ha llegado la broca. Actualmente, se cuenta con un cepario de 92 aislamientos de B. bassima, provenientes de diferentes localidades y condiciones agroecológicas. Este material está siendo sometido a una caracterización con el objetivo de conocer la relación existente entre ellos, y s u s fuentes de variación en cuanto a patogenicidad, producción de conidias, adaptación o tolerancia a factores abióticos. De estos aislamientos, 18 fueron suministrados por institutos como IIBC (Instituto Internacional de Control Biológico) y por países como Brasil, Ecuador y Guatemala; aproximadamente la mitad de estos aislamientos han mostrado actividad contra la broca (BustUos et al., 1991). Ai hacer evaluaciones de campo con el hongo B. bassima para el control de la broca del café en Colombia, observaron que al asperjar B. bassiana se logró un 36% de mortalidad; 48.1% después de tres aplicaciones y el efecto del hongo sobre la broca durante 119 días de evaluaciones y después de realizar seis aspersiones, la infección por el hongo se incrementó hasta alcanzar un promedio de 69%.
En Colombia, el efecto del hongo sobre poblaciones de broca se ha evaluado a través de d o s estrategias, una de introducción a sitios donde no existe y otra como un insecticida biológico aplicado en un programa de manejo integrado (MiP). La introducción ha sido exitosa y el hongo se manifiesta en diversos niveles en todas las áreas donde ha sido asperjado. Los niveles promedio de infección sobre la población de broca durante 1995 fueron del 4 5%.
En cuanto al uso del hongo como insecticida los resultados son erráticos, obteniéndose niveles bajos (20-30%) hasta niveles altos (60-7006). Este hongo sólo es efectivo cuando la broca entra en contacto con las conidias, al tratar de penetrar la cerem; si el insecto ya entró a la cereza es difícil que el hongo lo pueda afectar. Si B. bassiana se aplica antes de que la cereza esté susceptible a ser ovipositada por la broca o en caso contrario cuando ya ha afectado al fruto, entonces es dinero tirado porque realmente no hay ningún tipo de control.
5.2. El hongo entomopatógeno Beauvena bassiana
5.2.1. Antecedentes históricos y origen
Durante años, las observaciones de poblaciones de insectos han demostrado que los hongos son patógenos naturales, importantes y efectivos, reguladores de insectosl sin embargo, dependen en alto grado de la humedad y temperatura tanto ambiental como del microambiente que rodea al d o , razón por la cual en la mayoría de los casos no han sido considerados como una alternativa de confianza.
En Francia e Italia, donde la producción de seda fue importante en los siglos 16 y 17, se perdieron l as larvas de los gusanos de seda debido a la enfermedad anual de la “muscardina”; en 1835, el científico italiano Agostino Bassi de Lodi (el padre de la patología de insectos) mostrú que el problema que afectaba al gusano de seda era causado por un hongo que se multiplicaba en todo el cuerpo del insecto. Este fue el primer microorganismo en ser reconocido como un agente contagioso de enfermedad animal. Efectivamente, el primer patógeno para insectos, no para humanos. El hongo fue llamado más tarde “Beauveria bassim2 en honor de su descubridor. Son muy distintas y notables las momias blancas que afectan a las orugas llamadas muscardina, la cual es derivada de la palabra francesa bonbons la cual se parece a especímenes rnomificados.
Pnh;cin Elizabeth Baños Guevnro - 22

REVISIÓN DE LITERA'IWRA *
Actualmente el término muscardina se refiere a la enfermedad causada por hongos en insectos. Más tarde Balsamo Crivelli en 1935, ubicó al hongo en el género Botqvtis, nombrándolo Botrytis bassima. Posteriormente algunos investigadores observaron que el hongo presentaba características diferentes a Botrytis, estableciendo nuevas especies (Ferron, 1978).
Es el hongo más comúnmente aislado de insectos muertos y moribundos en la naturaleza, está ampliamente distribuido e infecta alrededor de 70 especies de insectoii. Las especies de B. bassima son cosmopolitas, entomopatógenos facultativos y tienen uno de los más amplios rangos de hospederos de cualquier entomopatógeno hyphomycete. Sin embargo, hay una amplia variación en especificidad entre aislamientos (Fargues, 1997).
5.2.2. Taxonomía y sinonimia
Los dos grupos m á s importantes de hongos son Deuteromycetes (órdenes: Moniliales y Sphaeropsidales) y Phycomycetes (órdenes: Blastocladiales, Chytridiales, y especialniente Entomopthorales).
Actualmente, el orden Moniliales está dividido dentro de cuatro familias debido al traslape de éstas en muchas características y a las similitudes en cultivo de muchas especies involucradas que de otra manera serían colocadas en dos o m á s de estas familias en vez de un género, todos los géneros asignados a este suborden están incluidos en una familia: la Modaceae. Este grupo y sus géneros presentan características generales tales como: hongos conidiales sin picnidios o acervuli. Conidióforos ausentes o bien desarrullados, separados, agrupados, o compactados dentro de sporocia o synnemata.
Las cuatro familias involucradas para definir el orden M o d a l e s tienen las siguientes características:
Familia: Modaceae, con conidia hialina y micelio; Familia: Dematiaceae, con conidia coloreada y/o conidióforos y micelio; Familia: Tubemlariaceae, con conidióforos agregados en esporodocios; Familia: Stilbellaceae, con conidióforos agregados en coremia o synnemata.
A su vez, los géneros que se encuentran dentro de la familia Moniliaceae son (Ferrun, 1978):
* Género: Beauveria (agente de la muscardina blanca) Especies: B. bassima
B. bmngniartii (B. tenella) * Género: Metmhizium (agente de la muscardina verde)
Especies: M. ffavo-viride M. anisopliae, variedad anisopliae y major
* Género: Paecilomyces Especies: P: fmnosus
?? fummo meus i? ameno meus
* Género: Akanthomyces Especies: A. gacilis
Género: Gibellula y Pseudogibellula
L - - Patricia Elizabeth Bañar Guevara 23

1
REMSIÓN DE LITERATURA
5.2.2.1. ClasiñcaciÓn taxonómica
Frazier (1976) menciona que los mohos son organismos del reino Fungi, que carecen de raíces, tallos y hojas; división Hongos, sin clorofila. Se encuadran en el tipo Eumicetos, hongos propiamente dichos y se subdividen de acuerdo a los criterios seguidos para la clasificación e identificación siguientes:
1. Que las hifas sean septadas o no
2. Que el micelio sea claro u obscuro (ahumado).
3. Que el micelio sea coloreado o incoloro
4. Que se produzcan esporas sexuales o no, y en caso de que se produzcan, de que tipo: oosporas, zigosporas o ascosporas.
5. Clase de esporas asexuales: esporangiosporas, conidios o artrusporas (oidios) .
6. Caracteres de las cabemelas de esporas a) Esporangios: tamaño, color, forma y localización. b) Conidios: conidios simples o en cadenas, conidios formados por gemación, o en
masas; forma y desposición de los estengmas o fiálides; apelmazamiento de los conidios.
7. Aspecto de los esporangióforos o conidióforos: simples o radicados, tipo de ramificación. Tamaño y forma de la columela apical del esporangióforo. Si los conidióforos están aislados o agrupados en haces.
8. Aspecto microsdpico de las esporas asexuales, especialmente de los conidios: forma, tamaño, color; lisos o rugosos; mono, bi o multicelulares.
9. Presencia de estructuras (o esporas) especiales: estolones, rizoides, células basales, apófisis, clamidosporas, esclerocios, etc.
La posición taxonómica del hongo B. bassima (Bals.) Vuill. citado por Madelin
División Amastigomycota (Eumycota) Subdivisión Deuterumycotina Clase Deuteromycetes (= Hyphomycetes) Orden Moniliales Familia Monilia c ea e Género Beau veria Especie bassima Nombre científico
Muchos aspectos biológicos de Beauveria bassima incluyendo el control de insectos plaga en la agricultura ha sido investigado mediante estudios que proveen información sobre la variabilidad genotipica de los marcadores genéticos, la distribución de los genotipos y sus frecuencias en las poblaciones, los hospederos, la agrupación o ramificación geográfica y la ploidia de cepas; y puede ser especialmente valorado cuando se utilice para la clasificación taxonómica de estos organismos (Grinstein et al., 1996).
(1963) es la siguiente:
Beauveria bassiana (Balsamo) Vuillemin, 191 1.
Patricia Elizabeth Baños Guevara 24

1
5.2.2.2. Sinonimia
Vuillemin en 19 12 c m el nuevo género de B. bassiana considerando la forma y tipo de conidióforo o simpodio; los esporangios fiiamentosos forman prolongaciones y la disposición de las esporas en Ug-zag hacen que Bofirtis bassima cambie a Beauveria bassiana (Balsamo) Vuillemin. Beauverie en 1914 concluye que Beauveria tiene cuatro especies, mencionándolos en forma cronológica, estas son (Ferrun, 1978):
Beauveria bmiana Beauveria effusa Beau veria densa Beau veria globulifera En 1954 MacLeod citado por Madelin (1963) realizó una exhaustiva revisión,
reduciendo a dos las 14 especies clasificadas en este género. Este autor concluye que Beauveria stephanoderes (Bally) Petch., Beauveria laxa Petch., Beauveria globulifera (Spega-) Picard, Beauveria effusa (Beauverie) Vuillemin, Beauveria dorphorae Poisson y Patay, Beauveria delacroixii (Saccardo) Petch., isaria vexans Pettit y Botrytis acridiorum Brugniart y Delacrok son razas de Beauveria bassiana (Balsamo) Vuillemin.
Mientras que Beauveria densa (Link) Picard, Beauveria brongni'ii (= Beauveria tenella) (Saccardo) Petch., Beauveria shiotae (Kuru) Langeron y Botrytis mefo/onfhae Saccardo, son razas de Beauveria tenelfa (Delacruk) Siemaszko; pero recientemente De Hoog incluyó a B. tenella en B. brongniartii.
(Balsamo) Vuillemin 191 1 (Beauverie) Vuillemin 19 1 1 (Link) Picard 1914 (Spega-) Picad 1914
Samson y Evans (1982), reportan dos nuevas especies de Beauveria en SudamCrica, Beauveria velata Nov. y Beauveria morpha.
5.2.2.3. Clave para identificación
B. bmiana se puede identificar según la clave descrita por Cooke (1963):
1. Esporas producidas simples o sucesivamente ................................................ 10 10. Esporas producidas en ramas hifales diferenciadas o no diferenciadas ...... 1 1 1 1 . Esporas producidas en ramas Wales diferenciadas ...................................... 23 23. Rama conidioforal produciendo una espora inmediatamente atras de
la punta, la cual continúa creciendo para producir esporas en una manera más o menos regular .......................................................................... 24
24. Las esporas nacen inmediatamente atrás de la punta apiculada en crecimiento ....................................................................................................... 25
25. Las esporas nacen en sucesión relativamente regular en la apiculación lateral ................................................................................................................ 27
27. Base de la rama conidioforal apiculada inflada, espacios cortos entre apiculados; conidias globosas o subglobosas de 2-3 x 2.0-2.5 nun, con conidióforo formado en racimos compactos.. .................................................................................. B. bassiana Conidias elipsoides de 2-3 x 1.5-2.5 mm, conidióforos escasos o raramente en racimos ................................................ B. bmngniarfii
. .
Patricia Elizabeth Baños Guevara 25

RFVISIÓN DE JJTERATLJRA
5.2.3. Morfología
La morfología, es decir la forma y estructura macro y microscópica de los hongos, sirve de base para su identificación y clasificación (Frazier, 1976).
En términos macroscópicos, 8. bassima se manifiesta en cultivo puro, a una edad de 2 1 días, por un crecimiento micelial blanco algodonoso, tiene una esponilación abundante de textura polvosa; cuando crece en el insecto a veces llega a envolverlo completamente, el crecimiento miceliar inicial es de color blanco, pero después puede cambiar a amarillento o rosado.
En términos microscópicos, el micelio se ramifica formando conidióforos simples, dilatados en la base, con un raquis alargado, denticulados, agrupados irregularmente o dispuestos en racimos verticiiados que pueden ser densos; la base del verticilo generalmente es más abultada sufriendo un adelgazamiento en el área donde se insertan los conidios, en la que se sitúan las esporas (simpodulosporas en sucesión acropétala) en forma de Ug-zag. Las colonias alcanzan de 0.6-2.3 cm de diámetro de ocho días a 20°C y son de apariencia polvosa, debido a la presencia de gran cantidad de conidios. Las conidias son hialinas, globosas o subglobosas, redondeados u ovales (2-3 x 2-2.5 mm); la cantidad de conidios aumenta con el transcurso del tiempo, de los 12 a los 15 días de edad la colonia presenta abundante esponilación de color crema (Samson, 1981 y McCoy et al., 1988, citados por Hernández, 1993).
El modo hifal de crecimiento proporciona a los hongos filamentosos mayor ventaja sobre los microorganismos unicelulares en la colonización de sustratos sólidos y la utilización de nutrientes aprovechables. El modo básico de crecimiento de los hongos filamentosos es la extensión apical de las hifas, además de la generación de nuevas puntas a través de ramificaciones. Las hifas pueden ser capaces de regular su frecuencia de ramificación en respuesta a la concentración del sustrato (Fdelstein y Segel, 1983 y Moo- Young et al., 1983, citados por Doelle et al., 1992).
5.2.4. Propáguios infectivos de B. bassiana
La reproducción asexual en hongos filamentosos resulta de un cambio en el equilibrio entre crecimiento vegetativo y esponilación. Este cambio puede deberse al estado nutritional del medio o factores ambientales tales como temperatura, aireación, pH, sales inorgánicas y luz. En 8. bassiana, la formación de esporas ha mostrado estar asociada con la actividad de agua, niveles de fosfato y, de acuerdo a datos, con carbono, nitrógeno y fosfato (Hegedus et al., 1990).
El hongo 8. bassima, en condiciones de campo, produce dos tipos de esporas: blastosporas en la hemolinfa, dentro del hospedero y conidias sobre la superficie del hospedero muerto; las diferencias se distinguen por la morfología y tamaño de ambas. Las blastosporas son relativamente largas, de pared lisa, usualmente de forma cilíndrica y tamaño irregular; son susceptibles a las condiciones ambientales y consecuentemente menos persistentes que las Conidias (Paaccola-Meirelles y Araujo, 1993; Aregger et al., 1988 citado por Aregger, 1992). Las conidias, en cambio, son pequeñas, esféricas, más uniformes en tamaño y a menudo completamente refráctiles o hialinas (Thomas et al., 1987).
- - Pahcia Elizabeth Bmim Guama 26

REVISIÓN DE i.iTERAlUR.4
B. bmiana puede producir tres tipos de esporas en condiciones de laboratorio: conidias aéreas, conidias sumergidas y blastosporas. Las características morfológicas y los análisis de proteínas proveen la evidencia de soporte para la distintividad de las tres esporas. La áspera superficie exterior de la conidia aérea puede proteger la espora de las condiciones ambientales. Las amidias sumergidas son producidas bajo condiciones limitadas de fosfato, y también muestran tales características de superficie aunque en menor grado (Thomas et al., 1987). Las blastosporas producidas en cultivos líquidos ricos en nutrientes no forman una capa exterior vemcosa que pueda ser observada por microscopio electrónico (Hegedus et al., 1990).
5.2.4.1. Fases de genninación a conidiación
Paccola-Meirelles y Araujo (1992) estudiaron y monitorearon el crecimiento de B. bmsiana de la fase conidial a la formación de micelio para determinar el mejor tiempo de cultivo necesario para la ocurrencia de la emergencia del tubo germinative y la primera división mitótica.
La primera etapa es una conidia uninucleada. En conformidad con los resultados obtenidos por B & & k a et al. (1987, citados por Paccola-Meirelles y Araujo, 1992), la conidia no tiende a agregarse. La duración de esta etapa depende del medio y finalizó en un promedio de 8 horas en varios medios de cultivo. La emergencia del tubo germinal (segunda etapa) ocurrió después de 8 a 16 horas. Aproximadamente del 42 a 43% de conidias emitieron un tubo germinal después de 8 horas en incubación en un Medio Completo y Medio Saboraud. Durante esta fase, el tubo germinal es unipolar y no aparece todavía septación.
Después de la emergencia del tubo germinal, la primera división nuclear ocurre, lo que caracteriza la tercera etapa.
ia cuarta etapa muestra la anafase y la telofase. El tiempo para la primera formación de septos varía entre cepas y entre medios.
La quinta etapa fue marcada por una segunda división nuclear y se inició después de 16 horas de incubación, se forma un núcleo de metafase.
En la sexta etapa, la formación del segundo septo ocurrió más frecuentemente después de 24 h de incubación, aunque después de 16 horas fue todavía posible observar esta figura.
En la kptima etapa existen ya ramificaciones Wales, formación de mnidios, migración nuclear, y una p f a s e del núcleo en el cual la cromatina puede observarse en la forma incondensada.
5.2.4.2. Formas de dispersión
La dispersión de los ppágulos infectivos de un nuevo hospedero representa una de las partes más peligrosas en el ciclo de vida del hongo. Los procesos de producción de esporas y acumulación, la dispersión de esporas, la sobrevivencia de la espora y su genninación frecuentemente dependen de condiciones ambientales (Hajek y Leger, 1994).
- _= Paíricia Elizabeth Baños Guevara 27

REVISIÓN DE LITERATURA
Femn (1978) también asevera la existencia de una correlación positiva entre el número de esporas infectivas y la mortalidad por micosis, pero aún no se ha demostrado que la influencia de bajas dosis no conduzca a un desarrollo letal de la enfermedad. La estimación de la LDw varía de acuerdo a la cepa del hongo y especies de un insecto dado y los modos de contaminación: por aplicación tópica del inóculo, por rociado de una suspensión de esporas directamente sobre el insecto o por plantas tratadas o sobre sustrato inerte en donde los insectos son colocados, por libre o f m d a ingestión, por contaminación de sustratos con crianza, particularmente para insectos subterráneos, por inmersión de insectos en una suspensión de esporas de concentración conocida. En términos generales, la LDw fue baja cuando la técnica usada por contaminación fue seguida por un contacto directo entre el insecto y el patógeno, lo cual ocurre generalmente con insectos aéreos.
En contraste al usar la técnica de contaminación indkcta especialmente para insectos subterráneos, como Melolontha meloiontha, se obtuvieron resultados satisfactorios con un inóculo 10 veces mayor que para insectos aéreos, conteniendo cerca de 3 x 1014 conidias de B. brongniarfii por ha. En el caso particular de un insecto de tamaño pequeño con una vida subterránea corta, se usó un inóculo más alto de B. bassima o M. anisopliae, con cerca de 107-108 conidios por cms de suelo o aproximadamente 10I6 o 1017 esporadha, considerando que sólo 10 cm de la capa superficial del suelo puede ser contaminada.
5.2.5. Mecanismos de acción
5.2.5.1. Modos de infección a insectos
Difinendo de las bacterias y virus, los hongos pueden infectar no sólo el intestino completo de insectos sino también toda la superficie del integumento; esta propiedad conduce directamente a la posibilidad teórica de infectarlos independientemente de su actividad alimenticia (Ferrun, 1978).
Madeiin (1963) menciona que existen varias formas por medio de las cuales la unidad infectiva de los hongos entomopatógenos, que es la espora o conidio, puede entrar al cuerpo de un insecto. Dentro de las principales formas de invasión considera las siguientes:
a) Vía oral; el organismo entra al insecto junto con el alimento, esto sucede por ejemplo con los virus, rickettsias, nemátodos, pmtozoarios y posiblemente algunos hongos.
b) A través del integumento íntegro o tráquea; esta es la vía más común que utilizan los hongos y algunos nemátodos, ya que la conidiospora puede emitir un tubo germinativo e invadir a través de la cuticula del msedo.
c) Por el tract0 digestivo. B. bassima es capaz de producir patogénesis cuando penetra a través de la p a d del intestino.
d) Vía parenteral o a través de las áreas intersepentales; cuando ocurre un trauma en el integurnento o existen heridas (Madelin, 1963).
e) A través del huevo; como en el caso de los virus, rickettsias, espimquetas y pmtozoarios.
- Pahlcia EJizabeth Bañar Gumwa 29

5.2.5.2. Micosis
Todas las etapas de desarrullo del insecto: huevo, larva, pupa e insecto adulto, son generalmente susceptibles a micosis. Existen datos contradictorios con respecto a la susceptibilidad de huevos, pero esto frecuentemente concierne una etapa de corta duración que después manifiesta la enfermedad. Este fenómeno de aplazar la mortalidad es muy frecuente en el caso de contaminación de hongos (Fargues, 1973, citado por Ferrun, 1978).
El ciclo biológico de 8. bassima comprende dos etapas de invasión: una patogénica y otra saprofítica. La fase de patogénesis ocurre cuando el hongo entra en contacto con el tejido vivo del hospedero y la humedad en el micmlima es del 85% o m á s (Bauved, 1997). Estas fases se pueden apreciar de mejor manera en las figura I.
5.2.5.2.1. Condiciones favorables para el desarrollo de una micosis
Se acepta, en general que todos los estados de desarrullo de los insectos son susceptibles a micosis, sin embargo es necesario tomar en cuenta que no todos los insectos son susceptibles al mismo hongo, ya que los diferentes hongos son caracterizados por cierta especificidad. Para que se ocurra una micosis es necesario que exista una cepa virulenta, un insecto hospedero susceptible, dosis correcta de esporas infectivas, estado fisiológico del huésped y las condiciones abióticas favorables. Los hongos entomopatógenos característicamente penetran su insecto hospedero hasta el integument0 en contraste con las bacterias y viruses que sólo entran por la vía del tract0 alimentario. La principal importancia es atribuida por ello, a la humedad relativa del ambiente atmosférico el cual puede controlar el desamllo del proceso infectivo del hongo imperfecto (Müller-Kogler, 1965, citado por Femn, 1977).
5.2.5.2.2. Desarrollo de la micosis
B. bassiana es un parásito facultativo, el cual posee conidias que constituyen la unidad infectiva del hongo. Con base en observaciones y en datos publicados por Ferron (1977) y Bauved (1997)., se han formulado hipótesis sobre las etapas del p r o p s o de una infección fungal
a) La primera fase de genninación de esporas y penetración de hifas al cuerpo del hospedero dura de 3 a 4 días. La penetración del hongo al hospedero ocurre a través de la cutícula o por vía oral. Cuando la penetración se da por la cutícula intervienen lipasas, quitinasas y proteasas. El tubo germinativo de la conidia invade directamente, produciendo apresorios que penetran la epicutícula, dando lugar a cuerpos hifales, los cuales se desarrullan en el hemocelo y circulan en la hemolinfa.
La patogenicidad del hongo sobre los insectos depende de una compleja relación entre la habilidad del hongo para penetrar la cutícula y la fortaleza del sistema kmunológico del insecto para prevenir el desarrollo del hongo. Esta relación se debe a factores muy concretos incluidos las diferencias cuticulares, la penetración cuticular y las reacciones inmunes. El desarrollo del hongo sobre el insecto puede ser influenciado por la eficacia de los hemocitos en encapsular y melanizar al patógeno.
- - Paíricia Elizabeth Bañar Guevara 30
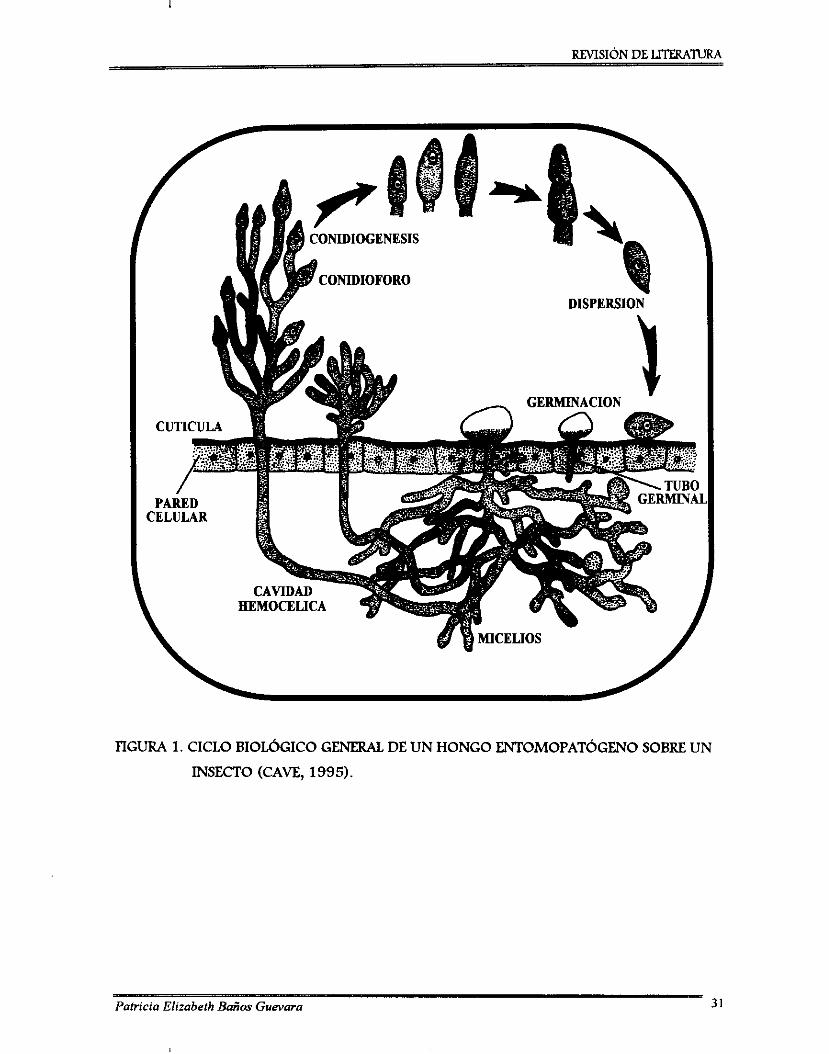
PARED CELULAR
@ 1 MICELIOS
FIGURA 1. CICLO BIOLÓGICO GENERAL DE UN HONGO ENTOMOPATÓGENO SOBRE UN INSECTO (CAVE, 1995).
- _c_
Patricia Elizabeth Bañar Guevara 31

&.si siempre los hematocitos se agregan al lugar de la penetración cuticular, formando algunas veces nódulos alrededor de las esporas inyectadas. En el interior de los insectos la genninación usualmente procede de esporas que están fuera de la agregación de hematocitos pero para que se desarrollen siempre deben de estar afuera del agregado.
b) La segunda fase es la reproductiva y saprúfita del hongo identificadas por invasión de los tejidos por parte del miceiio del hongo hasta causar la muerte del insecto, dura de 2 a 3 dias. Durante el proceso de invasión del hongo se producen una gran variedad de metaboiitos tóxicos. Los síntomas de la enfermedad en el insecto son la pérdida de sensibilidad, incordlliación de movimientos y parálisis. La muerte de los insectos marca el fin de la fase parasítica del desarrollo de los hongos.cuando el insecto queda momificado algunas veces se pueden presentar zonas de pigmentación localizadas que corresponden a los sitios de penetración de las conidias en el tegumento. Esta fase ocum independientemente de la humedad relativa.
Es importante recordar que el integument0 del insecto está compuesto esencialmente de proteínas y quitinas asociadas con iípidos y compuestos fenóiicos; la capa fina exterior o epicuticula contiene iípidos (ácidos grasos y parafina) cuya actividad antifungal ha sido demostrada, por la alta concentración que presentan en el integument0 del insecto. Una vez que la epicuticula es rota, prosigue la penetración del ápice hifal hasta la cutícula, la cual puede ser en mayor o en menor vía penetrada por la hifa, o invadida por estructuras muy extendidas lateralmente, produciendo una penetrante cuña. Estas expansiones laterales pueden causar fracturas a favor de la penetración y puede facilitar la dispersión de las enzimas del patógeno que degradan la cuticula.
La prucuticula actúa corno una barrera física por su irnpermeabilidad a las secreciones del patógeno, su resistencia a degradación por muchas enzimas patógenas, y su resistencia mecánica a penetración. El grado de resistencia depende de los efectos de combinación de la capa de la cuticula, la fue= de tensión impartida a la cuticula por el sistema de quitina laminar, y el grado de endurecimiento de la cuticula por esclerotización. insectos con segmentos de cuerpo fuertemente esclerotizados son usualmente invadidos vía membranas artrodiales.
Ya que el hongo invade el hemocele, el hospedero puede ser aniquilado por la combinación de daños mecánicos producidos por el crecimiento del hongo, agotamiento de nutrientes o por toxemia. La importancia relativa de estos mecanismos varía con el aislamiento del hongo y el hospedero. A las 48 horas de ocurrida la invasión, el hongo circula en la hemolinfa invadiendo toda la cavidad de la larva en forma de cuerpos hifales, generalmente las células sanguíneas no pueden detener la invasión masiva del hongo, ya que éste se multiplica rápidamente. Las hifas parecen tener preferencia por el tejido adiposo, pero también invaden el sistema muscular y nervioso (Hajek y Leger, 1994).
Wuif (1983) observó que el hongo B. b a s h a penetra a través de la cutícula del barrenador de la corteza Pityogenes chafcogaphus e infesta músculos y tejidos grasos. El cuerpo del insecto se infecta con hifas saprofíticas que causan la muerte de la plaga y el micelio emerge de la superficie del cuerpo.
PL
Patricia Elizabeth Baíim Gumma 32

1
RMSIÓN DE LITERATURA
c) Finalmente, en la tercera fase ocurre la esponilación y el inicio de un nuevo ciclo. El micelio del hongo se observa primero en las articulaciones y partes blandas de los insectos y en días posteriores se incrementa a todo el cuerpo hasta finalmente cubrirlo. Tras la muerte del insecto y bajo unas condiciones de humedad relativa alta (cercana al punto de saturación) las conidiosporas pueden extenderse a través del cuerpo cubriéndolo con material fungoso característico. Estas fases se aprecian de mejor manera en la figura 2.
5.2.5.3. Periodo de infección letal
5.2.5.3.1. Defensas del hospedero despues de la infección
El éxito de una infección depende del potencial genético del patógeno a crecer rápidamente, para penetrar las barreras inducidas por el hospedero, y la resistencia a químicos tóxicos. Estas cualidades pueden ser relacionadas a la fuerza inicial de las defensas constitutivas del hospedero y a la velocidad o magnitud de sus respuestas inducibles. La inmunidad de insectos es un área de investigación inexplorada. Antes se conocía que las propiedades de un hongo patógeno necesitan ser genéticamente provistas para control de insectos, la mayoría de esos factores son responsables de la susceptibilidad o resistencia del insecto al patógeno.
La melanización cuticular, inducida por daiio físico o p- 1,3 glucanos sobre la pared celular del hongo, es común. La frecuente melanización ocurre demasiado tarde, o en poca magnitud, detiene fuertemente o acelera el crecimiento de patógenos, pero determina el rol de las respuestas melánicas en la resistencia a la infección y es difícil debido al insuficiente conocimiento de los caminos en los cuales las reacciones melánicas y su cantidad pueden impedir el crecimiento del hongo. Si el micelio penetra todo el integument0 y puede ser favorecida por las esporas, entonces tal vez se involucran ambos mecanismos: físicos y enzimáticos; con o sin el desamlio de estructuras apresoriales. Esta penetración es frecuentemente seguida por una reacción melánica del integument0 a los sitios de infección, probablemente debido a los cambios en la actividad fenoloxidasa causada por el hongo.
La principal reacción hemocítica en el mecanismo de defensa celular antifungal es la encapsulación del hongo, el cual es rápidamente melanizado. Durante la encapsulación los granulocitos están atacando al hongo y pueden sumergirse (fagocitosis), entonces los plasmocitos son agrupados y forman un pseudotejido en capas concéntricas formando un granuloma (nódulo) en el cual el hongo puede ser lisado. La encapsulación sólo priovee protección para el hospedero contra patógenos débilmente virulentos. En el caso de cepas hipervirulentas de especies de hongos, los hospederos son incapaces de formar granulomas tipicos o los hongos rompen la encapsulación y continúan creciendo.
Por ejemplo, el exitoso crecimento de 8. bmima dentro de los hospederos está basado sobre la simple destrucción del hemocito de respuesta del hospedero; los númems de granulocitos son dramáticamente reducidos 3 días después del desafio. Se ha propuesto que la respuesta de defensa celular es el blanco inicial y las respuestas a fagocitosis persistan funcionales. Por parte del hongo se desamlla un mecanismo que tiende a frenar la reacción hemocítica; éste consiste en la producción de sustancias tóxicas o micotoxinas que hacen que el movimiento de los hemocitos hacia los cuerpos hifales disminuye y, por tanto, que la formación de granulomas se detenga hasta la muerte del insecto, inclusive antes de que el hongo hubiera colonizado el interior del insecto (Hajek y Leger, 1994).
Patricia Elizabeth Baños Guevara 33
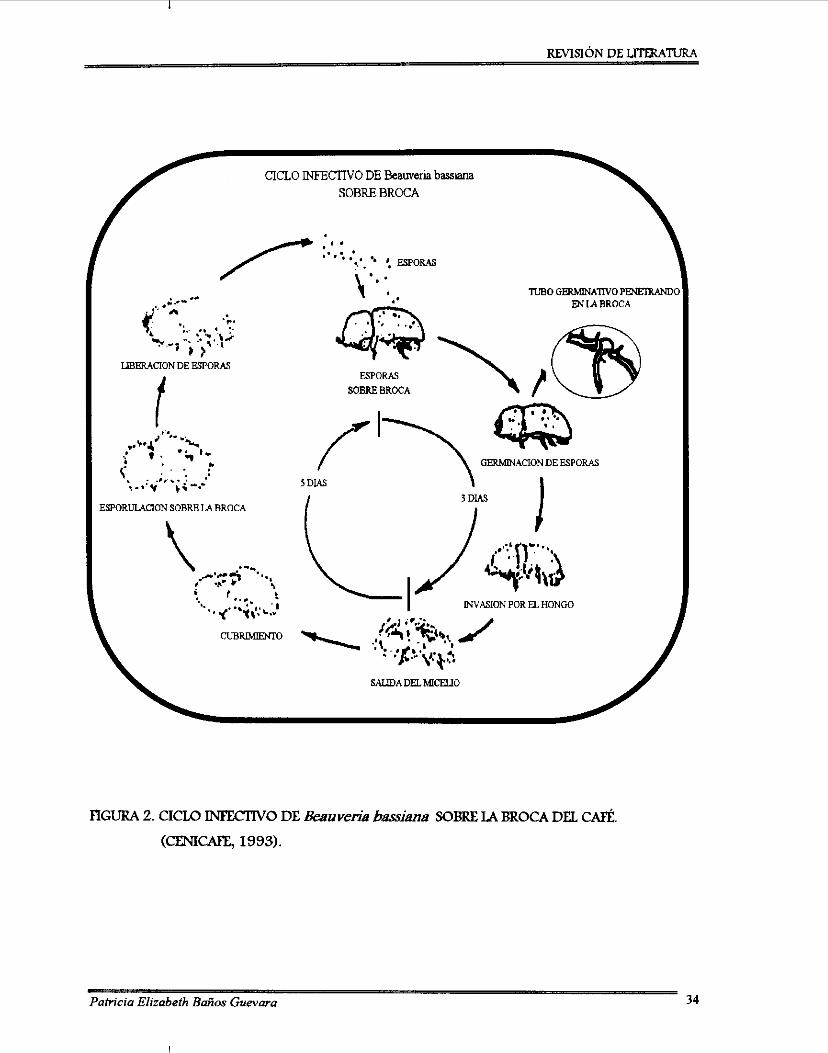
CICLO iNFECIlv0 DE &WCM bassiana SOBRE BROCA
-.L I .
i 5 m C I O N DE ESPORA3
'4,
c. . .;; . . . . * -. . . . . .
j ' t a,-
..*'& ,, .! * . ...'" ,<-.-
ESPORUWCION SOBRELABROCA
.
ESPORAS SOBRE BROCA
/ \ GERMNACIONDEESPORAS
5 DIAS 3 L S J I
FiGURA 2. CICLO I " 0 DE Beauveria bassiana SOBRE LA BROCA DEL C&.
(CENICAFE, 1993).
I CUBRIMlprrO
SALDA DEL MICEllO
Patricia Elizabeth Bañar Guevma 34

REVISIÓN DE LiTEXWi'üR.4
Una defensa biológica que presentan los artrópodos para rechazar la infección de los hongos entomopatógenos son los períodos de muda, por lo que la infección puede ser considerada exitosa únicamente cuando el hongo ha alcanzado la hipodermis, por lo que el proceso infeccioso puede variar como resultado de la mincidencia entre la penetración fungosa y el estado de muda. Este fenómeno es igualmente importante si el insecto en consideración sufre frecuentes mudas. En efecto, este proceso infeccioso puede variar mmo resultado de la coincidencia entre la penetración fungal y la época de muda. Por ejemplo, B. bassima puede atravesar todo el integumento del tercer instar larval de Lepfinofmsa decemlineata justo antes de mudar. En este momento la hifa y las blastosporas invaden el fluido exuvial e infectan el nuevo integumento formado, lo cual causa lisis localizadas de la hipodermis por el usual proceso de infección primaria de larvas entre mudas. Estas heridas son abiertas no sólo a elementos miceliales del fluido exuvial, sino también a bacterias que pueden causar septicemia letal.
De este modo, si la penetración dentro del integumento viejo es sólo superficial durante la muda, el insecto hospedero puede escapar a la infección al rechazar el inóculo infeccioso. Así B. bassima puede pasar a través del integumento de los instares larvarios justo antes de mudar, ya que la hifa y las blastosporas invaden el fluido exuvial e infectan nuevamente el integument0 formado, el cual causa una lisis localizada de la hipodermis por el proceso usual de infección primaria de larvas entre mudas, no obstante, si la penetración al integumento es Únicamente superficial durante la muda, el insecto puede escapar de la infección por rechazo al inóculo (Ferron, 1978).
5.2.5.3.2. Cambios de comportamiento de hospederos infectados letalmente
En muchos sistemas, una reducción en la alimentación es uno de los primeros cambios que ocurren en un hospedero infectado. Esta respuesta provee una constante vigilancia de los beneficios de los hongos patógenos infectando insectos plaga. Áfidos, hormigas, defoliadores de hierbas y plantas, y las moscas infectadas con diversos hongos son insectos conocidos por trasladarse a lugares lejanos antes de morir.
Es sabido que muchos hospederos mueren en posiciones con alas o élitros esparcidos lejos del cuerpo o con partes del cuerpo separadas del sustrato exponiendo las Areas esporuladas del cadáver. La mortalidad de los hospederos en lugares elevados 15 en posiciones expuestas podría claramente mejorar la dispersión de esporas. Todavía no son bien entendidos los comportamientos antes de morir, presiones selectivas sobre los hongos pueden tener evolución favorable de las interacciones hospedero-patógeno que promuevan definitivamente la dispersión de esporas (Hajek y Leger, 1994).
5.2.5.3.3. Desarrollo sapmfitico del hongo
Aun cuando una posible fase saprúfita no ha sido elucidada, B. bassima está comúnmente en suelos de todo el mundo y por consiguiente puede adaptar su crecimiento en dos hábitats completamente diferentes, suelo e insecto hospedero. Ciertamente el hongo ha sido observando creciendo sobre cadáveres de insectos en el suelo.
Paticia Elizabeth Baños Guevara 35

REVISIÓN DE !JlERATiJRA
Cuando el insecto hospedero muere, el hongo procede a crecer saprofíticamente invadiendo el interior del cadáver y se extiende virtualmente por todos los tejidos del insecto hasta colonizarlo completamente. La competencia entre los hongos y las bacterias de la flora intestinal ocurre. Finalmente emerge y, si la humedad relativa ambiental es alta, esporula (atmósfera saturada), sino permanece inactivo hasta que se presenten las condiciones adecuadas para esporular y completar su ciclo. Sin embargo, en la mayoría de los casos el cadáver es transformado en una momia resistente al deterioro bacterial aparentemente debido a antibióticos producidos por el hongo.
En aquellos lugares en donde l as esporas germinan la cuticula toma un wlor amarillento; después la parte exterior de la cuticula es m á s o menos destruida, entonces el área de infección se torna de un color pardo oscuro. Estas áreas aumentan de tamaño a medida que la infección progresa.
Por ello, los aislamientos adaptados a hospederos termófilos o a c h a s cálidos pueden también necesitar estar adaptados a continuar un crecimiento saprófito o sobrevivir bajo condiciones más frías, como se encontraron algunos aislamientos de acrididos. Por instancia, un aislamiento adaptado a un hospedem acridido puede también beneficiarse por crecer en el huésped muerto, el cual no podría termorregular largamente o saprófitarnente en el suelo, usualmente bajo muchas condiciones frías igual bajo condiciones tropicales, por ejemplo, en la noche (Fargues et al., 1997).
5.2.6. Producción de metaboiitos
En la mayoría de los casos, los metabolitos fúngicos asisten al patógeno con (Hajek y Leger, 1994):
a) Aspectos físicos de entrada o ingreso, ej. enzimas de degradación de la cutícula que activan la destrucción o modifican la integridad estructural del hospedero.
b) inhibición de procesos selectivos o de enzimas del hospedero.
c) interferencia con los sistemas regulatorios del hospedero
5.2.6.1. Producción de enzimas
indudablemente, muchas enzimas patógenas son importantes en la determinación de la virulencia debido a la capacidad de los patógenos a coexistir con el cambio de los procesos metabóiicos asociados con el estado de infección del hospedero. Todavía se tiene poca información acerca de cómo son asumidos los roles de una enzima particular o una toxina. El grueso de los disturbios fisiológicos asociados con los procesos de infección hace difícil distinguir entre el sitio inicial de la función perturbada y los eventos secundarios o falsos. Las técnicas de recombinación del Ácido Desoxiribonucléico (DNA) por aislamiento de genes específicos son, no obstante, un avance del conocimiento en este ámbito (Hajek y kger , 1994).
Patricia Elizabeth Baños Guevara 36

REVISIÓN DE LITERATURA
Se ha demostrado que los hongos entomopatógenos como B. bassima, B. brongnihrTii, M. anisopiiae, Cordyceps militark, Aspergilius flaws, y Lhtomophthora muscae producen varias clases de enzimas (Huber, 1958, citado por Leopold y Samskáková, 1970). Samsináková et al. (1971) mencionan que el hongo entomopatógeno B. bassima secreta dentro del medio de cultivo, ya sea sólido o líquido, varias enzimas: celulasas, lipasas, proteasas y quitinasas.
Los efectos destructivos de las proteasas del patógeno sobre la cuticula pueden ser atribuidos, al menos en parte, a la importancia estructural y accesibilidad enzimática de polímeros de proteína en la cuticula. Sin embargo, las fibras de quitina también participan como una barrera mecánica a la penetración y con un estabilizante de la matriz proteínica de la cuticula, evidenciado por el efecto sinérgico de las aplicaciones duales de DllnilUi, un inhibidor de la síntesis de quitina (Hajek y Leger, 1994).
La originalidad del equipo enzimático de los hongos entomopatógenos es debida a la capacidad de ataque al complejo proteo-quitina. Se ha estimado que sólo el 10% del total de proteínas del integument0 no están unidas. Por tanto, es necesario que tome lugar una hidrúlisis de proteínas antes de iniciar la acción de las quitinasas. Es sabido que en adición a las lipasas, proteasas, amilasas y quitinasas, que son efectivamente producidas por estos hongos, la actividad lipolítica y proteolítica precede el desdoblamiento de la quitina (Samsináková et al., 1971).
Kucera y Samsinakova (1968, citado por Kucera, 1971) detectaron que el hongo entomopatógeno B. bassiana produce componentes tóxicos de alto peso molecular los males se correlacionaron con su actividad proteolítica, este hongo pertenece al grupo de micrwrganismos en los cuales la producción proteolítica es estimulada por fuentes nitrogenadas de complejo natural.
Bidochka y Kachatourians (1987) señalan que el hongo entomopatógeno B. bassiana produce una proteasa extracelular con azocol como sustrato, la actividad de la proteasa fue óptima con un pH de 8.5, lo que indica que se puede considerar corno una proteasa alcaiina.
La temperatura óptima de actividad proteásica se encuentra entre 27 y 42OC. h proteasa fue estable a 27T, pero se inactiva a altas temperaturas. Aproximadamente 40% de la actividad permanece después de 30 minutos a 40° o después de 5 minutos a 50°C y a 60°C la proteasa fue rápidamente inactivada a los 5 minutos. Dicha p t e a s a de B. bassiana hidroliza la caseina, la gelatina, la elastina y en menor proporción al suero albúmina de bovino (Leopold y Samsináková, 1970).
El modo primario de infección de este hongo es transcuticular; se ha establecido que produce una mezcla de e-as hidrolíticas extracelulares, quitinasa, N-acetyl-D- glucosa-dasa y posiblemente lipasa u otras esterasas (Bidochka y Khachatourians, 1993) que contribuyen a la solubilización de la cuticula del insecto y la sucesiva penetración de la hifa fungal. En estudios específicos se han investigado principalmente las relaciones cuantitativas de la enzima quitinasa en ü. bassima, la cual es una de las m á s conocidas y éste el hongo entomopatógeno más común. Es un poiífago, destruye muchos tipos de insectos, y no está limitado a un hospedero específico; es un hongo capaz de penetrar igualmente a través de la superficie del cuerpo del organismo hospedero en la estructura, de la cual no participa la quitina (Cermáková y Samsináková, 1960, citados por Leopold y Samsináková, 1970).
P
Patricia Elizabeth Baños Guevara 37

1
REVISIÓN DE LlTEüAlUlZA
La epicuticula de los insectos es multiestratificada y cada capa tiene sus propiedades. El exterior de la epicuticula parece, en muchos insectos, ser mecánicamente frágil y puede ser penetrada con poca fuerza. su resistencia a la degradación enzimática y su impermeabiiidad sugieren que puede prevenir la entrada de las enzimas fúngicas que degradan la cuticula. El interior de la epicuticula está compuesta por polhero:; de lipopmteína y quinonas, composición que a pesar de ser rígida es degradada por las e m h a s producidas por los entomopatógenos (Hajek y Leger, 1994).
Cuando un insecto es atacado por hongos entomopatógenos, el parásito está asumiendo tener invadido el cuerpo de su hospedero para producir suficientes enzimas para así desintegrar la superficie del cuerpo de sus víctimas. Debido, en la mayoría de casos a que la penetración al caparadn quitinoso de los insectos es complicada, la atención se centra principalmente en la habilidad del parásito para producir la enzima quitinas. La cepa del hongo B. bassiana forma quitinasa, en cultivo sumergido y estacionario, y la libera al ambiente, lo cual significa que es una enzima extracelular, se asume que la formación de esta enzima toma lugar principalmente en el vástago de la conidia germinada, en la punta y findeiahifa.
Se cultivó una cepa del hongo entomopatógeno bajo condiciones estacionarias y sumergidas. En el micelio de cultivos estacionarios de edad variable y en el medio de cultivo sumergido de 3 días de edad, se descubrió la presencia de quitinasa, celulasa, proteasa y lipasa. B. basiana es capaz de liberar estas enzimas dentro del medio. Las protemas degradan peptona y gelatina pero no caseina. La más alta actividad de quitinasa y celulasa es encontrada en 4 o 5 días de crecido el micelio, mientras la máxima cantidad de protea,sas y lipasas son formadas después de 6 o 7 días del periodo de cultivo del hongo (Leopnld y Samsináková, 1370).
Duriez-Vaucelle et al. (1381) realizaron estudios electroforéticos para detenrkar los procesos enzimáticos de B. bassiana y determinaron veintitrés actividades enzimáticas intracelulares y exocelulares y cinco oxidoreductasas. Este hongo presenta dos tipos de actividades enzimáticas: las óxido-reductasas, como la glucosa 6-fosfato deshidmgenasa, la monofenol-monooxigenasa y la catalasa. Las otras enzimas se encuentran en el grupo de las hidmlasas dentro de l as cuales se mencionan las esterasas, desoxirribonucleasas, ribonucleasas, P-glucosidasas, B-glucoronidasas, quimiotripAa A y otras proteasas. Pearson et al. (1984) señala que las peptidasas son altamente específicas hacia varios péptidos cromogénicos sintéticos y fueron encontradas en el micelio de B. basiana después de atacar arlrópodos.
5.2.6.2. Producción de toxinas
Una vez que el hongo invadió el hemocele (aunque también cuando se propagan en medios de cultivo), el hospedero puede morir por alguna combinación de daño mecánico producido por el crecimiento del hongo, término de nutrientes y por la capacidad de los hongos de producir toxicosis. La importancia relativa de estos mecanismos vana con el aislamiento especifico del hongo o por el hospedero. Muchos hongos entomopatógenos producen toxinas que pueden matar al insecto, incluso antes de ser invadido. Tales daños, asociados con síntomas de infección, pueden ser producidos por enzimas del patógeno y por sus metabolitos de bajo peso molecular, toxinas (Hajek y Leger, 1994).
__PI
Paíricia Elizabeth Bañar Guevara 38

I
Después de atravesar la cutícula el hongo es afectado por las defensas de reacción celular del hospedero. Los plasmatocitos, normalmente dispersados en la hemolinfa, se acumulan alrededor del hongo, convirtiéndose en lamelas (granuloma). Pocos días después, estas formaciones hemocíticas pueden ser grandes y visibles, el grado de melanización está relacionado con el grado de virulencia de los hongos, en B. bassima la produccióii de toxinas, favorece la recuperación del micelio el cual reinicia su crecimiento normal e invade el hemocele por medio de las blastosporas, por ello el papel de las toxinas es considerado muy importante (Ferron, 1978).
habilidad y patogenicidad para vencer los mecanismos de defensa del hospedero, se debe en parte a la producción de toxinas, B. bassima produce el depsipéptido cíclico beauvericin, reportado como tóxico a larvas de mosquitos y adultos de mosca doméstica, pero no tóxico a larvas de lepidóptera, además no todos los aislamientos producen beauvericin, otros depsipéptidos cíclicos, incluyendo los beauverolides, bassianolides e isarolides son producidos también. El “Beauvericin” es un depsipéptido que fue separado del micelio de B. bassima, el cual contiene tres unidades, cada una consiste de (l)-N- metilfenilalanina y (d) -a-hidroxiácido isovalérico en un a d o macrucíclico (Lysenko y Kucera, 1971, citado por Méndez, 1990).
West y Briggs (1968) produjeron toxinas in vitro con cepas de B. bassima, mismas que fueron evaluadas mediante inyección en larvas de la palomilla Galleria mellonela. Se observó que las cepas provenientes de insectos produjeron una mayor cantidad de toxinas que aquellas cepas cultivadas continuamente sobre medio artificial; esto indica que el crecimiento reciente del hongo sobre insectos vivos, estimula la producción de toxinas.
Rojas (1982b) evaluó el efecto de la ingestión de toxinas producidas por B. bassma sobre la mortalidad de adultos y larvas de la pulga saltona de la papa Epirix spp. La máxima mortalidad de adultos almentados con hojas de para tratadas con la solución de toxinas se observó a los 4 días y larvas alimentadas con raiciiias con toxina murieron a los dos días.
Mediante la administración de las toxinas de B. bassima por via oral y por inyección en larvas de Ostrinia furnacalis se obtuvo que la inyección de las toxinas fue más efectiva que la administrada por la vía oral, asimismo se menciona que las toxinas aisladas del medio de cultivo produjeron mejor efecto que las aisladas del polvo de esporas y que la actividad de las toxinas puede mantenerse por varios años (Cheng y Xing, 1984).
5.2.6.3. Producción de ácidos metabólicos
La habilidad para hiperproducir ácidos metabólicos depende de la presencia de una fuente de carbono utilizable tal como glucosa, glicerol o trehalosa. La producción de acido metabólico en la presencia de glicerol por mutantes de B. bassima indicó que la biosíntesis de ácido puede proseguir s i n involucrar al ciclo del ácido tricarboxíiico (TCA). Sin embargo, una vez dentro del hemocele, la trehalosa y glucosa contenidas en la hemolinfa pueden ser metabolizadas y producir ácido oxálico que consecuentemente acidifica el medio de la hemolinfa y wntribuye a la mortalidad del insecto. En mutantes de B. bassima y otros hongos, la pmducción de ácido oxáíico ocurre en la presencia de ciertos carbohidratos. El estado nutricional de la cutícula del insecto influencia la fisiología del hongo y puede influenciar la producción de ácido oxálico (Bidochka y Khachatourians, 1993).
P
Patricia Elizabeth Baños Guevara 39

También se ha observado que B. bassiana produce ácido cítrico y ácido oxálico en cultivos líquidos conteniendo cuticula de minadores y que estos componentes son capaces de solubiiizar péptidos y proteínas de la cuticula de minador (Bidochka y Khachatourians, 1991 citado por Bidochka y Khachatourians, 1993). Posteriormente, los &adores pretratados con una solución ímM de ácido oxálico o cítrico mostró una respuesta con mortalidad sinérgica con infección de este hongo. La penetración de la cuticula del minador por germinación de B. bmima fue observada m á s frecuentemente cuando las cuticulas han sido pretratadas con ácido oxálico o ácido cítrico.
5.2.6.4. Producción de bactericidas y pigmentos
Los síntomas característicos que aparecen inmediatamente después de la muerte consisten de momificación de cadáveres, seguidas por la salida del micelio en crecimiento y la esporulación del hongo. En el caso de B. bassima, el cuerpo del micelio frecuentemente exhibe una coloración rosa debido a la producción de oosporeina por el micelio interno y blastosporas (Ferrun, 1981). Este pigmento rojo es responsable del cambio de color de los cadáveres, tiene una actividad bactericida.
En adición a oosporeina, se descubrieron dos pigmentos amarillos llamados bassianin y tenellin; su estructura y biosíntesis han sido objeto de estudio por muchos investigadores (Marcandier y Khachatourians, 1987).
El micelio de B. bassima produce la toxina bassianolide que causa alteración en el núcleo de las células y puede funcionar como un antibiótico, con actividad antimicrobiana, al igual que el beauvericin al prevenir la invasión de bacterias, lo que hace que exista una parálisis y momificación del insecto hospedero (Ferrun, 1977).
Rojas (1982a) observó que la ingestión de la toxina segregada por el micelio del hongo Beauveria sp. produjo en los adultos de la pulga saltona de la papa, Epiria sp., falta de apetito, inactividad y parálisis del insecto. En larvas ocasiona ennegrecimiento, hinchamiento y pérdida de peso; la mortalidad en los adultos se observó a los 4 días después de haber ingerido la toxina, mientras que las larvas mueren a los 2 días.
Smith y Grulla (1981, citados por Méndez, 1990) mencionan que el ácido caprílico está presente en larvas del gusano elotero, Heliothis zea, y del gusano soldado, Swoptera frugijxrdi, también observaron que el ácido caprííico inhibe la genninación de B. bassima. Otros ácidos grasos fibres, están presentes en las larvas de H zea y S. frugijxrda y se identificaron como los ácidos valérico y nonanóico que también poseen una actividad micostática hacia B. bassima.
5.2.7. Efecto de agroquímicos sobre B. bassiana
La agricultura es una actividad en la que frecuentemente se hace uso de productos químicos, ya sea insecticidas, fungicidas, herbicidas, etc. Sin embargo, algunos de ellos afectan la reproducción o el establecimiento de los enemigos naturales de las plagas agrícolas, por lo que es necesario conocer su efecto, en este caso sobre B. bassiana (Rosin, 1996).
- - Paíricia Elizabeth Bañar Guevma 40

5.2.7.1. Fertilizantes
Este hongo entomopatógeno puede persistir en suelo por largos penodos. La persistencia de la conidia, de cualquier modo, puede ser reducida por varios factores abióticos, incluyendo componentes nitrogenados.
Se condujo una investigación para evaluar los efectos del contenido de nitrógeno de los fertilizantes sobre la persistencia de B. bassima en suelo. Los fertilizantes nitrogenados fueron: (I) estiércol fresco de vaca, (2) composta y (3) urea (45-0-0).
El suelo que contenia estiércol fresco fue detrimental para B. bassima, mientras que altos rangos de estiércol composteado fue benéfico. La urea no tuvo un efecto significativo sobre el hongo comparado con los testigos. Por lo tanto, parece que hay ciertos factores en el estiércol que reducen la sobrevivencia de B. bassima, los cuales se pierden durante la descomposición. El incremento en unidades formadoras de colonias (CFUs) en suelo modificado con composta relativo a las CFUs del testigo puede ser explicado por la habilidad de B. bassima a existir como saprófito. Tal vez altos rangos de composta pueden incrementar la eficacia del hongo como un agente de control biológico. El material composteado, por tanto, es aceptable por tener bajo impacto sobre la microflora que otros modificados. Los efectos de los fertilizantes sobre B. bassima concordaron con los efectos de los fertilizantes sobre nemátodos entomopatógenos, aunque se observó gran variación en los estudios del hongo. Posteriores estudios serán requeridos para determinar si las tendencias observadas en este estudio son aplicables bajo condiciones de campo (Rosin, 1996).
5.2.7.2. Herbicidas
Méndez (1990) menciona varios trabajos de diversos autores que evaluaron la sensibilidad de B. bassima a 21 herbicidas selectivos en condiciones de campo y laboratorio. Las pruebas de germinación in vitro señalan al Glyphosate y Oryzalin como los únicos herbicidas que no causaron inhibición significativa en la germinación dt: los conidios, en tanto que Alaclor, Diclofop, Dinoseb, Diuron, Oxiluorfen, Paraquat, Pronamide, Sethoxydim, Simazine y 2,4-D resultaron altamente inhibidores de la germinación, en un 88 a 100%.
En pruebas de campo se observó que la mortalidad de Spadoptera fnrgiperda fue reducida en forma significativa cuando las larvas se expusieron a conidios y suelo tratado con Alaclor y Oryzalin, sin embargo, el número de colonias producto de conidios viables se redujo considerablemente en suelos tratados con Alaclor pero no en el tratado con Oryzalin.
4.2.7.3. Insecticidas
Los insecticidas constituyen el gnipo de agroquímicos que más investigaciones ha motivado. Steinhaus (1956, citado por Ramaraje et al., 1967) puntualizó la compatibilidad de patógenos y muchos insecticidas como una de las ventajas del control microbial. Se menciona la posibilidad de dos materiales asistiendo a otro, el patógeno desarruiiando la enfermedad suficiente de la plaga y logrando que pierda resistencia al químico y el químico en cambio debilitando a la plaga lo suficiente como para hacerla m á s susceptible a la infección por el patógeno ( Franz, 1961 y Hall, 1963, citados por Ramaraje et al., 1967:i.
Pahicia Elizabeth Bañar Guevara 41

REVISION DE LiTER.A1'uRA
Estos mismos autores probaron el efecto de los insecticidas: Dhecron, Folidol, Diclorodifedtricloroetano (DDT), Malation, Endrin y Isómero g a m a de 1, 2 , 3, 4 , 5, 6 hexacloro ciclohexano (BHC), a dosis de 0.04%, 0.0596, 0.06%, 0.1% y 0.5%, sobre el desarrollo de B. bmiana En este trabajo se obtuvo que el BHC 50% fue el más tóxico ya que inhibió el crecimiento del hongo en todas las concentraciones; a éste le siguieron el Endrin, DDT, Malation y Folidol; el Dhecron 100% fue el más inocuo. L a s diferentes concentraciones de insecticidas tuvieron diversos efectos sobre el crecimiento y esporulación del hongo, y también reacciona de distinta forma al mismo insecticida.
Para controlar la larva de kpfinotarsa decemlineafa se empleó una combinación de B. bassiana y bajas dosis de DDT, los individuos con baja resistencia fueron muertos por intoxicación resultado de las bajas dosis del insecticida, las larvas más resistentes murieron de septicemia bacterial como consecuencia de la multiplicación en el hemocele de formas patógenas de la flora intestinal. Los insecticidas modifican los hemocitos, favorecen la penetración del epitelio intestinal por la microflora del tract0 digestivo y causan alcalinización del pH de la hemolinfa. Esto permitió concluir que hubo una escasa acción sinérgica entre Beauveria y el DDT.
Para el control de esta plaga las dosis empleadas son: 2 kg de Boverin con 6 x lo9 conidiadg y una quinta parte de la cantidad normalmente aplicada de DDT o policlorfineno/ha. Las investigaciones en la ex-Unión Soviética parecen ahora ser orientadas hacia el uso de Boverin sólo en dosis más altas (1 - 1.5 kg de Boverin con 30 x. 1 O9 conidiadg o 3 a 4 veces más que la dosis previa) para obtener una reducción en la población larval sin insecticidas quimicos.
En Francia, para el mismo tipo de plaga se asocian reducidas dosis de Carbaryl con un inóculo de 3 x lo'* - I x 1015 conidias por 100 l i tros, al orden de alrededor de 400 a 900 litros por ha, a un total o máximo de 3 -9 x 1Ols esporadha (comparado con 9 x 10'3 esporas/ha en 2 aplicaciones para autores soviéticos), lo que no mostró un efecto sinérgico (Femn, 1978).
Le Pelley (1973) menciona que los primeros trabajos de investigación se realizaron con BHC y DDT, en donde el BHC resultó ser el mejor.
Este autor detectó otros efectos sinérgicos en larvas de Melolontha melofontha atacadas por la muscardina, causadas por B. bmngnibii en presencia de bajas dosis de organoclorados y organofosforados. Bajo estas condiciones, se increment6 la incidencia de muscardina y se aceleró el proceso de infección y la sensibilidad del insecto a micosis, lo que pa= que hay una concentración de esporas que causan mortalidad a pesar de la adición del insecticida. Es interesante notar que en estos casos particulares el sinergismo aparece sólo si la virosis o bacteriosis está ya en el insecto cuando la contaminación por Beauveria ocurre, y que bajo estas condiciones los dos tipos de micmrganismos se desarrollan simultáneamente.
En trabajos posteriores, h e i d a y Cavalvante (1964, citados por Méndez, 1990) obtuvieron que el Dieldrol (Dieldrin en aceite), Thiodan y Dieldrex (Dieldrin concentrado emulsionable) efectuaban un control satisfactorio de la broca, similar al testigo tratado con BHC al 1%. El instituto Brasileño del Café (1973), recomienda el Lindano 20% en las dosis de 2.0 y 2.5 l/ha y el Endosulfan (Thiodan 35%) a razón de 1.5 a 2.0 l/ha, repitiendo la aplicación 20 a 30 días después de la primera.
Pahicia Elizabeth Bañm Guevara 42

RFVISIÓN DE LiTERAlURA - Muñoz (1985, citado por Méndez, 1990) indica que en Honduras, se adoptó el uso
de la mezcla de Endosulfán al 35% en dosis de 15 cdgalón de agua más 3.5 cc del dispersante-adherente Agral 90, en aspersiones dirigidas al cafeto. Después de la cosecha, se efectuaron espolvoreos al suelo con BHC al 3% o Aldrin al 2.5% a razón de 150 lblmanzana; toda esto en un intento por erradicar la plaga. Después del fracaso de la erradicacióiz, se prohibió el uso del Dieldrin y el BHC por su alta persistencia en el ambiente y por el efecto negativo de este último en la calidad de la bebida.
En Centroamérica y México se mmiendaba la aplicación de Endosulfán más adherente para el combate químico de la plaga (ANACAFÉ, 1981; INMECAFÉ, 1983). Decazy (1988, citado por Méndez, 1990) reportó las diferentes dosificaciones de este insecticida en cada uno de los paises e indica que en México se aplican 210 g de ingrediente activoha, en Guatemala y Honduras 525 g, en El Salvador 750 g haciendo notar que en África la recomendación es de 1000 g. A partir de 1995, la Norma Oficial NOM-FITO-002- 1995 restringe el uso de Endosulfán, realizándose solamente en focos nuevos (infestaciones nuevas) supervisado por personal oficial o unidades de verificación.
Para controlar a la broca los productores de café usan polvos como el Malation, Paration o granulados como son Lorsban al 1.5% polvo o 3% granulado, algunos otros, poco usados pero que siguen siendo efectivos son el Volaton y el mismo Sevin, esto con la idea de usar los más baratos para abatir los costos. Por razones de costos no conviene utilizar productos como Furadan, Dysiston; estos son recomendables si se requiere controlar un complejo de plagas (gallina ciega, gusano de alambre, minadores, trips, pulgones, etc.).
5.2.7.4. Fungicidas
Olmert y Kenneth (1974) evaluaron 9 fungicidas sobre vanas cepas de B. bassiana, en donde se observó una gran variación en la sensibilidad del hongo a algún fungicida en particular. Entre los productos que más inhibición causaron en todas las cepas se encuentran el Benomyl y el Maneb; dentro de los que menos daño provocaron se encuentran el Oxiclomro de cobre, Dinocap y Daconil2787, en tanto que el Binapacryl no iuvo ningún efecto sobre la genninación de las esporas.
Furher (1980, citado por Méndez, 1990) indica que los fungicidas Euparen y Orthocid 50 son eficientes para el control del crecimiento de hongos sapryifitos sobre trozos de floema de abeto, el cual se usó como aiimento para piyogenes chalcographus en experimentos de laboratorio. Sin embargo, las infecciones de B. bassiana sobre este insecto fueron inhibidas cuando se les proporcionó alimento tratado con estos productos.
Tedders (1981) menciona que al evaluar los seis fungicidas in vitro más frecuentemente empleados en el cultivo del nogal pecanero contra el hongo B. bassiana, los cuales parasitan al picudo Curcuho caryae Horn, observó que el Hidróxido de trifenil fue el fungicida más tóxico para ambos patógenos, seguido de Benomyl, metil- l-buticarbamoyl-2- benzimidasolcarbamato, Zineb, Zinc etilenbis (ditiocahameto), Diodine n-dodecilguanidina acetato. Entre los productos menos tóxicos al hongo se encuentra el Azufre y Dinocap 2-(1- metdfenil)-4, 6 dinitrofenil crotonato.
Patricia Elizabeih Baños Guevara 43
1

REVISIÓN DE LITERATURA
Clark y colaboradores (1982) evaluaron los fungicidas más recomendados para el cultivo de la papa con el fin de conocer su efecto sobre B. bassima en cultivo líquido bajo condiciones de laboratorio. Los resultados indican que el Mancozeb inhibió por completo al hongo, el Metiram y Clomtalonil fueron los productos que mayor inhibición causaron sobre el hongo durante siete días de prueba, en tanto que el fungicida experimental CGA 48988 fue el menos tóxico debido a que mostiur una inhibición pequeña del hongo. Las evaluaciones en condiciones de campo dieron resultados similares a los obtenidos en laboratorio.
Rodriguez (1984) señala que al probar la acción de siete fungicidas (Tridiamefon, Benomyl, C a b , Captafol, Ferbam, Priracarbolid y Oxicloruro de cobre) que se usan en el cultivo del cafeto en Guatemala, a t res dosis: 0.5, 1.0 y 1.5 kg en el desarrollo del hongo 8. bassima, observó que todos los fungicidas tienen una influencia en el desarrollo del hongo y que de ellos sólo el oxicloruro de cobre inhibió menos al hongo. Los fungicidas que más inhibieron al entomopatógeno fueron Calixin, Benomyl y Captafol en sus t r e s dosis y el Ferbam en la dosis de 1.5 kg.
5.2.7.5. Reguladores de crecimiento y coadyuvantes
Storey y Gardner (1986) evaluaron 4 reguladores de crecimiento y 8 coadyuvantes, seleccionados por su uso potencial en jardines, praderas para pastoreo y huertos, en los cuales B. bassima puede ser usado como insecticida microbial. En esta investigación se obtuvo que los reguladores de crecimiento: Siiaid, Paclobutrazol y Flurprimidol inhibieron la germinación y el crecimiento del hongo, no así el Mefluidide que causó poco o ningún efecto sobre el hongo. La mortalidad de Spodopfera íiugiperda debido a B. bassima se redujo en forma significativa cuando las larvas se expusieron a los conidios en suelo tratado con Paclobutrazol, en tanto que la mortalidad registrada en larvas sobre suelo tratado con Mefluidide no fue signtficativamente diferente de la mortalidad causada por los conidios sin tratamiento. Respecto a la evaluación de coadyuvantes, el Triton CS-7 fue el único que causó inhibición en la genninación de los conidios.
5.2.8. Producción de hongos entomopatógenos
5.2.8.1. Caracterización y selección de cepas de hongos entomopatógenos
La virulencia y la especificidad de las cepas, la capacidad de producción masiva de formas resistentes, para un almacenamiento satisfactorio de las preparaciones biológicas, y su inocuidad para vertebrados son las principales propiedades que deben ser consideradas para el desarrollo de técnicas de control microbiológico de plagas de cultivos (Ferron, 1978).
5.2.8.1.1. Caracterización de cepas
La clasificación de hongos entomopatógenos está basada en el criterio morfológico; sin embargo, el conocimiento de la virulencia y la especificidad ha revelado la necesidad de caracterizaciones m á s precisas. Alguna información es obtenida por un estudio especializado de la conidiogénesis, or observaciones de la ultraestructura de las esporas en el microscopio electrónico, y en algunos casos por estudios fisiológicos, pero la correcta
Patricia Elizabeth Baños Guevara 44
I

REVISIÓN DE LlTERATuRA
caracterización de las cepas requiere nuevas técnicas. Para esto Fargues et al. (1975, citados por Ferron, 1978) usaron métodos electroforéticos e hunoelectroforéticos asociados con las reacciones características de la actividad enzimática en dos cepas de 8. bassima y Beauveria tenells Las técnicas serológicas son también usadas en la ex-Unión Soviética para caracterizar estos hongos.
5.2.8.1.2. Selección de cepas
En Colombia, para seleccionar los aislamientos más patogénicos, se desarrolló una técnica de bioensayo que permite conocer la actividad de la b m en el laboratorio, antes de ser utilizado en el campo. Se demostró también la importancia de infectar poblaciones de este insecto con B. bassima para reactivar su patogenicidad, porque cuando se cultiva este hongo en medios artificiales durante t res o m á s generaciones se reduce considerablemente la patogenicidad y se incrementa el tiempo promedio para causar mortalidad en la mitad de la población (Ti,,) en comparación con el hongo activado sobre broca (González et al., 1993, citados por Bustillo, 1997). Las evaluaciones de la virulencia de los aislamientos contra la broca, han mostrado que existen diferencias no sólo en el nivel de mortalidad, sino en el tiempo requerido para matar su hospedero. También existen diferencias entre aislamientos en cuanto a la producción de conidias.
5.2.8.1.3. Técnicas de aislamiento
Berlanga (1997) menciona que para identificar un agente causal, es necesario aislar los microorganismos asociados al insecto en colonias puras, utilizando especítnenes frescos, evitando el empleo de material descompuesto, puesto que en tales condiciones es muy común encontrar d o s o m á s especies de micmrganismos en donde proliferan más fácilmente contaminantes sapxdfitos. Las condiciones de los especítnenes colectados darán la pauta para determinar las técnicas o tratamientos que se utilizarán para lograr su purificación, conbsiderando las exigencias nutricionales y ambientales de algunos géneros de entomopatógenos.
A continuación se dexriben algunas de las técnicas de aislamiento de hongos:
* Siembra directa de esporas. Para realizar esta técnica es necesario que el insecto atacado esté esporulado y no tenga contaminantes; en caso de que no haya esporas, Alves (1986, citado por Berlanga, 1995) recomienda que el insecto sea colocado en cámara húmeda para inducir la esporulación. El proceduni ento consiste en observar al insecto a través del microscopio estereosujpico para detectar las áreas de mayor esporulación, con la ayuda de un asa bactereológica o aguja entomológica flameada, se toman las esporas del insecto micosado y se siembran en una caja Petri o tubo de ensayo distribuyéndose en forma de estria en el medio de cultivo. Esta técnica se usa comúnmente en el aislamiento de Metarhizium spp., Beauveria spp. y Paeciomyces spp.
. .
* Transferencia del insecto o partes de tejido atacado. Esta técnica se usa cuando no hay presencia de esporas en el insecto colectado o cuando se observa el crecimiento de contaminantes junto con las esporas del hongo que se desea purificar, recomendándose transferir tejido interno del insecto micosado, siempre y cuando la parte interna esté completamente invadida de micelio.
Patricia Elizabeth Bañw Guevara 45

REVISIÓN DE LiTEFLAWR.4
El insecto atacado o partes de éste se depositan directamente en cajas de Petri con medio de cultivo, la superficie del insecto se desinfecta por inmersión en una solución de hipoclorito de sodio. Las placas se incuban a una temperatura de 27OC hasta lograr un crecimiento y una esporulación óptima.
* Medios seledivos. El uso de medios de cultivo seledivos nos facilita la purificación de algunos géneros de hongos entomopatógenos que son difíciles de aislar ya que requieren de nutrientes específicos para acelerar su crecimiento y esporulación.
* Técnica del insecto trampa. Para la detección de hongos entomopatógenos en forma indirecta, se reah un muestreo de suelo colectanto 1.5 kg que se coloca en bolsas de plástico con los datos de indentificación correspondientes, evitando temperaturas elevadas durante su transporte. Ya en laboratorio, 0.5 kg de la muestra de suelo se colocan en un recipiente de plástico humedeciéndolo con agua destilada, posteriormente se colocan 5 larvas de palomilla de la cera (Galleria rnellonella) de la última etapa de crecimiento, se cierra la charola y se invierte colocándola a 28°C durante 5 dias. Después de este tiempo se extraen las larvas, separando aquellas que presentan síntomas de infección por hongos para ser colocadas en cámara húmeda hasta la esporulación del hongo (Hernández, 1994, citado por Berlanga, 1995).
* Aislamiento de B. bsssiana. Ampliamente esparcido pero no comúnmente encontrado:, este hongo es un parásito primarito de insectos (ej. escarabajos de la papa, gusanos de .seda, arañas, langostas, abejas, avispas, varias larvas y capullos). Aislado de insectos muertos y larvas momificadas; se esteriliza la superficie en una solución de hipoclorito de d i o o calcio al 2-5%. Se inoculan las placas de Petri con Saboraud Maltosa Agar (SMA); se aisla del micelio blanco que se desarrolla en la superficie del especimen o del agar. El cultivo sobre un agar con infusión de heno es bueno para conidióforus con reducción del micelio vegetativo, sobre SMA para abundante micelio, el cual en algunos casos inicia a espomlar después de pocas semanas (Russell, 1974).
Alcocer (1979) señala como un medio de cultivo adecuado al obtenido con gelosa- papa-peptona, con el cual se puede obtener i n h l o para efectuar pruebas de laboratorio. Chase, Osborne y Ferguson (1986, citados por Méndez, 1990) indican que el medio de cultivo formado por 20 g de cereal de avena para bebé, 20 g de agar, 0.55 g/l de dodine y 5 mg de clorotetracicha, permite la recuperación de B. bmima en forma pura. Además, señalan que la adición de 10 mg/l de cristales de violeta ayudan en la visibilidad de las colonias del hongo sin afectar la frecuencia de recuperación.
5.2.8.1.4. Conservación de aislamientos
Una vez que las cepas de hongos se han identificado y purificado, es necesario formar colecciones, en las cuales las cepas de intei.és deben conservarse en un estado metabólico adecuado, considerando como funciones principales preservar la viabilidad y pureza del aislamiento, conservar sus caracteristicas morfológicas y genéticas y mantener su patogenicidad al paso del tiempo. Cualquiera que sea el propósito de la conservación de micmrganismos, es necesario obtener la información precisa de s u s caracteres biológicos, morfológicos y metabólicos, además de que sirven para preservar las cepas de interés. También hay que tener en cosideración aspectos desde el punto de vista epizoótico como su origen, fecha de aislamiento, lugar y hospedero (Berlanga, 1997).
P - Pairicia Elizabeth Bañar Guevma 46

Para que las cepas no pierdan su virulencia, se recomienda, después de 5 transferencias por medio de cultivo, reactivarlos infectando al hospedante natural o a un alterno.
* Métodos de conservación de los aislamientos. La selección del método para conservar las cepas, depende de la intención de los servicios para los que sea requerida, la inversión con que se cuente, el equipo y las instalaciones; por lo que se consideran l as de largo y de corto plazo, tomando en cuenta que las de corto plazo presentan desventajas en relación al primer tipo, porque implica mayores posibilidades de contaminación, perdida de características morfológicas o mutaciones indeseables (Berlanga, 1997).
En seguida se mencionan algunas condiciones bajo las cuales se pueden mantener las esporas de B. bassiam
Temperatura ambiente: Los aislamientos deben ser colocados en espacios cerrados para protegerlos del polvo o contaminantes. Sin embargo, bajo estas condiciones el medio de cultivo se seca en corto tiempo, por lo que es necesario transferir cada seis meses en medio de cultivo nuevo (Berlanga, 1995).
Refrigeración: Las cepas son sembradas en medios de cultivo en tubo inclinado, el cual es sembrado en estrias con esporas del hongo e incubado a 27OC hasta que se presente una esponilación completa, posteriormente se colocan en refrigeración a temperaturas que van de 4 a 8OC en un medio de cultivo rico en nutrientes; es necesario hacer transferencias cada seis meses a un medio de cultivo nuevo.
Aceite mineral: El aislamiento es sembrado con la técnica del tubo inclinado hasta su esponilación, una vez que el hongo ha cubierto la superficie del medio de cultivo, se agrega el aceite mineral hasta una altura de 1 cm arriba del punto donde termina el medio, previa esterilización durante 20 minutos a í20°C, repitiéndose tres veces y exponiéndolo entre cada esterilización en un período de 24 horas a temperatura ambiente. Posteriormente se mantienen los tubos en posición vertical en donde pueden durar viables por un año o más dependiendo del género con que se esté trabajando.
Silica gel: Las conidias del hongo son depositadas en una solución a base de cloruro de sodio, Tween 80 y leche descremada a la que se le agrega silica-gel activada por esterilización. Posteriormente el material es depositado en matraces conteniendo papel filtro estéril para eliminar el exceso de solución en los trozos de silica-gel, después se deposita en tubos esterilizados y se sella con parafina. El almacenamiento es a temperaturas de 4 a -2OOC y su conservación es por más de un año.
Liofilización: Este proceso se basa en la eliminación rápida de agua a baja temperatura y en vacío. Los hongos deben mantenerse durante este proceso en una solución “soporte” para protegerlas de los daños que puede ocasionar el congelamiento, la deshidratación y el oxígeno. Se debe hacer una congelación rápida seguida de una deshidratación lenta a baja presión. Después de la desecación, los tubos son fechados y almacenados en nitrógeno líquido. Con esta técnica, los patógenos pueden permanecer viables por m á s de 5 años.
Desecación en papel filtro: En este procedimiento se emplea primeramente una liofilización de los micmrganismos para posteriormente ser depositada en t rozos de papel filtro estéril y almacenada de O a 8OC. Con esta técnica se tiene la ventaja de que las cepas pueden ser transferidas fácilmente e incluso pueden ser enviadas en un sobre a otro sitio. La conservación con esta técnica es de más de 3 años.
Pahcia Elizabeth Bañm Guevara 47 -

REVISION DE LiTEU”R.4
Beauveria bassima se puede conservan como una suspensión de esporas en agua destilada a 4°C. Cada dos meses el hongo se traspasa sobre Agar Dextrosa Saboraud por 8- 10 dias; las esporas fueron aisladas, lavadas y suspendidas en agua destilada en una concentración de 1 x 109 o 1 x 1010 esporas/mL y conservadas a 4 T . Para algún experimento se incuban a 27T . El criterio para la germinaciónes que la conidia forme un tubo de genninación de una longitud de almenos 1 y !h veces el diámetro de la conidia (Thomas et al., 1987).
5.2.8.1.5. Seguridad
La seguridad de los hongos entomopatógenos para los vertebrados no ha sido completamente resuelta todavía a pesar de varias y diferentes investigaciones que han examinado el problema. Se requiere un gran esfuerzo en este campo, tal vez guiado por las recomendaciones del control microbian0 propuesto por la Agencia de Protección Ambiental de Estados Unidos. Was consideraciones van más allá de los fenómenos alérgicos ya encontrados en la manipulación de grandes cantidades de esporas secas.
Elios también abarcan estudios sobre la contaminación oral de especks de vertebrados, probar con vertebrados y, particularmente, con cultivos de tejido humano, el riesgo de cáncer, estudios serológicos y genéticos, y pruebas para la infección de fauna silvestre Útil y fauna domesticada (Agricultura de las Américas, 1978).
5.2.8.2. Técnicas de producción de hongos
De acuerdo con Gana (1995) las técnicas de producción de hongos pueden clasificarse en:
1. In vivo Se utilizan insectos vivos, lo que tiene potencial limitado y es poco importante desde un punto de vista comercial o semicomercial. Se ha trabajado con la inoculación de brocas sanas con broca infectada y muerta por el hongo. En esta forma se logró incrementar el número de brocas muertas de dos a más de 200.
2. In vitro Usando medios de cultivo en sustratos sólidos, ííquidos, semisólidos (cultivo difásico).
5.2.8.2.1. Producción en cultivo sólido
El sustrato sólido está caracterizado generalmente por el crecimiento de micruorganismos sobre sustratos insolubles al agua, en presencia de cantidades importantes de agua libre. La confusión estriba en definir el agua libre. Moo-Young y colaboradores (1983, Citados por Doelle et al., 1992) proponen el término de fermentación en estado sólido a todos los prucesos los cuales utilizan materiales parcialmente insolubles en agua para crecimiento microbiano en ausencia de agua lib=. Con el aumento del agua libre, las fermentaciones sólidas pmgresan de sólidas a fermentaciones de suspensiones de partículas sólidas.
Pahicia Elizabeth Bañar Guevara 48

Las definiciones de fermentación de sustrato sólido y la de estado sólido no son fácilmente precisables en cuanto a cómo aparece el agua fibre en diferentes sustratos y a diferentes contenidos de agua; ejemplo, en cascarilla de maple a 40% de humedad y en paja de trigo a 75% de contenido de humedad (Cannel y Moo-Young, 1980, citados por DoeUe et al., 1992), no es fácil medir el parámetro ni distinguir entre ellos la diferencia; aunque la actividad de agua (Aw) es un candidato potencial.
Algunos sinónimos de la fermentación en estado sólido son: * Fementación de sustrato sólido * Fermentación de fase sólida * Cultivo en estado sólido * Fermentación semisólida * Cultivo en superficie * Fementación koji
5.2.8.2.2. Producción en cultivo líquido
* Cultivo líquido de superficie. El cultivo estático se puede definir como aquel proceso de fermentación que se realiza sobre la parte superficial del medio de cultivo, que en este caso es líquido; se desarrolla en botellas o en charolas descubiertas que contienen el medio de cultivo estático y la producción se ubica en salas estériles. Su principal ventaja radica en que es una tecnología sencilla, de baja inversión y no tiene problemas de aporte energético (Crueger y Crueger, 1984).
En este sistema las variables de mayor importancia son el área de cultivo y la profundidad de la charola, el cultivo es análogo al cultivo sólido, donde las esporas se forman en la interfase de la película de crecimiento-aire, mientras que el micelio crece en la interfase película-medio. Se emplea para cultivo de hongos. El cultivo es muy susceptible a contaminación, por lo que la manipulación es muy delicada modes y Fletcher, 1969).
* Cultivo líquido sumergido. otro sistema de cultivo de hongos entomopatógenos es el que se basa en un medio de cultivo cuyo estado físico es líquido y en el cual el hongo está inmerso. La técnica de producción sumergida tiene las obvias ventajas de que el cultivo y el proceso pueden hacerse bajo condiciones estériles controladas y el escalamiento es relativamente fácil. Por eso el método preferido para la producción en gran escala de material infeccioso es una etapa de cultivo sumergido del hongo para producir esporas en un tiempo relativamente corto.
Dependiendo de las condiciones ambientales, los hongos imperfectos pueden crecer en cultivo líquido formando micelio o en forma levaduriforme; en el primer caso, las células son alargadas y no existe separación de las células madres de las hijas y en el segundo caso, las células tienden a ser esféricas y la reproducción es por gemación. Algunas de las condiciones que influyen en la morfología de los hongos son: la temperatura, el pH, la naturaleza y concentración de las fuentes de carbono y nitdgeno, la actividad de agua (Aw), la concentración de oxígeno y dióxido de carbono (intercambio gaseoso), la iluminación y la concentración de ciertos iones (calcio, magnesio, fosfatos) (Thomas et al., 1987; Cárdenas, 1995).
Patricia Elizabeth Bañar Guevara 49

I
REVISION DE LITfRATüRA
Un importante avance fue realizado en la ex-URRS, en la producción directa de conidiosporas de B. bassima en cultivos sumergidos. Muchos detalles de la conidiación de este hongo en tanques de fermentación son encontrados en la literatura soviética. Entre 5 x ip a 2 x 109 conidias/ml fueron formadas dependiendo de la composición del medio y tecnología de la fermentación. No obstante, podría parecer que la conservación tal como la preparación también posee serios problemas (Femn, 1978).
B. bassiana en cultivo líquido puede producir blastosporas y ocasionalmente conidias sumergidas. Para uso corno un bioinsecticida, la conidia tiene ventajas definitivas. Numerosos estudios han evaluado la producción de conidias en cultivos líquidos usando medio de grado sintético e industrial suplementado con glucosa, también se ha estudiado el crecimiento y esponilación en microcultivos usando medio de crecimiento conteniendo monómeros de quitina (Hegedus et al., 1990). La producción masiva de blastosporas en cultivos sumergidos se desarrolló hace aproximadamente 20 años; sin embargo, esta se ha abandonado por las dificultades de almacenar este tipo de conidias (Femn, 1981).
Para determinar la máxima producción de conidias sumergidas se propone monitorear tres parámetros: (1) la germinación del inóculo conidial, (2) el rápido crecimiento micelial, y (3) la producción preferencial de conidias sumergidas y formación suprimida de blastosporas (Hegedus et al., 1990).
Aunque las blastosporas han sido exitósamente producidas a escala industrial en un medio aireado y agitado, la terminación del proceso y la conservación de la preparación (concentración, secado) generalmente reduce la virulencia de las esporas. Por lo tanto, las esporas sumergidas son usadas sólo como un inóculo del medio superficial de donde las conidias aéreas obtenidas son secadas para que permanezcan efectivas por un prolongado penodo (Samsináková y Kálalová, 198 1).
Algunas de las características del cultivo sólido en comparación con el cultivo líquido sumergido se presentan en el Cuadro 3.
5.2.8.2.3. Producción en cultivo difásico
La técnica más comúnmente usada para la producción masiva involucra un cultivo en superficie, o dos etapas de cultivo en las cuales el hongo es desamilado primero bajo condiciones sumergidas y entonces se le permite esporular conidias aéreas en cultivo de superficie en medio líquido estático, en medio sólido o semisókb (MacLeod, 1954 y Hall y Paperok, 1982, citados por Hegedus et ai., 1990). En estos métodos de cultivo para producir conidias, el prcducto anhelado es altamente insecticida, es decir, debe ser de gran virulencia y resitencia a condiciones ambientales adversas.
El cultivo difásico tiene la ventaja de aumentar l a producción por unidad de materia p-, su principal desventaja es la gran posibilidad de contaminación debido a la manipulación que implica el manejo de dos etapas diferentes (Cárdenas, 1995). Esta técnica mezclada se ha desamilado principalmente en el instituto Ucraniano para la Pmteccion de Plantas en Kiev, para la producción de ciertas conidias de B. bassima, M. anisopliae y K lecanii. En medio líquido contiene blastosporas del cultivo sumergico y se vacía sobre camas de salvado o semillas de cereales para promover el rápido d e m i i o de una capa micelial, con el fin de producir conidióforos. A pesar de las dificultades técnicas, esto p e m t e la producción de suficientes cantidades de conidias para permitir la estandarización (Femn, 1978).
Palricia Elizabeth Bañas Guevara 50

CUADRO 3. DIFERENCIAS ENTRE CULTIVO EN SUSTRATO SÓJLDO Y CULTIVO LíQUIDO
* M&O de cultivo sin tiujo. * profundidad del medio escasa, excepto en bi0ItaCtom. * Se emplea un sustrato sólido insoluble en agua que provee las fuentes de c h n o , nitxúgeno, minerales y e n e ~ a . * El sustrato sólido abs& agua, al aiiadir el agua y nutrientes se toma en un medio sólido húmedo. * El agua aprovechable es suficiente para obtener un óptimo crecimiento y metabolismos de cultivo y por lo tanto constituye una pomión del volumen total del medio. El sistema de cultivo involucra 3 fases: sólido,
liquido y gaseoso.
* La fase líquida es discontinua en el sitema. * El sistema de cultivo no es un medio aséptico, se tiene que cocinar o primero esterilizar, lo que impiica mayores gastos. * No es posible controlar estríctamente los parámetros durante la fermentación. * El rango de inóculo siempre es aito.
' El oxígeno para crecimiento y metabolismo se deriva en mayor parte de la fase gaseosa y en menor grado del disuelto en asociación con el agua del susirato. * La aireación provee oxígeno y también remueve el calor metabólico y los pxuductos gaseosos.
* El sistema puede o no involucrar agitación del medio. * El crecimiento fungal involucra una pfunda penetración de la hifa dentro de las pariículas del sustrato sólido. * El medio fermentado para cosechar está en forma sólida-húmeda, se utilizan directamente los sólidos fermentados y el producto se encuentra en forma concentrada.
Mitchel y bnsane, 1992, citados por Dwlle et
SUMERGIDO CULTIVO EN SUSIRATO S6LJDO I CULTIVO LfQUIDO SUlMERGIDO
* Simulicidad del medio de cultivo. 1 ' Medio de cultivo complejo. * ~ e d i o de cultivo siemp& con flujo. El medio es invariablemente PW~LUUIO.
Se emplean diferentes fuentes solubles en agua: excepto en algunos casos donde uno de eiios es insoluble en agua. ' Los nutríentes son uniformemente distribuidos en el medio y se pueden añadir en cualquier momento de la fermentación. * El agua aprovechable es abundante sin alguna estandarización para su requerimiento y, por la tanto, constituye la mayor porción del volumen total del medio. El sistema de cultivo involucra 2 fases: líquido y
gaseoso. A veces involucra la fase sólida, pero la concentración es baja por lo que multa en una suspensión en el medio.
Continuidad en la fase liquida. * Estos sistemas son siempre bajo condiciones asépticas, económicamente tiene un inegable efecto y pmductividad. * Es esencial un riguroso control de todo:; lor parámetros durante la fermentación. El rango de inóculo es bastante bajo, a menor
que lo requiera el pmeso. * El oxígeno es tomado sólamente del disuelto er elmedio.
* La aireación provee oxígeno y remueve productos gaseosos, el calor se elimina p a s a d agua por el fermentador. * La agitación homogeniza el medio, excepto er fermentación anaerobia. * Las células del micelio del hongo crecen er forma de peJieís miceiiales que son distribuidor en el cultivo agitado.
El medio fermentado es cosechado en e s a líquido y contiene pducto en conceniraciór mucho más baja comparativamente.
al., 1992

5.2.9. Producción de B. bassiana en cultivo sólido
5.2.9.1. Proceso de producción
En general los pasos involucrados en la producción en cultivo sólido consisten básicamente de (Lonsane et al., 1982 y 1985, citados por Doelle et al., 1992):
a) La preparación del sustrato sólido a menudo involucra un ptratamiento para disminuir el tamaño de partícula o incrementar el aprovechamiento de nutrientes en el sustrato para el organismo. Los nutrientes pueden ser adicionados y se ajusta el pH en esta etapa.
b) Etapa de cocimiento, en la cual se esteriliza o al menos se p a s t e u k el sustrato, y causa la absorción de agua dentro de las partículas del mismo.
c) El planteamiento de la cantidad adecuada de inóculo, ya sea por la técnica tradicional o por la técnica de cultivo puro.
d) La inoculación de sólidos húmedos
e) La incubación en recipientes adecuados
O El mantenimiento de las condiciones óptimas tanto como sea posible.
g) La cosecha de los sólidos.
h) El secado de los sólidos o extracción del producto de los sólidos.
i) El procesamiento posterior si es necesario.
5.2.9.2. Tipos de sustratos sólidos
El amplio rango de sustratos sólidos utilizados en el cultivo pueden ser clasificados en dos grandes categorías: 1) las partículas son al mismo tiempo el soporte y el sustrato, este es el caso de todos los materiales orgánicos, 2) la parte sólida sólo es soporte (minerales o materiales sintéticos) y deben ser humedecidos con diferentes soluciones nutritivas para el desarrollo del microorganismo (Durand et al., 1993, citados por Hemández et al., 1997). De acuerdo al trabajo de tesis desarrollado nos centraremos posteriormente en la primera clasificación.
Los sustratos sólidos empleados en la fementación sólida involucran un gran número de macmmoléculas tales como almidón, celulosa, hemicelulosa, pectina, iignina, proteínas y lípidos (Mudgett, 1986, citado por Doelle et al., 1992). Estas macmmoléculas usualmente proveen la fuente de carbono y eneqía para el crecimiento microbian0 además de poder enriquecer la formulación de los diferentes medios de cultivo adicionando nutrientes como: extractos de carne, levadura, peptona.
P
Patricia Ehabeth Bañar Guevara 52

5.2.9.2.1. Uso del almidón como sustrato para procesos de cultivo sólido
El almidón es un sutrato caro debido a que tiene muchos usos alternativos. Por ejemplo, el almidón puede incorporarse áirectamente a dietas humanas y animales. El almidón se ha empleado en diversas estudios de cultivo sólido y se han dirigido a incrementar el valor nutricional. ia principal ventaja del almidón es que es fácilmente degradable y puede ser utilizado por un amplio rango de micmorganismos. Los sustratos de almidón que han sido usados más múnmente en cultivo sólido son maíz, a m z , trigo, salvado de trigo, sorgo, malta, papa, harina de plátano, residuo de caramelo de papa, harina de centeno y salvado de semillas (Telenga, 1958, citado por Samsináková y Kálalová, 1981; G a m , 1995).
En esta investigación se empleó a m z como sustrato para el cultivo sólido dada la naturaleza particulada del a m z es ideal para este sistema de cultivo. Recientemente, It0 y colaboradores (1989, citados por Doelle et al., 1992) usaron arroz en estudios fundamentales de penetración de micelio en cultivo sólido. El arroz es un sustrato conveniente para estos estudios debido a que provee un medio nutricionalmente completo para crecimiento microbiano. Ninguno de estos autores tuvo la necesidad de adicionar nutrientes extras al a m z .
5.2.9.2.2. Estructura química del almidón
Los carbohidratos son la fuente principal de energía para los organismos vivos. En la alimentación de los seres vivos el almidón es de las principales fuentes de carbohidratos, es un polisacárido producido por las plantas durante la fotosíntesis. Las plantas pueden almacenar grandes cantidades de almidón dentro de sus propias células en tiempo de abundancia de nutrientes, para poder utilizarlo cuando existe una demanda de producción de energía o servir corno alimento para otros seres vivos.
Las unidades fundamentales del almidón son la D-glucosa y es el único monosacárido que se obtiene de la hidrúiisis total de estos monosacáridos. Los almidones son homopolisacáridos que están fomados por dos polimeros, la amilosa y la amilopectina, se encuentran presentes en cantidades variables (Santos, 1995).
Los rangos de amilosa y amiiopectina varían entre diferentes fuentes: maíz, papa y almidón de trigo tienen de 69 a 77% de amilopectina y de 23 a 31% de amilosa (Galliard y Bowler, 1987, citados por Doelle et al., 1992).
Amilosa: Es un polimeto compuesto de unidades de glucosa unidas por enlaces a-1,4, que se encuentran formando cadenas lineales que pueden tener 2000 a 2500 monosacáridos, siendo la maltosa la unidad de repetición. ia amiiosa era considerada completamente iineal, pero hay evidencia que indica que tiene la propiedad de adquirir una conformación tridimensional helicoidal. Esta propiedad de la amilosa de formar hélice, hace que al colocar yodo en solución, se forme un complejo amilosa-yodo, debido aparentemente por la inclusión de una molécula de yodo en la hélice de amilosa. Este complejo es de color azul intenso y se requiere de un mínimo de 40 moléculas de glucosa para desarrollar perfectamente el color azul, por lo que las cadenas cortas de amilosa p d u c e n un color rojo con el yodo (Corm y Stumpf, 1975; Santos, 1995).
- - Patricia Elizabeth Baños Guevara = 53

Amilopectina: Es otro polímero formado por unidades de glucosa, es largo y altamente ramificado. En su molécula existen cadenas cortas formadas por glucosas unidas en posición a-1,4 (aproximadamente de 15 a 30 unidades), que a su vez están enlazadas entre sí por uniones a- 1,6 y son estos enlaces los que producen la ramificación. El peso molecular de la axniiopectina puede variar mucho aún en la misma muestra de almidón y puede ser de 500,000 o m á s grande (Conn y Stumpf, 1975; Santos, 1995).
5.2.9.3. Pretratamiento de sustratos de almidón
Los sustratos de almidón son cosechados como productos agrícolas en la forma de raíces, tubérculos o granos. Esto puede. ser usado directamente como sustratos para cultivos de sustrato sólido con un mínimo pretratamiento, tales como picado o quebrado, seguido de gelatinización. En este caso es esencial que la estructura celular de la planta sea destmida previamente dado que los micwrganismos usados no degradan a lignocelulosa generalmente. Alternativamente los productos agrícolas pueden ser transformados para producir almidón o harina. La molienda causa daños físicos al gránulo de almidón, haciéndolo más susceptible a hidrólisis por e&s (Galliard y Bowler, 1987, citados por Doelle, 1992). En todos los casos, la gelatinización representa un paso de pretratamiento esencial. La destrucción del gránulo de almidón durante la gelatinización hace a la amilosa y amilopectina mucho más susceptible a hidrólisis enzimática.
En situaciones prácticas, la gelatinización se alcanza por cocimiento en agua o vapor. A gran escala la vaporización es la opción más factible. La gelatinización debe realizarse con suficiente agua para alcanzar el contenido de humedad deseado. Alternativamente, el exceso de agua puede ser removido por drenado o en estufa de secado. La gelatinización del almidón para cultivo de sustrato sólido puede representar problemas. El problema de la gelatinización puede incrementarse debido a la baja conductividad térmica del sustrato, la presencia de este problema se debe al relativamente bajo contenido de agua. La conductividad térmica puede ser aumentada por la adición de agua, pero también mucha agua puede afectar adversamente la textura para el cultivo de sustrato sólido. Los problemas también se pueden presentar con el efecto de la gelatinización en la textura del sustrato. La amilosa que queda fuera del gránulo de almidón durante la gelatinización hace que las partículas del sustrato se peguen y tiendan a aglomerarse si el sustrato es mezclado durante la gelatinización. Esto ocasiona que el sustrato quede limitado en espacios interpartidas que no son aprovechables para el cultivo sólido. Algunos investigadores evitaron parcialmente la aglomeración adicionando 5% de aceite vegetal antes de la gelatinización (Doelle et al., 1992).
5.2.9.4. Condiciones de desarrollo
Se han tenido serios problemas con respecto al mezclado, intercambio de calor, transferencia de oxígeno, control de humedad y la localización de gradientes de pH y nutrientes, así corno los niveles de producto corno una consecuencia de la heterogeneidad del sistema de cultivo. Debido a estos problemas, los micwrganismos que sean seleccionados para el cultivo sólido deben ser muy tolerantes a un amplio rango de condiciones de cultivo (Mudgett, 1986, citado por Doelle et a1.,1992).
- - Patricio Elizabeth Borios Guevara 54

5.2.9.4.1. Temperatura
La temperatura de crecimiento es la responsable de la considerable variedad entre aislamientos, algunos hongos tienen amplio rango de temperatura Óptima mientras otros están mucho m á s restngidos, la selección de cepas de acuerdo a los requerimientos térmicos pueden ser garantía cuando se escoge una cepa para desarrollarse wmo un agente de control microbian0 (Fargues et al., 1997).
Resultados de varios experimentos confuman que B. bmiana es mesofilico, capaz de crecer entre 5 y 37OC wn un óptimo de genninación de 25 a 28-3OoC; l a s temperaturas Óptimas mencionadas para las d o s especies de Beauveria son: 23OC para B. bmnpiartii y 25OC para B. bmiana (Hernández, 1993); y aun así se afirma que este hongo puede desarrollarse dentro de un abanico de temperaturas de O a 40°C, ya que ha infectado a especies invernantes que habitan a temperaturas cercanas a 0°C (Walstad et al., 1970).
Las esporas toleran variaciones a altas temperaturas (98"C durante cinco horas continuas) en atmósfera seca; pero en atmósfera húmeda y expuesta a 4OoC soportan sólamente 24 horas, a bajas temperaturas no sufren reacción alguna, siendo la temperatura óptima para su crecimiento y esporulación de 28OC (Alcocer, 1979). El punto térmico de muerte para esporas de B. bassiana fue de 50OC. Temperaturas menores a 10°C y mayores entre 35OC y 49OC retardaron el proceso de germinación e incapacitaron la esporulación. La más rápida esporulación ocurre a los 5 días a una temperatura de incubación de 25°C y 30°C. A 35°C el tiempo requerido para esporulación fue de 8 días; a 10°C el tiempo fue de 15 días.
Tales variaciones en la tolerancia a la temperatura pueden explicar el amplio rango climático que exhibe este hongo; asimismo, con propósitos de manejo, las cepas de climas cálidos tienen buen desarrullo a altas temperaturas, mientras que cepas de temperaturas templadas se adaptan mejor a bajas temperaturas (Barson, 1977). Este relativo amplio rango de tolerancia térmica de B. bassima provee a las especies de este hongo de una notable adaptación a condiciones con grandes fluctuaciones térmicas. incluso independientemente de los orígenes geoclimátiws, puede ser verificado que el hongo tiene un complejci de especies o genotipos diferentes (Fargues et al., 1997).
Los efectos de la temperatura influyen directamente sobre la genninación de la conidia y el crecimiento vegetativo (Fargues et al., 1997). La rapidez del desarrollo micelial y la evolución de las infección, dependen de la temperatura. Temperaturas mayores o menores a las óptimas retardan el desarrollo de la micosis sin causar necesariamente la mortalidad total de los wnidios. Es importante considerar que la penetración de la cutimla no siempre es Ilifluenciada por el desarrollo del micelio wmo una función de la temperatura, la temperatura óptima para el desarrullo del hongo no es la misma que para el desarrollo de la enfermedad y en general los hongos son m á s sensibles a las temperaíuras elevadas que a las bajas, al grado de que pueden resultar letales (Femn, 1978).
La temperatura de almacenamiento afecta también la sobrevivencia de las esporas del hongo muscardina por períodos de tiempo largo; a 8OC las esporas permanecieron viables por al menos 12 meses. Peru a 2 l0C l a s esporas de B. bassiana sobrevivieron bien por sólo medio año (Walstad et al., 1970).
~ 55 Pujriciu Elizabeth Bañm Guevaru

Las represiones térmicas no son sólo resultado de condiciones ambientales, sino también a través de eílas algunos hospederos logran la termorregulación. Por ejemplo, los insectos como los Acrididos elevari su temperatura del cuerpo cuando seleccionan su habitat o se d e n t a l al sol y tal actividad ha sido mostrada para reducir la incidencia de la enfermedad Entomophthora muscae (cohn) Fres. en moscas Acridophaga Henry, Enthomophaga grylli (Fres.) Batko, B. bassima (Bals.) Vuill. y Metarhizium flavoviride Gams en acrididos. Es por eso importante equiparar los requerimientos térmicos de un prospecto de agente de control microbian0 a las condiciones climáticas esperadas del ambiente y/o del nivel del hospedero (Fargues et al., 1997).
Walstad et al. (1970) observaron que la temperatura afecta la eficacia de la “muscadna blanca” como agente de control biológico en Hylobius pales. Las esporas de ambos hongo germinaron a temperaturas entre 15 y 35T, el pico máximo de germinación ocumó de 25 a 35OC y la muerte de esporas ocurre a temperaturas cercanas a 5OoC.
Barson (1977) evaluó la patogenicidad de B. bassima sobre el escarabajo grande de la corteza del olmo Scorytus scolytus, en todas las larvas tratadas a la concentración de 2.8 x 107 esporas/ml, la mortalidad a 5’C fue lenta con un tiempo letal de 45 días y rápida a 25’C con un tiempo letal de 6 días.
Doberski (1981 a, b) menciona que las larvas del barrenador grande del olmo S~01ytus scolytus se inocularon con dos cepas de hongos entomopatógenos B. bassima y se incubaron a temperaturas de 2, 6, 15 y 20°C. Las cepas de B. bassima causaron una infección regular a 2OC y fue mayor a 15 y 20°C. La infección ocurrió a las t res temperaturas, pero a 2 5 T , los adultos de la plaga murieron por infección bacterial.
5.2.9.4.2. Humedad relativa
Diferentes autores han reportado anomalías en la biología de esporas de Moniliales a 30-45% de Humedad Relativa (H.R). Riba y Marcandier (1984, citados por Marcandier y Khachatourians, 1987) han mostrado que la longevidad de las conidias de B. bassima está afectada por la H. R bajo la cual las esporas son almacenadas: 15% de germinación después de almacenar a 33% de H. R, contra 78% a 0% H. R y 99% a 100% H. R. Clerk y Madelin (1965, citados por Marcandier y Khachatourians, 1987) encontraron la menor longevidad bajo condiciones de almacenamiento con alta H. R. Es evidente que la longevidad de la espora depende de la H. R; las Conidias de B. bassima y Paecilomyces farinosus viven más largamente a H. R de 34% que a 75%.
Las altas humedades atmosféricas favorecen el desarrollo de micosis epizooticas, varias informaciones permiten distinguir entre el mol de la humedad atmosférica durante el proceso de infección y su infiuencia sobre los diversos fenómenos ep;oot;cos. reconoce que el desarrollo del hongo sobre el cadáver y la esporulación están asociadas con la humedad del punto de saturación. Bajo estas condiciones la automultiplicación del inóculo resulta en una elevada contaminación de insectos sanos, favoreciendo el desarrollo epizoótico de micosis (Marcandiex- y Khachatourians, 1987).
Paíricia Elizabeth Bañm Guevara 56

REVISIÓN DE LITERATURA I
En otro aspecto, se ha demostrado que la infección en campo de los insectos se puede dar independientemente de la humedad ambiental; hay conidios de B. bassiana que no germinan si la H. R no es cercatm ai punto de saturación ai 92%- 100%; aunque existen cepas que pueden germinar a 40%, lo cual demuestra que los hongos se encuentran adaptados a diferentes atmóferas, por ello la selección de éstas puede aumentar la posibilidad de éxito en el manejo de un insecto en particular. Esto podría conducir a la suposición de que el fenómeno físico de la capa limite del integument0 del insecto facilita la germinación de esporas aun si la atmósfera es prácticamente deshidrata (Hernández, 1993).
Las variaciones en la humedad relativa no afectan la mortalidad en forma significativa; sin embargo, el deceso del insecto es más rápido en temperaturas óptimas (25 y 30°C) asociadas a condiciories de alta humedad (92%-100%) para una mejor genninación, crecimiento y esporulación de este hongo sobre insectos muertos, 10 mal explica el desarollo de e p h t i a s naturales en climas húmedos, ya que el inónilo sobre los cadáveres se multiplica sólo cuando las condiciones climáticas como la humedad relativa del microambiente del hospedero es cercano al punto de saturación. Valores menorea a 90% impiden la genninación de la espora. En cuanto al tiempo de genninación, a 100% de H. R. las esporas se desarrollaron en 4 días pero requirieron de 5 días a más a H. R. de 92.5-98% (Walstad et al., 1970).
Barson (1977) menciona que la H. R es importante en la patogenicidad de B. bassiana y cita que a 100% de H. R y a una concentración de 2.8 y 107 esporadml, todas las larvas del barrenador de la corteza del olmo Scolpus scol’us murieron.
Doberski (1981a) evaluó seis cepas (dos cepas de cada hongo) de B. bassjana, M. anisopliae y Paecilomyces farinaus y observó el efecto de la humedad relativa en la infección de las larvas Scolpus sc~lytus por una cepa de cada uno de los entomopatógenos; el rango de H. R fue de 51, 74 ,86 ,90 , 97.5 y 100%. B. bassiana y M. anisopliae causaron infección en todas las H. R, mientras que farinmusno fue infective a 51 y a 74% de H. R. El porcentaje de mortaiidad fue menor a 95 y 97% que a 100% de H. R.
Ramoska (1984) estudió en bioensayos la patogenicidad de conidios de B. bassiana en adultos de la chinche Blissus leucopterus leucopterus a 30, 50, 75 y 100% de H. R. y observó que el micelio del hongo y los conidios sólo se observaron en el rango de 75 a 100% de H. R Marcandier y Khachatourians (1987) realizaron estudios en laboratorio con el chapulín migratorio Melanoplus sanguinipes Fab. (orthoptera: Acrididae) y observaron que fue susceptible a la infección de fl. bassiana, la mal ocurrió independientemente de la H. R (12, 33, 76 y 100%) pero la mortalidad después del tratamiento fue mayor a 76% que a 33% de H. R.
5.2.9.4.3. Luminosidad
Factores de tipo abiótico como la luz solar, la temperatura, el agua o la humedad y las sustancias químicas, determinan el éxito o fracaso de un agente de control biológico en el c~mpo. La radiación solar, especialmente la del tipo ultravioleta, desempeña un papel importante en la mortalidad de esporas de hongos. La vida media de los diferentes tipos de inóculo de hongos expuestos a la luz solar se estiman en un tiempo aproximado de una hora para el hongo entomopatógeno más susceptible y de 36 horas para el más resistente.
- Paíricia Elizabeth Bmim Guevara 57

I
La luz solar puede inactivar a los entomopatógenos en forma directa a través de delecciones, uniones cruzadas, ruptura de bandas y formación de sitios lábiles en la molécula del DNA; y en forma indirecta, debido a la formación de radicales altamente reactivos los cuales a su vez inactivan a los entomopatógenos reduciendo su persistencia en el campo vele2 y Montoya, 1993).
La radiación ultravioleta B (uv-B; 280-320 nm) es un componente detrimental de la luz para todos los micmrganismos. Un número de sustancias han sido usadas para proteger y aumentar la persistencia de los entomopatógenos.
El impacto de la radiación uv-B sobre poblaciones fungales en habitats naturales no ha sido extensivamente estudiado. B. bassima, un hongo hialino, es endémico del suelo y es raramente aislado del foliaje de plantas. Posteriormente, la sobrevivencia de la conidia de B. bassima aplicado sobre foliaje es muy pobre, y el parámetro más importante limitante de la sobrevivencia de las wnidias en hábitats epígeos es la luz. La radiación ultravioleta causa daños primarios (ej. mutaciones en el ácido nucléico) y secundarios (ej. fotorreacciones) al exponer a los micmrganismos, los cuales pueden conducir a muerte celular. Observamos que las conidias de B. bassima son altamente sensitivas a la radiación artificial de uv-B pero el mecanismo que causa la muerte es desconocido. Al parecer la extrema sensibilidad de 8. bassima a la porción uv-B del espectro solar limita su persistencia en hábitats epígeos. La formulación de B. bassiana provee protección de la luz ultravioleta la cual será necesaria para incrementar su sobrevivencia y eficacia en hábitats epígeos.
El rojo Congo, y todos los abriiiantadores probados en un experimento proveen un grado de protección de la radiación artificial uv-B. Los adyuvantes se probaron en el campo pero sólo la arcilla y el abrillantador edbeno, tnopal LPW, incrementaron consistentemente la persistencia de la conidia de B. bassima sobre hojas de trigo.
Los bloqueadores de luz solar han provisto efectiva protección a los hongos de radiación uv-B, y el uso de bloqueadores tales como arcilla, almidón, o carbono pueden ser preferibles debido a que son inocuos al medio ambiente o fácilmente descompuestos. Varios entomopatógenos, incluyendo B. bassiana, han sido aplicados en aceite a ultra bajos volúmenes WLV) en intentos para incrementar su eficacia. %dios en campo, no encontraron incremento en la sobrevivencia de las conidias de B. bassima aplicadas en aceite en comparación con las conidias asperjadas a ULV en agua a volúmenes convencionales. Sin embargo, en el experimento de laboratorio, se observó que las conidias de B. bassima expuestas a radiación uv-B sobreviven mejor en aceite que en un vaso con agua. Sin embargo, la utilización de protectores uv-B para incrementar la eficacia del control de insectos dependerá de los protectores solares para prolongar la sobrevivencia de la conidia suficientemente para aumentar su eficacia.
En campo, Daoust y Pereka (1986) citan que los conidios de B. bassima sobreviven por mucho tiempo, ya que el hongo en condiciones de campo se mantiene protegido en cadáveres de insectos sin pérdida de la viabilidad por 16 semanas y con una leve dis&uciÓn del poder germinative después de 24 semanas. Aunque en regiones templadas se señala que la vida media de las esporas sobre el follaje se reduce a dos días e incluso a menos de 24 horas, a los cinco o diez días después de la aplicación las esporas pierden el 100% de viabilidad en formulaciones en polvo, empleo de aditivos, utilización de una monocapa que cubra las esporas, la aplicación del hongo al atardecer y la selección, mediante técnicas de ingeniería genética, de genes que transfieran una coloración oscura a cepas prometedoras como micopesticidas, ya que especies como AspegiIlus niger con esporas oscuras muestran una vida media de 14.8 días comparada con 1.9 días de B. bassima con luz solar.
Patricia Elizabeth Baños Guevma 58

La luz solar es detrimental para los conidios, cualquiera que sea el tiempo de exposición. Sin embargo, los conidios protegidos sobreviven por mucho tiempo, ya que el hongo se encuentra dentro de los cadáveres de insectos en condiciones de campo se mantiene estable sin pérdida de viabilidad hasta por 16 semanas y con una leve disminución del poder genninativo después de 24 semanas (Damst y Pereira, 1986).
5.2.9.4.4. Agua
La mayoría de las especies de hongos entomopatógenos son hidmfílicos, el agua liquida o en vapor han sido reconocidas como esenciales para la genninación de esporas y además la alta humedad relativa es conocida por favorecer el desarrollo de una micosis epizootics (Ferrun, 1978).
Un p m s o en cultivo sólido puede ser definido por el crecimiento microbiano en partículas sólidas sin la presencia de agua libre. La presencia del agua en sistemas de cultivo sólido existe en una forma compleja dentro de la matriz del sólido o como una delgada [xipa que es absorbida sobre la superficie de las partículas o perdida fuertemente en los límites de las regiones capilares del sólido (Mudgett, 1986, citado por Doelle et al., 1992). El agua lih solo ocurre una vez que se ha excedido la capacidad de saturación de la matriz del sólido. El nivel de humedad a la cual el agua libre parece variar considerablemente entre sustratos, sin embargo, es dependiente de las características propias de su agua. Para muchos sustratos al alcanzar el nivel de humedad del 80% se presenta el agua libre. Los niveles de humedad en procesos de cultivo sólido generalmente varían entre 30 y 85%. El cultivo de micmrganismos a altos niveles de humedad puede provocar que los espacios entre las partículas sean llenados por agua cuyos límites de intemmbio de gas y rendimientos del sustrato los hacen más vulnerables a contaminación bacterial.
En otro aspecto, a bajos niveles de humedad se reducen: el crecimiento microbiano, la estabilidad enzimática, el hinchamiento del sustrato y la difusión de nutrientes. El contenido óptimo de humedad para el cultivo de micmrganismos en procesos de cultivo sólido es altamente dependiente de las propiedades de captación de agua del sustrato.
Actualmente se acepta que los requerimientos de agua de los microorganismos deben ser definidos en términos de la actividad de agua (Aw) del ambiente, que está mejor descrito que el contenido de humedad del sustrato sólido. Este parámetro está definido por la relación entre la presión de vapor de agua en el sustrato (p) y la presión de vapor del agua pura (p.) y a la misma tempetatura, A v PIP.. Este concepto está relacionado a otros parámetros tales como la humedad relativa (% H. R = 100 x Aw) y el potencial de agua (psi = RT/V ln Aw, donde R es la constante del gas ideal, T es la temperatura absoluta y v es el volumen en moles de agua) (Griffin, 1981, citado por Doelle et al., 1992).
La Aw del agua pues es 1.00 y la de una solución 1 molar del soluto ideal sería 0.9823. La Aw estará en equilibrio con una humedad relativa (H. R) de la atmósfera en torno al alimento 100 veces superior si la H. R se expresa corno porcentaje. Cuando la humedad relativa que rodea al alimento corresponde a una Aw inferior a la del propio alimento tenderá a desecar su superficie; y a la inversa, cuando la H. R es mayor que la Aw del alimento esta tenderá a aumentar en la superficie de dicho alimento (Frazier, 1976).
Cada microorganismo tiene una Aw máxima, Óptima y mínima para su crecimiento. Los factores que pueden afectar las necesidades de Aw de un microorganismo son:
Patricia Elizabeth Bañas Guevara 59

REVISIÓN DE LiTERATURA
1) La clase de soluto empleado para reducir la Aw. En númerosos micoorganismos y especialmente en los hongos, la Aw mínima para el crecimiento es prácticamente independiente del soluto utiiizado, otros microorganismos en cambio, ofrecen diferentes Aw m í n i m a s , de acuerdo con el soluto utilizado.
2) El valor nutritivo del medio de cultivo. En general, cuanto mejor sea el medio de cultivo más baja será la Aw mínima.
3) La temperatura. La mayor parte de los microorganismos toleran mejor bajas Aw a la temperatura óptima.
4) SuminiStt.0 de oxígeno. El crecimiento de los microorganismos aerobios tiene lugar a una Aw más baja en presencia de aire que en su ausencia; lo contrario ocurre con los anaerobios.
5) pH. La mayor parte de los microorganismos toleran Aw más bajas a valores de pH prúxitnos a la neutralidad que en medios ácidos o alcaiinos.
6) inhibidores. La presencia de inhibidores acorta el intervalo de Aw que permite el crecimiento de los microorganismos.
Para el control de Aw se pueden utilizar los siguientes métodos: (1) equilibrio con disoluciones control, (2) determinación de las isotennas de absorción de agua de los alimentos y (3) adición de solutos (Frazier, 1976).
Para la mayoría de hongos filamentosos la óptima actividad de agua para crecimiento está entre 0.93 y 0.98, aunque algunas especies crecen mejor a altos valores. La mínima actividad de agua para crecimiento tiene rangos de 0.7 a 0.94. La mayoría de los hongos filamentosos crecen razonablemente bien a bajo pH. Esto es ventajoso ya que el controldel pH es muy difícil en cultivo de sustrato sólido y el metabolismo del microorganismo a menudo causa acidificación.
Grandes cambios en el contenido de humedad en estado sólido pueden resultar de pequeños cambios en la humedad relativa de la fase gaseosa en equilibrio con los sólidos, dependiendo de las características de sorción-desorción del sustrato. La cantidad de agua producida por el metabolismo, sin embargo, es insuficiente para reemplazar el agua perdida por evaporación durante la aireación fonada (Mudget, 1986, citados por Doelle et al., 1992).
5.2.9.4.5. Oxígeno
La mayoría de los honxos son aerobios y B. bassima no es la excepción, aunaue tiene características facultativas, es decir, se adapta a condiciones parc&hente anaerobias (Frazier, 1976).
En trabajos de conducta con la fisiología de B. bassiana se prestó atención al problema de su respiración, la cual estudiada en conexión con el desamlio del hongo, podría ser un factor indicativo de la calidad infectiva del material.
El estudio de respiración en hongos ha sido una materia bastante complicada. La naturaleza de las sustancias de reserva no está completamente entendida, y los sustratos endógenos no se han usado uniformemente baja condiciones experimentales variables.
Patricio Elizabeth Baños Guevora 60 - - --

otro problema es la participación de otros caminos en la utilización de sustratos endógenos. Como los hongos entomopatógenos no han sido todavía investigados desde este punto de vista, el propósito fue averiguar el curso de la actividad respiratoria con relación al desarrollo de B. bassima y entonces determinar la posibilidad de usar diferentes azúcares como fuentes efectivas de ca&xjn durante el cultivo del hongo.
Durante un período de 72 horas de cultivo bajo submersión, el curso del oxígeno tomado (QOZ) por el hongo entomopatógeno B. bassima fue investigado por medio de la convencional técnica manométrica, ambos en la presencia y ausencia de sustratos de azúcar exógenos. Los valores absolutos de QOz disminuyeron con la edad de cultivo. En los sustratos exógenos probados, el mejor para la respiración fue la fructosa, seguida por el azúcar invertida y la glucosa, pero no la sacarosa. La fructosa, cuando estuvo presente en el medio de cultivo inicial, indujo un incremento en el peso seco del hongo y en la virulencia del hongo al ser probado por inyección de la larva de la mariposa nocturna Galleria mellonella (Samsinakova y Hrabetová, 1969).
5.2.9.4.6. p H
La mayoría de los hongos pueden crecer en un intervalo de pH muy amplio (2 a 8.5), pero casi todos lo hacen mejor a un pH ácido. En el caso de B. bassima se desarrulla fácilmente en un amplio rango de pH: 5.5. a 7.0 (Frazier, 1976; Rojas, 1982a).
El hecho de que a determinado pH, la germinación de esporas pueda ser estimulada o abatida depende de la naturaleza del medio de cultivo el cual puede tener implicaciones prácticas y debe considerarse cuando se formulen las preparaciones de esporas para su aplicación en campo. Es interesante notar que las conidias sumergidas germinan más rápidamente que las conidias aéreas a valores de pH de 3.5 a 6.0 en ambos tipos de cultivos: sólidos y líquidos (Thomas et al., 1987).
Para el éxito del cultivo liquido el pH inicial es importante, en un experimento el Óptimo pH al inicio del proceso fue 4.5; como el crecimiento del hongo se incrementó el pH se elevó a 5.1 y al finalizar el cultivo fue alrededor de 5.0 (Samsináková, 1966).
5.2.9.4.7. Nutrientes
En general los hongos utilizan diversos tipos de alimentos, tanto sencillos como complejos. Generalmente poseen gran cantidad de enzimas hidrolítica y algunos se cultivan para obtener sus a d a s a s , pectinasas, proteinasas y lipasas (Frazier, 1976).
El pnxeso de germinación ocurre en la presencia de una fuente utilizable de carbono y una fuente de nitrógeno es necesaria para crecimiento hifal. Los requerimientos para el mimiento de B. bmima no son complejos; en realidad, los componentes de la fuente de energía de carbono tienen gran variabilidad química (de glucosa a una cera compleja) que producirán un extensivo crecimiento del hongo. Los compuestos que pueden servir como fuentes de carbono y energía para la germinación incluyen, glucosa, N- acetilglucosamina, glucosamina, quitina, almidón, lanolina, carbohidratos en aceite crudo y algunos ácidos grasos de cadena larga (Smith y Grula, 1981). Como fuente de carbohidratos, B. bassima utiliza, D-melicitosa, pero esporula mejor sobre D-sacarosa, D- glucosa y D-trehalosa; es necesario mencionar que este último compuesto es el mayor componente de la hemolinfa de los insectos (Campbell et al., 1983).
Patricia Elizabeth Bañar Guevma 61

1
REVISIÓN DE I_ LITERATURA
En otros estudios evaluaron 2 1 fuentes de peptonas como recurso para el crecimiento y esponilación de B. bmim4 los productos m á s sobresalientes fueron: soytone, casitone, nwpeptona y caseina hidrolisada. También se han estudia& varios aminoácidos como fuentes de nitrúgeno, dentro de los cuales dedacan el triptofano y la ala-, aunque con ácido glutámico y KNOs (Nitrato de potasio) también se produjo una cantidad regular de esporas.
B. bassima se puede cultivar en los siguientes medios de cultivo:
* De laboratorio. Las conidias a6reas del hongo entomopatógeno B. bassima se pueden cultivar en medios basados únicamente en agar de Saboraud, maltosa, malta, dextrosa (Samsináková, 1966; Marcandier y Khachatourians, 1987).
Un medio de agar empleado para la producción de B. bassima es: 0.39 KHzo4; 1.42 NazHP04.12Hz0; 0.60 MgS04.7HzO; 1.00 KCI; 0.70 “SIOS; 10 glucosa; 5 extracto de levadura; 20 agar; suplementado con 0.5 cloranfenicol en g/l, e incubado a 25OC por 3 semanas. Las conidias fueron entonces raspadas de estas placas y propagadas en cajas Petri de 90 mm, e incubadas por 3 días a 25°C. Este cultivo fue usado para los estudios de crecimiento radial (Fargues et al., 1997).
Ling, Donaldson modificaron un medio selectivo de agar y contiene Saboraud Maltosa Agar suplementado con 1% de extracto de levadura, 4 5 5 mg/l de “dodine”, 0.1 g/l de penicilina, y 0.5 g/1 de sulfato de streptomicina (Kmeger et al., 1992).
Fargues y Robert (1985) emplearon el medio semisintético de gelosa, que comprende: 0.39 g de KHZO,, 1.42 g de NHzHPOI, 0.60 g de MgSO4, 7Hz0, 1 g de KCl, 10 g de glucosa, 0.70 g de NH4NOs, 20 g de agar, suplementado con 5 g de extracto de levadura, en 1000 ml de agua destilada, pH de 6.5, incubación por 3 semanas a 24 +- l0C y fotoperíodo de 16 horas luz y 8 horas de obscuridad.
* Sustratos de origen agrupecuano. Kucera (1971) al milizar una investigación sobre producción de toxinas por el hongo B. bassima utilizó 24 fuentes de nitrógeno, incluyendo nitrúgeno inorgánico, nitrúgeno orgánico no protéico y examinó medios naturales: esquilmo de trigo, salvado de trigo, harina de maíz, harina de frijol, salvado de frijol, esquilmo de chícharo, levadura seca, extracto de levadura, nata de leche y extracto de carne de res, para determinar el efecto estimulante sobre la producción de una toxina de B. bassima Las mejores fuentes fueron la harina de maíz, el extracto de levadura y el extracto de carne de res. La producción óptima de esas fuentes ocurrieron al tercer día de fermentación, la composición de esta fuente de proteasa puede ser intluenciada por la fuente de carbono.
En Guatemala se realizaron estudios con medios como: maíz cocido, arroz cocido, leche en polvo disuelta en agua y &da, micosel m á s agar, arroz con leche cocidos, copra de coco, agua de coco y salvado de trigo. De estos medios se tuvieron resultados positivos en cuanto a multiplicación y reproducción de esporas en todos, menos en agar más micosel. En donde hubo mejor cultivo de B. bassima en forma pura fue con los medios de arroz cocido y copra de coco (Rojas, 1982a).
La producción de hongos, generalmente ha sido a partir de cereales, se utilizancomo sustratos sólidos diferentes tipos de grano; mismos que proporcionan el sustento para su reproducción. Diversos estudios determinaron que el trigo, maíz, arroz y sorgo son alternativas para la producción masiva de B. bassima (Rojas, 1982a, b; Woods y Gnila, 1984; Gana, 1995). -
Patncia Elizabeth Bañar Gumma 62

o t r o s autores (Samsináková, 1966; Marcandier y Khachatourians, 1987) reportan haber trabajado con: papa, gahanzo, trigo, ~ j o l , avena, arruz, ma&, salvado, etc., y todavía se pueden enriquecer suplementándolos con extracto de levadura (0.2 gíl) y tintura de rosa de bengala (0.66 gíl).
La limitación del crecimiento por nutrientes es usualmente más severo en sustrato sólido debido a los limitados rangos de difusión de nutrientes y el limitado acceso del microorganismo a ellos. De particular importancia es la razón carbono:nitrógeno (C:N y el óptimo valor puede variar sobre un amplio rango de I O a 100 en diferentes cultivos de estado sólido, aunque la aprovechabilidad del cahono y nitrúgeno es también importante y puede afectar el rango óptimo.
La esporulación de los hongos filamentosos es también regulado por efectos metabólicos causados por la aprovechabilidad de nutrientes. Moo-Young y colaboradores (1983, citados por Doelle et al., 1992) apuntaron que la esporulación fue el factor limitante en la producción de biomasa fungal y que los cambios ambientales del cultivo sólido afectan la espontlación aumentando debido a la poca difusión y limitada convección de un microorganismo esencialmente inmóvil.
5.2.9.4.8. Control de contaminan tes
En un proceso de cultivo de hongos filamentosos, la contaminación bacterial puede ser minimizada o prevenida usando un pH bajo. La literatura indica que el pH debe mantenerse abajo de 4 para controlar la contaminación bacterial. Por la combinación de un bajo valor de pH con otras condiciones ambientales desfavorables para crecimiento bacterial, la presiónselectiva contra bacterias puede hacerse m á s rigurosa. Los antibióticos pueden ser efectivos en cultivo sólido para control de crecimiento bacterial (Doelle et al., 1992).
5.2.9.4.9. Viabilidad
Los diferentes factores ambientales incluyendo temperatura, humedad, luz y gases (oxígeno y dióxido de carbono) Ilifluyen en la estabilidad de los micmrganismos entomopatógenos y en la viabilidad de los conidios; las condiciones de almacenamiento deben ser específicas para conservar estas características. B. bmiana puede perder hasta un 50% de viabilidad entre 5 a 10 días después de haberse aplicado al follaje (Walstad et al., 1970).
Steinhaus (1960, citado por Méndez, 1990) concluye que hay una declinación muy notable en la viabilidad de los conidios a temperaturas superiores. Los puntos críticos para la mortalidad de los conidios de B. bmiana y M. anisophae se ubican entre los 45 y 4 9 ~ , respectivamente, temperaturas entre 35 a 49OC retienen el proceso de germinación.
La germinación de los conidios es un suceso natural bajo ciertas condiciones, afectado especialmente por la variación de la temperatura, ya que reduce la viabilidad de las unidades infectivas de B. bawana También se ha demostrado que a medida que la temperatura decrece de 37 a 4OC, la viabilidad de las unidades infectivas se incrementa notablemente (Damst et al., 1983). El poder germinativo de las esporas de 8. bmiana mantenidas a 4OC se conserva por 120 semanas, mientras que a 23OC se pierde después de 6 a 13 semansa y a 3OoC después de 4 a 7 semanas de almacenada, el rápido decaimiento en la genninación de las esporas puede evitarse por congelamiento y almacenado en seco. Patricia Elizabeth Baños Guevma 63

REVISIÓN DE LITERATURA
Walstad et al. (1970), mencionan que la temperatura letal para los conidios de B. bassima está cercana a los 5OoC, también establecen que los wnidios de B. bassiana y M. misopIiae almacenados a 21"C sobrevivieron poco tiempo de 0.5 y 2.5 meses respectivamente, mientras que los conidios almacenados a 8OC permanecieron viables mínimo un año. Los conidios pulverizados para su secado a 5OoC y cubiertos con bentonita tienen de 50 a 70% de viabilidad, en tanto que las blastosporas que también son sensibles a esta técnica fueron liofilizadas y mezcladas con leche en polvo suplementadas con glicerina, la viabilidad de las esporas aumentó en 89%; el almacenamiento de estas preparaciones se efectuaron a una temperatura de 5OC y la viabilidad de los conidios se conservó por un lapso mayor a los 18 meses de almacenamiento.
Generalmente se cree que la patogenicidad es retenida después del almacenamiento de los conidios bajo ciertas condiciones que permiten la retención de la viabilidad, sin embargo se ha reportado que en B. bassima la capacidad infectiva decrece más rápidamente que la capacidad de g e d a c i ó n (Daoust et al., 1983).
5.2.9.5. Patrones de crecimiento
La falta de entendimiento de los patrones de crecimiento y cinética de crecimiento en cultivo sólido es debido a tres problemas:
a) La complejidad del arreglo tridimensional del micelio y el sustrato b) La heterogeneidad del sustrato mismo. c) La dificultad en la obtención de medidas directas de la biomasa.
La caracterización de la cinética de crecimiento y el desarrollo de modelos matemáticos tienen varias consecuencias benéficas:
a) La posibilidad de predecir la fabricación del reactor, incluyendo rangos, concentraciones, pxwductividades y tiempos óptimos de cosecha.
b) incorporación de los efectos de los factores físicos externos y nutricionales ayudando así a optimizar el proceso.
c) Modelos mecánicos para entender principalmente los procesos fundamentales involucrados en el crecimiento en cultivo sólido, incluyendo los fenómenos físicos y biológicos.
Los hongos fiíamentosos son el grupo m á s importante de micxuorganismos que se cultivan en sustratos sólidos. Entre sus ventajas se encuentran su modo de crecimiento hifal; sus patrones de crecimiento son más complejos que los de las colonias bacteriales que crecen sobre cultivo de superficie. Un hecho específico del crecimiento fungal sobre sustratos sólidos el cual ha recibido alguna atención es la penetración, facilitada por grietas o fisuras existentes en la superficie del insecto a través de la cual es capaz de accesar al interior y remover la barrera física a la penetración (Doelle et al., 1992).
P
Pairicia Eiizabeih Baños Guevara 64

REVISI~N DE LITERATURA - 5.2.9.5.1. Degradación del almidón por microorganismos
Muchos micmorganismos son capaces de hidrulizar el almidón y pueden por lo tanto aprovecharse para aplicaciones en cultivo sólido. De las vanas enzimas que degradan el almidón se encuentran la a-amilasa, la glucoamilasa, la pululanasa y la isoamilasa; solo la a-amilasa y la glucoamilasa son importantes en aplicaciones de cultivo de sustrato sólido.
Los micwrganismos c m n generalmente mucho mejor en cultivo sólido con almidón gelatinizado que con almidón no gelatinizado como sustrato. Sin embargo, una considerable cantidad de energía es requerida para la gelatinización y podrían, por lo tanto, ser ventajosos los organismos que crecen bien sobre almidón crudo (no gelatinizado). La degradación de almidón crudo se incrementa grandemente por la acción sinérgica entre a- a d a s a y glucoadasa (Lbpez-Ulibarri et al., 1989, citados por Doelle et al., 1992).
5.2.9.5.2. Productos metabóiicos de la fermentación sólida
* Enzimas. Para aprovechar la alta producción de enzimas de B. bassiana, se recomienda en algunos casos el cultivo sobre sustratos sólidos naturales. Se han usado como buenas fuentes de nitrógeno: harina de cacahuate, salvado de arroz, harina de soya, extracto de levadura, peptona y caseína ácida hidrulizada, aminoácidos puros, medio de hígado, y nexayote del proceso de nixtamalización del maíz. Se relacionaron varias fuentes de nitrúgenos como posibles estimulantes de la formación de proteasa tóxica, tales como: nitrúgeno inorgánico, orgánico no protéico, proteínas y medio de complejos naturales (Kucera, 1971).
* Ácidos metabólicos. B. bassiana también produce ácidos metabólicos, los cuales se detectan en cajas de Petn con medio de agar al cual se le ajusta inicialmente el pH a 5.5 con HCl lM, se incuban a 2 7 T y las wnas amarillas que aparezcan alrededor de las colonias indican la presencia del ácido metabólico, el cual tiene un pH menor a 4 (Frazier, 1976; Bidochka y Khachatounans, 1993). Ciertos hongos elaboran sustancias que son inhibidoras para otros micmorganismos, algunas sustancias son micostáticas, es decir, inhiben el crecimiento de los hongos; tal ocurre con el ácido sórbico, propionatos y acetatos; y otros son específicamente fungicidas, pues matan a los hongos.
El pH de un cultivo puede cambiar en respuesta a actividades metabólicas microbiales debidas a diversas rawnes. La secreción más obvia de los ácidos orgánicos es el ácido acético y láctico, el cual causa decremento del pH. En otro aspecto la asimilación de ácidos orgánicos puede presentar en ciertos medios un incremento en el pH. La tendencia del pH se incrementa si los componentes orgánicos amino son degradados cuando se usan para crecimiento.
En cultivos en estado sólido es muy dificil controlar el pH; por lo tanto es deseable que el microorganismo usado deba tener un amplio rango de pH y un pH Óptimo para crecimiento. Los hongos filamentosos pueden crecer dentro de un rango de pH de 2 a 9, con una región Ó p h a de pH entre 4 y 6 (Mitchell, 1992, citado por Doelle et al., 1992).
* Transferencia de calor. Debido a la alta concentración de sustrato por unidad de volumen, la cual es tipica de los procesos de estado sólido, la generación microbiana de calor por volumen de unidad es mucho más grande que para cultivos iíquidos.
- Patricia Elizabeth Bañar Guevara 65
,

REVISIÓN DE LITERATURA - El rango de calor metabólico generado puede ser aproximado al rango de evolución
de dióxido de carbono (COZ). De acuerdo a Reid (1989, citado por Doelle, 1992) por cada mol de COZ producido durante la oxidación de lignina implica la relación de 475 kJ de energía calorífica. El rango de transferencia de calor puede ser iimitado por los rangos de transferencia intra e interparticula, por el rango en el cual el calor es transferido de la superficie de la partícula a la fase gaseosa, o por el rango al cual el calor en la fase gaseosa es removido. ias características térmicas del material orgánico y el bajo contenido de humedad en cultivo sólido c m n condiciones especialmente difíciles para la transferencia de calor.
* Oxígeno y dióxido de carbono. El consumo de oxígeno y la evolución del dióxido de carbono resultan de la respiración, que es un proceso metabólico por el cual los micmrganismos aerobios derivan la mayor parte de su energía para crecer. Mas actividades metabóiicas son por lo tanto asociadas al crecimiento y pueden ser usadas para la estimación de la producción de biomasa.
La aireación satisface 4 funciones principales en procesos de cultivo sólido: (1) mantener las condiciones aekbicas, (2) para la deserción de dióxido de carbono, (3) para regular la temperatura del sustrato, y (4) regular el nivel de humedad.
El rango de aireación óptima en procesos de estado sólido debe ser determinado por la naturaleza del microorganismo usado, el requerimiento particular de oxígeno para crecimiento y síntesis de producto a partir de nutrientes, la cantidad de calor metabólico a ser disipado, el grado en el cual el COZ y otros metablitos volátiles deben ser eliminados, el espesor del sustrato y su densidad (inversamente relacionado por el volumen del espacio del poro) y el contenido de humedad. En algunos casos un alto rango de aireación es esencial para una productividad Óptima (Lonsane et al., 1985, citados por Doelle et al., 1992).
5.2.10. Escalamiento piioto de producción de B. bassiana en cultivo sólido
Para la técnica de producción masiva de un hongo entomopatógeno de caractensticas óptimas, es necesario considerar aspectos tales corno (Samsináková y Kálalová, 1981; Woods y Gmla, 1984):
a) Selección de cepas capaces de producir una gran cantidad de esporas suficientemente virulentas.
b) Seleccionar el medio más económico y efectivo
c) Fácil producción a bajos costos, es decir, evitar la complicación de la técnica de producción del patógeno.
d) Que éste consewe su potencialidad de germinación.
e) Desarrollo óptimo en el medio sintético
0 Optimización en su espomlación.
9, Diseñar bien los procedunientos de la formulación y condiciones de almacenamiento para que en la elaboración del biopreparado, el hongo no pierda su virulencia al ser utilizado en campo.
Patricia Elizabeth Baños Guevara 66

Debido a su aplicación potencial contra una extensa variedad de insectos, hay un enorme interés en la producción a gran escala de este hongo para su aplicación en campo. Además de que B. bmima es un hongo que se adapta fácilmente a la manipulación del laboratorio y es por lo tanto susceptible para producción masiva, los productos formulados de mnidias son comercialmente aprovechables. En contraste un pequeño esfueno ha sido invertido en la producción en masa de B. bmngniartii (Samsináková y Kálalová, 1981; Thomas et al., 1987; Aregger, 1992).
Se han realizado investigaciones para el uso de materiales baratos con garbanzo, salvado, y materiales de wmposta bajo condiciones semiestériles. Estos experimentos han mostrado, siempre que B. bmima requiere condiciones estériles, y su crecimiento es lento, las contaminaciones por bacterias y/o hongos saprofíticos pueden desarroliarse rápidamente y crecer sobre el hongo entomófago (Samsináková y Hrabetová, 1969).
En la actualidad el grano de arroz es el demayor uso, ya que éste produce el mayor rendimiento de material infectivo y conserva las condiciones de viabilidad, patogenicidad y pmducciónde volúmenes altos de esporas libres. En unidades de producción de hongos entornopatógenos, el arroz puede ser la alternativa más confiable para la reproducción masiva, ya que este puede ser de primera o segunda (Gam, 1995).
En México, el hongo entomopatógeno B. bassiana se produce en el Centro Nacional de Referencia de Control Biológico DGSV-SAGDR, bajo el sistema tradicional de cultivo empleando grano de arroz pulido y obteniendo rendimiento de 1.6 x 10’ conidiadg, con una estabilidad confinnada de 40 días a 2OC formuladas en aceite de coco (Prior et al., 1988) o 6 meses a 7OC en una formulación de diatomita o bentonita (Berlanga, 1995; Hernández y Carrillo, 1997).
En Colombia se han desarrollado métodos para la producción masiva de estos hongos tanto a nivel artesanal, para ser utilizados por agricultores, como a nivel industrial, produciendo formulaciones en cooperación con la industria privada; actualmente hay cinco laboratorios que producen este insecticida biológico comercialmente (Bustill0 y Posada, 1997).
La metodología para producir B. bassima en las fincas de los cafeticultores es muy sencilla, y se utiliza un sustrato de arroz y agua que se introduce en botellas desechables de vidrio, a las cuales se les pone un tapón con algodón absorbente y se sometan a un proceso de esterilización en “baño maria” (Antía et al., 1992, citado por Bustillo y Posada, 1997). La producción de conidias en estas botellas, después de 24 días y a una temperatura de 25OC es de 4 x 10” wnidiadl00 g de sustrato. La producción de una botelia es suficiente para asperjar 100 áxúoles a una dosis de 5 x 10’ conidiasíáxúol. Durante los t res últimos años, Cenicafé y el Servicio de Extensión de la Federación Nacional de Cafeteros de Colombia, han entrenado en esta técnica a más de 20,000 agricultores, muchos de los cuales están produciendo el hongo eficientemente.
CENICAFE también ha puesto a disposición del cafetero una unidad de producción masiva del hongo B. bmima, usando metodología artesanal; en ésta se capacitan cafeteros y personas interesadas. Además, se suministra gratuitamente el hongo denominado “Czpa CENICAFE” para su reproducción. De 1994 a 1997 se han pmducido 60 toneladas de esta cepa, la cual se ha distribuido a los cafeteros.
Pamcia Elizabeth Bañar Gumma 67

Las formulaciones del hongo producido con esta metodología se somete a control de calidad, para asegurar que esté libre de contaminantes, que tenga la concentración apropiada, que su viabilidad sea del 1004, que posea una patogenicidad sobre broca (en laboratorio) superior al 80% y que provenga de una cepa del hongo recién activada y de buen comportamiento en el campo. ia anterior asegura al cafetero la producción de un hongo de excelente calidad para el control de la broca. Este protocolo ha servido de base para la reglamentación de la producción de entomopatógenos en el país por parte del ICA (instituto Colombiano de Agricultura) (Bustill0 y Posada, 1997).
A nivel industrial (Morales et al., 1991), la producción de este hongo se inicia con cultivos puros aislados de broca, los cuales se colocan en cajas Petri con medio SDA, luego este inóculo se emplea para el cmimiento del hongo en frascos que contienen un medio líquido, nutritivo y aséptico, el cual es mantenido bajo condiciones de fermentación y agitación a 110 rpm durante 72 horas. Este cultivo produce blastosporas, que sirven para inocular bandejas con un sustrato líquido químicamente definido para la producción de conidias aéreas. Después de 15-20 días (dependiendo de la temperatura) el hongo está listo para ser cosechado, homogeneizado, formulado y secado (polvo). Esta tecnología ha sido transferida a productores particulares para que se encarguen de la producción industrial del hongo. En la actualidad existen cinco compañías en Colombia con licencia del instituto colombiano de Agricultura (ICA), que suministran hongo formulado para control de la broca.
En Colombia, a nivel nacional, durante 1992 se utilizaron cinco toneladas del hongo, en una concentración de 1 x 108 conidiadg, con fines experimentales. En 1993, la producción fue de 60 toneladas (Posada, 1993, citado por Bustillo y Posada, 1997); en 1994 se estimó en 100 toneladas, y en 1995 llegó a 200 toneladas. A esta producción se debe adicionar la de varias unidades auspiciadas por los comités Departamentales o las Cooperativas Cafeteras en los departamentos de Antioquía, Caldas, Risaralda, Qukdío, Nariño y Valle del Cauca.
En México, los hongos producidos son entregados directamente a los productores, con lo cual automáticamente quedan como centros regionales y no se complican con el sistema de distribuidores, lo cual encarece el costo del p d u c t o . La carencia en México de una formulación, que le proporcione estabilidad a largo tiempo a los hongos, es otra de las razones por la cual los agentes producidos deben ser empleados en un tiempo relativamente corto después de su cultivo.
La mayoría de los laboratorios de producción, lo formulan sólo con talco, que es normalmente vehículo y prutector solar, por ser reflejante de la radiación solar. Esta es una situación nueva para México, ya que hasta 1984 se publicó la primera lista de productos autorizados para el empleo agrícola, donde existía el primer microorganismo como &@ente activo en una formulación: la bacteria Bacillus thuringiensis BERLINER. La legislación mexicana en materia de plaguicidas no tenía contemplado en las condicionantes una sección para la autorización de productos de esta naturaleza; hoy con la participación de la Sociedad Mexicana de Control Biológico en el Comité de Evaluación de productos para uso Agrícola, diversos puntos de vista sobre estos aspectos han cambiado (Hernández, 1995).
P - Patricia Elizabeth Baños Guevma 68

5.2.10.1. Formulaciones comerciales de B. bassiana El control de calidad de estas preparaciones, al menos de Beauveria, necesita del USO
de diversas técnicas incluyendo el conteo de esporas, estimación de la germinación, y estudios sobre su vinilencia. La concentración de esporas puede ser determinada pot’ un hemocitómetro después de la homogeneización del material y eventualmente la coloración de esporas. La germinación de esporas es estimada por técnicas de dilución sobre medios nutritivos en cajas de Petri o por técnica de cultivo en portaobjetos y en el microscopio. El contrul de la virulencia se presenta con el objeto de comparar estudios relacionados al descubrimiento de una tecnica que podría ser adoptada por todos los laboratonos involucrados. El problema de la especificidad de la cepa es mencionada previamente y la falta de referencias estándares de las preparaciones tienen que ser resueltas todavía (Femn, 1978).
Los resultados con los entomopatógenos tienden a mostrar que la eficacia se mejora al aumentar la dosis; esto crea la necesidad de desarrollar formulaciones de insecticidas biológicos mejores y más económicas. En café se recomienda usar B. bassima en dosis de 1 x 103 conidiasíárboi, en una formulación que incluya un aceite emulsificable, con el fin de lograr mayor control de la broca (Bustillo y Posada, 1997).
Los productos comerciales a base de B. bassima deben protegerse al ser aplicados en campo con una formulación apropiada que contenga protectores solares, que prolonguen su estabilidad sólo por períodos cortos de tiempo. El uso de cebos y otros sustratos que puedan proteger de los factores climáticos debería de ser probado posteriormente en estudios de patogenicidad en campo para confirmar estos resultados (Daoust y Pereira, 1986).
También hay necesidad de mejorar la eficiencia de la tecnología de aspersión. En Colombia, Cenicafé con la cooperación de la compañía inglesa Micron desarrolló recientemente el equipo Motax para aplicaciones a bajo volumen. En evaluaciones realizadas en condiciones de campo, se informó que este equipo produce gotas menores a 100 m, con una cobertura entre 79 y 179 gotas/cmz. El volumen de aspersión por ha fue de 56 y 69 litros y el rendimiento de aplicación por día fue de 7142 árboles sembrados a 1 x 1.4 m. Como se ha informado este equipo muestra alta eficiencia en la aspersión del hongo B. bassima Estos resultados son muy importantes, no sólo para mejorar la eficacia física y biológica de las aspersiones, sino para reducir los costos de las mismas (Bustill0 y Posada, 1997).
En la ex-URSS, la preparación de conidióforos de B. bassima llamado Beauverin o &ve& cuya concentración es de 2 x IO9 conidiasíg y lo estabaza un sustrato inerte de caolín fue la primer formulación de una preparación entomopatógena, la cual en aplicaciones generalizadas se encontraron problemas tecnológicos de producción masiva del principio activo de alta calidad. En Europa se han distribuido dos preparaciones denomhdas Boverol y Boverosil (C-SFR) (Aregger, 1992).
Actualmente en la antigua Unión Soviética una planta piloto en Krasnodar produce anualmente 22 toneladas de Boverin (conidias de B. bassima más una materia inerte, estandarizado a 6 x 109 wnidias/g) (Bustill0 y Posada, 1997).
En los Estados Unidos de Norteaménca, algunas empresas corno Nutrilite Products inc. en Buena Park California; Abbott Laboratones de North Chicago iü. y Sigma Chemical Co. han desarroliado algunas técnicas para producir biopreparados a base de los conidios de B. bassima como es el caso de las dos primeras empresas o formulado con la toxina denominada beauvencin, wmo es el caso de la última. Patnna Elizabeth Bañar Guevara 69 I
I

REVISIÓN DE LiTERA'iUR.4 - - Sin embargo, comercialmente no existe ningún producto de esta naturaleza que se
esté aplicando actualmente en ese país. Los antecedentes que se tienen al respecto, es que algunos investigadores han empleado algunas formulaciones del hongo proporcionados por esas empresas con fines experimentales (Méndez, 1990).
Se menciona al producto Naturalis-L como un micoinsecticida cuyo &@ente activo es 8. bassima, el cual ha sido probado para el control del picudo algodonero Anfhonornusgrandisy mosquita blanca. Cárdenas (1997) señala que en México, la empresa Apbionsa produce B. bassima, M. anisoplae y i? furnosomeus, usando la fermentación sólida en cereales, obteniendo los productos comerciales %-Sin, Meta-Sin y Pae-Sin.
De igual forma existen a nivel nacional algunos laboratonos rurales o universidades como el Instituto Tecnológico de Oaxaca que producen a nivel semicomercial un bioinsecticida a base de B. bassima. Uno de los laboratorios es el Laboratorio Repductor de Hongos Entomopatógenos de Zentla, que se encuentra en la Col. Manuel González Zentla en Huatusco, Veracruz, donde se está produciendo un micoinsecticida de alta calidad y efectividad en campo.
5.2.10.1.1. Formulación y conservación
El desarrollo de una adecuada formulación es esencial en el éxito de la utilización de micoinsecticidas. Propiedades biológicas y físicas de la formulación deben permanecer estables por al menos 1 año, pero preferiblemente por más de 18 meses, para que la comercialización ocurra. De principal importancia es la retención de la viabilidad y virulencia de las unidades infectivas durante el almacenamiento y la aplicación. La formulación es obligatoria para aumentar la aplicación de esporas y la eficacia. El tipo de formulación seleccionado depende de la biología y las propiedades físicas del patógeno asi como del lugar y hábitos del insecto blanco (Daoust et al., 1983).
Se encontró que los conidios pulverizados para su secado a 150OC y cubiertos con bentonita tienen de 50 a 70% de viabilidad, en tanto que las blastosporas que también son sensibles a esta técnica fueron liofilizadas y mezcladas conleche en polvo suplementada con glicerina, con este tratamiento, la viabilidad de las esporas aumentó a 89%. El almacenamiento de estas preparaciones se efectuó a una temperatura de 5 O C con la cual se preservó la viabilidad de las esporas por un lapso mayor a los 18 meses (Méndez, 1990). El micelio o esporas de 8. bassima una vez secas, pueden ser mezcladas con talco o algún otro material inerte como la perlita, kaolín, la bentonita o el almidón. Alcocer (1979) utilizrzó la harina de arroz estéril como vehículo para su preparación y la preservó a 4 O c .
En contraste para las formulaciones líquidas, las conidias almacenadas como 10% granulares o 20% en polvo retienen alta viabilidad ahdedor de un periodo de 12 meses a 4OC. Entre las formulaciones secas, los p d u c t o s granulares fueron más detrimentales para las conidias que crecieron en arruz almacenadas a 20OC. La viabilidad de las conidias mezcladas con aserrín y rastrojos de maiz, especialmente aquellos altamente secos, declinan más rápidamente mientras la viabilidad baja sólo lentamente después de que las conidias no formuladas son mezcladas con venniculita. La sobrevivencia de las conidias producidas en arruz en formulaciones granulares fue más alta a 4OC, pero baja en aquellas conidias no formuladas.
Paíricia Elizabeth Baños Guevara 70

Los aceites minerales han sido usados exitósamente en el almacenamiento prolongado de numerosas especies de hongos entomopatógenos. En un estudio en el cual 12 aislamientos de M. anisopliae, B. basiana, B. brongniarfii (=B. tenella), e Hirsutella spp., fueron almacenados a 4OC, todos los aislamientos sobrevivieron por 3 años a 4OC. Sin embargo en un experimento en particular, la viabilidad conidial declinó rápidamente en 12 aceites botánicos y 2 no botánicos en un período aproximado de 2 meses a 19 y 26°C. La sobrevivencia de las esporas en aceite de cacahuate y mineral fue d a r a todas las temperaturas aunque tenían viscosidades diferentes. Otros vehículos líquidos incluyen aceites con bases de petóleo, ácidos orgánicos y agua, que fueron m á s detrimentales para las conidias que los aceites botánicos (Daoust et al., 1983).
Velez y Montoya (1993) señalan que los aceites vegetales se usan para la aplicación de conidias de B. bassiana en el control de mosquito. Con el fin de proteger con el aceite a las conidias expuestas por un período de 90 minutos a la radiación de tipo ultravioleta. Asimismo las conidias se han mezclado con dos desecantes y pierden viabilidad más rápidamente que las mnidias almacenas separadas de los desecantes. También se han empleado agua destilada, talco, bentonita, talco m á s Carrier, aceite de soya, aceite de ondina y queroseno.
Posteriores investigaciones necesitan estabilizar la sobrevivencia conidd a altas temperaturas (> 20°C) desde la comercialización puede ser posible si la conidia fue almacenada por períodos prolongados (12- 18 meses) a temperatura ambiente (Daoust et al., 1983).
A nivel comercial existe otro producto cuyo ingrediente activo son conidias de B. bassiana cepa Laverlam llamado Bauveril WP, la concentración a la que es recomendado usarlo es 1 x 10" conidias por dosis como mínimo. Su formulación es a base de proteínas, sales y azúcares y es almacenado a temperaturas entre 2-g0C, mantiene su poder insecticida durante por lo menos 12 meses. Por ser un insecticida biológico puede ser afectado por temperaturas elevadas. A 27OC el porcentaje de viabilidad presenta una caída equivalente al 50% de la población inicial a los 15 días. A 37OC la caída es vertiginosa desde el día 8 de almacenamiento. Al aplicarlo en campo el producto no debe ser mezclado, con fungicidas, productos químicos o coadyuantes que alteren el rango de pH entre 6-7.5. En cualquier mezcla debe probarse previamente su compatibilidad y para lograr una mayor eficacia en la aplicación de Bauveril WP y reducir el riesgo de lavado por lluvias, se recomienda utilizar el portador Bioport, incluido como coadyuvante en el producto (Bauveril, 1997).
B. bassiana se puede aplicar en el campo usando máquinas de aspersión convencionales o equipos nebulizadores motorizados; así como en aspersoras de ultra bajo volumen. Recientemente, se han evaluado formulaciones en aceite, con las cuales se logra mayor dispersión de las conidias, mejor sobrevivencia y mayor efectividad en el control del insecto. Un factor importante en la eficiencia del hongo es la cobertura, por lo tanto el equipo que se utiliza debe estar bien calibrado para obtener una cobertura apropiada (Bustill0 y Posada, 1997).
5.2.10.1.2. Dosis recomendadas de algunos productos comerciales
Según los antecedentes, la ex-Unión Soviética y algunos países del bloque socialista como Checoslovaquia y Polonia son los lugares en donde se han logrado grandes avances en la investigación, producción y empleo masivo de B. bassiana en el control de las plagas agricoh, Boverin es un insecticida microbial formulado a base de conidios de este hongo, el cual es registrado, producido y empleado para el control de Lepfinofarsa decemlineata en Patricia Elizabeth Baños Gumma 71
P

dosis de 2 a 4 kg de Boverin/ha (formulación en polvo que contiene 1 x lo9 conidiasíg, equivalente a 1.2-2.4 x 1013 conidiasíha). El Boverin es un producto que se emplea también para controlar Loxmtege sticticalis a r h n de 3 kgíha, además es un producto que usualmente se aplica en combinación con dosis reducidas de Triclorfon o Malation (Méndez, 1990).
Para el control de insectos del suelo, como M. melolontha, se utiliza B. brongniartti en una dosis de 5 x 1 0 1 4 wnidias/ha; para otms insectos pequeños, de corta vida en el suelo, se requiere más inónilo (íO'6-lO'' conidiadha). Si se utilizan dosis bajas, la enfermedad se desarrolla lentamente y es posible que no se presenten epizootias (Bustiilo y Posada, 1997).
Del producto Bauveril WP, se recomienda usar la cantidad de 1 a 2 dosis por hectárea mezcladas en el volumen de agua recomendado. Aplicar con equipos convencionales eqauipados con boquillas cónicas de baja descarga. Calibrar el equipo antes de iniciar la aspersión. Agitar periódicamente el caldo durante la mezcla y aplicación. Utilizar el producto inmediatamente sea mezclado con el agua. Bauveril WP puede aplicarse con los metodos convencionales de aspersión. Las aplicaciones deben realizarse preferiblemente entre 6:OO y 1O:OO am y después de las 4:OO pm o a cualquier hora en días nublados para evitar al máximo los rayos ultravioleta de sol que afectan las conidias. El producto puede ser usado hasta el momento de la cosecha (Bauveril, 1997).
El Bauvenl WP no tiene ningún efecto nocivo sobre el operario y el medio ambiente. No deteriora la fauna benéfica y permite establecer programas de manejo integrado. Tiene efectos más prolongados de control y no Ocurren efectos tóxicos por acumulación en aplicaciones sucesivas. Puede aplicarse en cualquier época de desarrollo del cultivo. No se tiene evidencias de resistencias directas ni cruzadas. El producto puede ser ligeramente irritante para el sistema respiratorio o por contacto directo con los ojos. En caso de inhalación, ponerse al aire fresco. En caso de contacto con los ojos, lavar inmediatamente con agua durante 10 minutos (Bauvenl, 1997).
4= Paíricia EIizabeth Bañas Guevara 72
1

MATERIALES Y MÉTOWS
V. Materiales y métodos
5.1. Lugar y tiempo de la investigación
Este trabajo de tesis se desarrolló en el Laboratorio de investigación del Departamento de ingeniería Agroindustrial de la Universidad Autónoma Chapingo., en colaboración con el Laboratorio Reproductor de Hongos Fhtomopatógenos de Zentla (LARHEZ) ubicado en la colonia Manuel González Zentla, Huatusco, Ver. que pertenec.e el Comité Regional de Sanidad Vegetal de Huatusco, Veracruz. La duración del proyecto fue de 1 año de experimentación y 6 meses de trabajo de gabinete.
5.2. Fuentes de financiamiento
Esta tesis fue financiada por el Programa Universitario de investigación en Alimentos (F’üiA) de la Universidad Autónoma Chapingo, a través de la línea de Biotecnología de Alimentos y de manera externa por el Comité Regional de Sanidad Vegetal de Huatusco, Veracruz.
5.3. Origen y mantenimiento de la cepa del hongo
El microorganismo utilizado fue una cepa silvestre de B. bassiana aislada y purificada directamente de brocas (H. hampel) micosadas de plantíos de café de la región de Huatusco, Veracruz, posteriormente el Laboratorio LARHEZ antes mencionado proporcionó la cepa de trabajo para el Laboratorio de investigación donde se obtuvieron cultivos de trabajo, empleando un medio de cultivo sintético de Agar Dextrosa Saboraud y frascos de dilución inclinados; realizando la siembra de conidias por la técnica de estría. Los cultivos se incubaron durante 15 días a 28OC y posteriormente se refrigeraron para su conservación y subsecuente utilización.
5.4. Selección de sustratos y análisis químico proximal
Los sustratos seleccionados están clasificados dentro de los sólidos: el grano de arruz pulido y entero y el arruz residual, producto de la primera cosecha de esporas de la producción masiva del LARHEZ
A estos dos sustratos se les practicó el análisis bromatológico general que incluye la determinación de humedad, cenizas, extracto libre de nitrógeno, proteína, extracto etéreo y por diferencia la fibra cruda (AOAC, 1984). También se cuantificó el contenido de almidón por el método de Fehling (AOAC, 1990).
P - P
Patricia Elizabeth Bañar Guevara 73
I

MATERIALES Y MÉTODOC - 5.5. Pruebas preliminares de preparación del sustrato
5.5.1. Perfil de esporulación de B. bmsima Para conocer el comportamiento del crecimiento y esponilación del hongo a estudiar
fue necesario realizar un perfil de esporuiación bajo el sistema de cultivo tradicional empleado en el LARHEZ. La metodología fue la siguiente: en tubos de ensaye con tapón de baquelita, se pesaron 15 g de a m z pulido entero, se le añadió 25 ml de agua destilada y se remojaron por 30 minutos, después se invirtieron los tubos para escurrir el exceso de agua por un lapso de tiempo igual, posteriormente se esterilizaron a 121°C por 15 minutos, se enfriaron a temperatura ambiente y se inocularon con una suspensión de esporas producto de un cultivo de trabajo, la cantidad fue 1 ml con una concentración de 6 x lo6 esporas. Los tubos se incubaron a ZS°C durante 15 días, diariamente se muestrearon 2 tubos cada 24 horas y se cuantificó el número de esporas por cuenta directa en la cámara de Neubauer.
5.5.2. Ajuste de pH
Como una prueba preliminar hubo que realizar un ajuste de pH al agua de remojo del arruz entero y residual, en esta fase se empleó el método de producción tradicional, el cual consiste en pesar 15 g de a m z por unidad experimental y con las soluciones ajustadas a los siguientes valores de pH: 5.0, 5.5, 6.0, 6.5 y 7.0 (se empleó una solución O.1N de ácido fosfórico) se remojaron los sustratos por 30 minutos y se escurrieron por un lapso igual de tiempo para después esterilizar los tubos de ensayo a 12 l0C por 15 minutos, dejarlos enfriar a temperatura ambiente e inocularlos con un volumen y cantidad de inóculo constante; se dejaron incubar por 15 días a una temperatura de 2PC . El pH Óptimo se seleccionó con base en el nivel m á s alto de esponilación del hongo.
5.5.3. Curvas de ajuste de contenido de agua Estas curvas se elaboraron para saber el comportamiento de la absorción de
humedad de los sustratos empleados; el procedimiento empleado fue el siguiente: a una cantidad estándar (15 9> de sustrato se le agregó de 3 a 12 ml de agua destilada, posteriormente las bolsas de polipapel se seliaron y se esterilizaron a 12 í0C por 20 minutos, se dejaron enfriar a temperatura ambiente y el contenido de humedad se calculó por diferencia de pesos y por secado a la estufa.
5.5.4. Desarrollo de un método colorimétrico para determinación de almidón soluble
Para facilitar la cuantificación del almidón soluble residual se innovó una propuesta metodológica realizada por la necesidad de facilitar el análisis de las muestras en cuanto a costo y tiempo; este método se basa en la cuantificación del almidón soluble presente en la muestra, dado que el grano de arroz tiene un alto porcentaje de almidón soluble se adecuó por tanto el método de tinción cualitativo con lug01 para identificación de almidón a un método colorimétrico para determinación de almidón soluble descrito en los siguientes renglones.
P
Patricia Elizabeth Bañar Guevara 14

MATERIALES Y &TODOS
5.5.4.1. Preparación de la muestra
Después de cosechar las esporas, efectuar la cuenta o hacer la prueba de viabihdad en su caso; todas las unidades experimentales se almacenaron en congelación, para realizar todos estos análisis al final del perfil de esporulación.
Las muestras se descongelaron, se maceraron y se pesó determinada cantidad (Zg); después se sometieron a ebullición y agitación durante 5 minutos, al cabo de los cuales se procedió a filtrar la solución en tela de lino y enjuagar el residuo con agua caliente hasta iiegar a un aforo de 100 ml.
Posteriormente, las soluciones filtradas se sometieron a un nuevo calentamiento para facilitar la subsecuente filtración a vacío con papel Whatman No. 42. La filtración a vacío fue el mejor método de extracción del almidón soluble en comparación wn la centrifugación a 2750 rpm que también se probó.
Ya filtradas las soluciones se hicieron las respectivas diluciones de acuerdo a la concentración del almidón, al inicio del perfil de esporulación hay mayor cantidad de almidón soluble residual que al final.
5.5.4.2. Reacción colorimetrica y lectura de datos
Efectuadas las diluciones adecuadas, se llevó a cabo la reacción colotimétrica, la cual consiste en agregar en un matraz volumétriw de 100 ml: 30 ml agua destilada, 1 ml de muestra, 1 ml de Cloruro de Calcio (0.5 M), 0.2 ml de lugol y agua destilada hasta aforo.
Se hornogeneizó la solución y se tomaron lecturas de densidad óptica (absorbancia) en el espectrofotómetro a una longitud de onda de 600 nm. Calibrando previamente con agua destilada como blanco.
5.5.4.3. Análisis de áatos
Para comprobar la repducibilidad del método desarruiiado se realizaron varias curvas tipo empleando almidón soluble (reactivo analítico) y como muestra harina de arroz para correlacionar los datos. Las curvas tipo fueron ajustadas por el método de regresión lineal.
Las curvas tipo empleadas para el análisis de datos fueron las harinas correspondientes a cada uno de los sustratos: harina de arroz entero y harina de arroz residual; ajustadas también por regresión lineal.
Los resultados de absorbancia de las muestras se interpolaron con la curva tipo y se pudo apreciar así la concentración de almidón soluble residual por gramo de arroz entero.
- Patricia Elizabeth Bañm Gumma 75

MATEMALES Y &TODOS
COhTENrDO DE AGUA
(%) 45 55 65 75
5.5.5. Afinación del método para determinación de viabilidad Para realizar la determinación de viabilidad se probaron d o s tipos de cultivo
empleando dos medios sintéticos:
1) Cultivo en superficie, en placas de Petri conteniendo medio Saboraud Dextrosa Agar (SDA) más 0.1% de rosa de bengala para inhibir el crecimiento de micelio y poder así cuantificar más fácilmente las colonias del hongo. Las placas se inocularon con apmximadamente 50 esporas, se incubaron por al menos 4 días a 28OC.
2 ) Cultivo líquido sumergido, en tubos de ensaye con caldo nutritivo para hongos se inocularon también 50 esporas y se mantuvieron en las m i s m a s condiciones.
TEMPERATURA ("C)
20 25 30 35 I (01,24,48) U (08,29,40) iíí (10,18,46) iV (14,17,381 V (13,20,42) Vi (02,32,36) W (09,23,41) Wii (04,31,37) M (05,28,47) X (15,27,35) Xi (07,22,44) W (12,19,45) Wr (03,30,39) XIV (11,26,43) XV (16,25,33) XW (06,21,34)
5.6. Primera fase del experimento
5.6.2. Preparación de las unidades experimentales Las unidades experimentales fueron bolsas de polipapel de alta densidad a las cuales
se les añadieron 15 g de cada uno de los sustratos seleccionados, se ajustó el contenido de agua como ya se explicó anteriormente, en la parte terminal de la bolsa se hicieron dos dobleces hasta alcanzar las medidas de 12 x 14 cm, se engraparon y sellaron las bolsas con cinta adhesiva, se esteriliuiron a 12 l°C por 20 minutos, al saiir se desinfectaron las grapas y se taparon perfectamente para evitar cualquier pérdida de humedad, posteriormente se realizó una suspensión de inóculo empleando un cultivo de trabajo y el volúmen se ajustó a 1 ml con una concentración de 12 x lo6 esporas; la inoculación de las unidades experimentales se reaiizb en un cuarto aséptico y en una cámara de flujo laminar, se emplearon jeringas de 1 d, al terminar la inoculación se hornogeneizaron manualmente las bolsas y se incubaron a las temperaturas señaladas en el diseño experimental anteriormente descrito.
Patricia Elizabeth Baiíai Gumma 76

M A T E W E S Y MÉTOWS
Las estufas empleadas no tuvieron caractensticas estándares, ya que para la temperatura de 20 y 25OC se emplearon dos cámaras de g e k a c i ó n que regulan su temperatura en base a resistencias eléctricas que calientan determinada cantidad de agua y a través del vapor de agua se logra la homogeneización de la temperatura deseada; para las temperaturas de 30 y 35OC se utilizaron estufas de incubación que funcionan con base en resistencias eléctricas que calientan el aire ambiental. Por ello se tuvo mucha atención cn el correcto sellado de las bolsas de polipapel.
A los 15 días de incubación, todas las unidades experimentales se congelaron para detener toda actividad metabólica.
5.6.3. Cosecha
La cosecha se realizó manualmente vaciando el contenido de cada unidad experimental en frascos desinfectados, la bolsa se enjuagó con 30 ml del dispersante Tween 80 a una concentración de 0.1%, con una varilla de vidrio se agitaron las muestras y se tomó 1 ml de la suspensión de esporas para realizar la cuenta; si era necesario se realizaron diluciones utilizando agua destilada.
5.6.4. Medición de las variables de respuesta
5.6.4.1. Conteo de esporas
Se evaluó la esponilación del hongo por conteo directo de esporas al microscopio, empleando la cámara de Neubauer o hemocitómetro.
La concentración de esporas se evaluó utilizando la siguiente ecuación:
(esporas/mililitro) = (X) (25) (10,000) (F.D.)
donde: X F.D. = Factor de dilución. 10,000 = Las dimensiones de la cuadrícula graduada de la cámara de recuento son de 1 mm
x 1 nun por lado y como el cubreobjetos está a O. 1 nun sobre la cámara, el volumen de ésta es de 0.1 nun3. Y debido a que las cantidades de &Mas se interpretan en números por centímetros cúbicos (un centímetro cúbico equivale a 1 mililitro), el volumen del cuadro grande es 0.0001 c m s ; se llega a este volumen de la siguiente forma: 10 x O. 1 nuns= 1 m m 3 ; 1,000 x 1 mm3= Icm3; 10,000 x O. 1 nuns = 1 cm3.
= Promedio del número de esporas contadas en 5 cuadros grandes de la cuadrícula.
25 = Número de cuadros pequeños; cada cuadro pequeño contiene 16 microcuadros.
Patricia Elizabeth Baños Gumma 77

MATERiALES Y MÉTOWS
5.7. Segunda fase del experimento
5.7.1. Selección de los mejores tratamientos
De la fase anterior se realizó un análisis estadístico empleando el Análisis de Varianza (ANAVA) para seleccionar el mejor tratamiento de cada uno de los sustratos probados, con base en el máximo iuvel de esporulación, como se explica postenomente en el análisis estadístico.
5.7.2. Medición de las variables de respuesta
5.7.2. I. Conteo de esporas Como ya se ha mencionado anteriormente las esporas se cuantificaron en un
hemocitómetro o cámara de Neubnuer.
5.7.2.2. Determinación del sustrato residual
Se realizó como ya se mencionó en las pruebas preliminares.
5.7.2.3. Pruebas de viabilidad
Para esta prueba se emplearon cultivos en superficie, empleando el medio sintktico Saboraud Dextrosa Agar (SDA), enriquecido con 0.6% de harina de arroz y 1% de rosa de bengala a una concentración de I : 10,000; se inoculó 0.1 ml de la dilución correspondiente de la suspensión de esporas, las placas se incubaron a cada una de las temperatura óptima del tratamiento seleccionado.
Se optó por monitorear sólo 3 puntos a la par del desarrollo del perfil de crecimiento y esponilación de 8. bmiana debido a la insuficiencia de material y tiempo.
5.7.2.4. Cambio de pH El cambio de pH se detectó empleando un potenciómetro, previamente calibrado con
una solución reguladora a pH 7.
- Paíricia Elizabeth Bañar Guevara 78

MATERIALES Y h4ÉTODOS - __p_I -
5.8. Análisis estadístico
5.8.1. Primera fase
una vez que se tuvieron los datos de esponilación de esta fase se procedió a elegir el mejor sustrato con base en el máximo nivel de esporulación, la selección se hizo mediante un análisis de varia- y una prueba de Tukey; los datos se analizaron con el auxilio del software denominado comúnmente SAS (sistema de Análisis Estadístico, SAS Institute INC, 1988). También se compararon las varia- de los residuales del modelo (Anexo 11 .1 ) .
Ya seleccionados los mejores tratamientos (Gráficas 8 y 9) se realizó una prueba para comprobar si el mejor tratamiento del experimento correspondía al óptimo tratamiento en realidad (Anexo 11.2).
5.8.2. Segunda fase
Una vez seleccionados los mejores tratamientos se realizaron los perfiles de esporulación y los análisis de las variables de respuesta correspondientes. Una vez que se tuvieron los datos se procedió a realizar el ajuste de las cullras de esporulación (Gráficas 10 y 1 1 ) y almidón soluble residual (Gráficas I2 y 13).
Para el ajuste de l a s curvas de esporulación, en arroz entero y residual se realizó un análisis de regresión no lineal empleando la función logística; aunque se emplearon diferentes métodos de ajuste dado que los resultados experimentales fueron diferentes, para arroz entero se usó el método Maquardt y en arroz residual se ajustó mejor el método Dud (Anexo 11.3). Se graficaron los datos ajustados proporcionados por este modelo así como los residuales.
En cuanto al ajuste de los resultados de la determinación de arroz residual, se empleó
Y = (X) (X”2) (XZ)
el siguiente modelo matemático para los dos sustratos (Anexo 11.4).
donde:
Y = Variable de respuesta, concentración de almidón.
X = Variable días, corresponde a cada uno de los días del perfil de esporulación.
P Paíricia Elizabeth Baños Guevara 79
I
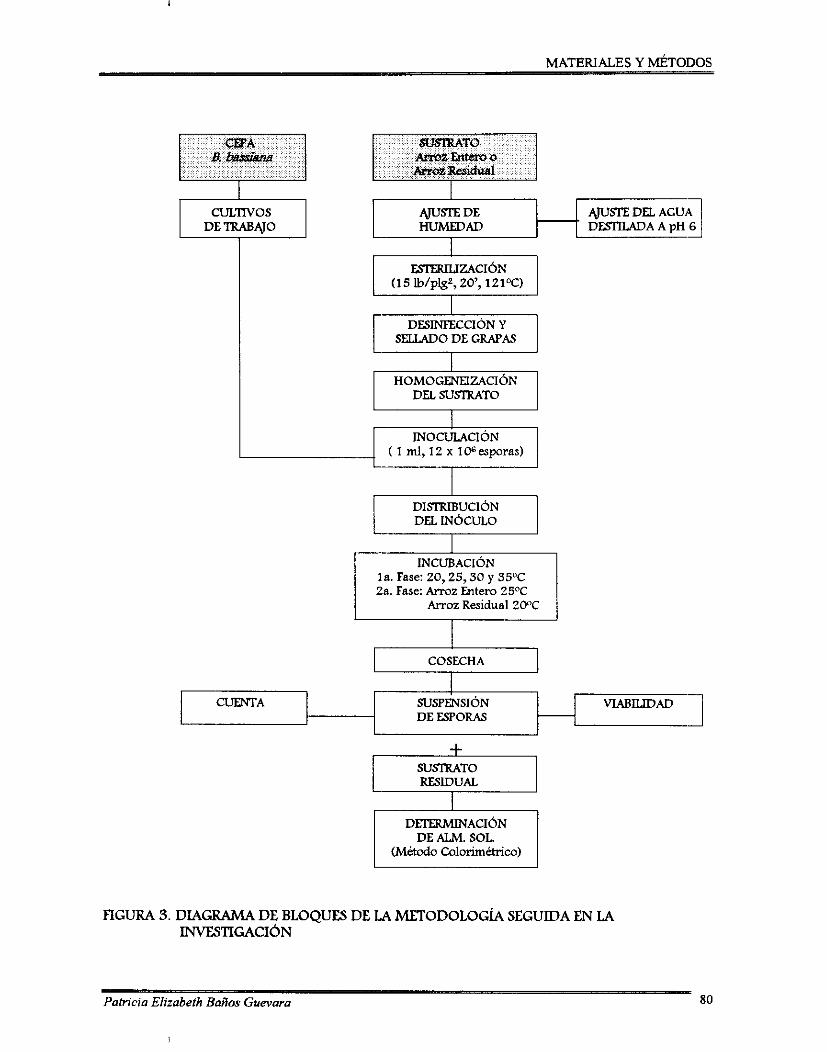
MATERIALES Y hCÉTODOS II - -
DE TRABqlO ,gum DEL A G U ~
HUMEDAD DESiILADA A pH 6 I
I I ,
ESITNUZACIÓN
SELLADO DE GRAPAS
DEL SUSTRATO
! INOCULACI~N
( 1 ml, 12 x ipesporas)
I M C U B A C I ~ N i l a . Fare: 20.25.30 v 35.T
2a. Fase: Arroz Entero 25OC Arroz Residual Z O T
---- I COSECHA I
I VIABILWAD 1 I J DEFSPORAS
SUSTRATO RESIDUAL
I
DFIZRMINACI~N DE ALM. SOL
(Método Colorimétrico)
FIGURA 3. DIAGRAMA DE BLOQUES DE LA h4ETODOLOGÍA SEGUIDA EN LA INVESTIGACI6N
- - P P
Patricia Elizabeth Bañar Guevara 80
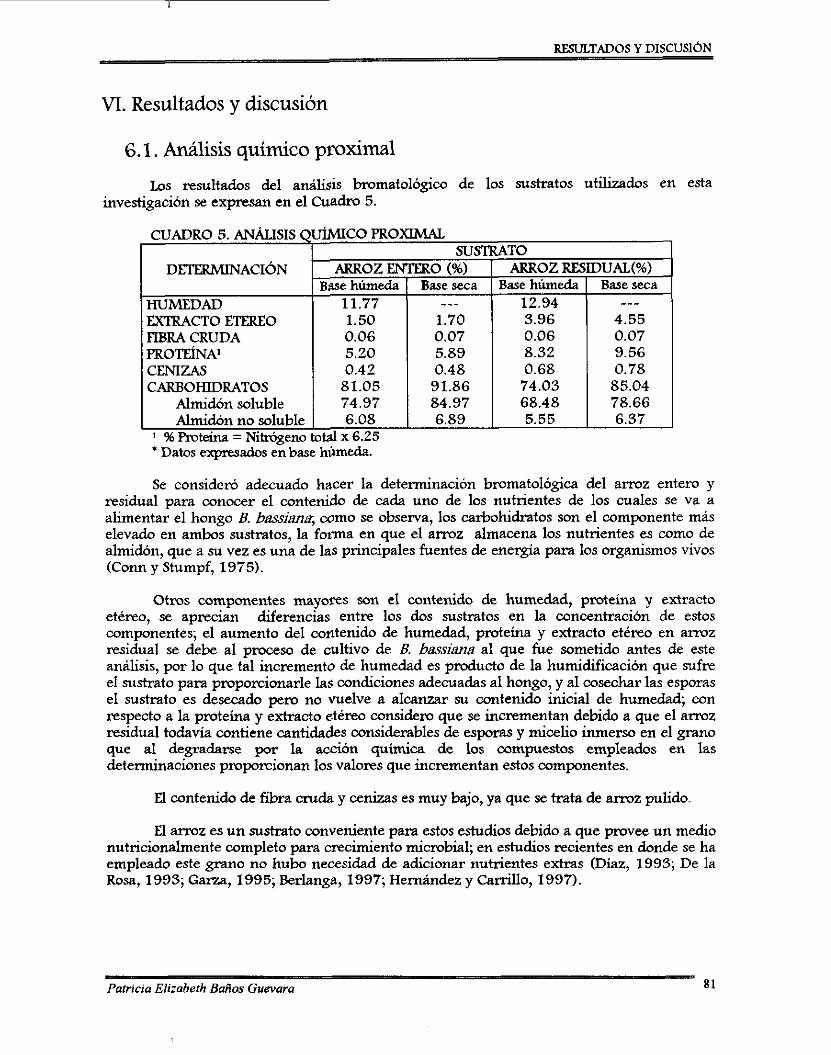
RESULTADOS Y DISCUSIÓN
Vi. Resultados y discusión
6.1. Análisis químico proximal Los resultados del análisis bmmatológico de los sustratos uti l izados en esta
investigación se expresan en el Cuadro 5.
CUADRO 5. ANÁLISIS QUÍMICO PROXIMAI, I I SUSI
XO (%) I DETERMINACIÓN
EXTRACTO ETEREO FIBRA CRUDA PROTE~NAI
CARBOHiDRATOS
Base húmeda 11.77
0.42 81.05
I Almidónsoluble I 74.97 Almidón no soluble I 6.08
I % F’ruteina = Nihóneno total x 6.25
Base seca
1.70 0.07 5.89 0.48
91.86 84.97 6.89
---
IT0 I ARROZ RE:
Base húmeda 12.94 3.96 0.06 8.32 0.68 74.03 68.48 5.55
>UAL(%)l Base seca
85.04 78.66
* Datos expresados &base húmeda.
Se consideró adecuado hacer la determinación brornatológica del arroz entero y residual para conocer el contenido de cada uno de los nutrientes de los cuales se va a alimentar el hongo B. bassiana, como se observa, los carbohidratos son el componente mas elevado en ambos sustratos, la forma en que el arroz almacena los nutrientes es como de almidón, que a su vez es una de las principales fuentes de energía para los organismos vivos (Conn y Stumpf, 1975).
Otros componentes mayores son el contenido de humedad, proteína y extracto etéreo, se aprecian diferencias entre los dos sustratos en la concentración de estos componentes; el aumento del coritenido de humedad, proteína y extracto etéreo en arroz residual se debe al proceso de cultivo de 6. bassima al que fue sometido antes de este análisis, por lo que tal incremento de humedad es producto de la humidificación que sufre el sustrato para proporcionarle lac condiciones adecuadas al hongo, y al cosechar las esporas el sustrato es desecado pero no vuelve a alcanzar su contenido inicial de humedad; con respecto a la proteína y extracto etéreo considero que se incrementan debido a que el arroz residual todavía contiene cantidades considerables de esporas y micelio inmerso en el grano que ai degraáarse por la acción química de los compuestos empleados en las determinaciones proporcionan los valores que incrementan estos componentes.
El contenido de fibra cruda y cenizas es muy bajo, ya que se trata de arroz pulido.
El arroz es un sustrato conveniente para estos estudios debido a que provee un medio nutricionalmente completo para crecimiento microbial; en estudios recientes en donde se ha empleado este grano no hubo necesidad de adicionar nutrientes extras (Diaz, 1993; De la Rosa, 1993; Gana, 1995; Berlanga, 1997; Hernández y C a d o , 1997).
P -- 81 -
Patricia Elizabeth Baños Guevara
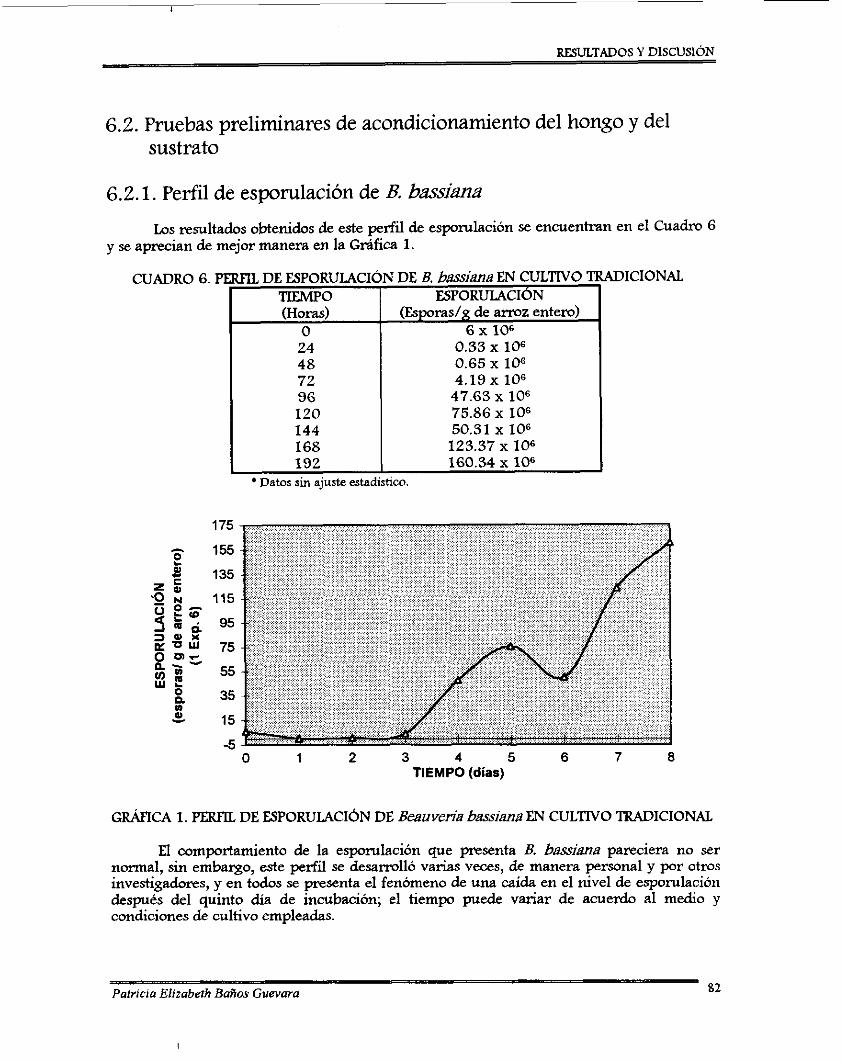
RESULTADOS Y DISCUSI~N -
TIEMPO (Horas)
O
6.2. Pruebas preliminares de acondicionamiento del hongo y del sustrato
ESPORULACI~N (Esporasíg de arroz entero)
6 x 10G
6.2.1. Perfil de esporulación de B. bassima Los resultados obtenidos de este perfil de esponilación se encuentran en el Cuadro 6
y se aprecian de mejor manera en la Gráfica 1.
24 48 72 96 120 144 168 192
0.33 x 106 0.65 x 10G 4.19 x 10G
47.63 x 10G 75.86 x IOG 50.31 x 10G 123.37 x 106 160.34 x 10G
175 - 155
8 135
115
O
2 s 9 N u g ú j 4 m . e 95
Y!!
3 a l Y K u W 75
2;s 55 O
01 al
n 35 15 - -5
O 1 2 3 4 5 6 7 a TIEMPO (días)
GRÁFICA 1. PFXFIL DE ESPORULACIÓN DE Beauveria bassima EN CULTIVO TRADICIONAL
El comportamiento de la esponilación que presenta B. bassima pareciera no ser normal, s i n embargo, este perfil se desarrolló vanas veces, de manera personal y por otms investigadores, y en todos se presenta el fenómeno de una caída en el nivel de esponilación después del quinto día de incubación; el tiempo puede variar de acuerdo al medio y condiciones de cultivo empleadas.
- L 82 Patricia Elizabeih Baños Gumara
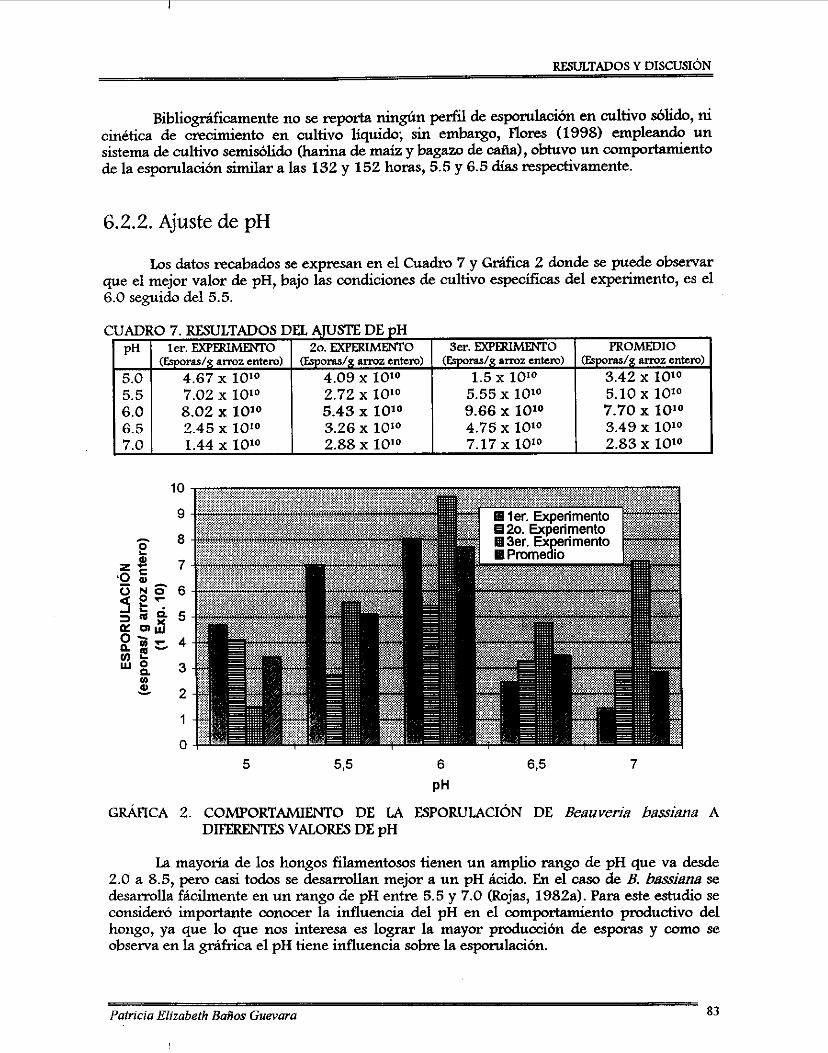
pH ler. EXP€XWEBTO 20. - 0
5.0 4.67 x 1O1O 4.09 x lozo 5.5 7.02 x lolo 2.72 x 1O1O 6.0 8.02 x 10'0 5.43 x 10'0 6.5 2.45 x loEo 3.26 x lolo 7.0 1.44 x 1O1O 2.88 x 1010
(Esporas/g arroz entero) (Erpors/g arroz entero)
10
9
Ser. -0 PROMEDIO (Esporadg arroz entero) (Erpo rs/g arroz entero)
1.5 x lolo 5.55 x 10'0 9.66 x 1Olo 4.75 x 10'0 7.17 x 1O1O
3.42 x 1O1O 5.10 x 10'0 7.70 x 1O1O 3.49 x 1 0 1 0
2.83 x lolo
1
O 5 5,5 6 6.5 7
PH
GRÁFICA 2. COMPORTAMIENTO DE LA FSPORULACIÓN DE Beauveria bassiana A DIFERENTES VALORES DE pH
La mayoría de los hongos filamentosos tienen un amplio rango de pH que va desde 2.0 a 8.5, pero casi todos se desarrollan mejor a un pH ácido. En el caso de B. bassi's se desarrolla fácilmente en un rango de pH entre 5.5 y 7.0 (Rojas, 1982a). Para este estudio se consideró importante conocer la influencia del pH en el comportamiento productivo del hongo, ya que lo que nos interesa es lograr la mayor producción de esporas y como se observa en la gráfrica el pH tiene influencia sobre la esporulación.
Paíricia Elizabeth Baños Guevara 83
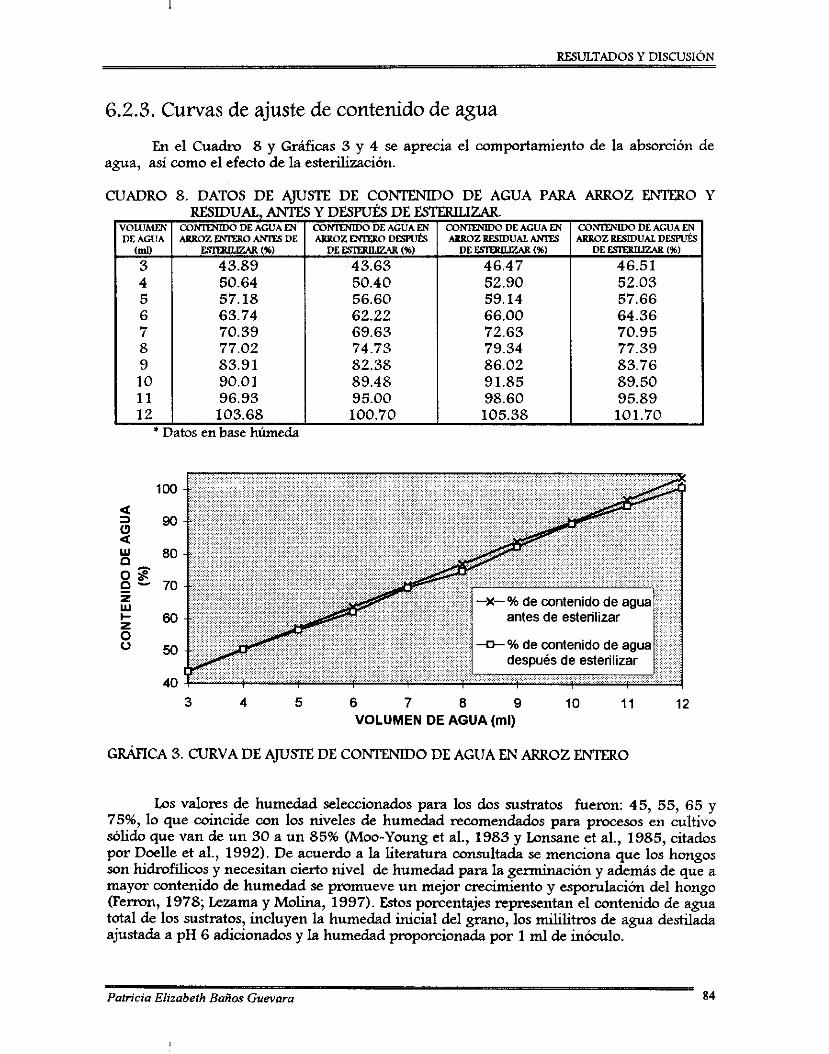
1
RESULTADOS Y DISCUSIÓN
6.2.3. Curvas de ajuste de contenido de agua En el Cuadro 8 y Gráficas 3 y 4 se aprecia el comportamiento de la absorción de
agua, así como el efecto de la esterilización.
CUADRO 8. DATOS DE AWSrr: DE CONTENIDO DE AGUA PARA ARROZ ENTERO Y
4 5 6 7 8 9 10 1 1
50.64 57.18 63.74 70.39 77.02 83.91 90.01 96.93
12 103.68 - * Datos en base húmeda
50.40 52.90 56.60 59.14 62.22 66.00 69.63 72.63 74.73 79.34 82.38 86.02 89.48 91.85 95.00 98.60 100.70 105.38
52.03 57.66 64.36 70.95 77.39 83.76 89.50 95.89 101.70
100
2 9 0 2
80
B E - 70 z w + 6 0 antes de esterilizar z O
0 -
+Oh de contenido de agua después de esterilizar
0 5 0
40
3 4 5 6 7 8 9 10 11 12 VOLUMEN DE AGUA (mi)
GRÁFICA 3. CURVA DE AJUSTE DE CONTENIDO DE AGUA EN ARROZ ENTERO
Los valores de humedad seleccionados para los dos sustratos fueron: 45, 55, 65 y 75%, lo que coincide con los niveles de humedad recomendados para procesos en cultivo sólido que van de un 30 a un 85% (Moo-Young et al., 1983 y Lonsane et al., 1985, citados por Doelle et al., 1992). De acuerdo a la literatura consultada se menciona que los hongos son hidmfílicos y necesitan cierto nivel de humedad para la genninación y además de que a mayor contenido de humedad se promueve un mejor crecimiento y esporulación del hongo (Femn, 1978; Lemma y Molina, 1997). Estos porcentajes representan el contenido de agua total de los sustratos, incluyen la humedad inicial del grano, los miiilitros de agua destilada ajustada a pH 6 adicionados y la humedad proporcionada por 1 ml de inóculo.
Patricia Elizabefh Baños Gumma 84
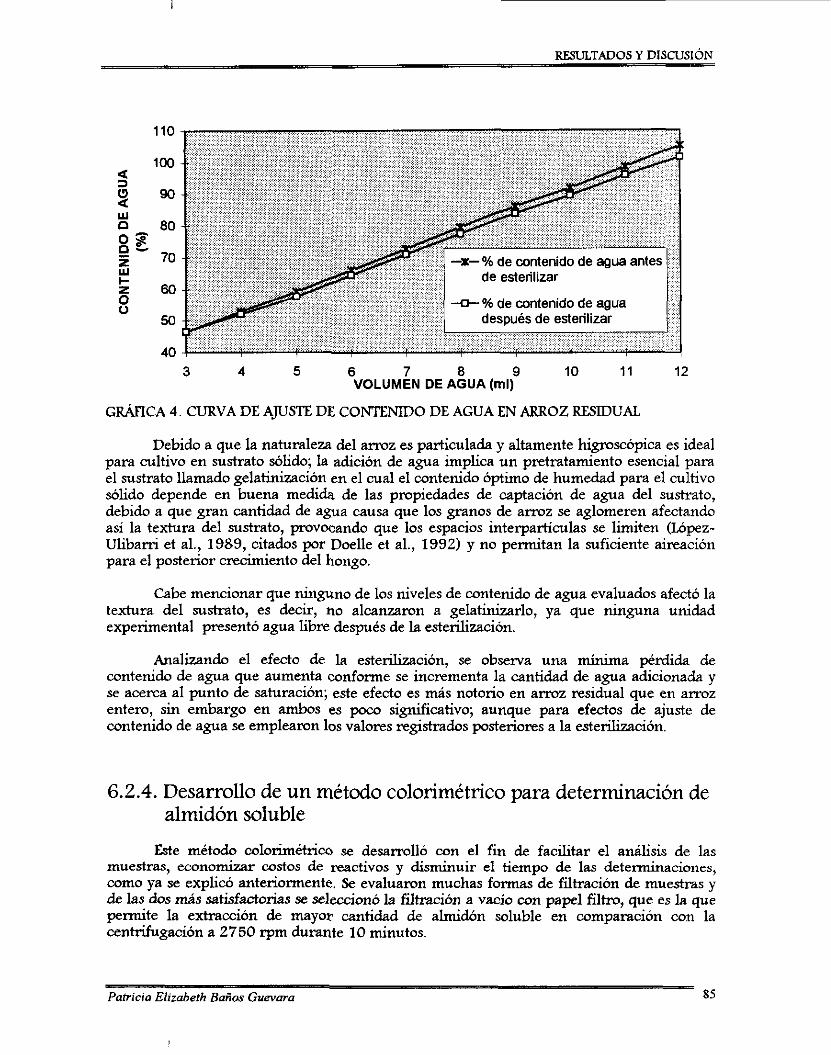
T
RESULTADOS Y DISCUSION
110
40 3 4 5 6 7 6 9 10 11 12
VOLUMEN DE AGUA (mi)
GRÁFICA 4. CURVA DE AJUSTE DE CONTEND0 DE AGUA EN ARROZ RESIDUAL
Debido a que la naturaleza del arroz es particulada y altamente higroscópica es ideal para cultivo en sustrato sólido, la adición de agua implica un pretratamiento esencial para el sustrato llamado gelatinización en el cual el contenido óptimo de humedad para el cultivo sólido depende en buena medida de las propiedades de captación de agua del sustrato, debido a que gran cantidad de agua causa que los granos de a m z se aglomeren afectando así la textura del sustrato, provocando que los espacios interpartículas se limiten (ibpez- Ulibarri et al., 1989, citados por Doelle et al., 1992) y no permitan la suficiente aireación para el posterior crecimiento del hongo.
Cabe mencionar que ninguno de los niveles de contenido de agua evaluados afectó la textura del sustrato, es decir, no alcanzaron a gelatinizarlo, ya que ninguna unidad experimental presentó agua libre después de la esterilización.
h a h a n d o el efecto de la esterilización, se observa una mínima pérdida de contenido de agua que aumenta conforme se incrementa la cantidad de agua adicionada y se acerca al punto de saturación; este efecto es más notorio en arroz residual que en arroz entero, sin embargo en ambos es poco significativo; aunque para efectos de ajuste de contenido de agua se emplearon los valores registrados posteriores a la esterilización.
6.2.4. Desarrollo de un métocio colorimétrico para determinación de almidón soluble
Este método color imé~co se desarrolló con el fin de facilitar el análisis de las muestras, economizar costos de reactivos y disminuir el tiempo de las determinaciones, como ya se explicó anteriormente. k evaluaron muchas formas de filtración de muestras y de las dos más satisfactohas se seleccionó la filtración a vacío con papel filtro, que es la que permite la extracción de mayor cantidad de almidón soluble en comparación con la centrifugación a 2750 rpm durante 10 minutos.
Patricia Elizabeth Bañas Guevara 85
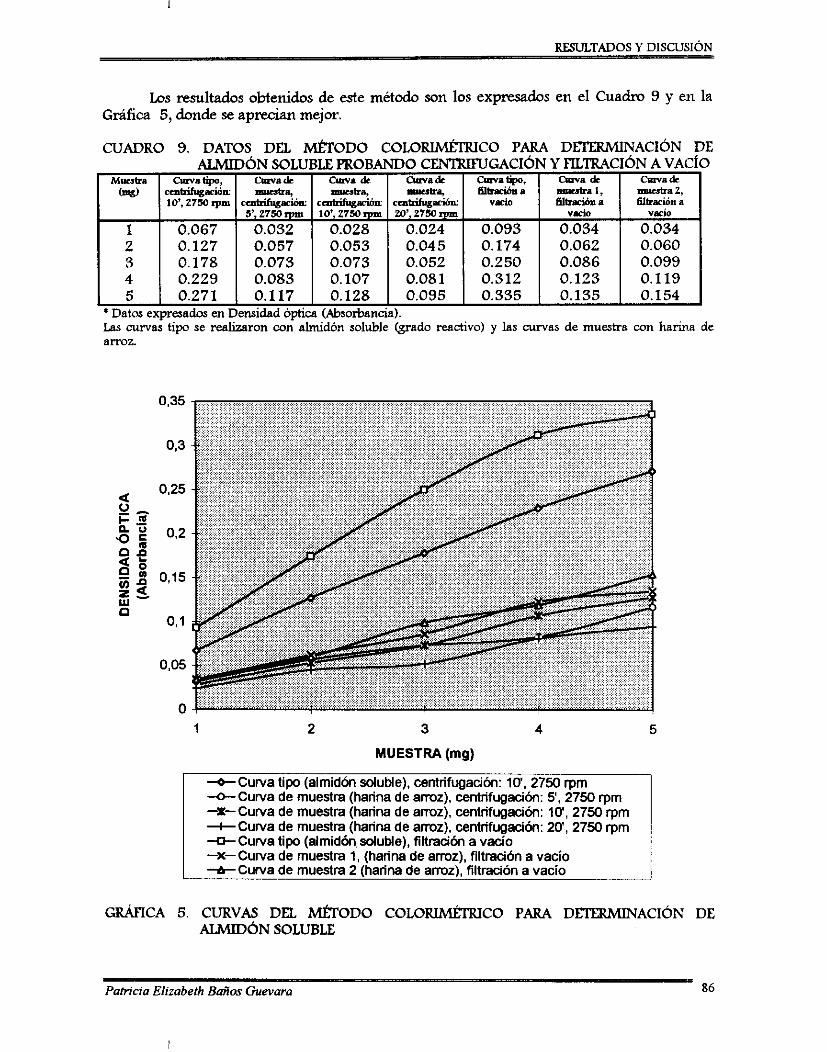
RESULTADOS Y DISCUSION -
Muwbn Cuma*, Cxwadc cwi de CnNPdC cwi*, c w a & 6i-n muubal,
1 0.067 0.032 0.028 0.024 0.093 0.034
6 i b d P (4 cmhih&& muuha,, y k 4 -5 10',2750rpm crnhihysoon: c m k w i q h emkwiqscik VA
S', 2750 ipm io', 2750- u)', 2750 ipm V r i O
Los resultados obtenidos de este método son los expresados en el Cuadro 9 y en la Gráfica 5, donde se. aprecian mejor.
cwndc muutra 2, 6 i k d B
ve io
0.034
Muwbn Cuma*, Cxwadc cwi de CnNPdC cwi*, c w a & 6i-n muubal,
1 0.067 0.032 0.028 0.024 0.093 0.034
6 i b d P (4 cmhih&& muuha,, -*4 -5 10',2750rpm crnhihysoon: c m h i h & h emhih&scik VA
S', 2750 ipm io', 2750- u)', 2750 ipm V r i O
CUADRO 9. DATOS DEI. &ODO COLORIMÉTRICO PARA DITERMINACI~N DE
cwndc muutra 2, 6 i k d B
ve io
0.034 2 0.127 0.057 0.053 0.045 0.174 0.062 3 0.178 0.073 0.073 0.052 0.250 0.086 4 0.229 0.083 O. 107 0.08 1 0.312 0.123 5 0.271 0.117 O. 128 0.095 0.335 0.135
0.060 0.099 0.119 0.154
0.25
0.2
0.15
o, 1
0,05
O
2 0.127 0.057 0.053 0.045 0.174 0.062 3 0.178 0.073 0.073 0.052 0.250 0.086 4 0.229 0.083 O. 107 0.08 1 0.312 0.123 5 0.271 0.117 O. 128 0.095 0.335 0.135
1 2 3 4 5
MUESTRA (mg)
+Curva tipo (almidón soluble). centrifugación: 10'. 2750 rprn &Curva de muestra (harina de arroz), centrifugación: 5,2750 rpm -X-Curva de muestra (harina de arroz). centrifugación: lU, 2750 rpm +Curva de muestra (harina de arroz). centrifugación: 20', 2750 rpm +Curva tipo (aimidón cduble), filtración a vacío -x-Curva de muestra 1, (harina de arroz), filtración a vacío +Curva de muestra 2 (harina de arroz). filtración a vacío
0.060 0.099 0.119 0.154
GRÁFICA 5. CURVAS DEL &ODO COLORIMÉTRICO PARA DETERMINACI~N DE ALMIDÓN SOLUBLE
P
Patricio Elizabeth Baños Guevaro 86
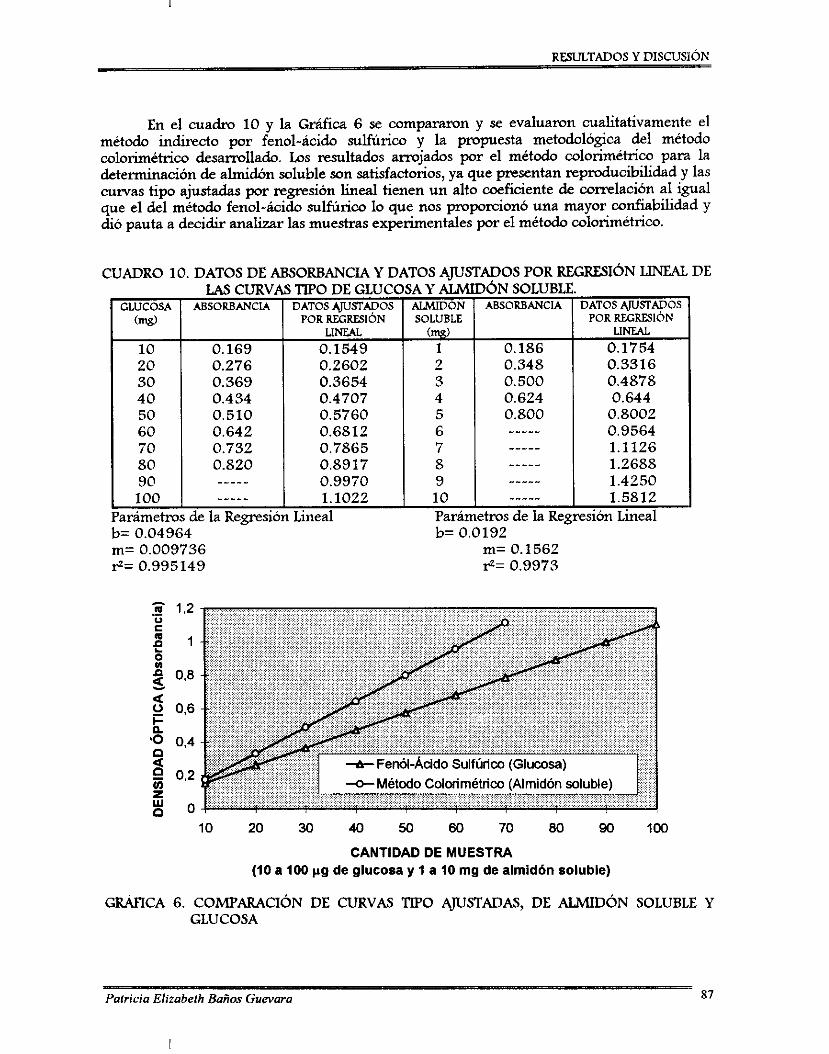
REWLTADOS Y DISCUSÓN
GLUCOSA ABSORBANCU DATOS wSl'A!JOS ALMIDON ABSORBANCIA POR REGRESION SOLUBLE (w)
10 O. 169 0.1549 1 O. 186 20 0.276 0.2602 2 0.348 30 0.369 0.3654 3 0.500 40 0.434 0.4707 4 0.624 50 0.510 0.5760 5 0.800 60 0.642 0.6812 6 ----- 70 0.732 0.7865 7 _ _ _ _ _ 80 0.820 0.8917 8 ---_- 90 ----- 0.9970 9 ----- 100 ----- 1.1022 lo -----
LINEAL (mg>
En el cuadro 10 y la Gráfica 6 se compararon y se evaluaron cualitativamente ei método indirecto por fenol-ácido sulfúrico y la propuesta metodológica del método colorimétrico desarrollado. Los resultados arrojados por el método colorimétrico para la determinación de almidón soluble son satisfactorios, ya que presentan reproducibilidad y las curvas tipo ajustadas por regresión lineal tienen un alto coeficiente de correlación al igual que el del método fenol-ácido sulfúrico lo que nos proporcionó una mayor confiabilidad y dió pauta a decidir analizar las muestras experimentales por el método colorimétrico.
DATOS qluSrA!JOS POR REGRESION
LINEAL 0.1754 0.3316 0.4878 0.644
0.8002 0.9564 1.1126 1.2688 1.4250 1.5812
10 20 30 40 50 60 70 80 90 100
CANTIDAD DE MUESTRA ( IO a 100 pg de glucosa y I a 10 mg de almidón soluble)
GRÁFICA 6. COMPARCIÓN DE CURVAS TIPO AJUSTADAS, DE ALMIDÓN SOLUBLE Y GLUCOSA
Paíricia Elizabeth Baños Guevara 87
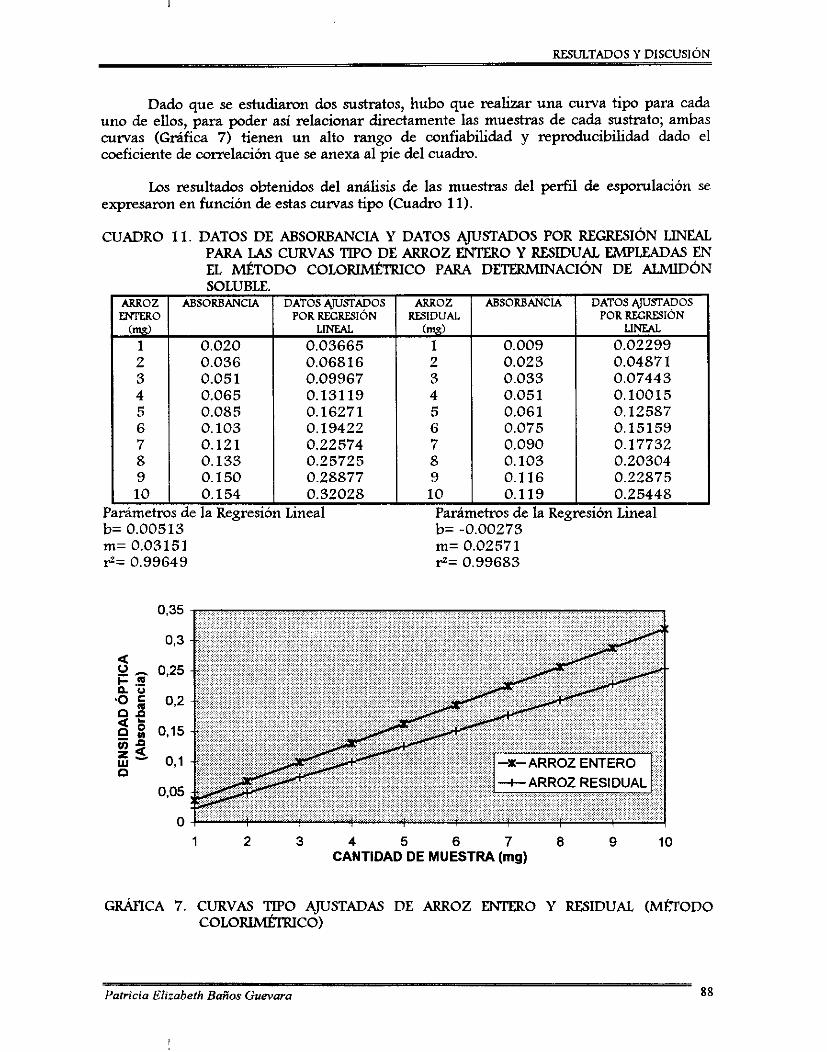
RESULTADOS Y DISCUSIÓN
Dado que se estudiaron dos sustratos, hubo que realizar una curva tipo para cada uno de ellos, para poder así relacionar directamente las muestras de cada sustrato; ambas curvas (Gráfica 7) tienen un alto rango de confiabilidad y repmducibilidad dado el coeficiente de correlación que se anexa al pie del cuadro.
Los resultados obtenidos del análisis de l as muestras del perfil de esporulación se expresaron en función de estas curvas tipo (Cuadro 11).
CUADRO 11. DATOS DE ABSORBANCIA Y DATOS AJUSTADOS POR REGRESIÓN LINEAL PARA LAS CURVAS TIPO DE ARROZ ENTERO Y RESIDUAL EMPLEADAS EN EL &TODO COLORIMÉTRICO PARA DETERMINACI~N DE WÓN
- ARROZ NITRO -s>
1 2 3 4 5 6 7 8 9 10 -
SOLUBLE ABSORBANCIA
0.020 0.036 0.051 0.065 0.085 O. 103 0.121 0.133 0.150
UNEAL 0.03665 0.068 16 O. O 9 9 6 7 0.13119 0.16271 0.19422 0.22574 0.25725 0.28877 0.32028 O. 154
ametros de la Regresion Lineal 0.00513
m= 0.03151 t3= 0.99649
0.35
0 3
0,25
02
0.15
0, l
0.05
O
ARROZ RESIDUAL (w)
1 2 3 4 5 6 7 8 9 10
ABSORBANCIA
0.009 0.023 0.033 0.051 0.061 0.075 0.090 0.103 0.116
DATOS qlusrAfOS POR REGRESION
UNEAL
0.02299 0.04871 0.07443 O. 1001 5 O. 12587 0.15159 0.17732 0.20304 0.22875
0.119 Parámetros de la Regresión Lineal - b= -0.00273 m= 0.02571 rZ= 0.99683
0.25448
1 2 3 4 5 6 7 0 9 10 CANTIDAD DE MUESTRA (mg)
GRÁFICA 7. CURVAS TIPO AJUSTADAS DE ARROZ ENTERO Y RESIDUAL (MÉTODO coLoRuMÉTRIco)
- Patricia Elizabeth Bañar Guwara 88

RESULTADOS Y DISCUSI~N
6.2.5. Método para determinación de viabilidad
Los resultados de ambos tipos de cultivos fueron diferentes: para el cultivo en superficie, el porcentaje de viabilidad fue de 25 a 30%, es un porcentaje muy bajo pem se debe a que la cantidad de rosa de bengala empleada en esta prueba preliminar fue mayor al recomendado en la literatura y se inhibió demasiado la germinación de las esporas.
Con respecto al cultivo líquido sumergido, debido a la escasa aireación y agitación las esporas sumergidas no germinaron y en cambio se formó una capa de micelio en la superficie del medio si el tubo de ensaye permanecía estático. De aquí se concluye que el primer método fue el más adecuado para determinar la viabilidad de B. bmiana, admando sólo una menor cantidad de rosa de bengala.
6.3. Primera fase del experimento
6.3.1. Selección de los mejores tratamientos
Durante la primera fase de este experimento se quiso conocer la influencia de dos de los principales factores ambientales que condicionan el desarrollo del hongo B. bassima en laboratorio, el contenido de agua del sustrato y la temperatura de incubación, así como sus interacciones y el efecto sobre el nivel de esporulación.
Estadísticamente los resultados de selección de los mejores tratamientos estuvieron en función de lo siguiente: el análisis de varianza mostró para ambos sustratos que P(F>Fc)= 0,0001, por tanto se rechaza la hipótesis de nula igualdad de tratamientos y se determinó que existen diferencias signtficativas entre tratamientos, con un nivel de significancia de a=0.05. También se observó que el coeficiente de variación fuera bajo, lo cual indica que la dispersión de los datos es mínima.
Dado que el análisis de varianza mostró diferencias entre tratamientos (Anexo 1 l.l), entonces se procedió a hacer una prueba de comparación de medias, por medio del método de Tukey. Para a m z entero hubo 3 grupos diferentes, de los cuales se escogió el tratamiento 10 (25OC de temperatura de incubación y 65% de contenido de agua) del grupo A por presentar el máximo nivel de esporulación. Para a m z residual señala 6 grupos, de los que se escogió el tratamiento 13 (20°C de temperatura de incubación y 75% de contenido de agua) por las mismas razones.
Evaluando las temperaturas, la prueba de Tukey en m z entero señala 2 grupos, siendo la temperatura de 2 5 T la que presenta la media mas alta; para a m z residual marca 3 grupos, y la temperatura de la media más alta es Z O T .
En cuanto a los contenidos de agua, esta misma prueba m a r a 2 grupos para arroz entero y también para arroz residual, siendo el contenido de agua de 75% el de la media más alta en ambos casos.
Las varianzas entre los residuales del modelo difieren considerablemente, lo cual se explica por la gran manipulación de las muestras y los errores cometidos durante el desamllo del experimento.
P _c_ - - 89 Pairicia Elizabeth Baños Guevara
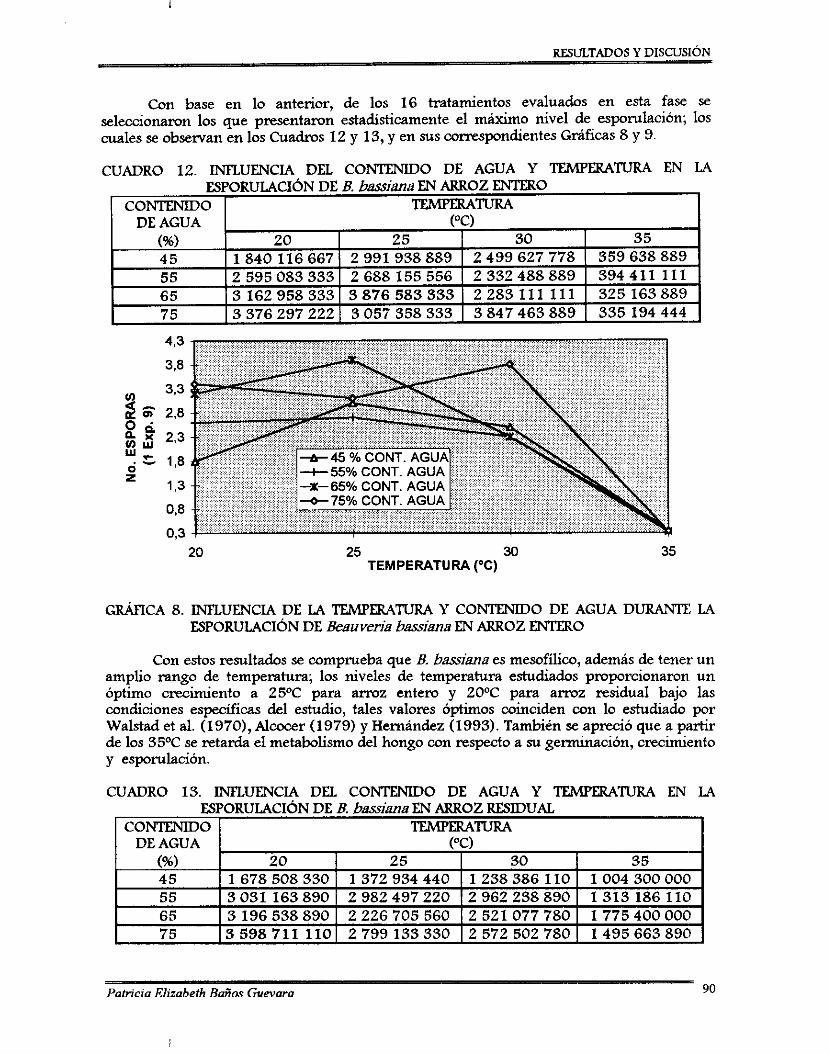
RESULTADOS Y DISCUSI~N
CONTENIDO TEMFmATURA DE AGUA (“C)
(%) 20 25 30 35 45 1840116667 2991938889 2499627778 359638889 55 2 595083333 2688155556 2332488889 394411111
- 65 3 162958333 3876 583333 2283111111 325163889 75 3376297222 3057358333 3847463889 335 194444
Con base en lo anterior, de los 16 tratamientos evaluados en esta fase se seleccionaron los que presentaron estadísticamente el máximo nivel de esporulación; los cuales se observan en los Cuadros 12 y 13, y en sus correspondientes Gráficas 8 y 9.
CUADRO 12. INFLUENCIA DEL CONTENIDO DE AGUA Y TEMPERATURA EN LA
DE AGUA (%) 45 55 65 75
(“C) 20 25 30 35
I678508330 1372934440 1238386110 1004 300000 3031163890 2982497220 2962238890 1313186110 3196538890 2226705560 2 521077780 1775400000 3 598711110 2799133330 2572502780 1495663890
4,3
3.8
+55% CONT. AGUA
20 25 30 TEMPERATURA (“C)
35
GRÁflCA 8. INFLUENCIA DE LA TEMPERATURA Y CONTENIDO DE AGUA DURANTI: LA ESPORULACIÓN DE Beauveria bassiana EN ARROZ ENTERO
Con estos resultados se comprueba que B. bmiana es mesofilico, además de tener un amplio rango de temperatura; los niveles de temperatura estudiados proporcionaron un óptimo crecimiento a 25OC para arroz entero y 2OoC para arroz residual bajo las condiciones específicas del estudio, tales valores óptimos coinciden con lo estudiado por Walstad et al. (1970), Alcocer (1979) y Hernández (1993). También se apreció que a partir de los 35OC se retarda el metabolismo del hongo con respecto a su genninación, crecimiento y esporulación.
CUADRO 13. INFLUENCIA DEL CONTENIDO DE AGUA Y TEMPERATURA EN LA ESPORULACIÓN DE B. bassima EN ARROZ RESIDUAL
I CONTENIDO I TEMPERATURA I
Paincia Elizabeth Bañm Guevara 90
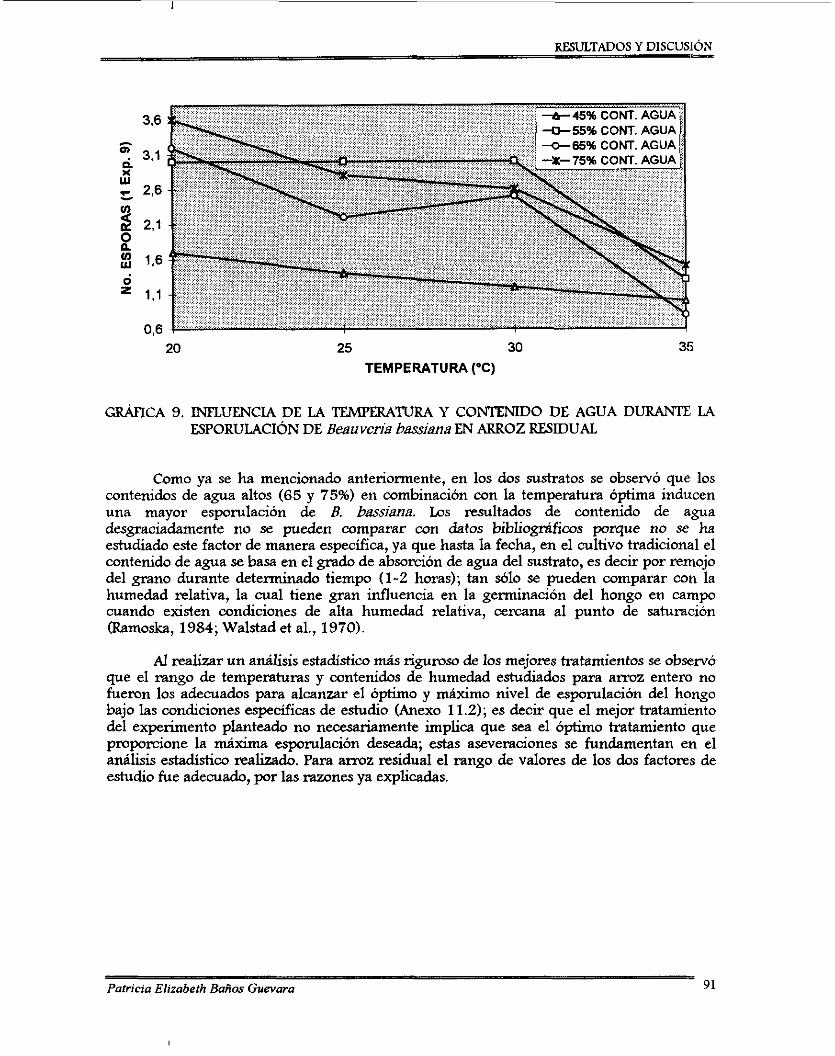
1
RESULTADOS Y DISCUSION -
20 25 30
TEMPERATURA (‘C)
35
GRÁFICA 9. iNFLUENCiA DE LA TEMPERATURA Y CONTENIDO DE AGUA DURANTE LA ESPORULACIÓN DE Beauveria bassima EN ARROZ RESIDUAL
Como ya se ha mencionado anteriormente, en los d o s sustratos se observó que los contenidos de agua altos (65 y 75%) en combinación con la temperatura óptima inducen una mayor esporulación de 6. bassiana. Los resultados de contenido de agua desgraciadamente no se pueden comparar con datos bibliográficos porque no se ha estudiado este factor de manera específica, ya que hasta la fecha, en el cultivo tradicional el contenido de agua se basa en el grado de absorción de agua del sustrato, es decir por remojo del grano durante determinado tiempo (1-2 horas); tan sólo se pueden comparar con la humedad relativa, la cual tiene gran influencia en la germinación del hongo en campo cuando existen condiciones de alta humedad relativa, cercana al punto de saturación (Ramoska, 1984; Walstad et al., 1970).
Al reaiizar un análisis estadístico m á s riguroso de los mejores tratamientos se observó que el rango de temperaiuras y contenidos de humedad estudiados para arroz entero no fueron los adecuados para alcanzar el óptimo y máximo nivel de esporulación del hongo bajo las condiciones específicas de estudio (Anexo 11.2); es decir que el mejor tratamiento del experimento planteado no nemsariamente impiica que sea el óptimo tratamiento que proporcione la máxima esponilación deseada, estas aseveraciones se fundamentan en el análisis estadístico realizado. Para a m z residual el rango de valores de los dos factores de estudio fue adecuado, por las rawnes ya explicadas.
Patricia Elizabeth Baños Guevara 91
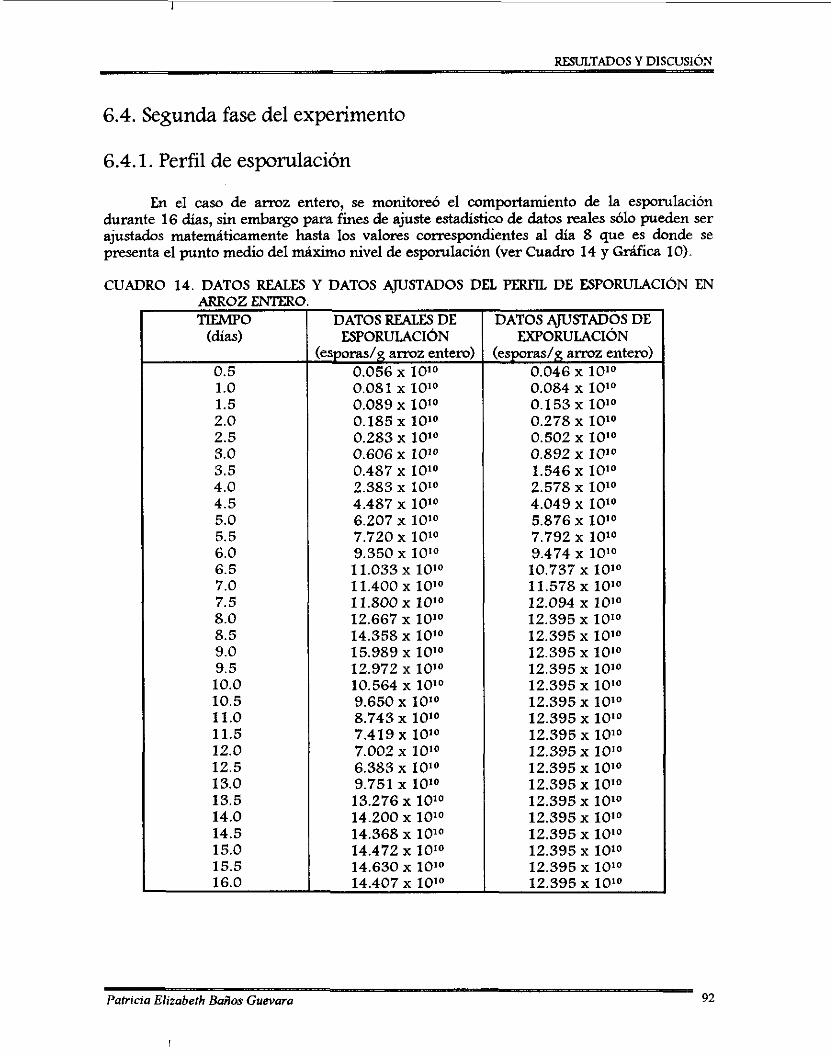
I
RESULTADOS Y DISCUSIÓN
6.4. Segunda fase del experimento
6.4.1. Perfil de esporulación
En el caso de arruz entero, se monitor& el comportamiento de la esponilación durante 16 días, sin embargo para fines de ajuste estadístico de datos reales sólo pueden ser ajustados matemáticamente hasta los valores correspondientes al día 8 que es donde se presenta el punto medio del máximo nivel de esponilación (ver Cuadro 14 y Gráfica 10).
CUADRO 14. DATOS REALES Y DATOS AJUSTADOS DEL PERFIL DE ESPORULACIÓN EN AFXOZ ENTERC TIEMPO
(días)
0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5 5.0 5.5 6.0 6.5 7.0 7.5 8.0 8.5 9.0 9.5 10.0 10.5 11.0 11.5 12.0 12.5 13.0 13.5 14.0 14.5 15.0 15.5
DATOS REALES DE ESPORULACI~N
:esporas/g arroz entero) 0.056 x 10'" 0.081 x 10'0 0.089 x 1010 0.185 x 10'" 0.283 x loao 0.606 x 1O1O 0.487 x 1010 2.383 x 1 O 1 O 4.487 x 1O1O 6.207 x 1O1O 7.720 x 1O1O 9.350 x lozo 11.033 x 1O1O 11.400 x 1O1O 11.800 x 10'" 12.667 x 1O1O 14.358 x 10'O 15.989 x 1O1O 12.972 x 1O1O 10.564 x 1O1O 9.650 x loxo 8.743 x 1010 7.419 x loao 7.002 x 1O'O 6.383 x loto 9.751 x 1Olo 13.276 x 1O1O 14.200 x 1010 14.368 x 1O1O 14.472 x 1O1O 14.630 x 10'" 14.407 x 10'O
DATOS AJUSTADOS DE FXPORULACI~N
(esporas/% arruz entero) 0.046 x 1O'O 0.084 x 1010 0.153 x 1O1O 0.278 x 1O1O 0.502 x 10'O 0.892 x 1O1O 1.546 x 1O1O 2.578 x lolo 4.049 x 1010 5.876 x 1O1O 7.792 x 1010 9.474 x 10'0 10.737 x 1010 11.578 x loxo 12.094 x 10'" 12.395 x 1O1O 12.395 x 10'" 12.395 x 1O1O 12.395 x 1O1O 12.395 x 10'" 12.395 x 1O1O 12.395 x 1010 12.395 x 1O1O 12.395 x 1010 12.395 x 10'" 12.395 x IOxo 12.395 x 12.395 x IO'O 12.395 x 1010 12.395 x 1010 12.395 x 1010 12.395 x 10'O
Patricia Elizabeth Bañar Guevma 92
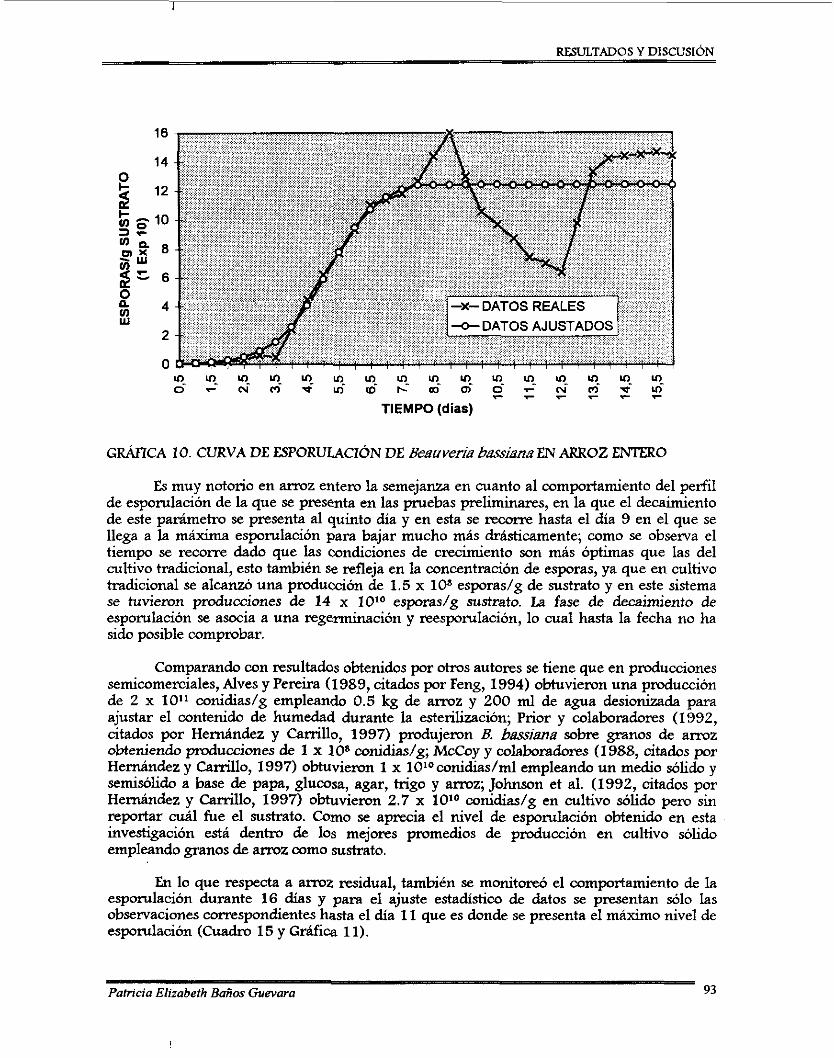
RESULTADOS Y DISCUSI~N
16
14
2
O
GRÁFICA 10. CURVA DE ESPORULACIÓN DE Besuveria bmsiiana EN ARROZ ENTERO
Es muy notorio en a m z entero la semejanza en cuanto al comportamiento del perfil de esponilación de la que se presenta en las pruebas preliminares, en la que el decaimiento de este parámetru se presenta al quinto día y en esta se recorre hasta el día 9 en el que se llega a la máxima esponilación para bajar mucho más drásticamente; como se observa el tiempo se recorre dado que las condiciones de crecimiento son más óptimas que las del cultivo tradicional, esto también se refleja en la concentración de esporas, ya que en cultivo tradicional se alcanzó una producción de 1.5 x lo8 esporas/g de sustrato y en este sistema se tuvieron producciones de 14 x lolo esporadg sustrato. La fase de decaimiento de esponilación se asocia a una regerminación y reesponilación, lo cual hasta la fecha no ha sido posible comprobar.
Comparando con resultados obtenidos por otros autores se tiene que en producciones semicomerciales, Alves y Pereira (1989, citados por Feng, 1994) obtuvieron una producción de 2 x 10" conidiadg empleando 0.5 kg de m z y 200 ml de agua desionizada para ajustar el contenido de humedad durante la esterilización; Prior y colaboradores (1992, citados por Hernández y Carrillo, 1997) produjeron B. bassima sobre granos de arroz obteniendo producciones de 1 x 10s wnidias/g; McCoy y colaboradores (1988, citados por Hemández y Carrillo, 1997) obtuvieron 1 x 10'O conidiadml empleando un medio sólido y semisólido a base de papa, glucosa, agar, trigo y m z ; Johnson et al. (1992, citados por Hemández y Carrillo, 1997) obtuvieron 2.7 x 10'O conidiadg en cultivo sólido pero sin reportar cuál fue el sustrato. Como se aprecia el nivel de esporulación obtenido en esta investigación está dentru de los mejores promedios de producción en cultivo sólido empleando granos de arroz como sustrato.
En lo que respecta a m z residual, también se monitoreó el comportamiento de la esponilación durante 16 días y para el ajuste estadístico de datos se presentan sólo las observaciones correspondientes hasta el día 11 que es donde se presenta el máximo nivel de esponilación (Cuadro 15 y Gráfica 1 1).
Paíricia Elizabeth Bañar Guevara 93
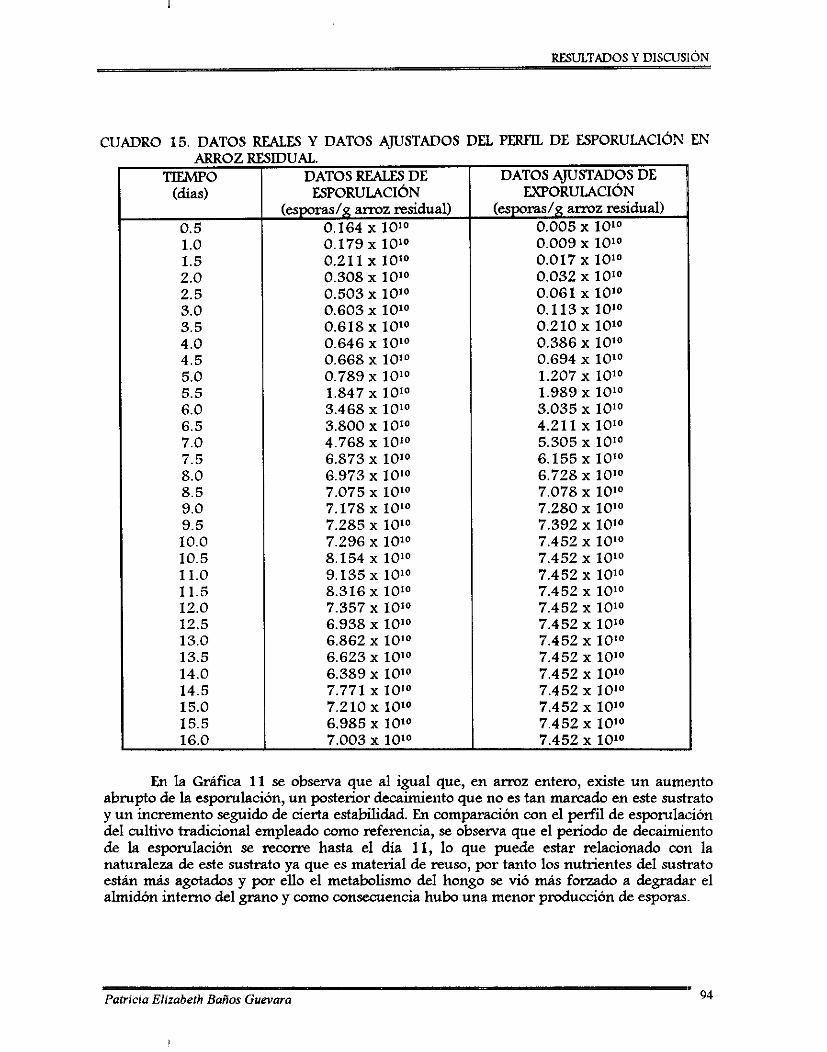
RESULTADOS Y DISCUSI~N
CUADRO 15. DATOS REALES Y DATOS AJUSTADOS DEL PERFIL DE ESPORULACIÓN EN ARROZ R
TIEMPO (dias)
O. 5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5 5.0 5.5 6.0 6.5 7.0 7.5 8.0 8.5 9.0 9.5 10.0 10.5 11.0 11.5 12.0 12.5 13.0 13.5 14.0 14.5 15.0 15.5 16.0
WUAL. DATOS REALES DE
ESPORULACI~N (esporadg a m z residual)
0.164 x loxo 0.179 x 10'O 0.211 x 10'0 0.308 x 10'O 0.503 x 1010 0.603 x 10'O 0.618 x 0.646 x 10'O 0.668 x 1010 0.789 x lolo 1.847 x loTo 3.468 x 10'O 3.800 x 1010 4.768 x 1010 6.873 x 1O1O 6.973 x 1010 7.075 x loro 7.178 x 10'" 7.285 x 1010 7.296 x 1010 8.154 x 1O1O 9.135 x 10'" 8.316 x 10Io 7.357 x 10'0 6.938 x 10'O 6.862 x 10'O 6.623 x 1O1O 6.389 x 1O1O 7.771 x 1O1O 7.210 x 1010 6.985 x loao 7.003 x 1010
DATOS AJUSTADOS DE EWORULACI~N
(esporas/% a m z residual) 0.005 x 1O1O 0.009 x 1O1O 0.017 x 1O1O 0.032 x loxo 0.061 x 10'" 0.113~ 1010 0.210 x 10'0 0.386 x 1O1O 0.694 x 10'O 1.207 x 1010 1.989 x 1010 3.035 x 1O1O 4.21 1 x 1OIo 5.305 x 1010 6.155 x 1010 6.728 x 1010 7.078 x 1010 7.280 x 10Io 7.392 x 1Ojo 7.452 x 1010 7.452 x 1010 7.452 x 1010 7.452 x 10'O 7.452 x 1010 7.452 x 1O1O 7.452 x 10'O 7.452 x 1O1O 7.452 x 1010 7.452 x 1O1O 7.452 x loxo 7.452 x 1010 7.452 x 1O1O
En la Gráfica 11 se observa que al igual que, en a m z entero, existe un aumento abrupto de la esporulación, un posterior decaimiento que no es tan marcado en este sustrato y un incremento seguido de cierta estabilidad. En comparación con el perfil de esporulación del cultivo tradicional empleado como referencia, se observa que el periodo de decaimiento de la esponilación se recorre hasta el día 11, lo que puede estar relacionado con la naturaleza de este sustrato ya que es material de reuso, por tanto los nutrientes del sustrato están más agotados y por ello el metabolismo del hongo se vió más fomdo a degradar el almidón interno del grano y como consecuencia hubo una menor producción de esporas.
Poíricio Elizabeth Baños Gumma - 94
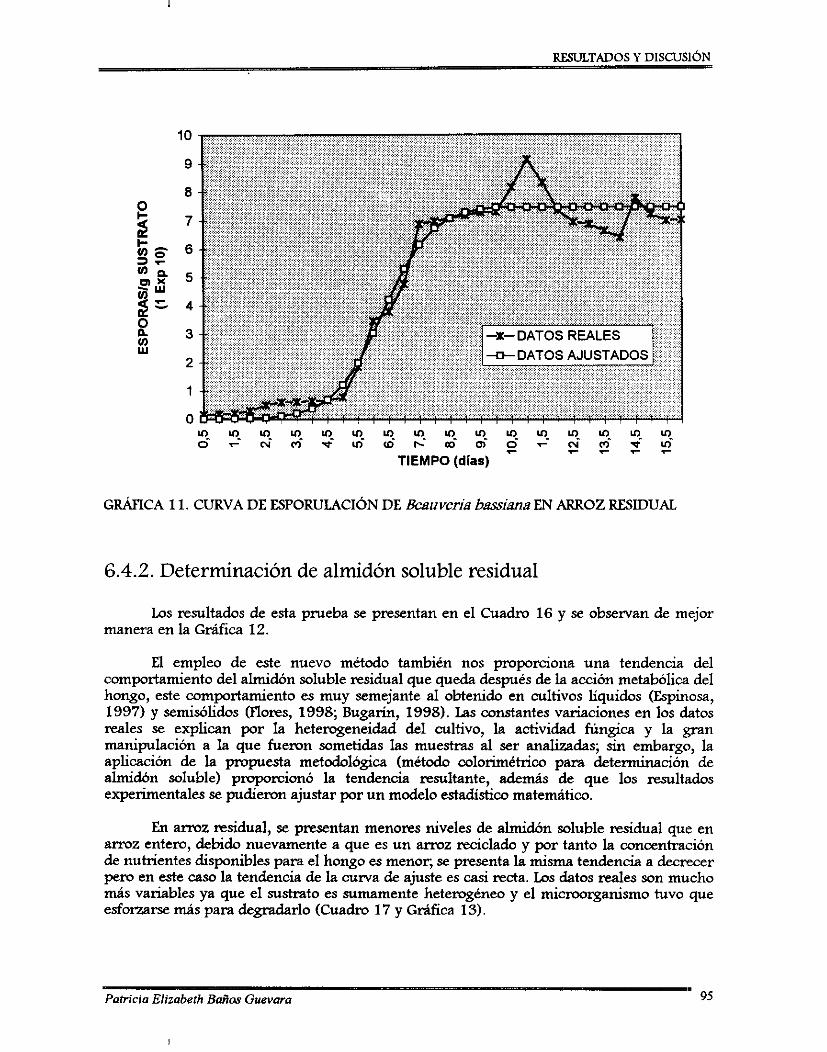
RESULTADOS Y DISCUSI~N P - -
y
TIEMPO (días)
GRÁFICA 1 1. CURVA DE ESPORULACIÓN DE Beauveria bassima EN ARROZ RESIDUAL
6.4.2. Determinación de almidón soluble residual
Los resultados de esta prueba se presentan en el Cuadro 16 y se observan de mejor manera en la Gráfica 12.
El empleo de este nuevo método también nos proporciona una tendencia del comportamiento del almidón soluble residual que queda después de la acción metabólica del hongo, este comportamiento es muy semejante al obtenido en cultivos iíquidos (Espinosa, 1997) y semisólidos (Flores, 1998; Bugarin, 1998). Las constantes variaciones en los datos reales se explican por la heterogeneidad del cultivo, la actividad fungica y la gran manipulación a la que fueron sometidas las muestras al ser analizadas; sin embargo, la aplicación de la propuesta metodológica (método colorimétrico para determinación de almidón soluble) proporcionó la tendencia resultante, además de que los resultados experimentales se pudieron ajustar por un modelo estadistico matemático.
En arroz residual, se presentan menores niveles de almidón soluble residual que en arroz entero, debido nuevamente a que es un arroz reciclado y por tanto la concentración de nutrientes disponibles para el hongo es menor; se presenta la misma tendencia a decrecer pero en este caso la tendencia de la curva de ajuste es casi &a. Los datos reales son mucho más variables ya que el sustrato es sumamente heterogéneo y el micmrganismo tuvo que esfonarse más para degradarlo (Cuadro 1 7 y Gráfica 1 3).
Pafricia Elizabeth Bañar Gumma 95
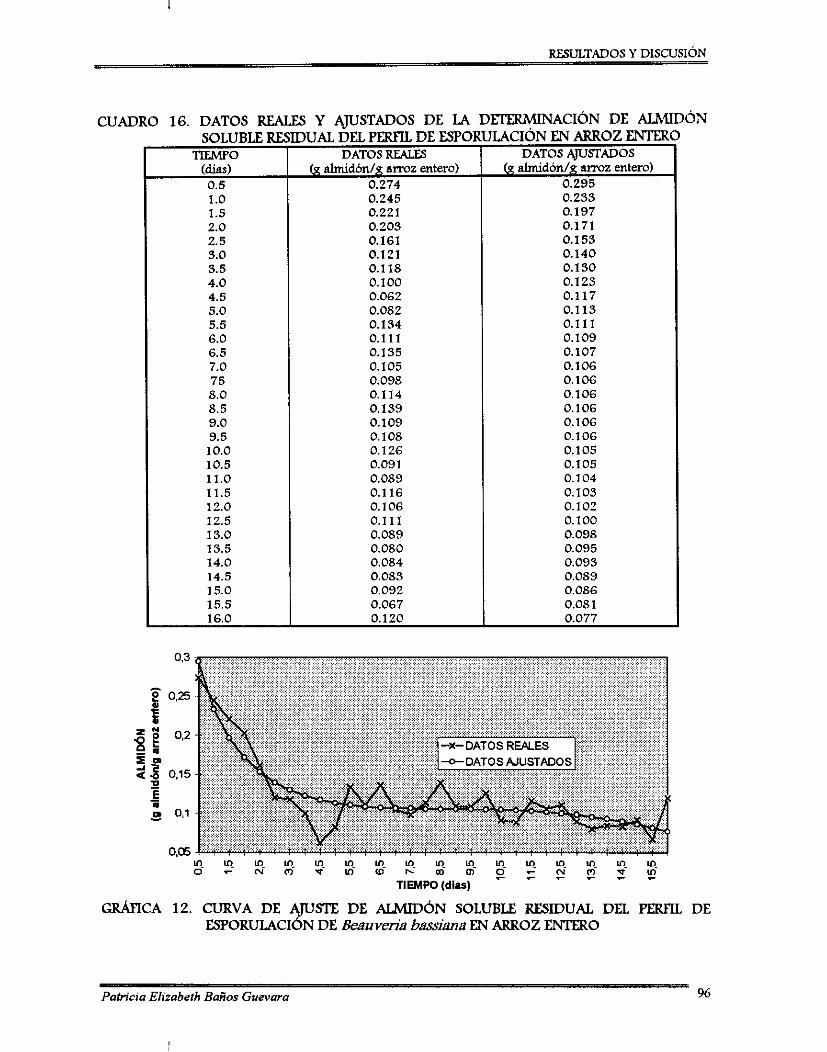
RESULTADOS Y DISCUSI~N -_
llEMP0 DATOS REALES (dias) í.g almidón/% arroz entero)
DATOS qTusrADOS Cs almidón/% a m z entero)
1.0 1.5 2.0 2.5 9.0 3.5 4.0 4.5 5.0 5.5 6.0 6.5 7.0 75 8.0 8.5 9.0 9.5 10.0 10.5 11.0 11.5 12.0 12.5 13.0 13.5 14.0 14.5 15.0
0.5
15.5 16.0
O. 2 74 I 0.295 0.245 0.221 0.203 0.161 0.121 0.118 o. 1 O0 0.062 0.082 0.134 0.111 0.135 0.105 0.098 0.114 0.139 O. 109 O. 108 O. 126 0.091 0.089 0.116 0.106 0.111 0.089 0.080 0.084 0.083
0.233 0.197 0.171 0.153 0.140 o. 1 SO 0.123 0.117 0.113 0.111 0.109 0.107 O. 106 O. 106 O. 106 O. 106 O. 106 O. 106 0.105 0.105 O. 104 O. 103 0.102 0.100 0.098 0.095 0.093 0.089 0.086 I 0.081
0.092 0.067
GRÁFICA 12. CURVA DE STE DE ALMIDÓN SOLUBLE RESIDUAL DEL PERFIL DE Beauveíia bassha EN ARROZ ENTERO
DE
P - Patricia Elizabeth Bañm Gumma 96
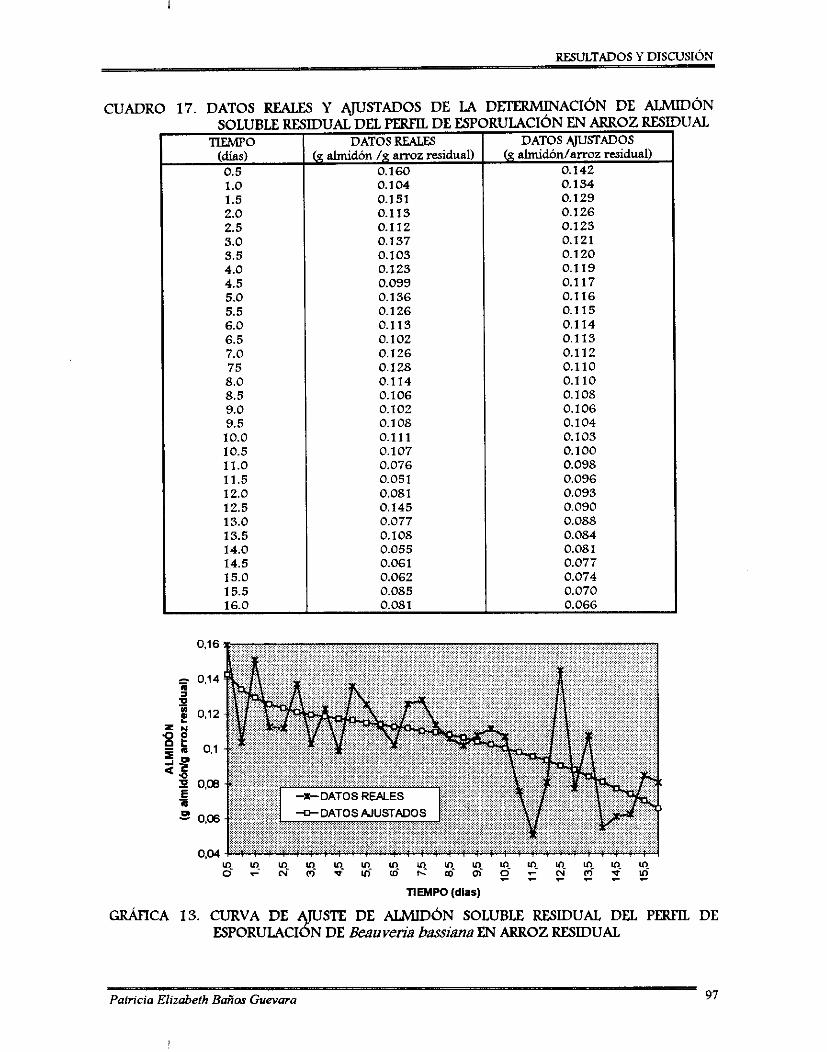
I
TIEMPO DATOS REALES (dias) IS almidón /x a m z residual)
RESULTADOS Y DISCUSION
DATOS W n A D O S IS almidódarroz residual)
O. 5 0.160 ~~~
1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5 5.0 5.5 6.0 6.5 7.0 75 8.0 8.5
O. 142
9.0 9.5 10.0 10.5 11.0 11.5 12.0 12.5 13.0 13.5 14.0 14.5 15.0 15.5 16.0
O. 104 0.151 0.113 0.112 0.137 0.103 0.123 0.099 O. 136 0.126 0.113 o. 1 o2 0.126 o. 128 0.114 0.106 o. I o2 o. 1 O8 0.1 11 O. 107 0.076 0.051 0.081 0.145 0.077 o. 1 O8 0.055 0.061 0.062 0.085 0.081
O. 134 O. 129 0.126 0.123 0.121 0.120 0.119 0.117 0.116 0.115 0.1 14 0.113 0.112 0.110 0.110 0.108 0.106 O. 104 O. I03 o. I O0 0.098 0.096 0.093 0.090 0.088 0.084 0.081 0.077 0.074 0.070 0.066
)UAL
~ . ~ . ~ . 9 9 ~ . 9 ~ . 9 ~ . - - - - - - o - ~ m ~ n w r - m r n ~ = ~ ~ ~ ~
TlEMPü (días)
GRÁFICA 13. CURVA DE STE DE ALMIDÓN SOLUBLE RESIDUAL DEL PERFIL DE DE Beauveria bassima EN ARROZ RESIDUAL
- 91 Patricia Elizabeth Bañas Guevara
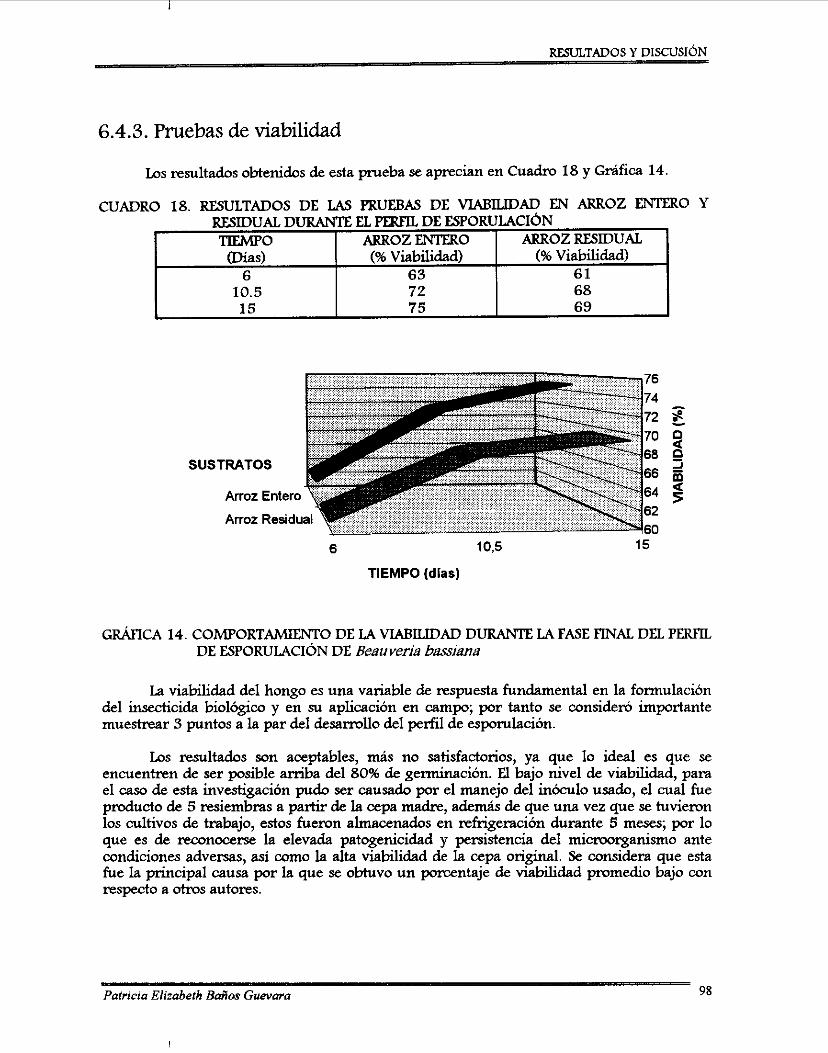
I
TIEMPO ARROZENTERO ARROZ RESIDUAL (Dias) (% Viabilidad) (% Viabilidad)
6 63 61 10.5 72 68 15 75 69
i
RESULTADOS Y DISCUSI~N
76 74 72 70 9 680 6 6 i CUCTRATOS
Arroz Entero 64 2 Arroz Residua
62 60
6 105 15
TiEMPO (dIas)
GRkFICA 14. COMPORTAMiENTO DE LA VIABILIDAD DURANTE LA FASE FINAL DEL PERFIL DE ESPORULACIÓN DE Beauveria bassjana
ia viabilidad del hongo es una variable de respuesta fundamental en la formulación del insecticida biológico y en su aplicación en campo; por tanto se consideró importante muestrear 3 puntos a la par del desamlio del perfil de esporulación.
Los resultados son aceptables, más no satisfactorios, ya que lo ideal es que se encuentren de ser posible arriba del 80% de germinación. El bajo nivel de viabilidad, para el caso de esta investigación pudo ser causado por el manejo del inirculo usado, el cual fue producto de 5 resiembras a partir de la cepa madre, además de que una vez que se tuvieron los cultivos de trabajo, estos fueron almacenados en refrigeración durante 5 meses; por lo que es de reconocerse la elevada patogenicidad y persistencia del microorganismo ante condiciones adversas, así como la alta viabilidad de la cepa original. Se considera que esta fue la principal causa por la que se obtuvo un porcentaje de viabilidad promedio bajo con respecto a otros autores.
P Pahiria Elizabeth Baños Guevara 98

RESULTADOS Y DISCUSI~N I_
SUSTRATO Arroz Entero Arroz Residual
En Colombia, por ejemplo, las cepas madre o puras, se almacenan bajo congelación a 4OC, hasta el momento de emplearlos para hacer nuevas siembras, pero no más de 3 meses; y para la producción del hongo se recomienda emplear cultivos puros de cepa o de segunda generación con lo cual se garantiza el vigoroso crecimiento del hongo, se evitan oportunidades de contaminación y se asegura la infdvidad contra la broca (Brocarta, 1993).
pH INICiAL pH FINAL 6.0 5.7 6.0 5.5
6.4.4. Cambio de pH Esta determinación se realizó para comprobar la modificación del pH con el
metabolismo del hongo en sustrato sólido. Aunque es una dete&ciÓn difícil de medir y no precisa debido a la heterogeneidad del sistema de cultivo y dado que no existe un medio acuoso para que logre una perfecta difusividad de nutrientes. Los resultados confirman lo estudiado por algunos autores en otros sistemas de cultivo, el pH baja; aunque en los sustratos sólidos estudiados, a m z entero y residual, no baja tan drásticamente, como se aprecia en el Cuadro 19.
El pH es un factor de estudio determinante para la germinación de las esporas, crecimiento micelial y esporulación, en cultivo sólido no fluctúa mucho debido a que en este sistema no existió agua libre ni otros nutrientes (peptona, sales minerales) que son los principales inductores para la producción de ácidos metabólicos; por ello B. bassiaria al tener sólo la suficiente cantidad de nutrientes para germinar, crecer y esporular no desvía tanto su metabolismo hacia la producción de otros productos secundarios, como podrían ser los ácidos.
CUADRO 19.
En resumen, la investigación desamliada aporta datos valiosos acerca de la importante infiuencia del efecto del contenido de agua del sustrato, la temperatura de incubación y pH, por tanto, de acuerdo a las condiciones de estudio, se puede señalar que el rango de temperatura Óptima oscila entre 20 y 25OC, el óptimo contenido de agua del sustrato entre 65 y 75% y el pH Óptimo es de 6.0.
Estas condiciones de cultivo afectan directamente el desamllo del hongo y como se apreció Ilifluyen en el nivel de esporulación del mismo, que es la variable de respuesta que nos interesa para poder producir el hongo Beaweria bassima a nivel comercial, por lo que estandarizando al máximo las condiciones ambientales se garantiza cierto nivel de esporulación, y como consecuencia alta viabilidad y elevada patogenicidad en campo.
Pnhicio Eliznbeth Bañar Guevnrn 99

CONCLUSIONES P
CONCLUSIONES
De acuerdo con los resultados obtenidos y con base en los objetivos planteados se líe& a las siguientes conclusiones:
1. El grano de arroz entero así como el arroz residual son buenos sustratos para la pducc ión de esporas de Beauveria b m i m a ya que reportan altos rendimientos en comparación con el sistema de cultivo tradicional. Aunque el arroz residual presenta menores rangos de esporulación que el arroz entero.
2. La temperatura de incubación es un factor condicionante para la esponilación de este hongo, y si se combina con un óptimo contenido de agua aumenta notablemente dicha variable de respuesta.
3. La máxima y óptima espomlación, bajo las condiciones específicas de estudio, en arroz entero fue con una producción de 12.7 x 10'0 esporas/g de sustrato, obtenidas a los 8 días de incubación, bajo unas condiciones de cultivo de 2 5 T de temperatura y 65% de contenido de agua. Para arroz residual la producción considerada como máxima y óptima es de 7.3 x 1 0 ' 0 esporadg de sustrato, obtenidas a los 10 días de incubación y con las condiciones de cultivo de 20°C de temperatura y 75% de contenido de agua.
4. El perfil de esporulación en ambos sustratos presenta al noveno día en arroz entero y al décimo primer día en arroz residual una drástica caida en la esporulación, producto de la naturaleza propia del hongo, las condiciones de cultivo y el tipo de sustrato; ya que esta también se presenta en el perfil de esponilación del cultivo sólido tradicional, cultivo liquido y en cultivo semisólido.
5. La viabilidad del hongo al término del perfil de espomlación fue de 75% en arroz entero y 69% en arroz residual, este parámetm de calidad se vió afectado por el manejo previo al que fueron sometidos los cultivos de trabajo.
6. El pH óptimo inicial para ajuste del contenido de agua de los dos sustratos fue de 6.0, el que tuvo un decremento mínimo no significativo al término del perfil de esporulación como consecuencia de la actividad metabólica propia del hongo.
7. La propuesta metodológica del método colorimétrico para determinación de almidón soluble proporcionó resultados repducibles y una tendencia aceptable para el caso particular de esta investigación.
Pahicia Elizabeth Baños Gumma -100

I
RECOMENDACIONES -
RECOMENDACIONES
1. Emplear cultivos de trabajo jóvenes, no más de 2 generaciones o re:..mbras, para asegurar un aceptable porcentaje de viabilidad y garantizar en consecuencia una alta patogenicidad en campo.
2. Estudiar un rango más amplio de factores de temperatura y contenido de agua, para saber cual es realmente el punto óptimo de producción, así como la interacción con otras condiciones tales como aireación, humedad relativa, luminosidad.
3. Profundizar y fundamentar más la propuesta metodológica del método colorimétrico para determinación de almidón soluble, ya que de avalarse implicaría un gran avance en las técnicas de análisis de muestras ricas en almidón.
4. Efectuar pruebas de patogenicidad en laboratorio y en campo de Beauveria bassima con esporas de las futuras investigaciones, para comprobar si conserva su virulencia tanto en condiciones controladas como en campo.
5. Antes de iniciar cualquier trabajo de investigación sobre este hongo es fundamental la realización de pruebas preliminares para estandarizar y dominar la metodología empleada y el manejo del hongo en laboratorio.
Patricia Elizabeth Baños Guevara 101
I

x. BIBLIOGRAFÍA
Agricultura de las américas. Marzo de 1978. Control biológico de plagas. Lo más natural es también económico. Colombia. Año 27. No. 3s. pp. 28-29,32-33.
Alcocer G., L. 1979. Incidencia del hongo Beauveria bassima en México y su reproducción en el laboratorio para el control microbiológico inducido. W reunión nacional de control biológico (DGSV-SARH). Veracruz, México. pp. 107-1 10.
A lom P., F. R 1985. Avances de un prog rama integrado de investigación contra la b m a . iii Congreso de manejo integrado de plagas. Guatemala, C. A. pp. 263-284.
ANA&. 1981. La broca del fruto del café Hpthenemus hampei F. Subgerencia técnica. Folleto técnico s/n. Guatemala, C. A p. 1 I.
AOAC. 1984. Official methods of analysis. Association of official analytical chemists. 14th. de. Washington D. C., U.S.A
AOAC. 1990. Official methods of analysis. Association of official analytical chemists. 14th. de. Washington D. C., U.S.A
Aregger, E. 1992. conidia pmduction of the fungus Beauveria brongniarfii on Barley and quality evaluation during storage alt 2°C. Journal of invertebrate pathology. 59: 2- 10.
Baker P., S. 1985. Biología e historia natural de la broca del café. En: Memonas del curso sobre manejo integrado de plagas del cafeto con énfasis en broca del fruto (Hypothenemus hampei, Ferr.). iiCA PROMECAFE. ANACAFE. Guatemala, C.A. pp. 105- 143.
Baker P., S; Barrera G., J. F. 1985. La distribución, ecologia y comportamiento de la broca del café en el Soconusco: la información necesaria para ensamblar un programa de control integrado. Memorias del iii congreso de manejo integrado. Guatemala, C. A. pp. 291- 296.
Barson, G. 1977. Laboratory evaluation of Beauveria bassima as a pathogen of the larval stage elm bark bettle, scolytusscolyíus. J. Inv. Pathol. 29: 361-366.
Bauveril. 1997. Internet. Bayer de México, S. A 1980. Café. Sugerencias para mayor producción. Folleto técnico dn.
México, D. F. Bergamin, J. 1944. O “repase” como método do controle da broca do café Hvpothenemus
hampei (Ferr., 1867) (Col. Ipidae). Arq. do inst. Biol. 15: 199-208. Berlanga Padilla, Angélica María. 1995. Aislamiento y conservación de hongos
entornopatógenos. En: Memorias del iii taller de producción masiva de hongos entomopatógenos. Centro nacional de referencia de control biológico (DGSV-SAGAR), Tecomán, Colima, México. pp. 3-9.
Berlanga Padilla, Angélica María. 1997. Aislamiento, identificación y conservación de hongos entomopatógenos. En: Memorias del ii curso-taller de producción masiva de agentes de control microbiano. Centro nacional de referencia de control biológico. h i e d a d mexicana de control biológico. Tecomán, Colima, México. pp. 18-30.
Bklochka, M. J.; Khachatourians, G. G. 1987. Purification and properties of an extracellular protease produced by the entomopatogenic fungus Beauvena bassima Appl. and envirun. microbiol. 53 (7): 1679-1684.
Bidochka, Michael J.; Khachatourians, George G. 1993. Oxalic acid hprpmductioii in Beauveria bassima mutants is related to a utilizable carbon soum but not to virulence. Journal of invertebrate pathology. 62: 53-57.
Bmarta. 1993. Folleto técnico s/n. Colombia. Bugarín López, Alejandro. 1998. Efecto de la concentración de sustrato (harina de arroz) y
de la humedad en la esporulación de Beauveria bassima, en cultivo semisólido. Tesis de licenciatura. Departamento de Ingeniería @industrial. Universidad Autónoma Chapingo. Texcoco, México.
- = Patricia Elizabeth Boiiár Guevara 102

T
Bustillo P., A. E. 1992. El control biológico como un componente en un p g r a m a de manejo integrado de la broca del café, Hpthenemus hampei. Ji Simposio nacional s o h control biológico en Colombia. Colombia. pp. 225-234.
Bustillo P., Alex E.; Posada F., Francisco J. 1997. El uso de entomopatógenos en el control de la broca del café en Colombia. internet.
Bustillos, A; Castillo, H.; Viilalba, D.; Morales, E.; Vélez, P. 1991. Evaluaciones de campo con el hongo Beauveria bassima para el control de la broca del café, Hpthenemus hmpeien Colombia. ASIC. 14O Colloque. San Francisco, Colombia. pp. 679-685.
CAB-international institute of Biological Control. 1989. News from CIBC. 12: 7. Campbell K., K.; Barnes G., L.; Cartwright, B., O; Eikenbary R, D. 1983. Growth and
sporulation of B. bassima and M. anisopliae in a basal medium containkg various cahohydmte sources. Journal of invertebrate pathology. 31 (3): 289-295.
Cárdenas C., H. M. 1995. Producción masiva de hongos entomopatógenos en medio liquido y difásico. En: Memoria del tercer taller de producción masiva de hongos entomopatógenos. Centro nacional de referencia de control biológico (DGSV-SAG.AR). Sociedad mexicana de control biológico. Tecomán, Colima, México. pp. 4 1-53.
Cárdenas C., H. M. 1997. Escalamiento industrial en la producción de entomopatógenos. En: Memoria del U curso-taller de producción masiva de agentes de control microbiano. Centro nacional de referencia de control biológico (DGSV-SAGAR). Tecomán, C o h a ,
Cave D., R 1995. Manual para la enseñanza del control biológico en América Latina.
Cenicafé. 1993. Boletín informativo sobre la broca del café. Colombia. No. 4. Cheng, 2.; Xing, H. P. 1984. Changes of hemolymph proteins in Ostrinia furnacalis with
spores or toxins of Beauveria bassianti Rev. of Appl. Entomol. (Series A) 1985. 73 (1 ) : 51 Clark R., A; Casagrande R, A; Wallace D., B. 1982. influence of pesticides on Beauveria
bassiana, a pathogen of the colorado potato beetle. Envirun. Entomol. 1 1 : 67-70. Conn, Eric E. y Stumpf, P. K. 1975. Bioquítnica fundamental. Segunda edición. Editorial
LiMUSA. México, D. F. Cooke Bridge, W. 1963. A laboratory guide to fungi. in polluted waters, sewage, and
sewage treatment systems. Their identification and culture. Ohio, E.U.A. Crueger, W.; Crueger, A. 1984. Biotechnology: a textbook of industrial microbiology.
ScienceTech. inc. Madison, Wisconsin,U.S.A pp. 116-117. Daoust, Richard A; Pereka, Roberto M. 1986. Stability of entomopathogenic fungi
Beauveria bmsiana and Metarhizium anisopliae on beetle-attracting tubers and cowpea foliage in Brazil. Envirunmental entomology. 15 (6): 1237-1243.
Daoust, Richard A; Ward, Michael G., Roberts, Donald W. 1983. Effect of formulation on the viability of Metarhizium anisopliae conidia. Journal of invertebrate pathology. 4 1:
De la Rosa R, W. 1993. Manejo del hongo entomopatógeno Beauveria bassima (Bals.) Vuill. y su efecto sobre la broca del café Hvpothenemus hmpzi Ferr. y su parasitoide Cephalonomia stephanodens Betrem. Tesis de maestría en protección vegetal. Departamento de parasitologia agrícola. Universidad Autónoma Chapingo. Texwco, México.
Diario Oficial de la Federación. 1997. Norma Oficial Mexicana NOM-002-FiTO-1995 por la que se establece la campaña contra la broca del café. Tomo DXX. No. 5. pp. 22-30.
Día2 V., V. M. 1993. Efecto del hongo entomófago Beauveria bmiana (Bals.) Vuiíl. en el control de la broca del grano de café Hpthenemus hampei Ferr. en Cacahotán, Chiapas. Tesis de maestría en ciencias en protección vegetal. Departamento de Parasitologia Agrícola. Universidad Autónoma Chapingo. Texcoco, México.
México. pp. 17-26.
Primera Edición. Tegucigalpa, Honguras.
151- 160.
Potricio Elizobeth Boiiár Guevmo 103
I

Doberski, J. W. 1981a. Comparative laboratory studies on three fungal pathogens of the elm bark beetle scolytus scol@tus effect of temperature and humidity on infection by Beauveria bassima, Metarhizium anisopíiae, and Paecilomyces fmnmus. Jainial of invertebrate pathology. 37: 195-200.
Doberski, J. W. 1981b. Comparative laboratory studies on three fungal pathogens of .the elm bark beetle scolyts scoíytus pathogenicity of Beauveria bassan4 Mefarhizium anisopliae, and Paecilomyces farinms to larvae and adults of S. scoíytus. Journal of invertebrate pathology. 3 7 188-194.
Doelle, Horst W.; Mitchell, David k and Rolz, Carlos E. 1992. Solid substrate cultivation. Usevier applied science. England.
Duriez-Vaucelle, T.; Fargues, J.; Robert P., H.; Popeye, R 1981. Etude enzymatique compaseé de champignons entomopathogénes des generes Beauveria et Methamhizirzm. Mycopathologia. 75: 109-126.
Espinosa Sánchez, Raúl. 1997. Efecto de la concentración del sustrato (almidón) en. la producción de esporas de Beauveria bassiana en cultivo líquido sumergido. Tesis de licenciatura. Departamento de Ingeniería Agroindustrial. Universidad Autónoma Chapingo. Texcoco, México.
Fargues, J.; Goettel M., S.; Smits, N.; Ouedraogo, A; Rougier, M. 1997. Effect of temperature on vegetative growth of Beauveria bassiana isolates from different origins. Mycologia. 89
Fargues, J.; Robert P., H. 1985. Persistance des wnidiospores des hyphomycetes entomopathogenes Beauveria bassiana (Bals.) Vuill., Metarhizium anisopíiae (Metsch.) Sor., Nomuraea riíeyi (F.) Samson et Paeciíomyces fumosoroseus wize dans le sol en conditions controleés. Agronomie. 5 81): 73-80.
(3): 383-392.
Feng. 1994. Producción de hongos entomopatógenos. Folleto técnico s/n. Colombia. Ferrun, P. 1977. influence of relative humidity on the development of fungal infection
caused by Beauveria bassiana (fungi imperfecti, Moniliales) in imagines of Acsnthoscelides obfecfus (Col Bruchidae). Entomophaga. 22 (4): 393-396.
Ferrun, P. 1978. Biological wntrol of insect pests by entomogenous fungi. Annual reviews entomology. 23: 409-420.
Femn, P. 1981. Pest control by the fungi Beauveria and Metazhizium. In: Microbial control of pests and plant diseases 1970- 1980. De. Burges, H. D. 24. Academic Pres. New York.
Flores Pérez, G. 1998. Estudio cinético de espomlación de Beauveria bassiana en maíz molido utilizando un sistema de cultivo semisólido. Tesis de licenciatura. Departamento de Ingeniería Agroindustrial. Universidad Autónoma Chapingo. Texcoco, México.
Frazier W., C. 1976. Microbiología de los alimentos. Editorial Acribia. Zaragoza, Espana. Segunda Edición. pp. 15-35.
Galán C., L. M.; Bodegas V., P. R. 1984. Estudio preliminar para el desarrollo de una estrategia de manejo integrado de la broca del cafeto (Hjpofhenemus hampei Ferr.) (Coleóptera: Scolytidae) para la región del Soconusco, Chiapas, México. En: Memorias del I1 congreso nacional de manejo integrado de plagas. Guatemala, C. A. pp. 195-207.
Garza G., E. 1995. Reproducción masiva de hongos entomopatógenos en sustratos sólidos. En: Memoria del 111 taller de producción masiva de hongos entomopatógenos. Centro nacional de regerencia de control biológico (DGSV-SAGAR). Tecoman, Colima, México.
Gorinstein, Shela; Paccola-Meirelles, Luna Doretto; Aparecida Belo, Valeria; Lucio hevedo, Joao. 1996. Characterization of Beauveria bassima, Metarhizium anisopíiae and Aspergiillus niduíms t h u g electmphoretic patterns of their protein fractions. Joounal of fermentation and bioengineering. 82 (2): 89-92.
pp. 465-482.
pp. 10-16.
Patricia Elizabeth BañÓs Guevara 104

Hajek E., A; Leger St., R J. 1994. Interactions between fungal pathogens and insect hosts. Annual reviews entomology. 39: 293-322.
Hegedus, Dwayne D.; Bidochka, Michael J.; Khachatourians, George G. 1990. Beauveria bassima submerged conidia production in a defined medium containing chitin, two hexosamines or glucose. Applied and mimbiology biotechnology. 33: 64 1-647.
Hemández Escareño, Alfonso Gerardo. 1993. Biología e identificación de Lugus Spp. (Hemíptera: Miridae) y pruebas de patogenicidad con cepas del hongo Beauveria bassima (Balsamo) Vuillemin. Tesis de maestria en ciencias. Colegio de postgraduados. Montedo, México. pp. 9-14.
Hernández Mendoza, José Luis. 1995. Situación actual del control mimbiano en México. En: memorias del iii taller de producción masiva de hongos entomopatógenos. Centro nacional de referencia de control biológico (DGSV-SAGAIL). Tecornán, C o k , MéXico. pp. 1-2.
Hemández Velázquez, Victor M. y C a d o Aguilar, Manuel. 1997. Producción masiva en sustrato sólido y formulación de hongos entomopatógenos. En: ii curso-taller de producción masiva de agentes de control mimbiano. Centro nacional de referencia de control biológico. h i e d a d mexicana de control biológico. Tecomán, C o b a , México. pp. 31-40.
instituto Brasilek do Cafe. 1973. Recomendawes para controle: ferrugem, broca do cafe, bico mine&, cexwsporiose, tombamento, cochodas, lagartas, acaros, micronutnentes. Folleto técnico s/n. p. 47.
INMECAFÉ (Instituto Mexicano del Café). 1983. Control de la broca del grano del café. Folleto. iM.C. Xalapa, Veracruz, México.
Jiménez Zilli, Jorge. 1995. Charlas de café ... sin broca. Boletín informativo. Comité Regional de Sanidad Vegetal “Huatusco”. Huatuxo, Veracruz, México.
Koch, K. C. 1989. Perspectivas en el control bioteconológico de la broca del café (Hypothenemus hampeiFerr.). ASIC. 13O Colloque. Paipa, Colombia. pp. 717-725.
Krueger, Stephen R; Nechols, James R; Ramoska, William k 1992. Habitat distribution and infection rates of the fungal pathogen Besuveria bassiana (Balsamo) Vuillemiri in endemic populations of the chinch bug, Blissus leucopterus (Say) (Hemiptera: Lygaeidae) in Kansas. Journal of the Kansas entomological society. 65 (2): 115-124.
Kucera, M. 1971. Toxins of the entomophagous fungus Beauveria bassiana ii effect of nitrogen sources on formation of the toxic protease in submerged culture. Journal of invertebrate pathology. 17: 211-215.
Leopold, J.; Samsináková, Anna. 1970. Quantitative estimation of chitinase and several otrher enzymes in the fungus Beauveria bassima 15: 34-42.
Le Pelley, R. H. 1973. Las plagas del café. Apicultura tropical. Editorial Labor, S. A. Barcelona, España. pp. 140-170.
Lemma Gutiérrez, Roberto; Molina Ochoa, Jaime. 1997. EpizoOtiología de agentes microbiales. En: ii curso-taller de producción masiva de agentes de control microbiano. Centro nacional de referencia de control biológico. sociedad mexicana de control biológico. Tecomán, Colima, México. pp. 10-17.
Lisansky S., G. 1985. Microbial insecticides. Biotechnology and its application to agriculture. 32: 145-151.
Madelin M., F. 1963. Desease caused by Hyphomycetous fungi. In: insect pathology and advances treatise. Volume 2. Edited by Eduard Steinhaus. Academic Press. pp. 233-264.
Marcandier, S.; Khachatourians G., G. 1987. Susceptibility of the migratory grasshopper, Melanoplus sanguinijxs (Fab.) (Orthoptera: Acrididae), to Beauveria bassima (Bals.) Vuillemin (Hyphomycete): influence of relative humidity. The canadian entomologist. 119: 901-907.
Pairicia Elizabeth Bañós Guevara 105

1
Méndez López, Ismael. 1990. Control micmbiano de la broca del fruto del cafeto Hypothenemus hampei Femri (Coleóptera: Scolytidae), con el hongo Beauvena bassiana (Bals.) Vuill. (Deutemmycetes) en el Soconusco, Chiapas. Tesis de maestría en ciencias. Colegio de Postgraduados.
Monterroso, J. L. 1981. incidencia de Beauveria bawiana sobre la broca del café y su reproducción en coco en Guatemala. Revista cafetalera ANACAFÉ. 6 (210): 10- 12.
Morales, E.; Cruz, F.; Ocampo, A; Rivera, G.; Morales, B.. 1991. Una aplicación de la biotecnología para el control de la broca del café. ASIC. 14" Colloque. San Francisco, Colombia. pp. 521-526.
Olmert, Y.; Kenneth R, G. 1974. Sensitivity of the entomopathogenic fungi Beauveria bawiana, Verticillium Iecanii and Verticillium spp. to fungicides and insecticides. Envkn. Entomol. 3 (1): 33-38.
Paccola-Meirelles L., D.; Araujo W., L. 1993. Mycelial development and nuclear differentiation in the Deutemmycete Beauveria bassiana cultivated in different liquid media. Micmbiologica. 16: 73-78.
Pearson, M, Hall, L.; Soderhall, K. 1984. Comparison of peptidase adivites in some fungi pathogenic to arthropods. Journal of invertebrate pathology. 44: 342-348.
Prior, C.; Jollands, P.; Le Patourel, G. 1988. infectivity of oil and water formulations of Beauveria bassima @eutemmywtina: Hyphomycetes) to the cocoa weevil pesi Pantorhfles plutus (Coleoptera: Curculionidae). Journal of invertebrate pathology. 52:
Quezada J., R 1985. Factibilidad del uso de enemigos naturales de la broca del café (Hypothenemus hampei). Actividades en Turrialba. lS(4): 1-8.
Ramaraje urs., N. V.; Govindu H., C.; Shivashankara Shastry, K. S. 1967. The effect0 of certain insecticides on the entomogenous fungi Beauveria bassiana and Metarrhizium anisopliae. Journal of invertebrate pathology. 9: 398-403.
Ramoska W., k 1984. La influence of relative humidity on B. bassiana infectivity and replication in the chinch bug, Blissus leucoptems. Journal of invertebrate pathology. 43:
Rendón S., G. 1995. Métodos estadísticos. Universidad Autónoma Chapingo. Texcoco, Mexico.
Rocha Garcia, Celia. 1996. La broca, plaga que afecta la parte esencial del café. Agronegocios en México. No. 19. pp. 11-14,
Rhodes, A,; Fletcher D., L. 1969. Principios de microbiología industrial. Editorial Acribia. Zaragoza, España. pp. 82-85.
Rodriguez C., E. 1984. Sensibilidad del hongo entomopatógeno Beauveria bassiana (Bals.) Vuill. a 7 fungicidas bajo condiciones de laboratorio. En: Memorias del ii congreso nacional de manejo integrado de plagas. Guatemala, C. k pp. 2 19-225.
Rojas C., J. J. 1982a. Efecto de la ingestión de toxinas producidas por el hongo Beauveria spp. sobre la mortalidad de adultos y larvas de la pulga saltona Epiírzx spp. Turrialba. 32 (I): 93-95.
Rojas C., J. J. 1982b. Efecto de la ingestión de toxinas producidas por el hongo Beauveria spp. sobre la mortalidad de adultos y larvas de la pulga saltona Epitrx spp. Turrialba. 32
Rosin, Faye. 1996. Note: effects of fertilizers on the survival of Beauveria bassima Journal
Ruales, Carlos. 1996. Manejo integrado de bruca del café (MiBC): la experiencia mexicana.
Russell B., Stevens. 1974. Mycology guidebook. University of Washington Press. EUA.
66-72.
389-394.
(1): 91-92.
of invertebrate pathology. 6 8 194-195.
En: Café & cacao: noticias. Nestle. 1 (2) : 2-3.
- - Patricia Elizabeth Bañ& Guevara '-106

1
BIBLIOGRAFh
Saito, T. 1984. Effect to pesticides on conidial germination and hyphal growth of the entomopathogenic fungus Beauveria bassiana. in: Entomology Abstracts. 1986. 17 (5): 42.
Samsináková, A 1966. Growth and sporulation of submersed cultures of the fungus Beauveria bassiana in various media. Journal of invertebrate pathology. 8: 395-400.
Samsináková, Anna; Hrabetová, Eva. 1969. Respiration of blastospores of the fungus Beauveria bassiana during submersed cultivation in the presence of certain sugars. Journal of invertebrate pathology. 13: 382-385.
Samsináková, Anna; Kálalová, S. 1981. Mass production of B. bassiana for regulation of Igptinotarsa decemiineata populations. Journal of invertebrate Pathology. 40: 299-300.
Samsináková, A; Misikova, S., Leopold, J. 1971. Action of emhat ic systems of Beauveria bassima on the cuticle of greater Wax moth larvae (Gaiieria rneiioneiia). Journal of invertebrate pathology. 18: 322-330.
Samson, Robert A; Evans H., C. 1982. Two news Beauveria spp. from South America. Journal of invertebrate pathology. Vol. 39. pp. 93-97.
Santos Moreno, Armando. 1995. Química y bioquímica de alimentos. Dirección general de difusión cultural. Primera Edición. Universidad Autónoma Chapingo. Chapingo, México.
SARH (Secretaria de Agricultura y Recursos Hidráulicos). 1980. Broca del Café Hpthenemus hampei (Ferran). Folleto Técnico s/n. México, D. F.
Serrano C., L. M. 1996. Notas del curso de metodología de la investigación. Departamento de ingeniena Agroindustrial. Universidad Autónoma Chapingo. Texcoco, México.
Smith, Rebecca J.; Grula E., A 1981. Nutritional requeriments for conidial germination and hyphal growth of Beauveria bassiana Journal of invertebrate pathology. 37 (3): 222- 230.
Storey G., K; Gardner A,, W. 1986. Sensitivity of the entomogenous fungus Beauveria bassiana to selected plant growth regulators and spray additives. Appl. and Env. Microbiol. 52 (1): 1-3.
Tedders W., L. 1981. In vibw inhibition of the entomopathogenic fungi Beauveria bassiana and Mefarhizium anisopliae by six fungicides used in pecan culture. Environ. Entomol.
Thomas C., K; Khachatounans G., G. and Ingledew M., W. 1987. Production and properties of Beauveria bassiana conidia cultivated in submerged culture. Can. J. Microbiol. 33 (1): 12-20
Vela- Pascual, Hermenegildo. 1995. La broca del café Hpothenemus hampei, Ferr. Su control efectivo aplicando manejo integrado. Programa de investigación y desarrollo en regiones cafetaleras. Universidad Autónoma Chapingo. Texcoco, México.
Velez Arango, Patricia Eugenia; Montoya R, Esther Cecilia. 1993. Supervivencia del hongo Beauveria bassiana bajo radiación solar en condiciones de laboratorio y campo.
Villanueva Manufo, Alfonso E. 1986. I Reunión intercontinental sobre broca del café. Centro de investigaciones ecológicas del sureste unidad Tapachula. Tapachula, Chiapas, México. pp. 13-15.
Walstad J., D.; Anderson R, F. and Stambaugh W., J. 1970. Effects of environmental conditions on two species of muscardene fungi (Beauveria bassiana and Metarhizium ani’piiae). J o u m l of invertebrate pathology. 16: 221-226.
West, E. J.; Briggs, J. D. 1968. in vitro toxins production by the fungus Beauveria bassiana and bioassay in greater wax moth larvae. J. of Econ. Entomol. 61 (3): 684-687.
Woods, Sue P.; Grula E., A 1984. Utilizable surface nutrients on Heliothis zea available for growth of Beauveria bassima Journal of invertebrate pathology. 43: 259-269.
10: 346-349.
CENICAFÉ. 44 (3): 111-122.
P
Patricia Elizabeth Bañ& Girevara -107

Wulf, k 1983. Investigations on the entomopathogenic fungus Beauveria bassima (Bals.) Vuill. parasiting the spruce wood engraver í'jfyogenes chalcographus L. (Col. Scolytidae). Entomology abstracts. 14 (6): 31.
Patricia Elizabeth Bañós Guevara 108

Xi. ANEXOS 11.1. Programa 1
Arroz entem
PROGRAMA DATA & INFTJT TRATS REFS TEMPS HUMS X x=x/100000; CARDS; ... , PROC MEANS MWN VAR nD; CLASS TRAT; PROC GLM; CLASS TRAT; MODEL X-íRAT; MEANSTRATiTUKNLINES; OUTPUT OüT=RESIDS R=RESIDS R=RX; PROC UNWARUTE NORMAL PLOT; VAR IR PRSC CLM DATA=& CLASS TEMF HUM, MODEL X 3 E M P HUM TEMP*HUM MEANS TEMP HUM TEMP*HUM/WKEY LINES. RUN;
RESULTADOS Analysis Variable : X TRAT NObs Mean Variance Std Dev
1 3 18401.17 10 3 3324.89 I1 3 22831.11 12 3 38474.64 13 3 3596.39 14 3 3944.11 15 3 3251.64 16 3 3351.94 2 3 25950.83 3 3 31629.58 4 3 33762.97 5 3 29919.39 6 3 26881.56 7 3 38759.17 8 3 30573.58 9 3 24996.28
2624432.85 1620.01 20309720.05 4506.63 62596494.21 7911.79 9898 12 1 O 9948.93
344898.18 587.28 178684.51 422.71
1181439.64 1086.94 96208 1.36 980.85 971996.97 985.89
29704663.08 5450.20 73929000.42 8598.20 45803631.02 6767.84 33546604.97 5791.94 09998562 10488.02 ~
9424208.52 3069.89 14368837.68 3790.62
Class Level Information Class Levels valuw
TRAT 16 1 1 0 1 1 1 2 I S 1 4 1 5 1 6 2 3 4 5 6 7 8 9 Number of observations in data set = 48
Dependent Variable: X SOUrCe DF SumofSquairs MeanSquare FVdue Pr>r Model 15 7031156909 468743734 14.85 O.OOO1 Error 32 1009852930 31557904 CorirctedTotal 47 8041009839
R - S q U X C.V. Roat MSE X Mean 0.874412 24.99165 5617.642 22478.0781
General linear Models P d u n Tukefs Síudcnázed Range (HSD) Test for variable: X Alpha= 0.05 df= 32 MSE= 31557904 Critical Value of Studentlzed Range= 5.244 Minimum S i c a n t Difference= I7008 Means with the same letter arc not signüícantiy different.
Patricia Elizabeth Bañar Guevara 109

Tukcy Grouping Mean A 98759
B B B B 8 B B B B B
A 98475 A 99769 A 31690 A 90574 A 29919 A 26882 A 25951 A 24996 A 29928 A 22891 C 18401 C 9944 C 9596 C 9952 C 9252
N 9 9 9 9 3 9 3 9 9 9 9 3 9 9 9 9
TRAT 7
12 4 3 8 5 6 2 9
10 11
1 14 13 16 15
General !inear Models Pmcedure Class Level Information
Class Levels Values TEMP 4 20253095 HUM 4 45556575 Number of obsewations in data set = 48
General linear Models P d u r e Dependent Variable: X
SoUrCe DF SumofSquarrs MeanSquare FValue Pr>F Model 15 7091156909 468749794 14.85 0.oOOI Emor 92 1009852930 31557904 CorrcdedTotal 47 8041009899
&Square C.V. Root W E X Mean 0.874412 24.99165 5617.642 22478.078 1
General linear Models Procedure Dependent Variable: X Sourie DF TypeIsS MeanSquair FValuc Pr>F TEMP 3 5876097408 1958699196 62.07 0.0001 HUM 9 429257152 149085717 4.53 0.0093 TEMP'HUM 9 725802350 80644706 2.56 0.0244
Tukey's Shidentized Range (HSD) Twt for variable: X Alpha= 0.05 df= 92 W E = SI557904 Critical Value of Shidentized Range= 9.832 Minimum Significant Difíenncc= 6219.6
Means with the same letter a n not sinnificantlv different. TukcyGmuping Mean N -TEMP
A SI539 12 25 A 27496 12 20 A 27407 12 SO
8 9596 12 95
Tukefs Shidentizsd Range (HSD) Tcst for variable: X Alpha= 0.05 df= 32 MSE= 91557904
Critical Value of Studentized Range= 9.892 Minimum Significant Diffenncc= 6213.6
T u k e y C m u ~ Mean N HUM Means with the same l&r are not significantly different.
A 26541 12 75 S A 24118 12 65 8 20025 12 55 B 19228 12 45
Patricia Elizabeth Bañar Guevara 110
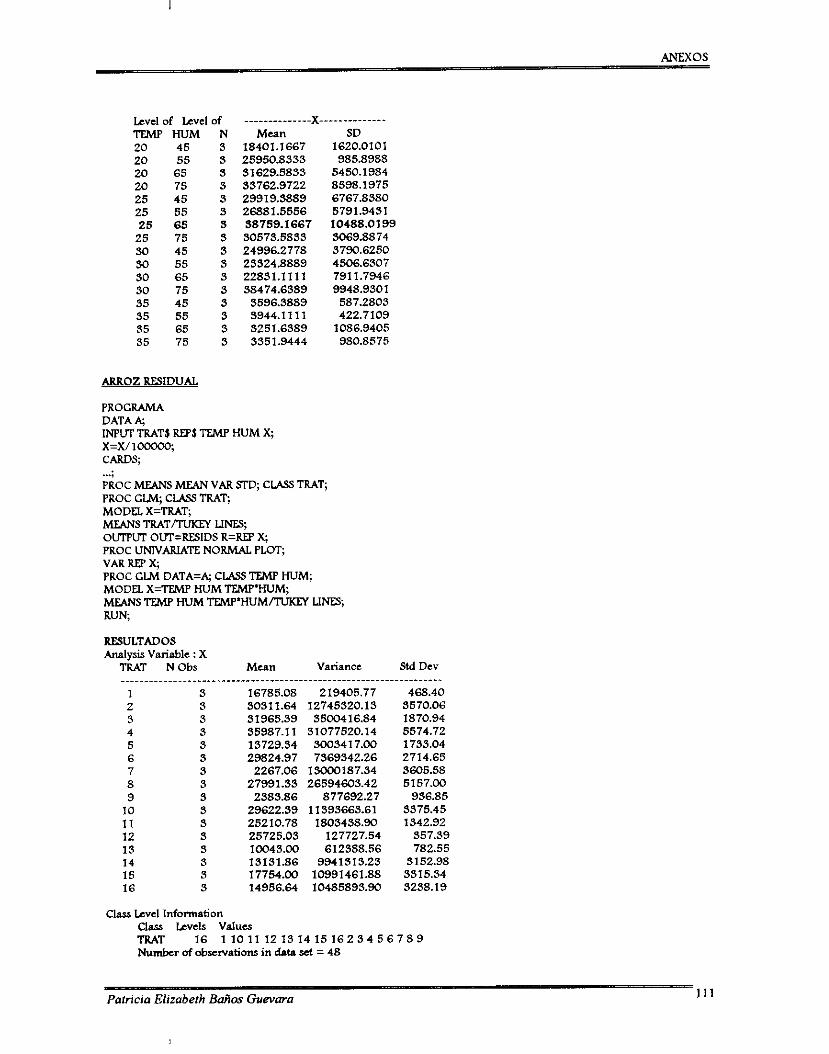
I
Level of Level of TEMP HUM N 20 45 3 20 55 9 ~
20 65 9 20 75 9 25 45 3 25 55 9 25 65 5 25 75 9 SO 45 9 so 55 9 SO 65 3 90 75 3 35 45 9 35 55 9 95 65 9 95 75 9
...----------- x.-------....-.
18401.1667 1620.01 O 1 25950.8333 985.8988 9 1629.5899 5450.1984 99762.9722 8598.1975 29919.9889 6767.8380 26881.5556 5791.9431 58759.1667 10488.0 I99 90575.5899 9069.8874 24996.2778 9790.6250 29924.8889 4506.6307
Mean SD
22831.1 1 1 1 791 1.7946 98474.6989 9948.9901
9596.3889 587.2803 9944.1111 422.7109 9251.6989 1086.9405 3351.9444 980.8575
ARROZ RESIDUAL
PROGRAMA DATA A: INPUT TRATS REPS TEMP HUM X; x=x/100000; CARDS; ..., PROC MEANS MEAN VAR STD; CLASS TRAT; PROC GLM, CLASS TRAT; MODEL X=TRAT; MEANS TRAT/TUKEY UNE% ~
OvíFuT OUT=RESIDS R=REP X; PROC UNNARIATE NORMAL PLOT. VARREPX; PROC GLM DATA=A; CLASS TEMP HUM MODEL X=TEMp HUM TEMP'HUM, MEANS TEMP HUM TEMP"UM/TUKEY LIm, RUN,
RESULTADOS Analysis Variable : X
TRAT NObs Mean Variance
1 3 16785.08 219405.77 2 9 50911.64 12745320.13 3 3 91965.39 9500416.84 4 3 95987.11 91077520.14 5 3 19729.94 9003417.00 6 9 29824.97 7969942.26
Std Dev
468.40 9570.06 1870.94 5574.72 1799.04 2714.65
7 8 9 10 1 1 12 13 14 15 16
3 2267.06 13000187.94 9605.58 3 27991.33 26594605.42 5157.00 9 2883.86 877692.27 936.85 9 29622.99 11999669.61 9375.45 3 25210.78 1803438.90 1342.92
~
3 25725.09 127727.54 357.39 9 10049.00 612988.56 782.55 3 19191.86 9941 91 3.23 SI 52.98 3 17754.00 10991461.88 3315.34 3 14956.64 10485895.90 3238.19
class Level Information Class Levels valucs TMT 16 1 IO 11 12 19 14 15 1 6 2 3 4 5 6 7 8 9 Number of ob~wations in dab set = 48
- 111 Pairicio Elirabeih Baños Guevara

I
ANEXOS
Dependent Variable: X
SOUKC DF SumofSquarcs MeanSquare FValue Pr> F Model 15 3056436575 203762438 22.68 o.ooo1 Enwr 32 287487586 8983987 ComctedTotal 47 9343924161
R-SqualY C.V. Rwt MSE X Mean 0.914027 19.40752 2997.330 22355.5927
General h e a r Models Pmcedure Dependent Variable: X SOUKC DF TypeISS Mean Square FValue Pr > i TRAT 15 3056436575 203762438 22.68 O.ooO1
Tukey’s Studentized Range (HSD) Test for variable: X Alpha= 0.05 df= 32 MSE= 8983987 Critical Value of Studentized Range= 5.244 Minimum S-ficant Difference= 9074.8
Means with the same letter are not sixnificantly different Tukey Grouping Mean
A 35987 B A 31965 B A C 30312 B A C 29825 B A C 29622 B A C 27991 B D C 25725 B D C 25211 E D C 22267 E D F 17754 E D F 16785 E F I4957 E F 13729
F 13132 F 12384 F I0043
3 3 3 3 3 3 3 3 3 3 3 3 3 3 3 3
4 3 2 6
10 8
12 1 1 7
I5 1
16 5
14 9
13
class kvel Information Class Levels values TEMP 4 20253035 HUM 4 45556575 Number of observations in data set = 48
Dependent Variable: X
SOUKC DF SumofSquarcs MeanSquare FValuc Pr > F Model 15 3056436575 203762438 22.68 o.oO01 Error 32 287487586 8983987 ComctcdTotai 47 3343924161
R-Square C.V. Rwt MSE X Mean 0.914027 15.40752 2997.330 22355.5927
Dependent Variable: X SOUKC DF TypeISS MeanSquan FValue Pr>F TEMP 3 1359840327 453280109 50.45 O.OOO1 HUM 3 1353679960 451226653 50.23 O.ooO1 TEMPWUM 9 342916288 38101810 4.24 0.0011
Tukeps Studentized Range (HSD) Test for variable: X Alpha= 0.05 df= 32 MSE= 8983987
Critical Value of Studentized Range= 3.832 Minimum Significant Difference= 3315.3
Means with the same letter are not significantly different.
Patricia Elizabeth Bañar Guevara 112

I
TukeyGrouping Mean N TEMP A 28762 12 20 8 23453 12 25 B 23236 12 30 C 13971 12 35
Tukey's Studentizcd Range (HSD) Tcst for variable: X Alpha= 0.05 df= 32 MSE= 8983987 Critical Value of Studentizcd Range= 3.832 Minimum Significant Difference= 3315.3
Mcam with the same letter are not significantly different. TukcyGrouping Mean N HUM
A 26165 12 75 A A
R
Level of Lcvel of TEMPHUM N 20 45 3 20 55 3 20 65 3 20 75 3 25 45 3 25 55 3 25 65 3 25 75 3 30 45 3 30 55 3 30 65 3 30 75 3 35 45 3 35 55 3 35 65 3 35 75 3
25723 12 55 24299 12 65 IS235 I2 45
Mean 16785.0833 30311.6389 31965.3889 35987.1 I 1 1 15729,3444 29824.9722 22267.0556 27991.3333 12383.8611 29622.3889 25210.7778 25725.0278 10043.oooO 13 131.861 1 17754.oooO 14956.6389
SU 4 68.4 O 7 7 O 3570.05884 1870.94009 5574.72153 1733.03633 2714.65325 3605.57726 5156.99558 936.85232 3375.45013 1342.921 78 357.38990 782.55259
3152.98481 3315.33737 3238.19300
11.2. Programa 2
ARROZ ENTERO PROGRAMA DATA A: INPUT TRATS REPS TEMP HUM X; x=x/1ooooo; CARDS: 1 A 20 45 1827100000 1 8 2 0 4 5 1685016667 1 C 20 45 2008233333 ...; PROC RSREG: MOD= X-3ÉMP HLJM/UCKFlT PRESS; RUN,
RESULTADOS *Coefficients for the independent Variables
Factor Subtractdoff Dividedby TEMP 27.500000 7.500000 HUM 60.000000 15.000000
Ruporve Surface for Variable x Ruporve Mean 22478 Root MSE 6355.791608 R-Square 0.7890 Coef. of Variation 28.2750 Sum of Squared Residuals 1696635652 Pirdided Resid SS (PRESS) 2142543477
PaMcia Elizabeth Brñios Guevara 113

R e g d o n Degrees of F d o m Typ I Sum of Squares R-Square F-Ratio Pmb > F Linear 2 3856583954 0.4796 47.735 O.oo00 Quadratic 2 2354819686 0.2928 29.147 O.oo00 Cmsspmiud 1 133617763 0.0166 3.908 0.0761
Regression D e w of F&m Type I Sum of Squares R-Square F-Ratio Prob > F TotalRegríss 5 654502 I402 0.7890 91.414 o.oo00
Residual D e w of F d o m Sum of Squares Mean Square F-Ratio Prob > F Lack of Fit 10 686786876 8678688 2.176 0.0466 F’ure Eimr 32 1009848776 5157774 TotalErmr 42 1696635652 40396087
Degrees of Paramckr Standard T for H O Parameter F d o m Estimate Enor Parameter=O Prob z IT I INPERCEPT I -184151 48539 -3.794 0.0005 TEMP 1 15468 2208.1231 16 7.005 O.oo00 HUM 1 507.1 71000 1175.391924 0.431 0.6683 TEMPTEMP 1 -279.696597 36.695180 -7.622 o.Ooo0 HUMTEMP 1 -26.695094 14.678072 -1.819 0.0761 HUM’HUM I 4.060503 9.173795 0.443 0.660
Parameter INTERCEPT 30711
Parameter Estimate from coded Data
TEMP HUM
-11374 3904.744792
Parameter TEMPTEMP -15733 HUMTEMP -3003.198 125
Parameter Fstimate from Coded Data
HUM-HUM 913.613281
Sum of Factor Degrees of Fmedom Squares MeanSquare F-Ratio Pmb>F TEMP 3 5930519794 1976839931 48.936 O.oo00 HUM 3 548119370 182706457 4.523 0.0078
Canonical Analysis of Response Surface (based on coded data)
Critical value Factor coded Uncodcd TEMP -0.136 162 26.4 78 788 HUM -2.360772 24.583423
Predicted value at Stationary point 26877
Canonical Analysis of Response Surface (based on coded data)
Fjgenvedors Einenvalues TEMP HUM 1647.980203 -0.089126 0.996020
-15867 0.996020 0.089126 stationary point is a saddle point.
PROGRAMA DATA A; INPUTTRATS REPS TEMP HUM X; x=x/100000, CARDS;
PROC RSREG; MODEL X=TtMP HUM/LACICFIT PRESS, RUN;
... ,
Patricia Elizabeth Bañas Guevara 114

1
RESULTADOS Coding Coefficients for the Independent Variables
Factor Subtracted off Divided by TEMP 27.500000 7.500000 HUM 60.000000 15.000000
Rwponse Surface for Variable X Rcsponse Mean 22356 Root MSE 4421.484586 R-SqUalt 0.7545 Coef. of Variation 19.7780 Sum of S q u a d Residuals 821080089 Predicted Resid SS (PRESS) 10225601 51
Regression D e w of Fncdom Type 1 Sum of Squaw R-Square F-Ratio Pmb>F linear 2 2030702795 0.6073 51.937 0.0000 Quadratic 2 585385980 0.1152 9.857 0.0003 CmsSprodUd I 106755297 0.0319 5.461 0.0243
Regression Degrees of Freedom Type I Sum of Squares R-Square F-Ratio Pmb>F TotalRegrrss 5 2522844071 0.7545 25.810 0.0000
Residual Degrees of Fnedom Sum of Squares Mean Square F-Ratio Prob > F Lack of Fit 10 533592504 53359250 5.939 0.0001 Pure Error 32 287487586 8983987 Total Error 42 821080089 19549526
Parameter Degress of Freedom Parameter Estimate Standard Error INlZRCEpT I -135860 33767 TEMP 1 2715.124197 1536.107998 HUM 1 4216.344039 817.675845 TEMPTEMP 1 59.550037 25.527453 HUMTEMP 1 -23.861299 10.21098 1 HUM'HUM 1 -26.554177 6.381863
Parameter INTERCEPT 26911 TEMP -6688,568124 HUM 604.856042
Parameter TEMPTEMP -2224.692969 HUMTEMP -2684.996125 HUM'HUM -5974.689843
Factor Degires of Freedom Sum of Squares Mean Squan F-Ratio TEMP 8 1346666764 448888921 22.962 HUM 3 1282932604 427644201 21.875
Parameter Ertimate from Coded Data
Parameter Estimate from coded Data
Canonical Analysis of Response Surface (based on coded data)
Critical Value Factor coded Wncoded TEMP -2.066296 12.002781 HUM 0.939258 75.998567
Predicted value at stationary paint 36436
Canonical Analysis of Response Surface (bascd on wdcd data)
Eigenvectors Eigenvalues TEMP HUM
Stationary point is a maximum.
-1793.804423 0.952158 -0.508667 -6405.578389 0.305667 0.952138
T for HO Parameter=@ Pmb > IT I -4.023 0.0002 1.768 0.0844 5.156 O.oo00 -1.549 0.1288 -2.337 0.0243 -4.161 0.0002
Pmb > F O.oo00 O.oo00
Patricia Elizabeth Bañar Guevara 115

ANEXOS - -
11.3. Programa 3
data 4 input dias esp; a*
pmc soit$y DIAS, pmc nün but=5 efonnat 84 mcthod=manpnrdt, pnimsA= 12.5to 13.4byO.I
... ,
B = O.ooO8 to 0.0017 by O.ooO1 C = 1.39 to 1.48 by 0.01;
"Función Logística; mods1 ESP = A / (I+Exp(B-C*DW));
title 'REiIDUALES DE LA CURVA qRlsrADa'; tine2 'DW';
Nn ;
RESULTADOS Non-linear Least Squares Grid Search Dependent Variable ESP
A B C Sum of Squares 1.250000E+01 1.700000E-03 1.480000E+00 3.321401E+03 1.250000E+OI 1.600000E-03 1.480000E+00 3.321442E+03 1.250000E+01 1.500000E-03 1.480000E+00 3.321482E+03 1.25ooOOE+Ol 1.400000E-03 1.480000E+00 3.321523E+03 1.250000E+01 1.500000E-03 1.480000E+00 3.321564E+03
Non-Linear Last Squam Iterative Phase Dependent Variable ESP Method: Maquardt
Iter A B C sum of souam O I 2 3 4 5 6 7 8 9 IO 11 12 13
1.25ooOOE+01 7.64 194 1E+W 5.260966E-01 2.245193E-O1 1.419932E-01 I.917469E-O1 2.41 1454E-01 6.928277E-O1 7.787IOOE+00 6.558214E+W 6.852253€+00 7.918396E+00 9.032449E+W 1.028053E+01
1.700000E-03 -I.O56240E+00 -7.577566E-O1 - 1.826620E-0 I 1.980235E-01 2.21208IE-O1 2.452028E-0 I 8.493239E-01 1.92987 lE+O 1 1.935832E+01 1.866007E+01 18034 18E+O 1 1.77441 IE+01 1.489384E+01
I . ~ ~ C O O O E + C ~ 3.92 14o i~+o3 -4.787885E+00 2 522262E+03 -3.984775E100 2.5 152 I2E-O3 - 1.750017E+00 2.3 13 158E-03 2.594630€+00 2.249005E-03 2.535422E100 2.226258Es03 2.2€8838E+00 2.205854E+03 1.796878E-01 2.132518E+03 l.O15154E+Ol 9.2438741+02 8.681089E+W 7.577410E+02 6.106515E+W 5.33541 IE+02 4.963659Es00 3.2539541+02 4.195439E-00 1.5781 12E*02 2.826376E*00 1.562352E-02
-116 Patricia Elizabeth Baños Guevma

I
ANEXOS
14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29
32 33 34 35 36 37 38
I . 1601 59~+01 I . 161012E*@l 1.16017SE+01 1.164056E+O 1 I . 169526E-01 1.1 75435E-01 1.181 754E+01 1.188460E+01 l.l95526E*Ol 1.20291 OE+01 12 10554E+0 1 1.2 18980E*01 1.226287E+OI 1.234 156E+OI 1.241854E+01 1.249237E-O1 1.256169E*01 1.262533E-01 1.268239E+O I 1.273237E*01 1.27751 7 ~ ~ 0 1 1.281 1058+01 1.278673€+0 I 1.277349E-01 1.277480E+01
1.379728E~01 2.5883398+00 l.SZ670SEt01 2.5360358~00 1.270761E~01 2.443929E*00 1.2129S6E+01 2.332889E+00 1.157530E-01 2.226108E+00 1.104548E+01 2.123675E~00 1.054019E+01 2.025805E-00 1.006000E-01 1.9327 14E+00 9.60576iE+00 1.844641E+00 9.178448E+00 1.761829E*00 8.779054E-00 1.684514E+00 8.408525E+0(i 1.612910E*00 8.067639E+00 1.547191E*00 7.75692 1 E+OO 1.487478E-00 7.4 7656ZE-00 1.4338 16E+00 7.226358Es00 1.386168Es00 7.00566iE+00 1.344400E~00 6.813393E+00 l.SO8277E~W 6.647997E-00 1.277469Es00 6.507550E+00 1.251558E+00 6.389808E+00 1.230060€+00 6.292321E+W 1.2 12454E-00 6.259074E-00 1.215236E+00 6.238918Es00 1.214235E+00 6.2345 1 1 E+OO 1.2 14782E -00
9.9531528+01 8.109186E+OI 7.346224E+OI 6.894968E+01 6.453238E+01 6,OSS700E+OI 5.6365 12E+01 5.263060E+OI 4.915355E+01 4.595683E+01 4.506277E+01 4.0490 12E+O 1 8.825 144E+O 1 8.695 1 I2E+01 3.4784 13E+01 3.3535728+01 3.258199E+OI 8.389124E+01 3.142598E+01 3.1 145498+01 3.10085SE+01 3.09761 5E+01 3.052983E+01 3.052364E+01 3.051756E+OI
39 1.277487E+OI 6.234450E+00 1.21479SE+00 3.051755E+@I 40 1.277493E+01 6.234389E+00 1.214804E+00 8.051754E+01 41 1.2774948+01 6.234382E+00 1.2I4805E+00 3.051754E+01
N O R Convergence criterion met.
Non-iincar Lurrt Squares Summary Statistics Dependent Variable ESP Source DF SumofSquam McanSquarc Regrwsion 3 2284.7019799 761.5673266 Residual 45 30.51 75430 0.678 I676 Unuirrcded Total 48 2315.2195229 (Conrctcd Total) 47 1149.9771250
Parameter Lrtimatc Asymptotic Asymptotic 95 96 Std. Ermr Confidence Interval
Lower Upper A 1.277494E+01 5.8184767E-O1 1.1603041E+01 1.3946836E+01 B 6.234382E+00 2.97536318-01 5.6351155E+00 6.8336495E+00 C 1.2 14805E+00 1.2068 1 78E-0 1 9.7 I 740 12E-01 1.4578697E+W
Asymptotic Correlation Matrix Corr A B C
A 1 -0.798674564 -0.869778828 B -0.798674564 1 0.9837033608 C -0.869778828 0.9837033608 1
Patricia Elizabeth Bañm Guevara 117
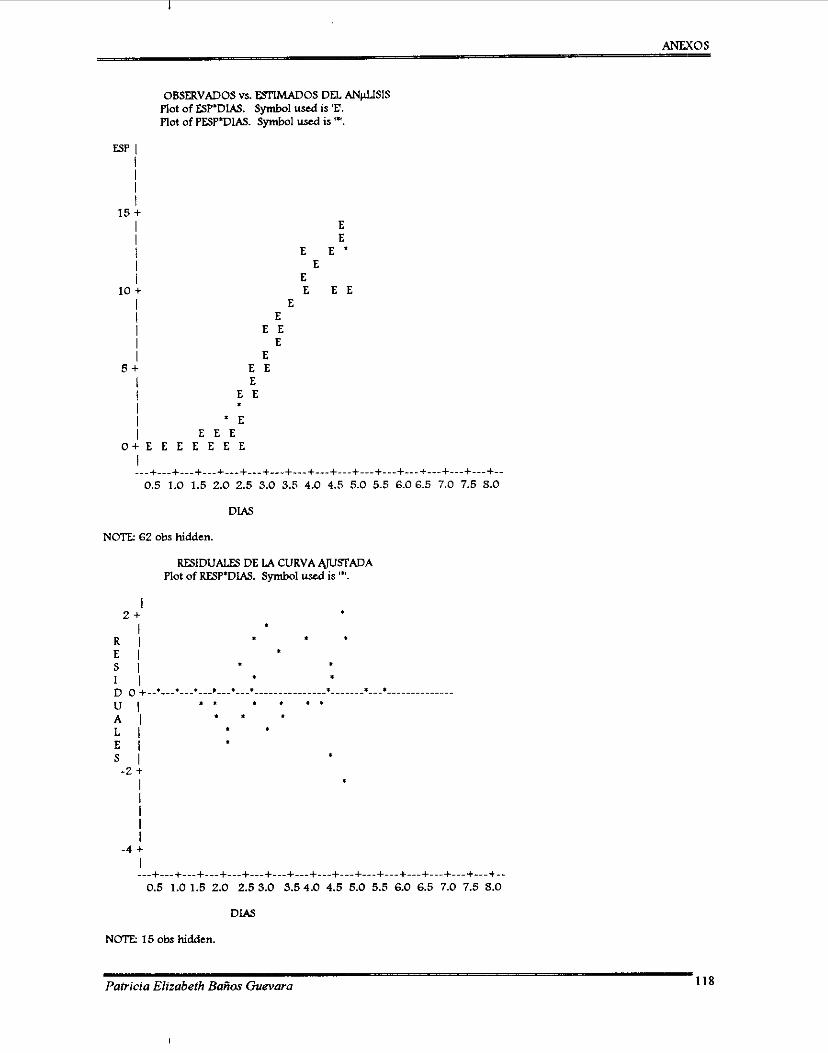
I
OBSERVADOS vs. -0s DEL ANIIUSIS not of ESP'DIAS. Plot of PESP'DIAS. Symbol used is I*.
Symbol used is 'F.
ESP I I I I I
I I I I I
15 i
10 +
E E
E E ' E
E E E E
* E
O + E E E E E E E
I I I
I
E E E
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5 5.0 5.5 6.0 6.5 7.0 7.5 8.0
DIAS
NOTE: 62 obs hidden.
RESIDUALFS D E LA CURVA qniSrADA Plot of RESP'DIAS. Symbol U d is I*'.
-2 + I
-4 + I ......................................................................
0.5 1.0 1.5 2.0 2.5 3.0 3.54.0 4.5 5.0 5.5 6.0 6.5 7.0 7.5 8.0
DIAS
NOTE: 15 o h hidden.
118 Paíricia Elizabeth Baños Guevara

ARROZ RESIDUAL
pNK plot data=GW; plot wp*dias=Y PFSP*DLG='" i overlay vpos=Z5; title 'OBSERVADOS vs. ETiivWJOS DEL ANALEIS'; title2 'DIAS',
dot mu*dias='" / mf=0 vfK1s=25: p m plot data=cRAF;
iitle ~ ~ I U A L E S DE LA CURVA ~ÚÚSTADA'; title2 'DIAS',
run;
RESULTADOS ANALlSlS REGRLSION NO LINWL / DIAS
Non-Linear Lcast Squares Grid Search Deucndent Variable ESP A B c Sum of Squares
6.900000€+00 1.500000E-03 8.800000E-01 1.1152896+03 6.900000E+00 1.400000E-03 8.800000E-01 1.1 15310E+03 6.900000E+W 1.300000E-03 8.800000E-01 1.1 15331E+03 6.900000Es00 1.200000E-03 8.800000E-01 1.115352Es03 6.900000E-00 1.100000E-03 8.800000E-01 1.115373E-O3
Non-tincar Least Squarcs DUD Initialization Dependent Variable ESP DUD A B C Sum of Squares -4 6.900000E+00 1.500000E-03 8.800000E-01 1.115289E+03 -3 7.590000E-00 1.5JXOGüE-03 2 6.900000E+00 1.650000E-03
- I 6.900000E+00 1.500000E-03 O 6.900000E-00 1.650000E-03 1 6.697975E-00 -1.506679E-01 2 6.626513€+00 -8.478652E-02 3 6.61921íE+00 -8.720673E-02 4 6.620719E+00 -8.7425718-02 5 6.806336€+00 2.005663E-01 6 7.993957E-00 2.600143E+00 7 8.097099E+00 4.221086E+W 8 7.664398E-00 5.219678E+00
8.800000E-O1 8800000E-01 9.680000E-01 8.800000E-O1 9.6284351-01 9.8348258-02 8.1 12756E-O2 8.6581 17E-O2 4.1259458-02 2.841792E-01 6.521936E-01 7.830271E-01
1.39s I65E-03 1.1 15257E+OS 1.146764E+03 1.1 15257E+03 7.563 175E-02 4.96531 IE+OZ 4.962744Es02 4.961 668E-02 4.81 3460Es02 2.630457E+02 4.989492M01 4.772 I I6E+O I
Poíricio Elizobeth Boños Guevaro 119
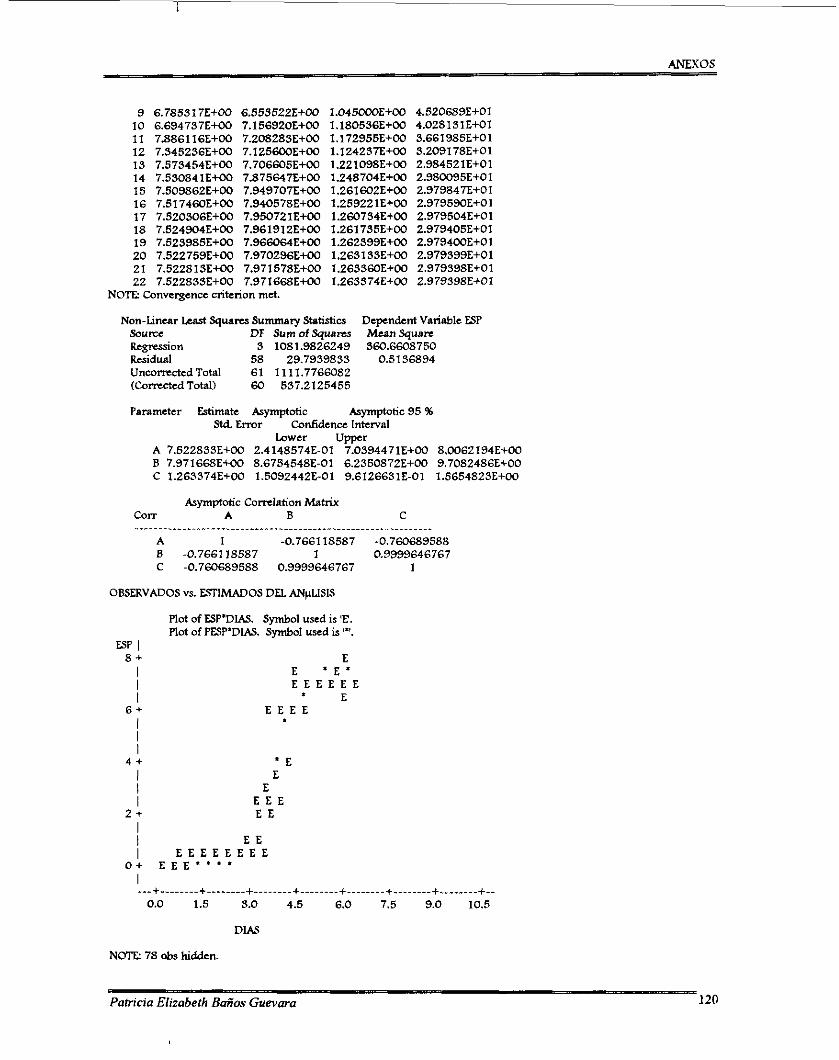
ANEXOS
9 6.785317E+OO IO 6.694737E+00 11 7.886116E+00 12 7.345236E+00 13 7.573454E+00 14 7.5$0841E+00 I5 7.509862E+00 16 7.517460E+00 17 7.520306E+00 18 7.524904E+00
6.55352ZE+00 7.156920E+00 7.20828SE+00 7.125600E+W 7.706605E+00 7.875647E+00 7.949707E+W 7.940578E+00 7.950721E+00 7.96 191 ZE+W
I.O45OOOE+00 1.180536E+00 1.172955E+W 1.124237E+00 1.22 1098E+W 1.248704E+W 1.261 602E+00 I.259221E+00 1.260794E+00 1.261735E+00
4.520689E+Ol 4.028131E+01 3.661985E+01 3.2091 78E+01 2.984521E+Ol 2.980095E+01 2.979847E+Ol 2.979540E+01 2.979504E+O 1 2.979405E+OI
~
19 7.523985E100 7.966064E-00 1.262399E+00 2.979400E*01 20 7.522759E-00 7.970296E+W 1.26313SE-00 2.979399E-01 21 7.522513E+00 7.971578E-00 1.263360E+00 2.9793981+01 22 7.52283SE-00 7.971668E+OO 1.263574E+00 2.979398E4l
N O n Cnnvemence cntenon met.
Non-Linear Least Squarc~ Sununary Statistiu Dependent Variable U P SOUrCC Di SumofSquam MeanSquarc Regrrssion 3 1081.9826249 360.6608750 Residual 58 29.7939835 0.5136894 Uncorrected Total 61 11 11.7766082 (ComctcdTotall 60 537.2125455
Parameter Estimate Asymptohc ASY7nptOhC 95 % Std Errur Confidence lntewal
Lower Upper A 7.52?833E*W 2.4148574E-01 7.0394471E+00 8.00621Y4E+00 R 7.971668E+00 8.6754548E-01 6.2350872E-00 9.7082486E-00 C 1.263374E-00 1.5092442E-01 9.6126631E-01 1.5654823Ec00
Asymptotic Cnmlahon Matnx Corr A R C
A 1 -0.7661 18587 -0.760689588 5 -0.7661 18587 I 0.9999646767 C -0.760689588 0,9999646767 I
OBSERVADOS vs. W W O S DEL ANuUSlS
I 8 +
I I I
I I I
I I I
I I I
I
6 1
4 +
2 +
O +
Plot of ESP'DIAS. Symbol used is 'E. Plot of PESP'DIAS. Symbol used is '*l.
E E ' E ' E E E E E E
E E E E E
' E E
E E E E E E
E E E E E E E E E E
E E E * * * '
Patricia Elizabeth Baños Guevara 120
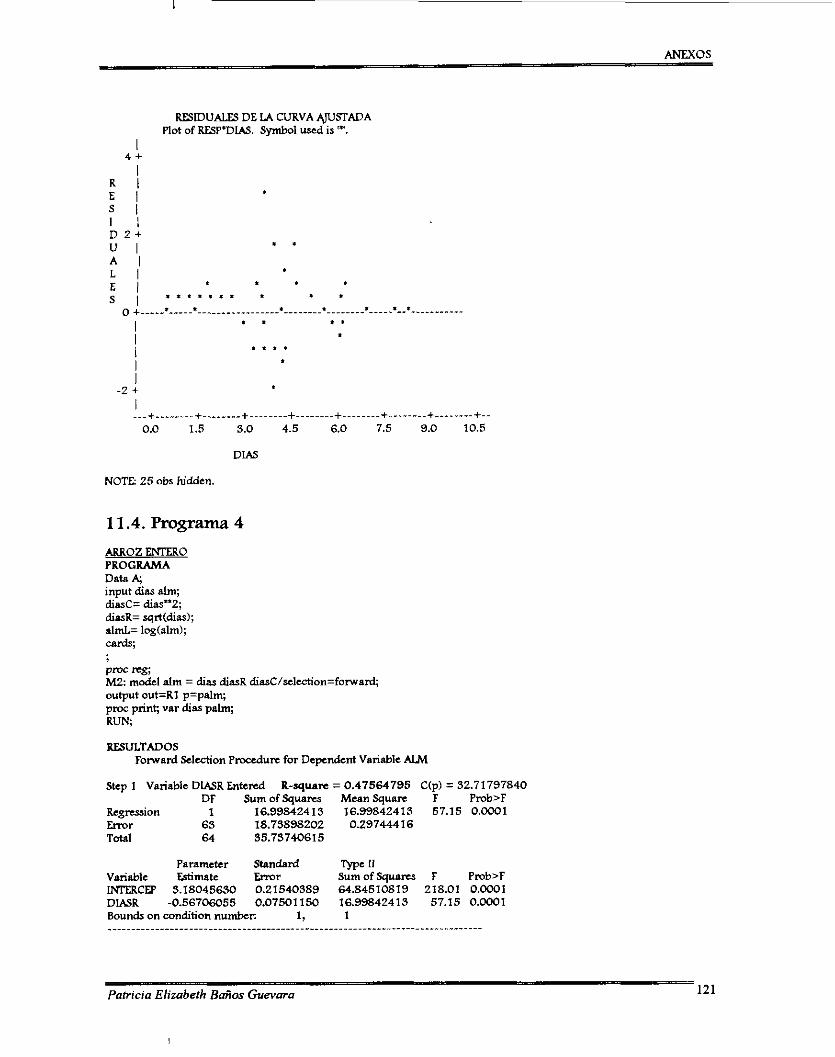
RESIDUALES DE LA CURVA qlVSTADA Plot of RESPz)iAS. Symbol used is '=.
I
I 4 +
R I E l S I 1 1 D 2 + U I * .
* . I
.*.. I I I
I ...+ ______.. +.- _ _ _ _ _ _ + ....____ + .___.... + .._._._. + .... --..+ .--..--. +--
0.0 1.5 5.0 4.5 6.0 7.5 9.0 10.5
DlAS
NOTE: 25 obs hidden.
11.4. Programa 4
ARROZ ENTERO PROGRAMA Data A; input dias aim; diasc= dias-2; dinsR= sqrt(dias); almL= log(alm); cards;
P W rrg; MZ: model aim = dias diasR diasC/selection=fomard; output out=RI p-palm; pmc print; var dias palm; RUN;
RESULTADOS Fomani Selection Procedure for Dependent Variable ALM
Step I Vaiiable D M R E n t e d R-qua= = 0.47564795 C(p> = 32.71797840 DF SumofSauam MeanSauare F Prob>F
Regresion 1 16.938424 13 16.99842413 57.1 5 O.OOO1 Ermr 63 18.73898202 0.2974441 6 Total 64 35.73740615
Parameter Standard Type 11 Variable W m ate Error SumofSquam F Prob>F WIZRCEF' 3.18045630 0.21540389 64.84510819 218.01 O.OOO1 DIASR -0.56706055 0.07501150 16.99842413 57.15 O.OOO1 Bounds on wndition number. 1 , 1
- Patricia Elizabeth Bañas Gumma -121

Step 2 Variable DUS Entend R-square = 0.60876818 C(p) = 10.92526483
DF SumofSquaw MeanSquare F Pmb>F Rcgrwsion 2 21.75579555 10.87789778 48.24 O.ooO1 Error 62 13.981 61060 0.22550985 Total 64 85.73740615
Parameter Standard Tnx I1 ,. Vanable h m a t e E r m r SumofSquaru f Pmb>F I"ECEP 4.95597995 0.42966539 30.00294224 133.04 0.0001 DIAS 0.33005817 0.07186041 4.75737142 21.10 0.0001 DUSR -2.21661598 0.36503273 8.31539390 36.87 0.0001
Bounds on condition number: 31.23546, 124.9418 ..............................................................................
Step 3 variable DUSC Entend R-rqure = 0.65870503 C(p) = 4.00000000 DF SumofSauam MeanSauare F Prob>F
Regression 3 23.54040914 7.84686305 39.24 0.0001 Error 61 12.19699702 0.19995077 TOW 64 35.7374061 5
Parameter Standard Type I1 Variable %mate Enwr SumofSquam F Pmb>F INERCEP 6.81045005 0.74094933 16.89264641 84.48 O.OOO1 DIAS I.238I4314 0.3II40025 3.16101746 15.81 O.ooO2 DIASR -4.86013975 0.94927180 5.24130502 26.21 0.0001 DUSC -0.02399918 0.00803315 1.78461358 8.93 0.0040
Bounds on condition number. 661.5288, 3081.904
No other variable met the 0.5000 significance level for entty into the model
Summary of Forward Selection Procedure for Dependent Variable ALM
Step Entered In R**2 R*'2 C(p> F Prob>F 1 DUSR 1 0.4756 0.4756 32.7180 57.1483 0.0001 2 DIAS 2 0.1331 0.6088 10.9253 21.0961 O.ooO1 3 DUSC 3 0.0499 0.6587 4.oooO 8.9253 0.0040
Variable Number Pattial Model
ARROZ RESIDUAL
PROGRAMA 4
Data A; input dias alm; diasc= dias"2; diasR= 4rt(dias); almL= log(a1rn); cards;
P W w; M2 model alm = dias diasR diasC/selection=forwani; output out=RI p=pnlm; proc p ~ $ var dias pdm; RUN,
... ,
RESULTADOS
Step 1 Variable DUS Entered R-squa~t = 0.38880121 C(p) = 1.58574206 DF SumofSquarcs MeanSquarc F Pmb>F
Re-ion 1 3.63509694 3.69509694 39.44 O.ooO1 Error 62 5.71440306 0.09216779 Total 63 9.3495oooo
Forward Selection Procedure for Dependent Variable ALM
c
Pahicia Elizabeth Baños Guevara 122

ANEXOS - Parameter Standard Type11
Variable Estimate ErmI SumofSquam F Pmb>F WíERCEP 1.79714718 0.07771242 49.29077036 534.79 O.ooO1 DlAS -0.05162390 0.00822020 9.69609694 39.44 O.ooO1 Bounds on condition number: 1, 1
Step 2 Variable DlASC Entered R-square = 0.99394842 C(p) = 3.06709853 DF SumofSquam MeanSquare F Pmb>F
Regresion 2 9.68322071 1.84161036 19.83 0.0001 E m r 61 5.66627929 0.09288982 Total 63 9.3495oooo
Parameter Standard Type 11 Variable Estimate Error SumofSquam F Pmb>F INERCEP 1.72979839 0.12182681 18.72722297 201.61 O.ooO1 DlAS -0.02785374 0.09403997 0.06219521 0.67 0.4164 DUSC -0.00144062 0.00200149 0.04812377 0.52 0.4744 Bounds on condition number: 17,01471, 68.05882
Step 3 Variable DlASR Entered R-square = 0.40453868 C(p) = 4.00000000
Regression 3 3.78223439 1.26074480 13.59 O.OOO1 Error 60 5.56726561 0.09278776 Total 63 9.3495oooo
DF SumofSquam MeanSquare F Prob>i
Parameter Standard Type II Variable Emmate Ermr SumofSquarcs F Pmb>F WPERCEP 2.23587997 0.50481625 1.82020795 19.62 O.ooO1 DlAS 0.18876075 0.21243556 0.07325878 0.79 0.3778 DlASR -0.66836369 0.64700940 0.09901368 1.07 0.3057 DlASC -0.00672215 0.00549018 0.13910186 1.50 0.2256 Sounds on condition number: 663.404, 3090.051
No dher variable met the 0.5000 significance level for enhy into the model
Summary of Forward Scledion Procedure for Dependent Vanable AiM Variable Number Partial Model
Step Entcrcd in R**2 R"2 C(p) F Pmb i i 1 DlAS 1 0.3888 0.3888 1.5857 39.4400 0.0001 2 DlASC 2 0.0051 0.3939 3.0671 0.5181 0.4744 3 DlASR 3 0.0106 0.4045 4.oooO 1.0671 0.3057
- ._=
Pahicia Elizabeth Baños Guevara 123





























