Embed Size (px)
Citation preview

ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS Vol. 242, No. 2, November 1, pp. 574-5’78,1985
The Uranic Acid Composition of Anticoagulantly Active and Inactive Heparin’
LOUIS ROSENFELD, STEVEN RADOFF, AND ISIDORE DANISHEFSKY’
Department of Biochemistry, New York Medical College, Valhalla, New York 10595
Received June 27,1985
Bovine heparin was fractionated with respect to chain length and anticoagulant ac- tivity. Analysis of each of these fractions for iduronic and glucuronic acids demonstrated that active heparin has a greater amount of glucuronic acid than inactive heparin. The ratio of the uranic acids in the respective fractions was the same for heparin with different molecular weights. Thus, active heparin with longer chain lengths have more additional glucuronate residues than are required for the antithrombin-binding domain. The results indicate that the active and inactive heparin species differ in more than one structural characteristic and suggest a considerable divergence in their respective bio- synthesis. 0 1985 Academic Press, Inc.
Heparin is a polymer of alternating units of glucosamine and uranic acid (1). The latter is comprised of glucuronic and idu- ronic acids. Although the amount of idu- ronic acid is always greater than that of glucuronic acid, the percentage of iduron- ate in terms of total uranic acid varies from 60 to 87 depending on the source of the heparin (2, 3). The glucuronic acid units appear to be components that have escaped a postpolymerization epimerization step to iduronate that occurs in the biosynthesis of heparin (4).
Heparin can be separated into antico- agulantly active and relatively inactive fractions by affinity chromatography on antithrombin-linked Sepharose (5,6). The affinity of active heparin to antithrombin has been attributed to a segment contain- ing a specific oligosaccharide sequence. Among other features this segment con- tains a glucuronic acid moiety (7,8). With short-chain porcine heparin, M, 7000, there
was an excess of approximately 1 mol gluc- uranic acid per chain. It was suggested that the glucuronic acid involved in the anti- thrombin-binding sequence of active heparin could account for this additional unit (7).
This paper describes results of a study of the glucuronic and iduronic acids content in active and inactive heparin. Utilizing two independent methodologies, it is dem- onstrated that the active heparin contains a higher ratio of glucuronic to iduronic acid. Moreover, the percentage of gluc- uranic acid in both active and inactive hep- arin does not vary with the chain length of the polysaccharide. This leads to the conclusion that high-molecular-weight ac- tive heparin contains more glucuronic acid, when compared to high-molecular-weight inactive heparin, than is required for the reported composition of a single anti- thrombin-binding sequence.
EXPERIMENTAL PROCEDURES
i This research was supported by Grant HL-16955 Materials. Beef lung heparin with an anticoagulant
from the National Institutes of Health. activity of 162 USP units/mg was supplied by Upjohn a To whom correspondence should be addressed. International through the courtesy of Dr. P. W.
0003-9861/85 $3.00 Copyright 0 1985 by Academic Press, Inc. All rights of reproduction in any form reserved.
574

URONIC ACIDS IN HEPARIN 575
O’Connell. Idose, phenyl glucuronide, and ED@ were purchased from Sigma Chemicals. Phenyl iduronide was obtained from Calbiochem-Behring. Sodium bo- rotritide (sp act, 532 mCi/mmol) was the product of New England Nuclear. Amberlite MB-1 (mixed bed resin) and Amerlite IR-120 (H+) were purchased from Mallinkrodt. Glucuronolactone was from K & K Lab- oratories, ptoluenesulfonylchloride was from Aldrich, pyridine was from Eastman, and dimethoxyethane was from Burdick & Jackson Laboratories. Other re- agents were the best available reagent grade.
Tritriated IPId was synthesized from glucurono- la&one by the method of Blanc-Muesser et al. (Cl), ex- cept that 2.5 mCi of sodium borotritide followed by 0.1 g sodium borohydride was used to reduce 0.2 g of 1,2-isopropylidene-5-tosyl-glucuronolactone. The IPId was converted to tritiated L-idose by hydrolysis with 0.05 M H2S04 at 50°C for 8 h (10). Tritiated AnId was prepared from the IPId by hydrolysis with 1.0 M H&SO4 at 100°C for 6 h (10).
Heparinfractions. Heparin was fractionated on the basis of chain length and then on the basis of affinity to an antithrombin-Sepharose column using two methods. For the samples studied by GC, the molec- ular weight species were separated on Bio-Gel P-100 following the method of Pixley and Danishefsky (11). The effluent was divided into five equal-weight frac- tions according to their elution sequence, and the first, third, and fifth fractions were rechromatographed on the gel-filtration column. Portions of each of these pools were further fractionated on antithrombin-se- pharose as described (11). Only the 0.05 and 1.0 M NaCl eluants were used.
The high-molecular-weight fractions (M, 19,500) employed for analyses by reduction and paper chro- matography were obtained by gel filtration of crude heparin on Sephadex G-206 (12). This material was then separated into active (640 units/mg) and inactive (<4 units/mg) fractions on antithrombin-Sepharose as previously described (11). To obtain the completely inactive material it was necessary to chromatograph the nonbinding heparin on the antithrombin-se- pharose a second time.
Analyses. Determinations of the concentrations of heparin were based on analyses for total uranic acid by the carbazole method (13). Analyses for carboxyl- reduced heparin were based on glucosamine assays by the amino acid analyzer (14) or the Elson-Morgan procedure (15). Anticoagulant activities were mea-
’ Abbreviations used: AnId, 1,6-anhydro-L-idopy- ranose; CR, carboxyl-reduced; EDC, 1-ethyl-3-[3-(di- methylamino)propyl]carbodiimide hydrochloride; GC, gas chromatography; HMDS, hexamethyldisilazane; IPId, 5,6-anhydro-1,2-0-isopropylidene-fl-L-idopyra- nose; TMCS trimethylchlorosilane; TMS, trimethyl- silyl.
sured by the effect of heparin in accelerating the neu- tralization of thrombin by antithrombin (16).
Quantitation of uranic acid components. Analyses for iduronic and glucuronic acids in heparin were per- formed by GC using the method of Inoue (3) with a minor modification, i.e., for the preparation of the TMS derivatives, the ratio of pyridine, HMDS, and TMCS (Pierce) was 10:4:2. Determinations were performed on a Perkin-Elmer Model 3920B gas chromatograph equipped with 0.4 X 200-cm glass column of 3.5% SE 30 on Gas Chrom Q (Applied Sciences Labs).
Another procedure for determination of the uranic acids in heparin involved preliminary reduction with sodium borotritide by a modification of the method of Taylor et al. (17). An aqueous solution of heparin, 5 mg/ml, was treated with four times its weight of EDC at pH 4.75. Excess reagent was removed by di- alofiltration using an Amicon PM-10 filter, and the material was concentrated to 10% of the original vol- ume. The pH was adjusted to 8.5 and the EDC-heparin was treated with 500 &i NaB’H,/mg heparin for 2 h at 50°C. Nine volumes of a solution of NaBH, (85 mg/ ml) was added and the mixture was kept for another hour at 50°C. The solution was acidified with glacial acetic acid and dialofiltered exhaustively, and the CR- heparin was precipitated by addition of 4 vol of 5% ethanolic sodium iodide. The preparation was then fractionated on Sephadex G-200. No indication of de- creased molecular weight or contaminating low-mo- lecular-weight radioactivity was observed.
In order to degrade the CR-heparin to obtain the respective hexoses it was first treated with NaNO, (10 mg/mg CR-heparin) at pH 1.0-1.7 at 0°C for 2 h. An equivalent amount of ammonium sulfamate was then added to destroy the excess nitrite, and the mixture was hydrolyzed with 1.0 N HCI for 2 h at 100°C. The mixture was then evaporated to dryness at reduced pressure, and the residue was redissolved in water and treated with 10 mg NaNOa/mg CR-heparin at a pH of 3.2-4.0 for 2 h at room temperature. Residual nitrite was decomposed with an equivalent amount of ammonium sulfamate, and the mixture was sub- jected again to hydrolysis with 1.0 N HCI for 1 h at 100°C. The mixture was taken to dryness, dissolved in water, and passed through an Amberlite MB-1 (mixed bed) resin. The effluent was concentrated and chromatographed on Whatman No. 1 paper, with the upper phase of a mixture of butanol:acetic acid:water, 5:1:4. The standards employed were glucose, idose, and the tritium-labeled idose and AnId synthesized as de- scribed above. The migration positions of the sugars were identified by staining with alkaline AgN03 and by radioactivity counting of individual segments of the paper. Chromatograms were run in isolated lanes. After development each lane was cut into 0.5-in. seg- ments. The segments were extracted for 2 h at 37°C with 1.0 ml of water. Nine milliliters of Liquiscint (National Diagnostics) was then added, and the ra-

576 ROSENFELD. RADOFF, AND DANISHEFSKY
dioactivity was determined with the paper still in the vial.
Additionally, phenyl glucuronide and phenyl idu- ronide were carboxyl-reduced by the procedure de- scribed for heparin, except that a 2.5-fold excess of EDC was employed and the reagent was not removed before reduction. The products were hydrolyzed under the same conditions as for CR-heparin and analyzed by paper chromatography.
RESULTS
The glucuronic and iduronic acid con- tents of different fractions of heparin from beef lung, as obtained by GC, are shown in Table I. The results are expressed as the percentage glucuronic acid. Total uranic acid was determined from the sum of gluc- uronate and iduronate. Statistical analysis of the data indicates that there is a sig- nificant difference between the antico- agulantly active and inactive heparins (P < 0.001). In active or inactive heparin there is no statistical difference in the uranic acid ratios between preparations of different molecular weights. The increased number of glucuronic acid residues in the active heparin as compared with the inactive can be calculated from the data and is also shown in Table I. For example, these cal- culations indicate that the highest-molec- ular-weight fraction of active heparin has about three more glucuronic acids than the
TABLE I
URONIC ACIDS IN HEPARIN FRACTIONS
Percentage giucuronic acid”
Glucuronates Mean M, Activeb Inactive in excess
7,500 17.9 7.8 1.2 14,500 17.9 8.4 2.2 22,500 16.6 9.7 2.5 Mean 17.4 f 0.3” 8.6 f 0.4”
D Percentage of total uranic acids as determined by GC. The percentage of iduronate is the difference be- tween the tabulated values and 100%. Each value is the result of two determinations. Commercial heparin contained 13.8 -C 0.7% (N = 5) glucuronic acid.
b Specific activities for these three fractions were, respectively 366,585, and 619 units/mg.
“SD;N= 6.
inactive heparin of the same molecular weight.
These results were confirmed by means of an alternate method for analyses of uranic acids. A heparin fraction with a mean M, of 19,500 was separated into active and inactive fractions. Both fractions were carboxyl-reduced with sodium borotritide. Acid hydrolyses were performed after deaminative degradations at pH 1.0-1.7 and at pH 3.2-4.0 in order to ensure max- imal release of hexoses. The components from the hydrolyzates were separated by a paper chromatographic system that dif- ferentiates standards of glucose, idose, and AnId (Fig. 1). This analysis revealed that although the mobility of a considerable amount of radioactivity corresponded with that of glucose and idose, the preponderant radioactive component was one which co- chromatographed with AnId (Fig. 2). Sim- ilar findings were observed with a hydroly- zate of carboxyl-reduced phenyl iduronide; however, phenyl glucuronide gives only glucose when treated in the same way. Calculations for the relative amounts of idose were based on the sum of the ra- dioactivities of idose and AnId. Typical chromatographic results for heparin frac- tions are shown in Fig. 2. The results and statistical analysis of numerous such ex- periments are given in Table II, and agree with those found by GC analyses.
DISCUSSION
The results in this paper demonstrate that anticoagulantly active bovine heparin differs from inactive heparin in that it contains a considerably higher content of glucuronic acid. Data for porcine heparin, with a molecular weight of 7000, and het- erodisperse mouse mastocytomal heparin, suggest that this difference occurs with heparins from other species (5,7). The ac- tual proportion of each uranic acid is not the same for heparin preparations from different animal species (2,3). Preparations of porcine heparin have a higher gluc- uranic-to-iduronic acid ratio than bovine material. Moreover, the percentage of glucuronic acid in inactive porcine heparin is greater than that found for the antico-
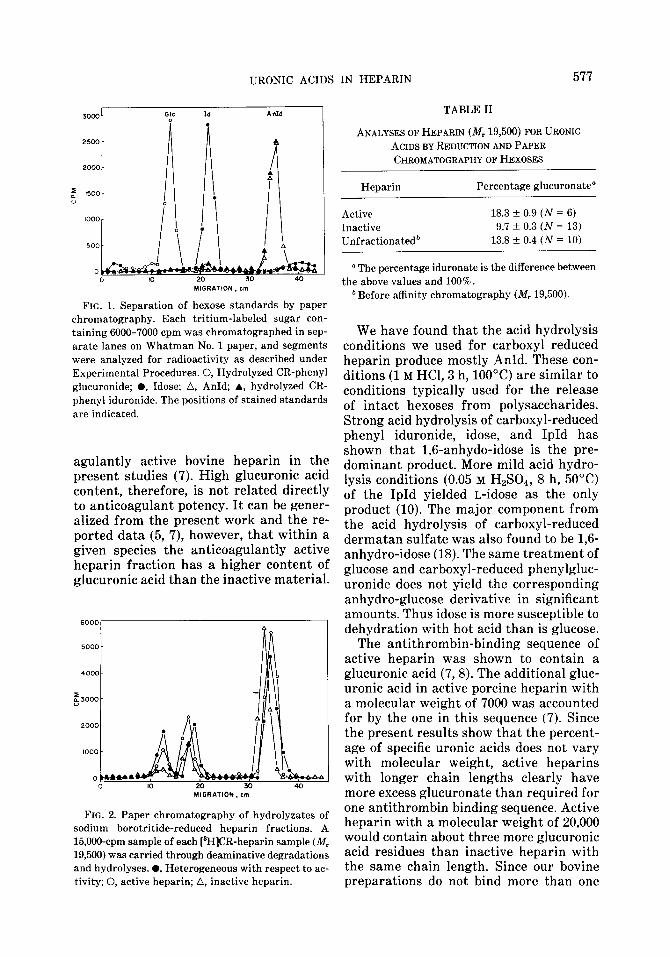
URONIC ACIDS IN HEPARIN 577
2500
2000
r: 1500 ”
IO00
500
0 0 20 JO 40
MIGRATKJN. cm
FIG. 1. Separation of hexose standards by paper chromatography. Each tritium-labeled sugar con- taining 6000-7000 cpm was chromatographed in sep- arate lanes on Whatman No. 1 paper, and segments were analyzed for radioactivity as described under Experimental Procedures. 0, Hydrolyzed CR-phenyl glucuronide; l , Idose; A, AnId; A, hydrolyzed CR- phenyl iduronide. The positions of stained standards are indicated.
agulantly active bovine heparin in the present studies (7). High glucuronic acid content, therefore, is not related directly to anticoagulant potency. It can be gener- alized from the present work and the re- ported data (5, 7), however, that within a given species the anticoagulantly active heparin fraction has a higher content of glucuronic acid than the inactive material.
MIGRATION, cm
FIG. 2. Paper chromatography of hydrolyzates of sodium borotritide-reduced heparin fractions. A 15,000-cpm sample of each [‘H]CR-heparin sample (kf, 19,500) was carried through deaminative degradations and hydrolyses. 0, Heterogeneous with respect to ac- tivity; 0, active heparin; A, inactive heparin.
TABLE II
ANALYSES OF HEPARIN (M, 19,500) FOR URONIC
ACIDS BY REDUCTION AND PAPER CHROMATOGRAPHY OF HEXOSES
Heparin Percentage glucuronate’
Active 18.3 f 0.9 (N = 6)
Inactive 9.7 f 0.3 (N = 13) Unfractionatedb 13.8 +_ 0.4 (iv = 10)
a The percentage iduronate is the difference between the above values and 100%.
b Before affinity chromatography (Mr 19,500).
We have found that the acid hydrolysis conditions we used for carboxyl reduced heparin produce mostly AnId. These con- ditions (1 M HCl, 3 h, 100°C) are similar to conditions typically used for the release of intact hexoses from polysaccharides. Strong acid hydrolysis of carboxyl-reduced phenyl iduronide, idose, and IpId has shown that 1,6-anhydo-idose is the pre- dominant product. More mild acid hydro- lysis conditions (0.05 M HaSOd, 8 h, 50°C) of the IpId yielded L-idose as the only product (10). The major component from the acid hydrolysis of carboxyl-reduced dermatan sulfate was also found to be 1,6- anhydro-idose (18). The same treatment of glucose and carboxyl-reduced phenylgluc- uronide does not yield the corresponding anhydro-glucose derivative in significant amounts. Thus idose is more susceptible to dehydration with hot acid than is glucose.
The antithrombin-binding sequence of active heparin was shown to contain a glucuronic acid (7,8). The additional gluc- uranic acid in active porcine heparin with a molecular weight of 7000 was accounted for by the one in this sequence (7). Since the present results show that the percent- age of specific uranic acids does not vary with molecular weight, active heparins with longer chain lengths clearly have more excess glucuronate than required for one antithrombin binding sequence. Active heparin with a molecular weight of 20,000 would contain about three more glucuronic acid residues than inactive heparin with the same chain length. Since our bovine preparations do not bind more than one

578 ROSENFELD, RADOFF, AND DANISHEFSKY
antithrombin (16), at least some of the ex- cess glucuronates are not involved in an- tithrombin binding. Apparently, in the biosynthetic process, the active heparin undergoes a lesser number of epimeriza- tions than the inactive variety.
It is conceivable that chains with a greater number of glucuronic acid residues are more susceptible to the biosynthetic processes that result in the unique anti- thrombin-binding domain. This would im- ply that much of the synthesis of both ac- tive and inactive heparin is identical, ex- cept that the specific modifications which confer anticoagulant activity occur more often in chains which have had fewer epi- merizations of glucuronic acid. Another possibility is that the two types of heparins are synthesized independently and there- fore exhibit more than one structural vari- ation. We have previously demonstrated that the antithrombin-binding sequence is located at or near the nonreducing termi- nus of the heparin chain (12). It will there- fore be of interest to determine whether the additional glucuronate residues in ac- tive heparin are distributed randomly or clustered at specific locations.
REFERENCES
1. RODEN, L., AND HOROWITZ, M. I. (1978) inThe Gly- conconjugates (Horowitz, M. I., and Pigman, W., eds.), vol. 2, pp. l-85, Academic Press, New York.
2. TAYLOR, R. L., SHIVELY, J. E., CONRAD, H. E., AND CIFONELLI, J. A. (1973) Biochemistry 12,3633- 3637.
3. INOUE, S. (1973) Biochim. Biophys. Acta 329,264- 270.
4. HOOK, M., LINDAHL, U., BACKSTROM, G., MALMS- TROM, A., AND FRANSSON, L.-A. (1974) J. Biol. Chem. 249,3908-3915.
5. HOOK, M., BJORK, I., HOPWOOD, J., AND LINDAHL, U. (1976) FEBS Lett. 66,90-93.
6. ANDERSON, L. O., BARROWCLIFFE, T. W., HOLMER, E., JOHNSON, E. A., AND SIMS, C. E. C. (1976) Thrvmb. Res. 9,575-583.
7. ROSENBERG, R. D., AND LAM, L. (1979) Proc. Natl. Acad. Sci. USA 76,1218-1222.
8. LINDAHL, U., THUNBERG, L., BACKSTROM, G., RIE- SENFELD, J., NORDLING, K., AND BJORK, I. (1984) J. Biol. Chem 259,12368-12376.
9. BLANC-MUESSER, M., DEFAYE, J., HORTON, D.,AND TSAI, J.-H. (1980) Methods Carbohydr. Chem. 3, 177-183.
10. BAGGEIT, N., AND SAMRA, A. (1984) Carbohydr. Rex 127,149-153.
11. PIXLEY, R., AND DANISHEFSKY, I. (1982) Thromb. Res. 26,129-133.
12. RADOFF, S., AND DANISHEFSKY, I. (1984) J. Biol. Chem. 259,166-172.
13. BI’ITER, T., AND MUIR, H. M. (1962) AnaL B&hem. 4,330-334.
14. DOWNS, F., AND PIGMAN, W. (1976) Methods Car- bohydr. Chem. 7.444-448.
15. BOAS, N. F. (1953) J. Biol. Chem. 204,553-563. 16. RADOFF, S., AND DANISHEFSKY, I. (1982) Arch.
B&hem. Biophys. 215,163-170. 17. TAYLOR, R. L., SHIVELY, J. E., AND CONRAD, H. E.
(1976) Methods Carbohydr. Chem. 7,149-151. 18. STOFFYN, P. J., AND JEANLOZ, R. W. (1960) J. Biol.
Chem. 235,2507-2510.





























