Embed Size (px)
Citation preview

Journal of Neurology, Neurosurgery, and Psychiatry, 1972, 35, 52-59
Transmission block in terminal nerve twigs:a single fibre electromyographic finding in man
ERIK STALBERG AND BARBARA THIELE
From the Department of Clinical Neurophysiology,Academic Hospital, Uppsala, Sweden
SUMMARY Single fibre electromyography has been performed in patients with partial nervelesions, amyotrophic lateral sclerosis, progressive spinal muscle atrophy, muscular dystrophy, anddistal hereditary myopathy. The recorded action potentials were often more complex than in thenormal muscle due to increased fibre density in the motor unit and the individual spike componentsshowed a large jitter and occasional blockings. Sometimes two or more spikes in a complex dis-appeared and reappeared simultaneously upon successive discharges. This phenomenon, called'paired blocking', has been further investigated. The jitter of the blocking potentials in relation tothe rest of the complex was large, up to 500 ttsec. The degree of blocking increased with increasinginnervation frequency until it eventually proceeded to total block during continuous activity. Some-times a slight effect on blocking was seen after edrophonium. This type of block is probably localizedin the terminal nerve twigs, perhaps in newly formed sprouts. The phenomenon of neurogenicblocking may contribute to the fatigue clinically experienced in different denervation-reinnervationcases.
With a multielectrode technique described byEkstedt and Stalberg (1963), Ekstedt (1964), andStalberg (1966), the electrical activity was re-corded from voluntarily activated human singlemuscle fibres. Normally the muscle fibres,belonging to one motor unit, are separated fromeach other so that an electrode of the type usedin this investigation, 25 ,u in diameter, recordsactivity from only one muscle fibre in a certainmotor unit in about 700% of the insertions. Theuptake radius of the electrode is about 200 ,u. Inabout 3000 of the insertions two or, exception-ally, more muscle fibres belonging to the samemotor unit can be recorded. The action poten-tials from different fibres are in these casesalways coupled to each other with a smallvariability in the interpotential interval from dis-charge to discharge-the jitter (Ekstedt, 1964;Stalberg, Ekstedt, and Broman, 1971). In thenormal muscle there is never any blocking of oneor the other potential in such a potential pair.However, in myasthenia gravis, for example, thejitter is increased and occasional blockings ofone or the other of the potentials are seen, morefrequent in patients with pronounced symptoms(Ekstedt and St'alberg, 1967; Blom and Ringqvist,
52
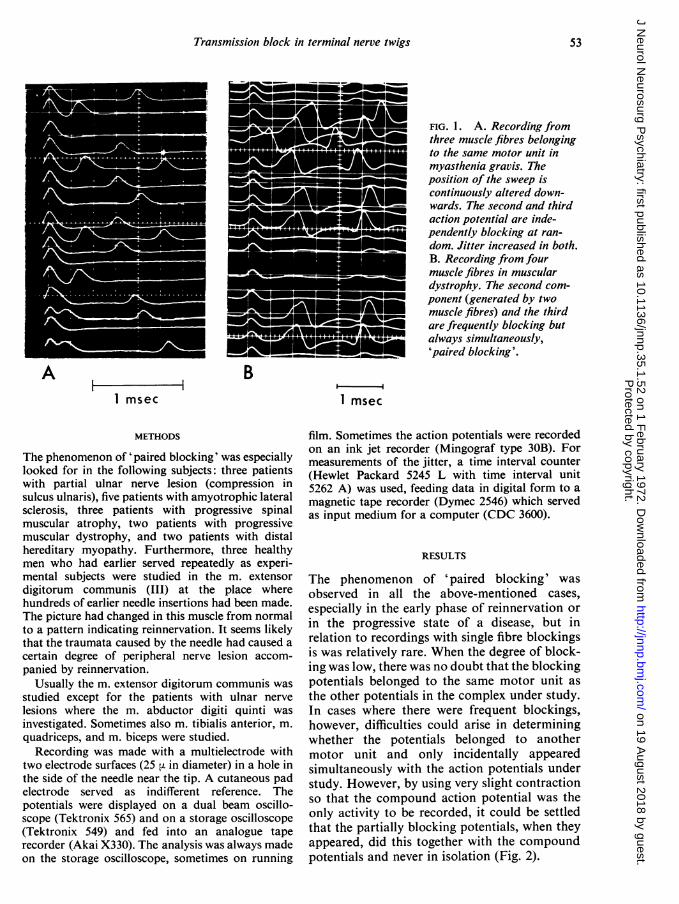
1971). If activity from three fibres from onemotor unit happens to be recorded in a myas-thenic muscle, it can be seen that the blockingappears independently in the different fibres(Fig. la).
In different pathological conditions, especiallyin lower motor neurone disorders, there is anincreased fibre density in the motor unit due toperipheral sprouting (Wohlfart, 1957; Kugelberg,Edstrom, and Abbruzzese, 1970) and thereforethe activity from many muscle fibres belongingto one motor unit can be recorded from thesame electrode. The number of simultaneouslyrecorded fibres in the same motor unit is typicallybetween three and 10. In such a complex thejitter between individual muscle fibre potentialsis increased and blockings are seen.
During the study of these cases with singlefibre electromyography it was sometimes seenthat in the compound action potentials two ormore blocking spike components were intimatelycoupled and none of these spikes blocked inde-pendently of the other(s). From now on this willbe called 'paired blocking' (Fig. 1 B). Thisinvestigation is aimed at studying the nature ofthis kind of blocking.
Protected by copyright.
on 19 August 2018 by guest.
http://jnnp.bmj.com
/J N
eurol Neurosurg P
sychiatry: first published as 10.1136/jnnp.35.1.52 on 1 February 1972. D
ownloaded from
Transmission block in terminal nerve twigs
FIG. 1. A. Recording fromthree muscle fibres belongingto the same motor unit inmyasthenia gravis. Theposition of the sweep iscontinuously altered down-wards. The second and thirdaction potential are inde-pendently blocking at ran-dom. Jitter increased in both.B. Recording from fourmuscle fibres in musculardystrophy. The second com-ponent (generated by twomuscle fibres) and the thirdare frequently blocking butalways simultaneously,'paired blocking'.
A BI m 1 m
I msec 1 msec
METHODS
The phenomenon of 'paired blocking' was especiallylooked for in the following subjects: three patientswith partial ulnar nerve lesion (compression insulcus ulnaris), five patients with amyotrophic lateralsclerosis, three patients with progressive spinalmuscular atrophy, two patients with progressivemuscular dystrophy, and two patients with distalhereditary myopathy. Furthermore, three healthymen who had earlier served repeatedly as experi-mental subjects were studied in the m. extensordigitorum communis (III) at the place wherehundreds of earlier needle insertions had been made.The picture had changed in this muscle from normalto a pattern indicating reinnervation. It seems likelythat the traumata caused by the needle had caused a
certain degree of peripheral nerve lesion accom-
panied by reinnervation.Usually the m. extensor digitorum communis was
studied except for the patients with ulnar nerve
lesions where the m. abductor digiti quinti was
investigated. Sometimes also m. tibialis anterior, m.
quadriceps, and m. biceps were studied.Recording was made with a multielectrode with
two electrode surfaces (25 p. in diameter) in a hole inthe side of the needle near the tip. A cutaneous padelectrode served as indifferent reference. Thepotentials were displayed on a dual beam oscillo-scope (Tektronix 565) and on a storage oscilloscope(Tektronix 549) and fed into an analogue taperecorder (Akai X330). The analysis was always madeon the storage oscilloscope, sometimes on running
film. Sometimes the action potentials were recordedon an ink jet recorder (Mingograf type 30B). Formeasurements of the jitter, a time interval counter(Hewlet Packard 5245 L with time interval unit5262 A) was used, feeding data in digital form to amagnetic tape recorder (Dymec 2546) which servedas input medium for a computer (CDC 3600).
RESULTS
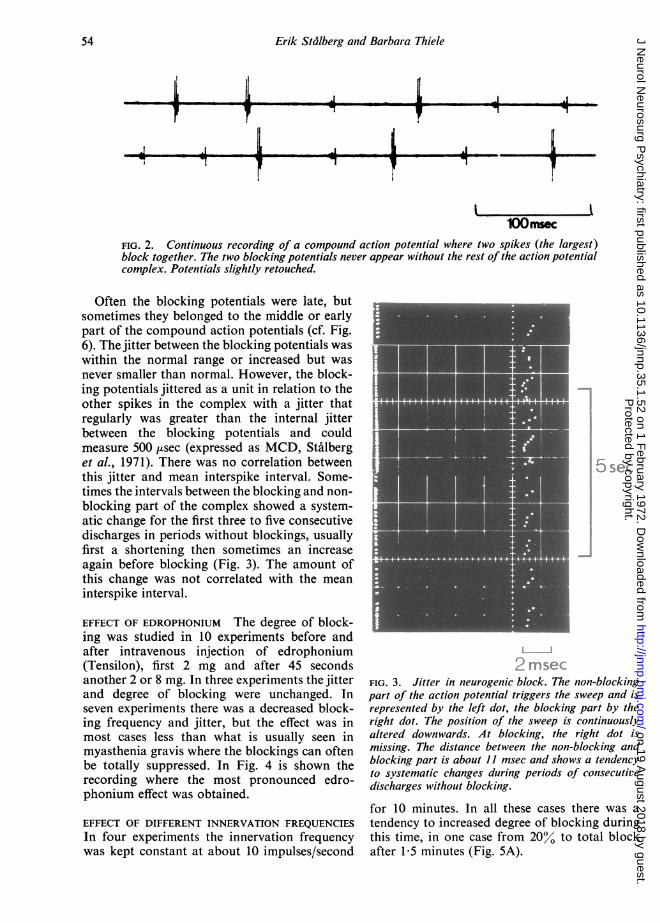
The phenomenon of 'paired blocking' wasobserved in all the above-mentioned cases,especially in the early phase of reinnervation orin the progressive state of a disease, but inrelation to recordings with single fibre blockingsis was relatively rare. When the degree of block-ing was low, there was no doubt that the blockingpotentials belonged to the same motor unit asthe other potentials in the complex under study.In cases where there were frequent blockings,however, difficulties could arise in determiningwhether the potentials belonged to anothermotor unit and only incidentally appearedsimultaneously with the action potentials understudy. However, by using very slight contractionso that the compound action potential was theonly activity to be recorded, it could be settledthat the partially blocking potentials, when theyappeared, did this together with the compoundpotentials and never in isolation (Fig. 2).
53P
rotected by copyright. on 19 A
ugust 2018 by guest.http://jnnp.bm
j.com/
J Neurol N
eurosurg Psychiatry: first published as 10.1136/jnnp.35.1.52 on 1 F
ebruary 1972. Dow
nloaded from
Erik Stalberg and Barbar-a Thiele
IL 11} I # §~~~~~-1 'I
,i , L i ,1 I'I -I,'I
IlOOmsec
FIG. 2. Continuous recording of a compound action potential where two spikes (the largest)block together. The two blocking potentials never appear without the rest of the action potentialcomplex. Potentials slightly retouched.
Often the blocking potentials were late, butsometimes they belonged to the middle or earlypart of the compound action potentials (cf. Fig.6). The jitter between the blocking potentials waswithin the normal range or increased but was
never smaller than normal. However, the block-ing potentials jittered as a unit in relation to theother spikes in the complex with a jitter thatregularly was greater than the internal jitterbetween the blocking potentials and couldmeasure 500 ,tsec (expressed as MCD, St'alberget al., 1971). There was no correlation betweenthis jitter and mean interspike interval. Some-times the intervals between the blocking and non-
blocking part of the complex showed a system-atic change for the first three to five consecutivedischarges in periods without blockings, usuallyfirst a shortening then sometimes an increaseagain before blocking (Fig. 3). The amount ofthis change was not correlated with the mean
interspike interval.
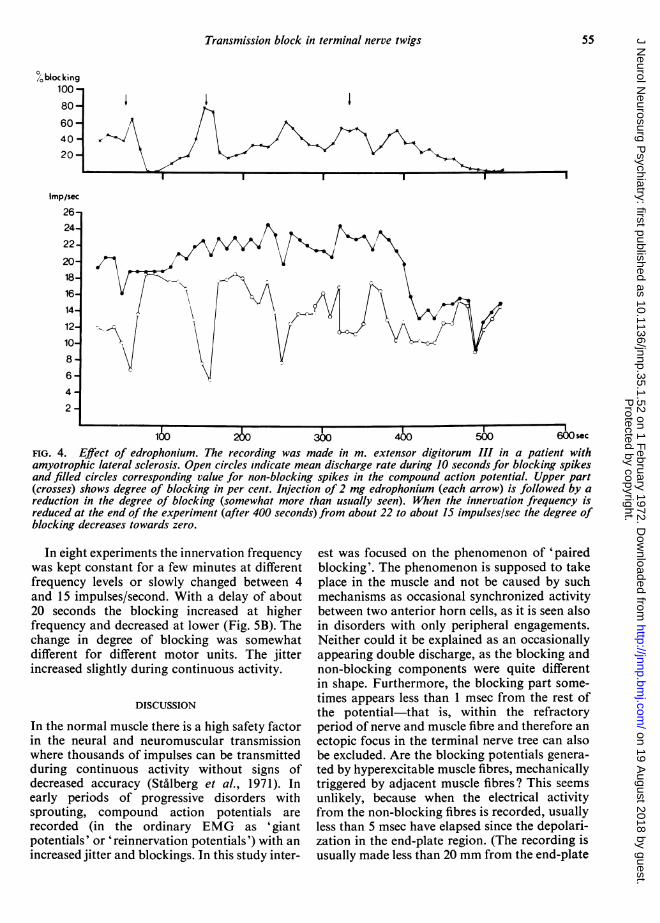
EFFECT OF EDROPHONIUM The degree of block-ing was studied in 10 experiments before andafter intravenous injection of edrophonium(Tensilon), first 2 mg and after 45 secondsanother 2 or 8 mg. In three experiments the jitterand degree of blocking were unchanged. Inseven experiments there was a decreased block-ing frequency and jitter, but the effect was inmost cases less than what is usually seen inmyasthenia gravis where the blockings can oftenbe totally suppressed. In Fig. 4 is shown therecording where the most pronounced edro-phonium effect was obtained.
EFFECT OF DIFFERENT INNERVATION FREQUENCIESIn four experiments the innervation frequencywas kept constant at about 10 impulses/second
FIG. 3. Jitter in neurogenic block. The non-blockingpart of the action potential triggers the sweep and isrepresented by the left dot, the blocking part by theright dot. The position of the sweep is continuouslyaltered downwards. At blocking, the right dot ismissing. The distance between the non-blocking andblocking part is about 11 msec and shows a tendencyto systematic changes during periods of consecutivedischarges without blocking.
for 10 minutes. In all these cases there was a
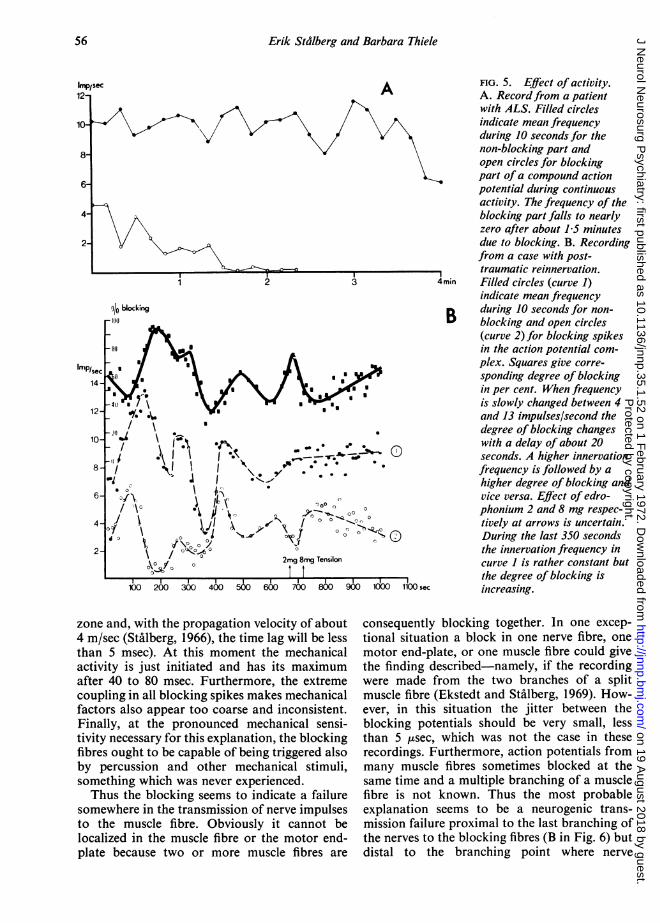
tendency to increased degree of blocking duringthis time, in one case from 20% to total blockafter 1-5 minutes (Fig. 5A).
A
54
IL
Protected by copyright.
on 19 August 2018 by guest.
http://jnnp.bmj.com
/J N
eurol Neurosurg P
sychiatry: first published as 10.1136/jnnp.35.1.52 on 1 February 1972. D
ownloaded from
Transmission block in terminal nerve twigs
Imp/sec
2624
22
2018
16
14//
12
10
8
64
2
:260 3M 4W ~~~~500 600 sec
FIG. 4. Effect of edrophonium. The recording was made in m. extensor digitorum III in a patient withamyotrophic lateral sclerosis. Open circles indicate mean discharge rate during 10 seconds for blocking spikesand filled circles corresponding value for non-blocking spikes in the compound action potential. Upper part(crosses) shows degree of blocking in per cent. Injection of 2 mg edrophonium (each arrow) is followed by areduction in the degree of blocking (somewhat more than usually seen). When the innervation frequency isreduced at the end of the experiment (after 400 seconds) from about 22 to about 15 impulses/sec the degree ofblocking decreases towards zero.
In eight experiments the innervation frequencywas kept constant for a few minutes at differentfrequency levels or slowly changed between 4and 15 impulses/second. With a delay of about20 seconds the blocking increased at higherfrequency and decreased at lower (Fig. SB). Thechange in degree of blocking was somewhatdifferent for different motor units. The jitterincreased slightly during continuous activity.
DISCUSSION
In the normal muscle there is a high safety factorin the neural and neuromuscular transmissionwhere thousands of impulses can be transmittedduring continuous activity without signs ofdecreased accuracy (St'alberg et al., 1971). Inearly periods of progressive disorders withsprouting, compound action potentials arerecorded (in the ordinary EMG as 'giantpotentials' or 'reinnervation potentials') with anincreased jitter and blockings. In this study inter-
est was focused on the phenomenon of 'pairedblocking'. The phenomenon is supposed to takeplace in the muscle and not be caused by suchmechanisms as occasional synchronized activitybetween two anterior horn cells, as it is seen alsoin disorders with only peripheral engagements.Neither could it be explained as an occasionallyappearing double discharge, as the blocking andnon-blocking components were quite differentin shape. Furthermore, the blocking part some-times appears less than 1 msec from the rest ofthe potential-that is, within the refractoryperiod of nerve and muscle fibre and therefore anectopic focus in the terminal nerve tree can alsobe excluded. Are the blocking potentials genera-ted by hyperexcitable muscle fibres, mechanicallytriggered by adjacent muscle fibres? This seemsunlikely, because when the electrical activityfrom the non-blocking fibres is recorded, usuallyless than 5 msec have elapsed since the depolari-zation in the end-plate region. (The recording isusually made less than 20 mm from the end-plate
55P
rotected by copyright. on 19 A
ugust 2018 by guest.http://jnnp.bm
j.com/
J Neurol N
eurosurg Psychiatry: first published as 10.1136/jnnp.35.1.52 on 1 F
ebruary 1972. Dow
nloaded from
Erik Stalberg and Barbara Thiele
sec A
91o blocking
FIG. 5. Effect of activity.A. Recordfrom a patientwith ALS. Filled circlesindicate mean frequencyduring 10 seconds for thenon-blocking part andopen circles for blockingpart ofa compound actionpotential during continuousactivity. The frequency of theblocking part falls to nearlyzero after about 15 minutesdue to blocking. B. Recordingfrom a case with post-traumatic reinnervation.
4min Filled circles (curve 1)indicate mean frequency
B during 10 seconds for non-B blocking and open circles
(curve 2) for blocking spikesin the action potential com-plex. Squares give corre-sponding degree of blockingin per cent. When frequencyis slowly changed between 4and 13 impulses/second thedegree of blocking changeswith a delay of about 20seconds. A higher innervationfrequency is followed by ahigher degree of blocking andvice versa. Effect of edro-phonium 2 and 8 mg respec-tively at arrows is uncertain.During the last 350 secondsthe innervation frequency incurve I is rather constant butthe degree of blocking is
sec increasing.
zone and, with the propagation velocity of about4 m/sec (Stalberg, 1966), the time lag will be lessthan 5 msec). At this moment the mechanicalactivity is just initiated and has its maximumafter 40 to 80 msec. Furthermore, the extremecoupling in all blocking spikes makes mechanicalfactors also appear too coarse and inconsistent.Finally, at the pronounced mechanical sensi-tivity necessary for this explanation, the blockingfibres ought to be capable of being triggered alsoby percussion and other mechanical stimuli,something which was never experienced.Thus the blocking seems to indicate a failure
somewhere in the transmission of nerve impulsesto the muscle fibre. Obviously it cannot belocalized in the muscle fibre or the motor end-plate because two or more muscle fibres are
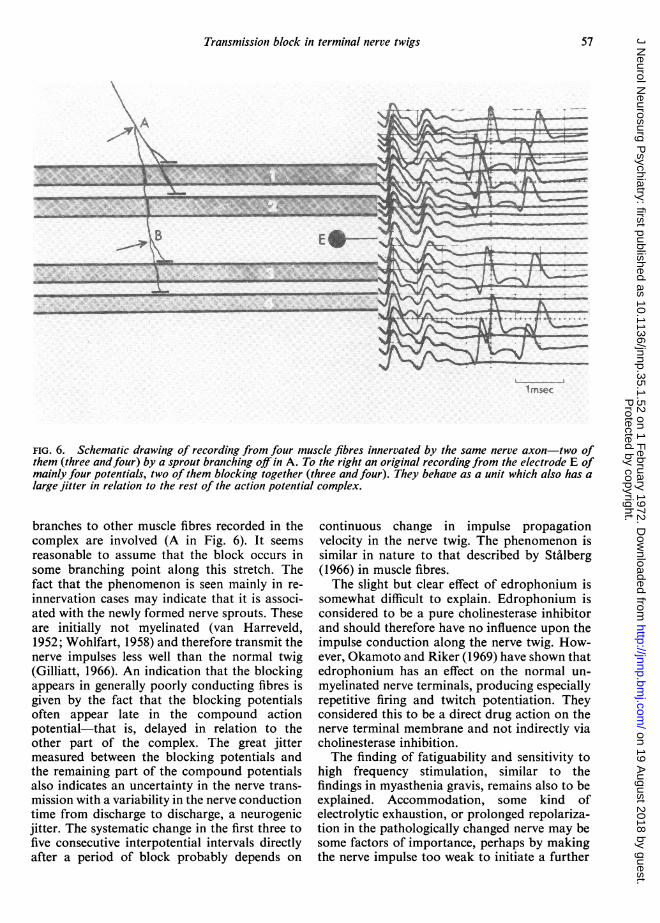
consequently blocking together. In one excep-tional situation a block in one nerve fibre, onemotor end-plate, or one muscle fibre could givethe finding described-namely, if the recordingwere made from the two branches of a splitmuscle fibre (Ekstedt and Stalberg, 1969). How-ever, in this situation the jitter between theblocking potentials should be very small, lessthan 5 usec, which was not the case in theserecordings. Furthermore, action potentials frommany muscle fibres sometimes blocked at thesame time and a multiple branching of a musclefibre is not known. Thus the most probableexplanation seems to be a neurogenic trans-mission failure proximal to the last branching ofthe nerves to the blocking fibres (B in Fig. 6) butdistal to the branching point where nerve
Imp/12-
10-
8-
6-
4-
2-
56P
rotected by copyright. on 19 A
ugust 2018 by guest.http://jnnp.bm
j.com/
J Neurol N
eurosurg Psychiatry: first published as 10.1136/jnnp.35.1.52 on 1 F
ebruary 1972. Dow
nloaded from
Transmission block in terminal nerve twigs
I msec
FIG. 6. Schematic drawing of recording from four muscle fibres innervated by the same nerve axon-two ofthem (three andfour) by a sprout branching off in A. To the right an original recording from the electrode E ofmainly four potentials, two of them blocking together (three andfour). They behave as a unit which also has alarge jitter in relation to the rest of the action potential complex.
branches to other muscle fibres recorded in thecomplex are involved (A in Fig. 6). It seemsreasonable to assume that the block occurs insome branching point along this stretch. Thefact that the phenomenon is seen mainly in re-innervation cases may indicate that it is associ-ated with the newly formed nerve sprouts. Theseare initially not myelinated (van Harreveld,1952; Wohlfart, 1958) and therefore transmit thenerve impulses less well than the normal twig(Gilliatt, 1966). An indication that the blockingappears in generally poorly conducting fibres isgiven by the fact that the blocking potentialsoften appear late in the compound actionpotential-that is, delayed in relation to theother part of the complex. The great jittermeasured between the blocking potentials andthe remaining part of the compound potentialsalso indicates an uncertainty in the nerve trans-mission with a variability in the nerve conductiontime from discharge to discharge, a neurogenicjitter. The systematic change in the first three tofive consecutive interpotential intervals directlyafter a period of block probably depends on
continuous change in impulse propagationvelocity in the nerve twig. The phenomenon issimilar in nature to that described by Stalberg(1966) in muscle fibres.The slight but clear effect of edrophonium is
somewhat difficult to explain. Edrophonium isconsidered to be a pure cholinesterase inhibitorand should therefore have no influence upon theimpulse conduction along the nerve twig. How-ever, Okamoto and Riker (1969) have shown thatedrophonium has an effect on the normal un-myelinated nerve terminals, producing especiallyrepetitive firing and twitch potentiation. Theyconsidered this to be a direct drug action on thenerve terminal membrane and not indirectly viacholinesterase inhibition.The finding of fatiguability and sensitivity to
high frequency stimulation, similar to thefindings in myasthenia gravis, remains also to beexplained. Accommodation, some kind ofelectrolytic exhaustion, or prolonged repolariza-tion in the pathologically changed nerve may besome factors of importance, perhaps by makingthe nerve impulse too weak to initiate a further
57P
rotected by copyright. on 19 A
ugust 2018 by guest.http://jnnp.bm
j.com/
J Neurol N
eurosurg Psychiatry: first published as 10.1136/jnnp.35.1.52 on 1 F
ebruary 1972. Dow
nloaded from

Erik Stdlberg and Barbara Thiele
propagation after a branching point or along anabnormal nerve branch. The systematic changeof interspike intervals at consecutive dischargesmay indicate some cumulative process. Someparallels may also be drawn with the finding ofBessou, Burgess, Perl, and Taylor (1971) whodescribed a decreasing response in C-mechano-receptors in cats during repeated mechanicalstimulation in the receptive field. The same typeof findings with exhaustion at iterative mechani-cal and electrical stimulation have now also beenmade in human C-mechanoreceptors (Hallin andTorebjork, 1970; Torebjork and Hallin, 1970),where the degree of blocking is positively corre-lated to stimulation frequency. Obviously thesethin and unmyelinated nerve structures behavein a way similar to some nerve twigs at re-innervation.Are the neurogenic blockings localized only in
the nerve tree where it is still branching or maythey also happen in more terminal parts? Theseparts are the last to be myelinated at reinnerva-tion and are also thinner and they ought there-fore to offer the lowest safety factor for trans-mission. In the present recordings, there wasoften block in only one spike component,reacting to high frequency and edrophoniumlike the recordings with 'paired blockings'.Here the block naturally could be situated in aproximal part of the nerve twig, providing therecording were made from only one of themuscle fibres innervated by a badly conductingnerve fibre. It may, however, also be explainedby a block in the very terminal nerve twigs.Possibly the greatest uncertainty during re-innervation is in the most peripheral parts, some-times extending proximally over the last branch-ing points of the nerve twig and so giving theneurophysiological possibility of confirming aneurogenic transmission failure in the form of'paired blocking'. Thus, part of the jitter andblockings seen in single spikes in the compoundaction potentials in muscles undergoing re-innervation may be of neurogenic origin. The' coupling discharge' in neurogenic muscularatrophy reported by Takahashi (1966)-that is,a small mono- or biphasic intermittently block-ing spike in a compound action potential-couldthus indicate a neurogenic block.
Might the described sensitivity to high innerva-tion frequencies with increasing blockings beassociated with the increased fatigue duringactivity clinically experienced in neuromusculardisorders, or is the degree of blocking usually of
minor importance quantitatively? It is plannedto investigate this.
Neurogenic blocking may also sometimes beresponsible for a decreasing muscle response atnerve stimulation, wrongly interpreted as 'myas-thenic reaction'.
It cannot at present be stated with certaintyfrom a recording with single fibre blockingwhether the block is neurogenic or situated inthe neuromuscular junction or in the musclefibre. One indication favouring neurogenicinvolvement might be the finding of a tendencyto a systematic change in the interpotentialintervals instead of a quite random one as seenafter curare or in myasthenia gravis. Further-more, tests with, for example, edrophonium canbe made when trying to sort out the neuro-muscular block, but a positive effect can notfully exclude a neurogenic cause for the block.It is, therefore, now of interest to discover othertests to increase the chances of differentiatingbetween neurogenic, neuromuscular, and myo-genic block.
The investigation was supported by the SwedishMedical Research Council (Grant No. 14X-135)(Stalberg) and the Deutsche Forschungsgemeinschaft(Thiele).
REFERENCES
Bessou, P., Burgess, P. R., Per], E. R., and Taylor, C. B.(1971). Dynamic properties of mechanoreceptors with un-myelinated (C) fibres. Journal of Neurophysiology, 34, 116-131.
Blom, S., and Ringqvist, I. (1971). Neurophysiological find-ings in myasthenia gravis. Single muscle fibre activity inrelation to muscular fatiguability and response to anti-cholinesterase. Electroencephalography and Clinical Neuro-physiology, 30, 477-487.
Ekstedt, J. (1964). Human single muscle fiber action poten-tials. Acta Physiologica Scandinavica, 61, Suppl. 226.
Ekstedt, J., and Stflberg, E. (1963). A method for recordingextracellular action potentials of single muscle fibres andmeasuring their propagation velocity in voluntarilyactivated human muscle. Bulletin of the American Associa-tion ofEMG Electrodiagnosis, 10, 16.
Ekstedt, J., and StAlberg, E. (1967). Myasthenia gravis.Diagnostic aspects by a new electrophysiological method.Opuscula Medica, 12, 73-76.
Ekstedt, J., and Stalberg, E. (1969). Abnormal connectionsbetween skeletal muscle fibers. Electroencephalography andClinical Neurophysiology, 27, 607-609.
Gilliatt, R. W. (1966). Nerve conduction in human andexperimental neuropathies. Proceedings of the RoyalSociety of Medicine, 59, 989-993.
Hallin, R. G., and Torebj6rk, H. E. (1970). Afferent andefferent C units recorded from human skin nerves in situ.Acta Societatis Medicorum Upsaliensis, 75, 277-281.
Harreveld, A. van (1952). Re-innervation of paretic muscleby collateral branching of the residual motor innervation.Journal of Comparative Neurology, 97, 385-407.
Kugelberg, E., Edstrom, L., and Abbruzzese, M. (1970).Mapping of motor units in experimentally reinnervated rat
58P
rotected by copyright. on 19 A
ugust 2018 by guest.http://jnnp.bm
j.com/
J Neurol N
eurosurg Psychiatry: first published as 10.1136/jnnp.35.1.52 on 1 F
ebruary 1972. Dow
nloaded from

Transmission block in terminal nerve twigs
muscle. Journal ofNeurology, Neurosurgery, andPsychiatry,33, 319-329.
Okamoto, M., and Riker, W., Jnr. (1969). Subacutedenervation: a means of disclosing mammalian motornerve terminals as critical sites of acetylcholine andfacilitory drug actions. Journal of Pharmacology andExperimental Therapeutics, 166, 217-224.
Stalberg, E. (1966). Propagation velocity in human musclefibers in situ. Acta Physiologica Scandinavica, 70, Suppl.287.
Stalberg, E., Ekstedt, J., and Broman, A. (1971). The electro-myographic jitter in normal human muscles. Electro-
encephalography and Clinical Neurophysiology. 31, 429-438.
Takahashi, K. (1966). The coupling discharge in neurogenicmuscular atrophy. Archives of Neurology, 14, 617-623.
Torebjork, H. E., and Hallin, R. G. (1970). C-fibre unitsrecorded from human sensory nerve fascicles in situ. ActaSocietatis Medicorum Upsaliensis, 75, 81-84.
Wohlfart, G. (1957). Collateral regeneration from residualmotor nerve fibers in amyotrophic lateral sclerosis.Neurology (Minneap.), 7, 124-134.
Wohlfart, G. (1958). Collateral regeneration in partiallydenervated muscles. Neurology (Minneap.), 8. 175-180.
59P
rotected by copyright. on 19 A
ugust 2018 by guest.http://jnnp.bm
j.com/
J Neurol N
eurosurg Psychiatry: first published as 10.1136/jnnp.35.1.52 on 1 F
ebruary 1972. Dow
nloaded from





























