Embed Size (px)
Citation preview

Vol. 5, 967-973, September 1994 Cell Growth & Differentiation %7
The Enterocyte-like Differentiation of the Caco-2Tumor Cell Line Strongly Correlates withResponsiveness to cAMP and Activationof Kinase A Pathway’
Sandro Pignata,2 Laura Maggini, Raffaele Zarrilhi,Antonio Rea, and Angela M. Acquaviva3
Cattedra di Gastroenterologia [S. P.], Dipartimento di Biologia e PatologiaCellulare e Molecolare, Centro di Endocrinologia e OncologiaSperimentale C. N. R. EL. M., R. Z., A. R., A. M. A.], Facolta di Medicinae Chirurgia, Universita di Napoli “Federico II,” via S. Pansini, 5801 31 , Naples, Italy
Abstract
We have investigated the expression of protein kinase C(PKC) and protein kinase A (PKA) during the phases ofgrowth and differentiation of the human coloncarcinoma Caco-2 cells. We studied whetherdifferentiation correlated with the responsiveness tocAMP and with an increased transport of the catalyticsubunit of PKA into the nucleus. Also, we evaluatedwhether this phenomenon was affected by PKC activity.High levels of activated PKC were found in the plasmamembranes of replicating cells. When the cells began todifferentiate, plasma membrane-activated PKCdecreased, while the cytosolic fraction increased. On thecontrary, PKA holoenzyme increased duringdifferentiation, along with the transport of its catalyticsubunit into the nucleus. Both types I and II kinase Aholoenzymes increased during differentiation, withmaximal type II activity found when cells were fullydifferentiated. In replicating preconfluent cells, theinhibition of PKC by high dose phorbol 1 2-myristate 13-acetate or sphingosine increased the amount of bothPKA catalytic subunit in the nucleus and sucrase activity.During differentiation, 8-Bromo-cAMP increased PKAcatalytic subunit in the nucleus and apoliprotein AlmRNA levels. These effects were inhibited by how-dosephorbol 1 2-myristate 1 3-acetate, which activates PKC inthe plasma membranes. Our data suggest that PKC isactivated in proliferating Caco-2 cells. The inhibition ofPKC induces the transport of PKA catalytic subunit intothe nucleus and the expression of the differentiationmarkers. Differentiated Caco-2 cells show a loweractivation of PKC and an increased transport of thecatalytic subunit of PKA into the nucleus. DifferentiatedCaco-2 cells are highly responsive to cAMP and the8-Br-cAMP analog is able to revert the inhibitory effectof PKC.
Received 3/30/94; revised 6/27/94; accepted 6/30/94.� This work was supported in part by a grant from Associazione Italiana perIa Ricerca sul cancro. S. P. is supported by a fellowship from Dottorato diRicerca in Scienze Gastroenterologiche, Universit#{224}“La Sapienza”, Roma.2 Present address: Divisione di Oncologia Medica B, Istituto Nazionale perIo Studio e Ia Cura dei Tumori, Fondazione C. Pascale, Napoli, Italy.3 To whom requests for reprints should be addressed.
IntroductionThe human colon carcinoma cell line Caco-2 represents auseful “in vitro” experimental model to study intestinaldifferentiation. This cell line was derived from a relativelywell-differentiated tumor and grows slowly in nude mice(1). Caco-2 cells differentiate spontaneously, depending oncell density (2). When seeded at high density either onpermeable filters on impermeable substrates (plastic orglass), cells form well-polarized monolayens showing, at theultrastructural level, tight junctions, well-developed apicalmicnovilhi features of mature small bowel entenocytes, and15th-week fetal colonocytes (1-6). Sucrase isomaltase, al-kahine phosphatase, and aminopeptidase, brush bonder en-zymes peculiar to the adult human small intestine, areproduced during differentiation (2, 3, 5). Also, the enzymediamine oxidase, present at high concentration in maturesmall bowel enterocytes, is produced and secreted by con-fluent Caco-2 cells (7, 8). Furthermore, synthesis and secre-tion of human apohipoproteins Al, B, E, and C duringdifferentiation has been demonstrated (8-1 3). The differen-tiation is a growth-related phenomenon that starts as soonas confluency is reached (2, 4). Whereas the crypt containsmultiple cell types, Caco-2 cells are remarkably homoge-nous, suggesting that they represent the neoplastic equiva-lent of crypt enterocytes committed to the absorptive cellline but arrested in differentiation at the crypt/fetal stage (4,5). For these reasons, Caco-2 cells are a unique model tounderstand how the neoplastic process interferes with theregulation of cellular differentiation.
Little information is available on the molecular mecha-nisms that regulate cell lineage commitment, cell-specificgene expression, and the process of differentiation duringmigration of cells from cnypts (undifferentiated compart-ment) to villi (differentiated compartment) in the intestinalepithehium (14). Activation of PKC4 is involved in the stim-ulation of colonic epithehial proliferation by bile salts andtumor-promoting phorbol esters (1 5-1 7). Also, the level ofthis kinase activity is higher in colonic mucosa from patientswith colonectal cancers than in normal subjects (1 7). On theother hand, some differentiation function of colonic epithe-hum, such as the opening of apical chloride channels (18-20), is related to activation of cAMP-dependent proteinkinase (PKA).
Whether protein kinase-dependent pathways are in-volved in the differentiation of Caco-2 cells has not beenevaluated. Therefore, this study was designed to assess theactivity of PKC and PKA during the spontaneous diffenenti-ation of Caco-2 cells.
4 The abbreviations used are: PKC, protein kinase C; PKA, protein kinase A;8-Br-cAMP, 8-bromo-cAMP; PMA, phorbol 12-myristate 13-acetate; bp,base pair(s); GPDH, glyceraldehyde-3-phosphate deydrogenase.
A8
6(0
0
0)0
2
00 5 10 15 20
Days
B
7
SUCRASE
ISOMALTASE
GPDH �. � � I- �
12345
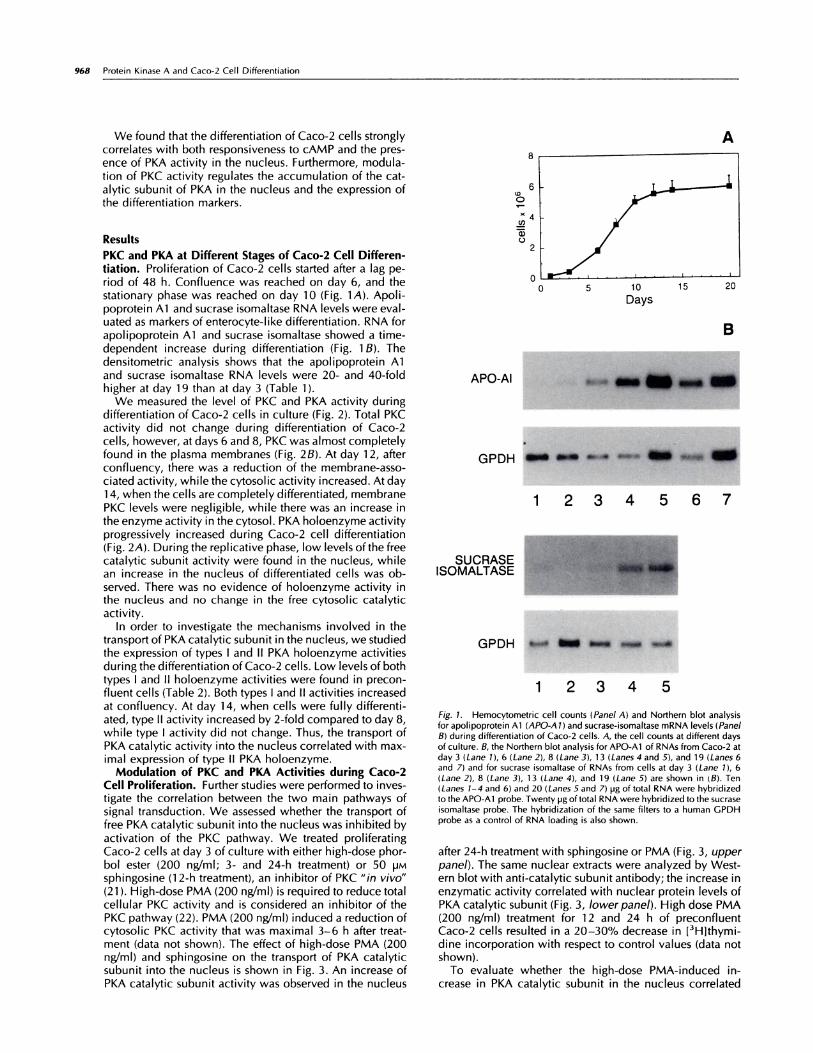
Fig. 1. Hemocytometric cell counts (Panel A) and Northern blot analysis
for apolipoprotein Al (APO-Al) and sucrase-isomaltase mRNA levels (PanelB) during differentiation of Caco-2 cells. A, the cell counts at different days
of culture. B, the Northern blot analysis for APO-Al of RNAs from Caco-2 atday 3 (Lane 1), 6 (Lane 2), 8 (Lane 3), 13 (Lanes 4 and 5), and 19 (Lanes 6and 7) and for sucrase isomaltase of RNAs from cells at day 3 (Lane 1), 6(Lane 2), 8 (Lane 3), 13 (Lane 4), and 19 (Lane 5) are shown in tB). Ten(Lanes 1-4 and 6) and 20 (Lanes 5 and 7) pg of total RNA were hybridizedto the APO-Al probe. Twenty pg oftotal RNA were hybridized to the sucrase
isomaltase probe. The hybridization of the same filters to a human GPDHprobe as a control of RNA loading is also shown.
968 Protein Kinase A and Caco-2 Cell Differentiation
We found that the differentiation of Caco-2 cells stronglycorrelates with both responsiveness to cAMP and the pres-ence of PKA activity in the nucleus. Furthermore, modula-tion of PKC activity regulates the accumulation of the cat-alytic subunit of PKA in the nucleus and the expression ofthe differentiation markers.
Results
PKC and PKA at Different Stages of Caco-2 Cell Differen-tiation. Proliferation of Caco-2 cells started after a lag pe-nod of 48 h. Confluence was reached on day 6, and thestationary phase was reached on day 10 (Fig. 1A). Apoli-poprotein Al and sucrase isomaltase RNA levels were eval-uated as markers of enterocyte-like differentiation. RNA forapolipoprotein Al and sucrase isomaltase showed a time-dependent increase during differentiation (Fig. 1 8). Thedensitometnic analysis shows that the apolipoprotein Aland sucrase isomaltase RNA levels were 20- and 40-foldhigher at day 19 than at day 3 (Table 1).
We measured the level of PKC and PKA activity during
differentiation of Caco-2 cells in culture (Fig. 2). Total PKCactivity did not change during differentiation of Caco-2cells, however, at days 6 and 8, PKC was almost completelyfound in the plasma membranes (Fig. 28). At day 12, afterconfluency, there was a reduction of the membrane-asso-ciated activity, while the cytosolic activity increased. At day14, when the cells are completely differentiated, membranePKC levels were negligible, while there was an increase inthe enzyme activity in the cytosol. PKA holoenzyme activityprogressively increased during Caco-2 cell differentiation(Fig. 2A). During the replicative phase, low levels ofthe freecatalytic subunit activity were found in the nucleus, whilean increase in the nucleus of differentiated cells was ob-served. There was no evidence of holoenzyme activity inthe nucleus and no change in the free cytosohic catalytic
activity.In order to investigate the mechanisms involved in the
transport of PKA catalytic subunit in the nucleus, we studiedthe expression of types I and II PKA holoenzyme activitiesduring the differentiation of Caco-2 cells. Low levels of bothtypes I and II holoenzyme activities were found in precon-fluent cells (Table 2). Both types I and II activities increasedat confluency. At day 14, when cells were fully differenti-ated, type II activity increased by 2-fold compared to day 8,while type I activity did not change. Thus, the transport ofPKA catalytic activity into the nucleus correlated with max-imal expression of type II PKA holoenzyme.
Modulation of PKC and PKA Activities during Caco-2Cell Proliferation. Further studies were performed to inves-tigate the correlation between the two main pathways ofsignal transduction. We assessed whether the transport offree PKA catalytic subunit into the nucleus was inhibited byactivation of the PKC pathway. We treated proliferatingCaco-2 cells at day 3 of culture with either high-dose phor-bol ester (200 ng/ml; 3- and 24-h treatment) or 50 �M
sphingosine (1 2-h treatment), an inhibitor of PKC “in vivo”
(21). High-dose PMA (200 ng/ml) is required to reduce totalcellular PKC activity and is considered an inhibitor of thePKC pathway (22). PMA (200 ng/ml) induced a reduction ofcytosolic PKC activity that was maximal 3-6 h after treat-ment (data not shown). The effect of high-dose PMA (200ng/ml) and sphingosine on the transport of PKA catalyticsubunit into the nucleus is shown in Fig. 3. An increase ofPKA catalytic subunit activity was observed in the nucleus
APO-Al -� �1I�uI_ a.
GPDH � � � � -. �
123456
after 24-h treatment with sphingosine or PMA (Fig. 3, upper
panel). The same nuclear extracts were analyzed by West-em blot with anti-catalytic subunit antibody; the increase inenzymatic activity correlated with nuclear protein levels ofPKA catalytic subunit (Fig. 3, !owerpanel). High dose PMA(200 ng/ml) treatment for 1 2 and 24 h of preconfluentCaco-2 cells resulted in a 20-30% decrease in [3H]thymi-dine incorporation with respect to control values (data notshown).
To evaluate whether the high-dose PMA-induced in-crease in PKA catalytic subunit in the nucleus correlated

10Ca)0a 8CR
�? 6
0
0
0.3
0E i
0
a The mean densimometric scan values and SE of at least three independent
experiments are reported. Maximal values were arbitrarily taken as 100.
Apo-Al , apohipoprotein Al ; SI, sucrase isomaltase.
A
Control PMA 3 hr PMA 24 hr Sphingosine
� 4.,
U)C
ci)0
a-
0)
�0
C)
0a-00
C
3-I-
0
Ea-
14
12
10
8
6
4
2
0
0
14
12 -
10
8
6
4
2
00
� #{149}..�S �#{149}:.
2 4 6 8 10 12 14
DaysB
2 4 6 8 10 12 14
a)
0
E:3
3(I)
2 3 4
Days
a PKA activity is expressed as pmol of incorporated ATP. Results are the
mean ± SD of three independent experiments.
with differentiation, preconfluent Caco-2 cells were treatedfor 3 days with 200 ng/mI PMA and analyzed for sucraseisomaltase activity. Treatment started at day 2, and sucraseisomaltase activity was assayed at days 3, 4, and 5; PMAwas added to the medium every 24 h. As shown in Fig. 4,treatment with 200 ng/ml medium PMA induced a time-
dependent increase in sucrase activity up to about 2-foldwhen compared to control values (P < 0.05).
Modulation of PKC and PKA Activities during Differen-tiation of Caco-2 Cells. Confluent Caco-2 cells at day 8 ofculture showed an increase in nuclear PKA catalytic activitythat correlated with the early expression of differentiationmarkers, including mRNAs for apohipoprotein Al and su-crase (Figs. 1 and 2). To investigate if modulation of PKCactivity was able to prevent the response to cAMP and the
Cell Growth & Differentiation 969
Table 1 Apo-Al and SI expression during differen tiation of Caco-2 cells
Days of cultureApo-Al/GPDH
mRNA levels’
SI/GPDH
mRNA levels’
3 4.1 ± 1.7 2.3 ± 0.8
6 8.9 ± 3.5 4.2 ± 1.8
8 27.9 ± 7.3 14.2 ± 5.4
13 100 93.4±9.8
19 88.4±5.4 100
Days
Fig. 2. PKA and PKC expression during differentiation of Caco-2 cells.Panel A, PKA cytosohic holoenzyme (s), nuclear holoenzyme ( #{149}), cytosohic-free catalytic activity (#{149}),and nuclear free catalytic activity (A). Panel B, PKC
activity in membrane (U) and cytosol 4). Results are the means of at least
three experiments; bars, SD.
Table 2 Types I and II PKA holoenzyme activitya during differentiation of
Caco-2 cells
Days of culture Type I holoenzyme Type 2 holoenzyme
3 0.3 ± 0.1 0.7 ± 0.2
8 6.2 ± 1.2 6.7 ± 1.4
14 6.1 ± 1.3 15.3 ± 3.6
123
Fig. 3. Upperpanel, effect of 200 ng/ml PMA (3 and 24 h treatment) and 50�M sphingosine (1 2 h treatment) on the activity of PKA free catalytic subunit
in the nucleus of Caco-2 cells at day 3 of culture. Results are the mean ± SDvalues of three experiments. Lower panel, Western blot analysis with anti
PKA catalytic subunit antibody of nuclear protein extracts from Caco-2 cellsat day 3 of culture after PMA (200 ng/ml) treatment Lane 1, untreated cells;Lane 2, treatment with 50 �M sphingosine for 1 2 h; Lane 3, treatment with
PMA (200 ng/ml) for 24 hJ. Arrow, the catalytic subunit reactive band.
Fig. 4. Effect of PMA (200 ng/ml) on sucrase activity in preconfluent Caco-2
cells (PMA, U; Control, #{149}).Treatment started at day 2 and assay wasperformed at days 3, 4, and 5. PMA was added to the medium every 24 h.Results are mean values of five experiments; bars, SD. ‘ P < 0.05 withrespect to control values. +, not significant with respect to basal values.
C’)Cci)00.
0):3.
�0ci)cci00.0C)C
0.
0E0.
10
8
6
4
2
0
Membrane Cytosol
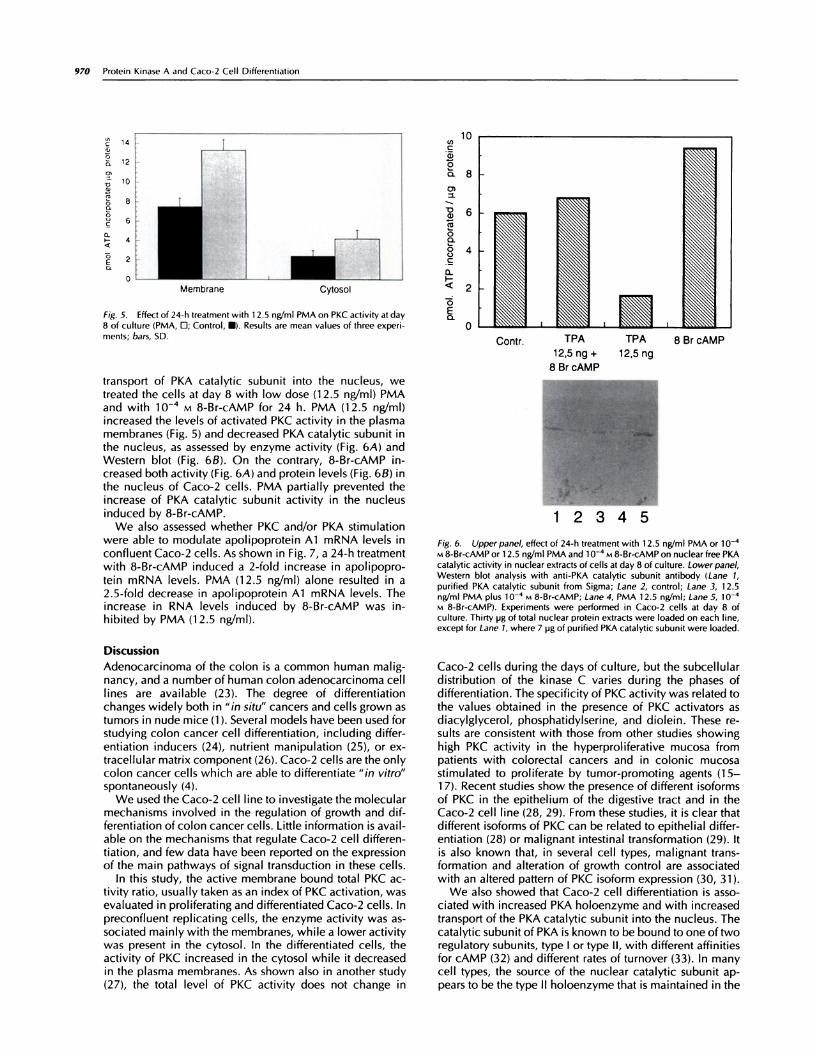
Fig. 5. Effect of 24-h treatment with 1 2.5 ngfml PMA on PKC activity at day
8 of culture (PMA, D; Control, U). Results are mean values of three experi-ments; bars, SD.
Contr. TPA TPA 8 Br cAMP12,5 ng12,5 ng +
8 Br cAMP
970 Protein Kinase A and Caco-2 Cell Differentiation
14
� 12
� 10‘1)
� 8
� 6
transport of PKA catalytic subunit into the nucleus, wetreated the cells at day 8 with low dose (1 2.5 ng/ml) PMAand with l0� M 8-Br-cAMP for 24 h. PMA (12.5 ng/ml)increased the levels of activated PKC activity in the plasmamembranes (Fig. 5) and decreased PKA catalytic subunit inthe nucleus, as assessed by enzyme activity (Fig. 6A) andWestern blot (Fig. 6B). On the contrary, 8-Br-cAMP in-creased both activity(Fig.6A) and protein levels (Fig. 6B) inthe nucleus of Caco-2 cells. PMA partially prevented theincrease of PKA catalytic subunit activity in the nucleusinduced by 8-Br-cAMP.
We also assessed whether PKC and/or PKA stimulationwere able to modulate apolipoprotein Al mRNA levels inconfluent Caco-2 cells. As shown in Fig. 7, a 24-h treatmentwith 8-Br-cAMP induced a 2-fold increase in apolipopro-tein mRNA levels. PMA (12.5 ng/ml) alone resulted in a2.5-fold decrease in apolipoprotein Al mRNA levels. Theincrease in RNA levels induced by 8-Br-cAMP was in-hibited by PMA (12.5 ng/ml).
Discussion
Adenocarcinoma of the colon is a common human mahig-nancy, and a number of human colon adenocarcinoma celllines are available (23). The degree of differentiationchanges widely both in “in situ” cancers and cells grown astumors in nude mice (1 ). Several models have been used forstudying colon cancer cell differentiation, including differ-
entiation inducers (24), nutrient manipulation (25), or ex-tracellular matrix component (26). Caco-2 cells are the onlycolon cancer cells which are able to differentiate “in vitro”spontaneously (4).
We used the Caco-2 cell line to investigate the molecularmechanisms involved in the regulation of growth and dif-ferentiation of colon cancer cells. Little information is avail-able on the mechanisms that regulate Caco-2 cell differen-tiation, and few data have been reported on the expressionof the main pathways of signal transduction in these cells.
In this study, the active membrane bound total PKC ac-tivity ratio, usually taken as an index of PKC activation, wasevaluated in proliferating and differentiated Caco-2 cells. Inpreconfluent replicating cells, the enzyme activity was as-sociated mainly with the membranes, while a lower activitywas present in the cytosol. In the differentiated cells, theactivity of PKC increased in the cytosol while it decreasedin the plasma membranes. As shown also in another study(27), the total level of PKC activity does not change in
.. .
12345
Fig. 6. Upper panel, effect of 24-h treatment with 1 2.5 ng/ml PMA or 1 0�M 8-Br-cAMP or 1 2.5 ng/ml PMA and 1� vi 8-Br-cAMP on nuclear free PKAcatalytic activity in nuclear extracts of cells at day 8 of culture. Lower panel,Western blot analysis with anti-PKA catalytic subunit antibody (Lane 1,
purified PKA catalytic subunit from Sigma; Lane 2, control; Lane 3, 1 2.5nWml PMA plus io-� M 8-Br-cAMP; Lane 4, PMA 12.5 nWmI; Lane 5, i#{252}�M 8-Br-cAMP). Experiments were performed in Caco-2 cells at day 8 ofculture. Thirty pg of total nuclear protein extracts were loaded on each line,
except for Lane 1, where 7 pg of purified PKA catalytic subunit were loaded.
Caco-2 cells during the days of culture, but the subcellulardistribution of the kinase C varies during the phases ofdifferentiation. The specificity of PKC activity was related tothe values obtained in the presence of PKC activators asdiacylglycerol, phosphatidylserine, and diolein. These re-suIts are consistent with those from other studies showing
high PKC activity in the hyperproliferative mucosa frompatients with colorectal cancers and in colonic mucosastimulated to proliferate by tumor-promoting agents (15-1 7). Recent studies show the presence of different isoforms
of PKC in the epithelium of the digestive tract and in theCaco-2 cell line (28, 29). From these studies, it is clear thatdifferent isoforms of PKC can be related to epithelial differ-entiation (28) or malignant intestinal transformation (29). Itis also known that, in several cell types, malignant trans-formation and alteration of growth control are associatedwith an altered pattern of PKC isoform expression (30, 31).
We also showed that Caco-2 cell differentiation is asso-ciated with increased PKA holoenzyme and with increasedtransport of the PKA catalytic subunit into the nucleus. Thecatalytic subunit of PKA is known to be bound to one of tworegulatory subunits, type I or type II, with different affinitiesfor cAMP (32) and different rates of turnover (33). In manycell types, the source of the nuclear catalytic subunit ap-pears to be the type II holoenzyme that is maintained in the

2
=a
0�... 1
6
0
Fig. 7. Effect of 48-h treatment with 1 2.5 ng/ml PMA, 8-Br-cAMP 1 O� M,
or 1 2.5 ng/ml PMA and 8-Br-cAMP 10’ M on APO Al mRNA expression inCaco-2 cells at day 8 of culture ( 1, control; 2, 8-Br-cAMP; 3, 1 2.5 ng/mlPMA; 4, 1 2.5 ng/ml PMA plus 1 0� M 8-Br-cAMP). Ten pg of total RNA wereloaded on each line. The histogram shows the densitometric analysis of theAPO-Al :GPDH ratio.
1 4
Cell Growth & Differentiation 971
I #{149} Apo-Al
!!!!GPDH
Golgi complex through the interaction between the type IIsubunit and specific anchoring proteins (34-37). The type Icomplex appears to be diffusely distributed in the cyto-plasm (34-37). In our experimental model, the increase inthe nuclear PKA catalytic subunit strongly correlates withthe increase in the type II regulatory subunit.
In replicating preconfluent Caco-2 cells, the activation ofPKC seems to interfere with the transport of the catalyticsubunit of PKA into the nucleus. In this phase, the inhibitionof PKC by sphingosine or by high-dose PMA increased theamount of nuclear PKA catalytic subunit. This effect wasassociated with an increased expression of the differentia-tion markers. Other studies (38) have shown that activationof PKC selectively increases the type ct regulatory subunitof cAMP-dependent protein kinase, which is not able totransport the catalytic subunit in the nucleus. The mecha-nisms underlying the effects of PKC on specific subtype ofPKA regulatory subunits in Caco-2 cells are largely Un-known and need to be investigated further.
After confluency, cells were highly responsive to cAMP.8-Br-cAMP increased PKA catalytic subunit in the nucleusand the levels of apolipoprotein Al mRNA. The activationof PKC activity by low-dose PMA partially prevented theincrease in nuclear PKA catalytic subunit induced by 8-Br-cAMP and reduced the levels of the differentiation markers.This effect of PKC has been demonstrated in other cell lines
(39), and it may represent a general mechanism mediatingthe effects of PKC on cellular growth and differentiation.The balance between PKC and PKA pathways may be rel-
evant for the regulation of replication and differentiation ofintestinal cells. Furthermore, the increase of cAMP levels
may regulate the cross-talk between the different PKC iso-forms expressed during the proliferative and differentiativephase of Caco-2 cells. Further experiments are needed toverify this hypothesis.
In conclusion, the present data suggest that PKC exerts anegative control on Caco-2 cell differentiation by inhibiting
the transport of the catalytic subunit of PKA into the nu-cleus. Our results indicate that some differentiation featuresof Caco-2 cells are cAMP dependent and suggest that
cAMP-dependent kinase may have a role in the activationof the genes expressed during differentiation of the entero-cytes.
Materials and Methods
Cell Culture
Caco-2 cells were routinely grown in plastic dishes at 37#{176}Cin a humidified incubator 5% CO2-95% air atmosphere in
Dulbecco’s modified Eagle’s medium supplemented withlO% fetal calf serum, glutamine (2 mM), penicillin (100units/mI), and streptomycin (100 pg/mI) and buffered withN-2-hydroxy-ethylpiperazine-N’-2-ethane sulfonic acid (20mM). Caco-2 cells were seeded at 5 X 1 0� cells/mI and wereroutinely subcultured when about 80% confluent. The cul-ture medium was changed every other day. Cells werealways >90% viable, as shown by trypan blue exclusion.Confluence was reached 6-8 days after the inoculum andthe stationary phase on day 10. Cell cultures were stoppedafter 1 5 passages.
Protein Kinase C Assay
Protein kinase C was measured as described by Ballesterand Rosen (22). Culture plates were washed with buffer A(20 mivt Tnis-HCI, 2 m�i EDTA, 50 m�i 2-mercaptoethanol,0.1 mM protease inhibitor (phenylmethylsulfonyl fluoride))and then scraped and homogenized in the same buffer. Thehomogenate was centrifuged at 2700 rpm for 5 mm, and thesupernatant was centrifuged again at 45,000 rpm for 1 h.The supernatant of the last centnifugation corresponded tothe cytosolic fraction. The pellet was resuspended in 1 mlbuffer A plus 0.l% Triton X-lOO. The samples were incu-bated overnight and then were centrifuged at 45,000 rpmfor 45 mm. The supernatant corresponded to membranefraction.
PKC activity was determined by measuring the transfer of32p from [‘y-32PJATP into lysine-rich histones (Ill 5). Thestandard reaction mixture (25 p1/tube) contained 3-5 pg ofprotein extracts, 20 m�i Tris HCI (pH 7.5), 10 mtvi MgCl2, 1mM CaCI2, 50 �M cold ATP, 80 pg/mI phosphatidylsenine,0.8 pg/mI dioleine, and 0.75 pCi of [y-32P]ATP. After 10mm incubation at 30#{176}C,the reaction was stopped by theaddition of 1 ml of 25% tnichloroacetic acid. Acid-precip-itable material was collected on Whatman GF/C filters byvacuum filtration and washed with 5 ml of 5% trichloro-acetic acid. Net PKC activitywas calculated by subtractingphosphorylation values obtained in the absence of phos-phatidylserine and dioleine from those measured in theirpresence.

972 Protein Kinase A and Caco-2 Cell Differentiation
cAMP-dependent Protein Kinase Assay
Nuclei and Cytosoh Preparation. Nuclei were preparedtreating cells with AT buffer [15 m�i NaCI, 60 m�i KCI, 14mM f3-mercaptoethanol, 2 mivi EDTA, 1 5 mM N-2-hydnoxy-ethylpiperazine-N’-2-ethane sulfonic acid (pH 7.9), and 0.3M sucrose] plus 1 % Triton X-1 00. The crude nuclear pelletwas prepared by spinning the extracts at 3500 rpm for 10mm at 4#{176}Con a 1 M sucrose cushion in AT buffer. Thesupernatant corresponded to the cytosohic fraction. Thenuclear pellet was resuspended in high-salt concentration(NaCI 1 50 mM) AT buffer. After salt extraction at 4#{176}C,thehomogenate was centrifuged at 12,000 rpm for 1 0 mm, andthe supennatant was assayed for kinase activity (no freecatalytic activity was detected in the pellet). The purity ofnuclei preparations was tested by electron microscopy andby quantitation of RNA content.
DEAE-52 Chromatography of cAMP-dependent ProteinKinase A. Type I and II PKA holoenzymes were preparedfrom the cytosohic fraction of Caco-2 cells according to
Costa (40), with the following modifications. The cytosolicfraction (300 pg proteins)was applied to a DEAE-52 column(0.5 X 5 cm) preequilibrated with 25 mt�i sodium potassiumphosphate buffer (pH 6.8) containing 1 mt�i EDTA, 15 mivi�3-mencaptoethanol, and 0.1 m�i dithiothneitoh. The columnwas washed with 10 ml of 25 mt�i phosphate buffer and alinear salt gradient from 25 mr’�i to 200 mi�.i sodium potas-sium phosphate buffer in the presence of 1 mr�i EDTA wasapplied. The flow rate was 0.5 mI/mm. Fractions of 0.5 mlwere collected from the column and were assayed for PKAactivity in absence or presence of cAMP. Types I and II PKAholoenzyme were ehuted between 60 and 1 20 m�’i, and 140and 200 mM sodium potassium phosphate buffer, nespec-tively.
Enzyme Assay. The assay (50 p1/tube) was performed for15 mm at 30#{176}C.The mixture contained 3-5 pg proteinextracts, 10 mM Tnis (pH 7.4), 1 0 mr�i MgCI2, 1 00 �M coldATP, and 0.75 pCi of [-y-32P]ATP, and 100 �M kempeptide(LRRASLG). The phosphorylation of kempeptide was deter-mined by spotting 30 pl of the incubation mixture on phos-phocellulose filter (Whatman p81 ) and washing it five timesin 0.75 mM phosphoric acid and once in 95% ethanol.Dried filters were counted by liquid scintillation. The phos-phoryhation of kempeptide was also determined by ion-exchange chromatography on AGI-X8 packed in Pasteurpipetes and equilibrated with 30% acetic acid. The assayfor holoenzyme activity was performed in the presence onabsence of 1 0� M cAMP (final concentration). The activityfound in the absence of 1 0� M cAMP in the assay mixturerepresents the activity of the free catalytic subunit of PKA.To determine PKA-specific activity, all samples were as-sayed with and without a specific inhibitor of PKA (finalconcentration, 50 pM) containing a PKA pseudophosphory-lation site (1P20 G1y10-Asp20-Lys11 from Sigma ChemicalCo.) (41).
Sucrase AssayThe culture plates were washed three times with ice-coldsaline. Cells were carefully scraped with a rubber police-man and homogenized in 0.1 M sodium phosphate buffer,pH 7.2, in a Dounce homogenizer.
Sucrase was assayed by Dahlqvist’s method (42) andproteins by Lowry’s method (43). Sucrase units are ex-pressed as pmol of glucose released in one mm at 3 7#{176}C,pH 7.2.
Northern Blot Analysis
Total RNA was extracted from Caco-2 cells by the methodof Chomczynski and Sacchi (44). The RNA was fractionatedin a 6% formaldehyde 1% aganose gel, transferred by cap-illary blotting to Hybond-N membrane (Amensham). Filterswere hybridized to apolipopnotein Al and sucrase isoma-Itase cDNA that were labeled with [a-32P]dATP (3000 Ci/mmol) using random hexamen primers and Klenow enzyme(Boeningher). Apolipoprotein Al cDNA probe was a 850-bpBamHI insert of a full-length human cDNA (45). Sucraseisomaltase probe was a 420-bp Sa!l-EcoRl insert isolatedfrom pHSl-l plasmid (46). GPDH probe was a 1 500-bpBamHI insert and included the full-length human cDNA(47). Filters were exposed to Kodak XAR-5 film for 1-3 daysat -80#{176}Cwith a Dupont Lightning Plus intensifying screen.RNA levels were quantified by densitometnic scanning ofthe autonadiognaphs using a Sony CCd videocamena linkedto a Macintosh Ihfx computer and the NIH IMAGE program(public domain).
Western Blot Analysis.
Nuclear extracts were fractioned on 1 0% sodium dodecylsulfate polyacrylamide gels, transferred to nylon mem-branes, and challenged with chicken-specific anti-PKA cat-alytic antibody (34). After incubation with the first antibody,the filter was incubated with anti-immunoglobuhin radishpenoxidase-hinked (Sigma). Final reaction was done in thepresence of 3,3’-diaminobenzidine. The specificity of reac-tion was performed by immunoprecipitation with antibodypreincubated with purified PKA catalytic subunit (Sigma).
AcknowledgmentsWe thank Dr. Peter G. Traber (University of Michigan Medical School) forkindly providing the pHSI-1 cDNA, Dr. Silvana Cassano for the anti-catalyticantibody, and Dr. Gianni Morrone for apolipoprotein Al cDNA, and Dr.Marco Romano for helpful suggestions.
References1 . Fogh, J., Fogh, J. M., and Orfeo, T. One hundred and twenty-sevencultured human tumor cell lines producing tumors in nude mice. J. NatI.Cancer Inst., 59: 221-226, 1977.
2. Pinto, M., Robine-Leon, S., Appay, M. D., Kedinger, M., Triadou, N.,Dussaulx, E., Lacroix, B., Simon-Assmann, P., Haffen, K., Fogh, J., andzweibaum, A. Enterocyte-like differentiation and polarization of the humancolon carcinoma cell line Caco-2 in culture. Biol. Cell., 47: 323-330, 1983.
3. Chantret, I., Barbat, A., Dussaulx, E., Brattain, M. G., and Zweibaum, A.Epithelial polarity, Villin expression, and enterocytic differentiation of cul-tured human colon carcinoma cells: a survey of twenty cell lines. CancerRes., 48: 1936-1942, 1988.
4. Neutra, M., and Louvard, D. Differentiation of intestinal cells in vitro. In:K. S. Matlin and Valentich L. (eds), Functional Epithelial Cells in Culture.Modern Cell Biology, Vol 8, pp. 363-398, New York: Alan R. Liss, Inc., pp363-398, 1989.
5. Hauri, H. P., Sterchi, E. E., Bienz, D., Fransen, J. A. M., and Marxer, A.
Expression and intracellular transport of microvillus membrane hydrolases inhuman intestinal epithelial cells. J. Cell Biol., 101:838-851, 1985.
6. Grasset, F., Pinto, M., Dussaulx, E., zweibaum, A., and Desjeux, J. F.Epithelial properties of human colonic carcinoma cell line Caco-2. Electricalparameters. Am. J. Physiol., 217: C260-C267, 1984.
7. D’Agostino, L., Daniele, B., Pignata, S., Gentile, R., Tagliaferri, P., Con-tegiacomo, A., Silvestro, G., Polistina, C., Bianco, A. R., and Mazzacca, G.Ornithine decarboxylase and diamine oxidase in human colon carcinomacell line Caco-2 in culture. Gastroenterology, 97: 888-894, 1989.
8. Daniele, B., and Quaroni, A. Polarized secretion of diamine oxidase byintestinal epithelial cells and its stimulation by heparin. Gastroenterology,
99: 1675-1687, 1990.

Cell Growth & Differentiation 973
9. Sasak, W. V., Buller, H. A., and Reinhold, R. Biosynthetic precursor (214kDa) of apolipoprotein B-48 is not secreted by Caco-2 cells and normalhuman intestine. Biochem. J., 264: 365-370, 1989.
10. Moberly, J. B., Cole, T. G., Alpers, D. H., and Shonfeld, G. Oleic acidstimulation of apolipoprotein B secretion from HepG2 and Caco-2 cellsoccurs post-transcriptionally. Biochim. Biophys. Acta, 1042: 70-80, 1990.
1 1 . Dashti, N., Smith, E. A., and Alaupovic, P. Increased production ofapolipoprotein B and its lipoproteins by oleic acid in Caco-2 cells. J. LipidRes., 31:113-123, 1990.
1 2. Jiao, S., Moberly, J. B., and Schonfeld, G. Editing of apolipoprotein Bmessenger RNA in differentiated Caco-2 cells. J. Lipid Res., 31: 695-700,1990.
13. Reisher, S. R., Hughes, T. E., Ordovas, J. M., Schaefer, E. S., andFeinstein, S. I. Increased expression of apolipoprotein genes accompaniesdifferentiation in the intestinal cell line Caco-2. Proc. NatI. Acad. Sci. USA,90:5757-5761, 1993.
14. Gordon, J. J. Understanding gastrointestinal epithelial cell biology: les-sons from mice with help from worms and flies. Gastroenterology, 104:
315-324, 1993.
15. Craven, P. A., and De Rubertis, F. R. Subcellular distribution of proteinkinase C in rat colonic epithelial cells with different proliferative activities.Cancer Res., 47: 3434-3438, 1987.
1 6. Craven, P. A., Pfanstiel, J., and De Rubertis, F. R. Role of activation ofprotein kinase C in the stimulation of colonic epithelial proliferation andreactive oxygen formation by bile acids. J. Clin. Invest., 79: 532-541 , 1987.
1 7. Sakanque, Y., Hatada, T., Kusunoki, M., Yanagi, H., Yamamura, 1., and
Utsunomiya, J. Protein kinase C activity as marker for colorectal cancer. Int.I. Cancer, 48: 803-806, 1991.
18. McRoberts, J. A., Beuerlein, G., and Darmsathaphorn, K. Cyclic AMPand Ca2’ activated K’� transport in a human colonic cell line. J. Biol. Chem.,261: 704-712, 1986.
19. Mandel, K. G., Dharmsathaphorn, K., and McRoberts, J. A. Character-ization of a cyclic AMP activated C1 transport pathway in the apical mem-
brane of a human colonic epithelial cell line. J. Biol. Chem., 261: 704-71 2,
1986.
20. Vaandragen, A. B., Bajnath, R., Groot, J. A., Bot, A. G. M., De Jonge, H.R. Ca2’ and cAMP activate different chloride efflux pathways in HT-29.cll9A colonic epithelial cell line. Am. J. Physiol., 261: G958-G965,1991.
21 . Hannun, Y. A., Loomis, C. R., Merril, A. H., and Bell, R. M. Sphingosineinhibition of protein kinase C activity and of phorbol dibutyrate binding in
vitro and in human platelets. J. Biochem. Chem., 261: 1 2604-1 2609, 1986.
22. Ballester, R., and Rosen, 0. M. Fate of immunoprecipitable proteinkinase C in GH3 Cells treated with phorbol 1 2-myristate 13-acetate. J. Biol.Chem., 260:15194-15199, 1985.
23. Fogh, J., and Treanpe, G. New human tumor cell line. ln:J. Fogh (ed),Human tumor cells in vitro, pp. 115-141. New York: Plenum PublishingCorp., 1975.
24. Gum, J. R., Kam, W. K., Byrd, J. C., Hicks, J. W., Sleisenger, M. H., andKim, Y. S. Effect of sodium butyrate on human colonic adenocarcinoma cellsinduction of placental-like alkaline phosphatase. J. Biol. Chem., 262: 1092-1097, 1987.
25. Pinto, M., Appay, M. D., Simon-Assmann, P., Chevalier, G., Dracopoli,N., Fogh, J., and Zweibaum, A. Enterocyte differentiation of cultured humancancer cells by replacement of glucose by galactose in the medium. Biol.Cell., 44:193-196, 1982.
26. Fukamachi, H., Mizuno, T., and Kim, Y. S. Morphogenesis of humancolon cancer cells with fetal rat mesenchymas in organ culture. Experientia(Basel), 42: 312-315, 1986.
27. RydeIl, E., Magnusson, K. E., Sjo, A., and Axelsson, K. Protein kinase Cand casein kinase II activities in two human colon carcinoma cell line HT 29and Caco-2: possible correlation with differentiation. Biosci. Rep., 10: 293-299, 1990.
28. Osada, S., Hashimoto, Y., Nomura, S., Kohno, Y., Chida, K., Tajima, 0.,Kubu, K., Akimoto, K., Koizumi, H., Kitamura, Y., Suzuki, K., Ohno, S., and
Kuroki, T. Predominant expression of nPKCi�, a Ca2�-independent isoform ofprotein kinase C in epithelial tissues, in association with epithelial differen-tiation. Cell Growth & Differ., 4: 167-175, 1993.
29. Delage, S., Chastre, E., Empereur, S., Wicek, D., Veissi#{233}re,D., Capeau,I., Gespach, C., and Cherqui, G. Increased protein kinase Ca expression inhuman colonic Caco-2 cells after insertion of human Ha-ras or polyomavirus middle T oncogenes. Cancer Res., 53: 2762-2770, 1993.
30. Persons, D. A., Wilkinson, W. 0., Bell, R. M., and Finn, 0. J. Alteredgrowth regulation and enhanced tumorigenicity of NIH 313 fibroblasts trans-fected with protein kinase C-I cDNA. Cell, 52: 447-458, 1988.
31 . Finkenzeller, G., Marm#{232},D., and Hug, H. Inducible overexpression ofhuman protein kinase a in NIH 3T3 fibroblasts results in growth abnormal-ties. Cell. Signal, 4: 1 63-1 77, 1992.
32. Hofmann, F., Beavo, J. A., Bechtel, P. J., and Krebs, E. G. Comparison ofadenosine 3’,S’-monophosphate-dependent protein kinases from rabbit skel-etal and bovine heart muscle. J. Biol. Chem., 250: 7795-7801, 1975.
33. Weber, W., and Hilz, R. cAMP-dependent protein kinases I and II:divergent turnover of subunits. Biochemistry, 25: 5661-5667, 1986.
34. Meinkoth, J. L., ii, Y., Taylor, S. S., and Feramisco, J. R. Dynamics of thedistribution of cyclic AMP-dependent protein kinase in living cells. Proc.Nat. Acad. Sci. USA, 87: 9595-9599, 1990.
35. Nigg, E. A., Hilz, H., Eppenberger, H. M., and Dutly, F. Rapid andreversible translation of the catalytic subunit of cAMP-dependent proteintype II from the Golgi complex to nucleus. EMBO J., 4: 2801-2806, 1985.
36. Bregman, D. B., Bhattacharuja, N., and Rubin, C. S. High affinity bind-ing protein for the regulatory subunit of cAMP-dependent protein kinase II�.Cloning, characterization and expression of cDNAs for rat brain P1 50. J.Biol. Chem., 264:4648-4656, 1989.
37. Bregman, D. B., Hirsch, A. M., and Rubin, C. S. Molecular character-ization ofbovine brain P75, a high affinity binding protein for the regulatory
subunit of cAMP-dependent protein kinase lIp. J. Biol. Chem., 266: 7207-7213, 1991.
38. Tasken, K., Kvale, D., Hansson, V., and Jahnsen, T. Protein kinase Cactivation selectively increases mRNA levels for one ofthe regulatory subunit(RIa) of cAMP dependent protein kinase in HT 29 cells. Biochem. Biophys.Res. Commun., 172:409-414, 1990.
39. Gallo, A., Benusiglio, E., Bonapace, L. M., Feliciello, A., Cassano, S.,Garbi, C., Musti, A. M., Gottesman, M. E., and Awedimento, E. V. v-ras andprotein kinase C dedifferentiate thyroid cells by down-regulating nuclearcAMP-dependent protein kinase A. Genes Dev., 6: 1621-1630, 1992.
40. Costa, M., Gerner, E. W., and Russel, D. Cell cycle specific activity oftype I and II cyclic adenosine 3’,S’ monophosphate-dependent proteinkinases in Chinese hamster ovary cells. J. Biol. Chem., 251: 3313-3319,1976.
41 . Cheng, H. G., Kemp, B. E., Pearson, R. B., Smith, A. J., Misconi, L., VanPatten, A. J., and Walsh, D. A. A potent synthetic peptide inhibitor of thecAMP-dependent protein kinase. J. Biol. Chem., 261: 989-992, 1986.
42. Dahlqvist, A. Method of assay of intestinal disaccharidases. Anal. Bio-chem., 7: 18-25, 1964.
43. Lowry, 0. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J. Proteinmeasurement with the Folin phenol reagent. J. Biol. Chem., 193: 265-275,1951.
44. Chomczynski, P., and Sacchi, N. Single-step method of RNA isolation byacid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Bio-chem., 162: 156-159, 1987.
45. Karathanasis, S. K., Zannis, V. I., and Breslow, J. L. Isolation and char-acterization of the human apolipoprotein A-I gene. Proc. NatI. Acad. Sci.USA, 80:6147-6151, 1983.
46. Traber, P. G., Yu, L., Wu, G. D., and Judge, T. A. Sucrase-isomaltasegene expression along crypt-villus axis of human small intestine is regulatedat level of mRNA abundance. Am. J. Physiol., 25: G123-G130, 1992.
47. Tso, J. Y., Sun, x. H., Kao, T., Reece, S., and Wu, R. Isolation andcharacterization of rat and human glyceraldehyde-3-phosphate dehydroge-nase cDNA: genomic complexity and molecular evolution of the gene.Nucleic Acids Res., 13: 2485-2502, 1985.





























