Embed Size (px)
Citation preview

Full Terms & Conditions of access and use can be found athttp://www.tandfonline.com/action/journalInformation?journalCode=tbis20
Bird Study
ISSN: 0006-3657 (Print) 1944-6705 (Online) Journal homepage: http://www.tandfonline.com/loi/tbis20
Trends in breeding phenology across ten decadesshow varying adjustments to environmentalchanges in four wader species
Hans Meltofte, Ole Amstrup, Troels Leuenhagen Petersen, Frank Rigét &Anders P. Tøttrup
To cite this article: Hans Meltofte, Ole Amstrup, Troels Leuenhagen Petersen, Frank Rigét& Anders P. Tøttrup (2018) Trends in breeding phenology across ten decades show varyingadjustments to environmental changes in four wader species, Bird Study, 65:1, 44-51, DOI:10.1080/00063657.2018.1444014
To link to this article: https://doi.org/10.1080/00063657.2018.1444014
Published online: 20 Mar 2018.
Submit your article to this journal
Article views: 49
View related articles
View Crossmark data

Trends in breeding phenology across ten decades show varying adjustments toenvironmental changes in four wader speciesHans Meltoftea, Ole Amstrupb, Troels Leuenhagen Petersenc, Frank Rigéta and Anders P. Tøttrupc
aDepartment of Bioscience, Aarhus University, Roskilde, Denmark; bAmphi Consult, Aalborg, Denmark; cCenter for Macroecology, Evolution andClimate, University of Copenhagen, Copenhagen, Denmark
ABSTRACTCapsule: During 1928–2016, initiation of egg-laying advanced in two wader species, remainedunchanged in one, and was delayed in one species. The changes across years and variationamong species can be explained by climatic variables and differences in migratory strategies.Aims: To document possible changes in initiation of egg-laying in common Danish wader speciessince the early part of the 20th century and seek possible correlations between egg-laying, timing ofarrival and environmental factors.Methods: Annual records of the first eggs and chicks found on the scientific reserve of Tipperne inwestern Denmark 1928–2016 were analysed using linear regression to determine patterns in timingof egg-laying, pre-breeding length and influence of climate factors.Results: Two short/medium-distance migrant wader species, Northern Lapwing Vanellus vanellusand Common Redshank Tringa totanus advanced breeding initiation by about one week, withwinter North Atlantic oscillation Index and spring temperature as important predictors. Bycontrast, two long-distance migrants, Black-tailed Godwit Limosa limosa and Ruff Calidris pugnax,did not advance egg-laying, and Ruff actually delaying it. As a result, the pre-laying period wassignificantly prolonged in both Black-tailed Godwit (21 days) and Ruff (52 days), while there wasno significant change for Common Redshank.Conclusion: Long-distance migrants are able to adjust spring arrival but unlike short/medium-distance migrants, do not necessarily adjust breeding initiation.
ARTICLE HISTORYReceived 8 May 2017Accepted 19 February 2018
Much research is aimed at gaining a more thoroughunderstanding of responses to global change at alltrophic levels of ecosystems (Parmesan 2007, Jørgensenet al. 2016, de Keyzer et al. 2017). This includesadjustments in reproductive phenology, and whetherchange is related to the climatic variations and otherfactors likely to affect the reproductive cycle (Parmesan2007). Birds need to time breeding in order for thechicks to optimally exploit peak food abundance (Kamet al. 2004, Both et al. 2006, but also see Jonzén et al.2007), which in turn is affected by soil productivityand climate. Phenotypic plasticity in birds allows, tosome extent, quick adjustments of phenology (Berthold2001, Fiedler 2009, Tøttrup et al. 2010) and a growingnumber of studies find evidence for micro-evolutionaryadaptations to current climatic changes (Brandsmaet al. 2017, Källander et al. 2017).
Spatial and temporal responses to climate changediffer between species and across trophic levels,because some organisms rely on endogenous factorsthat are more or less independent of the environment
(Berthold 2001, Both & Visser 2001, Walther et al.2002, Both et al. 2006, Jørgensen et al. 2016). Changesin breeding phenology of sedentary organisms (e.g.some birds, most invertebrates and plants) arepresumably affected differently from those of migratoryanimals, because the resident species will experiencethe entire annual cycle in the ecosystem and hencehave the opportunity to adjust their physiologythroughout the year (Berthold 2001, Fiedler 2009). Toavoid disadvantages in interspecific competition, forexample for best breeding sites, migrants need toadequately adjust timing of arrival and breeding to theconditions at the breeding grounds; conditions thatthey are not able to predict during the winter. Both &Visser (2001) showed that inadequate changes inbreeding phenology in the Pied Flycatcher Ficedulahypoleuca, a long-distance migrant, caused localpopulation declines of up to 90% due to mistiming andcompetition with Great Tits Parus major. Suchmaladaptation might cause a micro-evolutionarybottleneck by strong selection for early egg-laying
© 2018 British Trust for Ornithology
CONTACT Hans Meltofte [email protected] Department of Bioscience, Aarhus University, Frederiksborgvej 399, DK-4000 Roskilde, Denmark; AndersP. Tøttrup [email protected] Center for Macroecology, Evolution and Climate, Natural History Museum of Denmark, Universitetsparken 15, DK-2100Copenhagen, Denmark
BIRD STUDY, 2018VOL. 65, NO. 1, 44–51https://doi.org/10.1080/00063657.2018.1444014

(Both & Visser 2001, Both et al. 2006, but also see Jonzénet al. 2007). However, studies on a nearby populationalso showed that advances in egg-laying actuallybenefitted recruitment of offspring into the population(Tomotani et al. 2017).
In a previous study, we demonstrated advancedarrival time at the breeding area for three of the waderspecies analysed in this study – Black-tailed GodwitLimosa limosa (13 days earlier), Common RedshankTringa totanus (14 days earlier) and Ruff Calidrispugnax (42 days earlier) – over an 80-year period(1929–2008; Petersen et al. 2012), whereas arrival ofNorthern Lapwing Vanellus vanellus could not beestablished because the species was also frequentlypresent during winter.
In this study we present an analysis of breedinginitiation observations based on an even longer timeperiod (1928–2016) for the same four wader species inthe same area. We include analyses of changes inlength of the pre-breeding period and assess howclimate impacts the timing of breeding. Our datarepresent the longest available time series on egg-layingphenology of birds in Denmark and, except for theNorthern Lapwing (Both et al. 2005), possibly inwaders worldwide.
Methods
The study was carried out at the scientific reserve ofTipperne located in the brackish lagoon of RingkøbingFjord in western Denmark (55°53′ N 8°13′ E). Thereserve includes 700 ha meadows and reed beds largelywithout changes in habitat management during thestudy period. For details of the environment and thebirds of Tipperne, see Meltofte (1987), Thorup (1998)and Meltofte & Clausen (2011, 2016).
During most of the study years, the NorthernLapwing population numbered around 100–275 pairs,the Black-tailed Godwit population 40–200 pairs, theRuff population 25–300 breeding females and theCommon Redshank population 50–800 pairs (Thorup1998 and in litt.). For all four species the breedingpopulations varied around relatively moderate levels upto the late 1970s, after which they increasedsignificantly to a peak in around 1990, followed by areturn to more or less previous levels from around 2000.
Egg-laying data (date of first egg laid) for the fourspecies were compiled from observers’ diaries andbreeding phenology forms kept at the reserve during theentire study period. We have solely used dates of firstcompleted clutch and explicit observations of the firstnest this year to calculate first egg dates (laying intervals:Northern Lapwing: 42 hours, Ruff: 24 hours, Black-
tailed Godwit: 24 hours, Common Redshank: 38 hours(Cramp et al. 1983)). Similarly, we have extracted allavailable data on the first hatched young from the samesources and converted these to first egg dates using 33days for laying and incubation in Northern Lapwing, 27days in Black-tailed Godwit and Ruff, and 29 days inCommon Redshank (Thorup 1998 and in litt.).
Data on first egg dates are potentially affected by arelatively high predation pressure early in the seasoncompared to just a few weeks later (Thorup 1998) andto the fact that it is more difficult to find the first nestthan to find 15 nests one week later. However, theconstant effort exerted in locating nests mitigates thepotential bias in changes over time in our data.
Individual phenological repeatability in birds remainspoorly understood (but see Lourenço et al. 2011, Thorupet al. 2013). As a proxy measure for the pre-laying periodat the population level, we used the difference in numberof days between the first arrival and first egg date (seeHögstedt 1974, who used female arrival). Date ofarrival and hence length of the pre-laying period couldnot be determined in Northern Lapwing because, asmentioned, this species occurred irregularly throughoutthe winter on Tipperne (Meltofte & Clausen 2016).
Climate data for the entire period were available atnational level only and were obtained from the onlinearchives of the Danish Meteorological Institute(Cappelen 2016). As an attempt to integrate springconditions both covering the general characteristics ofwestern Europe and the local phenology forinvertebrate food and vegetation growth, we correlatedfirst egg dates with three-month averaged meanmonthly temperature and precipitation for January–February–March (JFM) for the early breedingNorthern Lapwing and Black-tailed Godwit and forFebruary–March–April (FMA) for later breeding Ruffand Common Redshank. Furthermore, we tested for acorrelation with the North Atlantic Oscillation (NAO)Index for the winter period of December through toMarch (DJFM) (Hüppop & Hüppop 2003, Hurrell2016), which is the dominant mode of climatevariability in the North Atlantic region (IPCC 2010).
Determination of the wintering area of thepopulations occurring in Denmark was based onBønløkke et al. (2007) using estimates of springmigration distances to Tipperne from Google Earth(2010, ver. 4.3) and classifying <4000 km as short/medium-distance and >4000 km as long-distance. Thisclassified Northern Lapwing and Common Redshankas short/medium-distance migrants and Black-tailedGodwit and Ruff as long-distance migrants (Petersenet al. 2012), that are wintering in western Europe/Mediterranean and tropical Africa, respectively.
BIRD STUDY 45

For the statistical analyses all data were converted toJulian days (where 1 = 1 January). Ordinary linearregression was then used to determine species-specifictemporal patterns in timing of egg-laying and pre-breeding length. Prior to the regression analyses,possible outliers were identified and removed, if theyhad influence values higher than Cooks Distance(Weisberg 1985). In order to assess possible year-to-year correlations between first egg dates and date ofobservation of the first individuals, the potentiallyconfounding temporal effect on both variables wasremoved. This was done by regression analyses of bothvariables on year and then extracting the residualsfrom these two regressions. A Spearman correlationanalysis of these two sets of residuals was then used totest the relationship between year-detrended first eggdates and observation of first individuals.
Results
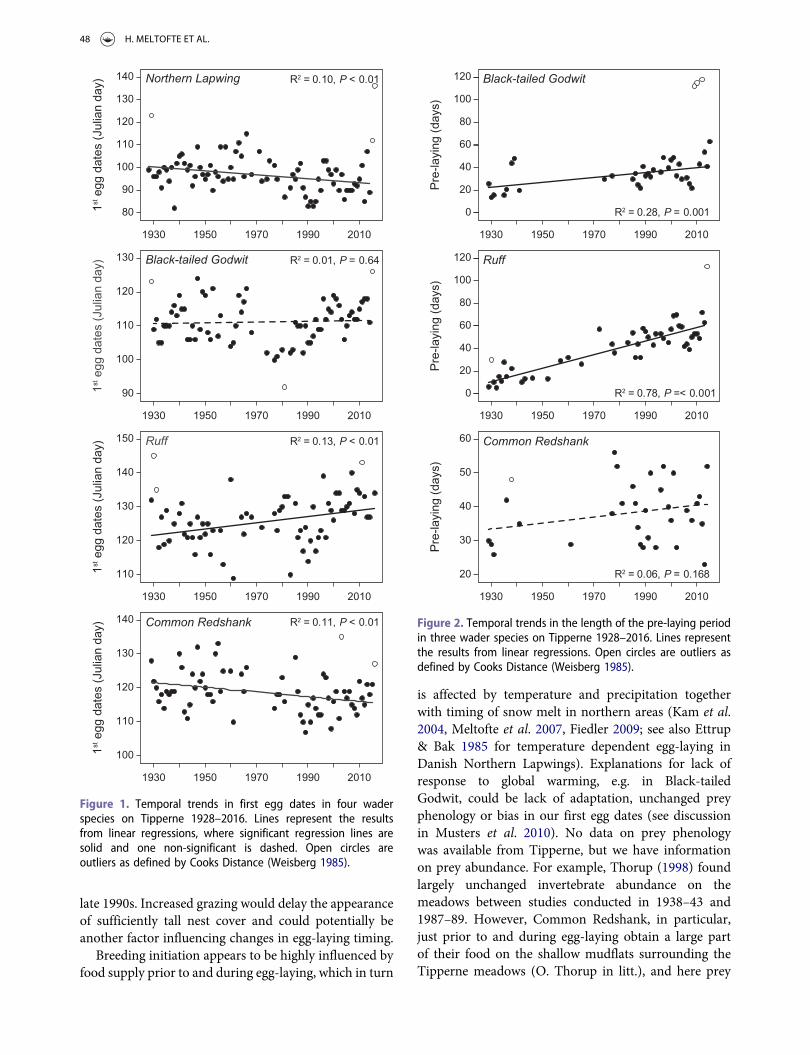
We found significant advances in first egg dates forNorthern Lapwing (−0.0858 days/year) and CommonRedshank (−0.0683 days/year) while a significant delaywas noted for Ruff (0.0908 days/year) (Table 1 andFigure 1). In total, the mean initiation of the earliestclutches advanced by 7.6 and 6.1 days, respectively,over the study period 1928–2016 for the two short/medium-distance migrants, Northern Lapwing (fromapproximately 10 April to 3 April) and CommonRedshank (from approximately 2 May to 26 April)during the 89-year study period. Initiation of egg-laying was delayed by 8.1 days in the long-distancemigrant, the Ruff (from approximately 2 May to 9May), and remained largely unchanged in the similarlylong-distance migrant, the Black-tailed Godwit (mean21 April; Figure 1).
Over the study period, the pre-laying period becamesignificantly longer in the long-distance migrants Ruff(0.596 days/year, P < 0.0001) and Black-tailed Godwit(0.214 days/year, P = 0.001) (Table 1 and Figure 2). InRuff, we found a longer pre-laying period in years withhigher spring temperatures (3.225 days/°C, P = 0.048)and more precipitation (0.448 days/mm, P = 0.039), butno effect of winter conditions (Table 1). We found nosignificant correlation between year-detrended first eggdates and first observation date of arrival for Black-tailed Godwit (P = 0.59) and Common Redshank (P =0.49), whereas such a correlation was found for Ruff(P = 0.001).
When changes in timing to the start of breeding werecompared with advances in arrival, the pre-laying periodwas prolonged by 21.1 days (from 22.3 to 43.4 days) inBlack-tailed Godwit and 52.4 days (from 9.5 to 61.9
days) in Ruff. The apparent prolonged pre-layingperiod of 7.7 days in Common Redshank was notstatistically significant (Table 1).
For all four species, timing of first egg was earlier insprings following mild winters (positive NAO values).Furthermore, breeding initiation was also earlier inyears with higher temperatures for Northern Lapwing,Black-tailed Godwit and Common Redshank. In Black-tailed Godwit and Common Redshank, we also foundthat breeding was earlier in years with moreprecipitation (Table 1).
Discussion
We demonstrate earlier initiation of egg-laying by oneweek for the two short/medium-distance migratorywaders, Northern Lapwing and Common Redshank,and unchanged or delayed initiation of egg-laying fortwo long-distance migrants, Black-tailed Godwit andRuff. The pre-laying period was consequentlyprolonged in the long-distance species while a smallexpansion in the short/medium-distance CommonRedshank was not statistically significant (we do nothave arrival data for Northern Lapwing). Overall,winter NAO and spring temperature were found to bethe most important predictors. This is in accordancewith our previous findings on spring arrival (Petersenet al. 2012); i.e. timing of arrival in waders can beexplained by local temperature and precipitation aswell as winter conditions (NAO, winter index) andspecies-specific migration distances.
Thorup (1998) found no change in Northern Lapwingand Common Redshank breeding phenology during1928–92 on Tipperne, but this is not necessarilycontradictory to our results because Thorup usedpopulation-wide breeding phenology, which may showa slower trend than analyses of first egg dates, where asingle nest counts. On the other hand, the data wepresent here are based on a time series that is 25 yearslonger than that of Thorup, and we consider first egg/chick dates to have been more consistently recorded onthe reserve than the population-wide laying phenology.Yet, as in our study, the long-distance migrant, Black-tailed Godwit, showed no change in breedingphenology in Thorup’s study (see also Kentie et al.2017), whereas breeding in Ruff was delayed accordingto a more recent analysis (Thorup 2016).
Given the marked advancement in its arrival, theunchanged breeding phenology in Black-tailed Godwitwas surprising (see below). Even more surprising wasdelayed egg-laying in Ruff, which Thorup (2016)ascribed to the likely effects of greatly increasedgrazing by Barnacle Geese Branta leucopsis since the
46 H. MELTOFTE ET AL.
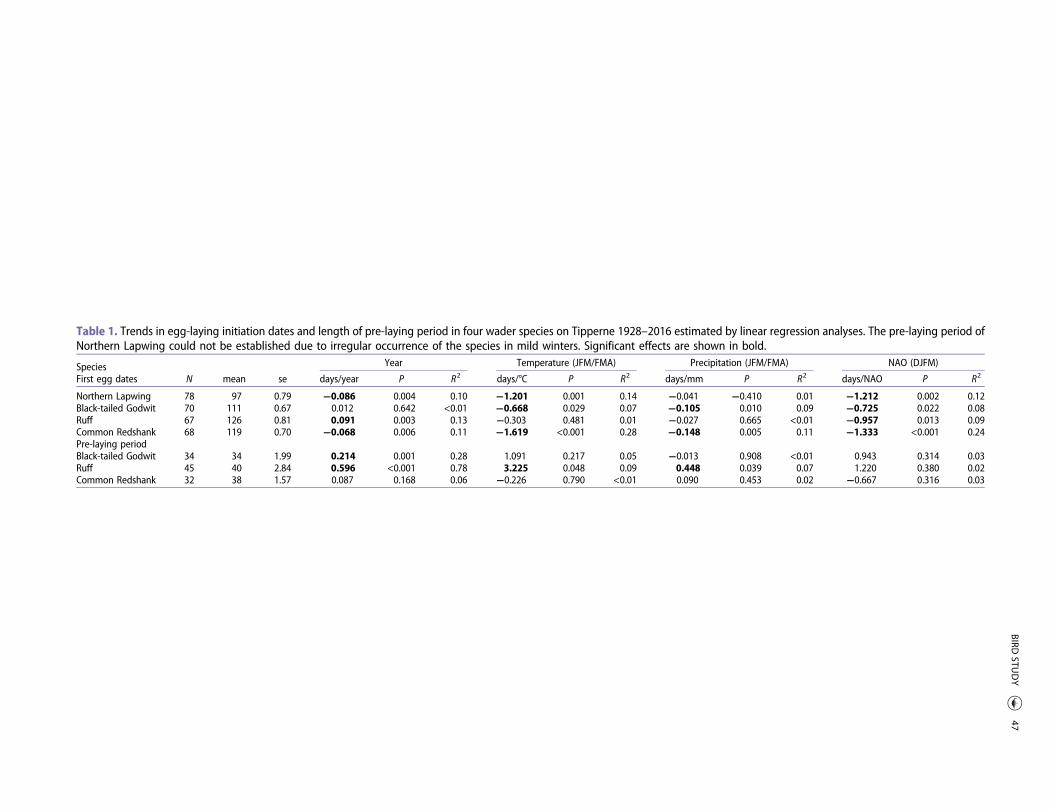
Table 1. Trends in egg-laying initiation dates and length of pre-laying period in four wader species on Tipperne 1928–2016 estimated by linear regression analyses. The pre-laying period ofNorthern Lapwing could not be established due to irregular occurrence of the species in mild winters. Significant effects are shown in bold.
SpeciesN mean se
Year Temperature (JFM/FMA) Precipitation (JFM/FMA) NAO (DJFM)
First egg dates days/year P R2 days/°C P R2 days/mm P R2 days/NAO P R2
Northern Lapwing 78 97 0.79 −0.086 0.004 0.10 −1.201 0.001 0.14 −0.041 −0.410 0.01 −1.212 0.002 0.12Black-tailed Godwit 70 111 0.67 0.012 0.642 <0.01 −0.668 0.029 0.07 −0.105 0.010 0.09 −0.725 0.022 0.08Ruff 67 126 0.81 0.091 0.003 0.13 −0.303 0.481 0.01 −0.027 0.665 <0.01 −0.957 0.013 0.09Common Redshank 68 119 0.70 −0.068 0.006 0.11 −1.619 <0.001 0.28 −0.148 0.005 0.11 −1.333 <0.001 0.24Pre-laying periodBlack-tailed Godwit 34 34 1.99 0.214 0.001 0.28 1.091 0.217 0.05 −0.013 0.908 <0.01 0.943 0.314 0.03Ruff 45 40 2.84 0.596 <0.001 0.78 3.225 0.048 0.09 0.448 0.039 0.07 1.220 0.380 0.02Common Redshank 32 38 1.57 0.087 0.168 0.06 −0.226 0.790 <0.01 0.090 0.453 0.02 −0.667 0.316 0.03
BIRDSTU
DY
47
late 1990s. Increased grazing would delay the appearanceof sufficiently tall nest cover and could potentially beanother factor influencing changes in egg-laying timing.
Breeding initiation appears to be highly influenced byfood supply prior to and during egg-laying, which in turn
is affected by temperature and precipitation togetherwith timing of snow melt in northern areas (Kam et al.2004, Meltofte et al. 2007, Fiedler 2009; see also Ettrup& Bak 1985 for temperature dependent egg-laying inDanish Northern Lapwings). Explanations for lack ofresponse to global warming, e.g. in Black-tailedGodwit, could be lack of adaptation, unchanged preyphenology or bias in our first egg dates (see discussionin Musters et al. 2010). No data on prey phenologywas available from Tipperne, but we have informationon prey abundance. For example, Thorup (1998) foundlargely unchanged invertebrate abundance on themeadows between studies conducted in 1938–43 and1987–89. However, Common Redshank, in particular,just prior to and during egg-laying obtain a large partof their food on the shallow mudflats surrounding theTipperne meadows (O. Thorup in litt.), and here prey
1st e
gg d
ates
(Ju
lian
day)
80
90
100
110
120
130
140
20101930 1950 1970 1990
1st e
gg d
ates
(Ju
lian
day)
90
100
110
120
130
20101930 1950 1970 1990
1st e
gg d
ates
(Ju
lian
day)
110
120
130
140
150
20101930 1950 1970 1990
1st e
gg d
ates
(Ju
lian
day)
100
110
120
130
140
20101930 1950 1970 1990
Black-tailed Godwit
Northern Lapwing
Ruff
Common Redshank
R2 = 0.10, P < 0.01
R2 = 0.01, P = 0.64
R2 = 0.13, P < 0.01
R2 = 0.11, P < 0.01
Figure 1. Temporal trends in first egg dates in four waderspecies on Tipperne 1928–2016. Lines represent the resultsfrom linear regressions, where significant regression lines aresolid and one non-significant is dashed. Open circles areoutliers as defined by Cooks Distance (Weisberg 1985).
Pre
-layi
ng (
days
)
20101930 1950 1970 1990
Pre
-layi
ng (
days
)
20101930 1950 1970 1990
Pre
-layi
ng (
days
)
20
30
40
50
60
20101930 1950 1970 1990
Black-tailed Godwit
Ruff
Common Redshank
R2 = 0.28, P = 0.001
R2 = 0.78, P =< 0.001
R2 = 0.06, P = 0.168
0
20
80
60
40
100
120
0
20
80
60
40
100
120
Figure 2. Temporal trends in the length of the pre-laying periodin three wader species on Tipperne 1928–2016. Lines representthe results from linear regressions. Open circles are outliers asdefined by Cooks Distance (Weisberg 1985).
48 H. MELTOFTE ET AL.

densities have increased considerably during the studyperiod (Meltofte & Clausen 2016). Predation pressuremight counteract earlier egg-laying because predationis most intense early in the breeding season (Thorup1998). On the other hand, advanced breeding initiationincreases the opportunity for more replacementclutches due to the longer breeding season.
In a Dutch study on meadow birds, Beintema et al.(1985) found that in the 1980s breeding phenology hadalready advanced by 2–3 weeks in all six wader speciesstudied, including the four species from this study.Spring temperature was discarded as the cause, and thephenological changes were attributed to intensifiedagricultural management, especially fertilizer applicationleading to soil fauna phenology advancement. Since thenegg-laying phenology in meadowbirds has been studiedextensively in the Netherlands with the aim to unravelthe impact of climate versus changes in habitatmanagement, e.g. in form of fertilizers, soil humidity,timing of mowing, etc. (Both et al. 2005, Kleijn et al.2010, Musters et al. 2010, Schroeder et al. 2012, Senneret al. 2015, Brandsma et al. 2017). The results show amixed picture with varying influence of both agriculturalpractices and climate variables in the individual speciesand periods of time. On Tipperne, effects of habitatmanagement can largely be excluded becausemanagement of the meadows has been restricted tograzing (from May at the earliest) and late summermowing (from July at the earliest), while fertilizers havenot been used (Thorup 1998). However, in theNetherlands a similar difference was found between egg-laying advancement during the last half century in theshort-distance migrant, Northern Lapwing, and nochange in the long-distance migrant, Black-tailed Godwit(Musters et al. 2010). Also in accordance with our results,Both et al. (2005) found that advancement of egg-layingin Northern Lapwing apparently began before the middleof the 20th century and has accelerated since then.
Gill et al. (2014) suggest that earlier breeding could bethe mechanism leading to earlier arrival in a populationof short-distance migrant Icelandic Black-tailed GodwitsLimosa l. islandica. This is driven by earlier born youngbeing able to select better wintering habitats and therebyarrive earlier in Iceland at the time they start breeding.Our data do not include individual data to further studythis mechanism. Still, for our short/medium-distancespecies, this could potentially be an explanation to studyfurther.
Unlike the long-distance migratory waders covered inthis study, the long-distance migrant, Pied Flycatcher,has shortened the pre-laying period on their Dutchbreeding grounds (Both & Visser 2001). The PiedFlycatcher breeds earlier now, but not early enough
compared to prey phenology. As shown by Both &Visser (2001), a pre-laying period which was at theminimum required, plus insufficient advancement ofarrival, hampered adequate advancement of egg-layingwhich subsequently led to severe population declines inthis species. The two short/medium-distance migrants,Northern Lapwing and Common Redshank, do laytheir eggs earlier and thus have an advantage ininterspecific competition. They gain more time forreplacement clutches and possibly are better able tosynchronize hatching with an advanced peak ininvertebrate abundance on the meadows, assuming thatprey phenology on Tipperne has also advanced.
Högstedt (1974) showed a strong negative correlationbetween the length of the pre-laying period and preyabundance in a Swedish Northern Lapwing populationin a study based on female arrivals and first egg dates.Högstedt suggested a minimum pre-laying period ofabout 13 days. However, given the increasinglysedentary behaviour of Northern Lapwing onTipperne, a minimum pre-laying period could hardlyhave been a limiting factor for Lapwing breedingphenology in the reserve in recent decades. We do notknow to which degree our breeders are sedentary, sothe year-round presence of the species makes furtherexploration impossible within this current data.
Thorup (2004) addressed different possible causes ofdeclining Ruff and Black-tailed Godwit populations inDenmark, and stressed that climate change is unlikely tobe the cause because Danish temperatures have not risenabove that of the former French, Hungarian andUkrainian breeding sites of these species. Risingtemperatures should nevertheless not be neglected as afuture cause of population decline as a result of a sloweradaptation to the changed breeding ground phenology(e.g. prolonged pre-laying periods) compared to that ofsedentary and short-distance migratory waders (thisstudy).
Invertebrate biomass varies between years at Tipperne(Thorup 1998) and abundance is especially low after asevere winter followed by a dry spring (Larsen 1949).Hence, less frequent winter frosts and increasedprecipitation (IPCC 2010) predict good conditions forinvertebrates. Warmer winters might be especiallyadvantageous for meadow breeders, because Black-tailed Godwit chick survival was higher with moderatetemperatures and precipitation (Groen & Hemerik2002). Higher temperatures are believed to beespecially favourable for Northern Lapwing because thechicks are much less dependent on brooding in periodswith higher temperatures (Beintema & Visser 1989).However, a potentially negative effect of warmersprings is that grass may by higher and denser at an
BIRD STUDY 49

earlier time, which could make it difficult for the birds tocapture their invertebrate prey.
Our results show that long-distance migrants mayadjust timing of spring arrival, but breeding initiation isnot necessarily adjusted, as it is in short/medium-distance migrants. This indicates that records of springarrival alone may not be sufficient to assess climatechange impacts on timing of breeding. Understandingthe link between timing of spring arrival and breedinginitiation time is essential if conservation or managementactions are to be implemented.
Acknowledgements
We thank Preben Clausen, Aarhus University, and Ole Thorup,Amphi Consult, for providing help and supplementary data andother information on Tipperne. Nick Quist Nathaniels kindlychecked our English, and two anonymous referees providedhighly valued suggestions for improvement of the manuscript.
Funding
APT thanks the Aage V Jensen Foundation for support and theDanish National Research Foundation for supporting theCenter for Macroecology, Evolution and Climate [grantnumber DNRF96].
References
Beintema, A.J. & Visser, G.H. 1989. The effects of weather ontime budgets and development of chicks of meadow birds.Ardea 77: 181–192.
Beintema, A.J., Beintema-Hietbrink, R.J. & Müskens,G.D.J.M. 1985. A shift in the timing of breeding inmeadow birds. Ardea 73: 83–89.
Berthold, P. 2001. Bird Migration. A General Survey, 2nd ed.Oxford University Press, Oxford.
Bønløkke, J., Madsen, J.J., Thorup, K., Pedersen, K.T.,Bjerrum, M. & Rahbek, C. 2007. The Danish BirdMigration Atlas. Rhodos A/S (in Danish, with Englishsummary).
Both, C. & Visser, M.E. 2001. Adjustment to climate change isconstrained by arrival date in a long-distance migrant bird.Nature 411: 296–298.
Both, C., Piersma, T. & Roodbergen, S.P. 2005. Climaticchange explains much of the 20th century advance inlaying date of Northern Lapwing Vanellus vanellus in TheNetherlands. Ardea 93: 79–88.
Both, C., Bouwhuis, S., Lessells, C.M. & Visser, M.E. 2006.Climate change and population declines in a long-distancemigratory bird. Nature 441: 81–83.
Brandsma, O.H., Kentie, R. & Piersma, T. 2017. Why didLapwings Vanellus vanellus in managed habitat advanceegg laying during a period without warming early springs?Ardea 105: 19–26.
Cappelen, J. (ed.) 2016. Denmark – DMI Historical ClimateData Collection 1768-2015. DMI Report No. 16-02.
Cramp, S., Simmons, K.E. & Ferguson-Lees, J. 1983.Handbook of the Birds of the Europe, the Middle East andNorth Africa. The Birds of the Western Palearctic. Vol. 3.Oxford University Press, Oxford.
de Keyzer, C.W., Rafferty, N.E., Inouye, D.W. & Thomson,J.D. 2017. Confounding effects of spatial variation on shiftsin phenology. Glob. Change Biol. 23: 1783–1791.
Ettrup, H. & Bak, B. 1985. Breeding season, clutch size andyoung production of Danish Lapwings Vanellus vanellus.Dansk Orn. Foren. Tidsskr. 79: 43–55 (in Danish, withEnglish summary).
Fiedler, W. 2009. Bird ecology as an indicator of climate andglobal change. In Letcher, T.M. (ed) Climate Change:observed impacts on planet earth, Ch. 9: 181–195. ElsevierB.V., Amsterdam.
Gill, J.A., Alves, J.A., Sutherland, W.J., Appleton, G.F.,Potts, P.M. & Gunnarsson, T.G. 2014. Why is timing ofbird migration advancing when individuals are not?P. Roy. Soc. B 281: 20132161.
Google Earth 2010. 4.3.7284.3916 (beta), online:kh.google.com (02-03-2010).
Groen, N.M. & Hemerik, L. 2002. Reproductive success andsurvival of Black-tailed Godwits Limosa limosa in adeclining local population in the Netherlands. Ardea 90:239–248.
Högstedt, G. 1974. Length of the pre-laying period in theLapwing Vanellus vanellus L. in relation to its foodresources. Ornis Scand 5: 1–4.
Hüppop, O. & Hüppop, K. 2003. North Atlantic Oscillationand timing of the spring migration in birds. Proc. R. Soc.Lond. B 279: 233–240.
Hurrell, J. 2016. North Atlantic Oscillation (NAO) Index(station-based). https://climatedataguide.ucar.edu/climate-data/hurrell-north-atlantic-oscillation-nao-index-station-based (13 May 2016).
IPCC 2010. Intergovernmental Panel on Climate Change.http://www.ipcc.ch/pdf/technical-papers/climate-changes-biodiversity-en.pdf (23-03-2010).
Jonzén, N., Hedenström, A. & Lundberg, P. 2007. Climatechange and the optimal arrival of migratory birds.Proc. R. Soc. Lond. B 274: 269–274.
Jørgensen, P.S., Bohning-Gaese, K., Thorup, K., Tøttrup,A.P., Chylarecki, P., Jiguet, F., Lehikoinen, A., Noble,D.G., Reif, J., Schmid, H., van Turnhout, C., Burfield,I.J., Foppen, R., Vořišek, P., van Strien, A., Gregory,R.D. & Rahbek, R. 2016. Continental-scale global changeattribution in European birds – combining annual anddecadal time scales. Glob. Change Biol. 22: 530–543.
Källander, H., Hasselquist, D., Hedenström, A., Nord, A.,Smith, H.G. & Nilsson, J.-Å. 2017. Variation in layingdate in relation to spring temperature in three species oftits (Paridae) and pied flycatchers Ficedula hypoleuca insouthernmost Sweden. J. Avian Biol 48: 83–90.
Kam, J. van de, Ens, B., Piersna, T. & Zwarts, L. 2004.Shorebirds. An Illustrated Behavioural Ecology. KNNVPublishers, Zeist.
Kentie, R., Marquez-Ferrando, R., Figuerola, J., Gangoso,L., Hooijmeijer, J.C.E.W., Loonstra, A.H.J., Robin, F.,Sarasa, M., Senner, N., Valkema, H., Verhoeven, M.A.& Piersma, T. 2017. Does wintering north or south of theSahara correlate with timing and breeding performance inblack-tailed godwits? Ecol. Evol. 7: 2812–2820.
50 H. MELTOFTE ET AL.

Kleijn, D., Schekkerman, H., Dimmers, W.J., van Kats,R.J.M., Melman, D. & Teunissen, W.A. 2010. Adverseeffects of agricultural intensification and climate changeon breeding habitat quality of Black-tailed GodwitsLimosa l. limosa in the Netherlands. Ibis 152: 475–486.
Larsen, E.B. 1949. The influence of the severe winters of 1939-1942 on the soil fauna of Tipperne. Oikos 1: 184–207.
Lourenço, P.M., Kentie, R., Schroeder, J., Groen, N.M.,Hooijmeijer, J.C.E.W. & Piersma, T. 2011. Repeatabletiming of northward departure, arrival and breeding inBlack-tailed Godwits Limosa l. limosa, but no dominoeffects. J. Ornithol. 152: 1023–1032.
Meltofte, H. 1987. The occurrence of staging waders Charadriiat the Tipperne reserve, western Denmark, 1928-1982.Dansk Orn. Foren. Tidsskr 81: 1–108 (in Danish, withEnglish summary).
Meltofte, H. & Clausen, P. 2011. The occurrence of swans,ducks, Coot and Great Crested Grebe in the TipperneReserve 1929-2007 in relation to environmental conditionsin the brackish lagoon, Ringkøbing Fjord, Denmark. DanskOrn. Foren. Tidsskr. 105: 1–120 (In Danish, with extensiveEnglish summary).
Meltofte, H. & Clausen, P. 2016. Trends in staging waders onthe Tipperne Reserve, western Denmark, 1929-2014 with acritical review of trends in flyway populations. Dansk Orn.Foren. Tidsskr. 110: 1–72.
Meltofte, H., Høye, T.T., Schmidt, N.M. & Forchhammer,M.C. 2007. Differences in food abundance cause inter-annual variation in the breeding phenology of High Arcticwaders. Polar Biol. 30: 601–606.
Musters, C.J.M., ter Keurs, W.J. & de Snoo, G.R. 2010.Timing of the breeding season of Black-tailed GodwitLimosa limosa and Northern Lapwing Vanellus vanellus inThe Netherlands. Ardea 98: 195–202.
Parmesan, C. 2007. Influences of species, latitudes andmethodologies on estimates of phenological response toglobal warming. Glob. Change Biol. 13:1860–1872.
Petersen, T.L., Meltofte, H. & Tøttrup, A.P. 2012.Advanced spring arrival of avian migrants on Tipperne,western Denmark, during 1929-2008. Dansk Orn. Foren.Tidsskr. 106: 65–72.
Schroeder, J., Piersma, T., Groen, N.M., Hooijmeijer,J.C.E.W., Kentie, R., Lourenco, P.M., Schekkerman, H.& Both, C. 2012. Reproductive timing and investment inrelation to spring warming and advancing agriculturalschedules. J. Ornithol. 153: 327–336.
Senner, N.R., Verhoeven, M.A., Hooijmeijer, J.C.E.W. &Piersma, T. 2015: Just when you thought you knew it all:New evidence for flexible breeding patterns inContinental Black-tailed Godwits. Wader Study 122:18–24.
Thorup, O. 1998. The breeding birds on Tipperne 1928-1992.Dansk Orn. Foren. Tidsskr 92: 1–192 (in Danish, withextensive English summary).
Thorup, O. 2004. Status of populations and management ofDunlin Calidris alpina, Ruff Philomachus pugnax andBlack-tailed Godwit Limosa limosa in Denmark. DanskOrn. Foren. Tidsskr. 98: 7–20.
Thorup, O. 2016. Timing of breeding in Ruff Philomachuspugnax: a crucial parameter for management and useof wet grassland in Western Europe. Wader Study 123:49–58.
Thorup, K., Vardanis, Y., Tøttrup, A.P., Kristensen, M.W.& Alerstam, T. 2013. Timing of songbird migration:individual consistency within and between seasons.J. Avian Biol. 44: 486–494.
Tomotani, B.M., van der Jeugd, H., Gienapp, P., De LaHera, I., Pilzecker, J., Teichmann, C. & Visser, M.E.2017. Climate change leads to differential shifts in thetiming of annual cycle stages in a migratory bird. Glob.Change Biol. DOI:10.1111/gcb.14006.
Tøttrup, A.P., Rainio, K., Coppack, T., Lehikoinen, E.,Rahbek, C. & Thorup, K. 2010. Local temperature fine-tunes the timing of spring migration in birds. Integr.Comp. Biol. 50: 293–304.
Walther, G., Post, E., Convey, P., Menzel, A., Parmesan, C.,Beebee, T.J.C., Fromentin, J., Hoegh-Guldberg, O. &Bairlein, F. 2002. Ecological responses to recent climatechange. Nature 416: 389–395.
Weisberg, S. 1985. Applied Linear Regression. 4th ed. Wiley,New York, NY.
BIRD STUDY 51





























