Embed Size (px)
Citation preview

The Shoot Stem Cell Niche in Angiosperms: Expression Patterns of WUSOrthologues in Rice and Maize Imply Major Modifications in theCourse of Mono- and Dicot Evolution
Judith Nardmann and Wolfgang WerrInstitut fur Entwicklungsbiologie, Koln, Germany
In Arabidopsis, stem cell homeostasis in the shoot apical meristem (SAM) is controlled by a feedback loop between WUSand CLV functions. We have identified WUS orthologues in maize and rice by a detailed phylogenetic analysis of the WOXgene family and subsequent cloning. A single WUS orthologue is present in the rice genome (OsWUS), whereas theallotetraploid maize genome contains 2 WUS paralogues (ZmWUS1 and ZmWUS2).
None of the isolated grass WUS orthologues displays an organizing center–type expression pattern in the vegetativeSAM as in Arabidopsis. In contrast, the grass-specific expression patterns relate to the specification of new phytomersconsistent with the transcriptional expression patterns of TD1 and FON1 (CLV1 orthologues of maize and rice, respec-tively). Moreover, the grass WUS and CLV1 orthologues are coexpressed in all reproductive meristems, where fasciationand supernumerary floral organs occur in td1 or fon1 loss-of-function mutants. The expression patterns of WUS ortho-logues in both grass species compared with those of dicots imply that major changes in WUS function, which are correlatedwith changes in CLV1 signaling, have occurred during angiosperm evolution and raise doubts about the uniqueness of theWUS/CLV antagonism in the maintenance of the shoot stem cell niche in grasses.
Introduction
Angiosperms, flowering plants, comprise the largestgroup of plants today with more than 235,000 species inover 300 families, with fossil records suggesting that manyof the modern families and genera may already have existed75 MYA. Angiosperms are considered as a monophyleticgroup, a true clade, comprising 2 classes—the monocoty-ledons and the dicotyledons—which diverged over 150MYA (Wikstrom and Kenrick 2001). The monophyletic or-igin relative to the tremendous number of species suggeststhat ancestral angiosperm genomes were prone to rapid di-versification in the course of evolution. The evolution ofmaize (Zea mays L.) is a special example because it wasforced by human selection through domestication overthe last 6,000–10,000 years (reviewed in Doebley 2004).Maize still hybridizes with its ancestral wild teosinte rela-tives Z. mays. ssp parviglumis or ssp. mexicana, and pio-neering genetic analyses led to the identification of fewmajor quantitative trait loci (QTL) that account for majorchanges in the plant architecture of maize compared withthe wild species such as the maize female inflorescence,the ear being unique in the plant kingdom. Some QTLs havebeen resolved into individual genes often encoding tran-scription factors such as teosinte branched1 (tb1; Doebleyet al. 1997), teosinte glume architecture1 (tga1; Wang et al.2005), and zfl2 (Z. mays FLORICAULA/LEAFY2), a can-didate gene for a QTL controlling ear rank differencesbetween maize and teosinte (Bomblies et al. 2003).
The gross morphological changes based on single lociraise the question how common are regulatory networkselaborated in the dicot model species Arabidopsis in mono-cots such as maize. Comparative approaches in the MADSbox gene family have provided some evidence that basicprinciples of the ABC model of flower development suchas B-function genes are conserved in angiosperms (Zahn
et al. 2005) and applicable to maize (Whipple et al. 2004)despite the different architecture of the grass flower.
In the shoot apex, a mechanism regulating the balancebetween indeterminate (meristem maintenance) and determi-nate growth (leaf development) also appears to be ancient.KNOX (knotted1-like homeobox) genes are expressed in ameristem-specific manner and are downregulated in primor-dial cells (Vollbrecht et al. 1993) by the activity of MYBorthologues (ARP: ASYMMETRIC LEAVES1, ROUGHSHEATH2, PHANTASTICA: Waites et al. 1998;Timmermans et al. 1999; Tsiantis et al. 1999; Byrne et al.2000), recently reviewed in Piazza et al. (2005). ThisKNOX–ARP pathway is not only conserved between mono-and dicots but was also independently recruited into micro-phyll development in lycophytes (Harrison et al. 2005). Incontrast, there are major differences in other aspects of leafdevelopment between mono- and dicots. Two paralogousloci in maize NARROW SHEATH1/NARROW SHEATH2(NS1/2) encode homologues to the Arabidopsis PRESSEDFLOWER (PRS) gene (Nardmann et al. 2004). NS1/2 andPRS have maintained their marginal expression domain inlateral organ primordia. The prs mutant phenotype, however,is mostly restricted to lateral sepals of the flower in Arabidop-sis, whereas the maize homologues contribute to the specifi-cation of a lateral leaf domain at the flank of the shoot apicalmeristem (SAM) during the vegetative phase (Scanlon 2000).
Some remarkable differences exist between meristemsof dicots and monocots. In dicot species including Arabidop-sis, the SAM appears to be 3 layered, with a tunica compris-ing 2 clonal layers (L1 1 L2) and the corpus commonlydesignated as the L3 layer (Szymkowiak and Sussex 1996;Evans and Barton 1997). In contrast, monocots such as maizehave only 1 histologically apparent single tunica layer (L1)and the inner corpus (Abbe et al. 1951; Steffensen 1968). Ra-dially, in theArabidopsis SAM, the central stem cell zone canbe distinguished from the peripheral zone, where cells beginto differentiate. Vegetative leaves in Arabidopsis originatefrom few founder cells specified in a spiral phyllotaxy inthe peripheral zone (Irish and Sussex 1992), whereas in con-trast, the maize leaf may be traced back to approximately 200leaf founder cells (Poethig 1984), which are recruited from
Key words: WUSCHEL orthologues, maize, rice, angiospermevolution.
E-mail: [email protected].
Mol. Biol. Evol. 23(12):2492–2504. 2006doi:10.1093/molbev/msl125Advance Access publication September 20, 2006
� The Author 2006. Published by Oxford University Press on behalf ofthe Society for Molecular Biology and Evolution. All rights reserved.For permissions, please e-mail: [email protected]
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/23/12/2492/974436 by guest on 20 Novem
ber 2018

the whole circumference of the shoot apex and give rise toan alternate phyllotaxy.
The maintenance of pluripotent stem cells in the SAMof Arabidopsis depends on the WUSCHEL/CLAVATAfeedback loop. The WUSCHEL (WUS) homeobox gene pro-vides a key function for the maintenance of stem cell ho-meostasis in the SAM (Mayer et al. 1998). WUS acts cellautonomously and gene expression is restricted to a fewcells in the L3 layer considered to be the stem cell–organizingcenter (OC). Whereas WUS promotes stem cell fate, it isantagonized by CLAVATA (CLV) signaling (Brand et al.2000). Acting in a single pathway, 3 CLV genes (CLV1–3)encode a heterodimeric transmembrane receptor kinase(CLV1/2; Clark et al. 1997; Jeong et al. 1999) and its corre-sponding polypeptide ligand (CLV3; Fletcher et al. 1999).WUS activity is restricted via CLV signaling via a yet un-known signal transduction pathway (Brand et al. 2000).
The recent cloning of THICK TASSELDWARF1 (TD1;Bommert et al. 2005) and FLORAL ORGAN NUMBER(FON1; Suzaki et al. 2004), CLV1 orthologues from maizeand rice, respectively, and of FASCIATED EAR2 (FAE2), aCLV2 orthologue from maize (Taguchi-Shiobara et al.2001), has suggested that CLV signaling is conserved inmonocots. The expression patterns of these genes as wellas the phenotypes of corresponding loss-of-function mu-tants, however, imply major modifications in the controlof meristem size in grasses. In Arabidopsis, CLV1 transcrip-tion is confined to the L2 and L3 layers of the SAM through-out the plant life cycle (Clark et al. 1997). In contrast, TD1transcripts are not detected in the vegetative SAM but foundin early leaf primordia (Bommert et al. 2005). During thereproductive phase, however, TD1 is transcribed throughoutthe inflorescence meristem (IM), the descending spikelet-pair meristems (SPMs), spikelet meristems (SMs), and floralmeristem (FMs). During later stages of floral development,TD1 transcripts accumulate in differentiated organs such asglumes, lemma, and palea, reminiscent of the expressionpattern in leaf primordia during vegetative phase (Bommertet al. 2005). Similarly, the rice orthologue FON1 is tran-scribed throughout the FM but also in differentiated lateralfloral organs (Suzaki et al. 2004).
As CLV1 signaling is thought to restrict WUS activity,the differences in the transcription patterns of CLV1 ortho-logues between maize and rice or Arabidopsis raised thequestion whether the expression pattern ofWUS orthologuesin grasses was also conserved. WUS is the founding memberof the so-called WOX (WUS homeobox) gene family en-coded by 15 genes in the Arabidopsis genome (Haeckeret al. 2004). Here, we describe the identification of WUSorthologues in maize and rice by a detailed phylogeneticcomparison of the WUS/WOX gene family between the di-cots Arabidopsis thaliana and Populus trichocarpa and 2monocot species Oryza sativa and Z. mays. The allotetra-ploid maize genome contains 2 WUS paralogues (ZmWUS1and ZmWUS2), whereas a single WUS orthologue is presentin the smaller rice genome (OsWUS). Consistent with TD1/FON1 expression data, the expression patterns of WUS or-thologues in both grass species imply that major changesduring angiosperm evolution have occurred and raise doubtsabout the uniqueness of the WUS/CLV antagonism in themaintenance of the shoot stem cell niche in grasses.
Materials and MethodsCloning of ZmWUS1 and ZmWUS2
To amplify the homeobox of the maize WUS ortho-logues, polymerase chain reaction (PCR) was performed ongenomic DNA of Rscm2 maize using the primer pair ZmHD1(5#-TGGACICCIACIACIGARCARAT-3#)andZmHD2 (5#-GCYTTRTGRTTYTGRAACCARTARAA-3#). Isolation oftotal RNA followed the protocol of Chomczynski and Sacchi(1987), and rapid amplification of the 3#-cDNA ends wasthen performed on RNA of Rscm2 female inflorescenceswith primers corresponding to the homeobox using the First-Choice RLM-RACE Kit (Ambion, Austin, TX) according tothe manufacturer’s protocol; 3#-RACE was performed withprimers ZmWUSa (5#-CTCTACTACGGCTGCGGCATCC-3#) and ZmWUSb (5#-GACAAGATCGAGGGTAA-GAAC-3#). Analysis of the 3#-RACE products revealed3#-ends of 2 different genes we named ZmWUS1 andZmWUS2. For5#-RACE, primers specific forZmWUS1wereused: ZmWUS1a (5#-CAATCGGAACTCGTGTCCAG-CATCAC-3#) and ZmWUS1b (5#GAACAACCTGAA-GAAGGCGACAACGAG-3#). 5#-RACE for ZmWUS2was performed with the primers ZmWUS2a (5#-CCAT-TATCGTGTCCGACGACGAGAAG-3#) and ZmWUS2b(5#-CATGTAGTCCTGCAGGAAGCATGTCTC-3#).
All PCR products were cloned into pCRII TOPO (Invi-trogen, Carlsbad, CA) and sequenced. ZmWUS2 was mappedto chromosome 10, between markers bnlg1450 and asg19b,using the DNA Kit of 94 IBM Lines (http://www.maizemap.org/dna_kits.htm).
Computational and Database Analysis
Analysis of DNA and protein sequences was per-formed using the Wisconsin GCG software package version7.0 (University of Wisconsin Genetics Computer Group).Homology searches were performed using TBlastN atNational Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov/blast/), DNA Data Bank Japan(http://www.ddbj.nig.ac.jp), or the Plant Genome Data-base (http://plantgdb.org) with default parameters. Multi-ple sequence alignment was performed using ClustalW(http://www.ebi.ac.uk/clustalw) and BOXSHADE (http://searchlauncher.bcm.tmc.edu/multi-align/multi-align.html).The PHYLIP (http://evolution.genetics.washington.edu/phylip.html) program was used for phylogenetic and mo-lecular evolutionary analyses based on the maximum likeli-hood method. Sequences selected for the phylogenetic treein figure 1A are as follows (accession numbers in parenthe-ses): ZmNS1 (AJ536578); ZmNS2 (AJ472083), and Os-WOX5 (Q8WOF1); AtWOX1–AtWOX14 as publishedrecently (Haecker et al. 2004). Accession numbers forthe remaining homeodomains (HDs) are listed in table 1.Accession numbers of the genes encoding the proteinsfor the sequence alignment are ZmWUS1 (AM234744),ZmWUS2 (AM234745), OsWUS (AB218894), ROA(AY162209), TER (AF481951), PtWUS (AM234747),and AtWUS (At2g17950).
In situ Hybridization
For nonradioactive in situ hybridization, sampleswere prepared following the protocol of Jackson (1991).
WUSCHEL Orthologues in Maize and Rice 2493
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/23/12/2492/974436 by guest on 20 Novem
ber 2018

For sections of maize embryos, kernels were trimmed onboth sides of the embryo axis for better penetration ofthe formaldehyde fixative and the wax solution. Paraffin-embedded tissue was sectioned by the use of the LeicaRM 2145 rotary microtome, and 7-lm sections were usedfor hybridization. Probes for in situ hybridization werecloned either in sense or antisense orientation to the T7 pro-moter and then used as a template for synthesis of digoxi-genin-labeled RNA probes by T7 RNA polymerase asdescribed (Bradley et al. 1993). In situ probes for the grassWUS genes were chosen to contain sequences downstreamof the homeobox and amplified via PCR: ZmWUS1 pos.294–951 of accession AM234744, ZmWUS2 pos. 267–978 of accession AM234745, OsWUS pos. 435–870 of ac-cession AB218894. In situ probes for TD1 and FON1 wereused as published (Suzaki et al. 2004; Bommert et al. 2005).The probe for KN1 corresponded to the mRNA sequence ofaccession AY312169 (Vollbrecht et al. 1993).
Light Microscopy and Image Processing
Imageswere takenusinganAxioskopmicroscopeequip-ped with anAxiocam camera (Zeiss, Oberkochen, Germany).Pictures were processed using Adobe Photoshop version 7.0.
ResultsWUS Orthologues in Rice and Maize
Whereas members of the WUS/WOX gene family fromrice could be identified from the genome sequence (Goffet al. 2002), the maize family members had to be isolatedusing a combinatorial approach: firstly, reverse transcrip-tase (RT)–PCR was performed on cDNAs prepared fromimmature embryos, roots, shoots, and inflorescences withdegenerate primers designed to amplify the conservedHD-encoding sequences of WUS/WOX relatives (Haeckeret al. 2004). Secondly, the available maize genome se-quences were screened for WUS/WOX HD-encoding se-quences. In total, 16 members of the WUS/WOX genefamily were identified and subjected to phylogenetic anal-yses based on the HD amino acid sequence (fig. 1A). In ad-dition to the identified rice members, the newly identifiedmaize (Zm: Zea mays) genes, and the founding Arabidopsisthaliana (At) family members, we included the WOX HDsequences from a second dicot Populus trichocarpa (Pt).Similar to the rice genome (Goff et al. 2002), the populusgenome has been fully sequenced (http://genome.jgi-psf.org/poptr1_1/poptr1_1.home.html) and therefore shouldprovide the full WOX gene complement. The resulting phy-logenetic tree unambiguously reveals 2 WUS paralogues toexist in maize, ZmWUS1 and ZmWUS2, and a single ortho-logue in rice, OsWUS. All 3 monocot WUS genes groupwithin 1 unique subbranch together with their 2 dicot ortho-logues, AtWUS and PtWUS. Only members of this WUS
branch encode HDs with the typical extra tyrosine residuein the loop between helix 1 and helix 2 (fig. 1B). The aminoacid similarity between ZmWUS1 or ZmWUS2 and At-WUS of Arabidopsis is 83% or 85%, respectively, andthe similarity is higher when compared with OsWUS(97% in each case). We also included 2 recently identifiedWUS orthologues, ROA (ROSULATA; Kieffer et al. 2006)and TER (TERMINATOR; Stuurman et al. 2002), from thedicot species Antirrhinum majus and Petunia hybrida, re-spectively, in this alignment. Even between the dicot spe-cies, there is little sequence conservation between the WUSorthologues outside the HD apart from a few short motifs:an acidic domain, the WUS box of unknown function(Haecker et al. 2004), and an ethylene-responsive elementbinding factor-associated amphiphilic repression (EAR)–like domain (Ohta et al. 2001). The EAR-like domain isonly found in WUS and its orthologues ROA and TERand not in other members of the WOX family in Arabidop-sis. The presence of this domain in ZmWUS1, ZmWUS2,and OsWUS, therefore, is further evidence for their beingWUS orthologues in these monocot species.
We were not able to identify polymorphisms in codingor immediate upstream and downstream sequences ofZmWUS1 in different mapping populations and were there-fore unable to map the gene so far. The map position ofZmWUS2 resides in a region on chromosome 10, which isduplicated on chromosome 2. The duplication of the maizegenome is also evident in the WOX2, 5, or 13 branches,where 2 maize paralogues group with single orthologuesin Oryza, Arabidopsis, or Populus (fig. 1A). Additionally,the duplicated narrow sheath1 and 2 loci, which encode pa-ralogues of PRS in Arabidopsis, support this genome dupli-cation and indicate conserved gene functions, althoughadapted to grass- or dicot-specific developmental programs(Matsumoto and Okada 2001; Nardmann et al. 2004). Othersignificant aspects of the phylogeny are that orthologues ofAtWOX1, 6, 7, 8, 10, and 14 are absent in the 2 grasses,whereas orthologues of AtWOX1, 6, and 7 are found in thesecond dicot genome (P. trichocarpa). Notably, orthologuesof AtWOX8, 10, and 14 are also absent from the Populus ge-nome, and for AtWOX10 and At WOX14, no expression wasdetected in Arabidopsis (Haecker et al. 2004). In contrast,however, the transcriptional activity of allmaize genes exceptZmWOX2B could be confirmed by spliced intron–spanningRT–PCR amplicons (data not shown).
OsWUS, the Single Rice WUS Orthologue Marks LeafMargins and Is Transiently Expressed in the SAM
As phylogenetic analyses revealed a single WUS ho-mologue in rice, OsWUS, RNA in situ hybridizations wereperformed in young rice seedlings, which uncovered re-markable differences between the expression patterns of
FIG. 1.—WUSCHEL/WOX genes in maize and rice. (A) Phylogenetic tree based on HD sequence information of newly isolated Z. mays (Zm) WUS/WOX relatives in comparison to the relatives encoded by the Arabidopsis thaliana (At), Oryza sativa (Os), and P. trichocarpa (Pt) genomes. (B) Proteinsequence comparison of Arabidopsis WUS (AtWUS) with its orthologue in Populus (PtWUS), Antirrhinum (ROA), Petunia (TER), the 2 maize WUSparalogues, ZmWUS1 or ZmWUS2, and the single rice WUS orthologue (OsWUS). The extra tyrosine residue specific for the WUS HD is indicated by anarrow. The positions of the HD, the WUS boxes, and the EAR-like domain are indicated by gray lines.
!
2494 Nardmann and Werr
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/23/12/2492/974436 by guest on 20 Novem
ber 2018

WUSCHEL Orthologues in Maize and Rice 2495
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/23/12/2492/974436 by guest on 20 Novem
ber 2018

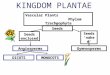
OsWUS and those established for the WUS gene in Arabi-dopsis (Mayer et al. 1998). OsWUS transcripts are detectedin young leaf primordia with a pronounced preference forlateral leaf margins. Depicted in figure 2A1 B are leaf pri-mordia P1, P2, and P3, and highest OsWUS transcript levelsare evident at the very tip of P2 and P3 lateral margins. In thecenter of the shoot apex, where WUS is exclusively ex-pressed in the Arabidopsis SAM, OsWUS transcripts wereundetectable in the majority of consecutive sections (total15) through the rice SAM. However, in a small number ofconsecutive sections (5 out of 15), OsWUS expression wasalso detected in the center of the vegetative SAM in 1–2sections at the height of the P0 phytomer (fig. 2A). Thismay be best explained by a transient expression of OsWUSin the shoot apex in contrast to the stable OC-type expres-sion pattern of WUS in Arabidopsis. During later stages ofphytomer development, OsWUS is expressed at the abaxialface of emerging axillary meristems (fig. 2C).
These differences in expression patterns and the iden-tification of 2 WUS ortholgues in maize, ZmWUS1 andZmWUS2, raised questions concerning the expression do-mains of the maize paralogues and whether there aregrass-specific alterations in the WUS expression domainscompared with Arabidopsis. The expression patterns ofthe maize ZmWUS1 and ZmWUS2 differ considerablyand will be described separately below.
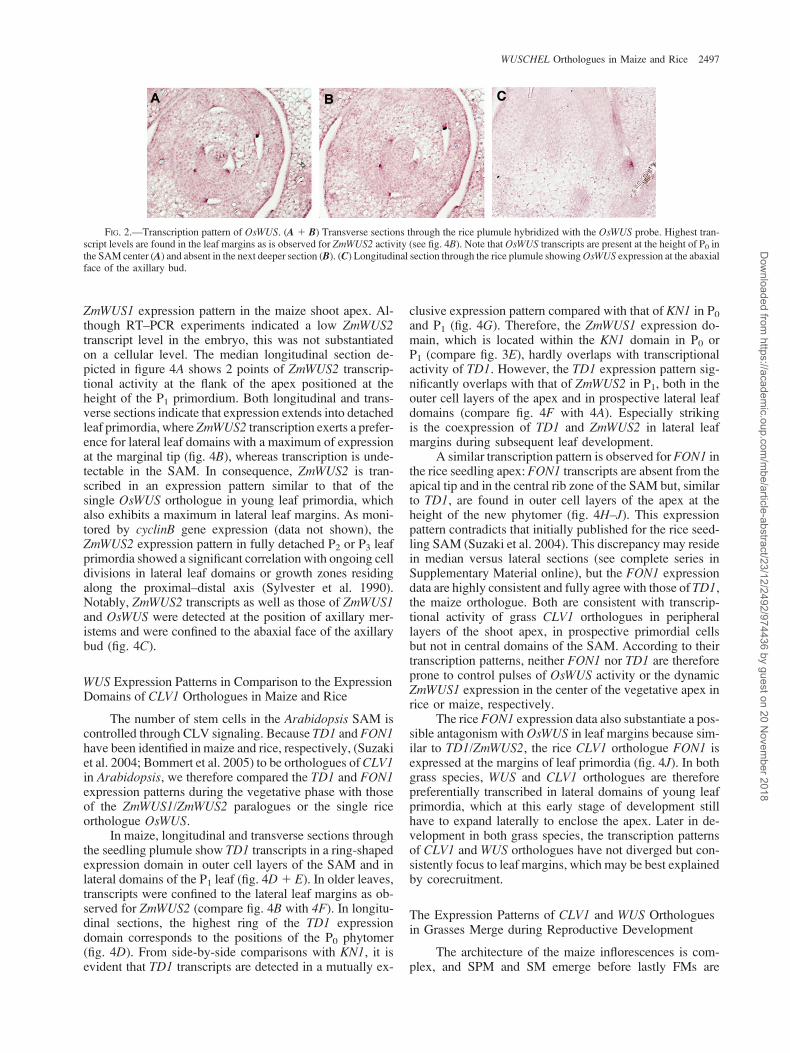
The ZmWUS1 Pattern in the Seedling SAM Is Dynamic
Transcriptional activity of ZmWUS1 was not detectedearlier than the coleoptilar stage embryo. ZmWUS1 wastranscribed in a few cells underlying the emerging coleop-tile (fig. 3A1 B) in a domain initially including the L1 layerbut later extending to subtending cell layers (fig. 3A–C).Expression then ceased after the initiation of the secondembryonic leaf, even though the maize embryo developsadditional 4 leaves prior to seed dormancy. ZmWUS1 tran-scriptional activity was again detected in the seedling SAMafter germination, although this expression was weak in thecenter of the apex with transcript maxima detectable at lat-eral positions of the apex (fig. 3D). Multiple series of lon-gitudinal and transverse sections showed that transcript
maxima dynamically differed between opposing flanks ofthe apex (data not shown). Moreover, consecutive (7 lmthick) transverse sections showed that the height of thisZmWUS1 domain in the center of the apex changes relativeto the apical tip in the course of development, as does therelative height of P1 and P0 (fig. 3E). The vegetative maizephytomer comprises a leaf attached to a node, an internode,and an axillary bud. Although located close to the midribposition of the subtending phytomer, the axillary meri-stem clonally relates to the internode above. The lateralZmWUS1 maxima, therefore, coincide either with the mid-rib positions of the emerging leaf primordia, P1 and P0, orwith the positions of the axillary meristems at the oppositeface of the apex.
To elucidate the position of the lateral ZmWUS1 max-ima, we performed side-by-side in situ hybridizationexperiments with a Knotted1 (KN1) probe (Vollbrecht et al.1993). The ZmWUS1 transcription domains at the height ofthe P0 and P1 phytomers in the shoot apex are shown infigure 3E, which also depicts adjacent sections hybridizedwith the KN1 probe. The P1 sections in figure 3E showa downregulation of KN1 expression at the left flank ofthe SAM corresponding with the midrib of the emergingP1 leaf. ZmWUS1 activity is absent in these primordial cellsbut overlaps with the KN1 expression in a central domain ofthe apex. However, in a deeper section, ZmWUS1 expres-sion extends toward the flank of the SAM opposite to the P1
midrib. Therefore, ZmWUS1 transcripts are absent infounder cells of the P1 leaf sheath/blade but are transcribedin cells of the prospective node/internode in the center ofthe shoot apex and in the subtending axillary meristem,which is located opposite to the P1 midrib. Consequently,the expression maxima at the flanks of the apex (see fig. 3D)relate to the formation of axillary meristems and not to thespecification of leaf founder cells at the prospective midribposition, and subsequently, the ZmWUS1 expression do-main is confined to the abaxial face of the axillary bud(fig. 3G).
In summary, the ZmWUS1 transcription domain in thecenter of the shoot apex initiates as a disc-shaped expres-sion domain at the height of the new P0 phytomers, which isreminiscent of the transient expression of the single OsWUSorthologue in the center of the rice apex. The expression ofZmWUS1 in these cells either persists or is reactivated dur-ing subsequent phytomer development, shifting to deeperlayers of the SAM. The transcription domain extends later-ally to the position of axillary meristems in P1 (fig. 3F).Transcription ceases thereafter, except for activity in axil-lary meristems (fig. 3G). Similar to the transient OsWUSexpression in the rice SAM, the dynamic ZmWUS1 expres-sion pattern in the maize shoot apex does not reflect the sta-ble OC-type expression domain of WUS in Arabidopsis.However, ZmWUS1 expression appears to be correlatedwith the initiation of leaf phytomers and associated withthe initiation of the SAM in the embryo or the establishmentof axillary meristems at the flank of the vegetative apex.
ZmWUS2 Is Activated in the P1 Leaf Primordium
The in situ hybridization results obtained with theZmWUS2 probe were in striking contrast to the dynamic
Table 1Accession Numbers of WOX HDs
Nameof HD
AccessionNumber
Nameof HD
AccessionNumber
ZmWOX2A AM234767 OsWOX9 AM234752ZmWOX2B AM234778 OsWOX9C AM234753ZmWOX4 AM234768 OsWOX11/12 AM234754ZmWOX5A AM234769 OsWOX13 AM234755ZmWOX5B AM234770 PtWOX1A AM234756ZmWOX9A AM234771 PtWOX1B AM234757ZmWOX9B AM234772 PtWOX2 AM234758ZmWOX9C AM234773 PtWOX4 AM234759ZmWOX11/12A AM234774 PtWOX5/7A AM234765ZmWOX11/12B AM234775 PtWOX5/7B AM234766ZmWOX13A AM234776 PtWOX6 AM234760ZmWOX13B AM234777 PtWOX9 AM234761OsNS AM234748 PtWOX11/12A AM234763OsWOX2 AM234749 PtWOX11/12B AM234764OsWOX4 AM234750 PtWOX13 AM234762
2496 Nardmann and Werr
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/23/12/2492/974436 by guest on 20 Novem
ber 2018
ZmWUS1 expression pattern in the maize shoot apex. Al-though RT–PCR experiments indicated a low ZmWUS2transcript level in the embryo, this was not substantiatedon a cellular level. The median longitudinal section de-picted in figure 4A shows 2 points of ZmWUS2 transcrip-tional activity at the flank of the apex positioned at theheight of the P1 primordium. Both longitudinal and trans-verse sections indicate that expression extends into detachedleaf primordia, where ZmWUS2 transcription exerts a prefer-ence for lateral leaf domains with a maximum of expressionat the marginal tip (fig. 4B), whereas transcription is unde-tectable in the SAM. In consequence, ZmWUS2 is tran-scribed in an expression pattern similar to that of thesingle OsWUS orthologue in young leaf primordia, whichalso exhibits a maximum in lateral leaf margins. As moni-tored by cyclinB gene expression (data not shown), theZmWUS2 expression pattern in fully detached P2 or P3 leafprimordia showed a significant correlation with ongoing celldivisions in lateral leaf domains or growth zones residingalong the proximal–distal axis (Sylvester et al. 1990).Notably, ZmWUS2 transcripts as well as those of ZmWUS1and OsWUS were detected at the position of axillary mer-istems and were confined to the abaxial face of the axillarybud (fig. 4C).
WUS Expression Patterns in Comparison to the ExpressionDomains of CLV1 Orthologues in Maize and Rice
The number of stem cells in the Arabidopsis SAM iscontrolled through CLV signaling. Because TD1 and FON1have been identified in maize and rice, respectively, (Suzakiet al. 2004; Bommert et al. 2005) to be orthologues of CLV1in Arabidopsis, we therefore compared the TD1 and FON1expression patterns during the vegetative phase with thoseof the ZmWUS1/ZmWUS2 paralogues or the single riceorthologue OsWUS.
In maize, longitudinal and transverse sections throughthe seedling plumule show TD1 transcripts in a ring-shapedexpression domain in outer cell layers of the SAM and inlateral domains of the P1 leaf (fig. 4D1 E). In older leaves,transcripts were confined to the lateral leaf margins as ob-served for ZmWUS2 (compare fig. 4B with 4F). In longitu-dinal sections, the highest ring of the TD1 expressiondomain corresponds to the positions of the P0 phytomer(fig. 4D). From side-by-side comparisons with KN1, it isevident that TD1 transcripts are detected in a mutually ex-
clusive expression pattern compared with that of KN1 in P0
and P1 (fig. 4G). Therefore, the ZmWUS1 expression do-main, which is located within the KN1 domain in P0 orP1 (compare fig. 3E), hardly overlaps with transcriptionalactivity of TD1. However, the TD1 expression pattern sig-nificantly overlaps with that of ZmWUS2 in P1, both in theouter cell layers of the apex and in prospective lateral leafdomains (compare fig. 4F with 4A). Especially strikingis the coexpression of TD1 and ZmWUS2 in lateral leafmargins during subsequent leaf development.
A similar transcription pattern is observed for FON1 inthe rice seedling apex: FON1 transcripts are absent from theapical tip and in the central rib zone of the SAM but, similarto TD1, are found in outer cell layers of the apex at theheight of the new phytomer (fig. 4H–J). This expressionpattern contradicts that initially published for the rice seed-ling SAM (Suzaki et al. 2004). This discrepancy may residein median versus lateral sections (see complete series inSupplementary Material online), but the FON1 expressiondata are highly consistent and fully agree with those of TD1,the maize orthologue. Both are consistent with transcrip-tional activity of grass CLV1 orthologues in peripherallayers of the shoot apex, in prospective primordial cellsbut not in central domains of the SAM. According to theirtranscription patterns, neither FON1 nor TD1 are thereforeprone to control pulses of OsWUS activity or the dynamicZmWUS1 expression in the center of the vegetative apex inrice or maize, respectively.
The rice FON1 expression data also substantiate a pos-sible antagonism with OsWUS in leaf margins because sim-ilar to TD1/ZmWUS2, the rice CLV1 orthologue FON1 isexpressed at the margins of leaf primordia (fig. 4J). In bothgrass species, WUS and CLV1 orthologues are thereforepreferentially transcribed in lateral domains of young leafprimordia, which at this early stage of development stillhave to expand laterally to enclose the apex. Later in de-velopment in both grass species, the transcription patternsof CLV1 and WUS orthologues have not diverged but con-sistently focus to leaf margins, which may be best explainedby corecruitment.
The Expression Patterns of CLV1 and WUS Orthologuesin Grasses Merge during Reproductive Development
The architecture of the maize inflorescences is com-plex, and SPM and SM emerge before lastly FMs are
FIG. 2.—Transcription pattern of OsWUS. (A 1 B) Transverse sections through the rice plumule hybridized with the OsWUS probe. Highest tran-script levels are found in the leaf margins as is observed for ZmWUS2 activity (see fig. 4B). Note that OsWUS transcripts are present at the height of P0 inthe SAM center (A) and absent in the next deeper section (B). (C) Longitudinal section through the rice plumule showing OsWUS expression at the abaxialface of the axillary bud.
WUSCHEL Orthologues in Maize and Rice 2497
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/23/12/2492/974436 by guest on 20 Novem
ber 2018
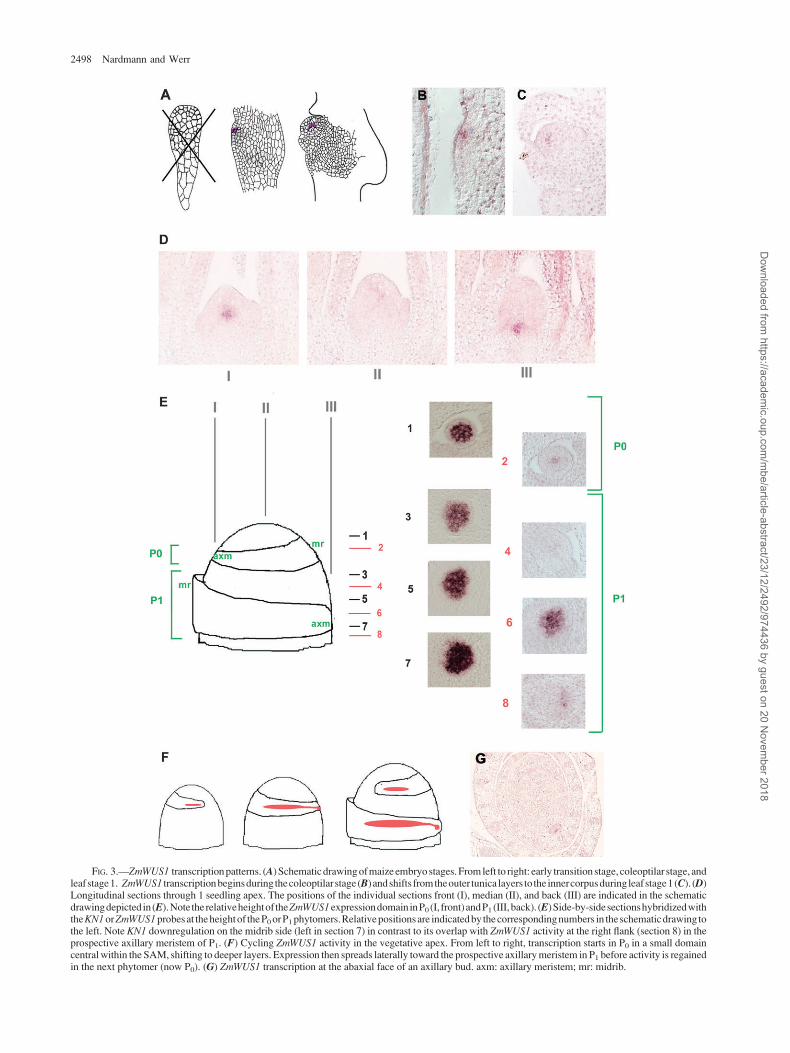
FIG. 3.—ZmWUS1 transcription patterns. (A) Schematic drawing of maize embryo stages.From left to right: early transitionstage, coleoptilar stage, andleaf stage1. ZmWUS1 transcriptionbeginsduring thecoleoptilar stage (B) andshifts fromtheouter tunica layers to the innercorpusduring leaf stage1 (C). (D)Longitudinal sections through 1 seedling apex. The positions of the individual sections front (I), median (II), and back (III) are indicated in the schematicdrawingdepicted in(E).Note the relativeheightof theZmWUS1expressiondomain inP0 (I, front) andP1 (III, back). (E)Side-by-sidesectionshybridizedwiththeKN1orZmWUS1probesat the height of the P0 orP1 phytomers.Relative positionsare indicatedby the correspondingnumbers in the schematic drawing tothe left. Note KN1 downregulation on the midrib side (left in section 7) in contrast to its overlap with ZmWUS1 activity at the right flank (section 8) in theprospective axillary meristem of P1. (F) Cycling ZmWUS1 activity in the vegetative apex. From left to right, transcription starts in P0 in a small domaincentral within the SAM, shifting to deeper layers. Expression then spreads laterally toward the prospective axillary meristem in P1 before activity is regainedin the next phytomer (now P0). (G) ZmWUS1 transcription at the abaxial face of an axillary bud. axm: axillary meristem; mr: midrib.
2498 Nardmann and Werr
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/23/12/2492/974436 by guest on 20 Novem
ber 2018

initiated (fig. 5A); secondary axes in the male tassel are dueto the activity of early branch meristems (BM). Both maizeWUS paralogues show meristem-specific expression dur-ing the reproductive phase (fig. 5B–G); however, signifi-cant differences exist between ZmWUS1 and ZmWUS2.Although active in axillary meristem anlagen (see fig.3G), no ZmWUS1 expression was detected in the matureIM of the male tassel or the female ear (fig. 5D). ZmWUS1transcripts were found in the descending SPM, SM, and FM(fig. 5B 1 C). In SPMs, SMs, and early FMs, ZmWUS1transcription is confined to a small OC-type central domain(fig. 5B) such as is observed in the early embryo. In the lateFM, the ZmWUS1 expression domain extends toward the
L1 layer similar to the shift of WUS expression from theL3 layer in the IM to the L2 layer in the FM in Arabidopsis(Mayer et al. 1998). The expression pattern, therefore, re-flects histological differences between 1 or 2 tunica layersin maize and Arabidopsis meristems, respectively. Later infemale flower development, ZmWUS1 expression wasdetected in the outer cell layers of the gynoecium in the up-per floret and still simultaneously in the center of the lowerfloret (fig. 5C), which subsequently aborts in the female earinflorescence.
In contrast, ZmWUS2 transcripts are found in the IM(fig. 5G) often with a slight preference for the L1 layer andwith mRNA being evenly distributed through tunica and
FIG. 4.—Transcription pattern of the ZmWUS2 paralogue and comparison with the expression of the CLV1 orthologues TD1 and FON1 in the veg-etative grass apex. (A) Longitudinal median section through the vegetative maize apex. ZmWUS2 is expressed in peripheral cell layers at the height of the P1
primordium. (B) Transverse section through the SAM at the height of P0 showing the absence of ZmWUS2 transcripts internally within the apex andexpression in P1. Note the strong preference in leaf margins of P2, already beginning in the upper margin of P1. (C) ZmWUS2 transcription at the abaxialside of the elaborated axillary bud (left), the signal at the right side of the apex corresponds to a younger axillary bud. (D) Longitudinal section through themaize SAM showingTD1 transcripts in P0 and P1. (E) Transverse section through P0 showing the ring-shapedTD1 expression domain at the SAM peripheryand the preference in lateral domains of P1. (F) TD1 marginal pattern in older leaf primordia. (G) Mutually exclusive transcription patterns of TD1 (left)and KN1 (right) in P0 (top) and P1 (bottom), as summarized in the schematic drawing to the right. (H1 I) Longitudinal and transverse sections throughthe rice apex showing FON1 transcripts at the periphery of the SAM. (J) Transverse section through a late P1 showing the FON1 preference in leafmargins.
WUSCHEL Orthologues in Maize and Rice 2499
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/23/12/2492/974436 by guest on 20 Novem
ber 2018
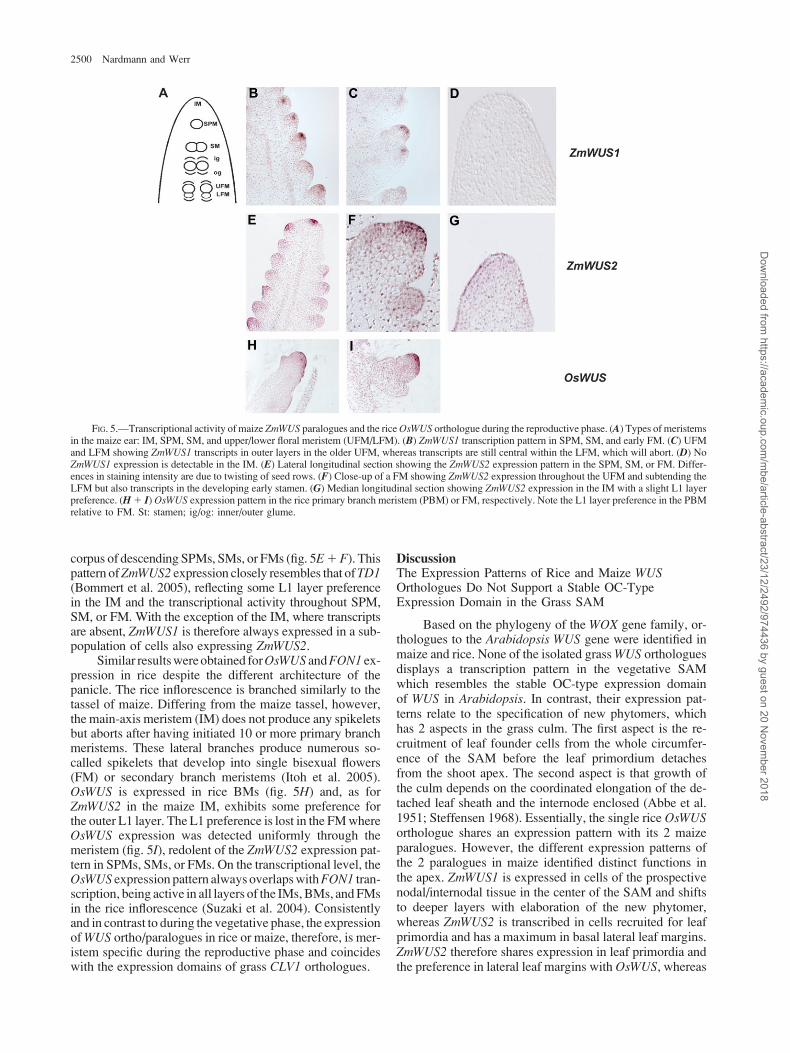
corpus of descending SPMs, SMs, or FMs (fig. 5E1F). Thispattern ofZmWUS2 expression closely resembles that ofTD1(Bommert et al. 2005), reflecting some L1 layer preferencein the IM and the transcriptional activity throughout SPM,SM, or FM. With the exception of the IM, where transcriptsare absent, ZmWUS1 is therefore always expressed in a sub-population of cells also expressing ZmWUS2.
Similar results were obtained forOsWUS andFON1 ex-pression in rice despite the different architecture of thepanicle. The rice inflorescence is branched similarly to thetassel of maize. Differing from the maize tassel, however,the main-axis meristem (IM) does not produce any spikeletsbut aborts after having initiated 10 or more primary branchmeristems. These lateral branches produce numerous so-called spikelets that develop into single bisexual flowers(FM) or secondary branch meristems (Itoh et al. 2005).OsWUS is expressed in rice BMs (fig. 5H) and, as forZmWUS2 in the maize IM, exhibits some preference forthe outer L1 layer. The L1 preference is lost in the FM whereOsWUS expression was detected uniformly through themeristem (fig. 5I), redolent of the ZmWUS2 expression pat-tern in SPMs, SMs, or FMs. On the transcriptional level, theOsWUS expression pattern always overlaps withFON1 tran-scription, being active in all layers of the IMs, BMs, and FMsin the rice inflorescence (Suzaki et al. 2004). Consistentlyand in contrast to during the vegetative phase, the expressionof WUS ortho/paralogues in rice or maize, therefore, is mer-istem specific during the reproductive phase and coincideswith the expression domains of grass CLV1 orthologues.
DiscussionThe Expression Patterns of Rice and Maize WUSOrthologues Do Not Support a Stable OC-TypeExpression Domain in the Grass SAM
Based on the phylogeny of the WOX gene family, or-thologues to the Arabidopsis WUS gene were identified inmaize and rice. None of the isolated grass WUS orthologuesdisplays a transcription pattern in the vegetative SAMwhich resembles the stable OC-type expression domainof WUS in Arabidopsis. In contrast, their expression pat-terns relate to the specification of new phytomers, whichhas 2 aspects in the grass culm. The first aspect is the re-cruitment of leaf founder cells from the whole circumfer-ence of the SAM before the leaf primordium detachesfrom the shoot apex. The second aspect is that growth ofthe culm depends on the coordinated elongation of the de-tached leaf sheath and the internode enclosed (Abbe et al.1951; Steffensen 1968). Essentially, the single rice OsWUSorthologue shares an expression pattern with its 2 maizeparalogues. However, the different expression patterns ofthe 2 paralogues in maize identified distinct functions inthe apex. ZmWUS1 is expressed in cells of the prospectivenodal/internodal tissue in the center of the SAM and shiftsto deeper layers with elaboration of the new phytomer,whereas ZmWUS2 is transcribed in cells recruited for leafprimordia and has a maximum in basal lateral leaf margins.ZmWUS2 therefore shares expression in leaf primordia andthe preference in lateral leaf margins with OsWUS, whereas
FIG. 5.—Transcriptional activity of maize ZmWUS paralogues and the rice OsWUS orthologue during the reproductive phase. (A) Types of meristemsin the maize ear: IM, SPM, SM, and upper/lower floral meristem (UFM/LFM). (B) ZmWUS1 transcription pattern in SPM, SM, and early FM. (C) UFMand LFM showing ZmWUS1 transcripts in outer layers in the older UFM, whereas transcripts are still central within the LFM, which will abort. (D) NoZmWUS1 expression is detectable in the IM. (E) Lateral longitudinal section showing the ZmWUS2 expression pattern in the SPM, SM, or FM. Differ-ences in staining intensity are due to twisting of seed rows. (F) Close-up of a FM showing ZmWUS2 expression throughout the UFM and subtending theLFM but also transcripts in the developing early stamen. (G) Median longitudinal section showing ZmWUS2 expression in the IM with a slight L1 layerpreference. (H1 I) OsWUS expression pattern in the rice primary branch meristem (PBM) or FM, respectively. Note the L1 layer preference in the PBMrelative to FM. St: stamen; ig/og: inner/outer glume.
2500 Nardmann and Werr
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/23/12/2492/974436 by guest on 20 Novem
ber 2018

ZmWUS1 may relate to the pulsing OsWUS transcription inthe SAM center. The expression patterns of the maize WUSparalogues therefore support the hypothesis that gene du-plications are preserved by subfunctionalization (Lynchand Force 2000). After duplication, mutations have resultedin a reduction in their joint expression levels and patterns ofactivity compared with that of the single ancestral WUSgene, which formerly may have been expressed similarlyto OsWUS, the single WUS orthologue in rice.
The different expression patterns of grass WUS ortho-logues compared with that of WUS in Arabidopsis acquiresignificance due to the correlated changes in the expressionpatterns of the grass CLV1 orthologues, TD1 and FON1 inmaize and rice, respectively (Suzaki et al. 2004; Bommertet al. 2005). Both orthologues are transcribed at the flank ofthe SAM in cells that have been recruited into leaf primor-dia but not in the center of the SAM where ZmWUS1 andOsWUS are transiently expressed. TD1 transcription ini-tiates at P0 and then overlaps with ZmWUS2 expressionin the P1 phytomer and lateral leaf domains after the detach-ment of the primordium. The activation of ZmWUS2 in P1
within the TD1 expression domain suggests that pluripo-tency is acquired de novo in cells already determined forthe primordial fate. The OsWUS and FON1 expression pat-terns in rice leaf primordia conform to this assumption aswell as to the assumption that the grass CLV1 orthologuesTD1 and FON1 act to antagonize ZmWUS2 or OsWUS ac-tivity in prospective primordial cells. Later in leaf develop-ment, ZmWUS2 and TD1 in maize and OsWUS and FON1in rice are cotranscribed in leaf margins. The marginal pat-terns of WUS and CLV1 orthologues in maize and rice arereminiscent of the narrow sheath1 and narrow sheath2(NS1/NS2) gene expression patterns in maize, encoding pa-ralogues of the PRS gene in Arabidopsis (Nardmann et al.2004). Maize ns1/ns2 double mutants are affected in thedevelopment of the basal lateral domains of leaves, andthe similarity in transcription pattern suggest that ZmWUS2and OsWUS may contribute to leaf development and beantagonized by TD1 and FON1, respectively.
The recent identification of ROSULATA (ROA), whichencodes the WUS orthologue in A. majus, also confirms thatin dicots, ROA is not only involved in SAM maintenancebut also acts outside the SAM (Kieffer et al. 2006). In theroa mutant, the petioles of leaves are frequently ventralizedor appear radially symmetrical and the leaf blades are lessexpanded than in wild type. The roamutants also have shortinternodes, which are comparable to deficiencies in theelongation of internodes in wus-1 mutant inflorescencesin Arabidopsis. WUS functions in dicots therefore contrib-ute to the elongation of the shoot axis, which could relate toZmWUS1 transcriptional activity within the SAM duringdevelopment of the maize leaf phytomer.
Grass WUS Orthologues Display a MeristematicExpression Pattern during Reproductive Growth
In contrast to the vegetative phase, ZmWUS1,ZmWUS2, and OsWUS are expressed in meristems duringthe reproductive phase. The expression patterns ofZmWUS2 and TD1 considerably overlap in all meristemtypes of the male or female inflorescence and exert a pref-
erence for the L1 layer in the IM, whereas transcriptionalactivity in the SPM, SM, and FM is constitutive throughoutthe meristems. In contrast, ZmWUS1 transcripts are absentin the ear or tassel IM but mark a small OC-type domain inSPM, SM, and early FM before the expression spreads tothe larger ZmWUS2/TD1 coexpression domain during laterstages of floral development. Concerning the absence ofZmWUS1 activity in the IM, it should be borne in mind thatboth maize paralogues are coexpressed during early stagesof axillary meristem development as is observed during theinitiation of SPMs, SMs, and FMs. Although indeterminate,the IMs of ear or tassel are elaborated meristems, andZmWUS2 activity may be sufficient for their maintenance.Similar to that in maize, FON1 and OsWUS are coexpressedin BMs and FMs of the rice panicle. WUS and CLV1 ortho-logues of maize and rice are therefore coexpressed in allreproductive meristems, where fasciation and supernumer-ary floral organs in td1 or fon1 loss-of-function mutantsprovide evidence for meristem enlargement (Suzaki et al.2004; Bommert et al. 2005).
Conforming to the stem cell concept in Arabidopsis,excessive WUS activity could explain the meristem enlarge-ment in clv mutants (Schoof et al. 2000; Waites and Simon2000). However, differences in the expression patterns ofmaize WUS paralogues could not be detected in the td1mutant background (data not shown), whereasWUS expres-sion shifts from the L3 to the L2 layer in Arabidopsis(Schoof et al. 2000). This may relate to differences inthe shoot meristem architecture between maize and Arabi-dopsis. Whereas the OC-type, WUS expression domain inthe Arabidopsis SAM subtends a 2-layered tunica, maizemeristems possess only a single tunica layer (Abbe et al.1951; Steffensen 1968). High levels of TD1 or ZmWUS2transcript are detected in this single tunica layer of the ma-ture IM, and both genes are cotranscribed throughout alllayers of the descending SPMs, SMs, or FMs. ZmWUS2transcription, therefore, is not restricted to a subdomainof the TD1 expression domain in reproductive meristemsbut overlaps with TD1 expression and shares its transcrip-tion specificity with IM L1 layer or leaf margins. A pref-erence for the tunica layer and the leaf margin suggeststhat an extracellular ligand must be preferentially perceivedfrom subtending cell layers, in contrast to ArabidopsisSAM, where CLV3 is presented top-down from the L1/L2 tunica to the L3 layer.
The expression patterns of TD1 and FON1 during thereproductive phase and the mutant phenotypes suggest thatthey may control activity of grass WUS orthologues in re-productive meristems. By analogy, this assumption also im-plies that TD1 andFON1may restrict ZmWUS2 andOsWUSactivity in cells recruited for leaf primordia, where the ex-pression domains overlap during the vegetative phase.Whereas the Arabidopsis leaf can be traced back to a fewfounder cells within the apex (Irish and Sussex 1992), cellsfor the grass leaf are recruited from the whole circumferenceof the SAM (Poethig 1984). The strikingly different expres-sion patterns of grass WUS/CLV1 orthologues relative tothose in Arabidopsis during the vegetative phase thereforemay relate to fundamentally different developmental pro-grams and the peculiar architecture of the grass culm. How-ever, the maize or rice vegetative expression patterns signify
WUSCHEL Orthologues in Maize and Rice 2501
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/23/12/2492/974436 by guest on 20 Novem
ber 2018

an evolutionary freedom to recruit the WUS/CLV1 feed-back circuit for developmental pathways at the peripheryof the SAM without affecting stem cell homeostasis.
The Evolution of the Stem Cell Niche andWUS/CLV1 Coexpression
The present data set based on 2 grass species raisesconcerns about the general applicability of a concept basedon 2 major signaling pathways to control stem cell homeo-stasis in the shoot as is suggested by the WUS/CLV antag-onism in Arabidopsis. The similarity between the variousrice and maize expression patterns implies that the differ-ences existed prior to the separation of the 2 grass lineagesOryzoideae and Panicoideae and may be specific forgrasses. It would be intriguing to establish the WUS/CLVexpression patterns in the third grass radiation, the Pooi-deae. However, information pertaining to an even moreancestral situation would be obtained from analysis of pre-sumptive basal angiosperms such as Amborella, consideredto be a sister to both the dicot and the monocot lineage ofangiosperms (Qiu et al. 1999; Soltis et al. 1999). A likelyscenario is that stem cell homeostasis in an ancestral angio-sperm precursor involved multiple pathways acting inparallel to specify stem cell fate and to maintain the stemcell niche. Recent data from Arabidopsis support this view,where STIMPY (STIP or WOX9) and SPLAYED (SYD) actthrough WUS and presumably contribute to the stem cell–promoting pathway (Kwon et al. 2005; Wu et al. 2005).Interestingly, our phylogenetic analysis revealed 3WOX9 orthologues to exist in maize and 2 in rice, whichmay contribute similar or additional functions. Alterna-tively, the class III AtHD-Zip protein CORONA (CNA)acts in parallel to CLV signaling and thus independentlylimits stem cell number (Green et al. 2005). These data in-dicate that redundantly acting pathways are functional inArabidopsis and the maintenance of stem cell homeostasisin the SAM by WUS/CLV signaling may represent onlyone outcome of evolutionary selection. In an ancestralangiosperm species, the expression of WUS and CLV1orthologues may consequently have been restricted todeveloping new phytomers without affecting stem cell ho-meostasis in the shoot apex. Cotranscription of ZmWUS2/TD1 and OsWUS/FON1 throughout the plant life cycleand the accompanying changes from leaf-associated tomeristem-specific patterns suggest that the entire feedbackloop may be an ancient phenomenon.
The absence of TD1 or FON1 transcripts in the centerof the vegetative apex raises the question how ZmWUS1 orOsWUS activity is controlled in the SAM. Phylogenetically,the TD1 and FON1 proteins group within 1 branch togetherwith CLV1 (Bommert et al. 2005; DeYoung et al. 2006).However, this unique CLV1 branch is rooted together witha second branch B (Bommert et al. 2005), which containsadditional and closely related transmembrane receptor ki-nase proteins encoded by the genomes of maize, rice, andArabidopsis. Three closely CLV1-related genes BAM1–3(barely any meristem1–3) have recently been described inArabidopsis to be involved in multiple developmental pro-cesses including leaf development (DeYoung et al. 2006).Still ongoing research on CLV1 relatives in maize and rice
indicates that the subbranch B contains additional grass-specific kinase gene family members (Durantini D and WerrW, unpublished data), which root independently from theirclosest dicot relatives. Depending on their expression do-main, such grass-specific transmembrane receptor kinasefamily members could be candidates for the control ofthe dynamic ZmWUS1 or the transient OsWUS expressionin the center of the grass SAM. However, the identificationof such grass-specific receptor kinase candidates also raisesthe question how stem cell homeostasis in the maize or riceSAM is positively controlled during the vegetative phase,when a stem cell OC is not provided by expression ofWUS orthologues in grasses. Possibly, one of the otherWOX gene family members has been recruited for this pur-pose, or other signaling pathways, which have not yet beenidentified, may be involved.
In summary, the transcription patterns of WUS andCLV1 orthologues in maize and rice imply that the feedbackmechanism controlling stem cell homeostasis in the Arabi-dopsis SAM has been subject to significant adaptationsin the course of plant speciation. The recruitment ofZmWUS2/OsWUS and TD1/FON1 transcription to leaf phy-tomer development during the vegetative phase in maize aswell as rice, without affecting SAM function, implies thatother pathways to maintain stem cell homeostasis exist ingrass shoot meristems. The expression patterns imply 2 dis-tinct functions in the grass phytomer: one in the center ofthe apex and the other at the SAM periphery, where cells arerecruited into primordia. The data from grasses raise someconcern about the generalization of a shoot stem cell nichebased on only 2 major antagonistic functions. However, thecorrelation of transcription patterns strongly supports thatthe CLV1/WUS antagonism is an ancient angiospermfeature and may still be used to control the activity ofWUS orthologues in rice and maize such as it does inArabidopsis.
Supplementary Material
Supplementary material is available at Molec-ular Biology and Evolution online (http://www.mbe.oxfordjournals.org/).
Acknowledgments
We thank Dr J. Chandler for suggestions and stimu-lating discussions and critically reading the manuscript.The maize work was supported by the European Commis-sion (QLK3-2000-00196 and QLRT-2000-00302) and theDeutsche Forschungsgemeinschaft (WE 1262/5-1).
Literature Cited
Abbe EC, Phinney BO, Baer DF. 1951. The growth of the shootapex in maize: internal features. Am J Bot. 38:744–752.
Bomblies K, Wang RL, Ambrose BA, Schmidt RJ, Meeley RB,Doebley J. 2003. Duplicate FLORICAULA/LEAFY homologszfl1 and zfl2 control inflorescence architecture and flower pat-terning in maize. Development. 130:2385–2395.
Bommert P, Lunde C, Nardmann J, Vollbrecht E, Running M,Jackson D, Hake S, Werr W. 2005. thick tassel dwarf1 encodesa putative maize ortholog of the Arabidopsis CLAVATA1
2502 Nardmann and Werr
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/23/12/2492/974436 by guest on 20 Novem
ber 2018

leucine-rich repeat receptor-like kinase. Development. 132:1235–1245.
Bradley D, Carpenter R, Sommer H, Hartley N, Coen E. 1993.Complementary floral homeotic phenotypes result from oppo-site orientations of a transposon at the plena locus of Antirrhi-num. Cell. 72:85–95.
Brand U, Fletcher JC, Hobe M, Meyerowitz EM, Simon R. 2000.Dependence of stem cell fate in Arabidopsis on a feedback loopregulated by CLV3 activity. Science. 289:617–619.
Byrne ME, Barley R, Curtis M, Arroyo JM, Dunham M, HudsonA, Martienssen RA. 2000. Asymmetric leaves1 mediates leafpatterning and stem cell function in Arabidopsis. Nature.408:967–971.
Chomczynski P, Sacchi N. 1987. Single-step method of RNA iso-lation by acid guanidinium thiocyanate-phenol-chloroformextraction. Anal Biochem. 162:156–159.
Clark SE, Williams RW, Meyerowitz EM. 1997. The CLAVATA1gene encodes a putative receptor kinase that controls shoot andfloral meristem size in Arabidopsis. Cell. 89:575–585.
DeYoung BJ, Bickle KL, Schrage KL, Muskett P, Patel K, ClarkSE. 2006. The CLAVATA1-related BAM1, BAM2 and BAM3receptor kinase-like proteins are required for meristem functionin Arabidopsis. Plant J. 45:1–16.
Doebley J. 2004. The genetics of maize evolution. Annu RevGenet. 38:37–59.
Doebley J, Stec A, Hubbard L. 1997. The evolution of apical dom-inance in maize. Nature. 386:485–488.
Evans MM, Barton MK. 1997. Genetics of angiosperm shoot api-cal meristem development. Annu Rev Plant Physiol Plant MolBiol. 48:673–701.
Fletcher JC, Brand U, Running MP, Simon R, Meyerowitz EM.1999. Signaling of cell fate decisions by CLAVATA3 inArabidopsis shoot meristems. Science. 283:1911–1914.
Goff SA, Ricke D, Lan TH, et al. (55 co-authors). 2002. A draftsequence of the rice genome (Oryza sativa L. ssp. japonica).Science. 296:92–100.
Green KA, Prigge MJ, Katzman RB, Clark SE. 2005. CORONA,a member of the class III homeodomain leucine zipper genefamily in Arabidopsis, regulates stem cell specification andorganogenesis. Plant Cell. 17:691–704.
Haecker A, Gross-Hardt R, Geiges B, Sarkar A, Breuninger H,Herrmann M, Laux T. 2004. Expression dynamics ofWOX genes mark cell fate decisions during early embryonicpatterning in Arabidopsis thaliana. Development. 131:657–668.
Harrison CJ, Corley SB, Moylan EC, Alexander DL, ScotlandRW, Langdale JA. 2005. Independent recruitment of aconserved developmental mechanism during leaf evolution.Nature. 434:509–514.
Irish VF, Sussex IM. 1992. A fate map of the Arabidosis embry-onic shoot apical meristem. Development. 115:745–753.
Itoh J, Nonomura K, Ikeda K, Yamaki S, Inukai Y, Yamagishi H,Kitano H, Nagato Y. 2005. Rice plant development: fromzygote to spikelet. Plant Cell Physiol. 46:23–47.
Jackson D. 1991. In situ hybridization in plants. In: Bowles DJ,Gurr SJ, McPerson M, editors. Molecular plant pathology:a practical approach. Oxford: Oxford University Press.p. 163–174.
Jeong S, Trotochaud AE, Clark SE. 1999. The Arabidopsis CLAV-ATA2 gene encodes a receptor-like protein required for the sta-bility of the CLAVATA1 receptor-like kinase. Plant Cell. 11:1925–1934.
Kieffer M, Stern Y, Cook H, Clerici E, Maulbetsch C, Laux T,Davies B. 2006. Analysis of the transcription factor WU-SCHEL and its functional homologue in Antirrhinum revealsa potential mechanism for their roles in meristem maintenance.Plant Cell. 18:560–573.
Kwon CS, Chen C, Wagner D. 2005. WUSCHEL is a primarytarget for transcriptional regulation by SPLAYED in dynamiccontrol of stem cell fate in Arabidopsis. Genes Dev. 19:992–1003.
Lynch M, Force A. 2000. The probability of duplicate gene pres-ervation by subfunctionalization. Genetics. 154:459–473.
Matsumoto N, Okada K. 2001. A homeobox gene, PRESSEDFLOWER, regulates lateral axis-dependent development ofArabidopsis flowers. Genes Dev. 15:3355–3364.
Mayer KF, Schoof H, Haecker A, Lenhard M, Jurgens G, Laux T.1998. Role of WUSCHEL in regulating stem cell fate in theArabidopsis shoot meristem. Cell. 95:805–815.
Nardmann J, Ji J, Werr W, Scanlon MJ. 2004. The maize duplicategenes narrow sheath1 and narrow sheath2 encode a conservedhomeobox gene function in a lateral domain of shoot apicalmeristems. Development. 131:2827–2839.
Ohta M, Matsui K, Hiratsu K, Shinshi H, Ohme-Takagi M. 2001.Repression domains of class II ERF transcriptional repressorsshare an essential motif for active repression. Plant Cell. 13:1959–1968.
Piazza P, Jasinski S, Tsiantis M. 2005. Evolution of leaf develop-mental mechanisms. New Phytol. 167:693–710.
PoethigS.1984.Cellularparametersof leafmorphogenesis inmaizeand tobacco. In: White R, Dickinson WC, editors. Contemporyproblems in plant anatomy. New York: Academic Press.p. 235–259.
Qiu YL, Lee J, Bernasconi-Quadroni F, Soltis DE, Soltis PS, ZanisM, Zimmer EA, Chen Z, Savolainen V, Chase MW. 1999. Theearliest angiosperms: evidence from mitochondrial, plastid andnuclear genomes. Nature. 402:404–407.
Scanlon MJ. 2000. NARROW SHEATH1 functions from two mer-istematic foci during founder-cell recruitment in maize leafdevelopment. Development. 127:4573–4585.
Schoof H, Lenhard M, Haecker A, Mayer KF, Jurgens G, Laux T.2000. The stem cell population of Arabidopsis shoot meristemsis maintained by a regulatory loop between the CLAVATA andWUSCHEL genes. Cell. 100:635–644.
Soltis PS, Soltis DE, Chase MW. 1999. Angiosperm phylogenyinferred from multiple genes as a tool for comparative biology.Nature. 402:402–404.
Steffensen DM. 1968. A reconstruction of cell development in theshoot apex of maize. Am J Bot. 55:354–369.
Stuurman J, Jaggi F, Kuhlemeier C. 2002. Shoot meristem main-tenance is controlled by a GRAS-gene mediated signal fromdifferentiating cells. Genes Dev. 16:2213–2218.
Suzaki T, Sato M, Ashikari M, Miyoshi M, Nagato Y, Hirano HY.2004. The gene FLORAL ORGAN NUMBER1 regulates floralmeristem size in rice and encodes a leucine-rich repeat receptorkinase orthologous to Arabidopsis CLAVATA1. Develop-ment. 131:5649–5657.
Sylvester AW, Cande WZ, Freeling M. 1990. Division and differ-entiation during normal and liguleless-J maize leaf develop-ment. Development. 110:985–1000.
Szymkowiak EJ, Sussex IM. 1996. What chimeras can tell usabout plant development. Annu Rev Plant Physiol PlantMol Biol. 47:351–376.
Taguchi-Shiobara F, Yuan Z, Hake S, Jackson D. 2001. The fas-ciated ear2 gene encodes a leucine-rich repeat receptor-likeprotein that regulates shoot meristem proliferation in maize.Genes Dev. 15:2755–2766.
Timmermans MC, Hudson A, Becraft PW, Nelson T. 1999.ROUGH SHEATH2: a Myb protein that represses knoxhomeobox genes in maize lateral organ primordia. Science.284:151–153.
Tsiantis M, Schneeberger R, Golz JF, Freeling M, Langdale JA.1999. The maize rough sheath2 gene and leaf developmentprograms in monocot and dicot plants. Science. 284:154–156.
WUSCHEL Orthologues in Maize and Rice 2503
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/23/12/2492/974436 by guest on 20 Novem
ber 2018

Vollbrecht E, Kerstetter R, Lowe B, Veit B, Hake S. 1993. Ho-meobox genes in plant development: mutational and molecularanalysis. Evolutionary conservation of developmental mecha-nism. Wiley-Liss, Inc. p. 111–123.
Waites R, Selvadurai HR, Oliver IR, Hudson A. 1998. The PHAN-TASTICA gene encodes a MYB transcription factor involved ingrowth and dorsoventrality of lateral organs in Antirrhinum.Cell. 93:779–789.
Waites R, Simon R. 2000. Signaling cell fate in plant meristems.Three clubs on one tousle. Cell. 103:835–838.
Wang H, Nussbaum-Wagler T, Li B, Zhao Q, Vigouroux Y, FallerM, Bomblies K, Lukens L, Doebley JF. 2005. The origin of thenaked grains of maize. Nature. 436:714–719.
Whipple CJ, Ciceri P, Padilla CM, Ambrose BA, Bandong SL,Schmidt RJ. 2004. Conservation of B-class floral homeoticgene function between maize and Arabidopsis. Development.131:6083–6091.
Wikstrom N, Kenrick P. 2001. Evolution of Lycopodiaceae(Lycopsida): estimating divergence times from rbcL genesequences by use of nonparametric rate smoothing. Mol Phy-logenet Evol. 19:177–186.
Wu X, Dabi T, Weigel D. 2005. Requirement of homeobox geneSTIMPY/WOX9 for Arabidopsis meristem growth and mainte-nance. Curr Biol. 15:436–440.
Zahn LM, Leebens-Mack J, DePamphilis CW, Ma H, Theissen G.2005. To B or not to B a flower: the role of DEFICIENS andGLOBOSA orthologs in the evolution of the angiosperms.J Hered. 96:225–240.
Neelima Sinha, Associate Editor
Accepted September 18, 2006
2504 Nardmann and Werr
Dow
nloaded from https://academ
ic.oup.com/m
be/article-abstract/23/12/2492/974436 by guest on 20 Novem
ber 2018