Embed Size (px)
Citation preview

163
Molecular and Cellular Biochemistry 232: 163–167, 2002.© 2002 Kluwer Academic Publishers. Printed in the Netherlands.
The transcription co-repressor TLE1 Interactedwith the intracellular region of gp130 through itsQ domain
Fei Liu, Yin Liu, Demin Li, Yong Zhu, Weiming Ouyang, Xin Xie andBoquan JinDepartment of Immunology, 4th Military Medical University, Xi’an City, Shannxi Province, P.R. China
Received 20 August 2001; accepted 20 November 2001
Abstract
As the common signal transducer for IL-6 family cytokines, gp130 interacts with various signal molecules. Our previous workfound the amino-terminal enhancer of split (AES) molecule interacts with gp130 intracellular region through its conservedglutamine-rich (Q) domain. The Q domain in AES shares high homology with those in the transcription co-repressor transducin-like enhancer of split (TLE) proteins. The yeast two-hybrid assay, gluthione S-transferase fusion protein pull-down assay andimmuno-co-precipitation assay indicated that the Q domain of TLE1 is capable of binding gp130 intracellular domain, and theintracellular membrane proximal region of gp130 containing conserved Box1 and Box2 motifs seemed essential for this inter-action. The interaction between gp130 and TLE1 indicated that molecules of TLE family might play a role in gp130 signaling.(Mol Cell Biochem 232: 163–167, 2002)
Key words: gp130, TLE1, protein interaction, signal transduction, Q domain
Abbreviations: AES – amino-terminal enhancer of split; Gro – groucho; FBS – fetal bovine serum; MAb – monoclonal anti-body; SDS-PAGE – sodium dodecyl sulfate polyacrylamide gel electrophoresis; TBS – Tris-buffered saline; TLE – transducin-like enhancer of split; X-gal – 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside
Short communication
Introduction
As the common signal transducer for IL-6, IL-11, LIF (leu-kemia inhibitory factor), CNTF (ciliary neurotrophic factor),CT-1 (cardiotrophin-1) and OSM (oncostain M) receptors,gp130 plays important roles in embryo development, haemat-opoiesis and immunoreaction (reviewed in ref. [1]). It hasbeen proved that gp130 mediated signal transduction by bothRas-MAPK pathway and JAK-STAT pathway. But whetherother unknown signal pathway or molecules are involved ingp130 signaling is not clear. Previous study indicated thathuman AES (amino enhancer of split) associated with thecytoplasmic domain of human gp130 [2]. AES contains aconserved glutamine-rich (Q) domain at the N-terminus and
a unique C-terminus region. As a wildly expressed protein,its function is largely disputable or unclear [3–5]. AES wasproved to interact with cytoplasmic membrane proximal partof gp130, but was not phosphorylated after IL-6 stimulation.Co-expression of AES, gp130 and JAK kinase in COS7 cellsinhibited the association of JAK kinase with gp130, and de-creased the tyrosine phosphorylation level of gp130 and JAKkinase [6]. These results indicate AES might play a role ingp130 signaling.
Recently we have found that AES interacted with gp130through its Q domain by yeast two-hybrid assay (unpublisheddata). The transcription co-repressor groucho/TLE (transduc-ing-like enhancer of split) molecules also contain the con-served Q domain in their N-terminus [7, 8]. Groucho is a
Address for offprints: B. Jin, Department of Immunology, 4th Military Medical University, Xi’an City, Shannxi Province, P.R. China 710032(E-mail: [email protected])
164
Drosophila molecule and TLEs are its mammalian homo-logues; they can not bind DNA directly, but can interact withsome DNA-binding proteins or transcription factors, such asHES (hairy enhancer of split), LEF-1/TCF (lymphoid en-hancer factor-1/T-cell factor), PRDI-BF1/Blimp-1 and HNF3(hepatic nuclear factor 3), and mediate transcription inhibi-tion of the corresponding target genes (reviewed in ref. [9]).Now four human TLE molecules, TLE1~ 4, were cloned,among them TLE1’s function is studied most widely, so wefirstly investigated if TLE1 could also interact with the in-tracellular region of gp130 through its Q domain.
Materials and methods
Vector construction
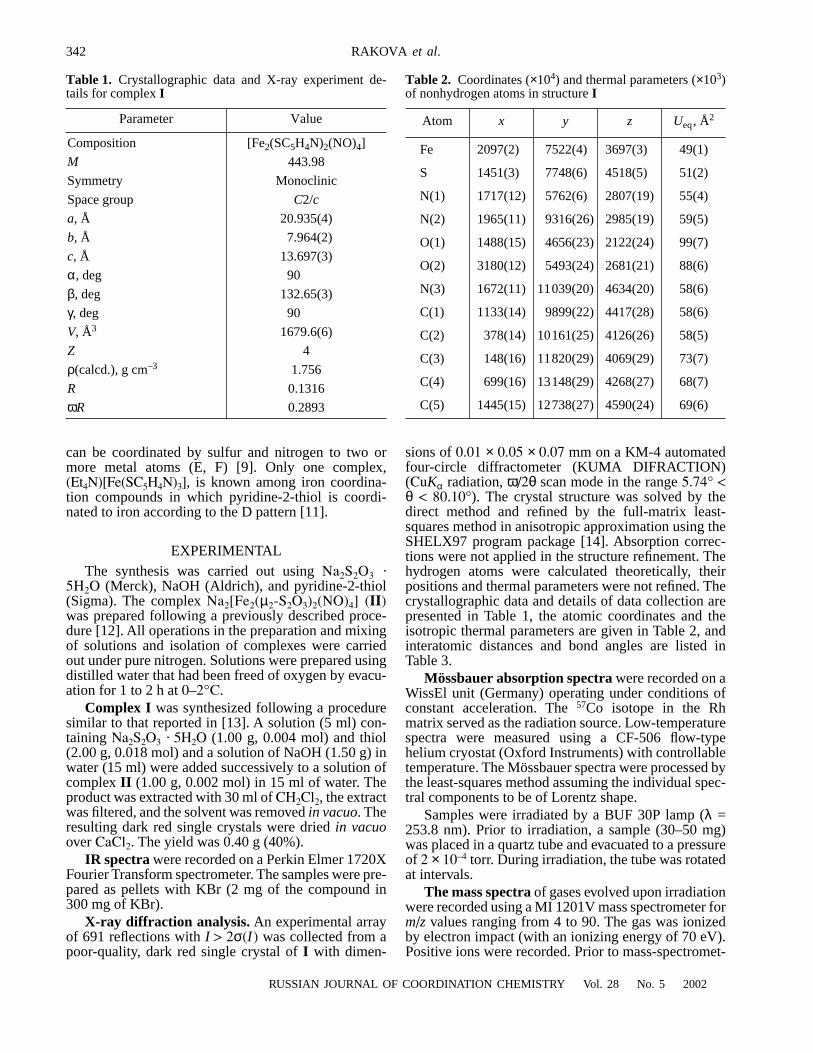
GST-fusion protein expression vectors pGEX1N-TLE1 (33–770 aa) and pGEX1N-HES1 (3–281 aa) [10] are kind giftsfrom Dr. Stefano Stifani of Montreal McGill University,Canada, pGEX2T-gp130IC (intracellular region of gp130,642–918 aa) [19] was a kindly gift from Dr. Tetsuya Taga ofTokyo Medical and Dental University. The cDNA fragmentsencoding membrane-proximal part or membrane-distal partof gp130 intracellular region and N-terminal or C-terminalregion of TLE1 were obtained by PCR amplification of corre-sponding cDNAs in this 2 vectors with primers contain-ing restriction enzyme sites. The primers were designedby Primer3 Program on http://www-genome.wi.mit.edu/cgi-bin/primer/primer3.cgi and synthesized by Sangon Inc.,Shanghai, China. The primers for gp130 were: left #1 GCG-AATTCAA TAAGCGAGACCTAA; right #1 GCGTCGAC-AGTGTTTTGTGAAGA; left #2 GCG-AAT-TCTCACAAAACACTTCGAGC; right #2 GCGTCGACTCACTGAGGC ATGTAGC.The primers for TLE1 were: left #1 GCGAATTCCAGTTCC-TGCAGG CG; right #1 GCGTCGACTTTGGCACGTTC-AAC; left #2 GCGAATTCCGGGAAA ATGGAATCG; right#2 CGGGATCCTAGCCTTCTTGTCCC. The full length ofTLE1 cDNA was directly digested from pGEX1N-TLE1 withEcoR I. Then these cDNA fragments were inserted into theyeast two-hybrid vectors pGAD424 (encoding the GAL4transcription activation domain, abbreviated to GAL4ad,Clontech Inc.), pGBT9 (encoding the GAL4 DNA-bindingdomain, abbreviated to GAL4bd) or GST-fusion protein ex-pression vector pGEX4T (Amersham-Pharmacia Inc.). Se-quences of these cDNA fragments were corroborated by DNAsequencing (auto-sequencer Type 310, ABI Inc.). The gp130constructs encode proteins containing amino acids 642–753,751–918 or 642–918 of human gp130 fused to the GAL4bdor GST respectively; the TLE1 constructs encode proteinscontaining amino acids 33–770, 269–770 or 33–112 of hu-man TLE1 fused to the GAL4ad (Fig. 1).
Yeast two-hybrid assay
The constructed pGAD424 and pGBT9 vectors were pairedas showed in Table 1, and co-transfected the yeast stainSFY526 following the protocol provided by Clontech Inc.The transformed yeast colonies were selected by syntheticdropout medium lacking tryptophan and leucine. The presentof gp130 and TLE1 cDNAs in the survived colonies wereverified by PCR with corresponding primers. Then the ac-tivities of the reporter gene β-galactosidase in the transformedcolonies were detected by colony-lift filter following theprotocol of Clontech Inc.
Preparation of GST-TLE1/gp130 fusion proteins and anti-serums
GST, all three GST-gp130IC fusion proteins, GST-TLE1 (33–770aa) and GST-HES1 (3–281aa) were expressed in E. Coli
Table 1. Interaction between TLE1 and intracellular region of gp130 in S.cerevisiae
Transformation β-Galactosidaseplasmid activity
GAL4 ad-TLE1 (33–770aa) + GAL4 bd-gp130 (642–918aa) +GAL4 ad-TLE1 (33–770aa) + GAL4 bd-gp130 (642–753aa) +GAL4 ad-TLE1 (33–770aa) + GAL4 bd-gp130 (751–918aa) –GAL4 ad-TLE1 (269–770aa) + GAL4 bd-gp130 (642–918aa) –GAL4 ad-TLE1 (33–112aa) + GAL4 bd-gp130 (642–918aa) +GAL4 ad-TLE1 (33–112aa) + GAL4 bd-gp130 (642–753aa) +GAL4 ad + GAL4 bd –
β-Galactosidase positive S. cerevisiae colonies appeared blue on the filterwithin 8 h, which indicating the products of co-transformed genes are ableto interact.
TLE1
33 -770
GP S/P CcN WD40
642 -918
gp130 IC (Intracellular region)
Box 1 2 3
Q
642 -753
751 -918
33 -112
269-
B GAL4bd TLE1 Constructs
A GAL4ad/GST gp130 Constructs
TLE1
33 -770aa
GP S/P CcN WD40
642 -918aa
gp130 IC (Intracellular region)
Box 1 2 3
Q
642 -753aa
751 -918aa
33 -112aa
- 770aa
B GAL4bd TLE1 Constructs
A GAL4ad/GST gp130 Constructs
TLE1
33 -770
GP S/P CcN WD40
642 -918
gp130 IC (Intracellular region)
Box 1 2 3
Q
642 -753
751 -918
33 -112
269-
B GAL4bd TLE1 Constructs
A GAL4ad/GST gp130 Constructs
TLE1
33 -770aa
GP S/P CcN WD40
642 -918aa
gp130 IC (Intracellular region)
Box 1 2 3
Q
642 -753aa
751 -918aa
33 -112aa
- 770aa
B GAL4bd TLE1 Constructs
A GAL4ad/GST gp130 Constructs
Fig. 1. The fragments of gp130 and TLE1 proteins inserted into the yeasttwo hybrid vectors.
-
-

165
strain JM109 and purified by glutathione-sepharose4B (Am-ersham-Pharmacia Inc.) affinity chromatography. With thepurified GST-gp130IC and GST-TLE1 fusion protein, therabbit anti-gp130 and anti-TLE1 serums were prepared fol-lowing conventional protocol. Briefly, each rabbit was firstlyimmunized subcutaneously with 1 mg purified GST fusionprotein or 0.5 mg purified GST mixed with complete Fre-und’s adjuvant (GibcoBRL), then boosted subcutaneously withidentical protein mixed with incomplete Freund’s adjuvant 3weeks later, once two weeks for 3 times; 1 week after the lastimmunization, the serums were harvested. Purified GST wascoupled with pre-activated sepharose4B (Amersham-Phar-macia Inc), then the anti-GST antibodies in the anti-serumswere absorbed by the GST-sepharose4B beads, and the spe-cificity of the anti-serums was evaluated by indirect ELISA.
Preparation of HeLa cell protein extract and GST pull-down assay
HeLa cell proteins extract was used to provide nature TLE1for it was proved containing a great amount of TLE1 [10].HeLa cells cultured in RPMI1640 containing 10% FBS wereharvested and homogenized in ice-cold buffer A (30 mMHepes, 60 mM KCl, 30 mM NaCl, 1 mM EDTA, 200 mM su-crose, 20 mM iodoacetamide, 1 mM PMSF, 2 µM leupeptin,2.5 µg/ml antipain, pH 7.6) by trituration through 6# needles,and lysed in the presence of 1% Triton X-100. Lysates werecentrifuged at 12,000 g for 15 min and the supernatant wasrecovered and used for the following assay. HeLa cell pro-tein extract (approximately 100 µg of total protein) wasincubated with each gp130IC-GST fusion protein or GST-HES1 (approximately 25 µg each) in 4°C for 2 h, then glu-tathione-sepharose4B beads were added and the incubationwent on for 1 h in 4°C. The beads were collected by briefcentrifugation and washed 5 times with Tris-buffed saline(TBS) and immediately incubated in 2X SDS-PAGE loadingbuffer. After SDS-PAGE and transfer to PVDF membrane,Western blotting with anti-TLE1 serum and BM Chemilu-minescence Kit (Boehringer Manneheim) was performed.Molecular size was indicated by RainbowTM colorful proteinmarker (200, 100, 68, 43, 30 kDa, Amersham-Pharmacia Inc.)on PVDF membrane.
Gene transfection and immuno-precipitation assay
Eukaryocyte expression vector pEF-BOS [20] containingfull-length cDNA of gp130 (pEF-BOS-gp130) and anti-gp130 MAb AM64 [19] are also kindly gifts from Dr TetsuyaTaga. HeLa cells was transfected with pEF-BOS-gp130 byDEAE-Dextran method; and harvested 48 h later, then lysedby ultrasonic to prepare whole cell extract. After pre-clean-
ing with Pansonin (Roche), the extract was incubated withanti-TLE1 serum, anti-gp130 serum, anti-GST serum or pre-immunized rabbit serum overnight at 4°C respectively. Af-ter washing with TBS for 3 times, the mixture of proteinA-agarose and protein G-agarose were added and incubatedfor 3 h in 4°C. Then the agarose beads were collected andwashed, the SDS-PAGE and Western blot assay of the com-plex bound with the beads were preformed, and the proteinswere blotted either with anti-TLE1 MAb 5F9 [21] or anti-gp130 MAb AM64, followed by visualization with BMChemiluminescence Kit.
Results
The yeast two-hybrid assay is a genetic test used to detectprotein interactions in vivo. It based on the fact that transcrip-tion factors consist of two separate functional domains, theDNA binding domain (bd) and the transcription domain (ad).Both components are required to activate transcription oftarget gene (β-galactosidase ). In this assay, the Q domain ofTLE1 (33–112 aa) was capable to interact with the intracel-lular region of gp130 (gp130IC) as well as the intact TLE1,while the Q domain deleted form (269–770 aa) could not (Ta-ble 1). On the other hand, the membrane proximal region ofgp130IC (642–753 aa) was essential to the interaction withTLE1, while the C-terminal region (751–918 aa) could notmediate this interaction.
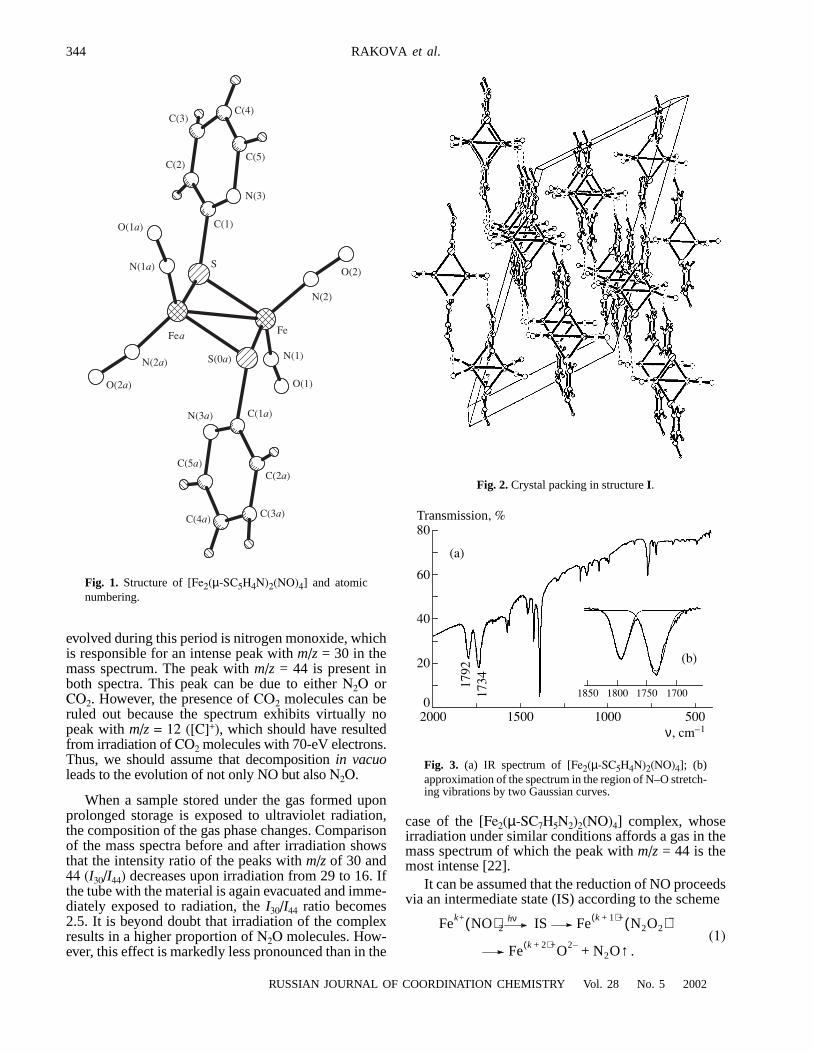
The results of the yeast two-hybrid assay were verified withprotein interaction assays using GST fusion proteins (GSTpull-down assay). The absorbed rabbit anti-TLE1 serum spe-cifically recognized TLE1 without cross-reaction with GSTor bacterial components from JM109 in ELISA, so did theanti-gp130IC serum. It was known that TLE1 could interactwith HES1 [10], so GST-HES1 fusion protein was used as apositive control. Western blot indicated the membrane proxi-mal region of gp130IC (642–753 aa) was capable to interactwith nature TLE1 as well as intact gp130IC (642–918 aa),while the C-terminal region (751–918 aa) could not mediatethis interaction (Fig. 2). Western bolt after immuno-precipi-tation also indicated that TLE1 in proteins extract of pEF-BOS-gp130 transfected HeLa cells was co-precipitated withgp130 by anti-gp130 serum, while over-expressed exogenesisgp130 could also be co-precipitated with TLE1 by anti-TLE1serum (Fig. 3).
Discussion
Our previous work showed that over-expression of AES in-hibited the phosphorylation of gp130 and JAK; M1 cellsover-expressing AES partly lost growth arrest response to
166
IL-6 [6, 11], indicating that AES might play a role in gp130signaling. By yeast two-hybrid assay, it was proved that AESinteracted with the intracellular membrane-proximal regionof gp130 through its Q domain. Considering the high homol-ogy (74%) between the Q domain of AES and that of TLE1,we presumed that TLE1 could also interact with the intrac-ellular region of gp130 and verified that by yeast two-hybridassay and protein binding assay. The Q domain of TLE/AEScould form 2 deduced leucin-zippers, which often participatein protein interaction [12]. It had been proved that Q domainof Gro is required for the formation of tetramer and its tran-scription repression activity [13].
Although there is only marginal evidence, it’s reasonableto believe that the Gro/TLE activity is under tight regulation.It was reported that Gro/TLE1 could be phosphorylated, whichenhanced their binding with nuclear components and mightup-regulate their capability to enter the nucleus [14]. Untilnow, the protein kinase responsible for this phosphorylation
is unknown. Based on the finding of the interaction of TLE1and gp130, we hypothesized that gp130 might be involvedin the phosphorylation of TLE.
It has been proved that gp130 mediated signal transduc-tion both by Ras-MAPK pathway and JAK-STAT pathway[15]. JAK kinases constitutively associated with the intrac-ellular region of gp130 (gp130IC), and are activated by crosstyrosine phosphorylation upon the dimerization of gp130.Activated JAKs phosphorylate tyrosines in gp130IC, andthese phosphorylated tyrosines (pY) serving as dock sites torecruit downstream signal molecules including STATs and SH2domain-containing tyrosine phosphatase2 (SHP2). SHP2serves as adaptor molecule to activate Ras, then the activatedRas triggers MAPKKK-MAPKK-MAPK cascade [16, 17].The intracellular region of gp130 contains 2 motifs conservedin members of the hematopoietic cytokine receptor family,namely Box1 and Box2. Box1 is a proline-rich motif of eightamino acid residues essential for JAK association. Box2, acluster of hydrophobic amino acid residues followed by posi-tively charged amino acids, also contributes to Jak-binding.Box3 is a motif of ten amino acid residues conserved betweengp130, LIF receptor and G-CSF receptor. SHP2 binds to phos-photyrosine-759 of gp130 that just precedes the Box3, whileSTATs, especially STAT3, are recruited by four YXXQ motifsin the C-terminal region starting from Box3 [18]. We demon-strated here that 642–753 aa of gp130 which containing Box1and Box2 is sufficient to mediate interaction between gp130and TLE1, while 751–918 aa of gp130 which containing Box3and four YXXQ motifs is not required for this interaction.
In conclusion, considering the key role of TLE in transcrip-tion regulation, the interaction between gp130 and TLE1might be a phenomenon of great consequence. Further inves-tigations on the role of TLE1 in gp130 mediated signalingare underway.
Acknowledgements
This work is supported by Chinese NkBRSF grant no. 200/CB510004 and NSF grants nos. 39880011 and 39800159. Wesincerely thank Dr Tetsuya Taga of Tokyo Medical and Den-tal University, Dr Stefano Stifani of Montreal McGill Uni-versity and Dr. Shigekazu Nagata of Jichi Medical School andfor their kind gift of vectors and advice.
References
1. Kishimoto T, Shizuo A, Narazaki M, Taga T: Interleukin-6 family ofcytokines and gp130. Blood 86: 1243–1255, 1995
2. Liu Y, Jin BQ, Liu F: Cloning and expression of a novel gp130 asso-ciated molecule. Zhonghua Weishengwuxue he Mianyixue ZaZhi 18:106–110, 1998
1 2 3 4 5
Fig. 2. Interaction between nature TLE1 protein and GST-gp130IC fusionprotein. Approximately 25 µg of GST-gp130642–918 (lane1), GST-gp130751–
918 (lane2), GST-gp130642–753 (lane 3), GST (lane 4) or GST-HES1 (lane 5)were incubated with approximately 100 µg HeLa cell proteins extract, fol-lowed by addition of glutathione-sepharose beads. The beads were re-covered and washed, and bound proteins were eluted and subjected toSDS-PAGE on a 10% gel. After electrophoretic transfer to a PVDF mem-brane, Western blotting was performed with rabbit anti-TLE1 serum (1:100).TLE1 proteins exhibit an apparent Mr of 90,000–93,000.
1 2 3 4 1 2 3 4
Fig. 3. Interaction between cellular TLE1 and gp130 proteins. Proteinsextract of pEF-BOS-gp130 transfected HeLa cells precipitated by anti-gp130 serum (lane1), anti-GST serum (lane2), anti-TLE1 serum (lane3) orpre-immunized rabbit serum (lane 4) were subjected to SDS-PAGE andtransferred to PVDF membrane. These precipitated proteins were firstlyblotted with anti-gp130 MAb AM64 (left panel), then blotted with anti-TLE1 MAb 5F9 (right panel) after striping. Molecular size standards areindicated in kilo-daltons (200, 100, 68, 43 kDa).

167
3. Miyasaka H, Choudhury BK, Li SSL: Molecular cloning and expres-sion of mouse and human cDNA encoding Drosphila enhancer of splitgroucho protein. Eur J Biochem 216: 343–352, 1993
4. Mallo M, Maguire MG, Harbison ML, Gridley T: Protein characteri-zation and targeted disruption of Grg. Dev Dyn 204: 338–347, 1995
5. Tetsuka T, Uranishi H, Imai H, Ono T, Sonta S, Takahashi N, AsamitsuK, Okamoto T: Inhibition of nuclear factor-kappaB-mediated transcrip-tion by association with the amino-terminal enhancer of split, a groucho-related protein lacking WD40 repeats. J Biol Chem 275: 4383–4390,2000
6. Liu Y, Jin BQ: gp130 Associated molecule inhibit the association ofJAK and gp130. Dankelong Kangti Tongxun (Chinese) 11: 48–51, 1995
7. Hartley DA, Preiss A, Artavanis-Tsakonas S: A deduced gene productfrom the Drosophila neurogenic locus, enhancer of split, shows homol-ogy to mammalian G-protein beta subunit. Cell 55:785–795, 1988
8. Stifani S, Blaumueller CM, Redhead NJ, Hill RE, Artavanis-TsakonasS: Human homologies of a Drosphila enhancer of split gene productdefine a novel family of nuclear proteins. Nature Genet 2: 119–127, 1992
9. Chen G, Courey AJ: Groucho/TLE family proteins and transcriptionalrepression. Gene 249: 1–16, 2000
10. Grbavec D, Stifani S: Molecular interaction between TLE1 and thecarboxyl-terminal domain of HES-1 containing the WRPW motif.Biochem Biophys Res Commun 223: 701–705, 1996
11. Liu F, Liu Y, Jin B: Preparation and characterization of monoclonalantibodies against AES protein, a novel gp130 associated molecule.Hybridoma 12: 371–378, 1999
12. Pinto M, Lobe CG: Products of Grg family can dimerize through theamino-terminal Q domain. J Biol Chem 271: 33026–33037, 1996
13. Chen G, Nguyen PH, Courey AJ: A role for groucho tetramerizationin transcriptional repression. Mol Cell Biol 18: 7259–7268, 1998
14. Husain J, Lo R, Grbavec D, Stifani S: Affinity for the nuclear com-partment and expression during cell differentiation implicate phos-phorylated groucho/TLE1 forms of higher molecular mass in nuclearfunctions. Biochem J 317: 523–521, 1996
15. Anhuf D, Weissenbach M, Schmitz J, Sobota R, Hermanns HM, RadtkeS, Linnemann S, Behrmann I, Heinrich PC, Schaper F: Signal trans-duction of IL-6, leukemia-inhibitory factor, and oncostatin M: Struc-tural receptor requirements for signal attenuation. J Immunol 165:2535–2543, 2000
16. Kim H, Baumann H: Dual signaling role of the protein tyrosine phos-phatase SHP-2 in regulating expression of acute-phase plasma proteinsby interleukin-6 cytokine receptors in hepatic cells. Mol Cell Biol 19:5326–5338, 1999
17. Fuhrer DK, Feng G-S, Yang Y-C: Syp associates with gp130 and januskinase 2 in response to interleukin-11 in 3T3-L1 mouse preadipocytes.J Biol Chem 270: 24826–24830, 1995
18. Ernst M, Inglese M, Waring P, Campbell IK, Bao S, Clay FJ, Alexan-der WS, Wicks IP, Tarlinton DM, Novak U, Heath JK, Dunn AR: De-fective gp130-mediated signal transducer and activator of transcription(STAT) signaling results in degenerative joint disease, gastrointestinalulceration, and failure of uterine implantation. J Exp Med 194: 189–204, 2001
19. Hibi M, Murakami M, Saito M, Hirano T, Taga T, Kishimoto T: Mo-lecular cloning and expression of an IL-6 signal transducer, gp130. Cell63: 1149–1157, 1990
20. Mizushima S, Nagata S: pEF-BOS, a powerful mammalian expressionvector. Nucleic Acids Res 18: 5322, 1990
21. Liu F, Liu Y, Jin B: Expression and purification of gp130-associatedmolecule and preparation of its monoclonal antibodies. J Cell MolImmunol (Chinese) 14: 43–46, 1998

168





























