Embed Size (px)
Citation preview

Tumor targeting of the IL-15 superagonist RLI by an anti-GD2antibody strongly enhances its antitumor potency
Marie Vincent1, Anne Bessard1, Denis Cochonneau2, G�eraldine Teppaz1, V�eronique Sol�e1, Mike Maillasson1,
St�ephane Birkl�e2, Laure Garrigue-Antar1, Agnes Qu�em�ener1 and Yannick Jacques1*
1 Centre de Recherche en Canc�erologie Nantes-Angers, Inserm, U892, CNRS, U6299, �equipe Cytokines et R�ecepteurs en Immuno-H�emato-Canc�erologie, Uni-
versit�e de Nantes, Nantes, France2 Centre de Recherche en Canc�erologie Nantes-Angers, Inserm, U892, CNRS, U6299, �equipe Radiobiologie et Ciblage de l’Endoth�elium, Universit�e de
Nantes, Nantes, France
Immunocytokines (ICKs) targeting cytokines to the tumor environment using antibodies directed against a tumor-associated
antigen often have a higher therapeutic index than the corresponding unconjugated cytokines. Various ICKs displaying signifi-
cant antitumoral effects in several murine tumor models have already been developed, and some of them, in particular inter-
leukin (IL)-2-based ICKs, are in Phase II clinical trials. Although sharing common biological activities with IL-2 in vitro, IL-15 is
now considered as having a better potential in antitumor immunotherapeutical strategies and has been shown to be less toxic
than IL-2 in preclinical studies. We previously developed the fusion protein RLI, linking a soluble form of human IL-15Ra-
sushi1 domain to human IL-15. RLI showed better biological activities than IL-15 in vitro as well as higher antitumoral effects
in vivo in murine and human cancer models. Here, we investigated, in the context of an ICK, the effect of associating RLI with
an antibody targeting the GD2 ganglioside, a validated tumoral target expressed on many neurectodermal tumors. Anti-GD2-
RLI fully retained the cytokine potential of RLI and the antibody effector functions (antibody-dependent cellular cytotoxicity
and complement-dependent cytotoxicity). It displayed strong antitumor activities in two syngeneic cancer models in immuno-
competent mice (subcutaneous EL4 and metastatic NXS2). Its therapeutic potency was higher than those of RLI and anti-GD2
alone or in combination. We suggest that this is related to its bifunctional (cytokine and antibody) nature.
Many cytokines have revealed antitumoral potencies, in lightwith the important role of the immune system in the preven-tion and treatment of cancer.1 Because of their rapid bloodclearance and their lack of tumor specificity,2 systemic high-dose cytokines administration is required to reach animmune response in the tumor microenvironment, and sucha regimen often led to severe toxic side effects limiting theirclinical use.3 Targeting cytokines to the tumor microenviron-ment using an antibody directed against a tumor-associatedantigen could lead to a more potent therapeutic benefit withacceptable side effects.4 Immunocytokines (ICKs) specific toantigens expressed on tumor cells are particularly attractivein this context. A large number of ICKs has been developedin preclinical studies, but only a few of them have enteredclinical trials (for reviews, see Refs. 5 and 6). Interleukin(IL)-2 fusion proteins are among the most advanced ICKs inclinical development, owing to the clinical approval of IL-2in the treatment of metastatic renal cancer and malignantmelanoma. However, they elicit adverse events resemblingthe ones observed with recombinant IL-2.7,8 Based on thepreclinical studies, IL-15, a cytokine structurally related toIL-2, does not appear to induce such side effects9 and exhib-its a better safety profile than IL-2, while showing similar orimproved efficacy.10 IL-15 is currently tested in several clini-cal trials for the therapy of cancer.11 It belongs to the four-a-helix-bundle family and generates its actions through the
Key words: interleukin-15, immunocytokine, GD2 disialoganglio-
side, cancer therapy
Abbreviations: ADCC: antibody-dependent cellular cytotoxicity;
CDC: complement-dependent cytotoxicity; ICK: immunocytokine;
IL: interleukin; mAb: monoclonal antibody; RTX: rituximab
The costs of publication of this article were defrayed in part by the
payment of page charges. This article must therefore hereby be
marked advertisement in accordance with 18 U.S.C. Section 1734
solely to indicate this fact.
*A.Q. and Y.J. contributed equally to this work.
Grant sponsors: INSERM, CNRS, Institut National du Cancer and
Canc�eropole Grand Ouest (MabImpact), OSEO Innovation and
R�egion Pays de Loire (CIMATH2), Ministere de l’Enseignement
Sup�erieur et de la Recherche, Association pour la Recherche sur le
Cancer, Ligue contre le Cancer
DOI: 10.1002/ijc.28059
History: Received 31 Oct 2012; Accepted 8 Jan 2013; Online 25 Jan
2013
Correspondence to: Agnes Qu�em�ener, Centre de Recherche en
Canc�erologie Nantes-Angers, Inserm, U892, CNRS, U6299, Groupe
de Recherche Cytokines et R�ecepteurs en Immuno-H�emato-
Canc�erologie, Institut de Recherche en Sant�e de l’Universit�e de
Nantes, 8 Quai Moncousu, BP 70721, F-44007 Nantes Cedex 1,
France, Tel.: 133-22-808-0306, Fax: 133-22-808-0204,
E-mail: [email protected]
Can
cerTherapy
Int. J. Cancer: 133, 757–766 (2013) VC 2013 UICC
International Journal of Cancer
IJC

binding to a receptor that shares the IL-2/IL-15Rb and com-mon g-chains as transducing components with the IL-2 re-ceptor.12 In addition, IL-2 and IL-15 can use private a-chainsconferring specificity.13,14 Despite functional redundanciesinitially demonstrated in vitro, both cytokines display distinctand often competing roles in many adaptive immuneresponses. Indeed, unlike IL-2, IL-15 does not inhibit CD81
effector T-cell responses through activation-induced celldeath and does not promote regulatory T cells that canattenuate antitumor immune responses.15–17 However, IL-15is crucial for the development of natural killer (NK) cells, thesurvival of memory T cells and is also required for the initia-tion of T-cell activation.18–20 It therefore plays a major rolein tumor immunosurveillance. The dominant mode of actionof IL-15 in vivo is an original mechanism called trans-presen-tation, in which IL-15Ra, expressed at the surface of IL-15producer cells (dendritic cells, macrophages and epithelialcells), presents IL-15 in trans to responder cells (NK ormemory CD81 T cells) bearing the IL-15Rb/g receptor.21–23
This trans-activation may also be mediated by a soluble IL-15Ra/IL-15 complex resulting from the proteolytical cleavageof membrane-anchored IL-15Ra by metalloproteases.24 Sev-eral in vitro and in vivo studies have revealed the higherstimulatory effect of such soluble IL-15Ra/IL-15 complexesover IL-15 alone.25–27 Binding of soluble IL-15Ra-Fc to IL-15was also shown to extend the half-life of IL-15 in vivo.27 Torecapitulate the complex of IL-15 with its a-receptor chainon a single molecule, we have previously engineered a fusionprotein (RLI) that consists of the NH2-terminal (amino acids1–77, sushi1) domain of human IL-15Ra linked via a 20-amino acid linker to human IL-15.28 This fusion protein wasshown to exert higher biological activities than IL-15 or eventhe noncovalent association of IL-15 with the soluble IL-15Ra in vitro to drive cell proliferation through the IL-15Rb/g receptor29 and in vivo to promote mobilization and expan-sion of NK cells.30 RLI had an increased half-life in vivo andshowed strong antitumoral effects in systemic B16 melanomamouse model and human HCT-116 colorectal cancer.31
The rational of our study was to evaluate the therapeuticalinterest of targeting the RLI molecule at the tumor site in theframe of an ICK. As a first tumor target, we have chosen theGD2 antigen, a sialic acid-bearing glycosphingolipidexpressed by many human neurectodermal tumors, such asmelanoma, glioma, neuroblastoma and small cell lung carci-noma,32 and in contrast, being only expressed at very low
levels in the peripheral nervous system33 and the cerebel-lum.34 GD2 is also involved in several biological functions35
such as cell recognition, cell matrix attachment and cellgrowth and differentiation, suggesting that tumor-associatedGD2 may play a significant role in the tumorigenic pheno-types of these cells. GD2 on tumor cell surfaces has beenshown to be a relevant target antigen for antibody-dependentcellular cytotoxicity (ADCC)36 and complement-dependentcytotoxicity (CDC).37 It has recently been ranked 12th amongall promising cancer antigens for cancer treatment or preven-tion.38 In our study, we have used the mouse/human chi-meric anti-GD2 antibody (c.60C3)39,40 to engineer an RLI-based ICK. This anti-GD2-RLI ICK was expressed in mam-malian cells, purified to homogeneity and evaluated for its bi-ological activities in vitro. It was further shown to displaystrong antitumoral activities in vivo against both primary(subcutaneously) and metastatic tumors in syngeneic immu-nocompetent murine models of cancer.
Material and MethodsReagents
Recombinant human IL-15 was purchased from Peprotech(Rocky Hill, NJ) and recombinant human IL-2 from Chiron(Suresnes, France). Murine IL-3, mouse anti-human IL-15monoclonal antibody (mAb; MAB247) and its biotinylatedform (BAM247) were obtained from R&D Systems (Lille,France). RLI was produced as described previously.28 Controlhuman isotype IgG was purchased from Santa-Cruz Biotech-nology (Heidelberg, Germany), peroxidase-conjugated poly-clonal goat anti-human IgG (H1L; 109-036-003) fromJackson Immuno Research (Suffolk, UK), anti-human IgG(H1L; UP892370) from Interchim (Montlucon, France) andmonoclonal mouse anti-human IL-15 antibody (B-E29) fromGen-Probe (Manchester, UK).
Cell lines
All cell lines were grown at 37�C under a humidified 5%CO2 atmosphere. Kit225 T-lymphoma human cells41 werecultured in RPMI-1640 medium (Life Technologies, Saint-Aubin, France) containing 6% heat-inactivated fetal calf se-rum (FCS; PAA, Austria), 2 mM glutamine and 5 ng/mlhuman IL-2. The EL4 murine T-lymphoma cell line (ATCCTIB-39) was cultured in RPMI-1640 medium containing 10%FCS and 2 mM glutamine. 32Db lymphoblast murine cells42
What’s new?
Many antibody-cytokine fusion proteins known as immunocytokines (ICKs) have been developed to deliver cytokines with anti-
tumoral potencies to the tumor microenvironment using antibodies directed against a tumor-associated antigen. Several ICKs
using notably interleukin 2 (IL2) present encouraging results in phase-II clinical trials. IL15, which belongs to the IL2 family,
and the superagonist RLI, which the authors previously developed, present many advantages over IL2 as immunological adju-
vant, such as chemotherapy potentiation and toxicity reduction. Here they show that a RLI-based ICK targeting the GD2 tumor
antigen has a higher therapeutic potency in vivo than RLI and antiGD2 alone or in combination.
Can
cerTherapy
758 RLI-based immunocytokine targeting GD2
Int. J. Cancer: 133, 757–766 (2013) VC 2013 UICC

were cultured in the same medium, supplemented with 0.4ng/ml murine IL-3, 10 mM 2-mercaptoethanol and 250 mg/mlgeneticin. Human neuroblastoma IMR32 cells (ATCC CCL-127) were cultured in DMEM medium (Life Technologies,Saint-Aubin, France) with 10% FCS and murine neuroblas-toma NXS2 cells, kindly provided by Dr. H.N. Lode (Univer-sit€atsklinikum Greifswald, Germany), and were cultured inthe same medium supplemented with 100 Units/ml penicillinand 100 mg/ml streptomycin. CD16-transduced NK92 cellsused as effector cells in the ADCC assays were kindly providedby Dr. B. Cl�emenceau (CRCNA, Nantes, France). They werecultured in RPMI-1640 medium containing 10% human serum,2 mM glutamine and 5 ng/ml human IL-2.
ICK plasmids construction
The cDNAs encoding the heavy and the light chain of themouse/human chimeric anti-GD2 antibody were generated asdescribed previously.40 The light chain and the heavy chainsequences were respectively cloned in pcDNA3 andpcDNA3.1 Hygro (Life Technologies, Saint-Aubin, France).The sequence of the human IL-15 superagonist RLI28 wasfused in frame at the 30 end of the anti-GD2 heavy chain. Anirrelevant ICK (anti-CD20-RLI) was also constructed withthe heavy and light chains of the rituximab (RTX) antibodyas templates (unpublished results).
ICK expression and purification
Expression plasmids pcDNA3 G418/anti-GD2-L andpcDNA3.1 HYGRO/anti-GD2-H-RLI were transfected intoCHO cells using a Lipofectamine 2000 Transfection Reagent kit(Life Technologies, Saint-Aubin, France). Cells were selected inthe presence of 0.5 mg/ml G418 and Hygromycin (Life Tech-nologies, Saint-Aubin, France). Double-resistant clones wereisolated by limiting dilution, and the supernatants of resistanttransfectants were screened by enzyme-linked immunosorbentassay (ELISA). ICK production was further conducted in AIM-V medium (Life Technologies, Saint-Aubin, France). ICK wereaffinity purified from culture supernatants by using a HiTrapProtein A HP column (GE Healthcare, Nantes, France). Theeluted ICKs were dialyzed against PBS for buffer exchange andsterile filtered (0.22 mm). ICK concentrations were determinedby measuring the absorbance at 280 nm, and their purity wasanalyzed by SDS-PAGE and Western blot.
SDS-PAGE and Western blotting analysis
Purified proteins were denatured for 5 min at 95�C in so-dium dodecyl sulfate loading buffer with (reducing condi-tions) or without (nonreducing [NR] conditions) 2-mercaptoethanol and analyzed on 4–12% Bis-Tris Gels (LifeTechnologies, Saint-Aubin, France). Proteins were revealedwith Coomassie blue reagent (Thermo Scientific, Illkirch,France), and for Western blot analysis, proteins were trans-ferred to Immobilon-P Polyvinylidene difluoride membrane(Millipore, Bedford, MA). Membrane was saturated with 5%dry milk powder, 0.05% Tween-20 in PBS, immunoblotted
with an anti-IL-15 mAb (B-E29) and washed in PBS 0.05%Tween-20 before being incubated with a secondary anti-mouse/anti-rabbit antibody conjugated with horseradish per-oxidase. Labeled proteins were detected using BM Chemilu-minescence Blotting Substrate (Roche, Meylan, France).
ELISA
Anti-GD2-RLI plasma levels were evaluated with two ELISAs.The first used an anti-human IgG (UP892370) as capture anti-body and the biotinylated anti-IL-15 (BAM247), also recogniz-ing the IL-15 moiety in RLI, as revealing antibody. The secondused the same capture antibody as above and another anti-IgGantibody (109-036-003) as revealing antibody. This latter ELISAwas also used to measure anti-GD2 plasma levels.
Flow cytometry analysis
Cells (2 3 105) were incubated for 1 hr at 4�C with 10 mg/ml anti-GD2-RLI, anti-GD2 or control isotype IgG. Afterreaction with 1.25 mg/ml PE-labeled goat anti-human IgG asa secondary antibody (BD Biosciences, Le Pont de Claix,France), cell fluorescence was measured in a Calibur flow cy-tometer, and data were analyzed using FlowJo Software (BDBiosciences, Le Pont de Claix, France).
Binding assays
RLI was radiolabeled with 125I-labeled iodine (specific radioac-tivity of �2,000 cpm/fmol) using a chloramine T method asdescribed previously.28 Cells were starved for 24 hr in mediumwithout cytokine and washed twice in PBS containing 0.5%bovine serum albumin and 2% FCS. To determine inhibitionof RLI binding, 32Db cells (IL-15Rb/g cells) or Kit225 cells(IL-15Ra/b/g cells) were incubated with 400 pM of radiola-beled RLI and increasing concentrations of IL-15, RLI or anti-GD2-RLI during 1 hr at 4�C. Nonspecific binding was deter-mined in the presence of a 100-fold excess of unlabeled RLIand subtracted from total binding. IC50 was determined fromexperimental data using one-site or two-site-Fit logIC50 non-linear regression curves with GraphPad Prism software.
Proliferation assays
The proliferative responses of Kit225 and 32Db cells to IL-15,RLI or anti-GD2-RLI was assessed by [3H]-thymidine incorpo-ration as described previously.28 Briefly, cells were starved inthe culture medium without cytokine during 24 hr for Kit225or 4 hr for 32Db. They were plated at 1 3 104 cells in 100 mland cultured for 48 hr in the medium supplemented with IL-15, RLI or anti-GD2-RLI. Cells were pulsed for 16 hr with 0.5mCi per well of [3H]-thymidine and harvested onto glass fiberfilters, and then cell-associated radioactivity was measured.
ADCC and CDC assays
Lysis of IMR32 cells was evaluated using 51Cr release assays.Target cells (1 3 106) were incubated at 37�C with 75 mCi51Cr (Na2
51CrO4; Perkin-Elmer, Courtaboeuf, France) during 1hr. Labeled cells (3 3 103) were incubated with increasing
Can
cerTherapy
Vincent et al. 759
Int. J. Cancer: 133, 757–766 (2013) VC 2013 UICC

concentrations of antibodies for 1 hr with 10% human serumfor CDC assays or first incubated for 20 min with antibodiesfollowed by a 4-hr incubation period in the presence of effec-tor cells at different effector to target cells (E/T) ratios forADCC assays. Released 51Cr in the supernatant was countedwith a scintillation counter (Perkin-Elmer, Courtaboeuf,France). The percentage of specific lysis was calculated as fol-lows: (Experimental release2 Spontaneous release)/(Totalrelease2 Spontaneous release) 3 100%. Spontaneous releasewas determined by incubation of target cells in culture me-dium in the presence of effector cells or complement but with-out antibodies. Total release was determined by incubatingtarget cells with 1% Triton X-100.
Pharmacokinetic experiments
Biodisponibilities were evaluated in male C57BL/6 mice after asingle intraperitoneal injection of anti-GD2-RLI (16 mg) or anti-GD2 (12 mg). At each time point (up to 312 hr) following injec-tion, the blood was collected (three mice per point) and immedi-ately centrifuged, and the plasma was frozen at 220�C. Anti-GD2-RLI and anti-GD2 plasma levels were evaluated withELISA (see above). Pharmacokinetic parameters were calculatedusing a one-compartment model with GraphPad Prism software.
Mice and tumor model
Seven- to eight-week-old male C57BL/6 mice (Janvier, Le GenestSaint Isle, France) or female A/JOlaHsd mice (Harlan, Gannat,France) were used and maintained under pathogen-free condi-tions, and experiments were carried out in accordance withFrench laws and regulations. Mouse lymphoma EL4 cells (1 3
106) were transplanted subcutaneously on the right flank ofC57BL/6 mice using peptide hydrogel (Puramatrix, BD, Le Pontde Claix, France). Two experiments were done independently. Inthe first experiment, mice were injected intraperitoneally with sa-line or equimolar doses of RLI (2 mg), anti-GD2 (12 mg) or anti-GD2-RLI (16 mg) at Days 1, 4, 7 and 11 after transplantation. Inthe second experiment, three additional injections were per-formed at Days 14, 18 and 21. Tumor growth was monitoredthrice a week by measuring tumor width (W) and length (L)with calipers and by determining tumor volume according tothe following equation: tumor volume5 (L 3 W2) 3 0.5. Ani-mals were sacrificed when the tumor volume exceeded 3,000mm3. For the neuroblastoma metastatic mice model, female A/JOlaHsd mice were injected intravenously with 2.5 3 105 NXS2cells. On Days 1, 4, 7 and 11, mice were treated intraperitoneallywith saline or equimolar doses of RLI (0.3 mg), anti-GD2 (1.8mg), coadministration of anti-GD2 and RLI and anti-GD2-RLI(2.4 mg). A higher dose of 2 mg of RLI was also tested. Twenty-six days after tumor cell inoculation, the mice were sacrificedand their liver was weighted for evaluation of tumor metastasis.
Statistical analysis
The data are presented as mean6 SEM. The animal survivaldata were analyzed using Kaplan and Meier survival analysis.Statistical analysis used log-rank test for survival curves,
Newman-Keuls multiple comparison test for tumor modelsand F-test for pharmacokinetic experiments. p-values of lessthan 0.05 were considered significant.
ResultsCharacterization of the anti-GD2 mAb and anti-GD2-RLI ICK
The ICK construction is depicted in Figure 1a, and itsthree-dimensional structure representation is illustrated inFigure 1b. The RLI moiety is fused by its N-terminus to theC-terminus of the antibody heavy chain. The ICK was pro-duced in stably transfected CHO cells and purified by ProteinA affinity chromatography. SDS-PAGE analysis under NRconditions revealed high degrees of purity with single bandsof 150 and 200 kDa for anti-GD2 and anti-GD2-RLI, respec-tively (Fig. 1c), corresponding to the predicted molecularmasses of the antibody and the fusion protein. The identity
Figure 1. Characterization of the anti-GD2 mAb and anti-GD2-RLI
ICK. (a) ICK construction; H: heavy chain; L: light chain; PL: pepti-
dic linker within RLI fusion protein.28 (b) Ribbon drawing of anti-
GD2-RLI, X-ray crystallographic structure of IgG1 (Protein Data Bank
code 1hzh) was used, as well as the RLI structural model previ-
ously described.28 The Connolly surface of IL-15 was represented
showing interaction interfaces with IL-15Rb/g in yellow. Blue and
orange: H and L chains of the antibody; gray: PL; purple: N-termi-
nal (sushi1) domain of IL-15Ra. (c) SDS-PAGE under nonreducing
conditions (NR). (d) Western blot analysis using an anti-IL-15 mAb
after SDS-PAGE under NR conditions (top) or reducing (R) condi-
tions (bottom). For (c) and (d): Lane 1: anti-GD2 and Lane 2: anti-
GD2-RLI. MW: molecular weight; kDa: kilodalton.
Can
cerTherapy
760 RLI-based immunocytokine targeting GD2
Int. J. Cancer: 133, 757–766 (2013) VC 2013 UICC

of anti-GD2-RLI (NR) and of its heavy chain (R) was furtherestablished by Western blot analysis with the B-E29 anti-IL-15 Mab (Fig. 1d).
Binding to GD2 or IL-15R
Anti-GD2-RLI bound to GD21 EL4 cells with similar meanbinding levels as anti-GD2 was revealed by flow cytometry(Fig. 2a, left panel). EL4 did not bind the irrelevant anti-CD20-RLI ICK (data not shown), excluding that part of theanti-GD2-RLI reactivity was through its cytokine moiety.Anti-GD2-RLI was also able to bind to the human lym-phoma cell line Kit225 that expresses endogenous IL-15Ra,IL-15Rb and IL-15Rg chains (Fig. 2a, right panel). By con-
trast, no binding was detected for anti-GD2, indicating thatanti-GD2-RLI was bound through its cytokine moiety.
The binding affinity of anti-GD2-RLI for IL-15R was thenevaluated through its ability to compete with RLI binding indifferent IL-15 receptor contexts (Fig. 2b and Table 1). Oncells that express the IL-15Rb/g heterodimer (Fig. 2b, leftpanel), RLI had a higher affinity than IL-15, as alreadyshown.28,29 Anti-GD2-RLI (IC50 5 0.42 nM) was found to beeven threefold more efficient than RLI (IC50 5 1.21 nM) tocompete with radioiodinated RLI binding. Finally, on Kit225cells that express both IL-15Ra/b/g and IL-15Rb/g43 (Fig.2b, right panel), IL-15 competed with RLI binding accordingto a biphasic pattern that corresponded to its binding at lowconcentrations to its high-affinity component (IL-15Ra/b/g;
Figure 2. Binding and activities of anti-GD2-RLI. (a) Specific binding of anti-GD2-RLI and anti-GD2 antibodies revealed by flow cytometry:
EL4 cells (left panel) and Kit225 cells (right panel) were used. Anti-GD2 (black line), anti-GD2-RLI (gray line) and control antibody (filled
gray). (b) Competition of anti-GD2-RLI with RLI binding to cell surface IL-15R: 32Db cells (left panel) or Kit225 cells (right panel) were used.
IL-15 (�), RLI (�) and anti-GD2-RLI (�). (c) Effects of ICK on cell proliferation through the cell surface IL-15Ra/b/g or IL-15Rb/g: 32Db cells
(left panel) or Kit225 cells (right panel) were cultured with increasing concentrations of human IL-15 (�), RLI (�), anti-GD2-RLI (�) or anti-
GD2 (�). (d) Antibody cytotoxic effector functions on IMR32 target cells: ADCC (left panel) at an E/T ratio of 30:1; CDC (right panel). DC:
Decomplemented human serum was used for negative control. RTX was used as an irrelevant mAb. For (b)–(d), data are expressed as
mean 6 SEM of triplicate determinations and are representative of at least three independent experiments.
Can
cerTherapy
Vincent et al. 761
Int. J. Cancer: 133, 757–766 (2013) VC 2013 UICC
IC50 5 0.07 nM; 61% of total signal) followed by its bindingat higher concentrations to its low affinity component (IL-15Rb/g; IC50 5 3.53 nM; 39% of total signal). RLI competedwith a single inhibition phase (IC50 5 0.96 nM), in agreementwith the fact that it has the same affinity for both types ofreceptors.43 Anti-GD2-RLI was also found to compete radio-iodinated RLI binding with a single inhibition phase,although in this case with a threefold lower efficiency thanRLI (IC50 5 2.97 nM).
Cytokine activities
On 32Db cells (IL-15Rb/g), RLI was about eightfold moreefficient than IL-15 (ED50 5 50 vs. 400 pM, respectively; Fig.2c, left panel). Unexpectedly, anti-GD2-RLI was fivefoldmore efficient (ED50 5 20 pM) than RLI in inducing 32Db
cells proliferation through IL-15Rb/g. In agreement with pre-vious reports,28,29,43 IL-15 and RLI induced the proliferationof Kit225 cells through IL-15Ra/b/g at similar low concen-trations (ED50 5 50 pM for both cytokines; Fig. 2c, rightpanel). On these cells, anti-GD2-RLI showed similar prolifer-ative dose-response curves as RLI. As negative control, anti-GD2 had no proliferative effect on either cell lines.
Antibody effector functions
The ADCC and CDC effector functions were evaluated on theGD2-positive IMR32 target cells. Anti-GD2-RLI induced similarADCC as anti-GD2. The effects were time dependent, effectorto target (E/T) cell ratio dependent (data not shown) and dosedependent (Fig. 2d, left panel). RTX, an anti-CD20 antibody,showed only background lysis, thereby demonstrating the anti-gen specificity of the assay. Anti-GD2-RLI, like anti-GD2, alsoelicited dose-dependent CDC on IMR32 cells in the presence ofhuman serum (Fig. 2d, right panel). Its effect was even fivefoldto tenfold stronger on a molar basis than that induced by theparental mAb. The specificity was assessed by the absence ofcytotoxicity when using decomplemented serum or RTX.
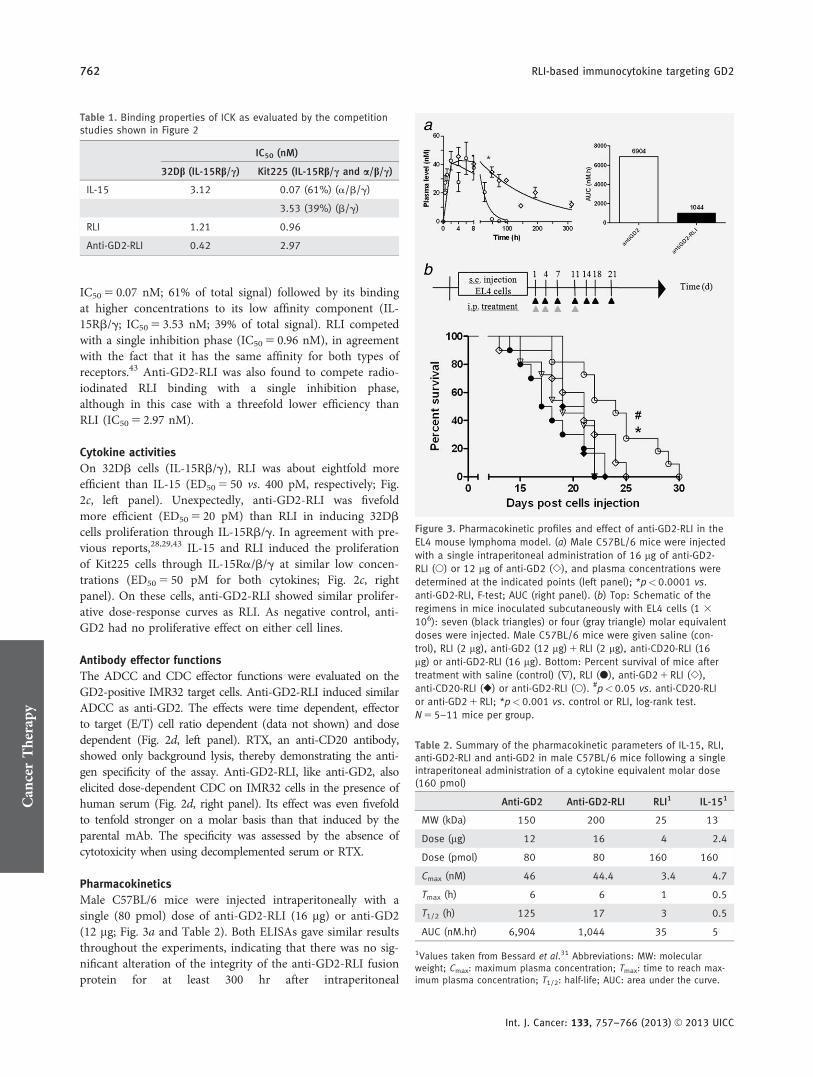
Pharmacokinetics
Male C57BL/6 mice were injected intraperitoneally with asingle (80 pmol) dose of anti-GD2-RLI (16 mg) or anti-GD2(12 mg; Fig. 3a and Table 2). Both ELISAs gave similar resultsthroughout the experiments, indicating that there was no sig-nificant alteration of the integrity of the anti-GD2-RLI fusionprotein for at least 300 hr after intraperitoneal
Table 1. Binding properties of ICK as evaluated by the competitionstudies shown in Figure 2
IC50 (nM)
32Db (IL-15Rb/c) Kit225 (IL-15Rb/c and a/b/c)
IL-15 3.12 0.07 (61%) (a/b/g)
3.53 (39%) (b/g)
RLI 1.21 0.96
Anti-GD2-RLI 0.42 2.97
Figure 3. Pharmacokinetic profiles and effect of anti-GD2-RLI in the
EL4 mouse lymphoma model. (a) Male C57BL/6 mice were injected
with a single intraperitoneal administration of 16 mg of anti-GD2-
RLI (�) or 12 mg of anti-GD2 (�), and plasma concentrations were
determined at the indicated points (left panel); *p<0.0001 vs.
anti-GD2-RLI, F-test; AUC (right panel). (b) Top: Schematic of the
regimens in mice inoculated subcutaneously with EL4 cells (1 3
106): seven (black triangles) or four (gray triangle) molar equivalent
doses were injected. Male C57BL/6 mice were given saline (con-
trol), RLI (2 lg), anti-GD2 (12 mg) 1 RLI (2 lg), anti-CD20-RLI (16
mg) or anti-GD2-RLI (16 mg). Bottom: Percent survival of mice after
treatment with saline (control) (r), RLI (�), anti-GD2 1 RLI (�),
anti-CD20-RLI (�) or anti-GD2-RLI (�). #p<0.05 vs. anti-CD20-RLI
or anti-GD2 1 RLI; *p<0.001 vs. control or RLI, log-rank test.
N 5 5–11 mice per group.
Table 2. Summary of the pharmacokinetic parameters of IL-15, RLI,anti-GD2-RLI and anti-GD2 in male C57BL/6 mice following a singleintraperitoneal administration of a cytokine equivalent molar dose(160 pmol)
Anti-GD2 Anti-GD2-RLI RLI1 IL-151
MW (kDa) 150 200 25 13
Dose (mg) 12 16 4 2.4
Dose (pmol) 80 80 160 160
Cmax (nM) 46 44.4 3.4 4.7
Tmax (h) 6 6 1 0.5
T1/2 (h) 125 17 3 0.5
AUC (nM.hr) 6,904 1,044 35 5
1Values taken from Bessard et al.31 Abbreviations: MW: molecularweight; Cmax: maximum plasma concentration; Tmax: time to reach max-imum plasma concentration; T1/2: half-life; AUC: area under the curve.
Can
cerTherapy
762 RLI-based immunocytokine targeting GD2
Int. J. Cancer: 133, 757–766 (2013) VC 2013 UICC

administration. The pharmacokinetic profile of anti-GD2 inmice (half-life of 125 hr) was comparable to that alreadydescribed for the mouse/human chimeric Ch14.18 anti-GD2antibody (half-life of 118 hr after 20 mg of intravenousadministration in mice).44 In comparison, anti-GD2-RLIshowed a strongly reduced bioavailability. Although display-ing a similar maximal peak of plasma concentration (Cmax ofabout 40 nM) as the parental antibody, its half-life (17 hr)and area under plasma concentration curve (AUC; 1,044nM.hr) were about sevenfold lower than those of anti-GD2(125 hr and 6,904 nM.hr). Nevertheless, the pharmacokineticparameters of anti-GD2-RLI were far higher than those pre-viously measured for the RLI cytokine (3 hr and 35 nM.hr),indicating that the antibody fusion procedure markedlyenhanced the bioavailability of RLI (Table 2).
Antitumoral activity in a primary tumor model
The subcutaneous growth of a primary tumor after EL4 lym-phoma cells injection was observed in all animals. Twoexperiments were done independently (Fig. 3b). In the firstexperiment, drugs were injected intraperitoneally at Days 1,4, 7 and 11 after EL4 inoculation, and in the second experi-ment, three additional injections were performed at Days 14,18 and 21. Within each group, no significant differences intumor growth were observed between the two experiments, aresult that allowed pooling the results. The mice survivalcurves were plotted according to the Kaplan-Meier methodand compared using log-rank test (Fig. 4c). Compared to themedian survival of vehicle-treated mice (18 days), mediansurvivals were not significantly modified after treatment withRLI, anti-GD2 plus RLI or the tumor antigen irrelevant ICKanti-CD20-RLI (median survivals of 21, 20 and 19 days,respectively). By contrast, anti-GD2-RLI ICK significantlyincreased the median survival to 24 days.
Antitumoral activity in a metastatic tumor model
The antitumoral efficacies of anti-GD2 and anti-GD2-RLI werecompared in the NXS2 neuroblastoma liver metastasis model.Flow cytometry showed that NXS2 cells did not express any ofthe IL-15Ra, IL-15Rb or IL-15Rg chains, and their proliferationin vitro was not affected by either IL-15 nor RLI, therefore rul-ing out any potential direct effect of the RLI moiety on the tu-mor cells (data not shown). In the absence of treatment (salinecontrol), neuroblastoma liver metastasis resulted in an increaseof the liver weight from 0.926 0.07 g (healthy control liver) to1.466 0.08 g (100% increase; Fig. 4). Treatment with the anti-GD2 mAb alone (1.8 mg) induced a partial (53%) inhibition ofthis tumor-related liver weight increase, whereas treatment withRLI alone (0.3 mg) had no significant effect. The association ofRLI with anti-GD2 did not enhance the effect of the antibody,but remarkably, treatment with anti-GD2-RLI (2.4 mg) resultedin a complete prevention of the liver weight increase with amean weight value (0.956 0.03 mg) similar to that of healthycontrols. However, treatment with a sevenfold higher dose ofRLI (2 mg) induced a partial (55%) inhibition effect.
DiscussionGD2 has been described as a relevant target for ADCCand CDC.40 We first showed that the fusion of RLI to the C-ter-minus of the anti-GD2 antibody heavy chain in anti-GD2-RLIdid not interfere with the recognition of the GD2 antigen asrevealed by flow cytometry. The cytotoxic effector functions(ADCC, CDC) of the antibody in vitro were then found to beconserved, and even increased in the case of CDC, suggestingthat fusing RLI to the antibody C-terminus did not decrease thebinding of the latter to Fc receptors or complement. The reasonfor the increased CDC activity remains to be understood. Adirect conformational effect of RLI on the binding of C1qappears unlikely, given the fact that the C1q binding site on theCH2 domain of the antibody heavy chain is located at a distanceof its C-terminus. The fusion process did not affect the recogni-tion by the RLI of the IL-15 receptors as revealed by flow cytom-etry. The affinity of anti-GD2-RLI for IL-15Rb/g was evenfourfold increased compared to unconjugated RLI. This increasedaffinity could reflect a bivalent mode of binding involving thetwo RLI moieties. We have also shown (unpublished results) thatthe addition of a flexible peptidic linker between the antibodyand RLI moieties did not modify the in vitro properties of theanti-GD2-RLI ICK, likely because the IL-15Ra domain of RLIplays the role of a linker, allowing a good presentation of the IL-15 binding interface of RLI to IL-15Rb/g.
The intracellular domains of the IL-15Rb and g-chains areconsidered to be the major actors responsible for the initiation
Figure 4. Effect of anti-GD2-RLI in the NXS2 mouse neuroblastoma
model. Top: Schematic of the regimen in mice inoculated intrave-
nously with NXS2 neuroblastoma cells (2.5 3 105). Bottom:
Female A/JOlaHsd mice were given molar equivalent doses of sa-
line (control), RLI (0.3 lg), anti-GD2 (1.8 lg) or anti-GD2-RLI (2.4
mg). At Day 26, mice were sacrificed and liver weights were meas-
ured. *p<0.05; **p<0.001 vs. control; #p<0.05, ##p<0.01,###p<0.001 vs. anti-GD2-RLI, Newman-Keuls multiple comparison
test. N 5 5–10 mice per group.
Can
cerTherapy
Vincent et al. 763
Int. J. Cancer: 133, 757–766 (2013) VC 2013 UICC

of signal transduction. Our team previously showed that IL-15cis- and trans-presentation mechanisms led to different dynamicsof receptor activation and signal transduction, with cis-presenta-tion (IL-15Ra/b/g activation) inducing fast and transientresponses and trans-presentation (IL-15Rb/g activation) induc-ing slower and more persistent ones.43 These differences werenot linked to the use of different signal transduction pathwaysbut rather related to differences in kinetics of cytokine–receptorcomplex internalization and persistence of functional receptors atthe cell surface, the intensity of the proliferative response beingcorrelated with the integration over time of the transduction sig-nal. On Kit225 cells, which express both IL-15Ra/b/g and IL-15Rb/g, IL-15 induces proliferation mainly through IL-15Ra/b/g activation (cis mode), whereas RLI acts through IL-15Rb/gactivation (trans-mode). The lower binding affinity of RLI forIL-15Rb/g than of IL-15 for IL-15Ra/b/g (0.96 vs. 0.07 nM,respectively) is therefore compensated by a more persistent re-ceptor activation in the case of RLI, resulting in similar prolifera-tive effects. On 32Db cells, IL-15 and RLI are both actingthrough IL-15Rb/g (trans-mode), and their proliferativeresponses therefore correlated with their respective binding affin-ities, RLI being eightfold more active than IL-15. Unexpectedly,the anti-GD2-RLI ICK was found to be fivefold more efficientthan RLI in inducing 32Db cell proliferation, despite displayingsimilar binding affinities as RLI on IL-15Rb/g. These results sug-gest that the ICK, may be due to its bigger molecular size and/or bivalent nature, displays even slower dynamics of receptorinternalization and more persistent activation than RLI alone.Further investigations are needed to check this hypothesis.
In two tumor models in vivo, the anti-GD2-RLI ICK dem-onstrated strong antitumoral activities that were higher thanthose elicited by the antibody alone, the cytokine alone andeven their combination, hence demonstrating the interest ofthis ICK approach.
In the tumor model in which EL4 lymphoma cells growas a primary tumor after subcutaneous injection,40 anti-GD2-RLI significantly inhibited tumor development and increasedmice mean survival, whereas RLI or the association of anti-GD2 and RLI, at equimolar doses, were without significanteffects. The irrelevant ICK anti-CD20-RLI was also withoutsignificant effect, suggesting an essential role of the anti-GD2component to target RLI to the tumor site, where the RLI-based immunostimulatory functions and antibody cytotoxicfunctions can synergize. ADCC triggered by human Fc inmouse tumor model has already been demonstrated in vivoin the case of RTX, another mouse/human chimeric IgG1directed against CD20, in experiments using FcRg chain-defi-cient mice45 or NK cell-depleted mice,46 in agreement with
the fact that mouse FcRgs efficiently bind human IgG sub-classes.47 However, an even higher ADCC effect of the RLI-based ICK may be expected in a human environment.Finally, the lower potency of human IL-15 compared to mu-rine IL-15 to generate NK cells in mouse48 might also con-tribute to a higher effect of the ICK in human.
The efficacy and interest of anti-GD2-RLI as antitumoralagent were confirmed in the experimental liver metastasis modelusing NXS2 cells. Anti-GD2-RLI was found to totally inhibitthe liver metastatic development, whereas at equivalent molardoses, RLI was without any effect, anti-GD2 was only partially(53%) efficient and the association of both did not increase thepartial effect of the antibody. These results demonstrate the in-terest of targeting RLI to the tumor site. The higher in vitroactivities of anti-GD2-RLI compared to RLI (fivefold higher af-finity for IL-15Rb/g and eightfold higher proliferative responsethrough IL-15Rb/g) or anti-GD2 (tenfold higher CDC) alsolikely participate to this increased in vivo antitumoral effect.Another parameter that could contribute to the higher efficacyof anti-GD2-RLI in vivo is the enhanced bioavailability of RLIin such ICK context. Indeed, the serum half-life and AUC ofRLI31 were increased by about sixfold and 30-fold, respectively,when fused to anti-GD2. This is in line with the fact that byincreasing the dose of RLI alone to sevenfold induced a signifi-cant (55%) antitumoral effect. In both in vivo models, the anti-tumoral effects of the cytokine part of the fusion protein werenot due to a direct action on the tumor cells that do notexpress any of the IL-15R chains and probably reflect an actionvia tumor-associated CD81 T cells26 and/or NK cells.31,49
In a recent report, a superior antitumoral activity of anscFv-RD-IL-15 fusion molecule was also associated with a morefavorable pharmacokinetics provided by the scFv moiety.50 Theinterest of targeting RLI to the tumor site by the mean of awhole antibody has not been reported so far, and our studyhighlights its therapeutical benefits. These benefits are related tothe improved pharmacokinetic properties of the cytokine pro-vided by the whole antibody (half-live of 17 hr for IgG vs. 8 hrfor scFv) together with the preservation of its effector properties(ADCC and CDC). This synergistic combination should allow areduction of the therapeutic doses and limit the side effects ofboth components. Therefore, such fusion proteins appear aspromising drugs in cancer immunotherapy.
AcknowledgementsM.V. was supported by fellowships from the Ministere de l’Enseigne-ment Sup�erieur et de la Recherche and the Association pour la Re-cherche sur le Cancer. The authors thank Virginie Maurier fortechnical assistance.
References
1. Meazza R, Azzarone B, Orengo AM, et al. Roleof common-g chain cytokines in NK celldevelopment and function: perspectives forimmunotherapy. J Biomed Biotechnol2011;2011:861920.
2. Carnemolla B, Borsi L, Balza E, et al.Enhancement of the antitumor properties ofinterleukin-2 by its targeted delivery to the tumorblood vessel extracellular matrix. Blood2002;99:1659–65.
3. Schwartz RN, Stover L, Dutcher J. Managingtoxicities of high-dose interleukin-2. Oncology(Williston Park, NY) 2002;16:11–20.
4. Halin C, Rondini S, Nilsson F, et al.Enhancement of the antitumor activity of
Can
cerTherapy
764 RLI-based immunocytokine targeting GD2
Int. J. Cancer: 133, 757–766 (2013) VC 2013 UICC

interleukin-12 by targeted delivery toneovasculature. Nat Biotechnol 2002;20:264–9.
5. Pasche N, Neri D. Immunocytokines: a novelclass of potent armed antibodies. Drug DiscovToday 2012;17:583–90.
6. Kontermann RE. Antibody-cytokine fusionproteins. Arch Biochem Biophys 2012;526:194–205.
7. Johannsen M, Spitaleri G, Curigliano G, et al.The tumour-targeting human L19-IL2immunocytokine: preclinical safety studies, phaseI clinical trial in patients with solid tumours andexpansion into patients with advanced renal cellcarcinoma. Eur J Cancer 2010;46:2926–35.
8. Gilman AL, Ozkaynak MF, Matthay KK, et al.Phase I study of ch14.18 with granulocyte-macrophage colony-stimulating factor andinterleukin-2 in children with neuroblastomaafter autologous bone marrow transplantation orstem-cell rescue: a report from the Children’sOncology Group. J Clin Oncol 2009;27:85–91.
9. Baluna R, Rizo J, Gordon BE, et al. Evidence fora structural motif in toxins and interleukin-2 thatmay be responsible for binding to endothelialcells and initiating vascular leak syndrome. ProcNatl Acad Sci USA 1999;96:3957–62.
10. Munger W, DeJoy SQ, Jeyaseelan R, Sr, et al.Studies evaluating the antitumor activity andtoxicity of interleukin-15, a new T cell growthfactor: comparison with interleukin-2. CellImmunol 1995;165:289–93.
11. Steel JC, Waldmann TA, Morris JC. Interleukin-15 biology and its therapeutic implications incancer. Trends Pharmacol Sci 2012;33:35–41.
12. Giri JG, Anderson DM, Kumaki S, et al. IL-15, anovel T cell growth factor that shares activitiesand receptor components with IL-2. J Leukoc Biol1995;57:763–6.
13. Anderson DM, Kumaki S, Ahdieh M, et al.Functional characterization of the humaninterleukin-15 receptor a chain and close linkageof IL15RA and IL2RA genes. J Biol Chem1995;270:29862–9.
14. Giri JG, Kumaki S, Ahdieh M, et al. Identificationand cloning of a novel IL-15 binding protein thatis structurally related to the a chain of the IL-2receptor. EMBO J 1995;14:3654–63.
15. Marks-Konczalik J, Dubois S, Losi JM, et al. IL-2-induced activation-induced cell death is inhibitedin IL-15 transgenic mice. Proc Natl Acad Sci USA2000;97:11445–50.
16. Waldmann TA, Dubois S, Tagaya Y. Contrastingroles of IL-2 and IL-15 in the life and death oflymphocytes: implications for immunotherapy.Immunity 2001;14:105–10.
17. Berger C, Berger M, Hackman RC, et al. Safetyand immunologic effects of IL-15 administrationin nonhuman primates. Blood 2009;114:2417–26.
18. Kennedy MK, Glaccum M, Brown SN, et al.Reversible defects in natural killer and memoryCD8 T cell lineages in interleukin 15-deficientmice. J Exp Med 2000;191:771–80.
19. Lodolce JP, Burkett PR, Boone DL, et al. T cell-independent interleukin 15Ra signals arerequired for bystander proliferation. J Exp Med2001;194:1187–94.
20. Li XC, Demirci G, Ferrari-Lacraz S, et al. IL-15and IL-2: a matter of life and death for T cells invivo. Nat Med 2001;7:114–18.
21. Dubois S, Mariner J, Waldmann TA, et al. IL-15Ra recycles and presents IL-15 in trans toneighboring cells. Immunity 2002;17:537–47.
22. Burkett PR, Koka R, Chien M, et al. Coordinateexpression and trans presentation of interleukin(IL)215Ra and IL-15 supports natural killer celland memory CD81 T cell homeostasis. J ExpMed 2004;200:825–34.
23. Mortier E, Advincula R, Kim L, et al.Macrophage- and dendritic-cell-derivedinterleukin-15 receptor a supports homeostasis ofdistinct CD81 T cell subsets. Immunity2009;31:811–22.
24. Mortier E, Bernard J, Plet A, et al. Natural,proteolytic release of a soluble form of humanIL-15 receptor a-chain that behaves as a specific,high affinity IL-15 antagonist. J Immunol2004;173:1681–8.
25. Dubois S, Patel HJ, Zhang M, et al.Preassociation of IL-15 with IL-15R a-IgG1-Fcenhances its activity on proliferation of NK andCD81/CD44 high T cells and its antitumoraction. J Immunol 2008;180:2099–106.
26. Epardaud M, Elpek KG, Rubinstein MP, et al.Interleukin-15/interleukin-15R a complexespromote destruction of established tumors byreviving tumor-resident CD81 T cells. CancerRes 2008;68:2972–83.
27. Stoklasek TA, Schluns KS, Lefrancois L.Combined IL-15/IL-15Ra immunotherapymaximizes IL-15 activity in vivo. J Immunol2006;177:6072–80.
28. Mortier E, Qu�em�ener A, Vusio P, et al. Solubleinterleukin-15 receptor a (IL-15R a)-sushi as aselective and potent agonist of IL-15 actionthrough IL-15R b/g. Hyperagonist IL-15 3 IL-15R a fusion proteins. J Biol Chem2006;281:1612–19.
29. Bouchaud G, Garrigue-Antar L, Sol�e V, et al. Theexon-3-encoded domain of IL-15ra contributes toIL-15 high-affinity binding and is crucial for theIL-15 antagonistic effect of soluble IL-15Ra. JMol Biol 2008;382:1–12.
30. Huntington ND, Legrand N, Alves NL, et al. IL-15 trans-presentation promotes human NK celldevelopment and differentiation in vivo. J ExpMed 2009;206:25–34.
31. Bessard A, Sol�e V, Bouchaud G, et al. Highantitumor activity of RLI, an interleukin-15 (IL-15)-IL-15 receptor a fusion protein, in metastaticmelanoma and colorectal cancer. Mol CancerTher 2009;8:2736–45.
32. Cahan LD, Irie RF, Singh R, et al. Identificationof a human neuroectodermal tumor antigen(OFA-I-2) as ganglioside GD2. Proc Natl AcadSci USA 1982;79:7629–33.
33. Yuki N, Yamada M, Tagawa Y, et al.Pathogenesis of the neurotoxicity caused by anti-GD2 antibody therapy. J Neurol Sci1997;149:127–30.
34. Svennerholm L, Bostr€om K, Jungbjer B, et al.Membrane lipids of adult human brain: lipidcomposition of frontal and temporal lobe insubjects of age 20 to 100 years. J Neurochem1994;63:1802–11.
35. Birkl�e S, Zeng G, Gao L, et al. Role of tumor-associated gangliosides in cancer progression.Biochimie 2003;85:455–63.
36. Mueller BM, Romerdahl CA, Gillies SD, et al.Enhancement of antibody-dependent cytotoxicitywith a chimeric anti-GD2 antibody. J Immunol1990;144:1382–6.
37. Imai M, Landen C, Ohta R, et al. Complement-mediated mechanisms in anti-GD2 monoclonalantibody therapy of murine metastatic cancer.Cancer Res 2005;65:10562–8.
38. Cheever MA, Allison JP, Ferris AS, et al. Theprioritization of cancer antigens: a nationalcancer institute pilot project for the accelerationof translational research. Clin Cancer Res2009;15:5323–37.
39. Handgretinger R, Anderson K, Lang P, et al. Aphase I study of human/mouse chimericantiganglioside GD2 antibody ch14.18 in patientswith neuroblastoma. Eur J Cancer 1995;31:261–7.
40. Alvarez-Rueda N, Leprieur S, Cl�emenceau B,et al. Binding activities and antitumor propertiesof a new mouse/human chimeric antibodyspecific for GD2 ganglioside antigen. Clin CancerRes 2007;13:5613s–5620s.
41. Hori T, Uchiyama T, Tsudo M, et al.Establishment of an interleukin 2-dependenthuman T cell line from a patient with T cellchronic lymphocytic leukemia who is not infectedwith human T cell leukemia/lymphoma virus.Blood 1987;70:1069–72.
42. Nakamura Y, Russell SM, Mess SA, et al.Heterodimerization of the IL-2 receptor b- andg-chain cytoplasmic domains is required forsignalling. Nature 1994;369:330–3.
43. Perdreau H, Mortier E, Bouchaud G, et al.Different dynamics of IL-15R activation followingIL-15 cis- or trans-presentation. Eur CytokineNetw 2010;21:297–307.
44. Kendra K, Gan J, Ricci M, et al.Pharmacokinetics and stability of the ch14.18-interleukin-2 fusion protein in mice. CancerImmunol Immunother 1999;48:219–29.
45. Clynes RA, Towers TL, Presta LG, et al.Inhibitory Fc receptors modulate in vivocytotoxicity against tumor targets. Nat Med2000;6:443–6.
46. Hernandez-Ilizaliturri FJ, Jupudy V, Ostberg J,et al. Neutrophils contribute to the biologicalantitumor activity of rituximab in a non-Hodgkin’s lymphoma severe combinedimmunodeficiency mouse model. Clin Cancer Res2003;9:5866–73.
47. Bruhns P. Properties of mouse and human IgGreceptors and their contribution to diseasemodels. Blood 2012;119:5640–9.
48. Eisenman J, Ahdieh M, Beers C, et al.Interleukin-15 interactions with interleukin-15receptor complexes: characterization and speciesspecificity. Cytokine 2002;20:121–9.
49. Rowley J, Monie A, Hung C-F, et al. Inhibitionof tumor growth by NK1.11 cells and CD81 Tcells activated by IL-15 through receptor b/common g signaling in trans. J Immunol2008;181:8237–47.
50. Kermer V, Baum V, Hornig N, et al. Anantibody fusion protein for cancerimmunotherapy mimicking IL-15 trans-presentation at the tumor site. Mol Cancer Ther2012;11:1279–88.
Can
cerTherapy
Vincent et al. 765
Int. J. Cancer: 133, 757–766 (2013) VC 2013 UICC





























