Embed Size (px)
Citation preview

Tryptophan-rich antimicrobial peptides: properties and applications
Rebecca L. Alfred*, Nadin M. Shagaghi*, Enzo A. Palombo and Mrinal Bhave
Environment and Biotechnology Centre, Faculty of Life and Social Sciences, Swinburne University of Technology, PO Box 218, Melbourne, Victoria 3122, Australia
*RLA and NMS have contributed equally to the manuscript.
There is a world-wide quest for development of new-generation antibiotics, due to the emergence of microbial pathogens that have developed resistance to the currently available antibiotics. The antimicrobial peptides (AMPs) are a strong candidate in this regard, due to minimal possibilities of emergence of resistance against these. The tryptophan (Trp)-rich peptides comprise an important class of AMPs. The amino acid Trp has unique biochemical properties that allow it to interact with and insert into biological membranes. This feature, together with the presence of basic residues in many AMPs that enable selective binding to microbial membranes, imparts strong antimicrobial properties on Trp-rich peptides. Such peptides thus display activity against a range of bacteria, fungi and viruses, and some are also active against protozoan pathogens and/or cancer cells. The mechanisms of action of the peptides may involve cell lysis or intracellular effects. The applications of such AMPs range from human health and hygiene to plant pathogen control.
Keywords Antimicrobial peptides; tryptophan; tryptophan-rich peptides; novel antibiotics.
1. Introduction
There is significant research worldwide to discover novel antibiotics for control of microbial infections, with the view of identifying those with minimal possibility of development of resistance. A great diversity of antimicrobial peptides (AMPs) has been identified in this quest, from various taxa of animals including humans, amphibians, insects, and from plants. A majority of these AMPs contain 12-50 amino acid residues, are cationic antimicrobial peptides (CAPs), having a net charge of +2 to +7 due to their high content of arginine (Arg), lysine (Lys) and/or histidine (His) residues. Many are also amphipathic (i.e., with distinct polar and non-polar regions), and often contain about 50% residues which are hydrophobic. The AMPs/CAPs can be classified into four main classes, based on sequence characteristics and secondary structures.
1. CAPs with a high degree of β-sheet structures, stabilized by disulphide bonds. This is the largest group and includes the α and β-defensins (vertebrates), drosomycin (fruit flies) and thionins (cereals).
2. α-helical peptides that are mostly unstructured until binding to a membrane, which induces formation of α-helical structures, e.g., cecropins (moths), cecropin-like peptides (pigs) and magainins (African frogs).
3. Peptides formed by the proteolytic digestion of larger proteins, e.g., bovine lactoferricin, from lactoferrin. 4. Peptides with a predominance of certain amino acids, particularly proline (Pro), and tryptophan (Trp). These
may or may not also be ‘CAPs’, based on their net charge. The Pro-rich peptides are exemplified by apidaegins (bee and wasps), and the Trp-rich peptides include indolicidin and the wheat puroindoline (PIN)-based peptides.
The structures and properties of AMPs have been reviewed elsewhere [1, 2]. This work focusses on Trp-rich peptides, and considers their unique characteristics and applications.
2. Trp-rich AMPs: relevance of the unique biochemistry of tryptophan
Trp residues, upon which this class of peptides takes its name, constitute the active core of these peptides. Tryptophan is an aromatic, hydrophobic, neutral amino acid, and is the largest amino acid with characteristic indole functional group. Trp has a strong tendency to insert into biological membranes and partition near the membrane-water interface, a feature which can cause it to affect lipid packing. The Trp residues appear to prefer to position specifically near the lipid carbonyl region [3]. These properties give it both hydrophilic and hydrophobic attributes, making it ideal for insertion into the membranes of microbial cells [4]. The large indole side chain of Trp is about a third of the thickness of a phospholipid monolayer, and the preference of Trp for the interfacial region makes it ideal as the hydrophobic face of a membrane-interacting amphipathic structure [3]. The Trp side chains have been strongly associated with antimicrobial activity due to this preference [5]. An important feature to note is that Trp residues seem to have many different and contrasting positions on the hydrophobicity scale of amino acids. In certain cases, Trp is considered hydrophobic due to its uncharged side-chain; however, it does not reside in the hydrocarbon region of lipid bilayers and is thus placed towards the hydrophilic side of the scale [5]. Further, Trp residues seem to play a role in protein folding, by native and non-native hydrophobic links even in denatured proteins [5].
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
1395

3. Trp-rich AMPs with antibacterial, antifungal and/or antiviral activities
3.1. Indolicidin and analogues:
First isolated from the cytoplasmic granules of bovine neutrophils [6], indolicidin (ILPWKWPWWPWRR-NH2), takes its name from the indole ring of Trp. It consists mainly of Trp (39%) and Pro (23%), has a naturally amidated (NH2) C-terminus and is unique in having the highest Trp content of any naturally occurring peptide reported as yet. It is one of the most extensively characterised Trp-rich peptides, showing a broad range of activities against bacteria [6, 7] and fungi [8] (summarised in Table 1). Indolicidin was found to permeabilise membranes of Escherichia coli, but not result in cell lysis [7]. It is too small to span the bilayer, but its ability to form an extended loop rather than α-helix appears to compensate for this [7]. It may have intracellular mechanisms of action (discussed later). Falla et al. [7] also investigated the significance of its natural N-terminal amidation, and found that indolicidin peptides with the amidation exhibited greater activity against both Gram-positive (2-fold) and Gram negative (4-fold) bacteria, compared to those without it. The greater activity is suggested to be due to the higher net positive charge, increasing its binding potential to the negatively-charged microbial membranes. This concept has been widely accepted, with a number of synthetic peptides made with this modification (e.g., our work) [9]. However, Dennison et al. [10] have challenged this view; assessment of C-terminally amidated and non-amidated isoforms of non-Trp-rich AMPs (e.g., Hectate, Shiva 10, FLAK 26, SB-37, Modeline-5) showed that the modification had no consistent effect; e.g., the potency of amidated isoforms of FLAK 26, SB-37 and Modelin-5 on Staplylococcus aureus was up eight-fold, down two-fold and similar to the non-amidated counterparts, respectively. It was hence suggested that the C-terminal amine may have different effects depending on the structure and sequence of a specific peptide. Similar investigations of Trp-rich AMPs are required. In addition to traditional antibacterial activity, antibiofilm activities of antimicrobial agents are important for treating biofilm-related infections, such as those caused by methicillin-resistant Staphylococcus aureus (MRSA). Indolicidin alone, and in combination with various antibiotics, has shown significant inhibitory activity against MRSA biofilm attachment and formation [11], indicating AMPs make good candidates for investigation into the treatment of biofilm-related infections. Similar to bacteria and fungi, the surfaces of many viruses are negatively charged, making them candidates for attack by CAPs (reviewed by Jenssen et al.) [2]. Of the Trp-rich AMPs, indolicidin was the first to be investigated for antiviral activity. Robinson et al. [12] assessed the activity of indolicidin and an analogue R12-OH (ILPWKWPWWPWR) against the enveloped Human Immunodeficiency Virus (HIV)-1. Indolicidin inactivated the virus within 1 hour of application; however at the same concentration (166 μg/mL) it was cytotoxic towards CD4+ expressing lyphoblastoid cell lines, suggesting a non-selective membrane disruptive mechanism for the anti-HIV activity. Interestingly, the R12-OH analogue was less effective, indicating the positive charge may be essential for antiviral activity, and it was also not cytotoxic to the cell line, suggesting removal of the natural C-terminal amidation of indolicidin (see above) results in increased selectivity for viral particles [12]. Further investigations on the mode of antiviral activity of indolicidin, its analogues and small Trp-rich peptides identified HIV intergrase [13, 14] as a specific target, rather than a membrane-disruptive mechanism. Native indolicidin has HIV-1 intergrase inhibitory activity and an analogue with the C-terminal Arg replaced by Pro (ILPWKWPWWPWPP) exhibited a 4-fold increase in the HIV-1 intergrase inhibitory activity [14]. Dimerisation of the analogue by connection of the C-terminal ends of monomers using Lys as a linker further increased HIV-1 intergrase inhibitory activity significantly, and the tetrameric peptide, prepared by connecting two dimers at C-ends was the most potent HIV-1 intergrase inhibitor, more potent than indolicidin [14]. The results indicated that the size and structure of the peptide are important for antiviral activity, not just sequence and charge. Indolicidin also exhibits strong activity against Herpes Simplex Virus (HSV), more so against HSV-2 than HSV-1, directly inactivating viral particles without cytotoxicity for Vero cells [15]. Further, two indolicidin analogues 10R (RRPWKPWWPWRR) and 11R (RWRRWPWWPWRRK) were shown to have activity against a plant virus, the Tobacco Mosaic Virus (TMV), with 10R being a 10-fold more active than 11R, despite 11R having a higher net positive charge [16]. It is unclear how this compares with native indolicidin, as there are no data for its activity against TMV. A short analogue of indolicidin called Omiganan (ILRWPWWPRRK-NH2) showed high activity against a number of clinically important microbial pathogens [17] (Table 1). Omiganan (1% w/w) aqueous gel was tested by applying at the catheter site and dressing changes [18]. It has completed Phase III clinical trials for prevention of catheter-related local and bloodstream infections, a major healthcare problem worldwide, the trial showing a reduction in microbial catheter colonisation [19]. Further, phase II trials against the non-infectious skin condition rosacea showed significant efficacy in reducing symptoms [20], and it is now in phase III trials.
3.2. Tritrpticin and analogues:
Tritrpticin (VRRFPWWWPFLRR) is derived from a precursor protein in the porcine bone marrow and has high homology to indolicidin, both having five aromatic residues, multiple Pro residues and comparable basic residues [21]. It has potent bactericidal and fungicidal activity against several clinically and environmentally important microorganisms [21, 22] (Table 1). In addition to the Trps, the positive charge seems important, as it facilitates the
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
1396

interaction with negatively charged microbial cell membranes. The double cationic Arg residues at/close to both termini of Tritrpticin appear to be important for its function, as deletion of even one Arg resulted in decreased antibacterial activity, while substitutions by Lys, also cationic, did not affect the activity [23]. However, as tested later, this change diminished the haemolytic activity, thus increasing the selectivity for microbial cells [24]. A number of Tritrpticin analogues have been created to elucidate the residues that are essential for its antimicrobial activity. C-terminal amidation resulted in a 2-fold increase in antibacterial activity [4], mirroring the findings on indolicidin [7]. Studies replacing the three Trps were also attempted; Phe was selected, as its side chain may promote deeper penetration into the membrane, and Tyr was selected as its side chain locates into the membrane interface [24]. The Trp to Phe substitution had no significant effect, whereas Trp to Tyr led to a 5-fold decrease in activity against E. coli and S. aureus. The analogues with Trp substituted by Phe/Tyr also had a considerably low haemolytic activity against mammalian red blood cells at high peptide concentrations (>1mg/mL) compared to Tritrpticin, suggesting the substitution of Trp by Phe may improve selectivity for microbial cells [24].
3.3. Lactoferricin and analogues:
These form a large class of CAPs found in different mammalian species, and produced by gastric-pepsin cleavage from the N-terminal part of the protein lactoferrin [25]. Partial degredation of bovine or human lactoferrin by gastric-pepsin produces two peptides, both of which have more antimicrobial activity than lactoferrin. However, the bovine lactoferricin (LFB17-41) (FKCRRWQWRMKKLGAPSITCVRRAF), composed of residues 17-41 of the full-length lactoferrin, has greater activity than human lactoferricin (47 amino acids) [25]. A shortened 15-mer analogue called LFB17-31 or LfcinB17-31 (FKCRRWQWRMKKLGA) also has strong activity against E. coli and S. aureus [4] and a variety of plant filamentous fungi [26] (Table 1). A further shortened 6-mer analogue designated LfcinB20-25 (RRWQWR) has also shown activity against pathogenic fungi but at reduced efficacy to LFB17-31 [26]. Unlike indolicidin and tritrpticin, LFB does not have a high proportion of Trp. Hence, to identify the residues important for its antibacterial activity, each residue was systematically replaced with alanine (Ala scan experiment) [4]. The results showed that the two Trps (Trp6 and Trp8) were most important, replacement of either with Ala abolishing the activity. The lactoferricins of pig (LFP), human (LFH), mouse (LFM) and goat (LFC) exhibit 33%, 40%, 53% and 67% similarity, respectively, to LFB at the protein level. Most importantly, these four lack the second Trp (Trp8), and show little or no antimicrobial activity. Introduction of a Trp at this position in these four peptides increased the activity against E. coli, and the second most important residue in LFB was Met10, a strongly hydrophobic residue. Interestingly, the replacement of the polar residues Cys3, Gln7 and Gly14 with Ala, a hydrophobic residue, resulted in increased antimicrobial activity [4]. Yang et al. [27] further worked on the antibacterial, anticancer and haemolytic activity of lactoferrin-derived peptides and found that the overall hydrophobicity affects the haemolytic activity; however, they did not investigate the effects of hydrophobicity on antimicrobial activity. Additional LFB-based peptides have also been found to have strong antimicrobial activities [22, 28, 29]. Lactoferricin and Lactoferrin both have been investigated as antiviral peptides. Lactoferrin (LF) exerts antiviral activity against Human Cytomegalovirus (HCMV) [30], hepatitis C virus [31], HIV [32] and HSV [33]. Hammer et al. [34] reported that bovine lactoferrin was more efficient than lactoferricin in inhibiting HSV-I/II replication. Hasegawa et al. [30] suggested that activity of lactoferrin HSV resulted from interference with early events of infection. The mode of action of lactoferrin and lactoferricin against HCMV seems to occur primarily at the cell surface level [35]. Lactoferrins from different species (human, goat, bovine) had different levels of activity against HCMV, the bovine form being most effective, indicating the differences in peptide sequences may affect antiviral activities (as well as for antimicrobial activity, see above), and lactoferrin was more efficient than lactoferricin, indicating that other regions of the full protein may be important [35]. Secondary structure of the peptide also seems important, as linear bovine lactoferricin had little anti-HCMV activity but the cyclic form with a disulphide between the Cys residues was more effective [35], mirroring the findings of Vorland et al. [36], and the antibacterial activity of lactoferricin B also decreased when the cysteine disulfides were reduced. Andersen et al. [35] also reported that lactoferrin and cyclic lactoferricin can inhibit HCMV by acting primarily at the cell surface and preventing the first contact between virus and host cell, possibly by blocking viral interactions with heparan sulphate (HS), the surface receptor for HCMV, and have no effect on viral protein production after it had entered the cell. Jenssen et al. [37] found that lactoferrin and lactoferricin analogues that exhibited anti-HSV activity bound to HS with high affinity. However, lactoferrins exhibited a greater activity than the smaller lactoferricins which had similar affinities for HS, and one analogue of lactoferricin with substitutions of six residues in the C-terminal had high affinity for HS but lower anti-HSV activity. The results support Andersen et al. [35] and show that in addition to ability to bind to HS, factors such as sequence, size and secondary structure may be important [37].
3.4. Brevinin-1EMa and analogues:
Brevinin-1EMa (originally called gaegurin 5), is a 24-residue AMP isolated from the skin of a Korean frog Glandirana emeljanovi [38]. In the search for its shortest bioactive analogue, the 11 residue N-terminal fragment (FLGALFKVASK-NH2) was found to be completely inactive. Its derivative GA-W2 with Trps at residues 4 and 8
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
1397

(FLGWLFKWASK-NH2) was found to be a potent AMP, but displayed strong haemolytic activity at higher concentrations. However, another derivative, GA-K4AL (FAKWAFKWLKK-NH2), displayed potent antimicrobial activity and little hemolytic activity, making it ideal as an AMP, possibly superior to omiganan (see above) [38].
3.5. Puroindoline-based peptides:
Two small, closely related proteins expressed in the wheat grain, called puroindoline-a (PINA) and puroindoline-b (PINB) [39] have been studied extensively in relation to their role in grain texture, a commercially critical phenotype (reviewed in [40]). They are ~13 kilo Dalton highly basic proteins with 55% similarity, and both contain a backbone of 10 Cys residues, forming 5 disulphide bonds, four α-helical sections, and a unique Trp-rich domain (TRD) comprised of five Trps in PINA and three in PINB. Wild type PINA and PINB bind to starch granules in the endosperm to effect soft texture, and mutations in or absence (due to natural gene deletion events) of either or both protein(s) results in the hard grain texture [40]. Aside from these effects, the commonly accepted hypothesis regarding the in vivo roles of these proteins is defence of the seed/seedling against pathogens. The first direct evidence of this potential was provided by in vitro studies using PINA and PINB purified from wheat against phytopathogenic fungi [41]. The synthetic peptides PuroA (FPVTWRWWKWWKG-NH2) and PuroB (FPVTWPTKWWKG-NH2) based on these proteins also have these activities, and these sequences were proposed to be the antimicrobial domains of PINA and PINB [42]. PINA and PINB proteins expressed in E. coli cells also showed antibacterial activities [43] and could kill intracellular Staphylococcus epidermidis without exhibiting haemolytic activity or toxicity to mouse macrophage cells [44]. We recently showed that peptides based on the TRD of PINA and PINB, the related wheat protein Grain Softness protein-1 (GSP-1) and the barley hordoindolines displayed strong activity against bacteria and phytopathogenic fungi [9], and the wheat PINB-2 proteins also share this property [45] (Table 1).
3.6. Rationally designed peptides:
In addition to the above natural AMPs and their synthetic analogues, rational design of peptides is an effective approach to improve the antimicrobial properties and selectivity [46] and reduce the manufacturing costs. In conjunction with Trp, other hydrophobic (His, Phe) and basic (Arg, Lys) residues have been used in combinatorial library or rational design approaches. PAF26 (Ac-RKKWFW-NH2) was identified from a hexapeptide combinatorial library designed and surveyed for activity towards phytopathogenic fungi that cause postharvest decay in citrus fruits [47] (Table 1). PAF26 and two derivatives containing N-terminal additions, PAF38 (Ac-RRKKWFW-NH2), PAF40 (Ac-HRKKWFW-NH2), as well as BM0 (Ac-RFWWFRRR-NH2, identified from an octapeptide library) have activity against Penicillium digitatum, Saccharomyces cerevisiae and E. coli [48]. P. digitatum causes postharvest green mold in citrus and shows resistance to many commercial fungicides; hence these peptides provide alternative novel plant disease control strategies [48]. Pac-525 (Ac-KWRRWVRWI-NH2) and its reversed sequence Pac-525rev (Ac-IWRVWRRWK-NH2) demonstrated antimicrobial activity against both Gram-positive and Gram-negative bacteria, and low hemolytic activity [22]. Pac-525 has four positively charged residues, distributed in an amphipathic structure made by the three Trp indole rings, and its activity was attributed to its ability to bind strongly and destabilize the negatively charged microbial membranes, not binding to a specific protein receptor. The amphipathic structures are propsoed to maximize both electrostatic and hydrophobic interactions with microbial membrane [22]. In a smiliar manner, two of the three Trp residues in PEM-2 (KKWRWWLKALAKK) form an apparent amphipathic conformation. PEM-2 is one of the series of short Trp-rich peptides derived from the C-terminal region of Bothrops asper myothoxin II, a Lys49 phospholipase A2 (PLA2), and exhibits activity against E. coli, S. aureus and Pseudomonas aeruginosa and low toxicity toward eukaryotic cells [49]. Using a more rational approach, Liu et al. [50] designed a series of peptides containing a repeated pattern of Arg-Trp ((RW)n, where n =1, 2, 3, 4, or 5) and varying in charge and hydrophobicity. Additionally, Gopal et al. extended this work by designing a second set of peptides containing a Lys-Trp repeated pattern ((KW)n where n = 2, 3 ,4 or 5) [51]. The RW series showed activity agianst E. coli and S. aureus [50], and Fusarium solani and F. oxysporum [51]. The antimicrobial activity increased with the peptide length, RW5 (RWRWRWRWRW-NH2) showing the highest activity; however, the hemolytic activity increased with the length as well. Therefore, (RW)3 (RWRWRW-NH2) and (RW)4 (RWRWRWRW-NH2) were identified as relatively nontoxic antibacterial and antifungal peptides, respectievly, that could be produced economically [50, 51]. Further, the KW series showed fungicidal activity against Fusarium species; but lower than that of the RW series [51]. The synthetic peptide PW2 is active against a number of fungi but not bacteria [52] and also anticoccidal activity (see below).
4. Trp-rich AMPs with anticoccidal (antiprotozoan) activity
Indolicidin shows anticoccidial activity against Giardia lamblia [53], possibly by binding and cell lysis by charged base interactions. PW2, a synthetic peptide (HPLKQYWWRPSI), isolated from a phage display library by a panning method using live sporozoites of Eimeria acervulina, exhibits activity against a number of fungi, but not bacteria [52] and
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
1398

anticoccidial activity against E. acervulina and Eimeria tenella, which are protozoan parasites that cause the economically important disease coccidiosis in poultry, and very low lytic activity against mammalian and avian host cells. The effectiveness against Eimeria sporozoites and absence of adverse effects indicates that PW2 may be used to develop new drugs for control of avian coccidiosis [52]. Tinoco et al. [54] suggested that its lack of activity against bacteria noted earlier [52] may be due to its low hydrophobicity (50%) compared to indolicidin (77%) and tritrpticin (69%), and that the low haemolytic activity may be due to the lower number of Trps, which correlates with the findings of Schibi et al. [24] for Tritrpticin (see above). Tinoco et al. [54] also proposed that the difference between PW2 and other Trp-rich AMPS in terms of the spectrum of activity (i.e., the ability of PW2 to inhibit protozoa and fungi but not bacteria) may be due to the distribution of positive charges over the length of the peptide. PW2 has two basic residues (Lys-4 and Arg-9) close to the aromatic region (WW), while indolicidin and tritrpticin have such residues nearer to the termini. The folding of PW2 seems to be similar to that of other Trp-rich AMPs and is attributed to the multiple Trps, forming a common motif, with one Trp being part of a hydrophobic core with other hydrophobic residues such as Pro and other Trps being on the surface and likely involved in membrane-binding [54].
5. Trp-rich AMPs with anticancer activity
A number of CAPs have been shown to exhibit cytotoxicity to a variety of cancer cells but not towards normal mammalian cells. The cell membranes of malignant cells are often negatively charged, unlike the zwitterionic membranes of normal cells, which may explain the selectivity. The negative charges are thought to come from the increased expression of the membrane phospholipid phosphotidylyserine (PS) and the glycoproteins, O-glycosylated mucines [55]. The anticancer activity of CAPs is thought to follow a mechanism similar to antimicrobial activity, i.e., membrane permeabilisation. Once inside the cells, a likely target is the negatively charged mitochondrial membrane; mitochondrial disruption is believed to induce apoptosis in cancer cells [56]. Such CAPs thus make an exciting new class of anticancer agents as, for cells to develop resistance to these, they would need to change their cellular and mitochondrial membrane structure. The main Trp-rich CAP to be studied for anticancer activities is the bovine lactoferricin (LFB). LFB was shown to bind to the negatively charged cancer cell membranes and disrupts them by formation of pores, allowing the peptide access to the mitochondria and inducing apoptosis [58]. Analogues of LFB (LFB14-31) (PEWFKCRRWQWRMKKLGA) were developed to elucidate the sequence and structural features essential for anticancer activity and compare to those responsible for its antimicrobial activities [27, 59]. An Ala scan experiment on LFB14-31 had found replacement of Cys6, Gln10 and Gly17 with the more hydrophobic residue Ala (peptide L2; PAWFKARRWAWRMKKLAA) increased anticancer activity against murine fibrosarcoma (methA) and human mammary carcinoma (MT-1) cell lines [27], possibly due to its higher hydrophobicity, suggesting this feature is important to anticancer activity as well as antimicrobial activity. LFB assumes an α-helical structure when bound to membranes. An analogue, designated L5 (PAWRKAFRWAWRMLKKAA), a showed significant increases in activity against MethA and MT-1 cells [27]. This peptide has all six positively charged residues in one sector of the α-helix, and this conformation may allow optimal interaction with the negatively charged cell membrane. Replacement of Trp3 of LFB14-31 with Ala resulted in a decrease in anti-tumour activity but increase in selectively for cancer cells over normal cells; however, replacement of Trp9 or Trp11 led to a significant loss of cytotoxic activity without improvement in selectivity [27]. Similar observations were made regarding the relevance of certain Trp residues to the activities of the related peptide LFB17–31 [4] and Tritrpticin [24] (see above). Peptide length was also found to be important, a length of 14 residues being optimal for high anti-tumour activity [59]. Recently, the bovine lactoferricin derivative, LTX-302 (WKKWDipKKWK-NH2, where ip is a bulky non-coded residue β-diphenylalanine) showed activity against A20 lymphoma by direct tumour cell lysis. Berge et al. [57] suggested that LTX-302 could induce long-term, specific immunity against the A20 lymphoma and confer a novel strategy for vaccination against cancer.
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
1399
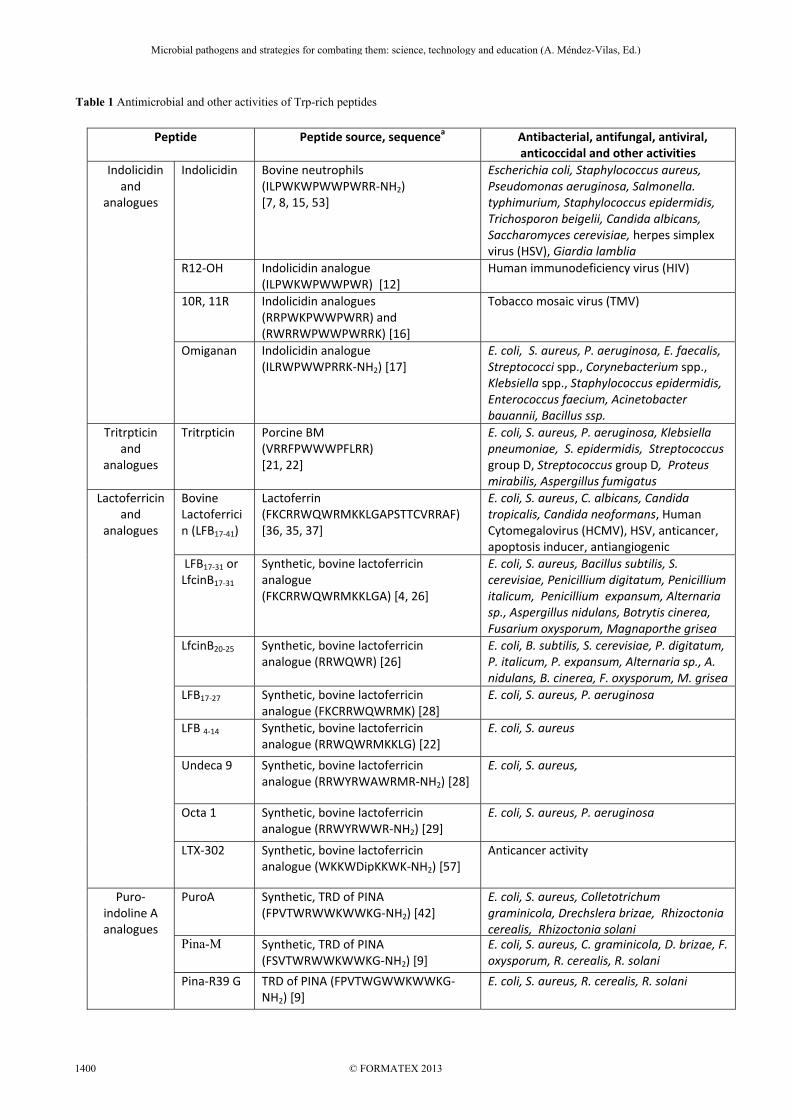
Table 1 Antimicrobial and other activities of Trp-rich peptides
Peptide Peptide source, sequencea Antibacterial, antifungal, antiviral,
anticoccidal and other activities Indolicidin
and analogues
Indolicidin Bovine neutrophils (ILPWKWPWWPWRR-NH2) [7, 8, 15, 53]
Escherichia coli, Staphylococcus aureus, Pseudomonas aeruginosa, Salmonella. typhimurium, Staphylococcus epidermidis, Trichosporon beigelii, Candida albicans, Saccharomyces cerevisiae, herpes simplex virus (HSV), Giardia lamblia
R12-OH Indolicidin analogue (ILPWKWPWWPWR) [12]
Human immunodeficiency virus (HIV)
10R, 11R Indolicidin analogues (RRPWKPWWPWRR) and (RWRRWPWWPWRRK) [16]
Tobacco mosaic virus (TMV)
Omiganan Indolicidin analogue (ILRWPWWPRRK-NH2) [17]
E. coli, S. aureus, P. aeruginosa, E. faecalis, Streptococci spp., Corynebacterium spp., Klebsiella spp., Staphylococcus epidermidis, Enterococcus faecium, Acinetobacter bauannii, Bacillus ssp.
Tritrpticin and
analogues
Tritrpticin Porcine BM (VRRFPWWWPFLRR) [21, 22]
E. coli, S. aureus, P. aeruginosa, Klebsiella pneumoniae, S. epidermidis, Streptococcus group D, Streptococcus group D, Proteus mirabilis, Aspergillus fumigatus
Lactoferricin and
analogues
Bovine Lactoferricin (LFB17-41)
Lactoferrin (FKCRRWQWRMKKLGAPSTTCVRRAF) [36, 35, 37]
E. coli, S. aureus, C. albicans, Candida tropicalis, Candida neoformans, Human Cytomegalovirus (HCMV), HSV, anticancer, apoptosis inducer, antiangiogenic
LFB17-31 or LfcinB17-31
Synthetic, bovine lactoferricin analogue (FKCRRWQWRMKKLGA) [4, 26]
E. coli, S. aureus, Bacillus subtilis, S. cerevisiae, Penicillium digitatum, Penicillium italicum, Penicillium expansum, Alternaria sp., Aspergillus nidulans, Botrytis cinerea, Fusarium oxysporum, Magnaporthe grisea
LfcinB20-25 Synthetic, bovine lactoferricin analogue (RRWQWR) [26]
E. coli, B. subtilis, S. cerevisiae, P. digitatum, P. italicum, P. expansum, Alternaria sp., A. nidulans, B. cinerea, F. oxysporum, M. grisea
LFB17-27
Synthetic, bovine lactoferricin analogue (FKCRRWQWRMK) [28]
E. coli, S. aureus, P. aeruginosa
LFB 4-14 Synthetic, bovine lactoferricin analogue (RRWQWRMKKLG) [22]
E. coli, S. aureus
Undeca 9 Synthetic, bovine lactoferricin analogue (RRWYRWAWRMR-NH2) [28]
E. coli, S. aureus,
Octa 1
Synthetic, bovine lactoferricin analogue (RRWYRWWR-NH2) [29]
E. coli, S. aureus, P. aeruginosa
LTX-302 Synthetic, bovine lactoferricin analogue (WKKWDipKKWK-NH2) [57]
Anticancer activity
Puro- indoline A analogues
PuroA
Synthetic, TRD of PINA(FPVTWRWWKWWKG-NH2) [42]
E. coli, S. aureus, Colletotrichum graminicola, Drechslera brizae, Rhizoctonia cerealis, Rhizoctonia solani
Pina-M
Synthetic, TRD of PINA (FSVTWRWWKWWKG-NH2) [9]
E. coli, S. aureus, C. graminicola, D. brizae, F. oxysporum, R. cerealis, R. solani
Pina-R39 G
TRD of PINA (FPVTWGWWKWWKG-NH2) [9]
E. coli, S. aureus, R. cerealis, R. solani
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
1400
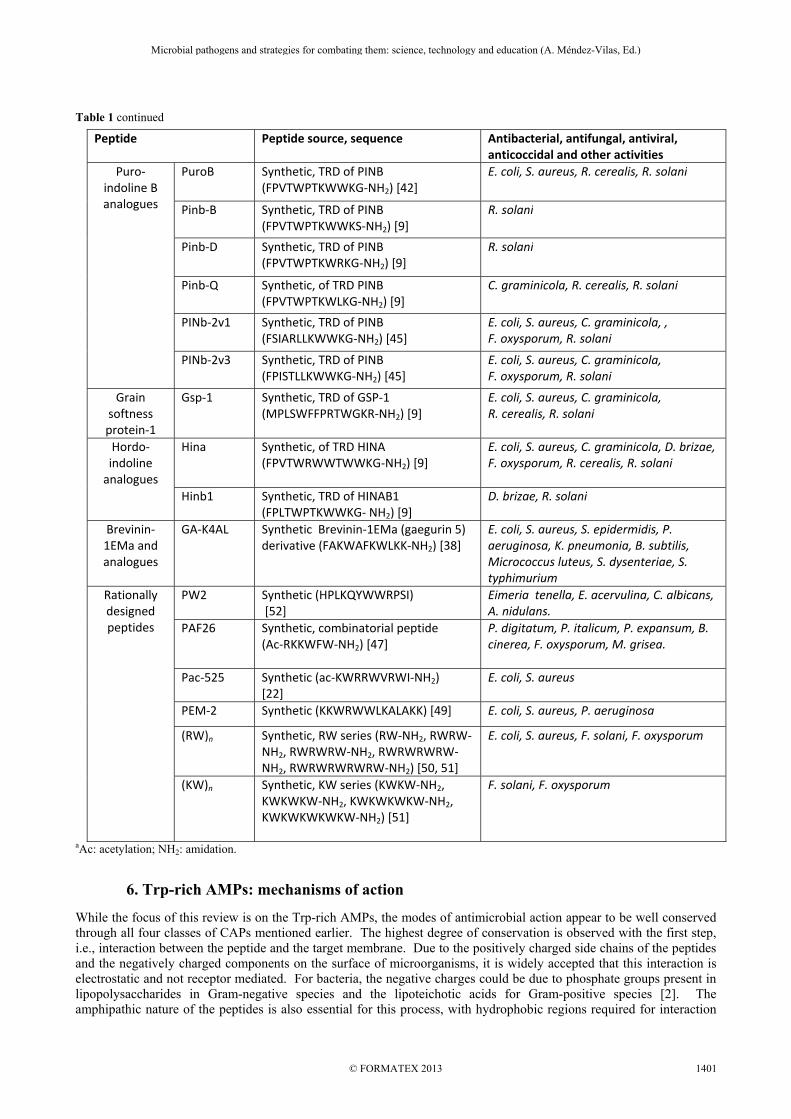
Table 1 continued
Peptide Peptide source, sequence Antibacterial, antifungal, antiviral, anticoccidal and other activities
Puro- indoline B analogues
PuroB
Synthetic, TRD of PINB (FPVTWPTKWWKG-NH2) [42]
E. coli, S. aureus, R. cerealis, R. solani
Pinb-B
Synthetic, TRD of PINB (FPVTWPTKWWKS-NH2) [9]
R. solani
Pinb-D
Synthetic, TRD of PINB (FPVTWPTKWRKG-NH2) [9]
R. solani
Pinb-Q
Synthetic, of TRD PINB (FPVTWPTKWLKG-NH2) [9]
C. graminicola, R. cerealis, R. solani
PINb-2v1
Synthetic, TRD of PINB (FSIARLLKWWKG-NH2) [45]
E. coli, S. aureus, C. graminicola, , F. oxysporum, R. solani
PINb-2v3 Synthetic, TRD of PINB (FPISTLLKWWKG-NH2) [45]
E. coli, S. aureus, C. graminicola, F. oxysporum, R. solani
Grain softness protein-1
Gsp-1 Synthetic, TRD of GSP-1 (MPLSWFFPRTWGKR-NH2) [9]
E. coli, S. aureus, C. graminicola, R. cerealis, R. solani
Hordo-indoline
analogues
Hina
Synthetic, of TRD HINA (FPVTWRWWTWWKG-NH2) [9]
E. coli, S. aureus, C. graminicola, D. brizae, F. oxysporum, R. cerealis, R. solani
Hinb1 Synthetic, TRD of HINAB1(FPLTWPTKWWKG- NH2) [9]
D. brizae, R. solani
Brevinin-1EMa and analogues
GA-K4AL Synthetic Brevinin-1EMa (gaegurin 5) derivative (FAKWAFKWLKK-NH2) [38]
E. coli, S. aureus, S. epidermidis, P. aeruginosa, K. pneumonia, B. subtilis, Micrococcus luteus, S. dysenteriae, S. typhimurium
Rationally designed peptides
PW2 Synthetic (HPLKQYWWRPSI) [52]
Eimeria tenella, E. acervulina, C. albicans, A. nidulans.
PAF26 Synthetic, combinatorial peptide (Ac-RKKWFW-NH2) [47]
P. digitatum, P. italicum, P. expansum, B. cinerea, F. oxysporum, M. grisea.
Pac-525 Synthetic (ac-KWRRWVRWI-NH2) [22]
E. coli, S. aureus
PEM-2 Synthetic (KKWRWWLKALAKK) [49] E. coli, S. aureus, P. aeruginosa
(RW)n Synthetic, RW series (RW-NH2, RWRW-NH2, RWRWRW-NH2, RWRWRWRW-NH2, RWRWRWRWRW-NH2) [50, 51]
E. coli, S. aureus, F. solani, F. oxysporum
(KW)n Synthetic, KW series (KWKW-NH2, KWKWKW-NH2, KWKWKWKW-NH2, KWKWKWKWKW-NH2) [51]
F. solani, F. oxysporum
aAc: acetylation; NH2: amidation.
6. Trp-rich AMPs: mechanisms of action
While the focus of this review is on the Trp-rich AMPs, the modes of antimicrobial action appear to be well conserved through all four classes of CAPs mentioned earlier. The highest degree of conservation is observed with the first step, i.e., interaction between the peptide and the target membrane. Due to the positively charged side chains of the peptides and the negatively charged components on the surface of microorganisms, it is widely accepted that this interaction is electrostatic and not receptor mediated. For bacteria, the negative charges could be due to phosphate groups present in lipopolysaccharides in Gram-negative species and the lipoteichotic acids for Gram-positive species [2]. The amphipathic nature of the peptides is also essential for this process, with hydrophobic regions required for interaction
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
1401

with membrane lipids, while hydrophilic regions associate with phospholipid headgroups. Following the initial interaction, several models have been proposed for the mechanisms by which AMPs/CAPs may permeabilise cell membranes. The leading models are; barrel-stave, toroidal pore, carpet-like (detergent-like), and aggregate formation [2, 60]. The ‘barrel-stave’ model (Fig. 1) is a proposed mechanism for α-helical peptides and involves four main steps: binding of the α-helical peptide monomers by hydrophobic interactions to the membrane; this can be due to charged phospholipids or zwitterionic membranes [61]; self-association of peptide monomers into bundles at the surface; insertion of peptide bundles of at least 2 monomers into the membrane to form a trans-membrane pore (the peptides must be at least 22 residues long for it to be able to transverse the bilayer); and finally, the expansion of the pore by recruitment of more monomers at the surface. Peptides that perturb membranes in this way typically exhibit a low minimum inhibitory concentration (MIC), as only a few pores are required to affect membrane integrity [62]. Indolicidin (detailed earlier) and a few CAPs that also have anticancer activities perturb membranes by this mechanism. Studies using model membranes show that indolicidin aggregates can form single channels in membranes, causing loss of cytoplasmic integrity [7].
A) Toroidal Pore B) Barrel-stave pore
C) Carpet
Fig. 1 Proposed mechanisms of action of antimicrobial peptides. The peptides are shown as cylinders, where the hydrophilic regions of the peptide are shaded dark grey and the hydrophobic regions are light grey. The phospholipids of the membrane bilayer are shown as hydrophilic heads (spheres) and hydrophobic tails. The ‘toroidal pore’ model (A) proposes that the peptides insert perpendicular to the plane of the bilayer, with the membrane curving inward. In the ‘barrel-stave’ model (B), the peptides insert in a perpendicular orientation to the plane of the bilayer, forming the ‘staves’ in a barrel-shaped cluster. The ‘carpet’ model (C) proposes that peptides aggregate parallel to the lipid bilayer, coating local areas in a “carpet”-like fashion. At a given threshold concentration, this is thought to result in a detergent-like activity, causing formation of micelle and membrane pores (Fig. based on [60]). The ‘toroidal pore’ is an alternative model for α-helical peptides and proposes that the peptide monomers aggregate at the surface and insert in a vertical orientation, and the membrane also curves inwards so that it lines the newly formed pore [2]. The barrel-stave pores are solely made of bundles of helical peptide monomers, while in toroidal pores, the peptides intercalate between the lipids, resulting in a pore that continuously bends from top to bottom [63]. Matsuzaki [64] proposed that this could occur due to strong ionic bonds forming between the anionic head groups on the phospholipids and the positively charged side chains of peptides, which may explain the curvature strain on the membrane as the peptides bind, resulting in pore formation. The ‘carpet-like’ (or ‘detergent-like’) model (Fig. 1) is the proposed mechanism for CAPs that have a specific membrane target range, e.g., microbial membranes and not the neutral/zwitterionic membranes of mammalian cells. The carpet-like mechanism model fits many CAPs, as it does not require the peptides to be of a particular length or sequence. Four major steps characterize the mechanism: the peptides bind via their positively charged basic residues to the negatively charged head groups of the cell membrane to cover it like a carpet; the hydrophobic regions of the peptide align with lipids in the hydrophobic core and the hydrophilic regions align with the phospholipid groups; more peptides bind until a threshold is reached, at which point there is induction of curvature in the membrane, which disintegrates and forms lipid micelle; and lastly, the micelle cause the formation of ‘toroidal’ pores. These allow
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
1402

passage of ions and low molecular weight compounds, ultimately resulting in cell lysis [62]. There some is debate over how predominant this model is; while Shai [62] suggest that the carpet-like model can be applied to many CAPs, Jenssen et al. [2] hypothesized that many CAPs may exhibit detergent (carpet-like) characteristics at high concentrations only due to the amphipathic nature of the peptides, and that there is little in vivo evidence that supports this mechanism occurring at minimum inhibitory concentration (MIC) values of a peptide against a microorganism. Jenssen et al. [2] propose an alternative ‘aggregate’ model, combining the concepts from the ‘carpet-like’ and ‘toroidal’ pore models. This less structured model explains membrane permeabilisation by pores followed by peptide translocation into the cytoplasm. It is proposed that peptides bond at the membrane surface, then insert into and span the membrane in an aggregate of lipid and peptide molecules called ‘micelle-like’ complexes. The result is the formation of channels of various sizes. The aggregates can also collapse, translocating the peptides across the lipid bilayer into the cytoplasm where they can impart intracellular effects [2]. Many studies on AMPs focus on their ability to attack cell membranes, leading to cell death by lysis; however, a several AMPs seem to (also) exhibit intracellular mechanisms leading to cell death. The intracellular targets include DNA, RNA, protein or cell wall synthesis and enzymatic activity [60]. Indolicidin has been observed to permeabilise membranes of E. coli, but not result in lysis [7]. It was found to be able to affect DNA synthesis [65], and it was proposed that it uses its membrane-binding ability to enter cells and affect targets within the cytoplasm, leading to antimicrobial effects. Further investigations into the DNA-binding action of indolicidin suggest a two-stage model, involving initial electrostatic binding of the positively charged amino acid residues to the phosphate groups of the DNA duplex, then insertion into the DNA groove [66].
7. Conclusions
In light of the ever-increasing emergence of antibiotic-resistant pathogens, antimicrobial peptides have great potential as novel antibiotics with very limited possibility of emergence of resistant microorganisms. They offer a great potential for wide spectrum activity, or alternatively, specificity against bacteria, fungi, viruses and/or protozoa of concern to human health, and some also with potential for cancer therapy. Peptides derived from natural sources also make them attractive agents for combating infectious agents transmitted in foods.
References
[1] Hancock REW, Diamond G. The role of cationic antimicrobial peptides in innate host defences. Trends on Microbiology. 2000;8:402-410.
[2] Jenssen H, Hamill P, Hancock REW. Peptide antimicrobial agents. Clinical Microbiology Reviews. 2006;19:491-511. [3] Schibli D, Epand R, Vogel H, Epand R. Tryptophan-rich antimicrobial peptides: comparative properties and membrane
interactions, Biochemistry and Cell Biology. 2002:80:667-677 [4] Strøm MB, Rekdal Ø, Svendsen JS. Antibacterial activity of 15-residue lactoferricin derivatives. Journal of Peptide Research.
2000;56:265-274. [5] Chan DI, Prenner EJ, Vogel HJ. Trytophan- and arginine-rich antimicrobial peptides: structures and mechanisms of action.
Biochimica et Biophysica Acta. 2006;1758:1184-1202 [6] Selsted ME, Novotny MJ, Morris WJ, Tang Y-Q, Smith W, Cullor JS. Indolicidin, a novel bactericidal tridecapeptide amide
from neutrophils. The Journal of Biological Chemistry. 1992;267:4292-4295. [7] Falla TJ, Karunaratne DN, Hancock REW. Mode of action of the antimicrobial peptide indolicidin. The Journal of Biological
Chemistry. 1996;271:19298-19303. [8] Lee DG, Kim HK, Kim SA, Park, SC, Jang SH, Hahm, KM. Fungicidial effect of indolicidin and its interaction with
phospholipid membranes. Biochemical and Biophysical Research Communications. 2003;350:350-310. [9] Phillips RA, Palombo EA, Panozzo JF, Bhave M. Puroindolines, Pin alleles, hordoindolines and grain softness proteins are
sources of bactericidal and fungicidal peptides. Journal of Cereal Science. 2011;53:112-7. [10] Dennison SR, Harris F, Bhatt T, Singh J, Phoenix DA, 2009. The effect of c-terminal amidation on the efficacy and selectivity
of antimicrobial and anticancer peptides. Molecular and Cellular Biochemistry 332, 43-50. [11] Mataraci E, Dosler S. In vitro activities of antibiotics and antimicrobial cationic peptides alone and in combination against
methicillin-resistant Staphylococcus aureus. Antimicrobial Agents and Chemotherapy. 2012;56:6366-6371. [12] Robinson WE, Mcdougall B, Tran D, Selsted ME. Anti-HIV-1 activity of indolicidin, an antimicrobial peptide from
neutrophils. Journal of Leukocyte Biology. 1998;63:94-100. [13] Krajewski K, Long Y-Q, Marchand C, Pommier Y, Roller PP. Design and synthesis of dimeric HIV-1 integrase inhibitory
peptides. Bioorganic & Medicinal Chemistry Letters. 2003;13:3203-5. [14] Krajewski K, Marchand C, Long Y-Q, Pommier Y, Roller PP. Synthesis and HIV-1 integrase inhibitory activity of dimeric and
tetrameric analogs of indolicidin. Bioorganic and Medicinal Chemistry Letters. 2004;14:5595-5598. [15] Albiol Matanic VC, Castilla V. Antiviral activity of antimicrobial cationic peptides against Junin virus and herpes simplex
virus. International Journal of Antimicrobial Agents. 2004;23:382-389. [16] Bhargava A, Osusky M, Hancock RE, Forward BS, Kay WW, Misra S. Antiviral indolicidin variant peptides: Evaluation for
broad-spectrum disease resistance in transgenic Nicotiana tabacum. Plant Science. 2007;172:515-523.
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
1403

[17] Sader HS, Fedler KA, Rennie RP, Stevens S, Jones RN. Omiganan pentahydrochloride (MBI 226), a topical 12-amino-acid cationic peptide: spectrum of antimicrobial activity and measurements of bactericidal activity. Antimicrobial Agents and Chemotherapy. 2004;48:3112-3118.
[18] Rubinchik E, Dugourd D, Algara T, Pasetka C, Friedland H. Antimicrobial and antifungal activities of a novel cationic antimicrobial peptide, omiganan, in experimental skin colonisation models, International Journal Of Antimicrobial Agents. 2009;34:457-461.
[19] Study of omiganan 1% gel in preventing catheter infections/colonization in patients with Central venous catheters: identifier NCT00231153. Clinical Trials: A service of the U.S National Institutes of Health. Available at www.clinicaltrials.gov. Accessed April 24, 2012.
[20] Study of antimicrobial activity of omiganan 1% gel vs. chlorhexidine 2% for topical skin antisepsis in healthy adult subjects: identifier NCT00608959. Clinical Trials: A service of the U.S National Institutes of Health. Available at www.clinicaltrials.gov. Accessed April 24, 2012.
[21] Lawyer C, Pai S, Watabe M, Borgia P, Mashimo T, Eagleton L, Watabe K. Antimicrobial activity of a 13 amino acid tryptophan-rich peptide derived from a putative porcine precursor protein of a novel family of antibacterial peptides. FEBS Letters. 1996;390:95-98.
[22] Wei SY, Wu JM, Kuo YY, Chen HL, Yip BS, Tzeng SR, Cheng JW. Solution structure of a novel tryptophan-rich peptide with bidirectional antimicrobial activity. Journal of Bacteriology. 2006;188:328-334.
[23] Nagpal S, Gupta V, Kaur KJ, Salunke DM. Structure-Function Analysis of Tritrypticin, an Antibacterial Peptide of Innate Immune Origin. Journal of Biological Chemistry. 1999;274:23296-23304.
[24] Schibli DJ, Nguyen LT, Kernaghan SD, Rekdal Ø, Vogel HJ. Structure-function analysis of tritrpticin analogs: potential relationships between antimicrobial activities, model membrane interactions, and their micelle-bound NMR structures. Biophysical Journal. 2006;91:4413-4426.
[25] Tomita M, Bellamy W, Takase M, Yamauchi K, Wakabayashi H, Kawase K. Potent antibacterial peptides generated by pepsin digestion of bovine lactoferrin. Journal of Dairy Science. 1991;74:4137-4142.
[26] Muńoz A, Marcoz JF. Activity and mode of action against fungal phytopathogens of bovine lactoferricin-derived peptides. Journal of Applied Microbiology. 2006;101:1199-1207.
[27] Yang N, Stensen W, Svendsen JS, Rekdal Ø. Enhanced antitumor activity and selectivity of lactoferrin-derived peptides. Journal of Peptide Research. 2002;60:187-197.
[28] Strøm MB, Rekdal Ø, Svendsen JS. The effects of charge and lipophilicity on the antibacterial activity of undecapeptides derived from bovine lactoferricin. Journal of Peptide Science, 2002a;7:36-43.
[29] Strøm MB, Rekdal Ø, Svendsen JS. Antimicrobial activity of short arginine- and tryptophan-rich peptides. Journal of Peptide Science. 2002b;8:431-437.
[30] Hasegawa ., Motsuchi W, Tanaka S, Dosako S. Inhibition with lactoferrin of in vitro infection with human herpes virus. Japanese Journal of Medical Science and Biology. 1994;47:73–85.
[31] Ikeda M, Sugiyama K, Tanaka T, Tanaka K, Sekihara H, Shimotohno K, Kato, N. Lactoferrin markedly inhibits hepatitis C virus infection in cultured human hepa-tocytes. Biochemical and Biophysical Research Communications. 1998;245:549-553.
[32] Harmsen M, Swart, P, Béthune M, Pauwels R, Clercq E, The T, Meijer D. Antiviral Effects of Plasma and Milk Proteins: Lactoferrin Shows Potent Activity against Both Human Immunodeficiency Virus and Human Cytomegalovirus Replication In Vitro, The Journal Of Infectious Diseases. 1995;172:380-383
[33] Van der Strate BWA, Beljaars L, Molema G, Harmsen MC, Meijer DKF. Antiviral activities of lactoferrin. Antiviral Research. 2001;52:225-239
[34] Hammer J, Haaheim H, Gutteberg TJ. Bovine lactoferrin is more efficient than bovine lactoferricin in inhibiting HSV-I/II replication in vitro. In: Shimazaki, K. (Ed.), Lactoferrin: Structure, Functions and Application. Amsterdam: Elsevier Science; 2000:239-243.
[35] Andersen JH, Osbakk SA, Vorland LH, Traavik T, Gutteberg TJ. Lactoferrin and cyclic lactoferricin inhibit the entry of human cytomegalovirus into human fibroblasts. Antiviral Research. 2001;51:141-149.
[36] Vorland L, Ulvatne H, Andersen J, Haukland H, Rekdal Ø, Svendsen J, and Gutteberg T. Lactoferricin of Bovine Origin is More Active than Lactoferricins of Human, Murine and Caprine Origin. Scandinavian Journal Of Infectious Diseases. 1998;30:513-517
[37] Jenssen H, Andersen JH, Uhlin-Hansen L, Gutteberg TJ, Rekdal Ø. Anti-HSV activity of lactoferricin analogues is only partly related to their affinity for heparan sulfate. Antiviral Research. 2004;61:101-109.
[38] Won HS, Kang SJ, Choi WS, Lee BJ. Activity Optimization of an undecapeptide analogue derived from a frog-skin antimicrobial peptide. Molecules and Cells. 2011;31:49-54.
[39] Gautier MF, Aleman ME, Guirao A, Marion D, Joudrier P. Triticum aestivum puroindolines, two basic cystine-rich seed proteins: cDNA sequence analysis and developmental gene expression. Plant Molecular Biology. 1994;25:43-57.
[40] Bhave M, Morris CF. Molecular genetics of puroindolines and related genes: allelic diversity in wheat and other grasses. Plant Molecular Biology. 2008;66:205-219.
[41] Dubreil L, Gaborit T, Bouchet B, Gallant DJ, Broekaert WF, Quillien L, Marion D. Spatial and temporal distribution of the major isoforms of puroindolines (puroindoline-a and puroindoline-b) and non specific lipid transfer protein (ns-LTP1e1) of Triticum aestivum seeds. Relationships with their in vitro antifungal properties. Plant Science. 1998;138:121-135.
[42] Jing W, Demcoe AR, Vogel HJ. Conformation of a bactericidal domain of puroindoline a: structure and mechanism of action of a 13-residue antimicrobial peptide. Journal of Bacteriology. 2003;185:4938-4947.
[43] Capparelli R, Palumbo D, Iannaccone M, Ventimglia I, Di Salle E, Capuano F, Salvatore P, Amoroso MG. Cloning and expression of two plant proteins: similar antimicrobial activity of native and recombinant form. Biotechnology Letters. 2006;28:943-949.
[44] Capparelli R, Ventimiglia I, Palumbo D, Nicodemo D, Salvatore P, Amoroso MG, Iannaccone M. Expression of recombinant puroindolines for the treatment of staphylococcal skin infections (acne vulgaris). Journal of Biotechnology. 2007;128:606-614.
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
1404

[45] Ramalingam A, Palombo EA, Bhave M. The Pinb-2 genes in wheat comprise a multigene family with great sequence diversity and important variants. Journal of Cereal Science. 2012;56:171-180
[46] Fjell CD, Hiss JA, Hancock REW, Schneider G. Designing antimicrobial peptides: Form follows function. Nature Reviews Drug Discovery. 2012;11:37-51.
[47] López-García B, Perez-Paya E, Marcos, JF. Identification of novel hexapeptides bioactive against phytopathogenic fungi through screening of a synthetic peptide combinatorial library. Applied Environmental Microbiology. 2002;68:2453-2460.
[48] Muñoz A, López-García B, Marcos JF. Comparative study of antimicrobial peptides to control citrus postharvest decay caused by Penicillium digitatum. Journal of Agricultural Food Chemistry. 2007;55:8170-8176.
[49] Yu HY, Huang KC, Yip BS, Tu CH, Chen HL, Cheng HT, Cheng JW. Rational design of tryptophan-rich peptides with enhanced antimicrobial activities and specificities. ChemBioChem. 2010;11:2273-2282.
[50] Liu Z, Brady A, Young A, Rasimick B, Chen K, Zhou C, Kallenbach NR. Length effects in antimicrobial peptides of the (RW)n series. Antimicrobial Agents and Chemotherapy. 2007;51:597-603.
[51] Gopal R, Hyungjong N, Seo CH, Park Y. Antifungal activity of (KW)n or (RW)n peptide against Fusarium solani and Fusarium oxysporum. International Journal of Molecular Sciences. 2012;13:15042-15053.
[52] Da Silva A, Kawazoe U, Freitas FFT, Gatti MSV, Dolder H, Schumacher RI, Juliano MA, Da Silva MJ, Leite A. Avian anticoccidial activity of a novel membrane-interactive peptide selected from phage display libraries. Molecular and Biochemical Parasitology. 2002;120:53-60.
[53] Aley SB, Zimmerman M, Hetsko M, Selsted ME, Gillin FD. Killing of Giardia lamblia by cryptdins and cationic neutrophil peptides. Infection and Immunity. 1994;62:5397-5403.
[54] Tinoco LW, Da Silva A, Leite A, Valente AP. NMR structure of PW2 bound to SDS micelles. The Journal of Biological Chemistry. 2002;277:36351-36356.
[55] Papo N, Shai Y. Host defence peptides as new weapons in cancer treatment. Cellular and Molecular Life Sciences. 2005;62:784-790.
[56] Hoskin DW, Ramamoorthy A. Studies on anticancer activities of antimicrobial peptides. Biochimica et Biophysica Acta. 2008;1778:357-375.
[57] Berge G, Eliassen L, Camilio K, Bartnes K, Sveinbjørnsson B, Rekdal Ø. Therapeutic vaccination against a murine lymphoma by intratumoural injection of a cationic anticancer peptide. Cancer Immunology, Immunotherapy. 2010;59:1285-1294
[58] Yoo YC, Watanabe R, Koike Y, Mitobe M, Shimazaki KI, Watanabe S, Azuma I. Apoptosis in human leukemic cells induced by lactoferricin, a bovine milk protein-derived peptide: involvement of reactive oxygen species. Biochemical and Biophysical Research Communications. 1997;237:624-628.
[59] Yang N, Strøm MB, Mekonnen SM, Svendsen JS, Rekdal Ø. The effects of shortening lactoferrin derived peptides against tumour cells, bacterial and normal human cells. Journal of Peptide Science. 2004;10:37-46.
[60] Brogden K. Antimicrobial peptides: pore formers or metabolic inhibitors in bacteria? Nature Reviews Microbiology. 2005;3:238-250.
[61] Oren Z, Shai Y. Mode of action of linear amphipathic α-helical antimicrobial peptides. Biopolymers. 1998;47:451-63. [62] Shai Y. Mode of action of membrane active antimicrobial peptides. Biopolymers. 2002;66:236-248. [63] Yang L, Harroun TA, Weiss TM, Ding L, Huang HW. Barrel-stave model or toroidal model? A case study on melittin pores.
Biophyical Journal. 2001;81:1475-1485 [64] Matsuzaki K. Why and how are peptide-lipid interactions utilized for self-defence? Magainins and tachyplesins as archetypes.
Biochimica et Biophysica Acta. 1999;1462:1-10. [65] Subbalakshmi C, Krishnakumari V, Nagaraj R, Sitaram N. Requirements for antibacterial and hemolytic activities in the bovine
neutrophil derived 13-residue peptide indolicidin. FEBS Letters. 1996;395:48-52. [66] Hsu CH, Chen C, Jou ML, Yueh-Luen Lee A, Lin YC, Yu YP, Huang WT, Wu SH. Structural and DNA-binding studies on the
bovine antimicrobial peptide, indolicidin: evidence for multiple conformations involved in binding to membranes and DNA. Nucleic Acids Research. 2005;33:4053-4064.
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
1405





























