Embed Size (px)
Citation preview

T H X JOURNAL OF BIOLOGICAL CHXMISTRY
Printed m U S A. Vol. 258, No. 1, Issue of January 10, pp. 67-54, 1983
Role of Carbohydrate in Human Chorionic Gonadotropin EFFECT OF DEGLYCOSYLATION ON THE SUBUNIT INTERACTION AND ON ITS IN VITRO AND I N VIVO BIOLOGICAL PROPERTIES*
(Received for publication, June 25, 1982)
Narender K. Kalyan and Om P. Bahlt From the Department of Biological Sciences, Division of Cell and Molecular Biology, State University of New York a t Buffalo, Amherst Campus, Buffalo, New York 14260
Previous studies from our laboratory have found that partial removal or modification of the carbohydrate of human chorionic gonadotropin (hCG) results in the impairment of the biological activity in vitro and in vivo without affecting its binding to the receptor. In this report, the complete removal of carbohydrate from hCG has been attempted by the application of trifluo- romethane sulfonic acid to the subunits followed by the reconstitution of the resulting deglycosylated subunits. The treatment with trifluoromethane sulfonic acid re- moved all the carbohydrates except the last peptide- linked hexosarnines, without affecting the polypeptide chains as shown by their amino acid compositions, NH2- terminal analysis, molecular weights, CD spectroscopy, and immunoreactivity. The reconstitution of the native and deglycosylated subunits resulted in one deglyco- sylated derivative, hCG-da+d@ and two mixed hybrids, hCG-da+@ and hCG-a+d@. All the reconstituted hCG derivatives not only bound to the testicular receptor with a high affinity of the order of lo1’ M-’, but also regained immunoreactivity (90 to 105%) comparable to that of hCG. None of the reconstituted hormones showed binding to the liver membrane receptor. How- ever, plasma clearance rate of radiolabeled hCG-da+d@ ( t l / z , 5 min) and hCG-a+d@ (t lIz, 15 min) was consider- ably faster than that of hCG (t lIz, 48 min). Organ distri- bution of the radioactivity showed that kidney, not the liver, was the major organ for the clearance of these derivatives. The data on the in vivo cellular responses of cAMP and progesterone production in corpus luteum cells and the in vivo response of depletion of ovarian ascorbic acid showed that the deglycosylation of one or both subunits led to a drastic drop in the biological activity of the hormone. While the deglycosylated hCG failed to stimulate hormonal activity, the mixed hybrids produced responses intermediate between those of the native and the deglycosylated hCG. In addition, the deglycosylated hCG was found to be a potent inhibitor of hCG activity in the above hormonal responses. These studies further demonstrate that the carbohydrate is not involved in the association of the subunits or in the antibody or receptor binding activity of the hormone although it is required for the expression of the hor- monal activity.
* This work was supported by United States Public Health Grant R01-HD-08766 and World Health Organization Grant H9/181/337. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore he hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
be sent. *‘To whom all correspondence regarding this manuscript should
Previous investigations from our laboratory have examined in vitro (1-3) and in vivo (4, 5 ) the role of carbohydrate in hCG.’ While the sequential removal of carbohydrate from hCG did not impair its receptor binding properties in vitro, its ability to stimulate cAMP and steroidogenesis in rat Leydig cells (1) and granulosa cells (2, 3) was considerably reduced. Similar results were obtained in vivo studies carried out with chemically modified analogs of asialo-hCG (4, 5). In all the hCG derivatives thus far tested, the carbohydrate was either partially removed or was modified. The sequential treatment with exoglycosidases hydrolyzed less than 60% carbohydrate from intact hCG or subunits, while the modifications removed no more than 30% carbohydrate, predominantly sialic acid (4). In the present work, the complete removal of carbohydrate has been attempted by the application of TFMS to the sub- units of hCG. This approach has enabled us to investigate the involvement of carbohydrate in the subunit interaction, in addition to its effect on the in vitro and in vivo biological properties of the hormone such as receptor binding, cAMP stimulation, steroidogenesis, plasma half-life, and ability to cause ovarian ascorbic acid depletion. Furthermore, by pre- paring the hybrids of the deglycosylated subunits with the corresponding native subunits, the role of carbohydrate in the individual subunits has been investigated. A preliminary re- port of this work has previously been made (6).
MATERIALS AND METHODS AND RESULTS2
Immunological and Receptor Binding Activity of Purified Reconstituted Deglycosylated and Hybrid hCG Derivatiues- Immunoreactivities of hCG and the reconstituted analogs were determined by homologous radioimmunoassay. As shown in Fig. 8, all the derivatives exhibited parallel dose- response curves in a lZ5I-hCG:anti-hCG radioimmunoassay system. Relative immunological potencies of various hormone analogs were determined from the dosage for 50% inhibition (IDSo) and are given in Table IV. Within experimental error, all derivatives had essentially identical immunoreactivities
’ The abbreviations used are: hCG, human chorionic gonadotropin or human choriogonadotropin; hCG-dol and hCG-dD, deglycosylated a- and 8-subunits of hCG; hCG-dol+dp, reconstituted deglycosylated hCG; hCG-da+P and hCG-a+dp, reconstituted mixed hybrids of hCG; TFMS, trifluoromethane sulfonic acid; PK-CAMP, protein kinase- b o y d CAMP.
Portion of this paper (including “Materials and Methods,” part of “Results,” Figs. 1-7, and Tables 1-111) are presented in miniprint a t the end of this paper. Miniprint is easily read with the aid of a standard magnifying glass. Full size photocopies are available from the Journal of Biological Chemistry, 9650 Rockville Pike, Bethesda, MD 20814. Request Document No. 82M-1692, cite the authors, and include a check or money order for $6.80 per set of photocopies. Full size photocopies are also included in the microfh edition of the Journal that is available from Waverly Press.
67

68
FIG. 8. Immunoreactivity ( A ) and receptor binding activity (B) of the reconstituted hCG derivatives. A, ho- mologous radioimmunoassay was per- formed using '251-hCG (30,000 cpm) and anti-hCG antiserum (1:30,000 dilution). Zero dose binding determined in the ab- sence of the hormone was about 9,OOO cpm and was taken as 100% binding. B, "'I-hCG (100,000 cpm) was incubated with crude testicular membrane (50 pg of protein) in the presence of varying concentrations of the hormones. In the absence of hCG, about 25,000 cpm were bound and assumed as 100% binding. Nonspecific binding was determined in the presence of 24 IU (2 pg) of hCG and was usually less than 2% of the total counts/min added.
Properties of Deglycosylated hCG
95 +3
90 +2
n + 1
$ 50 0 z
I- z & -
u" 20 - 1 3 (L
10 - 2
-3
I I I I
0.01 1
0.1 1.0 0.1 1 .o CONCENTRATION OF HORMONE [nM)
10.0
TABLE IV Comparison of immunoreactivity and testicular receptor binding
affinity of hCG and the reconstituted hCG derivatives .. ~ , ~~~ ~~
Immunoreactivity" Testicular receptor bind- ing affinityh
Hormone IDm x Relative rC, X 10"' Relative 10"' M potency' (mean f S.E.) potency'
hCG 0.97 100 1.60 k 0.17 100 hCG-a+p 1.05 92 2.00 f0.41 125 hCG-da+dp 1.15 hCG-da+P
85 1.40 k 0.19 88
hCGa+dp 1.10 0.90
88 2.05 f 0.45 128 107 1.60 f 0.18 100
The immunoreactivity of hormone derivatives was compared on the basis of ID,, (hormone concentration required for half-maximal inhibition) determined from the radioimmunoassay inhibition binding curves shown in Fig. 10. ' The apparent affinity constants (KO) of hormones for testicular
membrane receptor were calculated from Scatchard plots made from the equilibrium competitive binding assay shown in Fig. 10. Each value represents the mean k S.E. of two to three assays.
Immunoreactivity and receptor binding activity of hCG was as- sumed to be 100%.
ranging from 85 to 107% of hCG, suggesting the recovery of all antigenic sites on reassociation of the deglycosylated sub- units. Half-maximal inhibition was achieved at hormone con- centration around 10"' M.
Similarly, hCG and its reconstituted derivatives possessed identical receptor binding activities and gave parallel dose- response curves in the radioligand receptor assay shown in Fig. 8. The binding affinity constants (K,) of hCG and its analogs were estimated from Scatchard plots obtained from the competitive receptor binding data shown in Fig. 8. As is clear from Table IV, all reconstituted derivatives had identical binding constants, with K , values ranging from 1.4 to 2 X 10" M-', which are not significantly different from that of the native hCG (ICa, 1.6 X 10" "I). The above data clearly show that the carbohydrate of both or the individual subunit is not part of the antibody or the receptor binding site.
Stimulation of cAMP Production and Steroidogenesis by hCG and Its Reassociated Derivatives in Corpus Luteum Cells-In order to investigate the effect of deglycosylation of hCG in the post-receptor binding events of two cellular re- sponses, cAMP production and steroidogenesis were studied in luteum cells and are shown in Fig. 9. In the case of steroidogenesis, none of the reconstituted deglycosylated de- rivatives produced maximal response equivalent to that by 1 ng/ml of hCG. While the hybrids hCG-da+P and hCG-a+dP produced about 40% of the maximal response, the deglycosy- lated reconstituted derivative hCG-da+dp barely responded above the basal levels. In cAMP production, the mixed hy-
" C 15
w tn a c '"I
da +dB
0' I I 1 I
0 0.01 0.1 1.0 10.0 CONCENTRATION HORMONE/ DERIVATIVES
ng / ml
@ I P /
- 8 Jc.
4 /
hCG-do@
hCG-adB
hCG-do t dB
0 1 io 100 1000 CONCENTRATION HORMONE/ DERIVATIVES
ng /ml
FIG. 9. Stimulation of progesterone ( A ) and CAMP (B) pro- duction by hCG and its reconstituted derivatives in corpus luteum cells. A, varying amounts of hCG and its derivatives were incubated with 0.4 X lo6 cells/tube at 37 "C for 90 min. Both assays were done in triplicate and represent the mean f S.E. E, about 1 X lo6 cells were incubated in the presence of the indicated amounts of hormones at 37 "C for 90 min. The medium also contained 0.2 mM methyl 1,3-isobutylxanthine.

Properties of Deglycosylated hCG 69
brids of hCG at 1 p g / d produced only about 15% of the maximal response of hCG. In contrast, hCG-da+dp did not respond significantly above the basal level. In order to further investigate whether the inability of deglycosylated hCG to produce the cellular responses is also reflected in the levels of intracellular CAMP, PK-CAMP was estimated by a highly sensitive radioimmunoassay as described by Dufau et al. (15). As shown in Table V, hCG stimulated an increase in the level of PK-CAMP in a dose-response fashion at a hormone concen- tration of 0.1 to 100 ng/ml as previously reported (15). The deglycosylated hCG failed to produce any significant increase in the level of PK-CAMP up to a concentration of 1 p g / d , again showing that the derivative did not have any intrinsic hormonal activity.
Since hCG-&+dp had high binding affinity for the target tissue receptor without producing any hormonal response, it was assessed for its ability to inhibit hCG activity in the above cellular responses in luteum cells. The results of these studies are shown in Fig. 10 and Table V. The derivative caused dose- dependent inhibition of cAMP production and steroidogenesis by hCG. In case of cAMP production, the response produced by 10 and 100 ng of hCG was inhibited by 80 to 90% by an equal or a greater amount of the derivative. On the other hand, inhibition of the 50% steroidogenic response produced by 0.1 and 0.5 ng required 2 to 3-fold greater amount of the derivative. As is clear from Table V, the intracellular level of PK-CAMP measured in the above responses showed a corre- sponding inhibition by the hCG-da+dp. 2 pg of the deglyco- sylated derivative caused a decrease in the level of PK-CAMP from 305 f 11 fmol/tube, stimulated by 0.5 ng of hCG, to 220 rt 11 fmol/tube. The basal level of PK-CAMP in these studies was 188 5 7 fmol/tube.
Binding of 1251-labeled Deriuatives to Liver Plasma Mem- brane-In order to investigate the effect of deglycosylation of hCG on its uptake by the liver, direct binding studies using
I-labeled hCG derivatives and crude liver plasma mem- branes were performed as shown in Fig. 11. None of the deglycosylated and hybrid hCG derivatives bound to the liver membranes appreciably. As a reference, asialo-hCG was shown to bind specifically to the galactose receptor on liver membranes.
Plasma Clearance of 1251-labeled hCG, Deglycosylated hCG, and Mixed Hybrids in Mature Female Rats-Fig. 12 shows the plasma clearance curves of Iz5I-labeled hCG deriv- atives in rats, and Table VI lists the organ distribution of radioactivity 30 min after injection of the hormone. The
125
TABLE V Inhibition of hCG-induced intracellular PK-CAMP by the
reconstituted deglycosylated hCG, hCG-da+dp The results are expressed as mean f S.D. of three values.
Hormone Pk-CAMP fmol / loh cells
Control 188 f 7 hCG
0.1 ng 240 f 6 0.5 ng 305 f 11 10 ng 415 f 12 100 ng 430 f 15
100 ng lo00 ng
180 & 8 205 f 10
0.5 ng 240 f 12 2.0 ng 220 f 4
10 ng 210 f 5 100 ng 180 f 4
hCG-da+dp
hCG (0.5 ng) + hCG-da+dp
HCG (IO ng) + hCG-da+dp
t- z 0 (L
W
50
w 75 n t O . l n g hCG
I 1 I I I I I 0 0.5 1.0 1.5 2.00 10 100 1000
CONCENTRATION OF hCG-do+da (ng/ml)
FIG. 10. Inhibition of hCG stimulated progesterone ( A ) and CAMP (B) production in luteum cells by hCG-da+d/?. The derivative was mixed with hCG before addition to the cells. No hormone was added to the control tube. The tubes for cAMP studies also contained 0.2 m~ methyl 1,3-isobutylxanthine. The results are expressed as mean f S.E.
a
0 1 2 3 4 5 LIVER MEMBRANE PROTEIN (mg)
FIG. 11. Binding of hCG and its reconstituted derivatives to liver plasma membranes. To 5 ng (l00,ooO cpm) of 1Z51-labeled hormone were added varying amounts of crude liver membranes, and these were incubated at 37 "C for 1 h as described under "Materials and Methods."
'9
. !- z W u A hCG
W a a I I I
10 20 30 TIME (rnin)
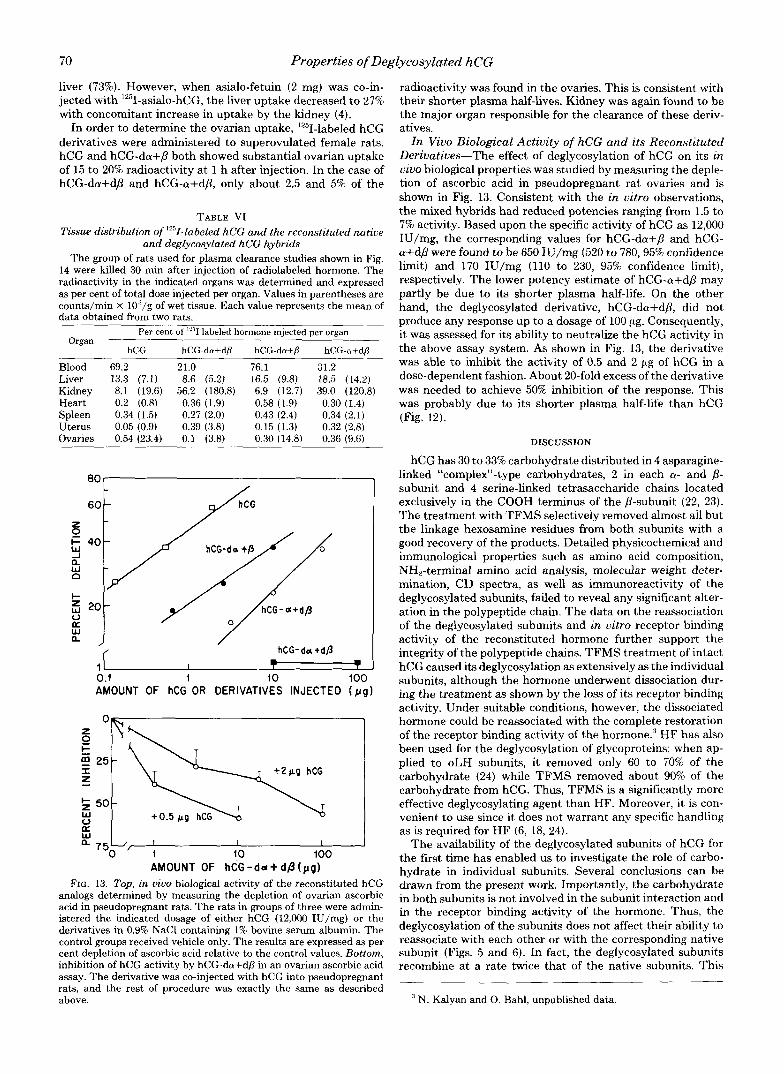
FIG. 12. Plasma clearance curves of hCG and its reconsti- tuted derivatives in female rats. About 0.5 X 10' cpm of Iz5I- labeled and 100 ng of unlabeled hormone were injected via tail vein, and blood samples were taken from the opposite tail vein at indicated time intervals. Radioactivity in 100 p1 of plasma was counted and expressed as per cent of the injected dosage/ml of plasma. Eachpoint represents the average from two animals.
plasma clearance rate of mixed hybrid containing native p- subunit, hCG-da+P (tIl2, 50 min), was found to be comparable to that of hCG (tIl2, 48 min). However, the plasma half-lives of hCG-da+dB and hCG-a+dp were considerably lower, with tllr of 5 and 13 min respectively, than that predicted from the in vitro liver receptor binding data. Distribution of the radio- activity in various organs showed that kidney contained about 56 and 39% radioactivity. As a reference, asialo-hCG with terminal galactosyl residue was found to be concentrated in
70 Properties of Deglycosylated hCG
liver (73%). However, when asialo-fetuin (2 mg) was co-in- jected with '2sI-asialo-hCG, the liver uptake decreased to 27% with concomitant increase in uptake by the kidney (4).
In order to determine the ovarian uptake, '251-labeled hCG derivatives were administered to superovulated female rats. hCG and hCG-da+p both showed substantial ovarian uptake of 15 to 20% radioactivity at 1 h after injection. In the case of hCG-da+dfi and hCG-a+db, only about 2.5 and 5% of the
TABLE VI Tissue distribution of '"I-labeled hCG and the reconstituted native
and deglycosylated hCG hybrids The group of rats used for plasma clearance studies shown in Fig.
14 were killed 30 min after injection of radiolabeled hormone. The radioactivity in the indicated organs was determined and expressed as per cent of total dose injected per organ. Values in parentheses are counts/min X 1O"/g of wet tissue. Each value represents the mean of data obtained from two rats.
Organ Per cent of '"1 labeled hormone injected per organ
hCG hCG-da+dp hCG-da+P hCG-a+d@
Blood 69.2 21.0 76.1 31.2 Liver 13.3 (7.1) 8.6 (5.2) 16.5 (9.8) 18.5 (14.2) Kidney 8.1 (19.6) 56.2 (180.8) 6.9 (12.7) 39.0 (120.8) Heart 0.2 (0.8) 0.36 (1.9) 0.58 (1.9) 0.30 (1.4) Spleen 0.34 (1.5) 0.27 (2.0) 0.43 (2.4) 0.34 (2.1) Uterus 0.05 (0.9) 0.39 (3.8) 0.15 (1.3) 0.32 (2.8) Ovaries 0.54 (23.4) 0.1 (3.8) 0.30 (14.8) 0.36 (9.6)
i l I t ? I 0.1 1 10 100 AMOUNT OF hCG OR DERIVATIVES INJECTED ( p g )
z t 0
E I
50- W 0
W a
750 '/ ; I I
10 100 AMOUNT OF hCG - d d + dfi ( p g )
FIG. 13. Top, in u i w biological activity of the reconstituted hCG analogs determined by measuring the depletion of ovarian ascorbic acid in pseudopregnant rats. The rats in groups of three were admin- istered the indicated dosage of either hCG (12,000 IU/mg) or the derivatives in 0.9% NaCl containing 1% bovine serum albumin. The control groups received vehicle only. The results are expressed as per cent depletion of ascorbic acid relative to the control values. Bottom, inhibition of hCG activity by hCG-da+dp in an ovarian ascorbic acid assay. The derivative was co-injected with hCG into pseudopregnant rats, and the rest of procedure was exactly the same as described above.
radioactivity was found in the ovaries. This is consistent with their shorter plasma half-lives. Kidney was again found to be the major organ responsible for the clearance of these deriv- atives.
I n Vivo Biological Activity of hCG and its Reconstituted Derivatives-The effect of deglycosylation of hCG on its in vivo biological properties was studied by measuring the deple- tion of ascorbic acid in pseudopregnant rat ovaries and is shown in Fig. 13. Consistent with the in vitro observations, the mixed hybrids had reduced potencies ranging from 1.5 to 7% activity. Based upon the specific activity of hCG as 12,000 IU/mg, the corresponding values for hCG-da+P and hCG- a+dP were found to be 650 IU/mg (520 to 780,95% confidence limit) and 170 IU/mg (110 to 230, 95% confidence limit), respectively. The lower potency estimate of hCG-a+dB may partly be due to its shorter plasma half-life. On the other hand, the deglycosylated derivative, hCG-da+dP, did not produce any response up to a dosage of 100 pg. Consequently, it was assessed for its ability to neutralize the hCG activity in the above assay system. As shown in Fig. 13, the derivative was able to inhibit the activity of 0.5 and 2 pg of hCG in a dose-dependent fashion. About 20-fold excess of the derivative was needed to achieve 50% inhibition of the response. This was probably due to its shorter plasma half-life than hCG (Fig. 12).
DISCUSSION
hCG has 30 to 33% carbohydrate distributed in 4 asparagine- linked "complex"-type carbohydrates, 2 in each a- and p- subunit and 4 serine-linked tetrasaccharide chains located exclusively in the COOH terminus of the P-subunit (22, 23). The treatment with TFMS selectively removed almost all but the linkage hexosamine residues from both subunits with a good recovery of the products. Detailed physicochemical and immunological properties such as amino acid composition, NH,-terminal amino acid analysis, molecular weight deter- mination, CD spectra, as well as immunoreactivity of the deglycosylated subunits, failed to reveal any significant alter- ation in the polypeptide chain. The data on the reassociation of the deglycosylated subunits and in vitro receptor binding activity of the reconstituted hormone further support the integrity of the polypeptide chains. TFMS treatment of intact hCG caused its deglycosylation as extensively as the individual subunits, although the hormone underwent dissociation dur- ing the treatment as shown by the loss of its recept,or binding activity. Under suitable conditions, however, the dissociated hormone could be reassociated with the complete restoration of the receptor binding activity of the hormone:' HF has also been used for the deglycosylation of glycoproteins; when ap- plied to oLH subunits, it removed only 60 to 70% of the carbohydrate (24) while TFMS removed about 90% of the carbohydrate from hCG. Thus, TFMS is a significantly more effective deglycosylating agent than HF. Moreover, it is con- venient to use since it does not warrant any specific handling as is required for HF (6, 18, 24).
The availability of the deglycosylated subunits of hCG for the fist time has enabled us to investigate the role of carbo- hydrate in individual subunits. Several conclusions can be drawn from the present work. Importantly, the carbohydrate in both subunits is not involved in the subunit interaction and in the receptor binding activity of the hormone. Thus, the deglycosylation of the subunits does not affect their ability to reassociate with each other or with the corresponding native subunit (Figs. 5 and 6). In fact, the deglycosylated subunits recombine at a rate twice that of the native subunits. This
N. Kalyan and 0. Bahl, unpublished data

Properties of Deglycosylated hCG 71
probably is due to the elimination of electronegative charge by the loss of sialic acid during deglycosylation. I t is worth noting that all the reconstituted hCG derivatives bind to the receptor with a high affinity of the order of 10" M-', compa- rable to that of hCG (Fig. 8 and Table IV). In addition, the deglycosylation of one or both subunits leads to a drastic drop in the biological activity of the hormone. Unlike the native hormone, the deglycosylated hCG fails to stimulate the in uitro cellular responses such as cAMP and progesterone pro- duction in rat luteum cells (Fig. 9). In uiuo, in rats, the biological response of ovarian ascorbic acid depletion is also impaired (Fig. 13). The mixed hybrids of deglycosylated and native subunits produce hormonal responses intermediate be- tween those of the native and the deglycosylated hCG. The deglycosylated hCG is a potent inhibitor of cAMP and pro- gesterone stimulation (Fig. 10). The inhibitory effect was much more pronounced in the in uitro system than in uiuo. This probably was due to its rapid clearance from plasma by kidney and not by liver (Figs. 11 and 12 and Table VI). The reasons for the rapid clearance by kidney are not clear at present. It may be due partly to the reduced size and charge. I t is interesting to note that the deglycosylated hCG does not bind in uitro or in vivo to the liver which has receptors for N - acetylhexosamines (25). It appears that the linkage hexosa- mines are not accessible to the cellular receptors due to steric effects.
Since the deglycosylated hormone binds to the receptor as well as hCG or better, the loss of carbohydrate does not seem to change the conformation of the receptor binding site of the hormone. This, however, does not rule out the possibility of the deglycosylated hormone-receptor complex undergoing a conformational change which may lead to the loss of its ability to form microclusters in the membranes and/or its ability to stimulate the subsequent steps in the hormone action as the activation of GTP or nucleotide binding protein and adenylate cyclase (26,27). Obviously, the precise site of the carbohydrate action in the currently available model of polypeptide hor- mone action is not known. Our preliminary data, however, indicate that the removal of carbohydrate from hCG affects an event(s) prior to the activation of nucleotide binding pro- tein and adenylate cyclase but on or after the hormone- receptor complex formation (28).
In summary, the present work clearly establishes the effi- cacy of TFMS in the selective removal of carbohydrate from hCG or hCG subunits without any detectable adverse effect on the polypeptide chains. It further demonstrates that the carbohydrate is not involved in the association of the subunits or in the antibody or receptor binding activity of the hormone. Finally, it is clearly shown that the deglycosylation of hCG results not only in the loss of biological activity, but it also imparts antagonistic properties to the hormone. The degly- cosylated hCG thus is important from both the fundamental and clinical point of view. While the derivative can serve as a probe in the study of the hormone action at a cellular and
molecular level, it has the potential of application to fertility regulation.
Acknowledgments-We wish to express our appreciation to James Stamos for preparing the figures and to Ursula Brunn for typing the manuscript.
REFERENCES 1. Moyle, W. R., Bahl, 0. P., and Marz, L. (1975) J. Biol. Chem.
2. Channing, C. P., and Bahl, 0. P. (1978) Biol. Reprod. 17,707-711 3. Channing, C. P., and Bahl, 0. P. (1978) Endocrinology 103,341-
4. Madnick, H. W., Kalyan, N. K., Segal, H. L., and Bahl, 0. P.
5. Kalyan, N. K., Lippes, H. A., and Bahl, 0. P. (1982) J . Biol.
6. Kalyan, N. K., and Bahl, 0. P. (1981) Biochem. Biophys. Res.
7. Carlsen, R. B., Bahl, 0. P., and Swaminathan, N. (1973) J. Bwl.
8. Lehnhardt, W. F., and Winzler, R. J. (1968) J . Chromatogr. 34,
9. Warren, L. (1959) J. Biol. Chem. 234, 1971-1975
250,9163-9169
348
(1981) Arch. Biochem. Biophys. 212,432-442
Chem. in press
Commun. 102,1246-1253
Chem. 248,6810-6827
471-479
10. Swaminathan, N., and Bahl, 0. P. (1970) Biochem. Biophys. Res.
11. Pandian, M. R., and Bahl, 0. P. (1977) Arch. Biochem. Biophys.
12. Ashwell, G., and Morell, A. G . (1974) Adu. Enzymol. Rel. Areas
13. Achord, D. T., Brot, F. E., and Sly, W. S. (1977) Biochem.
14. Ferguson, K. A. (1964) Metab. Clin. Ezp. 13, 985-1002 15. Dufau, M. L., Tsuruhara, T., Horner, K. A., Podesta, E., and Catt,
K. J. (1977) Proc. Natl. Acad. Sci. U. S. A. 74, 3419-3423 16. Harper, J. F., and Brooker, G. (1975) J. Cyclic Nucleotide Res. 1,
207-218 17. Mindlin, R. L., and Butler, A. M. (1933) J. Biol. Chem. 122, 673-
686 18. Edge, A. S. B., Faltynek, C. R., Hof, L., Reichert, L. E., Jr., and
Weber, P. (1981) Anal. Biochem. 118, 131-137 19. Bellisario, R., Carlsen, R. B., and Bahl, 0. P. (1973) J. Biol. Chem.
248,6796-6809 20. Holladay, L. A., and Puett, D. (1975) Arch. Biochem. Biophys.
171, 708-720 21. Bewley, T. A., Sairam, M. R., and Li, C. H. (1972) Biochemistry
22. Kessler, M. J., Reddy, M. S., Shah, R. H., and Bahl, 0. P. (1979)
23. Kessler, M. J., Mise, T., Ghai, R. D., and Bahl, 0. P. (1979) J .
24. Sairam, M. R. (1980) Arch. Biochem. Biophys. 204, 199-206 25. Kawasaki, T., and Ashwell, G . (1977) J . Biol. Chem. 252, 6536-
6543 26. Amsterdam, A., Berkowitz, A,, Nimrod, A., and Kohen, F. (1980)
Proc. Natl. Acad. Sci. U. S. A. 77,3440-3444 27. Abramowitz, J., Iyengar, R., and Birnbaumer, L. (1980) in Func-
tional Correlates of Hormone Receptors in Reproduction (Ma- hesh, V. B., Muldoon, T. G., Saxena, B. B., and Sadler, w . A,, eds) pp. 335-365, Elsevier/North-Holland, New York
28. Thotakura, N. R., and Bahl, 0. P. (1982) Biochem. Biophys. Res. Commun., 108, 399-405
Commun. 40,422-427
182,420-436
Md. Biol. 41.99-128
Biophys. Res. Commun. 77,409-415
11,932-936
J. Biol. Chem. 254,7901-7908
Biol. Chem. 254, 7909-7914

72 Properties of Deglycosylated hCG

Properties of Deglycosylated hCG 73
+
A 1 2 3 4 5 6 7 8 - "
1 .o
0.5
20 40 60 00 FRACTION NUMBER
Y E
0 1.8-
P Y
s 1.7- hCG-da CI,
1.6 I I I I I
6 8 10 12 14 16 PERCENT GEL CONCENTRATION
r
hCG-dafiYSOZYME
1 .o I I I
15 20 25 30 35 40 ( M ) 'I3

74 Properties of Deglycosylated hCG
WAVELENGTH (nml
Cl hCG- da + B Dl hCG-n +d/3
1 10 100 1 io 100 CONCENTRATION OF HORMONE (nM)
100
z P c 80 &
5 60 m
w
40 v Y
a 20
0 5 1.0 1.5 2.0 I
, zo FRACTION NUMBER
hCC-aihCG-B hCGdCllhCG-dB hCC-dCl+hCG-B hCGa+hCG-dB
1.3 3 . 0 1.7 1.5
37 1 7 28.5 31.5





























