Embed Size (px)
Citation preview

I Medische E;t:JliDtheek
!2oo.)" E.U.R. ~~ ~
The Use of Illusory Visual Information
in Perception and Action
Het gebruik van illusoire visuele informatie in perceptie en actie
Proefschrift
ter verkrijging van de graad van doctor aan de Erasmus Universiteit Rotterdam
op gezag van de rector magnificus Prof.dr. S.W.J. Lamberts
en volgens besluit van het College voor Promoties.
De openbare verdediging zal plaatsvinden op
donderdag 19 mei 2005 om 13.30 uur
door
Denise Desiree J osefina de Grave
geboren te Winterswijk

Promotiecommissie
Promotor:
Overige leden:
Copromotoren:
Prof.dr. J.G.G. Borst
Dr. M.A. Frens
Prof.dr. C.J. Erkelens
Prof.dr. C.I. de Zeeuw
Dr. E. Brenner
Dr. J.B.J. Smeets
Dit proefschrift werd mede mogelijk gemaakt door financiele steun van de Nederlandse
Organisatie voor Wetenschappelijk Onderzoek (NWO), gebied Maatschappij- en
Gedragswetenschappen (MaGW), dossiemummer 490-22-lSOb.

Contents
Chapter 1
General Introduction
Chapter 2
Are the original and the induced Roelofs effect
caused by the same shift in straight ahead?
Chapter3
Effects ofthe Ebbinghaus figure on grasping
are not only due to misjudged size
Chapter4
The coding of pointing movements
ChapterS
The coding of saccades
Chapter 6
Summary and conclusions
Hoofdstuk6
Samenvatting en conclusies
References
Dankwoord
Curriculum Vitae
Publications
Contents
5
17
31
45
61
75
87
95
107
109
Ill


Chapter 1
General introduction

Chapter 1
Humans constantly interact with objects in the environment (e.g. grasp a pencil for writing or
pick up a cup of tea) without making many mistakes in these performed actions. To guide
these actions, visual information is used. In order to accurately grasp and pick up the pencil,
vision provides information such as the size of the pencil and its position relative to the body
and other objects in the environment. But what happens if the interpretation of the visual
information differs from physical reality? When this happens, an illusion is experienced
(Gillam, 1998; Gregory, 1998). In our everyday experiences, we often perceive the world
inaccurately. For example, when a car drives by, at a certain speed it looks like the wheels are
turning backwards. Furthermore, distances appear shorter and hills appear steeper than they
really are (Proffitt et al., 1995). And estimating a distance from a map is inaccurate when
Muller-Lyer elements (figure 1.2b) are present in the map (Gillan et al., 1999). Are these
visual illusions also reflected in our actions? Sometimes they are and the performed action
does not have the intended goal. When flying an airplane, an illusion of motion direction can
make the pilot incorrectly adjust the flying direction, which can cause an accident (Shebilske,
1981). Visual illusions are a tool to fmd out how (illusory) visual information is processed
and how this information influences visually guided actions.
Scope of this thesis
Most theories of visual perception try to explain how the visual system acb,ieves normal
veridical perception. However, veridical perception carmot easily discriminate among such
theories. They can better be tested with stimuli that will result in an unrealistic percept in one
theory and not in others. In this thesis, geometrical-optical illusions are used to test different
theories that try to explain how the visual system processes (illusory) visual information.
Gillam (1998) defmed these illusions as simple line drawings in which one or another metric
property is wrongly perceived. In the next part, theories regarding the processing of (illusory)
visual information are described.
Two visual systems: "What" and "where"
Based on anatomical and neurophysiological work, Ungerleider and Mishkin (1982)
identified two cortical visual streams or pathways (figure 1.1 ), both projecting from the
primary visual cortex (Vl). The ventral stream projects from Vl through the inferior
temporal area (TEO) to the inferior temporal cortex (TE). The dorsal stream projects from Vl
to the posterior parietal cortex (PG). The ventral stream is proposed to be involved in object
recognition (properties such as colour, texture and shape) and the dorsal stream in spatial
6

General Introduction
discrimination (properties such as direction and movement speed), and thus were labelled
"what" and "where" respectively (Mishkin et. al., 1983). The main evidence for this
dichotomy came from lesion studies in monkeys. Lesions in the inferior temporal cortex
(interruption of the ventral stream) impaired the animal's ability to visually discriminate and
recognize objects without affecting the perception of spatial relations. Conversely, with a
lesion in the posterior parietal cortex (interruption of the dorsal stream) the monkey was
unable to spatially discriminate between objects, while the ability to recognize and identify
objects was still intact.
Figure 1.1: Lateral view of the left hemisphere of a monkey. The arrows represent two cortical visual
streams or pathways, each beginning in the primary visual cortex (OC), diverging in the prestriate
cortex (OB and OA), and then going ventrally into the inferior temporal cortex (TEO and TE) or
dorsally to the inferior parietal cortex (PG). Adapted from: Mishkin eta!., 1983.
"What" and "how"
More recently, Milner and Goodale (1995; Goodale & Milner, 1992) classified the two
streams in a rather different way. The distinction in the theory of Ungerleider and Mishkin
was primarily based on the differences in the stimulus attributes or features used by the two
systems. Milner and Goodale proposed that the anatomical distinction between the ventral
and the dorsal stream corresponds to the distinction between perceptual representation
(perception) and visuomotor control (action). In other words, the emphasis is now more on
differences in the way that information is transformed for different output purposes. The
primary function of the ventral stream is recognition and identification of objects in the visual
7

Chapter 1
environment ("what" are the visual attributes of an object), while the dorsal stream mainly
processes visual information needed for the control of actions directed at those objects
("how" can I grasp an object). To identify (and later recognize) an object in the environment,
visual information in the ventral stream must be independent of different viewing conditions
and irrespective of where the body is. Therefore properties of objects are encoded relative to
other objects in the environment ( allocentric frame of reference) and the information is stored
in memory for a longer time. In contrast, to grasp an object, it is not enough to know where
an object is relative to other objects; the spatial position relative to the body needs to be
determined (egocentric frame of reference). Since the relative positions of the observer and
the goal object can change from moment to moment, the egocentric coordinates of the
object's position must be computed every time an action occurs and updated during the
execution of that action. As a consequence the visuomotor system has a very short
"memory"; information about objects will last only for a couple of seconds. Extensive
research with brain lesioned patients as well as behavioural studies with normal subjects
seem to support this distinction between separate visual systems for perception and action. In
the following, first some neuropsychological cases will be described. Then we will look for
perception-action dissociations in normal subjects. And finally alternative theories for the
"two visual systems" hypothesis will be described.
N europsycbological disorders
In the normal and intact brain, the two visual systems are supposed to work together in a
unified manner, which makes it hard to determine the function of a specific brain area.
Studying deficits in neuropsychological patients makes it easier to infer the function of a
brain area. If a patient with a lesion in one area shows specific behavioural deficits, one might
conclude that the specific brain area is responsible for that behaviour. Milner and Goodale
studied a patient, D.F., who developed visual form agnosia after carbon monoxide poisoning
(e.g. Goodale et al., 1991; Goodale et al., 1994; Milner et al., 1991; Milner and Goodale,
1995). Due to damage in the ventrolateral region of the occipital cortex (lesion in the ventral
stream), D.F. is unable to recognize and discriminate between objects. When she was asked
to verbally report the orientation of an oriented slot, her answers showed no correlation with
the real orientation (Goodale et al., 1991). Despite her inability to report the orientation of the
slot, she was able to put her hand or post a card into the slot. In another task she was shown
blocks, which all had the same surface area, but differed in length-width ratio. D.F. was
unable to perceptually distinguish between them. When D.F. was asked to indicate the width
of a block with her index finger and thumb, performance did not improve. When asked to
8

General Introduction
grasp the above-mentioned objects, D.F.'s grip size was highly correlated with the size of the
block, as in normal visually guided movements (Milner & Goodale, 1995). In normal
grasping, the opening of the hand scales in proportion to the size of the object to be grasped.
The hand starts opening as soon as it begins to move towards the object and reaches a
maximum opening (peak grip aperture) at approximately two-thirds of the movement
(Jeannerod, 1986). In short, this patient is not able to perceive the perceptual properties of
objects (such as size, shape and orientation), but her visually guided actions toward these
objects are quite accurate.
As we have just seen, patients with visual form agnosia have difficulties recognizing and
discriminating between objects. To fmd the function of a brain structure it would be more
convincing to find cases of double dissociation. A patient with damage to a specific area may
show deficits on certain tasks and normal behaviour on another related task. Another patient
with a different lesion may show normal performance on the former task and impaired
performance on the latter task. In contrast with the visual form agnosia patients, are there
patients who can recognize objects, but are unable to accurately grasp them? Patients with
damage to the posterior parietal cortex (dorsal stream) show deficits in reaching and grasping
objects, but they have no difficulty recognizing or discriminating objects. The first
description of this visuomotor deficit was given by Balint (1909) and called optic ataxia.
Perenin and Vighetto (1988) showed that, in contrast with D.F., patients with optic ataxia
were unable to adjust the orientation of their hand when reaching toward and into an oriented
slot. When optic ataxic patients are asked to pick up objects, they are not able to adjust their
grip to the shape and size of the object (Goodale et al., 1994; Jakobson et al., 1991; Jeannerod
et al., 1994). However, their perceptual estimate of the size and shape of the object is quite
accurate. In short, the "two visual systems" hypothesis of Milner and Goodale seems
strengthened by this double dissociation in patients.
Perception-action dissociation in normal subjects
One problem with the evidence from neuropsychological patients is that lesions are not
necessarily restricted to one specific brain area. Damage in several neighbouring brain areas
could result in other deficits. Another problem is that the non-damaged areas could have
taken over part of the functioning of the damaged ones. This makes it hard to determine the
function of a specific area and with that the functioning of the dorsal and ventral stream. If
there is a dichotomy between perception and action, this should also be reflected in normal
subjects.
9

Chapter I
An interesting paradigm to test whether the dichotomy between the dorsal and the ventral
stream corresponds to a distinction between perception and action (in normal subjects) is the
study of visual illusions, e.g. the Ebbinghaus (also called Titchener circles) illusion (figure
1.2a). The standard version of this illusory figure consists of two central circles of equal
diameter presented side by side, one surrounded by an array of smaller circles and the other
surrounded by an array of larger circles. When giving a perceptual judgement, the circle
surrounded by the smaller circles is reported to be larger than the one surrounded by larger
circles. While our perceptual judgements are affected by this size contrast illusion, Milner,
Goodale and colleagues (e.g. Goodale & Haffenden, 1998; Goodale & Humphrey, 1998;
Milner & Goodale, 1995) assume that there is good reason to believe that visuomotor actions
are not influenced. In order to reach out and accurately grasp an object (i.e. a disk that is
superimposed on the central circle of the Ebbinghaus illusion), the visuomotor system
requires calculation of the objects position with respect to the body (egocentric frame of
reference) and computes the absolute properties of the disk (e.g. size, distance and orientation
of the object). In short, the physical size instead of the perceptual size of an object is
computed for calibrating the grip and therefore visuomotor performance is thought to be
insensitive to the illusion.
••• ••• ••• a
c
---·-e b
d
Figure 1.2: Ebbinghaus illusion (a): although both central circles are physically the same size, the one
surrounded by large flankers is perceived smaller than the one surrounded by small flankers. Miiller
Lyer illusion (b): the shaft with the ingoing fins is perceived smaller than the shaft with outgoing fins,
but both shafts have the same length. Horizontal-vertical illusion (c): the horizontal and the vertical
line have the same length. However the vertical line is perceived longer. The orientation illusion used
by Glover and Dixon, 2001 (d): when the grating is rotated slightly clockwise with respect to vertical,
the bar is perceived more counter-clockwise than it really is (and vice versa).
10

General Introduction
Whether perception and grasping in the Ebbinghaus illusion were influenced to a different
extent, was tested in an experiment of Aglioti et al. (1995). Subjects had to verbally judge
which of the two central target disks in the Ebbinghaus illusion was larger (or smaller), and
then grasp that disc. The influence of the illusion was larger on perceptual judgements than
on the peak grip aperture (PGA). Therefore they concluded that only the perceptual task was
influenced by the illusion and that grasping remained accurate. Haffenden and Goodale
(1998, 2000) found similar results using this illusion. In the perceptual task subjects did not
give a verbal judgement, but they matched the distance between their index fmger and thumb
to the diameter of the central target disk (manual size estimation). A target disk surrounded
by an array of large circles was estimated to be smaller than a same sized target disk
surrounded by an array of small circles. Thus the manual size estimates were influenced by
the illusion. When grasping the central target disk, the PGA showed a much smaller effect of
the illusion. Westwood et al. (2000a) used the Muller-Lyer illusion (figure 1.2b) to
demonstrate that the PGA was not affected by contextual cues that induce a length illusion.
Subjects were required to give a manual size estimate or to grasp a bar that was superimposed
on the shaft between the outgoing and the ingoing fins. Although the length of both shafts in
this illusion is physically equal, the shaft between the outgoing fms is perceived to be longer
than that between the ingoing fins. The large illusory effect that was found on manual size
estimation was not reflected in the PGA. Otto-de Haart et al. (1999) used the same illusion to
show that the PGA remained unaffected by the illusion under monocular viewing conditions.
Several other studies support the idea that pointing and grasping are relatively immune to
illusions (Danckert et al., 2002; Dyde & Milner, 2002; Haffenden et al., 2001). Differences in
illusion magnitudes between perceptual tasks and visuomotor tasks are not only found in
upper limb movements, but also in lower limb movements (Loomis et al., 1992; Wraga et al.,
2000), saccadic eye movements (Wong & Mack, 1981), ocular tracking (Bridgeman et al.,
1979) and in illusions presented in the haptic domain (Westwood & Goodale, 2003).
As described before, the perceptual system operates over a long timescale. On the other hand,
the absolute properties used for controlling an action, change rapidly from moment to
moment. Thus it makes sense to compute the required information immediately before
initiation of a movement, in real time. The representations in the visuomotor system will
rapidly decay and when a movement is initiated after a time delay, movement execution is
thought to depend on the stored perceptual representation. In that case visual illusions can
affect action. Several studies have shown increasing effects of illusions in action tasks after a
delay of several seconds (Bradshaw & Watt, 2002; Bridgeman et al., 1997; Gentilucci et al.,
1996; Hu & Goodale, 2000; Westwood et al., 2000a, 2000b, 2001; Wong & Mack, 1981).
11

Chapter 1
Theories challenging the perception-action dissociation
According to the "two visual systems" hypothesis visually guided movements are largely
immune to visual illusions. Indeed, often a smaller illusion magnitude was found on grasping
or pointing than on perception. However, a significant difference between perception and
action does not mean that the action remains accurate. In some studies a small effect on
action is found, even when there is no delay between presentation of the stimulus and the
performed action (Aglioti et al., 1995; Haffenden & Goodale, 1998). Haffenden and Goodale
(2000; Haffenden et al., 2001) suggested that this effect is not caused by the illusion, but that
two-dimensional elements, serving as an illusory background, can be treated by the
visuomotor system as potential obstacles and thereby influence the grasp. For instance, if the
surrounding circles in the Ebbinghaus illusion are treated as obstacles, which must be
avoided, the distance between the middle target circle and the array of surrounding circles
can be seen as a gap to reach into, in order to grasp the target. In the standard Ebbinghaus
illusion (figure 1.2a), the gap between the array of large circles and the central target disk is
large enough to put the fingers in. When picking up the target disk in this configuration the
fingers will open just enough to fit into the gap. However, the gap between the array of small
circles and the central target disk is too small for the fingers to fit into it. Thus when picking
up the disk surrounded by small circles the whole configuration is taken into account and the
fingers will open larger (Haffenden & Goodale, 2000).
Several other studies, which have found results that cannot easily be explained by the "two
visual systems" hypothesis, questioned the notion that the visuomotor system resists visual
illusions. The illusion magnitude in action tasks can equal the one in perception (Franz et al.,
2000; Pavani et al., 1999) or can even be larger than on perception (Glover & Dixon, 2001a;
Moo-Williams & Bull, 2000; Yarnagishi et al., 2001). In the next part alternative
explanations for the "two visual systems" hypothesis are presented.
The common source model
Franz et al. (2000) argued that in the study of Aglioti et al. (1995) the perceptual and the
grasping task were not properly matched, which resulted in an apparent dissociation between
perception and action. In the perceptual task of Aglioti et al. both Ebbinghaus configurations
were presented simultaneously. Subjects had to directly compare the size of both central
circles. However, in the grasping task of Aglioti et al. no such relative comparison is made
since the subjects grasped only one disk at a time. The differential effect of the illusion on
12

General Introduction
perception and action may depend on the fact that in the perceptual task subjects were
influenced by two configurations (relative comparison of the sizes of two objects), whereas in
the grasping task only one configuration influenced the peak grip aperture (absolute size),
resulting in a reduced illusion effect in grasping. To eliminate this problem Franz et al.
(2000) presented only one Ebbinghaus configuration at a time. They found similar effects of
the illusion on perceived size and the PGA, as did Pavani et al. (1999). Vishton et al. (1999)
confirmed that the difference in illusion magnitude between grasping and perceptual size
judgements disappears when making absolute instead of relative judgements. They let
subjects make perceptual judgements and a (pantomimed) grasp to one of the lines in the
horizontal-vertical illusion (figure 1.2c). Grip scaling showed only a very small effect of the
illusion. Perceptual judgements were affected to a much bigger extent when the length of one
line had to be expressed as a percentage of the other. However, perceptual judgements
directed at a single line, showed no difference in illusion magnitude between perception and
grasping. Since in grasping tasks the same illusion magnitude can be found as in perceptual
tasks, Franz et al. (2000) suggested that a common source of information, which is influenced
by illusions, is used to generate a visual percept as well as to guide grasping. In case of the
Ebbinghaus illusion this means that the perceptual illusory size is also used in grasping
movements. This interpretation will be referred to as the common source model.
The planning-control model
Glover and Dixon (200la) suggested that the difference in illusion effects found in action can
be understood by distinguishing between the planning and control phase of a movement. In
this planning-control model the planning and initiation of a movement is based on a context
dependent visual representation that is subject to visual illusion. During the execution of the
movement, errors in planning caused by visual illusions can be corrected on-line through
visual and proprioceptive feedback and an efference copy of the movement. This model
assumes that actions are always guided by two representations (a context dependent
representation in early stages of the movement and a context independent representation in
the later stages). Remember that the "two visual systems" hypothesis assumes that actions are
guided by a single, context independent representation. Support for the planning-control
model was found in experiments in which Glover and Dixon investigated the effect of an
orientation illusion on a reach to grasp movement. Subjects were required to reach out and
grasp a bar that was superimposed on an oriented background grating (figure 1.2d). When the
grating is tilted lOa clockwise from sagittal, the orientation of the bar is perceived slightly
more counter-clockwise than it actually is, and vice versa. In grasping, the orientation of the
13

Chapter 1
hand was measured throughout the reach. They found a dynamic illusion effect: the illusion
had a relatively large initial effect and decreased over time as the hand approached the target
(Glover & Dixon, 200la, 200lb). Many studies that measured the PGA, which occurs at
about two-third of the movement, concluded that grasping was not influenced by the illusion
(Aglioti et al., 1995; Haffenden & Goodale, 1998; Otto-de Haart et al., 1999; Westwood et
al., 2000a, 2000b ). According to Glover and Dixon, subjects have already largely corrected
their movement at the moment of PGA and thus it is not surprising that no effect is found.
Glover and Dixon (2001c, 2002) also observed the dynamic illusion effect when vision of the
hand and target was removed during reaching, showing that visual feedback is not necessary
for the on-line corrections. They interpret this as evidence that other sources of information
such as proprioceptive feedback can also be used. Contrary to the predictions of the planning
control model, Franz (2003) found that the effects of the Ebbinghaus illusion did not decrease
over time. Instead, the illusion effects stayed constant over time. Danckert et al. (2002) also
looked at the time course of the magnitude of the Ebbinghaus illusion on grasping, but found
no evidence for an effect even at the earliest stages of the movement. These inconsistent
results are mainly due to differences in analyses (Glover, 2002; Franz, 2003; Smeets et al.,
2003).
Inconsistent processing of attributes
There is another explanation for the finding that illusions can affect the PGA during grasping.
Smeets and Brenner (1995a) suggested that different spatial attributes (e.g. length, position)
are used in a perceptual task than in grasping. These spatial attributes, which are physically
related, are not necessarily perceived in a consistent manner because they are processed
independently (Gillam & Chambers, 1985; Gillam, 1998; Mack et al., 1985; Post & Welch,
1996; Smeets & Brenner, 1995a). For instance, the Milller-Lyer illusion induces
inconsistencies between length and position. This illusion changes the perceived length of the
shaft, without changing the perceived end-positions (figure 1.3). Such inconsistencies have
also been found between velocity and position (Smeets & Brenner, 1995a). Smeets and
Brenner stated that a task, irrespective of whether this task is classified as perception or
action, is only influenced by an illusion if an incorrectly perceived attribute is used to
perform that task. For example, when illusory length of the shaft of the Muller-Lyer illusion
is used in a task, e.g. in a perceptual size judgement task, an effect of the illusion is found.
When grasping a bar that is superimposed on the shaft of this illusion, the PGA is not
determined by the size of the bar, but by the positions at which the bar is grasped (Smeets &
Brenner, 1999) and thus no influence of the illusion is found. This was further tested in an
14

General Introduction
experiment (Brenner & Smeets, 1996) in which subjects were asked to pick up a disk on a
background of converging lines (Ponzo illusion). A disk placed on the converging side ofthe
background looks larger than the same disk placed on the diverging side. Brenner and Smeets
predicted that, as the object's size is used to estimate its weight, the illusory change in size
would affect the force with which the object is lifted. However the PGA would not be
influenced by this size illusion since in order to grasp the object the fmger and thumb are
directed towards positions on the object. Indeed, the results confirmed their predictions.
Jackson and Shaw (2000) also showed that the PGA was not influenced by the Ponzo
illusion, whereas another aspect of grasping, maximum grip force, was influenced. Many
studies in which the perceptual task is more strongly affected by an illusion than the PGA
during grasping, and which are therefore often considered as evidence for the "two visual
systems" hypothesis, can be explained with this approach (reviewed by Smeets et al., 2002).
a b
Figure 1.3: The length of the shafts in figure 1.3(a) and (b) is perceived differently, but the points of
the arrow in (a) seem perfectly aligned with the vertical lines in (b).
Short outline of chapters 2 through 5
A lot of studies used visual illusions to fmd out how (illusory) visual information influences
our activities. In this thesis we tried to distinguish between several theories: which theory can
best explain the large variety in experimental results? Broadly outlined, the two visual
systems hypothesis predicts an effect of illusions in perceptual tasks, not in visuomotor tasks.
The common source model states that the same illusory information is used in perception and
action and thus the same illusion effects are expected for both tasks. In the planning-control
model an effect of an illusion will only be present in the planning phase of a movement, not
in the online-control phase. If spatial attributes are inconsistently processed, illusion effects in
a task are expected if the affected attribute is used in that task.
15

Chapter 1
In chapter 2 we asked whether studies, that found a difference in the Roelofs effect between
perceptual and visuomotor tasks, can be a valid confirmation for the two visual systems
hypothesis. We investigated this by letting subjects perform two highly similar perceptual
tasks with the Roelofs effect
Not only did we investigate perceptual tasks, in chapter 3 we focused on visuomotor tasks.
As described before, sometimes effects of an illusion can be found on a visuomotor task.
Several theories have given an explanation for an illusion affecting grasping. According to
the common source model perceptual information, which is influenced by an illusion, is used
to perform a grasping movement. The explanation of Smeets and Brenner states that grasping
is expected to be influenced by an illusion only if an affected attribute is used in performing
the task. In a grasping experiment we evaluated these explanations.
The amount of illusion magnitudes that have been found in visuomotor tasks differs between
studies. According to the theory of Smeets and Brenner this could be caused by the use of a
combination of attributes (affected and unaffected ones). In chapter 4 we asked whether we
could manipulate the ratio of attributes that is used in a visuomotor task (and with that the
amount of illusory effect) by changing the task requirements. To answer this question
subjects were asked to point to the Brentano illusion under different visibility conditions.
In most studies grasping and/or pointing are used to perform an action. In chapter 5 we tried
to fmd out whether the effect of the Brentano illusion found in pointing generalizes to eye
movements.
16

Chapter 2
Are the original and the induced Roelofs effect caused
by the same shift in straight ahead?
Published as: De Grave, D.D.J., Brenner, E., & Smeets, J.B.J. (2002). Are the original
Roelofs effect and the induced Roelofs effect caused by the same shift in straight ahead?
Vision Research, 42, 2277-2283.

Chapter 2
Abstract
We investigated whether the original Roelofs effect and the induced Roelofs effect are caused
by the same shift in perceived straight-ahead. Subjects were presented with a target within a
frame in complete darkness. Target and frame could both be shifted to the left or right of
objective straight-ahead. On separate trials, subjects gave verbal estimates about the position
of either the target or the frame. The eccentricity of the frame was underestimated (the
original Roelofs effect). However, the perceived position of the target did not follow this
rnisjudgement of the eccentricity of the frame (the induced Roelofs effect was not present).
Thus, it is unlikely that both effects have a common origin in misjudging egocentric straight
ahead.
18

The original and the induced Roelofs effect
Introduction
Many studies have examined whether there 1s a difference between how susceptible
perception and action are to visual illusions (reviewed by Carey, 2001). Often, visual
illusions cause biases in perceptual tasks, but do not influence goal directed motor output.
One example of a dissociation between perceptual and motor tasks was reported by
Bridgeman et al. (1981), who showed that induced motion (apparent motion of a target when
the background moves) affected pointing less than it did perceptual judgements. The
perceptual judgement was whether the target had moved. In the motor task, subjects had to
point to the final target position.
Smeets and Brenner (1995a) noted that motion (or displacement) and position were
confounded in the above-mentioned study. They demonstrated that motion of the background
led their subjects to misjudge the movement of the target but that it did not have an effect on
their judgement of position. Therefore, they argued that the results of Bridgeman et al were
not an indication for a dissociation between perception and action, but for one between
position and motion (or displacement). A similar dissociation between position and size could
account for the apparent absence of an influence of illusions on grasping (Brenner & Smeets,
1996; Smeets & Brenner, 2001).
Bridgeman et al. (1997), Bridgeman and Huemer (1998) and Bridgeman et al. (2000) also
found a dissociation between perception and action in experiments that do not involve motion
or size. They induced a bias in a target's apparent position by placing a frame around the
target. An offset of the frame to one side of the subject's objective median plane caused a
bias in the perceived target position in the opposite direction (the induced Roelofs effect).
Goal directed arm movements towards that target remained accurate despite the perceptual
mislocalization. Their interpretation is that the results are caused by a dissociation between
perception and action. These results cannot easily be explained by the arguments of Smeets
and Brenner (1995a, 2001).
In order to investigate whether it is possible to explain Bridgeman's findings without
assuming that there are two visual systems, we would like to know what causes the induced
Roelofs effect. To do so, we first turned to the original Roelofs effect. Roelofs (1935) showed
that if a rectangular frame is presented with one of its edges on the subject's objective median
plane, that edge seems to be shifted to the side opposite to the rest of the frame. Werner et al.
(1953) showed that the original Roelofs effect was not only present when one edge of the
19

Chapter 2
frame was on the objective median plane, but also with a more subtle shift of the frame. They
presented a single luminescent frame in a darkened room The frame was shifted a little to the
right or to the left of the objective median plane. The subject's task was to fixate the centre of
the frame and indicate the position in space that appeared straight-ahead. Werner et al. (1953)
found systematic shifts of the position of the apparent straight-ahead in the direction of the
centre of the frame.
In the present experiment we investigate whether the original Roelofs effect and the induced
Roelofs effect could be caused by a shift in perceived straight-ahead (figure 2.1). If so, it
would be all the more surprising that such a shift does not influence action because an
illusory shift in perceived egocentric location is one of the most likely kinds of illusions to
influence action (Smeets & Brenner, 2001; Rossetti, 1998).
original Roelofs effect
induced Roelofs effect
stimulus
I I
0
•
response hypothesis
I I
0 0
• • 0
Figure 2.1: A hypothetical explanation for both the original and induced Roelofs effect. When a frame is shifted to the left, its eccentricity is underestimated; i.e. it is judged too far to the right (original Roelofs effect). The hypothesis is that the perceived straight-ahead (indicated by the head orientation) shifts to the left, in the direction of the frame. The same hypothesis can explain why the judged position of the target also shifts to the right (induced Roelofs effect).
If the original and induced Roelofs effects are both caused by a shift in perceived straight
ahead we can make two predictions. The first prediction concerns the induced Roelofs effect.
If the frame modifies the judgement of what is straight-ahead, we expect the order of
presentation to make a difference. In the present experiment three orders of presentation are
20

The original and the induced Roelofs effect
used: target shown before frame, both shown simultaneously or frame shown before target.
When the target is presented first, we expect target position (relative to straight-ahead) to
have been determined before the frame shifted the "straight-ahead", so no induced Roelofs
effect is expected. When the target and the frame are presented simultaneously, we expect to
fmd an induced Roelofs effect. When the frame is presented first the effect of the frame on
the straight-ahead might still be present, so we expect to fmd an induced Roelofs effect.
These predictions were tested in a first set of trials ("single task"), in which subjects had to
estimate the target position.
The second prediction concerns the relation between the induced and the original Roelofs
effect. If a shift in the judged straight-ahead underlies both effects, the magnitude of the
original and of the induced Roelofs effect should be the same. We tested this hypothesis
directly with a second set of trials ("dual task"), in which subjects had to estimate both the
position of the target and of the frame. Here too we varied the order of presentation.
Materials and methods
Subjects
Six subjects participated in the single task and six other subjects in the dual task. All had
normal or corrected-to-normal vision and were naive with regard to the purpose of this study.
Apparatus and stimulus
The experiments were conducted in a dark room. A chin-rest was placed 50 em from a
computer screen (39.2 x 29.3 em, 815 x 611 pixels, 120Hz) to help subjects keep their head
directed at the midpoint of the screen. All measures are given in centimetres relative to this
midpoint. At this distance, 1 centimetre corresponded to 1.15°. To prevent the subject from
seeing any reflections from the screen, subjects looked into a "box" made of black curtains,
and a red filter was placed in front of the computer screen.
The stimulus on the screen consisted of a small red target disk and/or a large red rectangular
frame. Both were presented at eye height, at various horizontal positions. The diameter of the
target disk was 0.75 em. The frame width was 18.3 em. Its height was 7.4 em. Line-width of
the frame was 0.3 em.
21

Chapter 2
Procedure
Single task
The experiment started with a training phase. During this phase only the target was shown.
On each trial of the training phase, subjects had to verbally estimate the target's position
relative to straight-ahead (in centimetres). Feedback about the real target position (in
centimetres) was then shown as a number on the screen. The range of target positions was
from 5 em to the left of the subject's objective median plane to 5 em to the right, in steps of
0.5 centimetres. Leftward offsets were considered negative. Thus, if the subject thought that
the target was three centimetres to the left of straight-ahead, he would answer minus three.
The training lasted at least 10 minutes. After this, the training ended when five consecutive
trials were considered correct.
Before participants started the test phase, they were told that no more feedback would be
given. Both the target and the frame were presented in each trial of the test phase. The target
could be presented at five positions: either straight-ahead, or 3 or 1.5 em to the left or right of
the objective midline. The frame could also be presented at five different positions either
straight-ahead, or 5 or 2.5 em to either side of the objective midline. The target and frame
were each visible for 500 ms and could be shown in three ways. In the simultaneous
condition both were presented together for 500 ms. In the target first condition the target was
presented for 500 ms, followed by a 100 ms dark interval and then the frame for 500 ms. For
the frame first condition, the order of presentation was reversed. Again, subjects had to
verbally estimate the position of the target (in centimetres) relative to straight-ahead. Each of
the five frame positions was combined with each of the five target positions in every order of
presentation, and each combination of positions was shown once. This gives a total of 75
trials: 5 frame positions, 5 target positions and 3 orders of presentation.
In order not to give the subjects any spatial reference when giving the response, the
experimenter registered the verbal responses on a keyboard. The subject could not see this
person (or anything else in the room). Total duration of the experimental session was about
half an hour.
Dual task
The training phase of this task was similar to that of the single task, except that either the
target or the frame was shown on each trial. The subject had to judge the position of the
presented object (target or frame). Subjects never saw both target and frame on a single trial
22

The original and the induced Roelofs effect
of the training phase. The order in which they were presented was random. The minimum
duration of the training was 15 minutes.
The stimuli in the test phase of the dual task were exactly the same as in the test phase of the
single task. Subjects had to estimate the position of either the target or the frame. They did
not know which position they had to estimate when they saw the stimulus. If a short sound
was presented 500 ms after the stimulus the estimate was for the frame. Otherwise, it was for
the target.
Every frame-target combination was presented twice: once to estimate the target position and
once to estimate the frame position. This gives a total of 150 trials: 5 frame positions, 5 target
positions, 3 orders of presentation and 2 estimates. All trials were presented in a random
order. Total duration of the experimental session was about forty-five minutes.
Data analysis
In order to quantify the influence of frame position on the subjects' judgements, we fit a
linear model to the data. Bridgeman et al. (1997, 2000) have shown that the relationship
between the target's position and the subjects' judgements of that position is linear. We
assume that the same is true for the relationship between the frame's position and the subjects'
judgements of the target's position, and that the effects of frame position and target position
are independent. We can therefore perform a multiple regression to determine how the
positions of the target and of the frame influence the perceptual judgements of the target's
position. This was done separately for every subject. A similar analysis was conducted for the
judgement of the frame's position. These analyses were performed for each order of
presentation. The regression coefficients (the frame and target gains) show how much the
response depends on a change in the independent variables (frame position and target
position). A gain of 1 means that the change in response is equal to the change in the
independent variable. A gain of 0 means that the response does not depend on the
independent variable.
Statistical tests were all conducted across subjects. One-sample t-tests were performed to
check whether the gains differed from 0 or I (to check whether there is any effect). If the
frame gain of the estimate of the target's position is significantly smaller than 0, there is an
induced Roelofs effect. If the frame gain of the estimate of the frame's position is smaller
than 1, i.e. if subjects underestimate the eccentricity of the frame, there is an original Roelofs
23
Chapter 2
effect. We are primarily interested in the effect of the frame (frame gains), but we also
examined whether subjects underestimated the eccentricity of the targets (target gain< 1). A
one way repeated measures ANOV A was performed on the frame gains to see whether the
order of presentation (frame first, simultaneous, target fust) made a difference.
Results
Single task
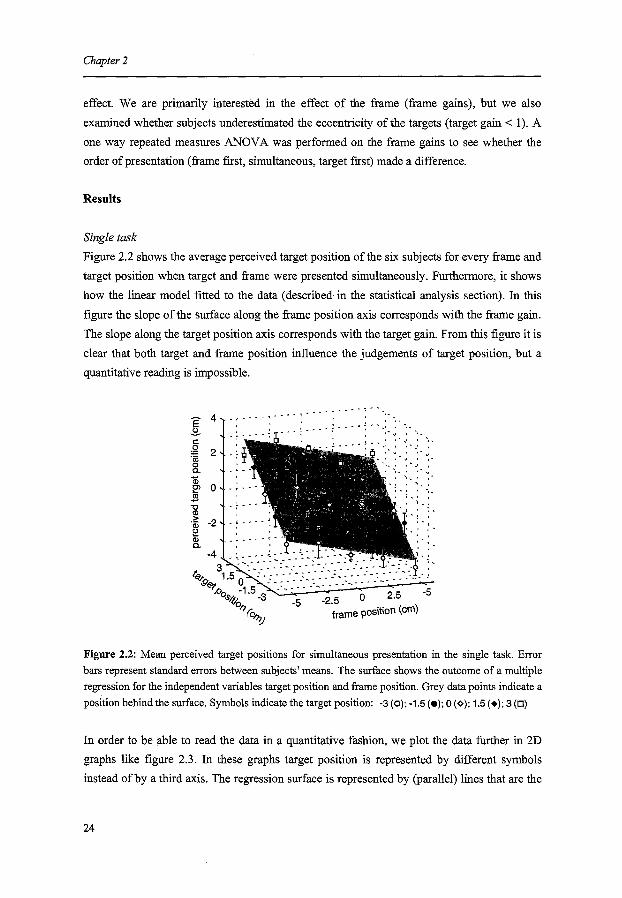
Figure 2.2 shows the average perceived target position of the six subjects for every frame and
target position when target and frame were presented simultaneously. Furthermore, it shows
how the linear model fitted to the data (described in the statistical analysis section). In this
figure the slope of the surface along the frame position axis corresponds with the frame gain.
The slope along the target position axis corresponds with the target gain. From this figure it is
clear that both target and frame position influence the judgements of target position, but a
quantitative reading is impossible.
"E 4 .3. c:
.Q 2 ~ 0 a. a; i 0 "C
~ ·a; -2 e Q) a.
-4
' ,. ~ ~ - ' ~· ' ... -
.-,, ' ·,_
Figure 2.2: Mean perceived target positions for simultaneous presentation in the single task. Error
bars represent standard errors between subjects' means. The surface shows the outcome of a multiple
regression for the independent variables target position and frame position. Grey data points indicate a
position behind the surface. Symbols indicate the target position: -3 (o); -1.5 (e); 0 (<>); 1.5 (+); 3 (D)
In order to be able to read the data in a quantitative fashion, we plot the data further in 2D
graphs like figure 2.3. In these graphs target position is represented by different symbols
instead of by a third axis. The regression surface is represented by (parallel) lines that are the
24
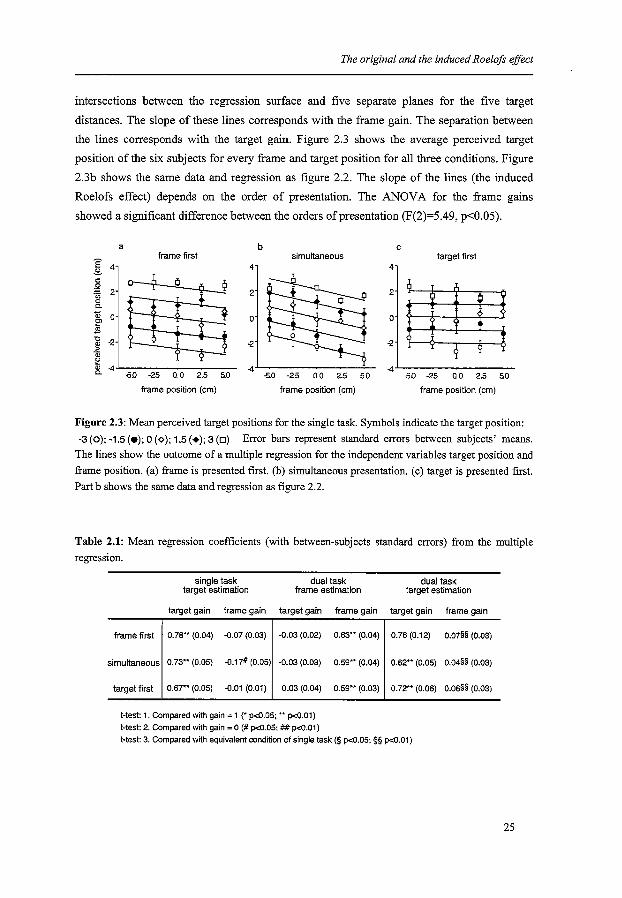
The original and the induced Roelofs effect
intersections between the regression surface and five separate planes for the five target
distances. The slope of these lines corresponds with the frame gain. The separation between
the lines corresponds with the target gain. Figure 2.3 shows the average perceived target
position of the six subjects for every frame and target position for all three conditions. Figure
2.3b shows the same data and regression as figure 2.2. The slope of the lines (the induced
Roelofs effect) depends on the order of presentation. The ANOV A for the frame gains
showed a significant difference between the orders of presentation (F(2)=5.49, p<0.05).
a b c
E frame first simultaneous target first
4 4 4 .£,
~ t::
.Q 2 2 ·]i 2
0 c. 05 0 0
"' 0
.$ -a .,2 .,2 .,2 Q)
.a: Q)
~ Q) -4 -4 -4 c. -6.0 -25 ao 2.5 50 <;Q -25 ao 2.5 50
frame position (em) frame position (em) frame position (em)
Figure 2.3: Mean perceived target positions for the single task. Symbols indicate the target position:
-3 (o); -1.5 (e); 0 (<>); 1.5 (+); 3 (D) Error bars represent standard errors between subjects' means. The lines show the outcome of a multiple regression for the independent variables target position and frame position. (a) frame is presented first. (b) simultaneous presentation. (c) target is presented first. Part b shows the same data and regression as figure 2.2.
Table 2.1: Mean regression coefficients (with between-subjects standard errors) from the multiple regression.
frame first
simultaneous
target first
single task target estimation
target gain frame gain
0.78 .. (0.04) -O.o? (0.03)
0.73** (0.05) -0.17# (0.05)
0.67*' (0.05) -O.Q1 (0.01)
dual task frame estimation
target gain frame gain
-0.03 (0.02) 0.63** (0.04)
-0.03 (0.03) 0.59** (0.04)
0.03 (0.04) 0.59 .. (0.03)
t-test: 1. Compared whh gain = 1 (* p<0.05; •• p<0.01)
t-test: 2. Compared whh gain = 0 (# p<O.OS: ## p<0.01)
dual task target estimation
target gain frame gain
0.76 (0.12) o.o7§§ (0.03)
0.62** (0.05) 0.04§§ (0.03)
0.72 .. (0.06) o.o6§§ (0.03)
t-test: 3. Compared whh equivalent condition of single task (§ p<0.05; §§ p<0.01)
25
Chapter 2
The leftmost column of table 2.1 shows the mean target and frame gains with their standard
errors. As expected, we found an induced Roelofs effect in the simultaneous condition (see
table 2.1). We found no induced Roelofs effect when the target was presented first.
Presumably, the frame position was ignored because subjects judged the target's position
before the frame was presented. There was also no significant induced Roelofs effect when
the frame was presented first. In all cases the eccentricity of the target was underestimated
(target gains significantly smaller than one).
Dual task
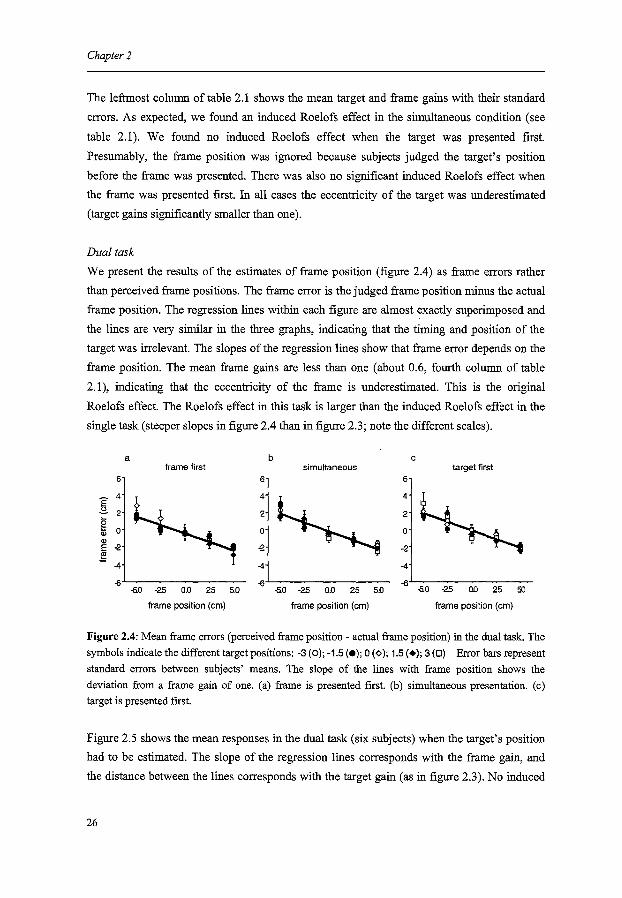
We present the results of the estimates of frame position (figure 2.4) as frame errors rather
than perceived frame positions. The frame error is the judged frame position minus the actual
frame position. The regression lines within each figure are almost exactly superimposed and
the lines are very similar in the three graphs, indicating that the timing and position of the
target was irrelevant. The slopes of the regression lines show that frame error depends on the
frame position. The mean frame gains are less than one (about 0.6, fourth column of table
2.1 ), indicating that the eccentricity of the frame is underestimated. This is the original
Roelofs effect. The Roelofs effect in this task is larger than the induced Roelofs effect in the
single task (steeper slopes in figure 2.4 than in figure 2.3; note the different scales).
6
E 4 .e 2 g "' 0 "' E -e _g
-4
a frame first
~·~--~--------~--6.0 -25 0.0 25 5.0
frame position (em)
b simultaneous
6
-4
~~----------------6.0 -25 0.0 25 5.0
frame position (em)
c target first
6
-4
"6'~-6.::-:0--:-25-:--0.0--:2::-:5--:50:-
frame position (em)
Figure 2.4: Mean frame errors (perceived frame position - actual frame position) in the dual task. The symbols indicate the different target positions: -3 (O); -1.5 (e); 0 (¢); 1.5 (+); 3 (D) Error bars represent standard errors between subjects' means. The slope of the lines with frame position shows the deviation from a frame gain of one. (a) frame is presented first. (b) simultaneous presentation. (c) target is presented first.
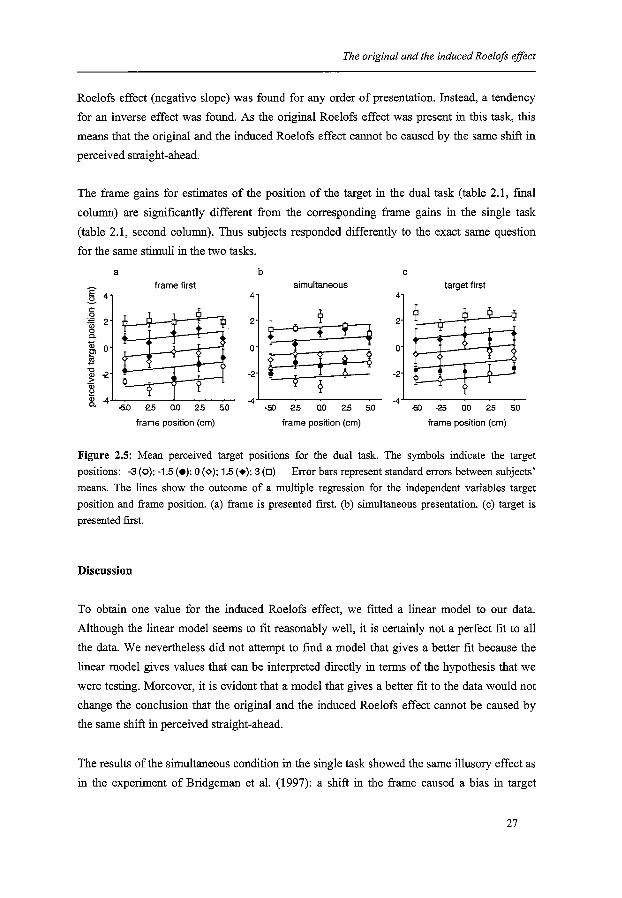
Figure 2.5 shows the mean responses in the dual task (six subjects) when the target's position
had to be estimated. The slope of the regression lines corresponds with the frame gain, and
the distance between the lines corresponds with the target gain (as in figure 2.3). No induced
26
The original and the induced Roelofs effect
Roelofs effect (negative slope) was found for any order of presentation. Instead, a tendency
for an inverse effect was found. As the original Roelofs effect was present in this task, this
means that the original and the induced Roelofs effect cannot be caused by the same shift in
perceived straight-ahead.
The frame gains for estimates of the position of the target in the dual task (table 2.1, fmal
column) are significantly different from the corresponding frame gains in the single task
(table 2.1, second column). Thus subjects responded differently to the exact same question
for the same stimuli in the two tasks.
E"4 .::!. c: 0
~ 2
8. 1i5 0
~ ~ -2 ·a; ~
a
2i 4 -50
frame first
frame position (em)
b c simultaneous target first
·50 -25 0.0 25 50 -50 -25 00 25 50
frame position (em) frame position (em)
Figure 2.5: Mean perceived target positions for the dual task. The symbols indicate the target positions: -3 (o); -1.5 (e); 0 (~); 1.5 (+); 3 (D) Error bars represent standard errors between subjects' means. The lines show the outcome of a multiple regression for the independent variables target position and frame position. (a) frame is presented first. (b) simultaneous presentation. (c) target is presented first.
Discussion
To obtain one value for the induced Roelofs effect, we fitted a linear model to our data.
Although the linear model seems to fit reasonably well, it is certainly not a perfect fit to all
the data. We nevertheless did not attempt to fmd a model that gives a better fit because the
linear model gives values that can be interpreted directly in terms of the hypothesis that we
were testing. Moreover, it is evident that a model that gives a better fit to the data would not
change the conclusion that the original and the induced Roelofs effect cannot be caused by
the same shift in perceived straight-ahead.
The results of the simultaneous condition in the single task showed the same illusory effect as
in the experiment of Bridgeman et al. (1997): a shift in the frame caused a bias in target
27

Chapter 2
position in the opposite direction to the offset of the frame (an induced Roelofs effect). The
frame gain in this condition (0.17) was similar to that in Bridgeman et al's comparable
studies (0.15 and 0.27 in the 1997 and 2000 papers). Target gain was slightly larger in our
study (0.73 rather than about 0.6). We also reproduced the original Roelofs effect in the
frame estimation trials of the dual task. When the frame was not straight-ahead its
eccentricity was underestimated, consistent with the fmdings of Roelofs (1935) and Werner
et al. (1953). This result was totally independent of the timing and location of the target.
Thus, we could replicate the results of the studies that were the basis of our hypothesis that
misestimation of straight-ahead is the common source of the original and induced Roelofs
effect.
If egocentric straight-ahead is misestimated, the extent of the mislocalization of target and
frame should be the same. In the dual task, the original and induced Roelofs effects were not
equal. The target did not even shift in the same direction as the frame. A common origin of
both effects in misjudging egocentric straight-ahead is thus very unlikely. The disappearance
of the induced Roelofs effect in our dual task has an important additional consequence.
The presence of an induced Roelofs effect in perceptual tasks and the failure of this
perceptual illusion to' influence action has been interpreted as evidence for separate visual
mechanisms for perception and action (Bridgeman et al., 1997; Bridgeman et al., 2000).
Perceptual judgements were reported by indicating the target position relative to the subject's
midline on a visible ruler (Bridgeman et al., 1997) or by estimating the target position in a 5
alternative forced choice task either verbally (Bridgeman et al, 2000) or by pressing a key on
a keyboard (Bridgeman et al., 1997). Despite these differences between the perceptual tasks,
there was always an effect of the illusion. Therefore, the absence of the induced Roelofs
effect in pointing tasks was regarded as a strong argument for a dissociation between
perception and action.
In our experiment we found that the influence of the illusion on perceptual judgements could
disappear when a small variation was introduced to the task, although exactly the same
stimuli and response method were used. The main difference between the target estimation
responses in the single task and the dual task is that subjects are more aware of the fact that
the frame could have different positions in the latter case. The frame estimation part of the
dual task emphasizes this. Consequently, whereas the frame might be regarded as a useful
reference in the single task, it is evident that it is not a reference in the dual task. Therefore
the only useful information to judge the target position in the dual task is egocentric
information. In pointing or jabbing it is evident from the task itself that egocentric
28

The original and the induced Roelofs effect
information about target location is more useful than information relative to a frame. The
critical difference between the tasks is therefore probably the extent to which allocentric
information can be considered useful information, rather than the kind of response (Vishton
et al., 1999). We conclude that the absence of the Roelofs effect in action should not be
considered as evidence for a dissociation between visual processing of spatial information for
perception and action.
29


Chapter 3
Effects of the Ebbinghaus figure on grasping are not
only due to misjudged size
Accepted as: De Grave, D.D.J., Biegstraaten, M., Smeets, J.B.J. & Brenner, E. Effects of the
Ebbinghaus figure on grasping are not only due to misjudged size. Experimental Brain
Research.

Chapter 3
Abstract
It is not evident how the small effects of the flankers of the Ebbinghaus figure on peak grip
aperture should be interpreted. One interpretation is that the flankers influence a size estimate
that is used in grasping. If this interpretation is correct, then no aspects of the grasping
movement should depend on the spatial positions of the flankers. An alternative interpretation
is that the effect on grip aperture is caused by a change in judgements of the required
precision, in which case various aspects could be influenced by the size and position of the
flankers.
We presented subjects with a display consisting of a central disk surrounded by four large or
small flankers. The array of circular flankers could be rotated by 45°. There were two tasks:
to reproduce the perceived size of the central disk, and to grasp the central disk. As in other
studies, the perceived size and the peak grip aperture were influenced by the size of the
flankers. The effect on perceived size settings was independent of the flankers' spatial
position. Nevertheless, the flankers' position did influence the final grip aperture and the grip
orientation at peak grip aperture and at movement offset. As the flankers changed more than
only the peak grip aperture, we conclude that the effect of the flankers on prehension cannot
only be due to a misjudgement of the central disk 's size.
32

Effects of the Ebbinghaus figure on grasping
Introduction
The Ebbinghaus illusion is the phenomenon that a central circle surrounded by large circular
flankers is perceived to be smaller than a similar central circle surrounded by small flankers.
Aglioti et al. (1995) performed an experiment in which subjects had to judge which of two
central target disks was larger (or smaller), and then to grasp that disk. The influence of the
illusion on peak grip aperture (PGA) was smaller than on the perceptual settings. They
concluded that despite the illusion the grasping movements remained accurate. A study by
Haffenden and Goodale (1998) gave similar results: two sizes of flankers had significantly
different effects on manual size estimation, but not on grasping. However, in both these
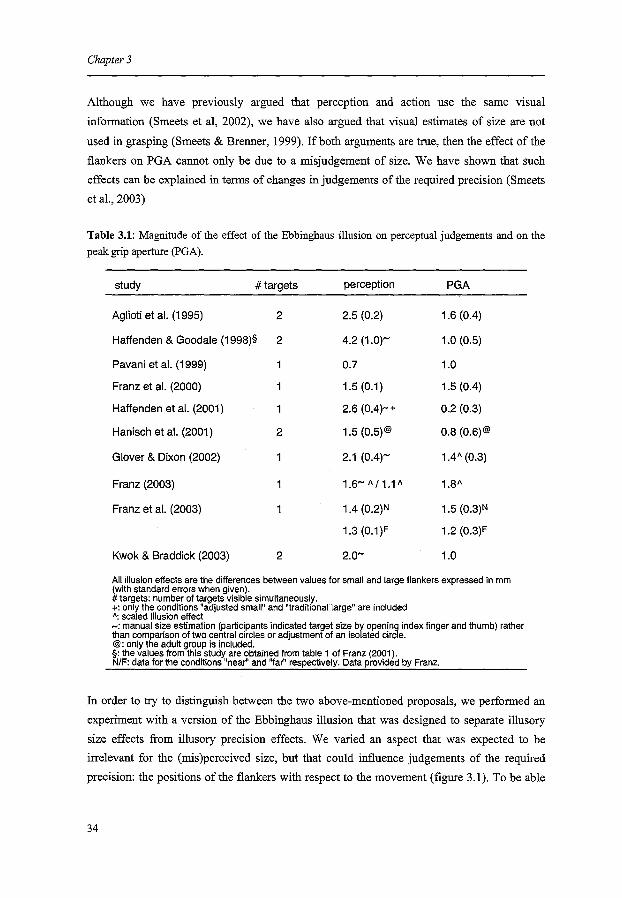
studies the PGA did show some effect of the illusion (see table 3.1). Wbat causes this effect
of the flankers on grasping?
Franz et al. (2000) proposed that the same illusory size information is used in perception and
in grip scaling. They showed that if the perceptual task is designed to more closely match the
grasping task (i.e. to involve a single target) the effects on grasping and perception are not
significantly different (Pavani et al., 1999; Franz et al., 2000, 2003). An overview of the
experimental results (table 3.1) seems to support the interpretation that the PGA and the
perceptual measure are influenced to the same extent if a single target is used (Haffenden et
al, 2001 is an exception). We will refer to this view as the "illusory size hypothesis".
Haffenden and Goodale (2000, also see Haffenden et al., 2001) proposed that the small effect
of the Ebbinghaus illusion on grip scaling does not result from misperceiving the size, but
from a direct influence of the flankers on the movement path (obstacle avoidance). They
predicted that two-dimensional rectangular flankers (potential obstacles) surrounding a target
disk would affect grasping if they were adjacent to the contact points of the digits, but not if
they were at other positions (Haffenden & Goodale, 2000). The predicted effect of the spatial
position of the flankers on the peak grip aperture did not reach significance. However, in a
later study Haffenden et al. (200 1) showed that the spatial position of the flankers in the
Ebbinghaus figure (distance between the target object and the flankers) can influence peak
grip aperture, providing some support for this hypothesis, which we will refer to as the
"illusory precision hypothesis". However, Franz et al (2003) suggested that the effects of the
Ebbinghaus illusion on grasping cannot be accounted for by non-perceptual effects (obstacle
avoidance). They found similar effects of the distance between the target and the flankers on
maximum grip aperture and perception.
33
Chapter 3
Although we have previously argued that perception and action use the same visual
information (Smeets et al, 2002), we have also argued that visual estimates of size are not
used in grasping (Smeets & Brenner, 1999). If both arguments are true, then the effect of the
flankers on PGA cannot only be due to a misjudgement of size. We have shown that such
effects can be explained in terms of changes in judgements of the required precision (Smeets
et al., 2003)
Table 3.1: Magnitude of the effect of the Ebbinghaus illusion on perceptual judgements and on the peak grip aperture (PGA).
study #targets
Aglioti et al. (1995) 2
Haffenden & Goodale (1998)§ 2
Pavani et al. (1999)
Franz et al. (2000)
Haffenden et al. (2001)
Hanisch et al. (2001)
Glover & Dixon (2002)
Franz (2003)
Franz et al. (2003)
2
Kwok & Braddick (2003) 2
perception PGA
2.5 (0.2) 1.6 (0.4)
4.2 (1.0)- 1.0 (0.5)
0.7 1.0
1.5 (0.1) 1.5 (0.4)
2.6 (0.4)-+ 0.2 (0.3)
1.5 (0.5)@ 0.8 (0.6)@
2.1 (0.4)- 1.4A (0.3)
1.6- A /1.1 A 1.8A
1.4 (0.2)N 1.5 (0.3)N
1.3 (0.1)F 1.2 (0.3)F
2.o- 1.0
All illusion effects are the differences between values for small and large flankers expressed in mm (with standard errors when given). #targets: number of targets visible simultaneously. +: only the conditions "adjusted small" and "traditional large" are included ": scaled illusion effect -:manual size estimation (participants indicated target size by opening index finger and thumb) rather than comparison of two central circles or adjustment of an isolated circle. @:only the adult group is included. §:the values from this study are obtained from table 1 of Franz (2001 ). NIF: data for the conditions "nea~' and "far" respectively. Data provided by Franz.
In order to try to distinguish between the two above-mentioned proposals, we performed an
experiment with a version of the Ebbinghaus illusion that was designed to separate illusory
size effects from illusory precision effects. We varied an aspect that was expected to be
irrelevant for the (mis)perceived size, but that could influence judgements of the required
precision: the positions of the flankers with respect to the movement (figure 3.1). To be able
34

Effects of the Ebbinghaus figure on grasping
to vary the spatial position of the flankers (potential obstacles) by a considerable amount
without changing their distance to the object, we used only four circular flankers. If the
illusory effect of the Ebbinghaus figure is caused by the flankers changing estimates of the
required precision, aspects of grasping such as grip orientation are likely to depend on the
spatial position of the flankers. If the illusory effect is caused by illusory size information, the
PGA should only depend on the spatial position of the flankers if the perceived size does, and
no other aspects of grasping should depend on the flankers' position. Since Franz et al. (2003)
showed that the perceptual judgement can change with the spatial position of the flankers, we
first tested whether the spatial position of four circular flankers surrounding the target disk
has an effect on perceptual size settings. After that, subjects were asked to grasp the target
disks. We examined the movement time (MT), the peak velocity (PV), the time to peak
velocity (TPV), the orientation of the grip at two instances, the PGA, the time to PGA and the
final grip aperture (FGA).
Materials and methods
Subjects
This study is part of an ongoing research program that has been approved by the local ethics
committee. Twelve right-handed colleagues volunteered to take part in the study after being
informed about what they would be required to do.
Apparatus and stimulus
The stimulus consisted of a single black central circle surrounded by four black (large or
small) circular flankers, all on a white background. The stimulus was projected from below
onto a projection surface. The resolution of the projected image was 1024 by 768 pixels, with
1 pixel corresponding with about 0.4 mm. The large and small flankers were 55 mm and 17
mm in diameter, respectively. The diameter of the central circle could be 28, 30 or 32 mm.
The distance from the edge of the central target disk to the nearest edge of the flanker was
12.5 mm, irrespective of flanker size. For each flanker size there were two possible
configurations of the Ebbinghaus figure. The only difference between them was that the
spatial position of the flankers was changed: the array of flankers could be in two
configurations differing by 45° in orientation (figure 3.1).
35

Chapter 3
small flankers
large flankers
• ••• •
• ••• • Configuration 1
• • •• •
• • •• • Configuration 2
Figure 3.1: The four combinations of flanker size and configuration used in the experiment. The only
difference between the two configurations is that the flankers have been rotated by 45 degrees.
In the perceptual task, an isolated comparison circle was displayed after the stimulus. The
position of the comparison circle was chosen at random within an area of about 16 mm
laterally and 24 mm in the sagittal direction. The centre of this area was 15 em to the right of
the centre of the original stimulus. The size of the comparison circle was initially set to a
random value between 10 and 50 mm.
In the grasping task a real black target disk was placed on top of the projected central target
circle. Its diameter was exactly the same as that of the projected central target circle. In order
to make the grasping task comparable with the perceptual task, the height of the target disk
was only 3 mm, which was hardly noticeable for the near-orthogonal viewing. However due
to the difference in material, subjects could clearly see that this was a real object. Because of
this difference in material of the stimulus in the perceptual setting and the grasping stimuli,
the magnitude of the illusion might differ slightly. We will therefore not directly compare the
magnitude of the influence of the illusion on grasping with that on perception. The projected
circular flankers were exactly the same as in the perceptual task. A black starting position
(diameter 2 mm) was projected 30 em to the right of the central circle. The set-up of the
grasping task is shown in figure 3.2.
36

Effects of the Ebbinghaus figure on grasping
:.· •
Figure 3.2: Top view of the experimental set-up. Subjects have their thumb and index fmger pinched
together at the starting position (30 em from the centre of the target disk) and their eyes closed. After
a verbal signal subjects open their eyes and grasp the thin central target disk.
Procedure
In each trial of the perceptual task a stimulus was presented for 1.5 seconds. Immediately
after it disappeared, the isolated comparison circle was presented. The subject's task was to
adjust the size of the comparison circle. They did so by moving the mouse horizontally
(changing the position of the mouse changed the size of the comparison circle). Subjects
adjusted its size until it was perceived to be the same size as the former central target circle.
When the subjects were satisfied with their setting they pressed the mouse button and after 1
second a new stimulus appeared. During the task subjects had unrestricted vision. There was
no time limit for the perceptual settings. Each subject performed 120 trials (3 sizes of the
target disk x 2 sizes of flankers x 2 configurations x 10 repetitions) in a completely random
order.
In the grasping task trajectories of finger movements were recorded using an Optotrak
System (sampling rate 250 Hz, resolution 0.01 mm). Infrared emitting diodes (IREDs) were
attached to the nail of the thumb and the index finger of the right hand. Before each trial the
starting position was presented. Subjects pinched their thumb and index fmger together at this
starting position and closed their eyes. The stimulus was then presented and the experimenter
put the target disk exactly on the projection of the central circle. The subject was instructed to
open his/her eyes, grasp the target disk at a comfortable pace, and place the disk at the right
side of the surface. The experimenter removed the target disk, and the starting position for the
37

Chapter3
next trial was presented. Each subject performed the same 120 trials as in the perceptual task
in a new random order.
Data analysis
Instantaneous velocity was computed from position samples of the IREDs. To do so we fit a
second order polynomial to 7 position samples (24 ms window) centred at each position.
Based on the parameters of the fit we estimated the finger's velocity at that instant.
Movement onset is defmed for each digit as the last frame before peak velocity in which the
tangential velocity was smaller than that on the preceding frame. Movement offset was
defined as the first frame after peak velocity in which the velocity component in the direction
perpendicular to the surface changed sign (i.e. when the digit started moving upwards). The
movement time (MT) was defmed as the time between the movement onset of the digit that
started moving first and the movement offset of the digit that stopped last This procedure
ensures that we include the whole movement. To characterize the velocity profile, we
determined the relative time to peak velocity (TPV), which is the fraction of the movement
time at which the peak velocity occurred.
Peak grip aperture (PGA) was defmed as the maximum distance between the thumb and the
index fmger IREDs. Smeets et al. (2003) predict that if a larger PGA is due to changed
accuracy constraints, it should occur earlier. We therefore determined the relative time to
peak grip aperture (TPGA) as the fraction of the MT at which the PGA occurred. Final grip
aperture (FGA) is the distance between the thumb and the index fmger IREDs at movement
offset. Grip orientation is defined as the orientation of the projection on the plane of the
Ebbinghaus figure of a straight line connecting the positions of the IREDs on the finger and
thumb (0° is to the right). This angle was determined both at PGA and at movement offset.
A scaled illusion effect was calculated for the perceived size and for the PGA (following
Franz et al., 2001). For each subject the effect of the size of the flankers was divided by the
slope relating the measures in question to the physical size, so that the influence of the
illusion could be expressed as an equivalent change in actual size.
An important prediction of the illusory size hypothesis is that subjects who are perceptually
very susceptible to the illusion should also have a large effect of the flankers on PGA. Such
correlations have been used to show that perception and action are based on the same visual
information (Lopez-Moliner et al., 2003). To test this, we calculated the correlation between
38

Effects of the Ebbinghaus figure on grasping
the magnitudes of the effect of the illusion on the perceptual judgements and on the PGA
across subjects.
Statistical tests were all conducted across subjects. In both tasks data were analysed with
repeated-measures ANOVAs with the factors target size (3 levels: 28, 30, 32 mm), size of the
flankers (2 levels: large, small) and spatial position of the flankers (2 levels: configuration 1
and 2). Values are presented as the mean± standard errors between subjects. A significance
level of u=0.05 was used for all statistical analyses. In the perceptual task the dependent
variable was the set size of the comparison circle. In the grasping task the dependent
variables were: PGA, FGA, grip orientation at PGA and at movement offset, movement time
(MT), peak velocity (PV), relative time to peak grip aperture (TPGA) and relative time to
peak velocity (TPV).
Results
In the perceptual task the set size of the comparison circle was influenced both by the real
target size (slope: 0.67 ± 0.11) and by the size of the flankers (figure 3.3). The unsealed
illusion effect (difference between a target surrounded by small flankers and one by large
flankers) is 1.8 ± 0.3 mm. The scaled illusion effect shows that surrounding a target disk by
large flankers rather than by small flankers has the same influence as making it 2. 7 mm
smaller (p<O.Ol). No difference in the set size was found between the two spatial positions of
the flankers (configuration 1 vs. configuration 2) and there were no significant interactions.
perceptual judgement of diameter of central disk (mm)
ns
31 p<0.01 JX(l.01 ,-------, ,-------,
30 f ! 29
28 ? ~ 27
26 ... ~ •• ·\· ·i·
Figure 3.3: Results for the perceptual judgement task averaged over the three target sizes. Error bars represent standard errors between subjects.
39
Chapter 3
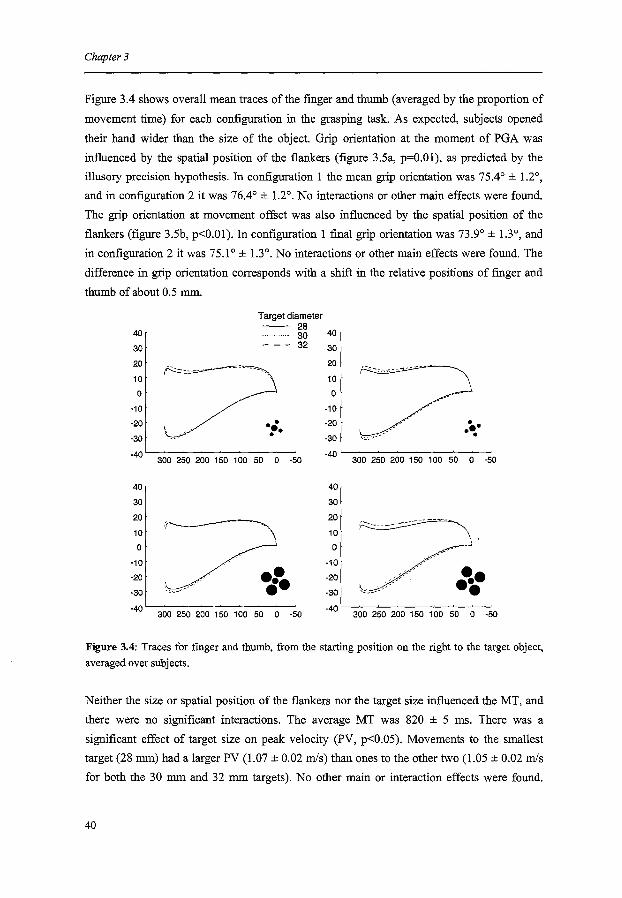
Figure 3.4 shows overall mean traces of the fmger and thumb (averaged by the proportion of
movement time) for each configuration in the grasping task. As expected, subjects opened
their hand wider than the size of the object. Grip orientation at the moment of PGA was
influenced by the spatial position of the flankers (figure 3.5a, p=0.01), as predicted by the
illusory precision hypothesis. In configuration 1 the mean grip orientation was 75.4° ± 1.2°,
and in configuration 2 it was 76.4° ± 1.2°. No interactions or other main effects were found.
The grip orientation at movement offset was also influenced by the spatial position of the
flankers (figure 3.5b, p<0.01). In configuration 1 fmal grip orientation was 73.9° ± 1.3°, and
in configuration 2 it was 75.1 o ± 1.3°. No interactions or other main effects were found. The
difference in grip orientation corresponds with a shift in the relative positions of finger and
thumb of about 0.5 mm.
Target diameter
40 28 30 40
30 32 30
20 20
10 10
0 0
·10 -10
·20 •• -20
·30 ~· -30
-40 300 250 200 150 1 00 50 0 ·50
-40 300 250 200 150 1 00 50 0 ·50
40 40
30 30
20 20
10 10
0 0
·10 ·10
·20 •••• -20 :-: ·30 -30
·40 300 250 200 150 1 00 50 0 ·50
-40 300 250 200 150 1 00 50 0 -50
Figure 3.4: Traces for finger and thumb, from the starting position on the right to the target object,
averaged over subjects.
Neither the size or spatial position of the flankers nor the target size influenced the MT, and
there were no significant interactions. The average MT was 820 ± 5 ms. There was a
significant effect of target size on peak velocity (PV, p<0.05). Movements to the smallest
target (28 mm) had a larger PV (1.07 ± 0.02 m/s) than ones to the other two (1.05 ± 0.02 m/s
for both the 30 mm and 32 mm targets). No other main or interaction effects were found.
40
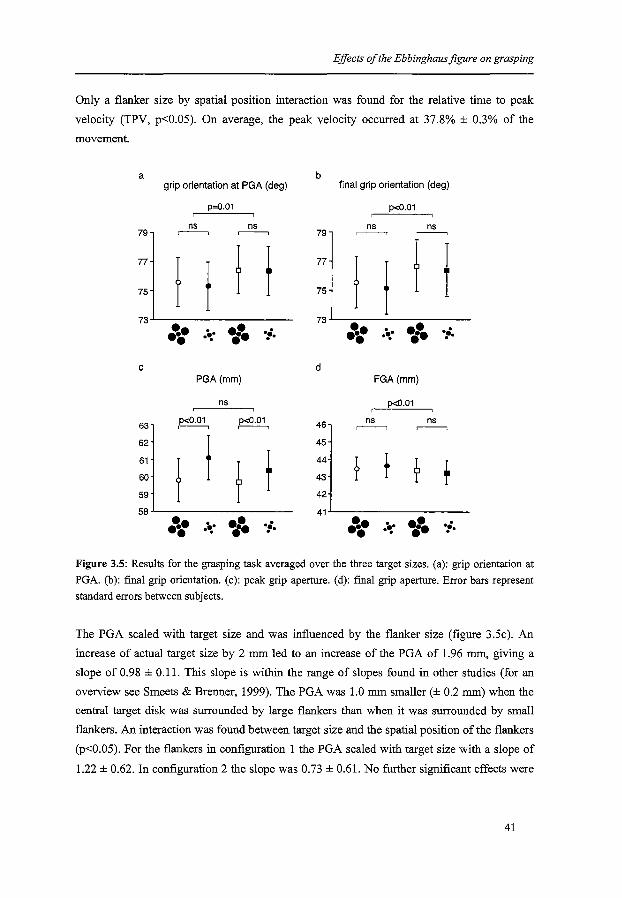
Effects of the Ebbinghaus figure on grasping
Only a flanker size by spatial position interaction was found for the relative time to peak
velocity (TPV, p<0.05). On average, the peak velocity occurred at 37.8% ± 0.3% of the
movement.
a b grip orientation at PGA (deg) final grip orientation (deg)
p=0.01 p<0.01
ns ns ns ns 79 79 ,------,
77
I I I I 77
I l l l 75 75
73 73
:-: '\ :-: ·\· '\ ·i· ·\· ·i·
c d PGA(mm) FGA(mm)
ns p<0.01
63 p<0.01 p<0.01 46 ns ns ,------, ,------, ,------, ,------,
62 ! 45
61
I ! I 44 f t ? f 60 43
59 42
58 41
:-: . ,. '\ ·i· :.· • ·\· ,. ·; .
Figure 3.5: Results for the grasping task averaged over the three target sizes. (a): grip orientation at
PGA. (b): final grip orientation. (c): peak grip aperture. (d): final grip aperture. Error bars represent
standard errors between subjects.
The PGA scaled with target size and was influenced by the flanker size (figure 3.5c). An
increase of actual target size by 2 mm led to an increase of the PGA of 1.96 mm, giving a
slope of 0.98 ± 0.11. This slope is within the range of slopes found in other studies (for an
overview see Smeets & Brenner, 1999). The PGA was 1.0 mm smaller(± 0.2 mm) when the
central target disk was surrounded by large flankers than when it was surrounded by small
flankers. An interaction was found between target size and the spatial position of the flankers
(p<0.05). For the flankers in configuration 1 the PGA scaled with target size with a slope of
1.22 ± 0.62. In configuration 2 the slope was 0.73 ± 0.61. No further significant effects were
41
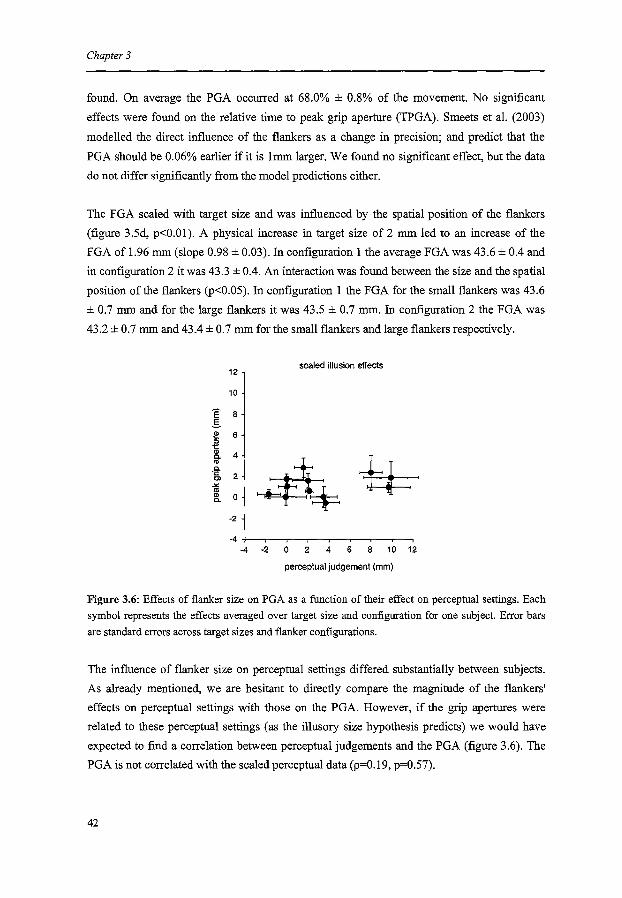
Chapter 3
found. On average the PGA occurred at 68.0% ± 0.8% of the movement. No significant
effects were found on the relative time to peak grip aperture (TPGA). Smeets et al. (2003)
modelled the direct influence of the flankers as a change in precision; and predict that the
PGA should be 0.06% earlier if it is lmm larger. We found no significant effect, but the data
do not differ significantly from the model predictions either.
The FGA scaled with target size and was influenced by the spatial position of the flankers
(figure 3.5d, p<O.Ol). A physical increase in target size of 2 mm led to an increase of the
FGA of 1.96 mm (slope 0.98 ± 0.03).1n configuration 1 the average FGA was 43.6 ± 0.4 and
in configuration 2 it was 43.3 ± 0.4. An interaction was found between the size and the spatial
position of the flankers (p<0.05). In configuration 1 the FGA for the small flankers was 43.6
± 0.7 mm and for the large flankers it was 43.5 ± 0.7 mm. In configuration 2 the FGA was
43.2 ± 0.7 mm and 43.4 ± 0.7 mm for the small flankers and large flankers respectively.
12 scaled illusion effects
10
-2
-4 +-~--,--,---r--r--r--~-, -4 -2 0 2 4 6 8 10 12
perceptual judgement (mm)
Figure 3.6: Effects of flanker size on PGA as a function of their effect on perceptual settings. Each
symbol represents the effects averaged over target size and configuration for one subject. Error bars
are standard errors across target sizes and flanker configurations.
The influence of flanker size on perceptual settings differed substantially between subjects.
As already mentioned, we are hesitant to directly compare the magnitude of the flankers'
effects on perceptual settings with those on the PGA. However, if the grip apertures were
related to these perceptual settings (as the illusory size hypothesis predicts) we would have
expected to find a correlation between perceptual judgements and the PGA (figure 3.6). The
PGA is not correlated with the scaled perceptual data (p=0.19, p=0.57).
42

Effects of the Ebbinghaus figure on grasping
Discussion
The difference between the perceptual judgements for a disk surrounded by large and small
flankers in this study is equivalent to a change in size of 2. 7 mm. Thus four flankers are
enough to obtain the Ebbinghaus illusion. Unlike a study by Franz et al. (2003), we found no
differences in perceptual judgements between the spatial positions of the flankers. Changing
the distance between the flankers and the target (Franz et al., 2003) affects perception in a
different way than changing the spatial position of the flankers with a rotation of the flanker
array.
The grip orientation at PGA and at movement offset were both influenced by the spatial
position of the flankers. Thus the flankers change the position at which the disk is grasped,
which is in favour of the illusory precision hypothesis. The spatial position of the flankers
also changed the slope of the relationship between PGA and object size. Again, this is in line
with the flankers directly interfering with the movements of the digits.
The PGA was influenced by the size of the flankers (1 mm difference between a target disk
surrounded by large flankers and one surrounded by small flankers). The magnitude of this
effect is within the range of effects that have previously been found in studies using the
classical Ebbinghaus configuration (table 3.1).
a b
Figure 3.7: Picking up a target with the fingernails parallel to the surface (a) can produce a different
marker distance than picking it up with the fingers perpendicular to the surface (b).
43

Chapter3
The effect of flanker orientation on the FGA means that subjects pick up the target object
with a different orientation of the digits. For example, subjects could pick up the target object
with the nails more perpendicular to the surface or with the nails more parallel to the object's
surface (figure 3.7). The FGA, defined as the absolute distance between the IREDs, will
differ when grasping in the manners shown in figures 3.7a and 3.7b, without changing the
actual distance between the contact points of the digits.
How exactly the subjects actually did pick up the targets cannot be determined using one
IRED on each digit. In the studies of Franz et al. (e.g. 2000, 2001) grip aperture was
measured with three IREDs on each digit. With this method, digit orientation can be
calculated and even the location of a point on the skin of the digits. However, the measured
grip size will still depend on how the object is grasped, because the object will not always be
grasped with the part of the digit for which the location is determined. Thus, although the
flankers' effects on grip orientation and FGA make us less confident about our measure of
PGA, this does not change the fact that the flankers influence the grasp in a manner that is
unrelated to perceived size.
Our study shows that conclusions should not be drawn on the basis ofPGA alone. In previous
studies conclusions were drawn about whether or not there is a dissociation between
information for perception and for action based on the presence or absence of an illusory
effect on the PGA (Haffenden & Goodale, 1998; Kwok & Braddick, 2003; Aglioti et al.,
1995). We show that other grasping parameters are also influenced, so the illusion cannot
only be influencing the movement through its influence on the perceived size. We therefore
conclude that the effect of the flankers on PGA need not only be due to the misjudgement of
the central disk's size, so that a direct comparison of the magnitudes of the effects on
perception and action is probably meaningless.
44

Chapter 4
The coding of pointing movements
Published as: De Grave, D.D.J., Brenner, E., & Smeets, J.B.J. (2004) Illusions as a tool to
study the coding of pointing movements. Experimental Brain Research, 155, 56-62.

Chapter4
Abstract
Pictorial illusions bias our judgements about certain visual attributes. Such illusions are
therefore only expected to influence a task if these attributes are used to perform the task.
When pointing to a position, different visual attributes could be used to guide the hand:
direction and distance (or length) of the required displacement (vector coding) or the final
position (position coding). In this study we used the Brentano illusion (an illusion of length)
to determine which attributes are used in pointing. Several conditions were tested in which
the visibility of the hand and the stimulus were varied. The illusion influenced movements
between two points along the shaft of the figure, but not movements perpendicular to the
shaft. When the hand and/or target were invisible during the movement, the influence of the
illusion increased. Pointing movements under different visual conditions were based on
different relative contributions of position and vector coding. The contribution of vector
coding was always rather modest.
46

The coding of pointing movements
Introduction
In Euclidean space, length is the distance between two positions. However, there is ample
evidence that people process visual information about these spatial attributes independently
(Gillam & Chambers, 1985; Gillam, 1998; Mack et al., 1985; Post & Welch, 1996; Smeets et
al., 2002). The Brentano illusion (figure 4.1), like other variants of the Muller-Lyer illusion,
primarily influences judgements of length. Thus, a task will only be influenced by this
illusion if the task requires a visual estimate of length.
This reasoning can also be reversed. The illusion can be used to determine which attributes
are used in pointing: the egocentric position of the endpoint (position coding: Bizzi et al.,
1992; Carrazzo et al., 1999; Feldman & Levin, 1995; Mcintyre et al., 1997; Mcintyre et al.,
1998; Van den Dobbelsteen et al., 2001) or the distance and direction ofthe target relative to
the starting position (vector coding: Bock & Eckmiller, 1986; Desmurget et al., 1998;
Messier & Kalaska, 1997; Rossetti et al., 1995; Vindras & Viviani, 1998). If an illusion that
influences the perceived length of a line also influences the endpoints of movements along
that line, we can conclude that the distance and direction of the required movement are used
to help determine the endpoint. The magnitude of this influence (in particular under open
loop conditions) can help us to determine the relative roles of vector coding and of position
coding in controlling our movements.
a b
• •
Figure 4.1: The Brentano illusion with the wings-in configuration at the top (a) and with the wingsout configuration at the top (b). Each subject performed one block oftrials for each configuration. The dots indicate the positions at which a target could appear (on one of the arrowheads or outside the illusion). Only one configuration was visible at a time.
47

Chapter4
In the present study we use the Brentano illusion to determine the extent to which the
distance and direction, rather than the egocentric position of the endpoint, contribute to the
control of pointing movements. We know that the arrowheads of the Brentano illusion
influence the judged length of the shaft. We assume that they have no effect on perceived
positions (Post & Welch, 1996) and directions (i.e. perpendicular alignment: Gillam &
Chambers, 1985). According to the vector coding hypothesis, when subjects point along the
shaft from one arrowhead to another (figure 4.1) they will use the distorted information about
the length of the shaft connecting the two arrowheads to guide their pointing movement. If
the movement is made perpendicular to the shaft from a position outside the illusion, the
shaft's (distorted) length is irrelevant. Thus such pointing movements will not be influenced
by the illusion. If the movement is made in any other direction we expect intermediate values.
In order to avoid making assumptions about how distances and directions outside the figure
are influenced by the arrowheads, we will concentrate on pointing movements that are either
along the shaft or perpendicular to the shaft of the Brentano figure. According to the position
coding hypothesis there will be no influence of the illusion at all.
A factor that can influence the kind of information that is used, and with that whether the
illusion will influence the task, is visibility of the hand and the stimulus. If subjects can see
both the target and their hand they will almost certainly detect initial errors and correct them
during the movement (Abrams et al., 1990; Prablanc and Martin, 1992). If the hand is not
visible during the movement, subjects may be less certain about the position of the hand, and
therefore find it harder to control its position. In such cases they are more likely to use length
information to control the distance to be covered by their pointing movements. Similarly, if
the end position is not visible they are more likely to consider the previously seen length as
the distance that they should move. If the information of both the hand and the target is
removed, the likelihood to use length information will even be larger. Thus the influence of
the illusion is expected to increase as feedback is removed.
Materials and methods
Subjects
This study is part of an ongoing research program that has been approved by the local ethics
committee. Thirteen right-handed colleagues volunteered to take part in the study after being
informed about what they would be required to do. All had normal or corrected-to-normal
vision.
48

The coding of pointing movements
Apparatus and stimulus
The stimulus consisted of a black Brentano figure and a red target dot on a white background
(figure 4.1 ). The two vertical shafts each had a length of 8 em. The length of the wings was
2.2 em. The inclination of the wings with respect to the shafts was 30°. Two configurations of
the Brentano illusion were used: one with the wings-in on top and one with the wings-out on
top. The target dot (diameter 0.2 em) could appear either on one of the three arrowheads of
the Brentano illusion (top, middle or bottom) or outside the figure. The target outside the
figure was presented 8 em to the right of the middle arrowhead. In each trial the figure and
only one target dot were presented.
a
0 I I
I I I I
I I
b
~ projection surface
mirror
box
graphics tablet graphics tablet
Figure 4.2: A schematic view of the apparatus. (a) In the hand visible conditions, the stimulus was projected directly on the graphics tablet. (b) In the hand invisible conditions subjects looked at the projected image via a mirror. They held the pen on the graphics tablet underneath the mirror. The task was always to bring the tip of the pen to the red target dot.
The stimulus was projected directly on a graphics tablet (W ACOM A2), or on the projection
surface of a box that was placed on the graphics tablet (figure 4.2). In the latter case subjects
could see the stimulus at the level of the tablet through a mirror without seeing their hand.
The resolution of the projected image was 1024 by 768 pixels, with 1 pixel corresponding
with about 0.5 mm. Pointing positions were registered as the positions at which the tip of the
49

Chapter4
pen stopped moving on the graphics tablet. This was defined as the position at the first
sample on which the tip of the pen had moved less than 1 mm during the preceding 300 ms.
Positions were sampled at 200 Hz with an accuracy of about 0.25 mm.
Procedure
All subjects performed four conditions of a pointing task in random order in separate
sessions. These conditions consisted of all possible combinations of the hand and target being
visible or invisible during the movement (four possible combinations). Each condition
contained two blocks of 400 trials, one for each configuration of the Brentano figure. The
order of the blocks was counterbalanced across subjects. The subjects were asked to bring the
tip of a pen to a red target dot, that was either on one of the three arrowheads or at a position
outside the illusion (figure 4.1). The endpoint of one movement was the starting position of
the pointing movement to the next target.
In the two stimulus visible conditions, the stimulus was continuously visible at the same
location. When the subject stopped moving, the red target dot jumped to a new position. In
the two stimulus invisible conditions, the stimulus was only visible before subjects started
their pointing movement. As soon as subjects moved the pen, the stimulus disappeared from
view. When the subject finished the pointing movement the stimulus appeared again with the
target dot at a new position.
In the two hand visible conditions, the image was projected onto the tablet, so that subjects
saw their hand all the time. If the target disappeared once the subject started to move, the
computer aligned the arrowhead to which the subject had just moved with the tip of the pen
when the stimulus re-appeared, so that it seemed to the subjects as if they never made a
mistake. If subjects drifted outside the range of the tablet the stimulus reappeared at the
middle of the tablet, and that trial was discarded. In the two conditions in which the hand was
invisible, the stimulus was projected on a back-projection screen at the top of a box (figure
4.2). The box was placed on the graphics tablet. Subjects looked at the projected image via a
mirror. They held the pen underneath the mirror. In this case there was no need to re-align the
stimulus.
After completing the four conditions of the pointing task subjects performed a length
matching task, in which they had to draw an invisible line of the same length as either the
wings-in part or the wings-out part of the Brentano illusion. The line had to be drawn
50

The coding of pointing movements
smoothly in one vertical stroke next to the illusion. The same stimulus as in the pointing task
was projected directly on the graphics tablet. Instead of one red target dot, two red dots were
presented. One red dot was always presented at the middle arrowhead and the other either at
the arrowhead at the top or at the bottom of the figure. Subjects had to draw a straight line of
the same length as the line between the target dots. They had to start at a red dot that was
presented 10 centimetres to the right of the figure. In order to make sure that subjects could
not align their drawing with elements of the figure this dot was not aligned with either
arrowhead, but 2.5- 3.7 em below the top or above the bottom arrowhead. If the upper part of
the figure had to be matched, the starting dot was near the top arrowhead. Otherwise it was
near the bottom arrowhead. The drawing direction was always toward the middle arrowhead
(from the top to the middle or from the bottom to the middle). When subjects started to move
the pen the figure and the starting dot disappeared from view, but the hand remained visible.
The trajectory of the drawing was recorded on the graphics tablet. This condition also
contained two blocks of 400 trials, one block for each configuration of the Brentano figure.
After having analyzed the data we asked subjects to perform two control pointing
experiments, one with movement time instructions and the other with "direction of approach"
instructions. The control experiment in which the movement time instruction (slow or fast)
was varied was performed in the condition in which the hand was visible and the stimulus
invisible. In the slow trials the movement time had to be between 900 and 1300 ms. In the
fast trials it had to be between 450 and 650 ms. Trials were now blocked by instructions for
the duration of the pointing movement. The configuration of the Brentano figure was
randomized across trials (this was possible because the stimulus always disappeared).
Auditory feedback was given on every trial that was not within the appropriate range of
movement times. Trials with movement times outside the range were deleted from data
analysis. Half the subjects performed the slow pointing movements before the fast ones.
The control experiment in which the "direction of approach" instruction was varied was
performed in both conditions in which the stimulus was invisible during the movement (with
and without visibility of the hand). The procedures in these conditions were identical to the
corresponding original pointing conditions except that the target dot was always on one of the
arrowheads. This means that movements were only made between the arrowheads. A beep
sounded in 50% of the trials in which the target appeared at the central arrowhead. In those
trials, the subjects' task was to draw a strongly curved path to the presented target. Otherwise
a straight path had to be drawn. A path was considered strongly curved when there was a
deviation of 8-12 em to the right of the shaft. A straight path had to be within a range of 2 em
51

Chapter4
to the left or right of the shaft. Auditory feedback was given for every path that was not
drawn in accordance with the demands of the trial. These trials were excluded from data
analysis.
Data analysis
The quantitative comparison of the influence of the illusion was based exclusively on the
three types of movements toward the target dot on the middle arrowhead (upward, downward
or leftward). In general subjects tend to misjudge the distance that is to be moved when
viewing of the hand and/or stimulus is prevented. Estimates ofthese distances differ between
subjects and between different spatial positions. Therefore a measure of the magnitude of the
illusion was computed for each subject, movement type and condition, which is independent
of the amplitude of the pointing movement and that always compares the influence of the
illusion at the same (average) spatial position.
For each type of movement we calculated the median vertical distance between the endpoints
of pointing movements for the two configurations (wings-in on top, wings-out on top)1. The
difference in distance between configurations was divided by the median vertical distance
between the pointing position for the top and bottom target to correct for general tendencies
to overestimate or underestimate the extent (in particular when no feedback of the hand was
provided). The result is the size of the illusion expressed as a percentage of the length of a
shaft of the Brentano figure.
Statistical tests were all conducted across subjects. For each pointing condition a repeated
measures ANOV A was performed on the illusion magnitudes to check whether the three
types of pointing movements were influenced differently by the illusion. One-sample t-tests
were performed to check whether the illusion magnitude in each of the three types of
movements differed from zero (to check whether there is any effect of the illusion) and paired
t-tests were used to determine which types of pointing movements differed. In the length
matching task one sample t-tests were conducted to check whether there was an effect of the
illusion for each direction of drawing (from the top to the middle and from the bottom to the
middle); a paired t-test was conducted to check whether the illusion differed between the
directions of drawing.
1 When the image was realigned in the hand visible and target invisible condition, the coordinate system for determining the positions was redefmed so that positions are always in relation to the figure on the screen.
52

The coding of pointing movements
Whether subjects used length information to the same extent in the four pointing conditions
and in the length matching task was tested with another repeated measures ANOV A. Since
the magnitude of the illusion for the upward and downward movements were not expected to
differ, these magnitudes were averaged for this analysis. Pointing movements from outside
perpendicular to the shaft were not expected to be influenced by the illusion at all and are
therefore not considered in the comparison between conditions.
For the movement time condition similar tests were conducted as for the original conditions.
The movements between the arrowheads were combined to compare the fast and slow
movements in a paired t-test. In the "direction of approach" conditions a factorial ANOVA
was performed to check for main effects (visibility condition and direction of approach) and
for an interaction between the two factors.
Results
Figure 4.3 shows the lengths of the pointing movements for individual subjects, movement
types (top to middle, bottom to middle), figure orientations (wings out at top or bottom) and
conditions (visibility of the hand and stimulus, length matching). The figure shows that there
are large deviations from the veridical distance (8 centimetres), especially when vision of the
hand is blocked. These deviations are not necessarily related to the illusion. The effect of the
illusion is evident from the fact that most points are below the unity line. The following
figures present the data in a way that is insensitive to systematically over- or underestimating
the distance (as described in the statistical analysis section).
The illusion influenced the fmal position of pointing movements that were along the shaft
(between the arrowheads) in all four conditions (figure 4.4). The position was shifted by less
than 1% of the length of the shaft in the full feedback condition, about 4% in the conditions
with either feedback of the hand or of the target, and about 8% in the condition with no
feedback. Movements perpendicular to the shaft showed no significant effect of the illusion,
except in the condition where the hand and the stimulus were both visible. In that case the
effect was extremely small, about 1/10 of the diameter of the red target. As the shift of the
endpoints of movements perpendicular to the shaft in the other conditions did not differ
significantly from this value, we regard this as our best estimate of the illusory effect on
perceived position. The shift of endpoints of movements up and down the shaft was always
larger than that of pointing movements from outside, perpendicular to the shaft (figure 4.4),
and was equal for the two directions. These results confirm our assumption that the illusion
influences the judged length of the line to a much larger extent than its endpoints.
53
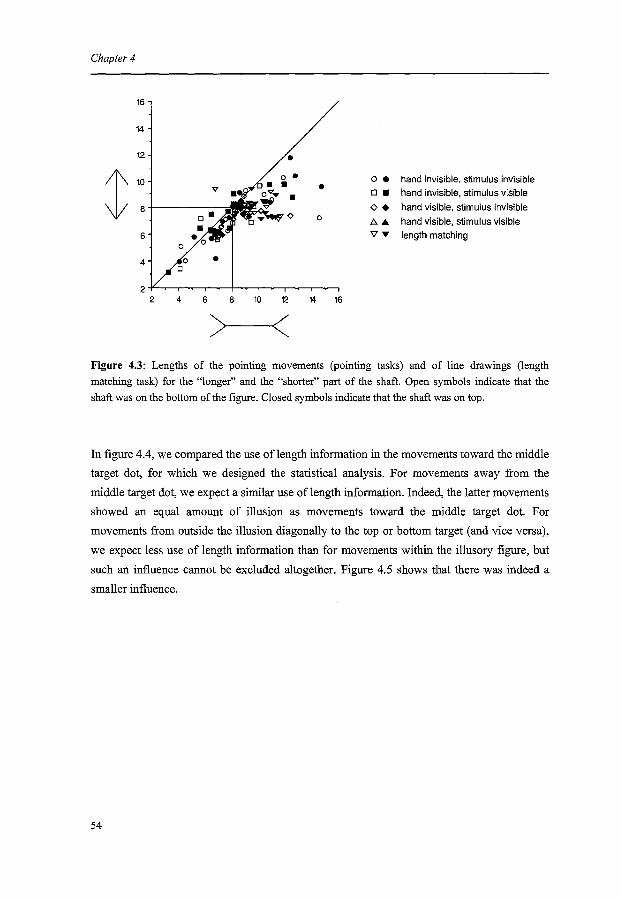
Chapter4
16
14
12
t 10
8
0. hand invisible, stimulus invisible • hand invisible, stimulus visible D. <>+ hand visible, stimulus invisible
0 Alt. hand visible, stimulus visible
6 VY length matching 0
4 •
14 16
>--< Figure 4.3: Lengths of the pointing movements (pointing tasks) and of line drawings (length
matching task) for the "longer" and the "shorter" part of the shaft. Open symbols indicate that the
shaft was on the bottom of the figure. Closed symbols indicate that the shaft was on top.
In figure 4.4, we compared the use oflength information in the movements toward the middle
target dot, for which we designed the statistical analysis. For movements away from the
middle target dot, we expect a similar use oflength information. Indeed, the latter movements
showed an equal amount of illusion as movements toward the middle target dot. For
movements from outside the illusion diagonally to the top or bottom target (and vice versa),
we expect less use of length information than for movements within the illusory figure, but
such an influence cannot be excluded altogether. Figure 4.5 shows that there was indeed a
smaller influence.
54
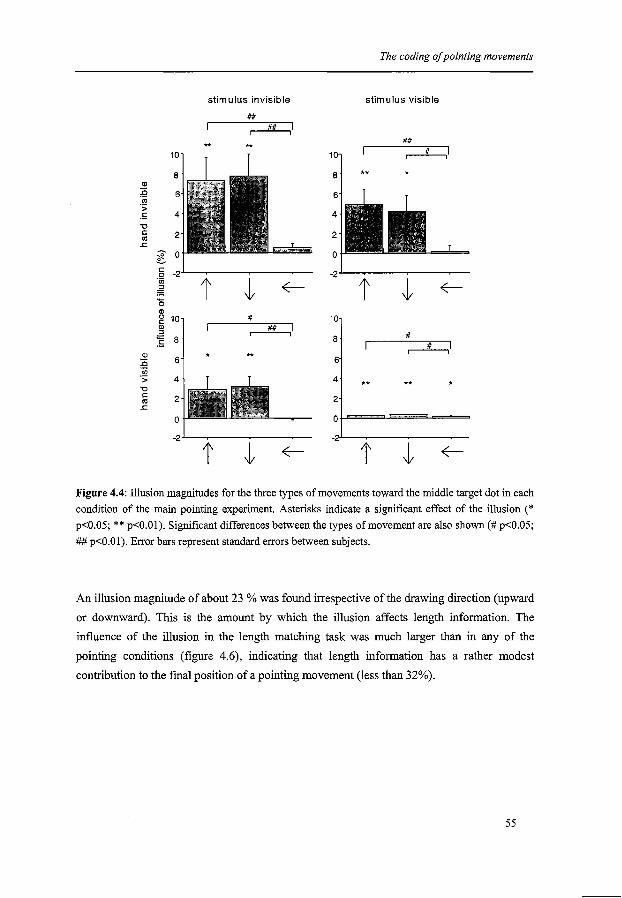
The coding of pointing movements
stimulus invisible stimulus visible
II# ##
II#
~ :-9 (I)
·;; .!: "0 c: (1j ~
~ c: 0 ·u;
t ~ ~ t ~ ~ :§ 0 Q) (.) 10 # 10 c: Q) ##
.=! 8 #
£ 8 #
~ 6 .c ·;;; ·;; 4 4 "0 c: 2 2 (1j
~
0 0
1' ~ ~ 1' ~ ~
Figure 4.4: Illusion magnitudes for the three types of movements toward the middle target dot in each condition of the main pointing experiment. Asterisks indicate a significant effect of the illusion (*
p<0.05; ** p<O.Ol). Significant differences between the types of movement are also shown(# p<0.05;
## p<O.O 1 ). Error bars represent standard errors between subjects.
An illusion magnitude of about 23 % was found irrespective of the drawing direction (upward
or downward). This is the amount by which the illusion affects length information. The
influence of the illusion in the length matching task was much larger than in any of the
pointing conditions (figure 4.6), indicating that length information has a rather modest
contribution to the fmal position of a pointing movement (less than 32% ).
55
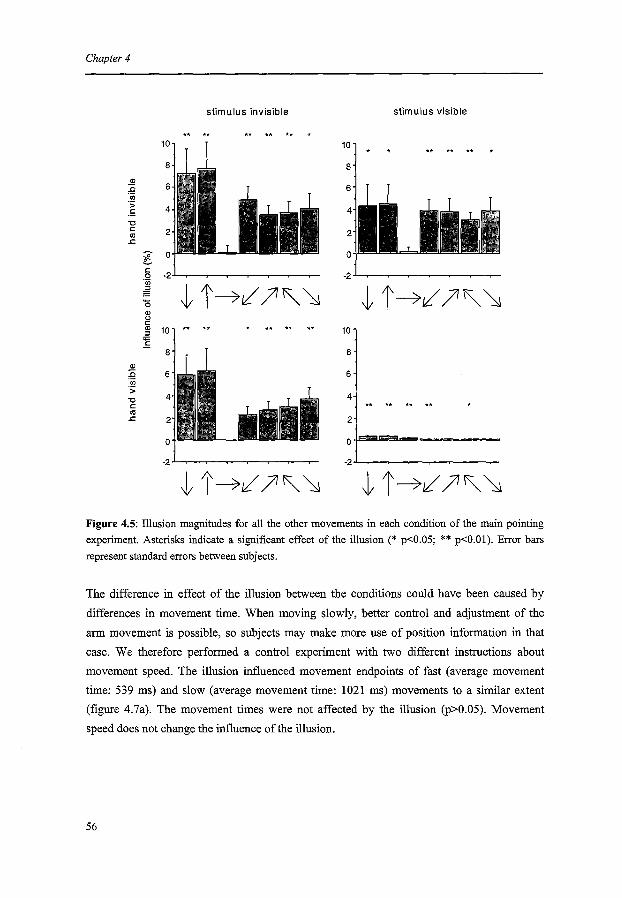
Chapter4
stimulus invisible stimulus visible
** 10
8
~ .0
:~ >
·"' ""C c: ro
.<::
~ c: 0 ·c;;
~ j-7t/' ?i'\ ~ g ~ j-7t/' ?i'\ ~ 0 (!) 0 c: "' 10 ::l
E 8
"' ;e 6 CJl ·;;:
4 ""C c: ro
.<:: 2
0
-2
~ j-7t/' /If\~ ~ j-7t/' ?i'\ ~
Figure 4.5: Illusion magnitudes for all the other movements in each condition of the main pointing
experiment. Asterisks indicate a significant effect of the illusion (* p<O.OS; ** p<O.Ol). Error bars
represent standard errors between subjects.
The difference in effect of the illusion between the conditions could have been caused by
differences in movement time. When moving slowly, better control and adjustment of the
arm movement is possible, so subjects may make more use of position information in that
case. We therefore performed a control experiment with two different instructions about
movement speed. The illusion influenced movement endpoints of fast (average movement
time: 539 ms) and slow (average movement time: 1021 ms) movements to a similar extent
(figure 4.7a). The movement times were not affected by the illusion (p>0.05). Movement
speed does not change the influence of the illusion.
56
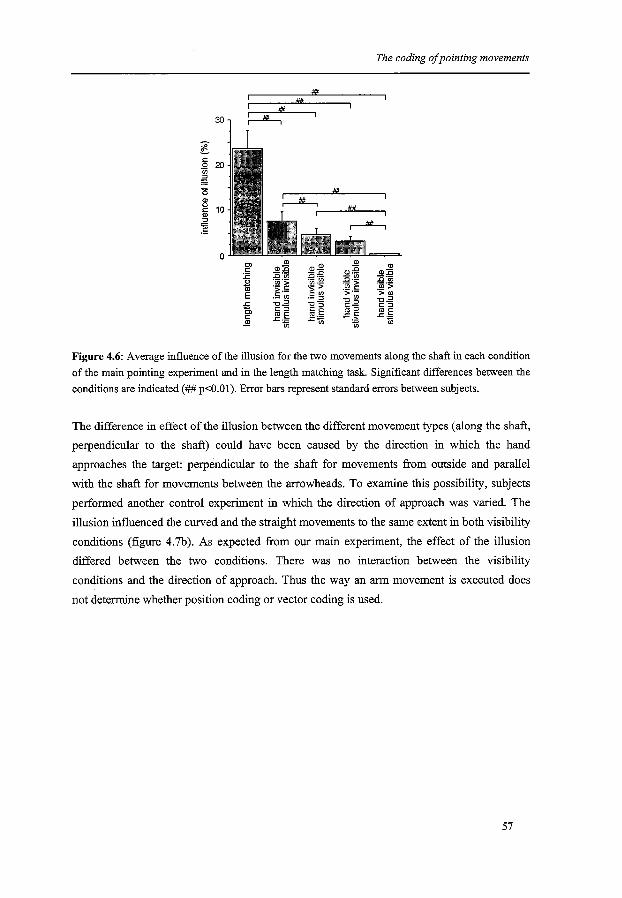
## 30 ~
~ <:: 0 20 ·u; § 0 2l <:: 10 Q) ::I
£
0
The coding of pointing movements
1111
Figure 4.6: Average influence of the illusion for the two movements along the shaft in each condition of the main pointing experiment and in the length matching task. Significant differences between the conditions are indicated(## p<O.Ol). Error bars represent standard errors between subjects.
The difference in effect of the illusion between the different movement types (along the shaft,
perpendicular to the shaft) could have been caused by the direction in which the hand
approaches the target: perpendicular to the shaft for movements from outside and parallel
with the shaft for movements between the arrowheads. To examine this possibility, subjects
performed another control experiment in which the direction of approach was varied. The
illusion influenced the curved and the straight movements to the same extent in both visibility
conditions (figure 4.7b). As expected from our main experiment, the effect of the illusion
differed between the two conditions. There was no interaction between the visibility
conditions and the direction of approach. Thus the way an arm movement is executed does
not determine whether position coding or vector coding is used.
57
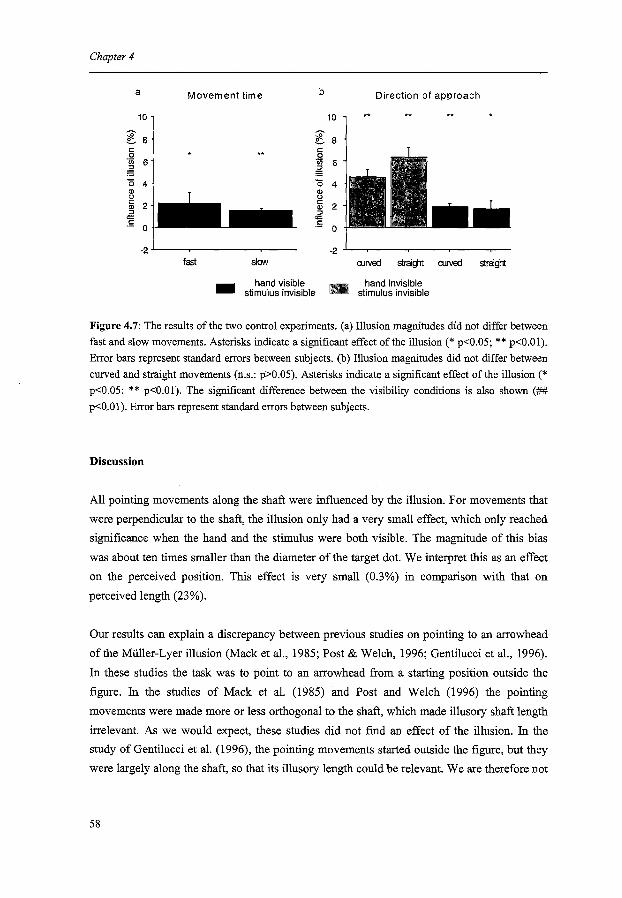
Chapter4
a
10
la <::: 0
·c;; 6 § 15 4 (I)
" iii 2 :::J
~ 0
Movement time b
fast - siCMI
10
~ 8 <::: 0
·c;; 6 § 15 4 (I)
" iii 2 :::J
E 0
hand visible stimulus invisible
Direction of approach
curved straight curved straight
hand invisible stimulus invisible
Figure 4.7: The results of the two control experiments. (a) Illusion magnitudes did not differ between
fast and slow movements. Asterisks indicate a significant effect of the illusion(* p<0.05; ** p<O.Ol).
Error bars represent standard errors between subjects. (b) Illusion magnitudes did not differ between
curved and straight movements (n.s.: p>0.05). Asterisks indicate a significant effect of the illusion(*
p<0.05; ** p<O.Ol). The significant difference between the visibility conditions is also shown (##
p<O.Ol). Error bars represent standard errors between subjects.
Discussion
All pointing movements along the shaft were influenced by the illusion. For movements that
were perpendicular to the shaft, the illusion only had a very small effect, which only reached
significance when the hand and the stimulus were both visible. The magnitude of this bias
was about ten times smaller than the diameter of the target dot. We interpret this as an effect
on the perceived position. This effect is very small (0.3%) in comparison with that on
perceived length (23%).
Our results can explain a discrepancy between previous studies on pointing to an arrowhead
of the Muller-Lyer illusion (Mack et al., 1985; Post & Welch, 1996; Gentilucci et al., 1996).
In these studies the task was to point to an arrowhead from a starting position outside the
figure. In the studies of Mack et al. (1985) and Post and Welch (1996) the pointing
movements were made more or less orthogonal to the shaft, which made illusory shaft length
irrelevant. As we would expect, these studies did not fmd an effect of the illusion. In the
study of Gentilucci et al. (1996), the pointing movements started outside the figure, but they
were largely along the shaft, so that its illusory length could be relevant. We are therefore not
58

The coding of pointing movements
surprised that in the latter study the illusion did affect pointing. Gentilucci et al. (1996)
ascribed the difference in results to the orientation of the illusion: horizontal in the former
two studies and vertical in the latter. Our results suggest that it is the relationship between the
starting point of the movement and the orientation of the figure that determines whether the
illusion has an effect.
It is well-established that the task that is used to evaluate the magnitude of an illusion can
influence the result. In an earlier study (de Grave et al., 2002) we showed that even for a
single perceptual task the influence of an illusion (the Roelofs effect) could depend on
seemingly irrelevant factors. In this study we show that the starting position of the hand
influences where one points when asked to indicate the vertices of an illusory figure (the
Brentano illusion). Thus the influence of illusions probably tells us more about the kind of
information that is used, than about what it is used for.
The planning and control model of Glover and Dixon (Glover & Dixon, 2001; 2002) states
that the planning of the movement is influenced by the illusion, but errors in planning are
corrected during on-line control of the movement. This is consistent with our finding that the
influence of the illusion is largest when there is least possibility to correct the movement.
However, this model predicts an influence of the movement time. If subjects make fast
pointing movements there is little time to make on-line corrections and therefore the
influence of the illusion should be larger. The model also predicts that the movement time
depends on the perceptual length of the shaft. If a movement is planned in advance for a
distance that seems shorter than it actually is (movement along the "short" shaft), the end
position will not have been reached by the time the planned movement time has passed. To
reach the end point, a longer movement must be made. Thus the movement time for the shaft
that looks shorter should be longer than that for the shaft that looks longer.
To test these hypotheses we performed a control pointing experiment in which movement
time instructions (slow or fast) were varied. We found no difference in the effect of the
illusion between fast and slow movements. We also found no differences in movement time
between the shaft that looks long and the shaft that looks short. Thus this model alone cannot
explain our data.
The results cannot be interpreted as either pure position coding or pure vector coding. For
pure position coding one would have to conclude that the perceived egocentric endpoint
depends on the starting point of the hand (within or outside the figure). For pure vector
59

Chapter4
coding, one would have to conclude that on-line control reduces the influence of initial errors
caused by relying on judged length. However, as mentioned above, if on-line control reduces
an initial error, the fast movements in our experiment should have shown a much larger effect
of the illusion than the slow movements, which they did not. Thus the endpoints of pointing
movements show effects of both position coding and vector coding.
In the length matching task subjects directly reported the perceived length. Therefore we
assume that the magnitude of the illusion found in this task (about 23%) is the maximum that
could be expected in the pointing tasks. In all pointing conditions the magnitude of the
illusion was much smaller (S 7.8%), which means that subjects never only used length
information. They did not even do so when no visual feedback was provided during the
movement. We conclude that length information probably normally plays a very minor role
in determining the endpoint of a movement.
60

Chapter 5
The coding of saccades
Submitted as: De Grave, D.D.J., Smeets, J.B.J. & Brenner, E. The coding of saccades studied
with the Brentano illusion.

Chapter 5
Abstract
Movements to a visual target could either be guided by visual judgements of direction and
distance (or length) of the required displacement (vector coding), or by estimates of the final
position (position coding), or both. In an earlier study, we showed that the Brentano illusion
(an illusion of length) only influences pointing movements if the illusory length is relevant
for the pointing task, demonstrating that vector coding is involved. The effect was small,
leading to the conclusion that the contribution of vector coding was rather modest. In this
study we used the Brentano illusion to determine whether length is used in a similar manner
to guide saccadic eye movements. Subjects made saccades between the vertices of the
Brentano illusion and between these points and a point outside the illusion. The illusion
influenced saccades between two points along the shaft of the figure, but not movements
perpendicular to the shaft. As may be expected, the eyes (saccades) rely more strongly on
vector coding than the hand (pointing).
62

The coding of saccades
Introduction
Several researchers have tried to answer the question whether saccades are susceptible to
illusions, such as the Milller-Lyer illusion. Wong and Mack (1981) shortly presented subjects
a target dot surrounded by a frame. After a second presentation of the stimulus, in which
displacement of the frame induced illusory displacement of the target. Subjects were asked to
make a saccade to the last presented target location. The saccades were not influenced by the
illusory change in target position. In contrast, in a study by Binsted and Elliott (1999a)
subjects had to make saccades between the vertices of the Muller-Lyer illusion in time with a
metronome. There was an influence of the illusion on the amplitude of the (first) saccades, as
in Festinger et al. (1968) and Yarbus (1967). So there is no simple answer to the question.
We have previously suggested that such results can be understood in terms of inconsistently
processed spatial attributes (Smeets et al., 2002). Physically related spatial attributes are not
necessarily perceived in a consistent manner, presumably because they are processed
independently. In the study of Wong and Mack (1981) the illusion is one of motion. The
perceived position might have remained unaffected, just as the perceived distance from the
present fixation position. Therefore it is not surprising that no effect of the illusion was
found.
Similarly, the Milller-Lyer illusion affects the perception of the length of the central part of
the figure (the shaft), but not that of the positions of its endpoints (Gillam & Chambers, 1985;
Gillam, 1998; Mack et al., 1985; Smeets et al., 2002). Saccadic eye movements to the
endpoints will only be influenced if the incorrectly perceived attribute, in this case length, is
used to perform that task. In the studies ofBinsted and Elliott (1999a), Festinger et al. (1968)
and Yarbus (1967) subjects made eye movements along the shaft of the MUller-Lyer illusion.
We propose that in that case subjects use the illusory length information to judge the required
amplitude of the saccade, and thus an effect of the illusion is found.
Various theories have been proposed for the relation between the Muller-Lyer illusion and
eye movements. The "centre of gravity" theory states that the illusion is caused by saccades
being directed to the centre of a stimulus configuration (Findlay, 1982; He & Kowler, 1989).
When making a saccade towards a wings-in vertex the movement is too short because the
saccade is made towards the centre of the arrowhead rather than to the tip. Saccades towards
a wings-out vertex are too long. Gregory (1968) proposed that the shaft with the wings-out
vertices can be seen as a two-dimensional projection of an inside corner so that the shaft is
63

Chapter 5
perceived as being the most distant part of the figure. The shaft with the wings-in vertices
represents the projection of an outside comer, which is judged to be the nearest part of the
figure. The difference in relative distance judgements gives rise to the illusion. We will refer
to this theory as the perspective projection theory. If this theory is correct the eyes should
diverge more when making saccades towards the shaft with the wings-out vertices of the
Mtiller-Lyer illusion, which is perceived as being further away, than when making saccades
towards the shaft with the wings-in vertices, which is perceived to be closer. This theory does
not predict an effect of the illusion on saccade amplitude, because the visual angle of a line
remains the same irrespective of the perceived distance. The illusion only arises when this
visual angle is converted into a length by considering the distance.
We recently demonstrated that whether an illusion influences a pointing movement depends
on whether the illusion affects the information that is used for that movement (De Grave et
a!., 2004). An illusion oflength, such as the Brentano illusion, will influence movements that
are based on the distance and direction of the target relative to the starting position (vector
coding), but not ones based on the egocentric position of the endpoint (position coding).
Moreover, such an illusion will only influence pointing movements along the line of illusory
length. We indeed found that the illusion influenced pointing movements between two points
along the shaft of the figure, but not movements perpendicular to the shaft. The effect was
much smaller than the perceptual effect, indicating that vector coding played only a minor
role in guiding pointing movements.
In the present experiment the Brentano illusion (figure 5.1) was used to identify the visual
information that guides saccades. If saccades and pointing movements use a common motor
command (Bekkering eta!., 1995; Bizzi eta!., 1971; Biguer eta!., 1982, 1984; Bock, 1986;
Reina & Schwartz, 2003), we can expect saccadic eye movements to rely on a combination of
vector coding and position coding. However, studies on the control of saccades generally
suggest that saccades are entirely vector coded (Robinson, 1972; Becker & JUrgens, 1979;
Mcilwain, 1991). This means that the eye and the hand use different information, which has
also been demonstrated in an experiment of Binsted and Elliott (1999b) using the Miiller
Lyer illusion. If we assume that saccades are purely vector coded, we would predict that the
end point of the first saccade will be more strongly affected by the illusion than the endpoint
of a pointing movement.
64

The coding of saccades
Materials and methods
Subjects
This study is part of an ongoing research program that has been approved by the local ethics
committee. Twelve subjects volunteered to take part in the study after being informed about
what they would be required to do. All had normal or corrected-to-normal vision.
Apparatus and stimulus
A bite-board was placed in front of a computer screen (38.4 x 28.8 em, 1024 x 768 pixels, 75
Hz) to keep the subject's head fixed at a viewing distance of 85 em. At this distance, 1 pixel
corresponds to 0.025°. The stimulus consisted of a vertical black Brentano figure and a red
target dot on a white background (figure 5.1). The length of each of the two vertical shafts
was 5.4°. The length of the wings was 1.SO. The inclination of the wings with respect to the
shafts was 30°. Two configurations of the Brentano illusion were used: one with the wings-in
on top and one with the wings-out on top. The target dot (diameter 0.13°) could appear either
on one of the three vertices of the Brentano illusion (top, middle or bottom) or outside the
figure. The target location outside the figure was 5.4° to the right of the middle vertex. In
each trial the figure and only one target dot were presented. Eye movements were registered
with an Eyelink eye tracker (SR Research Ltd.) with a temporal resolution of 250Hz and a
spatial resolution of 0.2°. The target jumped to a new position as soon as the subject kept his
or her eyes within a radius of 20 pixels around the target dot for 1 00 ms, after the eyes had
moved between 80 and 120% of the required distance from the start position of the saccade.
Procedure
All subjects performed two blocks of 400 trials, one for each configuration of the Brentano
figure. The order of the blocks was counterbalanced across subjects. The subjects were asked
to look at the red target dot, that was either on one of the three vertices or at a position
outside the illusion (figure 5.1). The Brentano figure was continuously visible at the same
location. When the red target dot jumped to a new position, subjects made a saccade to that
position. The target of each saccade was the starting position of the saccade to the next target.
After completing the eye movements task, subjects performed two perceptual judgement
tasks. The stimuli were similar to those used in the eye movements task. In the "relative
65

ChapterS
judgement" task they had compare the upper and the lower shaft and judge whether the upper
or the lower shaft appeared longer. The red dot remained stationary on the middle vertex and
remained visible during the entire experiment. Subjects had to fixate this dot while the
Brentano configuration was presented for 250 ms. They then had to press one of the "arrow"
buttons to indicate whether they thought that the upper shaft or the lower shaft was longer.
After 500 ms a new stimulus appeared. Two staircases of 250 trials, one for each
configuration, were randomly interleaved. If subjects pressed the up arrow button, the length
of the upper shaft in the next trial was decreased by 1% of its length (0.05°) and the length of
the lower shaft was increased by the same amount. Thus the overall length of the
configuration remained the same. The middle vertex was at the same position in each trial. If
subjects pressed the down button the converse occurred.
a b
• •
Figure 5.1: The Brentano illusion with the wings-in configuration at the top (a) and with the wings
out configuration at the top (b). Each subject performed one block of trials for each configuration. The
dots indicate the positions at which a target could appear (on one of the vertices or outside the
illusion). Only one configuration was visible at a time.
Only ten of the twelve subjects performed the "absolute judgement" task. A Brentano figure
was presented with a red dot on the middle vertex. In this task both shafts were always 5.4°.
After one second, a second red dot appeared either on the upper or lower vertex to indicate
which shaft length had to be judged. After 500 ms the stimulus with the two red dots
disappeared and a test line was presented for 500 ms. Two-third of the length of the test line
overlapped with the former position of the indicated shaft. Subjects had to press one of the
arrow buttons to indicate whether they thought the test line was longer or shorter than the
shaft between the two red dots. If subjects pressed the up arrow button, the length of the test
line on the next trial for that shaft and configuration was decreased by 1% (0.05°). If subjects
pressed the down button the length of the test line was increased by 1%. After 500 ms a new
66

The coding of saccades
stimulus appeared. Four staircases of 50 trials, one for each shaft in a configuration, were
randomly interleaved.
Data analysis
For the eye movement task only the first saccades after the target jumps were analysed. Trials
on which gaze shifted prior to the jump of the target or within 50 ms after target jump were
excluded from analysis. Trials on which the first saccade after target jump was not in the
required direction were also excluded from analysis. This resulted in a loss of 8% of all trials.
The quantitative evaluation of the influence of the illusion was based on the three types of
saccades toward the target dot on the middle vertex (upward, downward or leftward). For
each type of saccade we calculated the vertical distance between the median endpoints of
saccades for the two configurations (wings-in on top, wings-out on top). This difference was
divided by the vertical distance between the median endpoints of saccades to the top and
bottom target, to correct for general tendencies to overshoot or undershoot the extent. The
result is the size of the illusion expressed as a percentage of the length of the shaft of the
Brentano figure.
Statistical tests were all conducted across subjects. A repeated measures ANOVA was
performed on the size of the illusion, to examine whether the three different directions of eye
movements towards the middle vertex were influenced differently by the illusion. Paired t
tests were used to determine which eye movements differed. One-sample t-tests were
performed to check whether the illusion magnitude in each of the three types of movements
differed from zero (to check whether there was any effect of the illusion).
To test the perspective projection theory, we computed the difference in vergence between
the start and the end of the first saccade for each type of saccade. Vergence at the start of the
saccade was defined as the average vergence for 50 ms immediately preceding saccade onset.
Vergence at the end of a saccade was defined as the average vergence for 50 ms following
the end of a saccade. For each type of saccade we determined the difference in vergence
differences between the two configurations. A repeated measures ANOV A was performed on
these differences in vergence, to examine whether the different types of saccades showed
differences in vergence.
67

Chapter5
In the relative perceptual judgement task, we calculated how much longer the wings in part of
the illusion needed to be for it to appear to be as long as the wings out part (as a percentage of
shaft length). For each subject we determined the average percentage of how much the
stimulus with respect to the middle vertex was shifted during the last 150 trials. We did so for
each configuration, and divided the mean of the two averages by two to get an estimate of the
average influence of the illusion on a single shaft.
In the absolute perceptual judgement task, we also calculated the average difference in
perceived length between the wings in and the wings out part (expressed as a percentage of
the length of the shaft) for each subject. To do so we determined the difference between the
test line lengths during the last 80 trials of each configuration for the two configurations of
the figure. We did so for both the upper shafts and for the lower shafts, and divided the mean
of these two averages by two to get an estimate of the average influence on a single shaft.
The result is the size of the illusion expressed as a percentage of the length of the shaft of the
Brentano figure.
Results
The two panels in figure 5.2 show raw eye traces of the same subject with saccades to 1;he
middle target (five for each starting position) for the two configurations. For the wings in
configuration the subject's saccades undershoot the target For the wings out configuration
they overshoot the target Corrective saccades are made after about 200 ms. When a saccade
is made from outside the figure it ends at the target position (thin traces).
Figure 5.3 shows the size of the effect of the illusion on saccades toward the middle target
dot. The ANOV A showed a significant difference between the types of saccades (F(2)=3.95,
p<0.05). Saccades between the vertices (along the shaft) showed a significant effect of the
illusion (13% for saccades from the bottom to the middle target and 8% for ones from the top
to the middle target). No significant effect of the illusion (-1%) was found for saccades
perpendicular to the shaft. These results indicate that the saccades are based on an estimate of
distance, which in tum is based on the judged length of the shaft if the movement is along the
shaft.
As the figure was continuously visible during the experiment, subjects received feedback
about their performance, so one might expect a decrease in illusory effect during a block of
trials. The amplitude of saccades adapts to the fmal distance of a target if feedback specifies
that the saccade was incorrect (e.g. because the target jumps during the saccade; Frens & Van
68
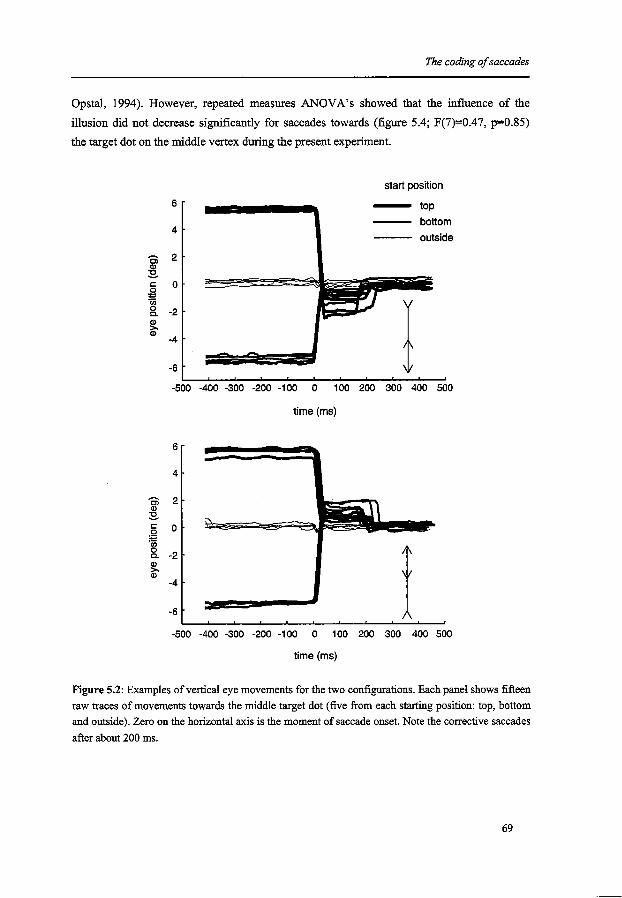
The coding of saccades
Opstal, 1994). However, repeated measures ANOVA's showed that the influence of the
illusion did not decrease significantly for saccades towards (figure 5.4; F(7)=0.47, p=0.85)
the target dot on the middle vertex during the present experiment.
Ci CD
:!:!. c 0 E Ol 0 c. ~ CD
Ci CD
:!:!. c 0
:;::::; ·u; 0 c. CD >-CD
6
4
2
0
-2
-4
-6
start position
top
bottom
outside
f -500 -400 -300 -200 -1 00 0 1 00 200 300 400 500
time (ms)
6
4
2
0
f -2
-4
-6
-500 -400 -300 -200 -100 0 100 200 300 400 500
time (ms)
Figure 5.2: Examples of vertical eye movements for the two configurations. Each panel shows fifteen
raw traces of movements towards the middle target dot (five from each starting position: top, bottom
and outside). Zero on the horizontal axis is the moment of saccade onset. Note the corrective saccades
after about 200 ms.
69
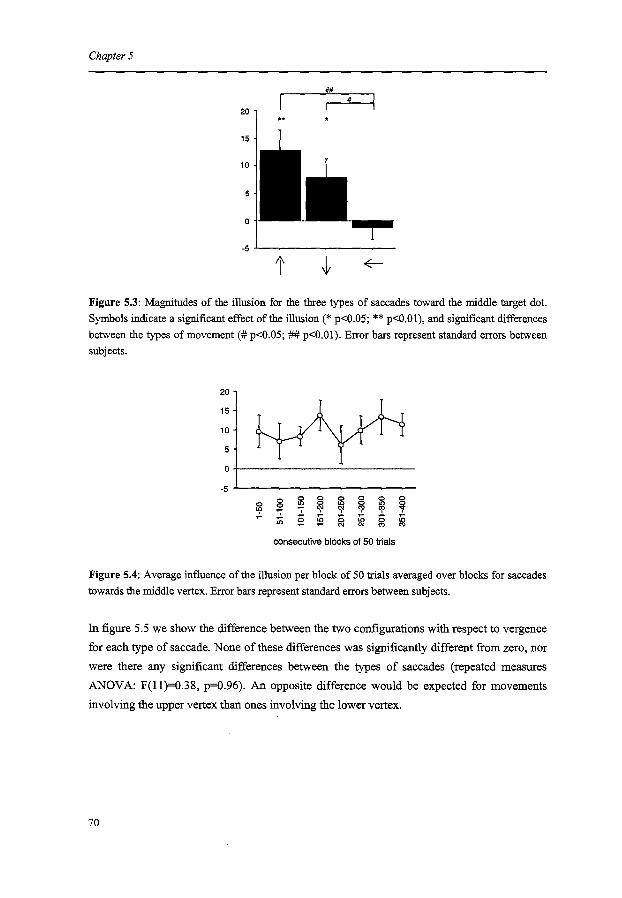
Chapter 5
## /!.
20
15
10
5
0
·5
t {, ~
Figure 5.3: Magnitudes of the illusion for the three types of saccades toward the middle target dot.
Symbols indicate a significant effect of the illusion(* p<O.OS; ** p<O.Ol), and significant differences
between the types of movement(# p<O.OS; ## p<O.Ol). Error bars represent standard errors between
subjects.
0 0 0 0 0 0 0 0 ':f 0 "' 0 "' 0 0 'l' 'l' 'l' 'l' -r ~ ~
0 ~
~ ~ 0 ;:;; ;:;; "' "' ~ "' "' "'
consecutive blocks of 50 trials
Figure 5.4: Average influence of the illusion per block of 50 trials averaged over blocks for saccades
towards the middle vertex. Error bars represent standard errors between subjects.
In figure 5.5 we show the difference between the two configurations with respect to vergence
for each type of saccade. None of these differences was significantly different from zero, nor
were there any significant differences between the types of saccades (repeated measures
ANOVA: F(ll)=0.38, p=0.96). An opposite difference would be expected for movements
involving the upper vertex than ones involving the lower vertex.
70
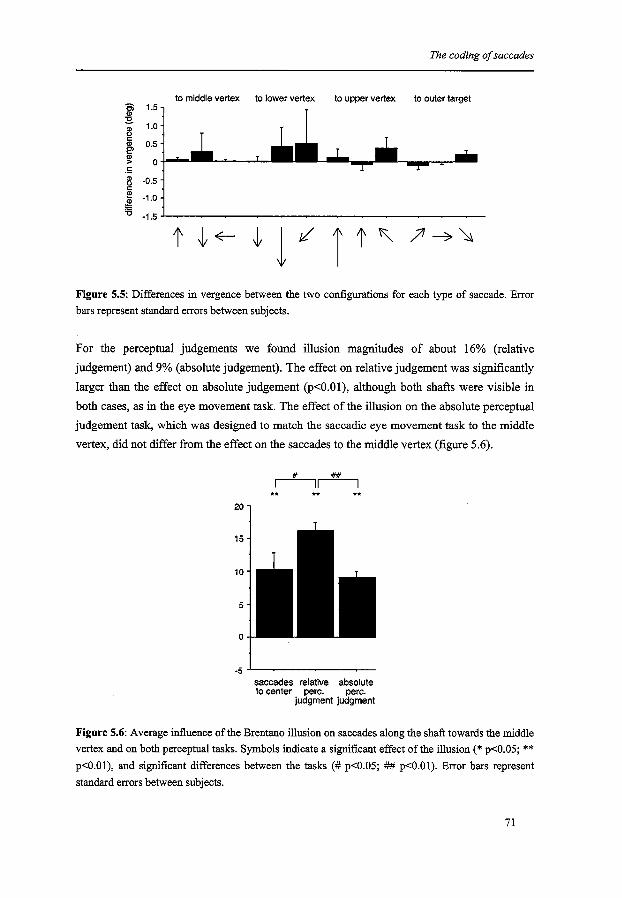
The coding of saccades
to middle vertex to lower vertex to upper vertex to outer target Oi 1.5
"' :)!!. 1.0
"' 0 c:
0.5 "' E' ~ 0 .5
"' -0.5 0 c: ~ -1.0 ~ '6 -1.5
t t~ t l t?' I t ~ 7'~~ Figure 5.5: Differences in vergence between the two configurations for each type of saccade. Error
bars represent standard errors between subjects.
For the perceptual judgements we found illusion magnitudes of about 16% (relative
judgement) and 9% (absolute judgement). The effect on relative judgement was significantly
larger than the effect on absolute judgement (p<O.Ol), although both shafts were visible in
both cases, as in the eye movement task. The effect of the illusion on the absolute perceptual
judgement task, which was designed to match the saccadic eye movement task to the middle
vertex, did not differ from the effect on the saccades to the middle vertex (figure 5.6).
saccades relative absolute to center perc. perc.
judgment judgment
Figure 5.6: Average influence of the Brentano illusion on saccades along the shaft towards the middle
vertex and on both perceptual tasks. Symbols indicate a significant effect of the illusion(* p<0.05; **
p<O.Ol), and significant differences between the tasks (# p<0.05; ## p<O.Ol). Error bars represent
standard errors between subjects.
71

ChapterS
Discussion
Saccadic eye movements along the shaft were influenced by the illusion. Saccades
perpendicular to the shaft were not. According to the "centre of gravity" theory (Findlay,
1982; He & Kow1er, 1989) the illusory effect is caused by saccades being directed to the
centre of the pattern near the vertex. This theory would predict the same error in saccadic end
position for saccades perpendicular to the shaft (the centre of gravity does not change with
starting position). This is inconsistent with our data, because the illusion had no effect on
saccades perpendicular to the shaft.
According to the perspective projection theory the illusion should not influence the amplitude
of the saccades, which it did. Moreover, the shaft with the wings out vertices (inside comer)
is presumably perceived to be further away than the shaft with the wings in vertices (outside
comer) so the eyes should diverge when making saccades from the isolated point to a vertex
of the wings-out shaft, and converge when making saccades from the isolated point to a
vertex of the wings-in shaft. Thus the perspective projection theory is also inconsistent our
data.
McCarley et al. (2003) found that voluntary saccades are influenced by the Miiller-Lyer
illusion to a similar extent as perceptual judgements, whereas reflexive saccades are only
modestly affected by the illusion. Our procedure was similar to the one that they used to elicit
reflexive saccades, and should therefore have given rise to small effects of the illusion. In
order to evaluate whether this was so, we can compare the influence on saccades with the
perceptual judgement tasks. The relative judgement task that we used was similar to the
classical experimental paradigm in perceptual judgement tasks (e.g. Aglioti et al., 1995;
Danckert et al., 2002; Haffenden & Goodale, 1998), in which two percepts are compared
directly. When we do that, a significant difference in illusion magnitude is found between
saccadic eye movements and perception. However, according to several authors (Franz et al.,
2000, 2001; Pavani et al., 1999; Vishton et al., 1999) this difference could be caused by the
fact that in the perceptual task subjects were influenced by two shafts being relevant
simultaneously (relative size) whereas only one shaft was relevant for the saccadic eye
movements (absolute size). Therefore the absolute perceptual judgement task was performed.
In this task subjects matched the perceived length of a single shaft to a neutral reference. In
this case the magnitude of the illusion is significantly smaller, indicating that the perceptual
illusion itself depends on the task details. We assume that the magnitude of the illusion found
in this task (about 9%) is what we would find for the saccadic eye movement task if it were
72

The coding of saccades
based on the judged distance (based on estimated shaft length) alone. The magnitude of the
illusion on saccades did not differ from the absolute judgement task, which means that
subjects mainly used length information to determine the saccade amplitude. However in
pointing movements length information had a rather modest contribution (less than 32%) to
the final position of a pointing movement (de Grave et al., 2004). This means that saccades
rely more strongly on vector coding than pointing.
In this study we showed that the starting position of the eye influences the end position of the
saccade. The influence of the starting position in relation to the Brentano illusion shows that
vector coding is used to determine the end positions of saccades. We can conclude that
saccades are mainly guided by the distance (vector coding).
73


Chapter 6
Summary and conclusions
The part "Can an apparent compression explain the difference between the original and
induced Roelofs effect?" is adapted from: De Grave, D.D.J., Brenner, E., & Smeets, J.B.J.
(2004). An apparent compression cannot explain the difference between the original and the
induced Roelofs effect. Vision Research, 44, I 031-l 032.
The part "Reconciling two visual systems with two theories of perception" is adapted from:
De Grave, D.D.J., Smeets, J.B.J., & Brenner, E. (2002). Ecological and constructivist
approaches and the influence of illusions. Behavioral and Brain Sciences, 25, 103-104.

Chapter 6
In tbis thesis, we investigated how visual illusions that distort our perception influence
visuomotor performance. There are several theories about how (illusory) visual information is
processed in the brain and how tbis information affects visuomotor performance. We have
tested some of these theories using visual illusions.
The original and induced Roelofs effect are not caused by the same shift in straight
ahead
The most influential theory regarding the influence of illusions on perception and behaviour
is undoubtedly the "two visual systems" hypothesis of Milner and Goodale, wbich assumes a
dissociation between perception and action. Support for tbis theory is found in experiments in
which perceptual tasks are influenced by visual illusions, whereas visuomotor actions are
performed accurately. Bridgeman and colleagues (Bridgeman et al., 1997, 2000; Bridgeman
& Huemer, 1998) investigated effects of the induced Roelofs effect on perception and action.
The induced Roelofs effect becomes evident when a target dot is presented within a
rectangular frame, that has an offset to one side of the subject's objective median plane. When
reporting the position of the dot a bias is found in the opposite direction of the offset of the
frame. In most cases, goal directed arm movements towards that target remained accurate
despite the perceptual bias. Bridgeman et al. 's interpretation was that the results are caused
by a dissociation between perception and action.
In chapter 2 we investigated whether it was possible to explain Bridgeman's findings without
assuming that there are two visual systems. Therefore we would like to know what causes the
induced Roelofs effect. To do so, we first turned to the original Roelofs effect. If a
rectangular frame is presented with an offset to the left of the subject's objective median
plane, the frame is perceived more to the right of the objective median plane than it really is.
Could the induced (as well as the original) Roelofs effect be caused by a sbift in perceived
straight ahead? Subjects performed two tasks with the induced Roelofs effect. In one task
subjects knew they always had to report the position of the target with respect to straight
ahead and in another task they did not know in advance whether to report the position of the
target or of the frame. These tasks resulted in two different magnitudes of the induced
Roelofs effect. We concluded that, based on just a difference in illusion magnitude between
perception and action, no conclusions should be drawn regarding two separate visual systems.
76

Summary and conclusions
Can an apparent compression explain the difference between the original and induced
Roelofs effect?
Dassonville et al. (2004) also tried to fmd out whether the two visual systems hypothesis or a
shift in straight ahead can explain the induced Roelofs effect. They also rejected the two
visual systems hypothesis but did fmd support for a shift in straight ahead. This raised the
question what causes the difference between our conclusion and theirs. Dassonville and Bala
(2004) argued that a training-related elimination of the Roelofs effect could explain our
finding that no induced Roelofs effect is found in the dual task. By providing a training
period before each task, subjects got feedback about their performance in localizing either a
single target or frame. In the practice trials of the dual task in which the position of the frame
had to be estimated, this feedback could have served to overcome the induced Roelofs effect.
Based on the corrective feedback the subject would adjust the perceptual scaling and the shift
in straight ahead disappears.
We did not conclude this from our data because in our dual task the eccentricity of the frame
is still underestimated Dassonville and Bala (2004) explained this by an apparent
compression of visual space. Errors in target localization caused by a shift in straight ahead
and those caused by an apparent compression can be measured independently (by varying
target and frame eccentricity, respectively). However, errors in frame localization caused by
these two factors cannot be measured separately, because both effects depend on the
eccentricity of the frame. The original Roelofs effect, that is still found in our dual task, could
be caused by an apparent compression of visual space rather than by a shift in perceived
straight ahead.
The apparent compression is a general phenomenon that applies to the target as well as the
frame, and should be present regardless of the temporal order of the frame and the target.
Such an apparent compression is indeed evident in the target gains of our target estimation
tasks (these gains do not equal 1). As our data showed clear signs of apparent compression,
we agree with Dassonville and Bala that this is a better framework to analyse our data. The
question is: can we reject the hypothesis that the induced Roelofs effect and the shift
component of the original Roelofs effect are both caused by the same shift in perceived
straight ahead? In other words, does our conclusion still hold if we take the apparent
compression into account?
77

Chapter 6
The effect of a combination of apparent compression and shift in straight ahead can be
formalised as follows: the estimated frame position (Pr ') depends on a shift in perceived
straight-ahead that is proportional to the frame position (b1*Pr) and a gain of the apparent
compression of positions (g*Pr). It is assumed that the target position does not influence the
estimated frame position (the mean gain of the influence of the target position on that of the
frame was -0.01; see table 2.1, 3rd column) The estimated target position (P1 ')is determined
by a shift in perceived straight-ahead (bz*Pr) and the same gain for the apparent compression
of positions (g*P1). Thus:
Pr' = (b1 + g)*Pr + O*Pt
Pt' = bz*Pr + g*Pt
(1)
(2)
In the training session of our dual task experiment (de Grave et al., 2002) accurate feedback
about the frame position was given. According to Dassonville and Bala (2004), this could
have eliminated the shift in perceived straight-ahead (b1 = b2 = 0), without eliminating the
apparent compression (g < 1). As the apparent compression is part of the original Roelofs
effect, and not of the induced Roelofs effect, this might qualitatively explain our results. To
test whether we can quantitatively explain our data with a single apparent compression g and
a single (possibly zero) shift in straight-ahead b1 = bz, we will calculate both values for the
shift in straight-ahead (b~, b2) from our data of the dual task experiment. Following the
reasoning of Dassonville and Bala (2004), there should be no difference between our three
orders of presentation. In order to have enough statistical power we will therefore average the
data over these three conditions.
The first calculation is based on equation (2) alone. As explained above, the frame gain for
the judgements of the target position (i.e. the induced Roelofs effect) is assumed to represent
the shift in straight-ahead: b2 = 0.059. For our estimate of b~, we subtract the apparent
compression g (obtained from the target gain for judgements of target position following
equation 2) from the frame gain of the frame judgements (the original Roelofs effect; (b1 +g)
in equation 1). This yields a different value for the shift in straight-ahead: b1 = -0.095. A
paired t-test showed that b1 f. b2 (p<O.Ol), so we can reject the hypothesis ofDassonville and
Bala (2004). Thus, also when taking into account the observed apparent compression, we can
reject the hypothesis that the original Roelofs effect and the induced Roelofs effect are based
on the same shift in straight-ahead.
78

Summary and conclusions
The Ebbinghaus figure changes more than perceived size
Several interpretations have been given for an effect of an illusion on grasping. Franz et al.
(2000) proposed in the common source model that an effect of the Ebbinghaus illusion on
grasping can be understood because the same illusory size infonnation is used in perception
and in grasping. Haffenden and colleagues (Haffenden & Goodale, 2000; Haffenden et al.,
200 1) stated that an effect of the illusion on grasping was not caused by the perceived illusory
size, since perception and action are dissociated. Rather, the circular flankers of the
Ebbinghaus illusion are treated as obstacles, which interfere with the grasping movement.
Smeets and colleagues have shown that an effect of the flankers on the PGA can be explained
in tenns of changes in judgements of the required precision (Smeets et al., 2003). We tried to
distinguish between those interpretations in chapter 3 by manipulating the spatial position of
the flankers around the central target in the Ebbinghaus illusion. Subjects were presented with
the Ebbinghaus illusion consisting of a central disk surrounded by four large or small
flankers. The array of flankers could be rotated by 45°. When grasping the central target disk,
the common source model predicts no effect of the spatial position of the flankers on
grasping, if perception is not affected by this spatial position. If the flankers somehow
interfere with the grasping movement, both other interpretations (Haffenden et al., 200 1;
Smeets et al., 2003) do expect to fmd an effect on grasping. The results show that the
perceptual size of the central target does not change with the spatial position of the flankers.
However, several grasping parameters were affected by the change in spatial position of the
flankers. Thus the effect of the flankers on prehension cannot only be due to a rnisjudgement
of the central disk's size as the common source model claimed.
In another experiment (Biegstraaten et al., submitted) we also found that non-illusory effects
are partly responsible for the influence of illusions on grasping. We let subjects grasp a bar
that was superimposed on the shaft of a Muller-Lyer figure. The configuration of the Milller
Lyer figure (ingoing or outgoing fins, see figure 1.2b) and the starting position of the hand
(below or besides the Milller-Lyer figure) affected the peak grip aperture, its timing and the
movement time. The configuration also affected the fmal grip aperture, although the
influence was very small. Thus also in this case the graphical elements influenced the
movement in other ways than by changing the perceived size alone, which makes it more
difficult to determine exactly what attributes are used in a motor task.
79

Chapter 6
The coding of pointing movements and saccades
According to the theory of Smeets and Brenner (1995a) visual illusions bias our judgements
about certain visual attributes and not of other {physically related) attributes. For example,
the Miiller-Lyer illusion changes the perceived length of a line, but not the perceived position
of the endpoints (figure 1.3). Visual illusions are therefore only expected to influence a task if
these affected attributes are used to perform the task. When pointing to a position, different
visual attributes could be used to guide the hand: direction and distance (or length) of the
required displacement (vector coding) or the fmal position {position coding). In chapter 4 we
determined what attributes are used in a pointing task. If a combination of vector coding and
position coding is used, the visibility of the hand and the stimulus can change the proportion
to which attributes are used, and with that the amount of illusory effect. Subjects were asked
to point (with a pencil) between the vertices of the Brentano illusion and a target dot outside
the illusory figure. The illusion influenced movements between two points along the shaft of
the figure, but not movements perpendicular to the shaft. When the hand and/or target were
invisible during the movement, the influence of the illusion increased. Pointing movements
under different visual conditions have different relative contributions of position and vector
coding. The contribution of vector coding was always rather modest.
In chapter 4 we found that pointing movements rely on a combination of vector coding and
position coding. But how are saccadic eye movements coded? One possibility is that saccades
and pointing movements use a common motor command. In that case a similar combination
of vector coding and position coding is expected for saccades as for pointing movements.
Alternatively, it has been suggested that saccades are entirely vector coded. This means that
the eye and the hand use different information and then a different influence of the illusion is
expected. In chapter 5 we used the Brentano illusion to determine whether saccadic eye
movements are coded in a similar manner as pointing movements. Subjects made saccades
between the vertices of the Brentano illusion and between these points and a point outside the
illusion (similar to the pointing task in chapter 4). The illusion influenced saccades along the
shafts, which means that length information (vector coding) is used. Unlike pointing
movements, saccades relied more strongly on vector coding.
However, it is still possible that hand and eye movements share a common motor command.
In our pointing task a tool (a pen) was used for pointing instead of a digit, which could
explain the result that pointing relies more on position coding. The use of this tool could have
made the hand use a different strategy to perform the movement than when pointing with a
80

Summary and conclusions
digit. To code the distance and direction of the required movement of the tip of the pen a
complicated vector must be determined since the tip of the pen can be at different positions
with regard to the body. When holding a pen, it may be easier to bring the tip of the pen to
the target by coding both positions relative to the body. Whether the use of a tool changes the
way a goal-directed movement is coded, should be tested .in further experiments.
(;eneralremarks
We have tested several theories regarding the processing of (illusory) visual information. The
highly influential two visual systems hypothesis of Milner and Goodale (1995) is not
supported in our studies. It has been a matter of debate for years whether or not there exists a
dissociation between perception and action that can be mapped onto the ventral and dorsal
stream, respectively. Some studies suggested that perceptual judgement and visuomotor
control are based on the same visual processes (Franz et al., 2000, 2001; Pavani et al., 1999;
Smeets & Brenner, 1995a; Vishton et al., 1999; Vishton & Fabre, 2003). Others have stated
that only some aspects of visuomotor performance are affected by illusions (Brenner &
Smeets, 1996; DeLucia et al., 2000; Jackson & Shaw, 2000; Smeets & Brenner, 1999; Smeets
et al., 2002; Van Donkelaar, 1999; Westwood et al., 2000c). Glover and Dixon (200la,
2001 b, 2001 c, 2002) argued that the effects of illusions are apparent at different phases of a
movement. Taken together, these studies indicate that the dissociation between perception
and action is not as strict as originally assumed in the two visual systems hypothesis.
Visuomotor performance is not always immune to illusions, while perceptual judgements are
not always susceptible to them.
There are several other pieces of evidence that support the link between perception and
action. For instance, studies on motion perception and smooth-pursuit eye movements have
demonstrated a tight link between the perceptual input and motor output. A lesion in area MT
of a macaque brain (or its human equivalent area V5) results in deficits in the perception of
motion and in the ability to produce accurate smooth-pursuit eye movements (e.g. Newsome
& Pare, 1988; Lekwuwa & Barnes, 1996). Moreover, Van Donkelaar et al. (2000)
demonstrated that smooth-pursuit adaptation is associated with changes in the perception of
motion. Thus, in this case, perception and action go hand in hand. Schwartz et al. (2004)
showed a link between motor areas and perception from the perspective of the ventral
premotor cortex (PMv). They found that neurons in PMv, known to be closely related to the
generation of arm movements, traced the perceptual path of the arm instead of the actual hand
movement.
81

Chapter 6
Refining the perception-action hypothesis
Although the body of evidence against the two visual systems hypothesis is growing larger,
the proponents of this theory think it remains to be established whether these fmdings should
be taken as evidence against the two visual systems hypothesis (reviewed by Bruno, 2001;
Carey, 2001; Glover, 2002; Plodowski & Jackson, 2001). There have been attempts to
reconcile the recent findings of perceptual illusions influencing the control of object-directed
actions with the two visual systems hypothesis (reviewed in Goodale & Westwood; 2004).
For example, Haffenden and Goodale (2000) argued that the sensitivity of grasping to the
Ebbinghaus illusion can be understood in terms of the surrounding circles being treated as
obstacles. However, when changing the gap size between the central target disk and the large
or small surrounding circles of the Ebbinghaus illusion (figure 1.2a), Franz et al. (2003) have
shown that the effects on grasping are also present in perception.
b
Figure 6.1: The simultaneous tilt illusion (a): the central gratings are parallel, but appear tilted in a direction opposite to the surrounding grating. The rod-and-frame illusion (b): the central lines are parallel, but appear tilted in a direction opposite to the surrounding frame.
Another reason why an illusion might affect a visually guided action, is given by Dyde and
Milner (2002). They have suggested that the effect of an illusion on action depends on the
level at which the illusion is generated in the brain. Eysel (2003) has shown that depending
on the type of visual illusion, an illusion can already be represented in V1. If an illusion is
82

Summary and conclusions
generated at an early stage in the visual system, i.e. before the separation of the dorsal and
ventral stream, the illusory information will flow into both streams causing effects on
perception as well as on action. The influence of an illusion generated at a higher level should
be restricted to the ventral processing stream and thus only have a strong effect on visual
perception. Dyde and Milner (2002; Milner & Dyde, 2003) have tested this hypothesis with
the simultaneous tilt illusion (STI; figure 6.la), and the rod-and-frame illusion (RFI; figure
6.1 b). The STI is assumed to be generated at a low level in the visual system and the RFI at a
higher level. As they predicted, the results showed equivalent effects of the STI on the
orientation of the hand and on matching. The RFI only showed an effect on the perceptual
matching task. A problem with this interpretation is how to find out exactly at what level in
the brain an illusion is generated. Another difficulty is formed by the results of our Roelofs
experiment (chapter 2). According to Dyde and Milner the Roelofs effect is generated at a
high level and thus would always affect a perceptual task. The disappearance of the Roelofs
effect in our dual task (chapter 2) cannot easily be explained with this theory.
Reconciling two visual systems with two theories of perception
Norman (2002) has tried to reconcile two competing theories in visual perception (the
constructivist and the ecological approach) with the two visual systems. According to the
constructivists perception is an active process in which information (cues and constancies)
that reaches our senses from the outside world is combined with stored knowledge. In this
way observers construct a representation of the world. An intelligent perceptual system, in
which memory and knowledge play an important role, is needed to process the incoming
information. To identify and later recognize objects in the environment, which is the fimction
of the ventral stream, information needs to be stored in and retrieved from memory. Norman
linked the constructivist approach to the ventral stream.
Ecologists argue that the information in the environment is sufficient. We pick up
information from the surrounding world without the need of mental processes. Perception is a
direct and immediate process. Neither representations nor memory are necessary because the
affordances (potential use of objects) and invariants (aspects of the visual environment that
remain the same as we move around) can be perceived directly from the objects in the
environment. Norman linked the ecological approach to the dorsal stream, where information
needed for the control of actions is processed with little or no conscious awareness and
memory.
83

Chapter 6
An important point for Norman's distinction is that the ventral stream is believed to consider
contextual information, while the dorsal stream is believed not to do so. This difference in
processing contextual information is what distinguishes ecologist's invariants from
constructivist's cues and constancies. Thus the influence of illusions, which often arise from
misinterpreting the context, can be considered critical for this debate. Many studies have
compared information processing in the dorsal and ventral stream by comparing the influence
of illusions in perceptual and motor tasks. In perceptual tasks, assumed to be processed by the
ventral system, illusions obviously show an influence on the measured variables (otherwise,
they would not be illusions). In motor tasks, assumed to be processed by the dorsal system,
often no influence is found.
Although they have received less attention, many experiments show that motor tasks can be
influenced by illusions. When hitting a moving target with ones hand, a moving background
can lead to changes in the hand's speed (Smeets & Brenner, 1995a) and in the hand's
direction (Smeets & Brenner, 1995b). Bridgeman et al. (1997) studied pointing movements
towards a target within a frame. For half of their subjects, an offset of the frame from the
subject's objective median plane caused a bias in the pointing movements in the opposite
direction. Brenner and Smeets (1996) demonstrated that the force exerted to lift an object is
influenced by the Ponzo illusion. Jackson and Shaw (2000) found the same for grip force.
Yamagishi et al. (2001) showed that pointing movements towards a small window with a
moving grating displayed behind it were biased in the direction of the grating's motion.
These experiments show that when performing motor tasks, as used by the ecologists to study
invariants, subjects can be fooled by illusions.
Individual illusions also do not influence performance in all perceptual tasks. Smeets and
Brenner (1995a) showed that background motion influences the perceived motion of a target,
but not the perceived position. Similarly, the Muller-Lyer illusion influences perceived size
but not the perceived positions of the endpoints (Gillam & Chambers, 1985). Vishton et al.
(1999) showed that the horizontal-vertical illusion is reduced considerably if the perceptual
judgement is an absolute judgement of a single element of the display instead of a relative
judgement of two elements. Similar results were obtained for the Ebbinghaus illusion (Pavani
et al., 1999) and the Muller-Lyer illusion (Franz et al., 2001). These experiments show that
when performing perceptual tasks, as used by constructivists to study cues, the effect of the
illusion can be absent.
84

Summary and conclusions
Whether an illusion affects a task does not depend on whether the task is an ecologist's motor
task or a constructivist's perceptual task, but largely on the question asked or variable
studied. However, the influence of illusions is not even fixed within a single experimental
paradigm for a single question. We showed this in chapter 2 using a constructivist's paradigm
based on the induced Roelofs effect (figure 2.3 & 2.5) and in chapter 4 using an ecologist's
paradigm based on moving a pencil to a target.
We argue that the illusory influences on both perception and action depend on the aspect of
the task that is studied and on the circumstances under which this is done. Since contextual
illusions are generally linked to the ventral stream, the ecological and constructivist approach
cannot correspond with the dorsal and ventral stream respectively.
Overall, it has become clear in this thesis that the "two visual systems" hypothesis of Milner
and Goodale, with strictly separate processing for perception and action, is not supported by
our data. Also the "common source" model of Franz et al. and the "planning-control" model
of Glover and Dixon cannot explain all our results. The theory of Smeets and Brenner about
the inconsistent processing of related attributes seems to be the best theory to explain the
experimental results.
85


Hoofdstuk6
Samenvatting en conclusies

Hoofdstuk6
In dit proefschrift is onderzocht hoe visuele illusies, die onze waarnerning (perceptie)
verstoren, het uitvoeren van handelingen (actie) kunnen bemvloeden. Er bestaan
verschillende theorieen over de verwerking van visuele informatie in het brein en hoe deze
informatie ons handelen bei"nvloedt. De meest invloedrijke theorie is de "two visual systems"
theorie van Milner en Goodale (1995). Deze theorie is gebaseerd op het feit dat er twee
visuele stromen (of paden) in de hersenen zijn gevonden: de ventrale en de dorsale stroom.
De functie van de ventrale stroom is het herkennen en identificeren van objecten in de
omgeving (bijvoorbeeld de kleur, textuur en vorm van een object). Om een object later
nogmaals te kunnen herkennen, moet de visuele informatie onafhankelijk zijn van de hoek
waaronder naar een object wordt gekeken of de positie waar het lichaam zich bevindt. Dit
betekent dat visuele informatie in de ventrale stroom wordt opgeslagen relatief ten opzichte
van andere objecten in de omgeving (allocentrisch referentiekader). De dorsale stroom
verwerkt de visuele informatie die nodig is voor het uitvoeren van handelingen naar die
objecten. Bijvoorbeeld om een object te kunnen oppakkenlgrijpen, is het niet voldoende te
weten waar het object zich bevindt ten opzichte van andere objecten in de omgeving; de
positie ten opzichte van het lichaam moet ook worden bepaald. Informatie in de dorsale
stroom wordt daarom opgeslagen in een egocentrisch referentiekader (ten opzichte van het
lichaam). Omdat de visuele informatie in de ventrale stroom ("perceptie") afhankelijk is van
de omgeving en die in de dorsale stroom ("actie") niet, wordt volgens deze theorie een
perceptuele taak, uitgevoerd door de ventrale stroom, wei door visuele illusies bei"nvloed en
een actie taak, uitgevoerd door de dorsale stroom, niet.
Toch blijkt deze "two visual systems" theorie niet altijd alle resultaten te kunnen verklaren.
In sommige studies werd een effect van illusies op actie gevonden dat net zo groot was als
het effect op perceptie (Franz et al., 2000; Pavani et al., 1999). Als altematief voor de "two
visual systems" theorie, stelden Franz en collega's (2000) voor dat er een
gemeenschappelijke bron van informatie wordt gebruikt voor zowel het genereren van een
visueel percept als voor het grijpen van een object (actie). Als deze informatie is bei"nvloed
door een visuele illusie, dan worden in perceptie en actie dezelfde illusie effecten gevonden.
Deze interpretatie zal worden aangeduid met de term "common source" model.
Glover en Dixon (2001a) kwamen met een model waarin de resultaten van studies met en
zonder effect van illusies op actie, verenigd kunnen worden. In dit zogenaamde "planning
control" model wordt een onderscheid gemaakt tussen de planningsfase van een beweging en
de controlefase. Om een object te kunnen oppakken!grijpen, moet van tevoren een planning
worden gemaakt hoe erheen te bewegen. Deze planningsfase en het begin van de beweging
88

Samenvatting en conclusies
zijn gebaseerd op context afhankelijke visuele informatie. Met andere woorden de planning
en bet begin van de beweging kunnen worden beinvloed door visuele illusies. Gedurende de
beweging kunnen fouten in de planning, veroorzaakt door visuele illusies, on-line worden
gecorrigeerd door visuele en proprioceptieve feedback. Het relatief grote effect dat een
visuele illusie heeft aan bet begin van een beweging, neemt af hoe dichter de hand bet doel
nadert.
Smeets en Brenner (1995a) hebben een andere verklaring voor bet feit dat illusies
grijpbewegingen kunnen beinvloeden. Zij suggereren dat verschillende spatiele attributen
worden gebruikt in perceptie en in actie (grijpen). Dit kan het beste worden uitgelegd aan de
hand van een voorbeeld. Fysiek gezien zijn lengte en positie aan elkaar gekoppeld; bet
verschil tussen twee posities komt overeen met de lengte van een lijnstuk tussen die twee
posities. Maar deze spatiele attributen (lengte en positie), die onafhankelijk van elkaar
worden verwerkt in de hersenen, hoeven niet noodzakelijkerwijs consistent te worden
waargenomen. Bijvoorbeeld, de Muller-Lyer illusie verandert de waargenomen lengte van de
lijn, zonder dat de waargenomen eindposities van die lijn veranderen (figuur 1.3). Wanneer
de illusoire lengte van de lijn wordt gebruikt om een taak uit te voeren, b.v. voor het
perceptueel inschatten van de lengte van de lijn, dan vind je een effect van de illusie. Voor
bet grijpen van een staafje dat op de lijn van de Muller-Lyer illusie is gelegd, zijn de posities
waar het staafje vastgepakt wordt van belang en niet de waargenomen lengte van dat staafje.
En aangezien deze posities niet door de illusie waren beinvloed, zal ook geen effect van de
illusie op bet grijpen te vinden zijn. In hun theorie gaan Smeets en Brenner gaan er vanuit dat
een taak:, onafhankelijk of die taak als perceptie of actie geclassificeerd wordt, aileen wordt
bemvloed door een illusie als een incorrect waargenomen attribuut wordt gebruikt om die
taak uit te voeren. Met behulp van verschillende visuele illusies hebben we de voorgenoemde
theorieen getest omtrent de invloed van illusies op visuele perceptie en actie.
Het "originele" en "geinduceerde" Roelofs effect
De "two visual systems" theorie van Milner en Goodale gaat er vanuit dat er een volledige
scheiding is in de verwerking van informatie voor perceptie en die van informatie voor actie.
Ondersteuning voor deze theorie komt voort uit experimenten, waarin perceptuele taken door
visuele illusies worden bemvloed, terwijl visuomotor taken nauwkeurig worden uitgevoerd.
Bridgeman en collega's (Bridgeman et al., 1997, 2000; Bridgeman & Huemer, 1998) hebben
de invloed van het "gemduceerde" Roelofs effect onderzocht op perceptie en actie. Bij het
"gemduceerde" Roelofs effect wordt in het donker een stip gepresenteerd, omgeven door een
89

Hoofdstuk6
rechthoekig frame. Dit frame is iets naar links of naar rechts verschoven ten opzichte van
"recht vooruit". W anneer naar de positie van de stip wordt gevraagd, wijkt het antwoord af in
de richting tegenovergesteld aan de verschuiving van het frame. Bijvoorbeeld: de stip staat
precies in het midden ("recht vooruit") en het frame is naar links verschoven ten opzichte van
"recht vooruit". W anneer nu naar de positie van de stip wordt gevraagd antwoordt de
proefpersoon dat deze iets rechts van "recht vooruit" staat. Echter, een doelgerichte
armbeweging naar de positie van diezelfde stip is nauwkeurig, ondanks de perceptuele
afwijking. Met deze dissociatie tussen perceptie en actie leken Bridgeman en coilega's de
"twee visuele systemen" theorie van Milner en Goodale te bevestigen. In hoofdstuk 2
onderzochten we of het mogelijk was de resultaten van Bridgeman te verklaren zonder twee
visuele systemen te verondersteilen. We wilden daarom weten wat de oorzaak is van het
"gei"nduceerde" Roelofs effect. Om hier achter te komen, richtten we ons eerst op het
"originele" Roelofs effect. Het "originele" bestaat in tegensteiling tot het "gemduceerde"
Roelofs effect aileen uit een frame. Als een rechthoekig frame wordt gepresenteerd met een
verschuiving naar links ten opzichte van "recht vooruit", dan wordt het frame meer naar
rechts waargenomen dan het werkelijk is. Zou het "gei:nduceerde" (alsmede het originele)
Roelofs effect veroorzaakt kunnen worden door een verschuiving in de waarneming van wat
"recht vooruit" is? We hadden twee taken waarin het "geihduceerde" Roelofs effect aan
proefpersonen werd getoond. In de ene taak moest altijd de positie van de stip ten opzichte
van "recht vooruit" worden gerapporteerd, terwijl in de andere taak niet van tevoren bekend
was of proefpersonen de positie van de stip of die van het frame moesten rapporteren. Deze
twee taken resulteerden in een verschillende invloed van de Roelofs illusie. Net als
Bridgeman en coilega's vonden we een "gei"nduceerd" Roelofs effect wanneer aileen naar de
positie van de stip werd gevraagd. Dit effect verdwijnt wanneer we in hetzelfde experiment
ook naar de positie van het frame vragen. We concludeerden dater geen conclusies getrokken
moeten worden over het bestaan van twee visuele systemen aileen op basis van een verschil
in invloed van de iilusie tussen perceptie en actie.
De Ebbinghaus figuur verandert meer dan aileen de waargenomen grootte
Er zijn verschiilende verklaringen mogelijk voor een effect van een (grootte)iilusie op
grijpen. Franz et al. (2000) stelde in het "common source" (gemeenschappelijke bron) model,
dat een effect van de Ebbinghaus iilusie op grijpen kan worden verklaard doordat dezelfde
illusoire grootte inforrnatie wordt gebruikt in perceptie en in actie. Volgens Haffenden en
collega's (Haffenden & Goodale, 2000; Haffenden et al., 2001) kan een effect van de illusie
op grijpen niet veroorzaakt worden door de iilusoir waargenomen grootte, omdat perceptie en
90

Samenvatting en conclusies
actie gescheiden zijn. Zij denken dat de omliggende cirkels van de Ebbinghaus illusie kunnen
worden gezien als obstakels, die interfereren met het grijpen. Smeets et al. hebben laten zien
dat een effect van de omliggende cirkels op de maximum handopening (maximale afstand
tussen vinger en duim tijdens een grijp beweging) kan worden verklaard in termen van een
verandering in de benodigde precisie (Smeets et al., 2003).
We proberen een onderscheid te maken tussen deze interpretaties in hoofdstuk 3, door de
spatiele positie van de omliggende cirkels ten opzichte van de centrale cirkel in de
Ebbinghaus illusie te veranderen (figuur 3.1). Proefpersonen kregen een afbeelding van een
Ebbinghaus illusie te zien, bestaand uit een centrale rniddencirkel omringd door vier grote of
kleine cirkels. De spatiele posities van de ornliggende cirkels kon 45° geroteerd zijn ten
opzichte van de rniddencirkel. Er waren twee verschillende taken voor de proefpersoon, die
onafhankelijk van elkaar werden uitgevoerd. In de eerste taak, een perceptuele taak, werd aan
de proefpersoon gedurende 1 seconde een van de Ebbinghaus figuren getoond. Daarna werd
een losse referentie cirkel getoond (zonder omringende cirkels). De proefpersonen moesten
met de computermuis deze cirkel even groot in te stellen als de rniddencirkel die ze net in de
Ebbinghaus figuur hadden gezien. In de andere taak, een grijptaak, werden dezelfde
Ebbinghaus figuren gebruikt. Maar nu bleef de Ebbinghaus figuur zichtbaar en werd er bij
iedere aanbieding van een figuur een dun zwart schijfje op de rniddencirkel gelegd. De taak
van de proefpersoon was om dit schijfje op te pakken en weg te leggen. Met behulp van
kleine infrarood led-jes, die op de vinger en duim van de proefpersoon werden geplakt, werd
het verloop van de grijpbeweging gemeten. Voor de perceptuele taak voorspellen alle
theorieen ("common source" model van Franz et al.; Haffenden et al. en Smeets et al.) dat de
referentie cirkel groter wordt ingesteld wanneer een Ebbinghaus figuur is omringd door
kleine dan door grote cirkels. Geen van de theorieen heeft een duidelijke voorspelling voor
een effect van de spatiele positie van de ornliggende cirkels op perceptie. Voor het grijpen
van de rniddelste cirkel, voorspelt het "common source" model geen effect van de spatiele
positie op het grijpen, als perceptie er ook niet door wordt beihvloed (het effect op perceptie
is gelijk aan het effect of actie volgens deze theorie). Als de ornliggende cirkels op de een of
andere marrier interfereren met het grijpen, dan verwachten beide andere interpretaties
(Haffenden et al. and Smeets et al.) een effect op het grijpen te vinden. De resultaten laten
zien dat de waargenomen grootte van de rniddelste cirkel (perceptie) alleen wordt beihvloed
door de grootte van de omliggende cirkels en niet door de spatiele positie ervan. Dat wil
zeggen dat, ongeacht de positie van de ornliggende cirkels, de rniddelste cirkel groter wordt
waargenomen wanneer deze is omringd door kleine cirkels dan dezelfde cirkel omringd door
grote cirkels. Bij het grijpen vonden we dat de maximum handopening aileen wordt
91

Hoofdstuk6
bei:nvloed door de grootte van de omliggende cirkels. Kortom, de illusoire grootte van de
middelste cirkel wordt weerspiegeld in de maximum handopening. De orientatie van de hand
tijdens de beweging en de handopening aan het eind van de grijpbeweging hangen alleen af
van de spatiele positie van de omliggende cirkels. Dat wil zeggen dat de hand op een andere
manier naar de middelste cirkel beweegt en deze oppakt als de positie van de omliggende
cirkels is veranderd. De gevonden invloed van de omliggende cirkels op grijpen kan dus niet
alleen veroorzaakt worden door een verkeerd ingeschatte grootte van de middelste cirkel,
zoals het "common source" model voorspelt.
Ook in een ander experiment (Biegstraaten et al., submitted) hebben we gevonden dat non
illusoire effecten deels verantwoordelijk zijn voor de invloed van illusies op grijpen. In dat
experiment lieten we proefpersonen een staafje grijpen dat hoven op een geprojecteerde
Miiller-Lyer figuur was gelegd. Zowel de configuratie van de Miiller-Lyer figuur (naar
binnen of naar buiten wijzende pijlen, zie figuur 1.2b) als de start positie van de hand (onder
of opzij van de figuur) bemvloedden de maximum handopening, tijd tot maximum
handopening en de totale bewegingstijd. De configuratie had ook nog een effect op de
handopening op het eind van de beweging. Dus ook in dit geval bemvloedden de grafische
elementen de beweging op een andere manier dan door het veranderen van de waargenomen
grootte alleen. Dit maakt het moeilijker om exact te bepalen welke attributen worden gebruikt
in een motor taak.
De codering van wijsbewegingen en saccades
Volgens de theorie van Smeets en Brenner bei:nvloeden visuele illusies ons oordeel over
bepaalde visuele attributen. Bijvoorbeeld, de Miiller-Lyer illusie verandert wel de
waargenomen lengte van een lijn maar niet de waargenomen posities van de eindpunten van
die lijn (figuur 1.3). Daarom verwachten we dat visuele illusies aileen invloed hebben op een
taak, wanneer deze incorrect waargenomen attributen worden gebruikt om de taak uit te
voeren. Voor het wijzen naar een bepaald punt kunnen verschillende visuele attributen
worden gebruikt om de hand aan te sturen: de richting en afstand (of lengte) van de
verplaatsing (vector codering) of de positie van het eindpunt van de beweging (positie
codering). In hoofdstuk 4 bepaalden we welke attributen worden gebruikt in een wijstaak. Als
een combinatie van vector codering en positie codering wordt gebruikt, dan kan de
zichtbaarheid van de hand en de stimulus de verhouding van de gebruikte attributen doen
veranderen en daarmee ook de invloed van de illusie. Aan de proefpersonen wordt gevraagd
met de punt van een pen heen en weer te wijzen tussen stippen die op de pijlpunten van de
92

Sarnenvatting en conclusies
Brentano illusie (figuur 4.1) verschijnen en een stip buiten de figuur. Alle proefpersonen
voerden vier condities van deze wijstaak uit, waarin het zicht op de hand en/of stimulus bij
het begin van de beweging kon worden ontnomen (vier mogelijke combinaties). De illusie
blijkt wijsbewegingen tussen de pijlpunten te bemvloeden, wat duidt op het gebruik van
lengte informatie in de wijsbeweging. Bewegingen die van buiten komen (loodrecht op de
Brentano figuur) worden niet door de illusie befuvloed. Wanneer de hand en/of doel
onzichtbaar zijn tijdens de beweging, neemt de invloed van de illusie toe.
Om beter te begrijpen welke informatie wordt gebruikt in de verschillende condities van de
wijstaak, hebbben we de resultaten vergeleken met een perceptuele taak, waarin alleen lengte
informatie gebruikt kan worden. In deze taak werd dezelfde Brentano figuur getoond als in de
wij staak. Proefpersonen moesten rechts naast de figuur een ( onzichtbare) lijn trekken, die in
lengte overeenkwam met het lijnstuk tussen de "naar binnen" of "naar buiten" wijzende
pijlen. Bij het tekenen van een lijn met een bepaalde lengte, is het waarschijnlijk dat aileen de
lengte informatie van het betreffende lijnstuk uit de Brentano figuur wordt gebruikt. Daarom
is het effect dat we in deze perceptuele taak vinden, het maximale effect dat we op de
wijstaak kunnen verwachten. De invloed van de illusie in de condities van de wijstaak was
veel kleiner dan die op de perceptuele taak (maximaal 1/3 van het perceptuele effect), wat
betekent dat de bijdrage van vector codering in alle condities van de wijstaak gering was.
In hoofdstuk 4 vonden we dat wijsbewegingen zijn gebaseerd op een combinatie van vector
codering en positie codering. Maar hoe zijn saccades (snelle oogbewegingen waarbij nieuwe
informatie op de fovea wordt gebracht) gecodeerd? Een mogelijkheid is dat saccades en
wijsbewegingen gemeenschappelijk worden aangestuurd ("common motor commands"). In
dat geval verwachten we eenzelfde combinatie van vector codering en positie codering voor
saccades als voor wijsbewegingen. Een andere mogelijkheid is dat saccades geheel vector
gecodeerd zijn. Dat betekent in dat geval dat het oog en de hand verschillende informatie
gebruiken. We verwachten dan een verschillend effect van de illusie op saccades dan op
wijsbewegingen. In hoofdstuk 5 gebruiken we dezelfde Brentano illusie om te bepalen of
saccades op eenzelfde marrier gecodeerd worden als wijsbewegingen. Proefpersonen werd nu
gevraagd om saccades te maken (heen en weer kijken) tussen stippen die op de pijlpunten van
de Brentano illusie verschenen en een stip buiten de figuur. De illusie beYnvloedt saccades
tussen de pijlpunten, maar niet de saccades van buiten de figuur. Ook in dit experiment werd
weer een perceptuele taak uitgevoerd om te kijken hoeveel vector informatie in saccades
wordt gebruikt. Proefpersonen kregen weer dezelfde Brentano figuur te zien met een rode stip
op de middelste pijlpunt en een op de onderste ofbovenste pijlpunt. Na een korte presentatie
93

Hoofdstuk6
van de Brentano figuur verscheen een lijn zonder pijlpunten of stippen, waarvan beoordeeld
moest worden of die lijn langer of korter was dan het lijnstuk tussen de rode stippen dat even
ervoor getoond was. Het perceptuele illusie effect verschilde niet van het effect op saccades.
Hieruit kunnen we concluderen dat, in tegenstelling tot wijsbewegingen, saccades
voomamelijk vector gecodeerd zijn.
In dit proefschrift hebben we een aantal theorieen over de verwerking van (illusoire) visuele
informatie getest met behulp van visuele illusies. Uit onze experimenten is gebleken dat de
"two visual systems" theorie van Milner en Goodale, waarin gescheiden verwerking wordt
verondersteld tussen perceptie en actie, niet wordt ondersteund. Ook het "common source"
model van Franz en collega's en het "planning-control" model van Glover en Dixon kunnen
niet al onze data verklaren. De theorie van Smeets en Brenner met betrekking tot
inconsistente verwerking van attributen, lijkt het beste onze experimentele data te kunnen
verklaren.
94

References
References
Abrams, R.A., Meyer, D.E., & Kornblum, S. (1990). Eye-hand coordination: oculomotor
control in rapid aimed limb movements. Journal of Experimental Psychology: Human
Perception and Performance, 16, 248-267.
Aglioti, S., DeSouza, J.F., & Goodale, M.A. (1995). Size-contrast illusions deceive the eye
but not the hand. Current Biology, 5, 679-685.
Balint, R. (1909). Seelenliimung des 'Schauens', optische Ataxie, riiumliche Storung der
Aufmerksamkeit. Monatschriftfor Psychiatrie und Neurologie, 25, 51-81.
Becker, W., & Ji.irgens, R. (1979). An analysis of the saccadic system by means of double
step stimuli. Vision Research, 19, 967-983.
Bekkering, H., Abram, R.A., & Pratt, J. (1995). Transfer of saccadic adaptation to the manual
motor system. Human Movement Science, 14, 155-164.
Biegstraaten, M. de Grave, D.D.J., Smeets, J.B.J., & Brenner, E. (submitted). Grasping the
Muller-Lyer illusion: More than just a change in length.
Binsted, G., & Elliott, D. (1999a). The Muller-Lyer illusion as a perturbation to the saccadic
system. Human Movement Science, 18, 103-117.
Binsted, G., & Elliott, D. (1999b). Ocular perturbations and retinaVextraretinal information:
the coordination of saccadic and manual movements. Experimental Brain Research, 127,
193-206.
Biguer, B., Jeannerod, M., & Prablanc, C. (1982). The coordination of eye, head and arm
movements during reaching at a single visual target. Experimental Brain Research, 46, 301-
304.
Biguer, B., Prablanc, C., & Jeannerod, M. (1984). The contribution of coordinated eye and
head movements in hand pointing accuracy. Experimental Brain Research, 55,462-469.
Bizzi, E., Kalil, R.E., & Tagliasco, V. (1971). Eye-head coordination in monkeys: evidence
for centrally patterned organization. Science, 173, 452-454.
95

References
Bizzi, E., Hogan, N., Mussa-Ivaldi, F.A., & Giszter, S. (1992). Does the nervous system use
equilibrium point control to guide single and multiple joint movements? Behavioral and
Brain Sciences, 15, 603-613.
Bock, 0. (1986). Contribution of retinal versus extraretinal signals towards visual localization
in goal-directed movements. Experimental Brain Research, 64,476-482.
Bock, 0., & Eckmiller, R. (1986). Goal-directed arm movements in absence of visual
guidance: evidence for amplitude rather than position control. Experimental Brain Research,
62, 451-458.
Bradshaw, M.F., & Watt, S.J. (2002). A dissociation of perception and action in normal
human observers: the effect of temporal delay. Neuropsychologia, 40, 1766-1778.
Brenner, E., & Smeets, J.B.J. (1996). Size illusion influences how we lift but not how we
grasp an object. Experimental Brain Research, Ill, 473-476.
Bridgeman, B., Lewis, S., Heit, G., & Nagle, M. (1979). Relation between cognitive and
motor-oriented systems of visual position perception. Journal of Experimental Psychology:
Human Perception and Performance, 5, 692-700.
Bridgeman, B., Kirch, M., & Sperling, A. (1981). Segregation of cognitive and motor aspects
of visual function using induced motion. Perception and Psychophysics, 29, 336-342.
Bridgeman, B., Peery, S., & Anand, S. (1997). Interaction of cognitive and sensorimotor
maps of visual space. Perception and Psychophysics, 59,456-469.
Bridgeman, B., & Huemer, V. (1998). A spatially oriented decision does not induce
consciousness in a motor task. Consciousness and Cognition, 7, 454-464.
Bridgeman, B., Gemmer, A., Forsman, T., & Huemer, V. (2000). Processing spatial
information in the sensorimotor branch of the visual system. Vision Research, 40, 3539-3552.
Bruno, N. (2001). When does action resist visual illusions? Trends in Cognitive Sciences, 5,
379-382.
Carey, D.P. (2001). Do action systems resist visual illusions? Trends in Cognitive Sciences,
5, 109-113.
96

References
Carrazzo, M., Mcintyre, J., Zago, M., & Lacquaniti, F. (1999). Viewer-centered and body
centered frames of reference in direct visuomotor transformations. Experimental Brain
Research, 129, 201-210.
Danckert, J.A., Sharif, N., Haffenden, A.M., Schiff, K.C., & Goodale, M.A. (2002). A
temporal analysis of grasping in the Ebbinghaus illusion: planning versus online control.
Experimental Brain Research, 144, 275-280.
Dassonville, P., & Bala, J.K. (2004). Are the original Roelofs effect and the induced Roelofs
effect confounded by the same expansion of remembered space? Vision Research, 44, 1025-
1029.
Dassonville, P., Bridgeman, B., Bala, J.K., Thiem, P., & Sampanes, A. (2004). The induced
Roelofs effect: two visual systems or the shift of a single reference frame? Vision Research,
44, 603-611.
De Grave, D.D.J., Brenner, E., & Smeets, J.B.J. (2002). Are the original Roelofs effect and
the induced Roelofs effect caused by the same shift in straight ahead? Vision Research, 42,
2277-2283.
De Grave, D.D.J., Brenner, E. & Smeets, J.B.J. (2004). Illusions as a tool to study the coding
of pointing movements. Experimental Brain Research, 155, 56-62.
DeLucia, P.R., Tresilian, J.R., & Meyer, L.E. (2000). Geometrical illusions can affect time
to-contact estimation and mimed prehension. Journal of Experimental Psychology: Human
Perception and Performance, 26, 552-567.
Desmurget, M., Pelisson, D., Rossetti, Y., & Prablanc, C. (1998). From eye to hand: planning
goal-directed movements. Neuroscience Biobehavioral Review, 22, 761-788.
Dyde, R.T., & Milner, A.D. (2002). Two illusions of perceived orientation: one fools all of
the people some of the time; the other fools all of the people all of the time. Experimental
Brain Research, 144, 518-527.
Eysel, U.T. (2003). Illusions and perceived images in the primate brain. Science, 302, 789-
790.
97

References
Feldman, A.G., & Levin, M.F. (1995). The origin and use of positional frames of reference in
motor control. Behavioral and Brain Sciences, 18, 723-806.
Festinger, L., White, C.W., & Allyn, M.R. (1968). Eye movements and decrement in the
Muller-Lyer illusion. Perception & Psychophysics, 3, 376-382.
Findlay, J.M. (1982). Global visual processing for saccadic eye movements. Vision Research,
22, 1033-1045.
Franz, V. (2003). Manual size estimation: a neuropsychological measure of perception?
Experimental Brain Research, 151, 4 71-4 77.
Franz, V.H., Gegenfurtner, K.R., Billthoff, H.H., & Fahle, M. (2000). Grasping visual
illusions: no evidence for a dissociation between perception and action. Psychological
Science, 11, 20-25.
Franz, V.H., Fahle, M., Billthoff, H.H., & Gegenfurtner, K.R. (2001). Effects of visual
illusions on grasping. Journal of Experimental Psychology: Human Perception and
Performance, 27, 1124-1144.
Franz, V., BUlthoff, H.H., & Fahle, M. (2003). Grasp effects of the Ebbinghaus illusion:
obstacle avoidance is not the explanation. Experimental Brain Research, 149, 470-477.
Frens, M.A., & Van Opstal, A.J. (1994). Transfer of short-term adaptation in human saccadic
eye movements. Experimental Brain Research, 100, 293-306.
Gentilucci, M., Chieffi, S., Daprati, E., Saetti, M.C., & Toni, I. (1996). Visual illusion and
action. Neuropsychologia, 34, 369-376.
Gillam, B., & Chambers, D. (1985). Size and position are incongruous: measurements on the
MUller-Lyer figure. Perception & Psychophysics, 37, 549-556.
Gillam, B. (1998). Illusions at century's end. In: Hochberg, J. ( ed). Perception and cognition
at century's end. Academic Press, London, 95-136.
Gillan, D.J., Schmidt, W., & Hanowski, R.J. (1999). The effect of the Muller-Lyer illusion on
map reading. Perception & Psychophysics, 61, 1154-1167.
98

References
Glover, S. (2002). Visual illusions affect planning but not control. Trends in Cognitive
Sciences, 6, 288-292.
Glover, S., & Dixon, P. (200la). Dynamic illusion effects in a reaching task: Evidence for
separate visual representations in the planning and control of reaching. Journal of
Experimental Psychology: Human Perception and Performance, 27, 560-572.
Glover, S., & Dixon, P. (200lb). Motor adaptation to an optical illusion. Experimental Brain
Research, 137, 254-258.
Glover, S., & Dixon, P. (200lc). The role of vision in the on-line correction of illusion effects
on action. Canadian Journal of Experimental Psychology, 55, 96-103.
Glover, S., & Dixon, P. (2002). Dynamic effects of the Ebbinghaus illusion in grasping:
support for a planning/control model of action. Perception & Psychophysics, 64, 266-278.
Goodale, M.A., Milner, A.D., Jak.obson, L.S., & Carey, D.P. (1991). A neurological
dissociation between perceiving objects and grasping them. Nature, 349, 154-156.
Goodale, M.A., & Milner, A.D. (1992). Separate visual pathways for perception and action.
Trends in Neurosciences, 15, 20-25.
Goodale, M.A., Meenan, J.P., BUlthoff, H.H., Nicolle, D.A., Murphy, K.J., & Racicot, C.I.
(1994). Separate neural pathways for the visual analysis of object shape in perception and
prehension. Current Biology, 4, 604-610.
Goodale, M.A., & Haffenden, A.M. (1998). Frames of reference for perception and action in
the human visual system. Neuroscience & Biobehavioral Reviews, 22, 161-172.
Goodale, M.A., & Humphrey, G.K. (1998). The objects of action and perception. Cognition,
67, 181-207.
Goodale, M.A., & Westwood, D.A. (2004). An evolving view of duplex vision: separate but
interacting cortical pathways for perception and action. Current Opinion in Neurobiology, 14,
1-9.
Gregory, R.L. (1968). Visual illusions. Scientific American, 219, 66-76.
99

References
Gregory, R.L. (1998). Eye and Brain: The Psychology of Seeing. Oxford University Press,
Oxford.
Haffenden, A.M., & Goodale, M.A. (1998). The effect of pictorial illusion on prehension and
perception. Journal of Cognitive Neuroscience, 10, 122-136.
Haffenden, A.M., & Goodale, M.A. (2000). Independent effects of pictorial displays on
perception and action. Vision Research, 40, 1597-1607.
Haffenden, A.M., Schiff, K.C., & Goodale, M.A. (2001). The dissociation between
perception and action in the Ebbinghaus illusion: Nonillusory effects of pictorial cues on
grasp. Current Biology, 11, 177-181.
Hanisch, C., Konczak, J., & Dohle, C. (2001). The effect of the Ebbinghaus illusion on
grasping behaviour of children. Experimental Brain Research, 137, 237-245.
He, P., & Kowler, E. (1989). The role of location probability in the programming of
saccades: implications for "center-of-gravity" tendencies. Vision Research, 29, 1165-1181.
Hu, Y., & Goodale, M.A. (2000). Grasping after a delay shifts size-scaling from absolute to
relative metrics. Journal of Cognitive Neurosciences, 12, 856-868.
Jackson, S.R., & Shaw, A. (2000). The Ponzo illusion affects grip-force but not grip-aperture
scaling during prehension movements. Journal of Experimental Psychology: Human
Perception and Performance, 26, 418-423.
Jakobson, L.S., Archibald, Y.M., Carey, D.P., & Goodale, M.A. (1991). A kinematic analysis
of reaching and grasping movements in a patient recovering from optic ataxia.
Neuropsychologia, 29, 803-809.
Jeannerod, M. (1986). The formation offmger grip during prehension. A cortically mediated
visuomotor pattern. Behavioural Brain Research, 19, 99-116.
Jeannerod, M., Decety, J., & Michel, F. (1994). Impairment of grasping movements
following a bilateral posterior parietal lesion. Neuropsychologia, 32, 369-380.
Kwok, RM., & Braddick, O.J. (2003). When does the Titchener Circles illusion exert an
effect on grasping? Two- and three-dimensional targets. Neuropsychologia, 41, 932-940.
100

References
Lekwuwa, G.U., & Barnes, G.R. (1996). Cerebral control of eye movements. I. The
relationship between cerebral lesion sites and smooth pursuit deficits. Brain, 119, 473-490.
Loomis, J.M., Da Silva, J.A., Fujita, N., & Fukusima, S.S. (1992). Visual space perception
and visually directed action. Journal of Experimental Psychology: Human Perception and
Performance, 18, 906-921.
Lopez-Moliner, J., Smeets, J.B.J., & Brenner, E. (2003). Similar effects of a motion-in-depth
illusion on manual tracking and perceptual judgements. Experimental Brain Research, 151,
553-556.
Mack, A., Heuer, F., Villardi, K., & Chambers, D. (1985). The dissociation of position and
extent in Miiller-Lyer figures. Perception & Psychophysics, 37, 335-344.
McCarley, J.S., Kramer, A.F., & DiGirolamo, G.J. (2003). Differential effects of the Miiller
Lyer illusion on reflexive and voluntary saccades. Journal of Vision, 3, 751-760.
Mcilwain, J.T. (1991). Distributed spatial coding in the superior colliculus: a review. Visual
Neuroscience, 6, 3-13.
Mcintyre, J., Stratta, F., & Lacquaniti, F. (1997). Viewer-centered frame of reference for
pointing to memorized targets in three-dimensional space. Journal of Neurophysiology, 78,
1601-1618.
Mcintyre, J., Stratta, F., & Lacquaniti F. (1998). Short-term memory for reaching to visual
targets: Psychophysical evidence for body-centered reference frames. Journal of
Neuroscience, 18, 8423-8435.
Messier, J., & Kalaska, J.F. (1997). Differential effect of task condition on errors of direction
and extent of reaching movements. Experimental Brain Research, 115, 469-4 78.
Milner, A.D., Perrett, D.l., Johnston, R.S., Benson, P.J., Jordan, T.R., Heeley, D.W.,
Bettucci, D., Mortara, F., Mutani, R., & Terazzi, E. (1991). Perception and action in "visual
form agnosia". Brain, 114,405-428.
Milner, A.D., & Goodale, M.A. (1995). The visual brain in action. Oxford University Press,
Oxford.
101

References
Milner, A.D., & Dyde, R. (2003). Why do some perceptual illusions affect visually guided
action, when others don't? Trends in Cognitive Sciences, 7, 10-11.
Mishkin, M., Ungerleider, L.G., & Macko, K.A. (1983). Object vision and spatial vision: two
cortical pathways. Trends in Neurosciences, 6, 414-417.
Mon-Williams, M., & Bull, R. (2000). The Judd illusion: evidence for two visual streams or
two experimental conditions? Experimental Brain Research, 130, 273-276.
Newsome, W.T., & Pare, E.B. (1988). A selective impairment of motion perception
following lesions of the middle temporal visual area (MT). The Journal of Neuroscience, 8,
2201-2211.
Norman, J. (2002). Two visual systems and two theories of perception: An attempt to
reconcile the constructivist and ecological approach. Behavioral and Brain Sciences, 25, 73-
144.
Otto-de Haart, E.G., Carey, D.P., & Milne, A.B. (1999). More thoughts on perceiving and
grasping the Muller-Lyer illusion. Neuropsychologia, 37, 1437-1444.
Pavani, F., Boscagli, I., Benvenuti, F., Rabufetti, M., & Fame, A. (1999). Are perception and
action affected differently by the Titchener circles illusion? Experimental Brain Research,
127, 95-101.
Perenin, M.T., & Vighetto, A. (1988). Optic ataxia: A specific disruption in visuomotor
mechanisms. Brain, 111, 643-674.
Plodowski, A., & Jackson, S.R. (2001). Vision: getting to grips with the Ebbinghaus illusion.
Current Biology, 11, R304-306.
Post, R.B., & Welch, R.B. (1996). Is there dissociation of perceptual and motor responses to
figural illusions? Perception, 25, 569-581.
Prablanc, C., Martin, 0. (1992). Automatic control during hand reaching at undetected two
dimensional target displacements. Journal of Neurophysiology, 67, 455-469.
Proffitt, D.R., Bhalla, M., Gossweiler, R., & Midgett, J. (1995). Perceiving geographical
slant. Psychonomic Bulletin & Review, 2, 409-428.
102

References
Reina, G.A., & Schwartz, A.B. (2003). Eye-hand coupling during closed-loop drawing:
Evidence of shared motor planning? Human Movement Science, 22, 137-152.
Robinson, D.A. (1972). Eye movements evoked by collicular stimulation in the alert monkey.
Vision Research, 12, 1795-1808.
Roelofs, C.O. (1935). Die optische localisation. Archiv for Augenheilkunde, 109, 395-415.
Rossetti, Y. (1998). Implicit short-lived motor representations of space in
brain damaged and healthy subjects. Consciousness and Cognition, 7, 520-558.
Rossetti, Y., Desmurget, M., & Prablanc, C. (1995). Vectorial coding of movement: vision,
kinaesthesia, or both? Journal of Neurophysiology, 74, 457-463.
Schwartz, A.B., Moran, D.W., & Reina, G.A. (2004). Differential representation of
perception and action in the frontal cortex. Science, 303, 380-383.
Shebilske, W.L. (1981). Visual direction illusions in everyday situations: implications for
sensorimotor and ecological theories. In: Fisher, D.F., Monty, R.A., & Senders, J.W. (ed).
Eye movements: cognition and visual perception. Lawrence Erlbaum Associates Publishers,
Hillsdale, New Jersey, 95-110.
Smeets, J.B.J., & Brenner, E. (1995a). Perception and action are based on the same visual
information: distinction between position and velocity. Journal of Experimental Psychology:
Human Perception and Performance, 21, 19-31.
Smeets, J.B.J., & Brenner, E. (1995b ). Prediction of a moving target's position in
fast goal-directed action. Biological Cybernetics, 73, 519-528.
Smeets, J.B.J., & Brenner, E. (1999). A new view on grasping. Motor Control, 3, 237-271.
Smeets, J.B.J., & Brenner, E. (2001). Action beyond our grasp. Trends in Cognitive Sciences,
5, 287.
Smeets, J.B.J., Brenner, E., De Grave, D.D.J., & Cuijpers, R.H. (2002). Illusions in action:
consequences of inconsistent processing of spatial attributes. Experimental Brain Research,
147, 135-144.
103

References
Smeets, J.B.J., Glover, S., & Brenner, E. (2003). Modelling the time-dependent effect of the
Ebbinghaus illusion on grasping. Spatial Vision, 16, 311-324.
Ungerleider, L.G., & Mishkin, M. (1982). Two Cortical Visual Systems. In: Ingle, D.J.,
Goodale, M.A., & Mansfield, R.J.W. (ed). Analysis of Visual Behavior. MIT Press,
Cambridge, 549-586.
Van den Dobbelsteen, J.J., Brenner, E., & Smeets, J.B.J. (2001). Endpoints of arm
movements to visual targets. Experimental Brain Research, 138, 279-287.
Van Donkelaar, P. (1999). Pointing movements are affected by size-contrast illusions.
Experimental Brain Research, 125, 517-520.
Van Donkelaar, P., Miall, R.C., & Stein, J.F. (2000). Changes in motion perception following
oculomotor smooth pursuit adaptation. Perception & Psychophysics, 62, 378-385.
Vindras, P., & Viviani, P. (1998). Frames of reference and control parameters in visuomanual
pointing. Journal of Experimental Psychology: Human Perception and Performance, 24,
569-591.
Vishton, P.M., Rea, J.G., Cutting, J.E., & Nunez, L.N. (1999). Comparing effects of the
horizontal-vertical illusion on grip scaling and judgment: relative versus absolute, not .
perception versus action. Journal of Experimental Psychology: Human Perception and
Performance, 25, 1659-1672.
Vishton, P.M., & Fabre, E. (2003). Effects of the Ebbinghaus illusion on different behaviors:
one- and two-handed grasping; one- and two-handed manual estimation; metric and
comparitive judgment. Spatial Vision, 16, 377-392.
Werner, H., Wapner, S., & Bruell, J. H. (1953). Experiments on sensory-tonic field theory of
perception: VI. The effect of position of head, eyes and of object on the position of the
apparent medium plane. Journal of Experimental Psychology, 46, 293-299.
Westwood, D.A., Chapman, C.D., & Roy, E.A. (2000a). Pantomimed actions may be
controlled by the ventral visual stream. Experimental Brain Research, 130, 545-548.
Westwood, D.A., Heath, M., & Roy, E.A. (2000b). The effect of a pictorial illusion on
closed-loop and open-loop prehension. Experimental Brain Research, 134,456-463.
104

References
Westwood, D.A., Dubrowski, A., Carnahan, H., & Roy, B.A. (2000c). The effect of illusory
size on force production when grasping objects. Experimental Brain Research, 135, 535-543.
Westwood, D.A., McEachern, T., & Roy, B.A. (2001). Delayed grasping of a Miiller-Lyer
figure. Experimental Brain Research, 141, 166-173.
Westwood, D.A., & Goodale, M.A. (2003). A haptic size-contrast illusion affects size
perception but not grasping. Experimental Brain Research, 153, 253-259.
Wong, E., & Mack, A. (1981). Saccadic programming and perceived location. Acta
Psychologica, 48, 123-131.
Wraga, M., Creem, S.H., & Proffitt, D.R. (2000). Perception-action dissociations of a
walkab1e Miiller-Lyer configuration. Psychological Science, 11, 239-243.
Yamagishi, N., Anderson, S.J., & Ashida, H. (2001). Evidence for a dissociation between the
perceptual and visuomotor systems in humans. Proceedings of the Royal Society of London
B, 268, 973-977.
Y arbus, A.L. (1967). Eye movements and vision. Plenum Press, New York.
105


Dankwoord
Inleiding
De afgelopen jaren is er ook onderzoek gedaan naar de aanwezigheid van illusies in het AIO-bestaan. Een algemeen aanvaarde opvatting is dat een AIO-baan saai is. Verder wordt vaak verondersteld dat een AIO lange dagen moet maken. In dit experiment is onderzocht of deze aannames illusies zijn. De AIO wordt geacht binnen vier jaar een oordeel te geven over eventuele aanwezigheid van deze illusies.
Methode
Subjects Betrouwbare resultaten kunnen aileen verkregen worden met voldoende proefpersonen; daarom is het aantal AIO's in dit experiment verdubbeld. Procedure Veel pauzes (met koekjes), conferenties en andere "social events" zijn gepland temidden van gezellige college's. Om de dagelijkse bezigheden van de AIO tot een succes te maken wordt tevens de hulp van Eli's kinderen ingeroepen.
Resultaten
Figuur 1 laat zien dat een AIO-baan niet saai hoeft te zijn (p<0.05). Er is tijd voor ontspannende theepauzes, waarin overigens ook over het werk gesproken kan worden. Vanwege de vrijheid om eigen werktijden te bepalen, kan de AIO tijdens zijn meest productieve uren gaan werken, waardoor geen extra lange dagen gemaakt hoeven te worden om te kunnen promoveren. De resultaten van beide AIO's verschillen niet (p>0.83). Figuur 1: Theepauze.
Discussie & conclusie
De begrippen "saai" en "lange werkdagen" komen niet overeen met de werkelijkheid van het AIO-schap en kunnen daarmee als een illusie worden beschouwd. Dankzij goede begeleiding is gebleken dat o.a. artikelen schrijven, programmeren (of iets wat daar op lijkt) en het maken van een poster op een leuke manier geleerd kunnen worden. Daarnaast hebben hulp en (computer)advies van verschillende college's bijgedragen aan het succes van dit experiment. Verder is ook de steun en het vertrouwen van familie en vrienden van essentieel belong geweest. Tot slot werden de welbekende laatste loodjes verlicht door een leuk begin van een post-doc baan.
Referenties Marianne & John, Eli, Jeroen, (voormalige) collega's, Glenn, Jan & Hanny, Jan, Leonie & Harm, Tessa & Arno.
Bedankt!
107


Curriculum vitae
Curriculum Vitae
Denise de Grave werd geboren in Winterswijk op 25 augustus 1977. In 1995 haalde
zij haar VWO diploma aan "De Driemark" te Winterswijk. In datzelfde jaar began zij
met de studie psychologie aan de Katholieke Universiteit Nijmegen met als
afstudeerrichting functieleer. In haar afstudeerscriptie onderzocht zij de aanwezigheid
van helderheidsverschillen in een illusoire kubus. In 2000 began zij haar
promotieonderzoek bij Eli Brenner en Jeroen Smeets op de afdeling
Neurowetenschappen (voormalig Fysiologie) van de Erasmus Universiteit Rotterdam.
Sinds augustus 2004 is zij werkzaam op de afdeling "Allgemeine Psychologie" van de
Justus-Liebig Universiteit te Giessen.
109


Publications
Publications
Articles
Biegstraaten, M., de Grave, D.D.J., Smeets, J.B.J., & Brenner, E. (submitted) Milller-Lyer:
More than just a change in length.
De Grave, D.D.J., Brenner, E., & Smeets, J.B.J. (2002) Are the original Roelofs effect and
the induced Roelofs effect caused by the same shift in straight ahead? Vision Research,
42(19), 2277-2283.
De Grave, D.D.J., Smeets, J.B.J., Brenner, E. (2002) Ecological and constructivist approach
and the influence of illusions. Behavioral and Brain Sciences, 25(1 ), 103-104.
De Grave, D.D.J., Brenner, E., & Smeets, J.B.J. (2004) An apparent compression cannot
explain the difference between the original and induced Roelofs effect. Vision Research,
44(10), 1031-1032.
De Grave, D.D.J., Brenner, E., & Smeets, J.B.J. (2004) Illusions as a tool to study the coding
of pointing movements. Experimental Brain Research, 155(1), 56-62.
De Grave, D.D.J., Biegstraaten, M., Smeets, J.B.J., & Brenner, E. (in press) Effects of the
Ebbinghaus figure on grasping are not due to misjudged size. Experimental Brain Research.
De Grave, D. D. J., Smeets, J. B. J. & Brenner, E (submitted). The coding of saccades studied
with the Brentano illusion.
Smeets, J.B.J., Brenner, E., de Grave, D.D.J., Cuijpers, R.H. (2002) Illusions in action:
consequences of inconsistent processing of spatial attributes. Experimental Brain Research,
147, 135-144.
Smeets, J.B.J., Van Beers, R.J., Van den Dobbelsteen, J., de Grave, D.D.J., & Brenner, E.
(submitted). Optimal sensory integration predicts movement drift.
111

Publications
Refereed abstracts
Biegstraaten, M., De Grave, D.D.J., Smeets, J.B.J., & Brenner, E. (2003). Miiller-Lyer and
grasping: Illusion or obstacles? Proceedings of the 12th International Conference on
Perception and Action (ICP A), 23.
De Grave, D.D.J., Brenner, E., & Smeets J.B.J. (2001). How a simple question can reverse
the Roelofs effect. Investigative Ophtalmology and Visual Science 42:8618.
De Grave, D.D.J., Smeets J.B.J., & Brenner, E. (2001). Gebruik van positie en lengte
informatie in de Brentano illusie. NieuwsbriefNVP, v67, 45.
De Grave, D.D.J., Brenner, E., & Smeets J.B.J. (2002). Pointing towards the Brentano
illusion. Journal ofVision, 2(7), 721.
De Grave, D.D.J., Brenner, E., & Smeets J.B.J. (2002). Misperceived length influences
pointing. Perception (suppl), 31, s87.
De Grave, D.D.J., Biegstraaten, M., Brenner, E., & Smeets, J.B.J. (2003). The Ebbinghaus
figure is more than a size illusion. Proceedings of the 12th International Conference on
Perception and Action (ICPA), 67.
De Grave, D.D.J., Biegstraaten, M., Brenner, E., & Smeets, J.B.J. (2003). The Ebbinghaus
figure is more than a size illusion. Proceedings of the second Helmholtz retreat, 26.
De Grave, D.D.J., Biegstraaten, M., Brenner, E., &.Smeets J.B.J. (2003). De Ebbinghaus
figuur is meer dan alleen een grootte illusie. NieuwsbriefNVP, v34, 33.
De Grave, D.D.J., Biegstraaten, M., Brenner, E., & Smeets, J.B.J. (2004). The Ebbinghaus
figure is more than a size illusion. Journal of Vision, 4(8), 836a.
Puts, M.J.H., de Grave, D.D.J., & de Weert, C.M.M. (2000). Brightness differences in an
illusory cube. Perception (suppl), 29, s9.
112

![Extrinsic grouping factors in motion-induced blindness · In addition, we investigated whether the perception of an illusory Kanizsa triangle [27] is accompanied by a perceptual grouping](https://img.pdfslide.us/doc/110x75/5fccf16407295729ed5d4490/extrinsic-grouping-factors-in-motion-induced-blindness-in-addition-we-investigated.jpg)







![SEPTEIIBI,N 2OO - rrcahistory.orgrrcahistory.org/images/0872.pdf · \.OLU N{]J 1? ALTCUST SEPTEIIBI,N NUX4BER 2OO Chuck smead at the summit of Pike's Peak tance oI the August 13](https://img.pdfslide.us/doc/110x75/5d1ac09588c993e6408ca91f/septeiibin-2oo-olu-nj-1-altcust-septeiibin-nux4ber-2oo-chuck-smead.jpg)



















