Embed Size (px)
Citation preview

Abstract This research examined token-mediated tool-use in a tufted capuchin monkey (Cebus apella). We con-ducted five experiments. In experiment 1 we examinedthe use of plastic color-coded chips to request food, and inexperiments 2–5 we examined the use of color-coded chipsto request tools. Our subject learned to use chips to requesttools following the same general pattern seen in great apesperforming analogous tasks, that is, initial discriminationfollowed by an understanding of the relationship among to-kens, tools, and their functions. Our findings are consistentwith the view that parallel representational processes under-lie the tool-related behavior of capuchins and great apes.
Key words Capuchin · Cebus · Representation · Tool use
Introduction
Behavior that reflects mental representation presents com-parative psychologists with unique opportunities for un-derstanding theoretically significant cognitive processes,including abstraction, reasoning, and learning (Klüver1933; Piaget 1952; Rumbaugh 1977). Early research inthis area showed that great apes use tokens to request spe-cific foods from human experimenters (Kohts 1935; Wolfe1936), whereas later studies showed that apes combinesymbols in more elaborate communicatory contexts(Savage-Rumbaugh 1986). Research on analogous capac-ities in monkeys has been less conclusive (Carpenter andLocke 1937), contributing to the view that monkeys donot have representational intelligence comparable to thatof apes (for a review of research on primate representa-tional abilities see Tomasello and Call 1997).
Tufted capuchins (Cebus apella) are New World mon-keys whose manipulative skills rival those of great apesand human children (Chevalier-Skolnikoff 1989; Visal-berghi 1990). Captive capuchins use tools in a variety ofcontexts, including sticks used as probes to extract syrupfrom enclosed containers, and stones as hammers to openhard-shelled nuts (Westergaard and Fragaszy 1987;Anderson 1990). Capuchins exhibit many tool-relatedskills that have been associated with mental representa-tion in apes (Köhler 1927), including appropriate selec-tion of tools from an array of objects, the use of tool-sets,retrieval of tools from a site visually separate from the siteof use, modification of tools prior to their use, and tool-combination (Westergaard and Fragaszy 1987; Wester-gaard and Suomi 1993a, 1994; Anderson and Henneman1994; Westergaard et al., in press). On the other hand E.Visalberghi has argued that capuchins lack representa-tional intelligence and therefore do not understand thecausal aspects of their tool-using actions (Visalberghi andTrinca 1989; Visalberghi and Limongelli 1994, 1996; Visal-berghi et al. 1995). This hypothesis is based on researchindicating that capuchins do not perform as well as apes ina task that requires the use of a stick to push food througha tube. However, the apes used in E. Visalberghi’s studieswere reared with a greater level of human interaction andtraining than were the capuchins, and despite this differ-ence only a subset of the apes performed more skillfullythan did the monkeys. Further, E. Visalberghi’s experi-ments use a conceptually complex task whose sensitivityfor assessing representational intelligence in capuchinsand apes has not been established. Indeed, human childrenare not proficient in variations of this task until about 30months of age, well past the age at which causal under-standing first emerges (Piaget 1952; Visalberghi andLimongelli 1996). Finally, it must be noted that capuchinsperform as well as apes in problems that assess causal un-derstanding of spatial support (Spinozzi and Poti 1989),and the relations among tool-size, material, and orienta-tion (Natale 1989).
Here we examined token-mediated tool-use in a tuftedcapuchin. We conducted five experiments. In experiment 1
Gregory Charles Westergaard · Chanya Liv ·Tara J. Chavanne · Stephen J. Suomi
Token-mediated tool-use by a tufted capuchin monkey (Cebus apella)
Anim Cogn (1998) 1 :101–106 © Springer-Verlag 1998
Received: 11 March 1998 / Accepted after revision: 19 August 1998
ORIGINAL ARTICLE
G. C. Westergaard (Y) · C. Liv · T. J. Chavanne · S. J. SuomiLaboratory of Comparative Ethology, National Institute of Child Health and Human Development, P.O. Box 529, Poolesville, Maryland, MD 20837, USAe-mail: [email protected], Tel.: +1-301-4960444, Fax: +1-301-4960630

we examined a capuchin's use of color-coded chips to request food, and in experiments 2–5 we examined thesame capuchin’s use of color-coded chips to request dif-ferent tools (a metal bolt and a stone) to meet the require-ments of different tasks (probing and nut-cracking). Wepropose that the use of tokens by a capuchin to make com-municative requests for specific context-dependent toolswould provide evidence for representational intelligencein the instrumental behavior of this species. Such a find-ing would be consistent with the view that parallel mentalprocesses underlie the instrumental behavior of capuchinsand great apes (Anderson 1996; Chevalier-Skolnikoff1989; Parker and Gibson 1977; Westergaard and Fragaszy1987), and inconsistent with a discontinuous view of pri-mate representational processes related to instrumentalbehavior (Visalberghi and Trinca 1989; Visalberghi andLimongelli 1994, 1996; Visalberghi et al. 1995).
Methods
Subject and housing
The subject was a captive-born adult male tufted capuchin monkey(“Virgil”), aged 9 years at the onset of this study. Virgil was expe-rienced in the use of tools but had no prior experience in the use ofcolor-coded chips or other tokens prior to this research. Before theonset of testing we taught Virgil to trade with humans by givinghim a grape each time he handed a chow biscuit (PMI Feeds No.5045) directly to an experimenter. This training took place over a1-week period in Virgil’s home cage, which he shared with fiveother capuchins. Virgil was chosen as the subject for this researchbecause of his willingness to participate in the experimental proce-dures. During formal test sessions we housed Virgil alone in a sec-ond indoor cage (2 × 1 × 2 m). This cage contained two adjacentmetal bowls (each 18 cm in diameter and 7 cm in depth) located onthe floor at one end of the cage.
Apparatus
We used plastic color-coded disks (each 4 cm in diameter and 3 g inweight) as tokens, and two enclosed plastic containers (each 10 ×10 cm) as tool-sites to examine probing and nut-cracking skills. Bothcontainers had a single narrow opening (1.5–1.8 cm in diameter)on top. The probing apparatus was partially filled with sweet syrup(Westergaard and Suomi 1994). The nut-cracking apparatus held ahazel-nut in a fixed position (Westergaard and Suomi 1993b).
General procedure
Two experimenters conducted test sessions. At the onset of eachtrial one experimenter simultaneously placed one chip into each ofthe two bowls on the cage floor (we randomly determined the bowlinto which each chip was placed). The experimenter then lookedstraight ahead or up towards the ceiling so as not to provide visualcues regarding chip placement. A second experimenter sat outsidethe opposite end of the test cage. To complete a trial Virgil had to“select” a chip, that is, remove a chip from a bowl, carry it acrossthe cage floor, and then hand it directly to the second experi-menter. This experimenter then handed Virgil either a food reward,an object that corresponded to the color of the selected chip, ornothing, depending on the test condition. Food and tools were keptout of Virgil’s view until after he handed a chip to the second ex-perimenter. The first experimenter noted the color of the chip thatVirgil selected.
Analysis
We set a performance criterion of at least 80% correct responseswithin a 50-trial block. Further, before we proceeded with a subse-quent experiment we required that Virgil meet this criterion withineach of at least two consecutive 50-trial blocks. We used the bino-mial test (P < 0.05, two-tailed) to examine Virgil’s performanceagainst a null hypothesis distribution with a mean of 50% correct.
Experiment 1
Procedure
At the onset of each trial an experimenter placed a whitechip and a red chip into the bowls. If Virgil selected awhite chip he was given a grape. If Virgil selected a redchip he was not given a grape and a new trial was initi-ated. We conducted 450 trials over a 10-day period.
Results
Virgil’s performance is summarized in Fig. 1. Virgil se-lected the correct (white) chip in 44% of the trials duringthe first 50-trial block. This rate did not differ signifi-cantly from the rate expected by chance (P > 0.05).Virgil’s performance improved in subsequent trial blocksand he reached the performance criterion during the sev-enth 50-trial block.
Discussion
In this experiment Virgil used color-coded chips to re-quest food from a human experimenter. Virgil’s behaviorfollowed the same general learning pattern seen in pri-mates in more traditional discrimination paradigms (Harlow1951). Virgil initially selected the correct chip at the levelexpected by chance and over a number of trials his per-formance improved steadily until he reliably selected the
102
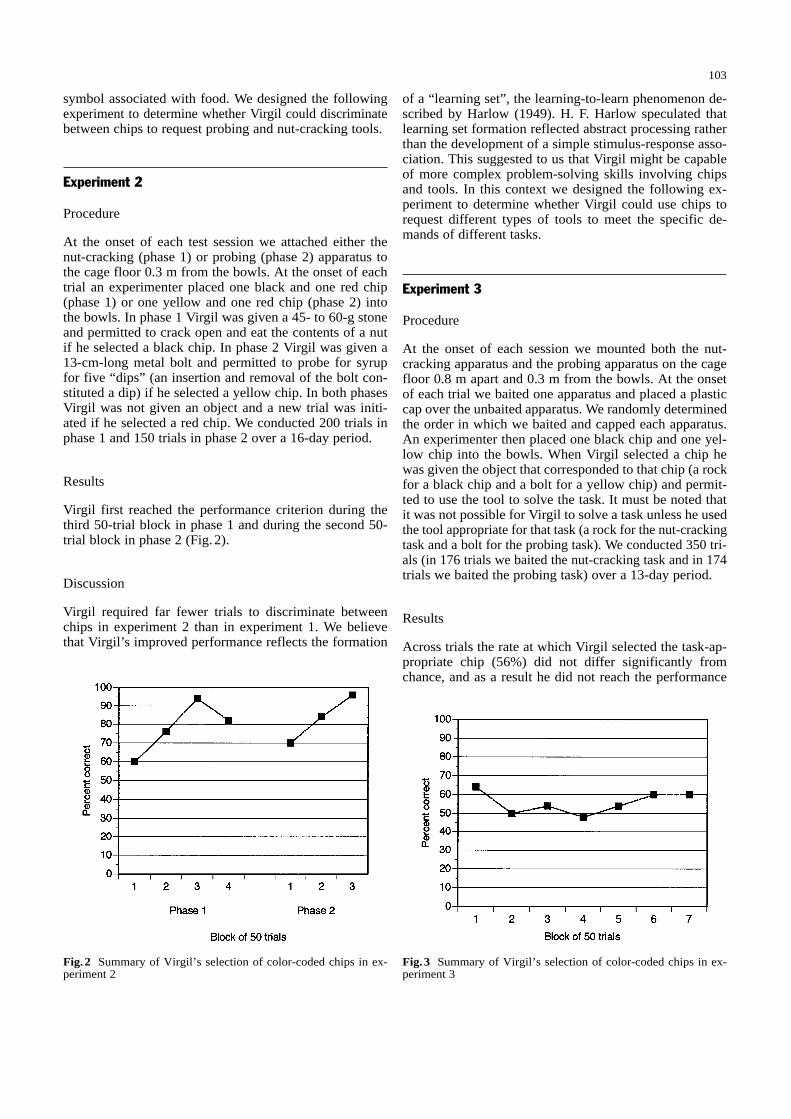
Fig.1 Summary of Virgil’s selection of color-coded chips in ex-periment 1
symbol associated with food. We designed the followingexperiment to determine whether Virgil could discriminatebetween chips to request probing and nut-cracking tools.
Experiment 2
Procedure
At the onset of each test session we attached either thenut-cracking (phase 1) or probing (phase 2) apparatus tothe cage floor 0.3 m from the bowls. At the onset of eachtrial an experimenter placed one black and one red chip(phase 1) or one yellow and one red chip (phase 2) intothe bowls. In phase 1 Virgil was given a 45- to 60-g stoneand permitted to crack open and eat the contents of a nutif he selected a black chip. In phase 2 Virgil was given a13-cm-long metal bolt and permitted to probe for syrupfor five “dips” (an insertion and removal of the bolt con-stituted a dip) if he selected a yellow chip. In both phasesVirgil was not given an object and a new trial was initi-ated if he selected a red chip. We conducted 200 trials inphase 1 and 150 trials in phase 2 over a 16-day period.
Results
Virgil first reached the performance criterion during thethird 50-trial block in phase 1 and during the second 50-trial block in phase 2 (Fig. 2).
Discussion
Virgil required far fewer trials to discriminate betweenchips in experiment 2 than in experiment 1. We believethat Virgil’s improved performance reflects the formation
of a “learning set”, the learning-to-learn phenomenon de-scribed by Harlow (1949). H. F. Harlow speculated thatlearning set formation reflected abstract processing ratherthan the development of a simple stimulus-response asso-ciation. This suggested to us that Virgil might be capableof more complex problem-solving skills involving chipsand tools. In this context we designed the following ex-periment to determine whether Virgil could use chips torequest different types of tools to meet the specific de-mands of different tasks.
Experiment 3
Procedure
At the onset of each session we mounted both the nut-cracking apparatus and the probing apparatus on the cagefloor 0.8 m apart and 0.3 m from the bowls. At the onsetof each trial we baited one apparatus and placed a plasticcap over the unbaited apparatus. We randomly determinedthe order in which we baited and capped each apparatus.An experimenter then placed one black chip and one yel-low chip into the bowls. When Virgil selected a chip hewas given the object that corresponded to that chip (a rockfor a black chip and a bolt for a yellow chip) and permit-ted to use the tool to solve the task. It must be noted thatit was not possible for Virgil to solve a task unless he usedthe tool appropriate for that task (a rock for the nut-crackingtask and a bolt for the probing task). We conducted 350 tri-als (in 176 trials we baited the nut-cracking task and in 174trials we baited the probing task) over a 13-day period.
Results
Across trials the rate at which Virgil selected the task-ap-propriate chip (56%) did not differ significantly fromchance, and as a result he did not reach the performance
103
Fig. 2 Summary of Virgil’s selection of color-coded chips in ex-periment 2
Fig.3 Summary of Virgil’s selection of color-coded chips in ex-periment 3

criterion within any of the seven 50-trial blocks (Fig. 3).Further analysis indicated that Virgil selected the yellowchip in 64% of the trials in which we baited the probingapparatus and in 53% of the trials in which we baited thenut-cracking apparatus, indicating Virgil's performancewas above chance level when we baited the probing appa-ratus but not when we baited the nut-cracking apparatus.
Discussion
Virgil preferentially selected the chip associated with themetal bolt when we baited the probing task, but showedno preference in his selection of chips when we baited thenut-cracking task. It appeared to us that Virgil did notcomprehend the task recognition requirement for this ex-periment. To address this issue we conducted the follow-ing experiment in which we baited each task for a numberof consecutive trials. We speculated that this procedurewould facilitate task recognition and association of tasks,appropriate tools, and their corresponding chips.
Experiment 4
Procedure
The procedure used in experiment 4 was similar to thatused in experiment 3 except that we baited the same ap-paratus (and capped the other apparatus) for either ten(phase 1) or five (phase 2) consecutive trials. At the endof each ten- or five-trial block we baited the other appara-tus for the same number of consecutive trials. We con-ducted 200 trials in phase 1 and 150 trials in phase 2 overa 21-day period.
Results
Virgil reached the performance criterion during the sec-ond 50-trial block in phase 1, and during the first 50-trialblock in phase 2 (Fig.4). Overall, Virgil selected the cor-rect chip in 89% of the test trials, including 91% of the tri-als in which we baited the probing apparatus and 86% ofthe trials in which we baited the nut-cracking apparatus.Further analysis indicated that Virgil’s performance on thefirst trial of each consecutive trial set (in which we baitedthe same apparatus) was initially at the level expected bychance (7 correct on trial 1 within the first 14 trial sets)but improved to over 97% correct on trial 1 of 34 subse-quent trial sets.
Discussion
The results of this experiment indicate that Virgil devel-oped a chip selection strategy based on task recognition.In the following experiment we evaluated the effective-
ness and flexibility of this strategy by again randomly pre-senting the probing and nut-cracking tasks.
Experiment 5
Procedure
The procedure used in experiment 5 was similar to thatused in experiment 3. We conducted 250 trials (in 125 tri-als we baited the nut-cracking apparatus and in 125 trialswe baited the probing apparatus) over a 14-day period.We randomly determined the order in which we baitedeach apparatus. In trials 201–250 we reversed the locationof each apparatus in order to control for the possibilitythat Virgil’s chip selection was based on a position cuerather than his understanding of the representational na-ture of the task. In addition to noting whether Virgil se-lected correct or incorrect chips the experimenters alsonoted whether Virgil placed tools in contact with thecapped apparatus, and whether Virgil attempted to use aninappropriate tool in trials in which he selected the incor-rect chip.
Results
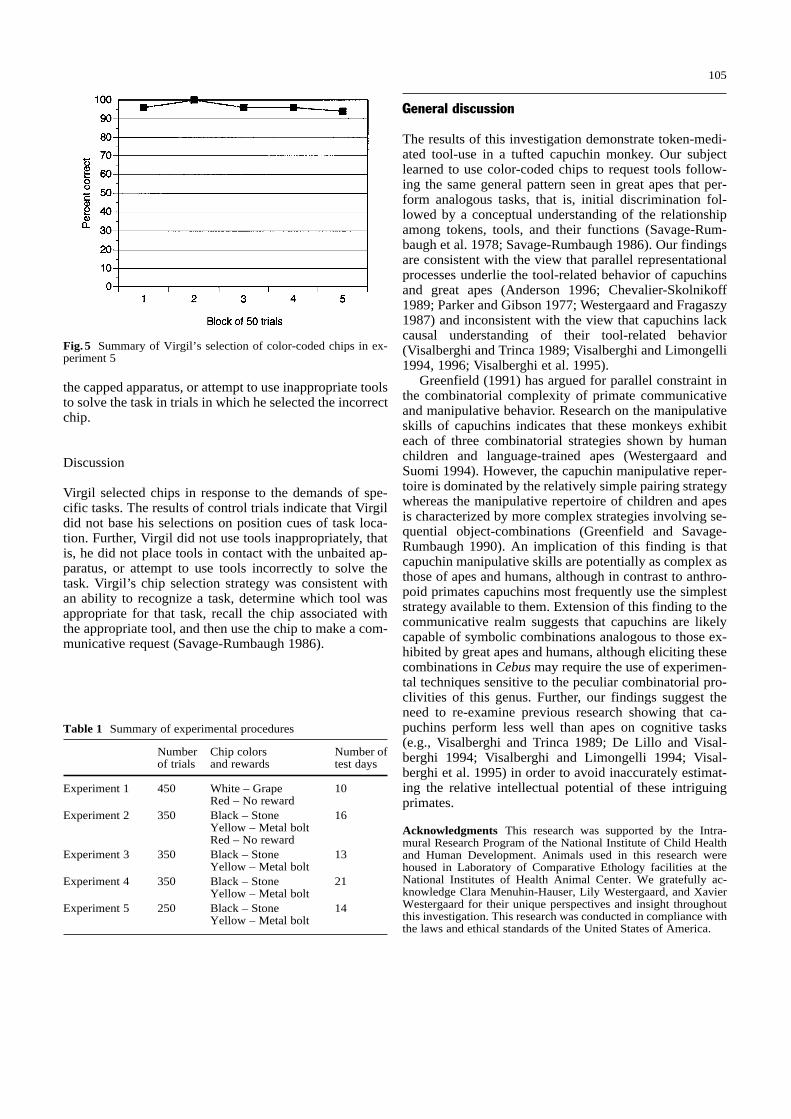
Virgil first reached the performance criterion during thefirst 50-trial block (Fig. 5). Overall, Virgil selected thecorrect chip in 241 of the 250 trials (96%). Further analy-sis indicated that Virgil selected the correct chip in 97% ofthe trials in which we baited the probing apparatus and in96% of the trials in which we baited the nut-cracking ap-paratus. Virgil’s performance in trials 201–250 after weswitched the location of each task (94% correct) did notdiffer significantly from his performance in trials 1–200(97% correct). Virgil did not place tools in contact with
104
Fig.4 Summary of Virgil’s selection of color-coded chips in ex-periment 4
the capped apparatus, or attempt to use inappropriate toolsto solve the task in trials in which he selected the incorrectchip.
Discussion
Virgil selected chips in response to the demands of spe-cific tasks. The results of control trials indicate that Virgildid not base his selections on position cues of task loca-tion. Further, Virgil did not use tools inappropriately, thatis, he did not place tools in contact with the unbaited ap-paratus, or attempt to use tools incorrectly to solve thetask. Virgil’s chip selection strategy was consistent withan ability to recognize a task, determine which tool wasappropriate for that task, recall the chip associated withthe appropriate tool, and then use the chip to make a com-municative request (Savage-Rumbaugh 1986).
General discussion
The results of this investigation demonstrate token-medi-ated tool-use in a tufted capuchin monkey. Our subjectlearned to use color-coded chips to request tools follow-ing the same general pattern seen in great apes that per-form analogous tasks, that is, initial discrimination fol-lowed by a conceptual understanding of the relationshipamong tokens, tools, and their functions (Savage-Rum-baugh et al. 1978; Savage-Rumbaugh 1986). Our findingsare consistent with the view that parallel representationalprocesses underlie the tool-related behavior of capuchinsand great apes (Anderson 1996; Chevalier-Skolnikoff1989; Parker and Gibson 1977; Westergaard and Fragaszy1987) and inconsistent with the view that capuchins lackcausal understanding of their tool-related behavior(Visalberghi and Trinca 1989; Visalberghi and Limongelli1994, 1996; Visalberghi et al. 1995).
Greenfield (1991) has argued for parallel constraint inthe combinatorial complexity of primate communicativeand manipulative behavior. Research on the manipulativeskills of capuchins indicates that these monkeys exhibiteach of three combinatorial strategies shown by humanchildren and language-trained apes (Westergaard andSuomi 1994). However, the capuchin manipulative reper-toire is dominated by the relatively simple pairing strategywhereas the manipulative repertoire of children and apesis characterized by more complex strategies involving se-quential object-combinations (Greenfield and Savage-Rumbaugh 1990). An implication of this finding is thatcapuchin manipulative skills are potentially as complex asthose of apes and humans, although in contrast to anthro-poid primates capuchins most frequently use the simpleststrategy available to them. Extension of this finding to thecommunicative realm suggests that capuchins are likelycapable of symbolic combinations analogous to those ex-hibited by great apes and humans, although eliciting thesecombinations in Cebus may require the use of experimen-tal techniques sensitive to the peculiar combinatorial pro-clivities of this genus. Further, our findings suggest theneed to re-examine previous research showing that ca-puchins perform less well than apes on cognitive tasks(e.g., Visalberghi and Trinca 1989; De Lillo and Visal-berghi 1994; Visalberghi and Limongelli 1994; Visal-berghi et al. 1995) in order to avoid inaccurately estimat-ing the relative intellectual potential of these intriguingprimates.
Acknowledgments This research was supported by the Intra-mural Research Program of the National Institute of Child Healthand Human Development. Animals used in this research werehoused in Laboratory of Comparative Ethology facilities at theNational Institutes of Health Animal Center. We gratefully ac-knowledge Clara Menuhin-Hauser, Lily Westergaard, and XavierWestergaard for their unique perspectives and insight throughoutthis investigation. This research was conducted in compliance withthe laws and ethical standards of the United States of America.
105
Fig. 5 Summary of Virgil’s selection of color-coded chips in ex-periment 5
Table 1 Summary of experimental procedures
Number Chip colors Number ofof trials and rewards test days
Experiment 1 450 White – Grape 10Red – No reward
Experiment 2 350 Black – Stone 16Yellow – Metal boltRed – No reward
Experiment 3 350 Black – Stone 13Yellow – Metal bolt
Experiment 4 350 Black – Stone 21Yellow – Metal bolt
Experiment 5 250 Black – Stone 14Yellow – Metal bolt

References
Anderson JR (1990) Use of objects as hammers to open nuts by ca-puchin monkeys (Cebus apella). Folia Primatol 54:138–145
Anderson JR (1996) Chimpanzees and capuchin monkeys: Com-parative cognition. In: Russon AE, Bard KA, Parker ST (eds)Reaching into thought: the minds of the great apes. CambridgeUniversity Press, Cambridge, pp 23–56
Anderson JR, Henneman MC (1994) Solutions to a tool-use prob-lem in a pair of Cebus apella. Mammalia 58:351–361
Carpenter CR, Locke NM (1937) Notes on symbolic behavior in aCebus monkey (Capucinus appella). J Genet Psychol 51:267–278
Chevalier-Skolnikoff S (1989) Spontaneous tool use and sensori-motor intelligence in Cebus compared with other monkeys andapes. Behav Brain Sci 12:561–627
De Lillo C, Visalberghi E (1994) Transfer index and mediationallearning in tufted capuchins (Cebus apella). Int J Primatol15:275–287
Greenfield PM (1991) Language, tools, and the brain: the on-togeny and phylogeny of hierarchically organized sequentialbehavior. Behav Brain Sci 14:531–595
Greenfield PM, Savage-Rumbaugh ES (1990) Grammatical com-binations in Pan paniscus: processes of learning and invention.In: Parker ST, Gibson KR (eds) “Language” and intelligence inmonkeys and apes: comparative developmental perspectives.Cambridge University Press, Cambridge, pp 540–578
Harlow HF (1949) The formation of learning sets. Psychol Rev56:51–65
Harlow HF (1951) Primate learning. In: Stone CP (ed) Compara-tive psychology. Prentice-Hall, Englewood Cliffs, pp 183–238
Klüver H (1933) Behavior mechanisms in monkeys. University ofChicago Press, Chicago
Köhler W (1927) The mentality of apes. Vintage, New YorkKohts N (1935) Infant ape and human child (in Russian with
English summary). Scientific Memoirs of the Museum Darwin-ianum, Moscow
Natale F (1989) Causality. II. The stick problem. In: Antinucci F(ed) Cognitive structure and development in nonhuman pri-mates. Lawrence Erlbaum, Hillsdale, pp 121–133
Parker ST, Gibson KR (1977) Object manipulation, tool use andsensorimotor intelligence as feeding adaptations in Cebus mon-keys and great apes. J Hum Evol 6:623–641
Piaget J (1952) The origins of intelligence in children (translatedby M Cook). International Universities Press, New York
Rumbaugh DM (1977) Language learning by a chimpanzee: theLana project. Academic Press,New York
Savage-Rumbaugh ES (1986) Ape language: from conditioned re-sponse to symbol. Columbia University Press, New York
Savage-Rumbaugh ES, Rumbaugh DM, Boysen S (1978)Linguistically mediated tool use and exchange by chimpanzees(Pan troglodytes). Behav Brain Sci 4:539–554
Spinozzi G, Poti P (1989) Causality. I. The support problem. In:Antinucci F (ed) Cognitive structure and development in non-human primates. Lawrence Erlbaum, Hillsdale, pp 113–119
Tomasello M, Call J (1997) Primate cognition. Oxford UniversityPress, New York
Visalberghi E (1990) Tool use in Cebus. Folia Primatol 54:146–154
Visalberghi E, Limongelli L (1994) Lack of comprehension ofcause-effect relations in tool-using capuchin monkeys (Cebusapella). J Comp Psychol 108:12–22
Visalberghi E, Limongelli L (1996) Acting and understanding:tool use revisited through the minds of capuchin monkeys. In:Russon AE, Bard KA, Parker ST (eds) Reaching into thought:the minds of the great apes. Cambridge University Press,Cambridge, pp 57–79
Visalberghi E, Trinca L (1989) Tool use in capuchin monkeys: dis-tinguishing between performing and understanding. Primates30:511–521
Visalberghi E, Fragaszy DM, Savage-Rumbaugh ES (1995) Per-formance in a tool-using task by common chimpanzees (Pantroglodytes), bonobos (Pan paniscus), an orangutan (Pongopygmaeus), and capuchin monkeys (Cebus apella). J CompPsychol 109:52–60
Westergaard GC, Fragaszy DM (1987) The manufacture and useof tools by capuchin monkeys (Cebus apella). J Comp Psychol102:159–168
Westergaard GC, Suomi SJ (1993a) Use of a tool-set by capuchinmonkeys (Cebus apella). Primates 34:459–462
Westergaard GC, Suomi SJ (1993b) Hand preference in the use ofnut-cracking tools by tufted capuchin monkeys (Cebus apella).Folia Primatol 61:38–42
Westergaard GC, Suomi SJ (1994) Hierarchical complexity ofcombinatorial manipulation in capuchin monkeys (Cebusapella). Am J Primatol 32:171–176
Westergaard GC, Lundquist AL, Haynie MK, Kuhn HE, Suomi SJ(in press) Why some capuchin monkeys (Cebus apella) useprobing tools (and others do not). J Comp Psychol
Wolfe JB (1936) Effectiveness of token-rewards for chimpanzees.Comp Psychol Monogr 12:72
106
![Give What You Get: Capuchin Monkeys (Cebus apella) and ... Files/Leimgruber_et...(Cebus apella). While there is evidence that capuchin monkeys [36– 38] and young children [39–42]](https://img.pdfslide.us/doc/110x75/614290aed9e4dc11f47f21a3/give-what-you-get-capuchin-monkeys-cebus-apella-and-filesleimgruberet.jpg)




























