Embed Size (px)
Citation preview

GENOMICS 34, 205–212 (1996)ARTICLE NO. 0267
The Sp4H Deletion May Contain a New Locus Essentialfor Postimplantation Development
JANE FLEMING,* ANGELA PEARCE,* STEPHEN D. M. BROWN,† AND KAREN P. STEEL*,1
*MRC Institute of Hearing Research, University Park, Nottingham, NG7 2RD, United Kingdom; and†MRC Mouse Genome Centre, Harwell, Oxon OX11 0RD, United Kingdom
Received January 3, 1996; accepted March 27, 1996
different mutations of this locus have been described:Sp4H is a semi-dominant mutation that maps to Sp, Spr, Sp1H, Sp2H, Sp4H, Spd, SpJ, Sp2J, and Sp3J
mouse chromosome 1. Heterozygous mice exhibit (Auerbach, 1954; Russell, 1947; Beechey and Searle,white spotting of the belly, whereas the fate of the ho- 1986; Dickie, 1964). SpJ, Sp2J, and Sp3J are now extinct.mozygous embryos is unknown. We have previously All heterozygous mutant mice are characterized byshown that the entire coding region of the Pax3 gene spotting of the belly, while homozygous mice exhibitis deleted in the Sp4H mutant. In this study, we have varying degrees of neural tube and neural crest cellanalyzed the fate of the Sp4H homozygous embryos. No defects. Alterations in the Pax3 gene have been demon-Sp4H homozygotes were detected by Southern blot or strated to be responsible for the Splotch mutations.PCR analysis in 82 E9–E13-day embryos. We have also This gene has been reported to be expressed at betweendocumented a significant increase in the number of
8 and 17 days of gestation in the dorsal neural tube,resorption sites in Sp4H heterozygous matings com-the tissue affected in Splotch mice (Goulding et al.,pared to control litters. Sections of the resorption sites1991), and RT-PCR has been used to show that Pax3(moles) suggest that postimplantation development istranscripts are present from 6.5 days onward (D. Hen-arrested prior to gastrulation. We have mapped thederson, S. Conway, and A. J. Copp, London, pers.extent of the deletion to a maximum of 1.53 { 0.6 cMcomm., 1996). The two DNA binding domains of theusing markers flanking the Pax3 locus. Four anony-gene, the paired and the homeodomain, have both beenmous markers, D1Mit215, D1Mit253, D1Mit332, andshown to bind to the Drosophila even-skipped gene pro-D1McG156, have been shown to be deleted in the Sp4H
mutation. Further nondeleted markers have been used motor in vitro, although target sequences for Pax3 into extend the linkage map of this region. A total of 22 vivo are unknown (Chalepakis et al., 1994a,b).loci were analyzed in a Splotch intraspecific back- To date, three unique molecular mutations in thecross. Using these data and deletion mapping data, we Spd, Sp2H, and Sp alleles and two deletions around thepredict the following order of markers: (D1Mit46, Vil)– Pax3 locus in Spr and Sp4H have been identified. In the(D1Mit79)–(D1Mit132, D1McG153)–(D1Mit332)–(D1- least affected allele, the Spd mutants, Spd/Spd embryosMcG156, D1Mit253, D1Mit215, Pax3)–(D1Mit134)–(D1- can survive to birth with evidence of spina bifida onlyMit8, D1Mit9, D1Mit44, D1Ler3, D1Mit183)–(D1Mit53, in the lumbrosacral region and only minor defects inD1Mit82, D1Mit182)–(Bcl2). As the deletion is large the spinal ganglia (Moase and Trasler, 1989, 1992).enough to include other genes, and it seems that dele-
These mice carry a single basepair substitution (G totion of Pax3 is not likely to account for the early deathC) leading to an amino acid change of Gly to Arg in theof the embryos, we suggest that another developmen-paired box region (Vogan et al., 1993), which alters thetally important gene may be deleted in the Sp4H mousecharge of the paired domain and may lower the levelmutant and that this may be responsible for the earlyof Pax3 mRNA as reported in 12-day embryosdeath of the homozygous mutant embryos. q 1996(Goulding et al., 1993). In the Sp2H, Sp1H, and Sp mouseAcademic Press, Inc.
mutants, homozygous embryos die at 13–16 days ofgestation with spina bifida, exencephaly, and meningo-coele. Sp/Sp mice also exhibit dysgenesis of neuralINTRODUCTIONcrest cell-derived structures (Auerbach, 1954), Sp1H/Sp1H mice have disturbance of muscle development inThe Splotch mouse mutation is a semi-dominant dis-the limb buds (Franz et al., 1993), and Sp1H/Sp1H andorder mapped to mouse chromosome 1 between the vil-Sp2H/Sp2H mice have malformations in structures oflin (Vil ) and leaden (ln) loci (Hillyard et al., 1992). Ninethe heart (Franz, 1989; Copp, 1995). Sp2H mice havea 32-bp deletion in the homeodomain region of Pax31 To whom correspondence should be addressed. Telephone: /44
115 922 3431. Fax: /44 115 942 3710. (Epstein et al., 1991), which is believed to give rise to
2050888-7543/96 $18.00
Copyright q 1996 by Academic Press, Inc.All rights of reproduction in any form reserved.
AID Genom 4118 / 6r16$$$461 05-06-96 07:10:04 gnma AP: Genomics

FLEMING ET AL.206
Genotyping embryos. Genomic DNA from 9- to 13-day embryosa truncated form of the Pax3 protein product with DNAand from embryonic and extraembryonic membranes minus the ec-binding possible at the paired box alone. The Sp1H mu-toplacental cone of 7.5-day embryos was prepared using phenol/chlo-
tation has since been shown to carry a mutation identi- roform extraction or by the method of Estibeiro (Estibeiro et al.,cal to that in Sp2H (Vogan et al., 1993). In the original 1990). Southern analysis was carried out using a Pax3 gene probe
and an Omp gene probe as a loading control, as described previouslySplotch, Sp, mutation, a 10- to 11-bp rearrangement at(Goulding et al., 1993). Some embryos were genotyped by PCR tothe intron 3/exon 4 border produces aberrant splicing ofdetect markers that were known to be deleted in the Sp4H mutation,the Pax3 gene. This leads to the production of mainlyalong with nondeleted markers as a control (see Results for details).
shorter transcripts of Pax3 lacking exon 4 (Goulding etDeletion analysis. Sp4H// mice were crossed to CBA/Ca, C57BL/
al., 1993). Exon 4 spans 14 amino acids in the carboxy 10J, and BALB/c inbred strains to produce F1 progeny. Parentalterminus of the paired box and includes the octapeptide DNA from tail and pinna skin was then typed using PCR or Southern
analysis for loci listed in Table 1 to identify polymorphisms. DNAregion. In the most severely affected mutant, Spr, ho-from the F1 progeny from the relevant cross was then typed to iden-mozygous embryos do not implant and heterozygoustify any loss of bands that would indicate that the marker was deletedmice are smaller than control littermates. These mice in the Sp4H stock. Allele sizes are given in Table 1.
have a 14- to 16-cM cytogenetically visible deletion that Deletion analysis of Pax3, Vil, and Bcl2 has been described pre-extends from the Vil to the Akp3 loci, including the viously (Goulding et al., 1993). The anonymous markers D1McG153
and D1McG156 were 2.3- and 0.23-kb clones isolated from a genomicentire coding region of the Pax3 gene plus several geneslibrary constructed from microdissected fragments from mouse chro-important in the production of the cytoskeleton (Evansmosome 1 (cytogenetic bands C2–C5), cloned into the EcoRI site ofet al., 1988; Beechey and Searle, 1986). Similar types l phage insertion vector NM1149 (Vidal et al., 1992). D1Ler3 was a
of mutation have been described for the human homo- 0.8-kb EcoRI/HindIII fragment of anonymous genomic DNA clonedinto pTZ18R (LeRoy et al., 1992). These three markers were usedlogue of the Splotch mutation, Waardenburg syndromefor Southern analysis of parental and F1 mouse DNA. The remainingtype I (WS1) (Tassabehji et al., 1994; Lalwani et al.,loci listed in Table 1 were typed using PCR primers flanking micro-1995). The only documented case of a homozygoussatellite regions using standard techniques (Dietrich et al., 1994).
PAX3 mutation in a WS1 patient is an amino acid sub-Linkage analysis. Sp// mice on a C57BL/6J background were
stitution in the paired box of the PAX3 protein (Zloto- crossed to the CBA/Ca inbred strain. F1 mice showing a clear bellygora et al., 1995). This may produce an effect similar spot were then backcrossed to CBA/Ca, and the progeny with a belly
spot were typed. Complete analysis of Pax3, Vil, and Bcl2 has beento that of the Spd mouse mutation, where reduced Pax3reported previously in 117 Sp// backcross progeny (Goulding et al.,DNA binding activity is predicted (Goulding et al.,1993). The backcross has since been extended to 412 mice, and the1993; Underhill et al., 1995). It is interesting to notegenotyping has included /// mice. Due to the variable penetrance
that the individual appears to have had sufficient PAX3 of the belly spot (approximately 61% of the expected number of miceactivity to escape severe neural tube defects and has carrying the Sp allele had a belly spot; Goulding et al., 1993), all of
the progeny were genotyped using Southern analysis with a Pax3survived past birth.probe as described previously (Goulding et al., 1993). Other markersThe Sp4H allele arose in an X-irradiated C3H/HeH 1used for linkage analysis are described under Results, and details101/H F1 hybrid male. An increase in the number of of allele sizes are given in Table 1. As Sp was on a C57BL/6J back-
resorption sites in Sp4H// 1 Sp4H// matings was re- ground, the allele sizes for markers derived from this stock are veryported, suggesting that the homozygous embryos died similar to those given in the C57BL/10J column of Table 1.postimplantation (Cattanach et al., 1993). We have pre-viously demonstrated that the entire coding region of RESULTSthe Pax3 gene is deleted (Goulding et al., 1993), but noother genes or markers so far studied have been shown Appearance of Embryosto be deleted (Goulding et al., 1993; Peters et al., 1995).In this report, we document the fate of the Sp4H mu- In an initial study into the viability of Sp4H homozy-
gote embryos, litters from Sp4H// 1 Sp4H// matings attants and define in greater detail the extent of the dele-tion on chromosome 1. 10–13 days of gestation were examined under the light
microscope for evidence of neural tube defects. In 13litters containing 92 embryos no abnormal embryosMATERIALS AND METHODSwere found, even though embryos exhibiting spina bi-fida and exencephaly have been observed around thisMice. Sp4H// mice on a C3H/HeH/101/H genetic background to-time in both Sp// 1 Sp// and Sp2H// 1 Sp2H// mat-gether with the parental C3H/HeH and 101/H inbred mice were origi-
nally obtained from the MRC Radiobiology Unit (Harwell, UK). Sp ings in this laboratory and elsewhere (Goulding et al.,mice on a C57BL/6J background were obtained from the Jackson 1993). In addition, there was a significant excess inLaboratory (Bar Harbor, ME). Timed matings were used to obtain the number of resorption sites when Sp4H// 1 Sp4H//embryos. Both Sp4H// 1 Sp4H// and /// 1 /// matings were used
matings were compared to controls on the same geneticto generate embryos. Only Sp4H// parents with distinct white bellybackground (Table 2). In litters at 10–13 days of gesta-spots were used for matings. Control parents had no spotting on the
belly, feet, or tails. tion there was a significantly increased resorption rateHistology. Resorption sites (moles) were either dissected out and for the Sp4H heterozygote matings at 27%, compared
examined under a dissecting microscope or removed and fixed in 4% with the control rate of just 11% (x2Å 6.24, PÅ 0.0125).paraformaldehyde, dehydrated, cleared, and embedded in paraffin The resorption rate was relatively high at 11%, but thewax for sectioning, using standard protocols. Five moles were serial-
total number we examined was relatively small. Theresectioned at 10 mm and stained with hematoxylin and eosin for exam-ination. is no significant difference between the observed num-
AID Genom 4118 / 6r16$$$461 05-06-96 07:10:04 gnma AP: Genomics
Sp4H DELETION 207
TABLE 1
Allele Sizes and Deletion Status of Markers Flanking the Pax3 Locus
CBA allele size C57 allele size Sp4H allele size RFLP or PCR Marker deleted(bp) (bp) (bp) analysis RFLP enzyme in Sp4H
Vil 18,000 7,500 RFLP XbaI NoD1Mit46 220 246 250 PCR NoD1Mit79 166 170 172 PCR NoD1Mit132 134 144 166 PCR NoD1Mit134 138 140 140 PCR NoD1McG153 1,050 1,500 1,500 RFLP RsaI NoD1Mit332 110 124 124 PCR YesD1McG156 2,200 2,000 2,200 RFLP MspI YesD1Mit253 130 160 160 PCR YesPax3 5,400 3,500 RFLP BamHI Yes
2,300D1Mit215 150 150 156 PCR YesD1Mit8 220 290 290 PCR NoD1Mit9 140 156 156 PCR NoD1Mit44 206 198 198 PCR NoD1Mit183 130 126 126 PCR NoD1Ler3 10,000 12,000 11,000 RFLP TaqI NoD1Mit53 115 131 131 PCR NoD1Mit182 138 150 150 PCR NoD1Mit82 196 240 124 PCR NoBcl2 132 154 PCR NoD1Mit216 126 126 120 PCR NoD1Mit162 134 130 130 PCR NoD1Mit49 140 140 134 PCR NoD1Mit80 146 146 140 PCR NoD1Mit83 180 166 166 PCR No
Note. Progeny typed are from Sp4H// 1 CBA/Ca or Sp4H 1 C57B1/10J crosses. Most of these markers were also used for the linkageanalysis of Sp// on a C57BL/6J background backcrossed to CBA/Ca. Markers D1Mit216, D1Mit215, D1Mit49, D1Mit80, and D1Mit83 werenot included in the linkage analysis.
ber of resorptions (32) and the number predicted based was more advanced in development, had a roundershape, and appeared to have extraembryonic mem-on a 25% loss of Sp4H homozygotes plus 11% of the
remainder (31 / 10.2 Å 41.2) (x2 Å 3.09, P Å 0.1). branes. Two of the five moles, both at 11 days of gesta-tion when sectioned, appeared to be undergoing resorp-Five moles from Sp4H heterozygote matings at 10–
11 days of gestation were sectioned. Four were pear- tion of tissue in the presumptive embryo-derived spacebecause of the presence of blood cells and debris (datashaped and had evidence of an ectoplacental cone and
implantation site. The fifth (at 10 days of gestation) not shown).Embryos at 7.5 days of gestation were examined to
investigate the time of death of the resorbing embryosTABLE 2from heterozygous matings. From a total of 41 decidual
Examination of Embryos for Neural Tube Defects swellings dissected from five pregnant females, only24 embryos were recovered, suggesting again that aAge Total No. of Abnormal Normal Resorption % Resorptionsignificant number of embryos are failing to develop.(d.p.c.) implantations embryos embryos sites sites
No evidence of any embryonic material was obvious inExamination of embryos from Sp4H// 1 Sp4H// matings the empty decidual tissue examined by dissection.
for neural tube defects
13 27 0 24 3 11Genotype of Embryos12 11 0 9 2 18
11 20 0 14 6 3010 66 0 45 21 33 DNA from 82 of a total of 97 embryos aged between
Totals 124 0 92 32 27 9 and 13 days of gestation was examined for evidence ofhomozygosity for the Sp4H deletion. Forty-nine embryos
Examination of embryos from /// 1 /// (control mice) matings were typed by Southern analysis to detect the presencefor neural tube defects or absence of the Pax3 gene using a cDNA probe, and
13 24 0 17 6 26 Omp was used as a positive control (data not shown).11 9 0 8 1 11 A further 33 embryos were genotyped using PCR prim-10 58 0 55 3 5
ers for DNA markers (D1Mit253 or D1Mit332) knownTotals 90 0 80 10 11 to be deleted around the Pax3 region in the Sp4H mu-
AID Genom 4118 / 6r16$$$461 05-06-96 07:10:04 gnma AP: Genomics
FLEMING ET AL.208
tant mice, and the nondeleted markers (D1Mit8 orD1Mit9) were used as a positive control (see later fordetails of deleted and nondeleted markers). None of the82 embryos tested were homozygous for the deletion.DNA was also extracted from some of the 7.5-day em-bryos, and these were genotyped using PCR as outlinedabove. Eight of these young embryos have been success-fully genotyped with at least one deleted and one non-deleted marker, and again no homozygous mutantshave been identified (data not shown).
Extent of the Sp4H Deletion
The extent of the Sp4H deletion around the Pax3 locushas been examined by deletion mapping using micro-satellite markers and anonymous DNA probes (Table1). Polymorphisms were identified between the Sp4H
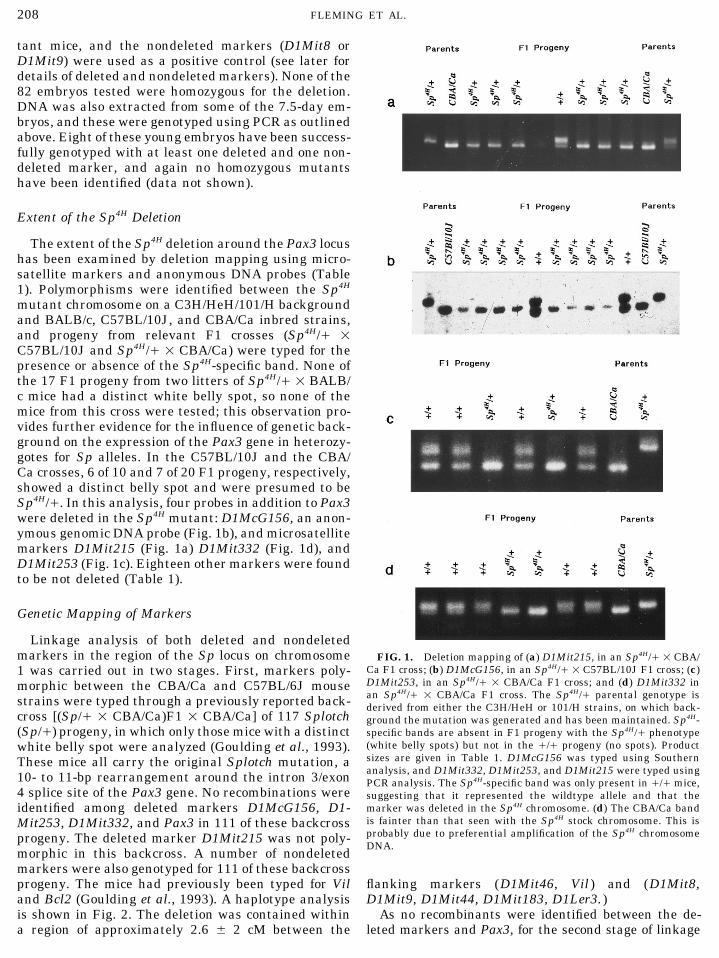
mutant chromosome on a C3H/HeH/101/H backgroundand BALB/c, C57BL/10J, and CBA/Ca inbred strains,and progeny from relevant F1 crosses (Sp4H// 1C57BL/10J and Sp4H// 1 CBA/Ca) were typed for thepresence or absence of the Sp4H-specific band. None ofthe 17 F1 progeny from two litters of Sp4H// 1 BALB/c mice had a distinct white belly spot, so none of themice from this cross were tested; this observation pro-vides further evidence for the influence of genetic back-ground on the expression of the Pax3 gene in heterozy-gotes for Sp alleles. In the C57BL/10J and the CBA/Ca crosses, 6 of 10 and 7 of 20 F1 progeny, respectively,showed a distinct belly spot and were presumed to beSp4H//. In this analysis, four probes in addition to Pax3were deleted in the Sp4H mutant: D1McG156, an anon-ymous genomic DNA probe (Fig. 1b), and microsatellitemarkers D1Mit215 (Fig. 1a) D1Mit332 (Fig. 1d), andD1Mit253 (Fig. 1c). Eighteen other markers were foundto be not deleted (Table 1).
Genetic Mapping of Markers
Linkage analysis of both deleted and nondeletedmarkers in the region of the Sp locus on chromosome FIG. 1. Deletion mapping of (a) D1Mit215, in an Sp4H// 1 CBA/
Ca F1 cross; (b) D1McG156, in an Sp4H// 1 C57BL/10J F1 cross; (c)1 was carried out in two stages. First, markers poly-D1Mit253, in an Sp4H// 1 CBA/Ca F1 cross; and (d) D1Mit332 inmorphic between the CBA/Ca and C57BL/6J mousean Sp4H// 1 CBA/Ca F1 cross. The Sp4H// parental genotype isstrains were typed through a previously reported back- derived from either the C3H/HeH or 101/H strains, on which back-
cross [(Sp// 1 CBA/Ca)F1 1 CBA/Ca] of 117 Splotch ground the mutation was generated and has been maintained. Sp4H-(Sp//) progeny, in which only those mice with a distinct specific bands are absent in F1 progeny with the Sp4H// phenotype
(white belly spots) but not in the /// progeny (no spots). Productwhite belly spot were analyzed (Goulding et al., 1993).sizes are given in Table 1. D1McG156 was typed using SouthernThese mice all carry the original Splotch mutation, aanalysis, and D1Mit332, D1Mit253, and D1Mit215 were typed using10- to 11-bp rearrangement around the intron 3/exon PCR analysis. The Sp4H-specific band was only present in /// mice,
4 splice site of the Pax3 gene. No recombinations were suggesting that it represented the wildtype allele and that themarker was deleted in the Sp4H chromosome. (d) The CBA/Ca bandidentified among deleted markers D1McG156, D1-is fainter than that seen with the Sp4H stock chromosome. This isMit253, D1Mit332, and Pax3 in 111 of these backcrossprobably due to preferential amplification of the Sp4H chromosomeprogeny. The deleted marker D1Mit215 was not poly-DNA.
morphic in this backcross. A number of nondeletedmarkers were also genotyped for 111 of these backcrossprogeny. The mice had previously been typed for Vil flanking markers (D1Mit46, Vil ) and (D1Mit8,
D1Mit9, D1Mit44, D1Mit183, D1Ler3. )and Bcl2 (Goulding et al., 1993). A haplotype analysisis shown in Fig. 2. The deletion was contained within As no recombinants were identified between the de-
leted markers and Pax3, for the second stage of linkagea region of approximately 2.6 { 2 cM between the
AID Genom 4118 / 6r16$$$461 05-06-96 07:10:04 gnma AP: Genomics

Sp4H DELETION 209
FIG. 2. Haplotype analysis of 111 Splotch intraspecific backcross progeny from [(C57BL/6J Sp// 1 CBA/Ca)F1 1 CBA/Ca]. Only thosemice with a distinct white belly spot were analyzed. The loci that have been typed are shown to the left. The numbers of each haplotypechromosome type are listed below. Open boxes represent a C57Bl/6J (the Sp chromosome) allele, and black boxes represent a CBA/Ca allele.
analysis the number of backcross animals was increased DISCUSSIONto 412 and included mice without a clear belly spot. Atotal of 392 of these backcrossed mice were typed forPax3 In Sp4H heterozygous matings, 25% of embryos are
expected to be homozygous. We postulate that theseby Southern analysis, as well as forD1Mit332, D1Mit253,D1Mit79, D1Mit8, and D1McG156, and further markers are seen only as resorption sites, since no abnormal
embryos are evident and normal embryos all have afrom this region (D1Mit9, D1Mit44, D1Mit183, D1Ler3,D1Mit132, D1Mit134, and D1McG153) were mapped heterozygous or control genotype. There is an excess of
resorption sites in Sp4H matings compared with controlthrough a panel of 10 mice with recombinations betweenD1Mit79 and D1Mit8. A haplotype analysis for the en- matings of normal mice with the same genetic back-
ground, and this excess is not significantly differentlarged cross is shown in Fig. 3. This analysis places theclosest flanking markers, D1Mit79 and D1Mit134, only from the 25% predicted if homozygous progeny die after
implantation. Resorption rates for laboratory mice vary1.53{ 0.6 cM apart, defining the maximum extent of thedeletion. Figure 4 presents the gene order established according to genetic background, but approximately
12% has been documented in most inbred strains (Leo-using the genetic linkage and deletion mapping data pre-sented here. nard et al., 1971). West (1993) reported a resorption
FIG. 3. Haplotype analysis of a total of 392 intraspecific backcross mice. The loci that have been typed are shown to the left. Thenumbers of each haplotype chromosome type are listed below. Open boxes represent a C57BL/6J allele; black boxes represent a CBA/Caallele. Some markers have been typed using only the 10 mice showing recombinations between D1Mit46 and the cluster of markers aroundD1Mit8, and the missing data are indicated by the absence of a box in the table.
AID Genom 4118 / 6r16$$$461 05-06-96 07:10:04 gnma AP: Genomics

FLEMING ET AL.210
gous embryos have been shown to arrest at E7.5–E8,by which time gastrulation has proceeded as indicatedby the presence of a mesoderm layer. At E9.5, althoughsmaller than their littermates, these embryos also havesome evidence of a developing notochordal plate (Bucanet al., 1995). Therefore, it seems that Rw/Rw embryosdie at a later stage than the Sp4H/Sp4H embryos. Asimilar pattern of lethality as seen in the Sp4H muta-tion is reported in Gpi-deficient mice, which probablydo not develop past E6.5 (West, 1993). Sections throughGpi-deficient moles reveal no embryonic tissue, al-though trophectoderm and placental tissue appear nor-mal at E9.5. Gpi-deficient embryos die due to a nutri-tional deficiency and have been proposed as a modelfor some anembryonic pregnancies. It seems likely fromthis study that the Sp4H/Sp4H embryos fail around thesame stage, from E6 onward.
We have mapped a total of 22 markers in the regionof the Sp4H deletion using an intraspecific backcross[(C57BL/6J Sp//1 CBA/Ca)F11 CBA/Ca] of 392 prog-eny and deletion mapping using two F1 crosses [Sp4H//1 CBA/Ca and Sp4H// 1 C57BL/10J]. No differencesin gene order were found between our linkage map andthat of other published analyses of linkage in this areaof chromosome 1 (Malo et al., 1993; Dietrich et al.,1994). However, Vidal et al. (1992) have succeeded inseparating D1McG156 from the cluster around Pax3,locating this marker 1.2 { 0.7 cM proximal of Pax3 inan interspecific backcross of 251 mice. The interspecificcross used by Vidal and colleagues shows systemati-
FIG. 4. A genetic map of the region of mouse chromosome 1 cally greater genetic distances between markers com-around the Sp4H deletion, showing markers mapped using the back-pared with our intraspecific cross, as discussed pre-cross and deletion analysis described. The maximum extent of the
deletion is shown. The position of D1Mit215 within the deletion is viously (Goulding et al., 1993). Their placement ofnot known. The map is not to scale. D1McG156 proximal of Pax3, together with our place-
ment of D1McG156 distal of D1Mit332, suggests theorder: centromere–(D1Mit132, D1McG153)–(D1Mit-rate of 11.6% for mice in a C57BL stock, which is simi-
lar to the 11% reported here for our control mice on a 332)–(D1McG156)–(Pax3, D1Mit253, D1Mit215)–(D1Mit134). We cannot rule out the possibility thatC3H/HeH 1 101/H background.
In Copp’s recent review of embryo loss, he suggests D1Mit215 or D1Mit253 might be located within thePax3 gene, but none of the primer sequences for D1Mit-three important stages of development for the peri-im-
plantation embryo: formation of a blastocyst, establish- 215 or D1Mit253 shows any homology with Pax3 geno-mic sequence reported so far in mouse or human. Asment of a connection between trophoblast and mater-
nal decidua, and gastrulation (Copp, 1995). Our obser- D1Mit215 could be mapped using deletion analysisalone, we do not know the exact location of this markervations on sectioned moles suggest that a functional
blastocyst and fetal–maternal communication are es- within the deleted region.We have found five markers to be deleted in the Sp4Htablished in Sp4H/Sp4H embryos. Implantation sites
and embryo-sized spaces are visible, and some evidence mutation: D1Mit332, D1McG156, D1Mit253, D1Mit-215, and Pax3. In our intraspecific backcross, the dis-of extraembryonic membrane has been documented in
the most advanced mole isolated. This is unlike the tance between the closest nondeleted flanking markersthat we could separate genetically, D1Mit79 and D1-situation in early postimplantation lethals, such as in
Fgf4 null embryos, which are totally resorbed by E6.5 Mit134, and hence the maximum size of the deletionin Sp4H, is estimated to be 1.53 { 0.6 cM, which isdue to failure of the inner cell mass to proliferate (Feld-
man et al., 1995). Presumed Sp4H/Sp4H embryos also equivalent to about 3 Mb of DNA. As the mouse Pax3gene is estimated to extend over about 100 kb of geno-differ from homozygous Blind embryos, in which the
trophectoderm does not fuse with the deciduum (Van- mic DNA (Goulding et al., 1993), other genes may wellbe contained within the deletion region. We suggestkin and Caspari, 1979), and tw73 homozygotes, in which
the trophectoderm and ectoplacental cone fail to form that the early lethality we have seen in Sp4H/Sp4H mu-tants is most likely to be due to deletion of another,a close association with the decidua (Spiegelman et al.,
1976). In the rump-white mutation, Rw/Rw homozy- unidentified, gene contained within the deleted region,
AID Genom 4118 / 6r16$$$461 05-06-96 07:10:04 gnma AP: Genomics

Sp4H DELETION 211
Auerbach, R. (1954). Analysis of the developmental effects of a lethalrather than being due to a loss of Pax3. We think itmutation in the house mouse. J. Exp. Zool. 127: 305–329.unlikely that deletion of Pax3 is responsible for the
Bailey, D. W., and Bunker, H. P. (1972). Located histocompatabilitylethality because the Sp2H allele, with a 32-bp deletiongenes. Mouse Newslett. 47: 18.
within the Pax3 gene, appears to be a null allele, likeBaran, V., Vesela, J., Rehak, P., Koppel, J., and Flechon, J. E. (1995).
Sp4H must be, but does not cause such early embryonic Localisation of fibrillarin and nucleolin in nucleoli of mouse preim-death. Sp2H homozygotes die at least 5 days after the plantation embryos. Mol. Reprod. Dev. 40: 305–310.Sp4H/Sp4H embryos. An Sp2H-like peptide construct ex- Beechey, C. V., and Searle, A. G. (1986). Mutations at the Sp locus.
Mouse Newslett. 75: 28.hibited no DNA binding activity at all in vitro, sug-Bucan, M., Nagler, D. L., Hough, R. B., Chapman, V. M., and Lo,gesting that the Sp2H mutation may lead to a total loss
C. W. (1995). Lethality of Rw/Rw embryos during early postim-of Pax3 function (Chalepakis et al., 1994a,b). Loss ofplantation development. Dev. Biol. 168: 307–318.function mutations in another Pax gene, Pax6, in the
Cattanach, B. M., Rasberry, C., Burtenshaw, M. D., and Evans,small eye mutants, produce effects similar to a com-E. P. (1993). A further mutation at the Splotch locus. Mouse Ge-
plete deletion of the Pax6 gene (Hill et al., 1991). These nome 91: 115.observations support the suggestion that deletion of Chalepakis, G., Goulding, M. D., Read, A. P., Strachan, T., and Gruss,Pax3 alone is not likely to cause the early death of Sp4H
P. (1994a). Molecular basis of Splotch and Waardenburg Pax-3mutations. Proc. Natl. Acad. Sci. USA 91: 3685–3689.homozygotes.
Chalepakis, G., Jones, F. S., Edelman, G. M., and Gruss, P. (1994b).A number of genes have been localized close to Pax3,Pax-3 contains domains for transcriptional activation and tran-but most map outside the region known to be deletedscriptional inhibition. Proc. Natl. Acad. Sci. USA 91: 12745–in the Sp4H mutant. Nucleolin (Ncl ) (Aitman et al., 12749.
1991) and H54 (Bailey and Bunker, 1972) both map Copp, A. J. (1995). Death before birth: Clues from gene knockoutswithin this region, but neither has been precisely local- and mutations. Trends Genet. 11(3): 87–93.ized. Ncl encodes a DNA, RNA, and ATP-binding pro- Dickie, M. M. (1964). New Splotch alleles in the mouse. J. Hered.tein (Miranda et al., 1995) localized to the nucleoli of 55: 97–101.mouse preimplantation embryos, where it is associated Dietrich, W. F., Miller, J. C., Steen, R. G., Merchant, M., Damron,
D., Nahf, R., Gross, A., Joyce, D. C., Wessel, M., Dredge, R. D.,with the activation of the nucleolar precursor body inMarquis, A., Stein, L. D., Goodman, N., Page, D. C., and Lander,its synthesis of rRNA (Baran et al., 1995). If Ncl wereE. S. (1994). A genetic map of the mouse with 4,006 simple se-deleted in Sp4H, it might be predicted to have deleteri- quence length polymorphisms. Nature Genet. 7: 220–339.
ous preimplantation effects. H54 is a histocompatabil- Epstein, D. J., Vekemans, M., and Gros, P. (1991). Splotch (Sp2H), aity gene, encoding a cell surface antigen, and the effect mutation affecting development of the mouse neural tube, shows
a deletion within the paired homeodomain of Pax-3. Cell 67: 767–of deletion of a histocompatability gene on early embryo774.development is not known. Two recessive lethal alleles
Estibeiro, P. J., Copp, A. J., Cockcroft, D. L., Brown, N. A., and(l1Rk2 and l1Rk5) produced by irradiation have alsoClarke, D. O. (1990). Extraction of macromolecules from embryonicbeen mapped to this region (Seldin et al., 1993), butmaterial. In ‘‘Postimplantation Mammalian Embryos’’ (A. J. Copp
only by two point crosses, and their relationship with and D. L. Cockcroft, Eds.), pp. 173–204, Oxford Univ. Press, Lon-the postimplantation lethality in Sp4H is not clear. don.
In conclusion, the Sp4H mouse mutant has a small Evans, E. P., Burtenshaw, M. D., Beechey, C. V., and Searle, A. G.(1988). A splotch locus deletion visible by Giemsa banding. Mousegenomic deletion covering less than 1.53 { 0.6 cM andNews Lett. 81: 66.is associated with early death of homozygous mutants.
Feldman, B., Poueymirou, W., Papaioannou, V. E., DeChiara, T. M.,Sp4H/Sp4H embryos appear to implant but do not pro-and Goldfarb, M. (1995). Requirement of FGF-4 for postimplanta-ceed to gastrulation. Deletion of Pax3 alone is probablytion mouse development. Science 267: 246–249.
not sufficient to account for the early postimplantationFranz, T. (1989). Persistent truncus arteriosis in the Splotch mutant
death of homozygotes, so we suggest that another locus mouse. Anat. Embryol. 180: 457–464.in the deletion may be responsible for the lethality in Franz, T., Kothary, R., Surami, M. A. H., Halata, Z., and Grim, M.Sp4H/Sp4H mice. (1993). The Sp mutation interferes with muscle development in
the limbs. Anat. Embryol. 187: 153–160.ACKNOWLEDGMENTS Goulding, M. D., Chalepakis, G., Deutsch, U., Erselius, J. R., and
Gruss, P. (1991). Pax-3, a novel murine DNA binding protein ex-We thank Dr. B. Cattanach for access to the Sp4H mice, Professorspressed during early neurogenesis. EMBO J. 10: 1135–1147.J.-L. Guenet and H. LeRoy for the D1Ler3 probe, Professor P. Gros
Goulding, M. D., Sterrer, S., Fleming, J., Balling, R., Nadeau, J.,for the D1McG156 and D1McG153 probes, and Dr. M. Goulding forMoore, K. J., Brown, S. D. M., Steel, K. P., and Gruss, P. (1993).the Pax3 probe. We also thank Hazel Taylor for advice on dissectingAnalysis of the Pax-3 gene in the mouse mutant Splotch. Genomicsembryos, Annemarie Kelly for comments on the manuscript, David17: 355–363.Marshall for statistical advice, Andrew Copp and colleagues for per-
mission to cite unpublished findings, Matt Ward and Louise Kew for Hill, R. E., Favor, J., Hogan, B. L. M., Ton, C. C. T., Saunders,excellent technical help, and Neil Glenn for animal care. This work G. F., Hanson, I. M., Prosser, J., Jordan, T., Hastie, N. D., andhas been supported by the Hearing Research Trust and Medical van Heyningen, V. (1991). Mouse small eye results from mutationsResearch Council. in a paired-like homeobox-containing gene. Nature 354: 522–525.
Hillyard, A. L., Doolittle, D. P., Davisson, M. T., and Roderick, T. H.REFERENCES (1992). Locus map of the mouse. Mouse Genome 90: 11–12.
Lalwani, A. K., Brister, J. R., Fex, J., Grundfast, K. M., Ploplis, B.,Aitman, T. J., Hearne, C. M., McAleer, M. A., and Todd, J. A. (1991).Mononucleotide repeats are an abundant source of length variants San Augustin, T. B., and Wilcox, E. R. (1995). Further elucidation
of the genomic structure of PAX3, and identification of two differ-in mouse genomic DNA. Mamm. Genome 1: 206–210.
AID Genom 4118 / 6r16$$$461 05-06-96 07:10:04 gnma AP: Genomics

FLEMING ET AL.212
ent point mutations within the PAX3 homeobox that cause Waar- Seldin, M. F., Hunter, K., and Watson, M. L. (1993). Mouse chromo-some 1. Mamm. Genome 4: S10–30.denburg’s syndrome type I in two families. Am. J. Hum. Genet.
56: 75–83. Spiegelman, M., Artzt, K., and Bennett, D. (1976). Embryologicalstudy of a T/t locus mutation (tw73) affecting trophectoderm devel-Leonard, A., Deknudt, Gh., and Linden, G. (1971). Ovulation andopment. J. Embryol. Exp. Morphol. 36: 373–381.prenatal losses in different strains of mice. Exp. Animale (Paris)
Tassabehji, M., Newton, V. F., Leverton, K., Turnbull, K., Seema-4: 1–6.nova, E., Kunze, J., Sperling, K., Strachan, T., and Read, A. P.LeRoy, H., Simon-Chazottes, X., and Guenet, J-L. (1992). A set of(1994). PAX3 gene structure and mutations: Close analogies be-anonymous DNA clones as markers for mouse gene mapping.tween Waardenburg syndrome and the Splotch mouse. Hum. Mol.Mamm. Genome 3: 244–246.Genet. 3: 1069–1074.
Malo, D., Vidal, S. M., Hu, J., Skamene, E., and Gros, P. (1993). Underhill, D. A., Vogan, K. J., and Gros, P. (1995). Analysis of theHigh-resolution linkage map in the vicinity of the host resistance mouse Splotch-delayed mutation indicates that the Pax-3 pairedlocus Bcg. Genomics 16: 655–663. domain can influence homeodomain DNA-binding activity. Proc.
Natl. Acad. Sci. USA 92: 3692–3696.Miranda, G. A., Chokler, I., and Aguilera, R. J. (1995). The murinenucleolin protein is an inducible DNA and ATP binding protein Vankin, G. L., and Caspari, E. W. (1979). Developmental studies ofwhich is readily detected in nuclear extracts of lipopolysaccharide- the lethal gene Bld in the mouse. J. Embryol. Exp. Morphol. 49:treated splenocytes. Exp. Cell Res. 217: 294–308. 1–12.
Vidal, S. M., Epstein, D. J., Malo, D., Weith, A., Vekemans, M., andMoase, C. E., and Trasler, D. G. (1989). Spinal ganglia reduction inGros, P. (1992). Identification and mapping of six microdissectedthe Splotch-delayed mouse neural tube defect mutant. Teratologygenomic DNA probes to the proximal region of mouse chromosome40: 67–75.1. Genomics 14: 32–37.
Moase, C. E., and Trasler, D. G. (1992). Splotch locus mouse mutants:Vogan, K. J., Epstein, D. J., Trasler, D. G., and Gros, P. (1993).Models for neural tube defects and Waardenburg syndrome type
The Splotch-delayed (Spd) mouse mutant carries a point mutationI in humans. J. Med. Genet. 29: 145–151.within the paired box of the Pax-3 gene. Genomics 17: 364–369.
Peters, J., Glenister, P., Moseley, H., and Roberts, A. (1995). Geno- West, J. D. (1993). A genetically defined animal model of anembry-typic and phenotypic analysis of Del(1)Spr3H and Pax3Sp-4H. Mouse onic pregnancy. Human Reprod. 8: 1316–1323.Genome 93: 1041–1043. Zlotogora, J., Lere, I., Bar-David, S., Ergaz, Z., and Abeliovich, D.
(1995). Homozygosity for Waardenburg Syndrome. Am. J. Hum.Russell, W. L. (1947). Splotch, a new mutation in the house mouseMus musculus. Genetics 32: 102. Genet. 56: 1173–1178.
AID Genom 4118 / 6r16$$$461 05-06-96 07:10:04 gnma AP: Genomics





























