Embed Size (px)
Citation preview

Cell Tiss. Res. 148, 57--67 (1974) �9 by Springer-Verlag 1974
The Uhrastructure of the Accessory Sex Organs of the Male Rat
VI. The Ef fec t of Cypro t e rone A c e t a t e on the Dorsa l Lobe
and the Coagu la t ing Gland
Erik Dahl a n d / ~ s m u n d Kjae rhe im
University of Oslo, Departments of Anatomy and Pathology, Dental Faculty, Blindern, Oslo, Norway
Received December 17, 1973
Summary. The dorsal lobe and the coagulating gland of the rat prostate were studied after administration of the anti-androgenic compound Cyp A for varying periods (7-20 days). There was a general involution of the organs with macroscopic atrophy and reduced amount of secretory material. Ultrastructurally the cells were pale, contained less organelles as com- pared with the controls, loss of cytoplasm and reduction both of cell height and width. The major changes also included reduction of the Golgi area, the rough endoplasmic reticulum and the number of free ribosomes. Within the dorsal lobe prominent nuclear changes were found, comprising different types of intranuclear inclusions of membranous configurations. The alterations observed in the present study are similar to the changes which were found in castrated animals and are consistent with atrophy. The results are discussed in relation to the available biochemical and autoradiographic data on the different lobes.
Key words: Prostate - - Rat - - Cyproterone - - Alterations - - Electron microscopy.
Introduction
During the past few years, several ant i -androgenic substances have been syn-
thesized. So far, invest igat ions on the effect of these ant i-androgens have been
confined to bioassay or clinical studies (see N eum ann et al., 1970). Previous
reports f rom this labora tory have defined the fine s t ructure of the accessory glands
of normal and cast ra ted rats, as well as the u l t ras t ruc tura l effects of adminis t ra t ion
of tes tosterone to cas t ra ted and infanti le rats (Dah], Tveter , Kjaerheim, 1973).
Wi th the results obta ined in these invest igat ions as a baseline, i t has been possible
to s tudy the effect of different ant i -androgenic substances. The purpose of the
present invest igat ion was to eva lua te the influence of adminis t ra t ion of the anti-
androgenic compound, cyproterone ace ta te (Cyp A) on the u l t ras t ruc ture of the
dorsal pros ta te and the coagulat ing gland of the rat.
Materials and Methods
Eleven adult albino rats (Charles River CDF strain) 4-6 months old were used. Four rats were given 5 mg Cyp A as subcutaneous injections every second day over a period of 5 days {total 15 mg). Four rats received 15 mg Cyp A as subcutaneous injections daily for 18 days (total 270 mg). The remaining three animals, used as controls, received 1 ml of the solvent solution as subcutaneous injections over a period of 18 days. All the animals were killed two days after the last injection. While in nembutal and aether anaesthesia, the aorta was can~ulated, and intraaortic fixation perfusion was carried out as described elsewhere (Dahl,

58 E. Dahl and •. Kjaerheim
Fig. 1. Survey electron micrograph of the dorsal lobe seen after Cyp-A t rea tment for 18 days. There was a general reduction of the cytoplasm, Golgi area (G) and the rough endoplasmic reticulum (RER). Dense bodies (Db) were found in an increased number. Different types of intranuclear alterations were seen adjacent to the nueleolus, either as pale areas (arrows central nucleus) or as electron-opaque fibrillar areas (arrows right nucleus). Note the small blebbings (Bl) at the cellular surface. Lu aeinar lumen, Bm basement membrane, N nucleus, BC basal
cell. • 8800

Effect of Cyproterone on Accessory Sex Organs 59
Kjaerheim, Tveter, 1973). The dissection of the prostate, the preparation of the samples and the electron microscopic techniques were according to the principles already reported (Dahl et al., 1973).
Observations
Injections of Cyp A subcutaneously for 18 days did not seem to influence the general condition of the animals. At the time of sacrifice, all the animals appeared to be in excellent condition. While no definite reduction in the gross anatomy of the prostatic complex was seen after treatment with cyproterone for 7 days, a general involution, with macroscopic atrophy and reduced amount of secretory material was prominent after 20 days. Although there were some differences between the various organs as with regards to the effect of Cyp A, several common ultrastructural features were prominent. Generally the cells were pale, contained less organellcs as compared with the controls, with reduction both of the cell height and width, and loss of cytoplasm (Figs. 1, 14, 15). Furthermore, the major changes also included reduction of the Golgi area and the rough endoplasmic reticulum (RER), in addition to a reduction of the number of free ribosomes. However, it should be stressed that quantitative variations were observed, both from cell to cell within the separate lobe, and also within the different organs. This was re- flected by some differences in the size and the amount of organelles of the epi- thelial cells--obhquity of the sections taken in consideration. Except for primordial intranuclear alterations, and also some questionable variations of the RER and the blebbing at the surface of the epithelial cells of the dorsal lobe, no convincing changes were observed after 7 days. After 20 days, however, the following altera- tions were found:
The Dorsal Lobe
The Golgi Area. Generally this structure was markedly reduced in size (Fig. 1). In some cells, only remnants were found. In other cells, however, a relatively well developed Golgi apparatus was still encountered (Fig. 2).
The Endoplasmic Reticulum. The I~ER cisternae were reduced considerably, both in number and size (Fig. 1). In addition, there also seemed to be a numerical reduction of free ribosomes. Large dilated cisternae, which regularly were seen in the normals, were never encountered. Whorl formations, which consisted of concentric, lamellated configurations of RER profiles were a common feature (Figs. 2, 3). Lysosomes and vacuoles were found in the central portion of these characteristic profiles (Figs. 2, 3).
Autophagic Vacuoles and Dense Bodies. Autophagic vacuoles of different sizes were infrequently found. They contained vesicles, dense bodies, membranes and other cytoplasmic debris (Fig. 4). Dense bodies of the lysosome-series were regu- larly found in an increased number (Figs. 2, 4).
Fig. 2. Whorl formations (WF) of the RER were regularly encountered. They were built up of concentric lamellated configurations and characterized by centrally located dense bodies (arrows), collapsed cisternae and vacuoles. Cyp-A treated for 18 days. N nucleus, G Golgi area,
with empty vacuoles, but of an almost normal size. x 18000

60 E. Dahl and A. Kjaerheim
Fig. 3. High magnification of a whorl formation of RER with collapsed cisternae in the juxta- nuclear region. Dense bodies (Db) of different sizes and cisternae with a ra ther l ight content (arrow) were regularly encountered. (Dorsal lobe, 18 days Cyp-A treated). N nucleus, x 30000
Fig. 4. A large autophagic vacuole located in the Golgi area. Small membrane-bound vesicles within the large vacuole were found, as well as dense bodies, membrane fragments and other cytoplasmic debris. N nucleus, Nu nucleolus. Note the autophagic vacuole does not exceed the uni t membrane of the neighbouring cells (arrows). (Dorsal bole, 18 days Cyp-A treated).
X 12 500

Effect of Cyproterone on Accessory Sex Organs 61
Blebbin9. At the surface of the cells, blebbing was less pronounced than in the normals, and also markedly reduced in size by the experimental period (Fig. 1).
Nuclear Inclusions. A characteristic feature of the dorsal lobe was the appear- ance of different types of intranuclear alterations which were regularly encountered (Figs. 1,5-13). They were of various sizes, usually comprising different forms of inclusions of membranous configurations. Some of them were composed of con- centric lamellar membranes {Figs. 10-13) while others were surrounded by a smooth, single membrane and contained rather opaque (fibrillar) material, part ly surrounded by heterochromatine (Figs. 1, 9). Vacuoles with a rather flocculent matrix were also found (Figs. 8, 13). Finally, circular, pale areas without any specific demarcation from the regular content of the nucleus were also rather often observed {Figs. 1, 8).
The Coagulating Gland
The Golgi Area. The whole Golgi apparatus was markedly reduced in size. I t occupied only a small part of the apical, juxta-nuclear region of the cell. Granu- lar, secretory material was not encountered within the Golgi area (Fig. 14).
The Endoplasmic Reticulum. There was a marked reduction of the R E R cisternae, R E R profiles and also a reduction of the number of free ribosomes (Figs. 14, 15). Even dilated and rather larger cisternae still were found, they never reached the enormous sizes which were a pathognomonic feature in the normals. Furthermore, the content of the cisternae appeared more electron dense than in the normals (Fig. 15).
i Autophagic Vacuoles and Dense Bodies. Autophagic vacuoles were only occasion- ally encountered. They were rather small of size. More often circular membranes, suggesting segregation of the epithelial cytoplasm were found (Fig. 14). Dense bodies were observed in an increased number, but never to the similar extent as in the dorsal prostate.
Intranuclear Inclusions. Intranuclear inclusions of the similar morphology as found in the dorsal prostate were never encountered. On the other side, inclusions of the nuclear body series were regularly observed (Fig. 16). Some of these con- tained a rather coarse, dense, granular material (Fig. 17).
Discussion
In the present study, the effect of Cyp A is reflected by macroscopic involution of the prostatic complex and reduced amount of secretory material. Ultrastructur- ally there are alterations in the fine structure as compared with the control animals with reduction both in the cell height and width, as well as quanti tat ive and qualitative changes of the organelles of the cells. When the present results are compared with previous studies (Flickinger, 1971 ; Dahl et al., 1973), it is evident that Cyp A is capable of inducing changes of the fine structure of the dorsal prostate and the coagulating gland. Furthermore, this alterations are similar to the changes which are found after castration (Brandes, GySrkey, Groth, 1962; Itelminen and Ericsson, 1971 ; Dahl and Kjaerheim, 1973 ; Dahl and Tveter, 1973) and seem to be compatible with the well documented anti-androgenic properties

62 E. Dahl and A. Kjaerheim
Fig. 5. Survey micrograph of a nucleus of a epithelial cell from the dorsal lobe, demonstrat ing different kinds of intranuclear alterations: A fine granular area without any membranous demarcation from the rest of the nuclear chromatin is seen to the left (arrows). Fibril lar material (Era) was found in increased amount in the area of the suggested original nucleolus. Nuclear body-like inclusions (open arrow). Nu fragments of the nucleolus. Cyp-A. 18 days.
• 17600 Fig. 6. Higher magnification of a par t of the nucleolar region in fig. 5 demonstrat ing the fibrillar material which part ly may have an amorphous appearance. Note the remnants of a single membrane (arrow) and the difference between the fibrillar and the granular material
within the nucleolus (Nu). Cyp-A 18 days. • 60000

Effect of Cyproterone on Accessory Sex Organs 63
of this drug (Neumann et al., 1970). I t may therefore be concluded that the admini- stration of Cyp A to experimental animals exerts an inhibitory effect on the pro- static complex consistent with atrophy.
The origin of the intranuclear inclusions in the dorsal lobe is unclear. The double membrane-bound structures containing a cytoplasmic matrix strongly suggest tha t a portion of the cytoplasm has been incorporated into the nucleus. However, the pale areas and the inclusions which are part ly surrounded by a single membrane may suggest tha t some of the intranuclear alterations may be intermediate stages of some of the inclusions observed, and formed de novo within the nucleus. The inclusions which are observed in the dorsal lobe in the present study are morpho- logically quite different from the structures usually termed nuclear bodies (Krishan, Uzman and Hedley-Whyte, 1967 ; Dahl, 1970), and seem to be without any morpho- logical connection with this structure.
The functional significance of the intranuclear alterations is obscure. There are at least two possible hypotheses to explain their origin (Nakayama and Nicherson, 1972; Nickerson, 1973). The first is that they arise as a response to degenerative changes, while the second alternative possibility is that they reflect a general stimulation of the cell. That the present inclusions found in the dorsal lobe most likely represent involutionary changes is supported by the observations that Cyp A induces a general a t rophy of the cells with reduced synthesis of protein and secre- tory material.
Correlation o/ Structure and Function. Autoradiographic and subcellular fractionation studies have indicated that androgen is selectively retained by the nuclei of the epithelial cells of the prostate (Tveter, and Attramadal, 1969; Sar, Liao and Stumpf, 1970; Tveter, 1970). This selective retention of androgen is due to the interaction of the hormone with specific hormonal binding sites, or receptors. Anti-androgenic compounds, such as Cyp A, are able to depress the uptake of testosterone in the prostate. Furthermore, they will significantly reduce the binding of androgen to the hormonal receptors by competing for the same cellular binding sites (Stern and Eisenfeld, 1969; Fang and Liao, 1969 ; Tveter and Aakvaag, 1969). If the association of androgen with these receptors is related to the mode of action of androgenic hormones, the reduced binding of the hormones caused by anti- androgens may be intimately connected with their biological effects. Since androgen is associated with receptors both in the cytoplasm and in the nuclei, t reatment with
Fig. 7. Detail of the nuclear body-like inclusions made of a fibrillar core surrounded by a granular periphera] ]ayer. Note the membrane-bound structure (arrow) most like]y representing
cytoplasmic material. Cyp-A 18 days. • 60000
Fig. 8. Intranuclear inclusions comprising configurations of several vacuoles (arrows). The matrix of the vacuoles is floccuIent and rather lightly electron-opaque. Some of the vacuoles
are partly surrounded by fragments of the nucleolus (Nu). Cyp-A 18 days. • 24000
Fig. 9. Intranuclear inclusions of a vacuolar fibrillar type (Fro) which occupies a large portion of the central part of the nucleus (cf. Fig. 1). The vacuole is surrounded by a single membrane (arrows) which in some places is disrupted (open arrow). N nuclear chromatin. Nu fragments
of the original nucleolus. Cyp-A 18 days. • 21000
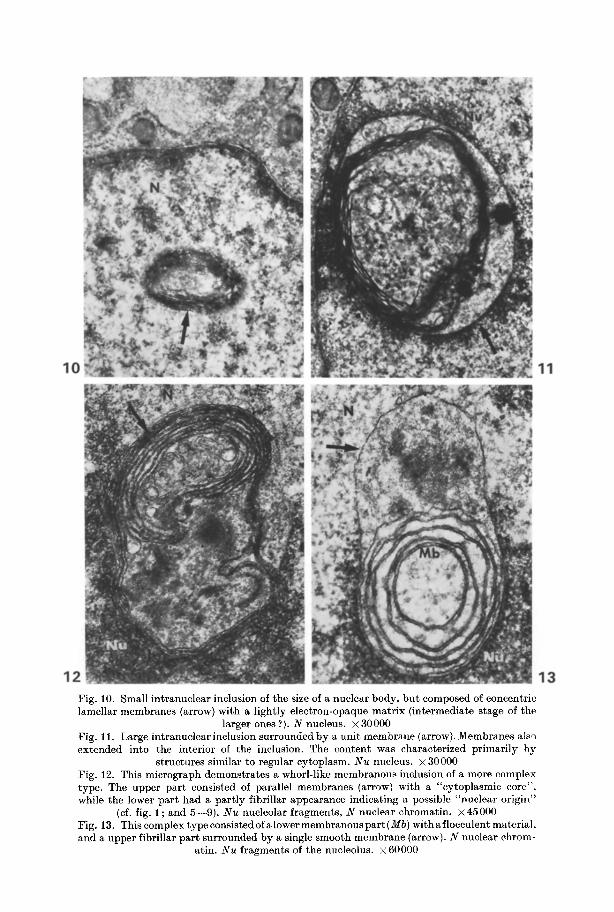
Fig. 10. Small intranuclear inclusion of the size of a nuclear body, bu t composed of concentric lamellar membranes (arrow) with a lightly electron-opaque matr ix (intermediate stage of the
larger ones ?). N nucleus. • 30000 Fig. l i . Large intranuclearinclusion surrounded by a unit membrane (arrow). Membranes also extended into the interior of the inclusion. The content was characterized primarily by
structures similar to regular cytoplasm. N u nucleus. • 30000 Fig. 12. This micrograph demonstrates a whorl-like membranous inclusion of a more complex type. The upper par t consisted of parallel membranes (arrow) with a "cytoplasmic core", while the lower par t had a part ly fibrillar appearance indicating a possible "nuclear origin"
(cf. fig. 1; and 5--9). N u nucleolar fragments, N nuclear chromatin, x45000 Fig. 13. This complex type consistedof alower membranouspar t (Mb) withaf locculent material, and a upper fibrillar par t surrounded by a single smooth membrane (arrow). N nuclear chrom-
atin. Nu fragments of the nucleolus, x 60000
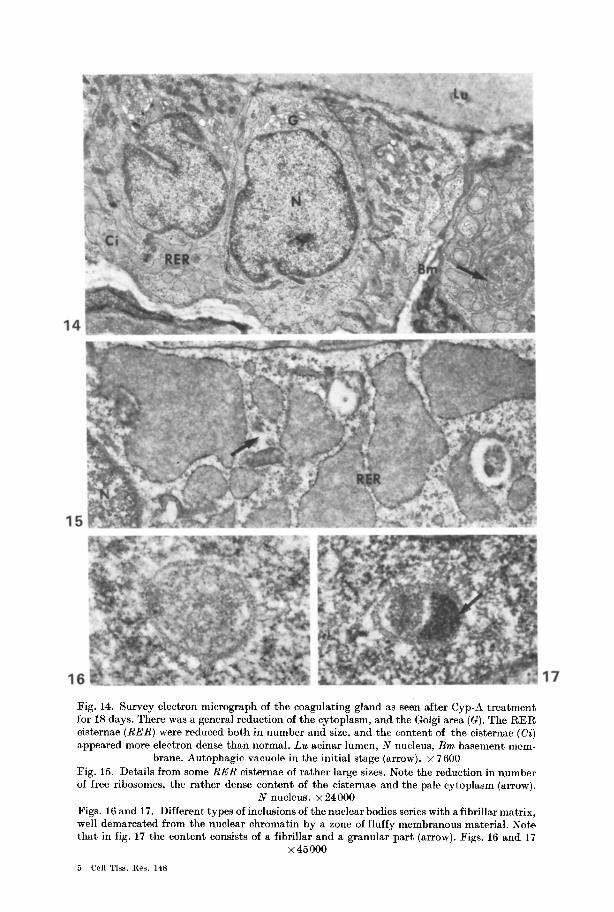
Fig. 14. Survey electron mierograph of the coagulating gland as seen after Cyp-A treatment for 18 days. There was a general reduction of the cytoplasm, and the Golgi area (G). The RER cisternae (RER) were reduced both in number and size, and the content of the cisteraae (Ci) appeared more electron dense than normal. Lu acinar lumen, N nucleus, Bm basement mem-
brane. Autophagic vacuole in the initial stage (arrow). • 7600 Fig. 15. Details from some RER cisternae of rather large sizes. Note the reduction in number of free ribosomes, the rather dense content of the cisternae and the pale cytoplasm (arrow).
N nucleus. • 24000 Figs. 16 and 17. Different types of inclusions of the nuclear bodies series with a fibrillar matrix, well demarcated from the nuclear chromatin by a zone of fluffy membranous material. Note that in fig. 17 the content consists of a fibrillar and a granular part (arrow). Figs. 16 and 17
• 45 000
5 Cell Tiss. Kes. 148

66 E. Dahl and A. Kjaerheim
ant i -androgens may directly influence the funct ion of the nuclei as well as of the cytoplasm. The present s tudy has demonst ra ted prominent changes, both within the cytoplasmic organelles as well as wi thin the nuclei. I t is now generally accepted t ha t the nucleus is the pr imary site of the action of androgen. The nuclear steroid- receptor complex is located in the genome (Liao et al., 1965) and is probably directly involved in the RNA synthesis at the t ransscript ional level. The specific effect on RNAs thus formed, will in t u r n regulate the synthesis of s tructural , enzymat ic and secretory proteins. Cyp A seems to inhibi t the binding of androgen to the chromat in and thus prevent RNA formation. Some of the dist inct nuclear al terat ions seen in the present invest igat ion ma y therefore represent the histo- logical correlation to these biochemical effects of Cyp A.
References
Brandes, D., Gy6rkey, F., Groth, D.: Fine structural and histochemical study of the effect of castration on the rat prostatic complex. Lab. Invest. 11, 339-349 (1962)
Dahl, E.: The fine structure of nuclear inclusions. J. Anat. (Lond.) 106, 255-262 (1970) Dahl, E., Kjaerheim, A.: The ultrastructure of the accessory sex organs of the male rat.
II. The postcastration of the ventral, lateral and the dorsal prostate. Z. Zellforsch. 144, 167-178 (1973)
Dah], E., Kjaerheim, A., Tveter, K. J.: The ultrastructure of the accessory sex organs of the male rat. I. Normal structure. Z. Zellforsch. 137, 345-359 (1973)
Bah], E., Tveter, K. J.: The ultrastructure of the accessory sex organs of the male rat. III. The post-castration involution of the coagulating gland and the seminal vesicle. Z. Zellforseh. 144, 179-189 (1973)
Dahl, E., Tveter, K. J., Kjaerheim, A.: The ultrastructure of the accessory sex organs of the male rat. 4. Effect of testosterone. Aeta endoer. (Kbh.) 74, 605-614 (1973)
Fang, S., Liao, S.: Antagonistic action of anti-androgens on the formation of a specific di- hydrotestosterone-receptor protein complex in rat ventral prostate. Molee. Pharmacol. 5, 428431 (1969)
Flickinger, C. J.: Ultrastructural observations on the postnatal development of the rat prostate. Z. Zellforsch. 113, 157-173 (1971)
Helminen, H. J., ~riesson, J. L.E.: Ultrastructural studies on prostatic involution in the rat. Mechanism of autophagy in epithelial cells, with special reference to the rough-surfaced endoplasmic reticulum. J. Ultrastruct. l~es. 36, 708-724 (1971)
Krishan, A., Uzman, B. G., Hedley-Whyte, E. T.: Nuclear bodies: a component of cell nuclei in hamster tissues and human tumors. J. Ultrastruct. ges. 19, 563-572 (1967)
Liao, S., Leininger, K. R., Sagher, D., Barton, 1~. W.: Rapid effect of testosterone on ribo- nucleic acid polymerase activity of rat ventral prostate. Endocrinology 77, 763-765 (1965)
Nakayama, I., Nickerson, P.A.: Intranuclear inclusions in mammotrophs of the female mongolian gerbil. Amer. J. Anat. 135, 93-104 (1972)
Neumaml, F. 1~., yon Bcrswordt-Wallrabe, W., Elger, H., Steinbeck, J. D., Hahn, ]~I., Kramer, I.: Aspects of androgen-dependent events as studied by antiandrogens. In: l~ecent progress in hormone research, vo]. 26 (E. B. Astwood, ed.), p. 337. New York: Academic Press 1970
Nickerson, P. A.: Induction of intranuclear inclusions by estrogen in mammotrophs of the mongolian gerbil. J. Ultrastruct. P~es. 44, 41~18 (1973)
Sar, M., Liao, S., Stumpf, W. E.: Nuclear concentration of androgens in rat seminal vesicles and prostate demonstrated by dry-mount autoradiography. Endocrinology 86, 1008-1011 (1970)
Stem, J. M., Eisenfeld, A J.: Androgen accumulation and binding to macromolecules seminal vesicles: Inhibition by cyproterone. Science 166, 233-235 (1969)
Tveter, K. J.: An autoradiographic study on the localization of androgen on the prostate gland and the seminal vesicles of the male rat. Acta endocr. (Kbh.) 63, 207-215 (1970)

Effect of Cyproterone on Accessory Sex Organs 67
Tveter, K. J., Aakvaag, A.: Uptake and metabolism in vivo of testosterone-l,2-aH by accessory sex organs of male rats; influence os some hormonal compounds. Endocrinology 85, 683- 689 (1969)
Tveter, K. J., Attramadal, A.: Autoradiographic localization of androgen in the rat ventral prostate. Endocrinology 85, 350-354 (1969)
Dr. Erik Dahl University of Oslo Department of Anatomy Dental Faculty P.O. Box 1052 Blindern, Oslo Norway
5*