Embed Size (px)
Citation preview

MOLECULAR AND CELLULAR BIOLOGY, June 1992, p. 2662-26720270-7306/92/062662-11$02.00/0Copyright © 1992, American Society for Microbiology
The Promoter of the CD19 Gene Is a Target for the B-Cell-SpecificTranscription Factor BSAP
ZBYNEK KOZMIK, SHIRLEY WANG, PETRA DORFLER, BEN ADAMS,' AND MEINRAD BUSSLINGER*
Institute of Molecular Pathology, Dr. Bohr-Gasse 7, A-1030 Vienna, Austria
Received 5 December 1991/Accepted 4 March 1992
The CD19 protein is expressed on the surface of all B-lymphoid cells with the exception of terminallydifferentiated plasma cells and has been implicated as a signal-transducing receptor in the control ofproliferation and differentiation. Here we demonstrate complete correlation between the expression pattern ofthe CD19 gene and the B-cell-specific transcription factor BSAP in a large panel of B-lymphoid cell lines. Thehuman CD19 gene has been cloned, and several BSAP-binding sites have been mapped by in vitro protein-DNAbinding studies. In particular, a high-affinity BSAP-binding site instead of a TATA sequence is located in the-30 promoter region upstream of a cluster of heterogeneous transcription start sites. Moreover, this site isoccupied by BSAP in vivo in a CD19-expressing B-cell line but not in plasma or HeLa cells. This high-affinitysite has been conserved in the promoters of both human and mouse CD19 genes and was furthermore shownto confer B-cell specificity to a f-globin reporter gene in transient transfection experiments. In addition, BSAPwas found to be the only abundant DNA-binding activity of B-cell nuclear extracts that interacts with the CD19promoter. Together, this evidence strongly implicates BSAP in the regulation of the CD19 gene.
The differentiation of B lymphocytes from progenitor cellsto immunoglobulin-secreting plasma cells is a multistepprocess which is characterized by the sequential expressionof specific cell surface markers involved in cell-cell interac-tion and signal transduction (29). Among them is the B-cell-specific antigen CD19. This protein is expressed throughoutB-cell ontogeny from the early progenitor cell up to themature B-cell stage and is lost from the cell surface only atthe terminal stages of B-cell differentiation. The CD19 mol-ecule is present on both normal and malignant B-lymphoidcells and hence is the most reliable diagnostic surface markerof the human B-cell lineage (23, 35). The CD19 gene codesfor a 95-kDa protein with two extracellular immunoglobulin-like domains, a short transmembrane region, and a largecytoplasmic tail (33, 34, 38). These features suggest that theCD19 protein is a membrane receptor involved in signaltransduction of B lymphocytes. No natural ligand for CD19is as yet known; instead, monoclonal anti-CD19 antibodieshave been used to study the physiological function of theCD19 molecule. In agreement with the receptor hypothesis,antibody binding leads to rapid internalization of CD19 (7,35) and to mobilization of free intracellular calcium ions (17,27, 35). CD19 antibodies strongly inhibit proliferation ofresting mature B cells in response to stimulation with anti-immunoglobulin M (IgM) antibodies (11, 27, 35). Theyprovide, however, a positive proliferative signal for earlyprecursor B cells (17), indicating that CD19 is involved inpositive or negative control of proliferation, depending onthe stage of B-cell differentiation. Transmembrane signalingvia the CD19 receptor is also important for late events ofB-cell differentiation, as CD19 antibodies block in vitrodifferentiation of mature B cells to plasma cells (7, 11). CD19specifically associates with at least four other cell surfaceproteins to form a multimolecular complex which includesthe B-cell-specific complement receptor type 2 (CD21) in
* Corresponding author.t Present address: University of Sussex, Falmer, Brighton BN1
9RN, England.
mature B cells (20). Activation of the CD19-CD21 complexhas recently been shown to potentiate the primary responseof B cells to antigen in vivo (14).
Regulation of B-cell-specific gene expression is broughtabout by several distinct transcription factors (reference 3and references therein). One of them is the B-cell lineage-specific activator protein (BSAP). We have identified thistranscription factor as a mammalian homolog of the sea
urchin protein TSAP, which is responsible for the develop-mental and tissue-specific regulation of two nonallelic pairsof late histone H2A-2 and H2B-2 genes (2). The four TSAP-binding sites present in these histone gene promoters are
recognized by TSAP with equal affinity, and yet they differconsiderably in primary sequence, allowing the definition ofonly a degenerate consensus sequence (2). The mammalianfactor BSAP interacts with all four TSAP-binding sites in a
manner indistinguishable from that of the sea urchin protein.Within the hematopoietic system, BSAP is exclusively ex-
pressed in the B-cell lineage with equal binding activity inpro-B, pre-B, and mature B cells, while it is undetectable interminally differentiated plasma cells. BSAP was shown toact as a positive transcription factor on an artificial promotercontaining a sea urchin TSAP-binding site. Known regula-tory regions of immunoglobulin and class II major histocom-patibility complex genes, however, lack high-affinity BSAP-binding sites, suggesting that BSAP regulates a different setof B-lymphoid-specific genes (3).Here we describe the cloning and characterization of the
human CD19 gene and show by in vitro and in vivo protein-DNA binding studies that the CD19 promoter is a target forthe B-cell-specific transcription factor BSAP.
MATERIALS AND METHODS
Cell lines. The cell lines Raji, Ramos, MC/CAR, IM-9,HS-Sultan, and RPMI 8226 were obtained from AmericanType Culture Collection (Rockville, Md.) and grown inRPMI 1640 medium supplemented with 10% fetal calf serum.The origin of the other cell lines used is described byBarberis et al. (3).
2662
Vol. 12, No. 6
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

REGULATION OF THE CD19 GENE BY BSAP 2663
Screening of cosmid library, subcloning, and DNA sequenc-ing. A human cosmid library constructed in the vectorpcos2-EMBL (8) was obtained from A.-M. Frischauf (Lon-don, England) and screened with randomly primed insertDNA of a CD19 cDNA clone (33) according to standardprocedures (19). The restriction fragments shown in Fig. 2were subcloned into plasmid pUC18. The DNA sequences ofboth DNA strands were determined on an automated se-quencer (Applied Biosystems, model 373A) or by the stan-dard dideoxy technique (31).
Cloning of mouse CD19 promoter. The mouse CD19 pro-moter was cloned by ligation-mediated polymerase chainreaction (PCR) as described in detail by Fors et al. (9).CD19-specific oligonucleotide primers were derived from thepublished mouse CD19 cDNA sequence (38). PstI-digestedDNA of NIH 3T3 cells and the oligonucleotide CATGGCAGCACAACATTGC were used for the Sequenase reaction,and the double-stranded linker described by Mueller andWold (22) was used for the T4 DNA ligase reaction. Theoligonucleotide TACCTCCACCAGTAAGGACTlCTGG wasused for the first amplification and the overlapping oligonu-cleotide ACGAGGATCCACCAGTAAGGACTTCTGGGGCCTG was used for the second amplification by PCR,followed by EcoRI and BamHI digestion. The 670-bp PCRfragment was cloned into plasmid pSP65 and analyzed byDNA sequencing.
In vitro protein-DNA binding assays. Standard and multiplebandshift assays were performed as described by Barberis etal. (2) and by Kozmik and Paces (16), respectively. DNase Ifootprint reactions were carried out as described previously(la). For methylation protection analysis, a 3'-end-labeledDNA fragment (-1 fmol; 3,000 cpm) was incubated with-40 ,ug of nuclear protein and 1 ,ug of poly(dI-dC) in 20 ,ul ofbandshift buffer containing 10 mM N-2-hydroxyethylpipera-zine-N'-2-ethanesulfonic acid (HEPES; pH 7.9), 100 mMKCl, 4% Ficoll, and 1 mM EDTA. After 10 min at roomtemperature, 1 ,ul of dimethyl sulfate (DMS; diluted 1:50 indistilled H20) was added for 1 min, the reaction was inacti-vated by the addition of 4 ,ul of 1 M ,-mercaptoethanol-1.5M sodium acetate, and the mixture was subjected to byphenol extraction, ethanol precipitation, piperidine cleav-age, and analysis on an 8% sequencing gel.
In vivo footprinting. The in vivo footprinting reaction wascarried out according to the protocol of Mueller and Wold(22). The oligonucleotide GGTAAGTTACTCAGCCAAGGACACC was used as genomic primer for the Sequenasereaction, the internal oligonucleotide TACCTTCCACCTTCACCACTAGAGG was used for exponential amplification byPCR, and the 5'-end-labeled oligonucleotide TTCCACCTTCACCACTAGAGGTTCCTC was used for detection of thesequence ladder by linear PCR amplification. The double-stranded linker described by Mueller and Wold (22) was usedfor the T4 DNA ligase reaction.
Si nuclease and RNase protection analyses. Si nuclease andRNase protection experiments were carried out as describedby Busslinger et al. (5) and Vitelli et al. (36), respectively.The human CD19 probe used for the experiment shown inFig. 1 was generated by subcloning a 513-bp SmaI-MscIfragment of the CD19 cDNA clone (33) into the Hindll site ofpSP64 in antisense orientation relative to SP6 transcription.For S1 mapping, a 539-bp NcoI-PvuII fragment of thissubclone was labeled at the NcoI site. The mouse CD19 Siprobe was obtained by annealing and cloning of the twooligonucleotides AAYICTATGA-93 nucleotides-CTCCTTCTCCtgca and GGAGAAGGAG-93 nucleotides-TCATAGderived from the mouse CD19 cDNA sequence (34; 5' and 3'
nucleotides indicated only) into the EcoRI and PstI sites ofpSP64. A 262-bp NcoI-PvuII fragment of this subclone was5' end labeled at the NcoI site and used as the S1 probe.RACE analysis. The rapid analysis of cDNA ends (RACE)
analysis was performed as described in detail by Frohman etal. (10). The CD19-specific oligonucleotide TTGAGAGACGTTGAAGATGA was used to prime the reverse transcriptionof -5 ,ug of poly(A)+ RNA from BJA-B cells, followed byRNA hydrolysis with 150 mM NaOH for 20 min at 60'C,cDNA isolation by chromatography on a Sephadex SP-C50column, and ethanol precipitation. The cDNA was tailedwith dATP and terminal deoxynucleotidyltransferase, fol-lowed by second-strand synthesis with Klenow DNA poly-merase I and the primer TTGAGGCACTGCAGCA(T)17.CD19 sequences were amplified by PCR with the oligonu-cleotide TTGAGGCACTGCAGCA, and the PstI-digestedPCR products were cloned into the PstI site of plasmidpSP64. CD19 cDNA clones were identified by colony hy-bridization with the end-labeled oligonucleotide TGACTTCCATGGGGGTGA and sequenced.
Transient transfection assay. Nine micrograms of the testplasmid OVEC-S or OVEC-CD19 and 1 ,ug of the controlplasmid OVEC-Ref were transfected into BJA-B, J558L, andSP2/0 cells as described previously (3). Cytoplasmic RNAwas extracted 48 h later, and 10 ,ug of RNA was analyzed byRNase protection assay with a rabbit 3-globin SP6 probe asdescribed previously (3).
Nucleotide sequence accession number. The DNA sequenceshown in Fig. 2 has been submitted to the GenEMBL database and assigned accession number M84371. The humanand mouse CD19 DNA sequences (Fig. 9) have been sub-mitted to the GenEMBL data base and assigned accessionnumbers M84371 and M84372.
RESULTS
Strict correlation between CD19 gene expression and BSAPactivity in B-cell lines. A literature search previously indi-cated that the expression pattern of the CD19 gene may besimilar to that of the B-cell-specific transcription factorBSAP, suggesting an involvement of BSAP in the regulationof this gene (3). As a first step towards testing this hypoth-esis, we have investigated this correlation in more detail. Alarge panel of murine and human B-lymphoid cell linespreviously analyzed for BSAP-binding activity (3) was thusanalyzed for CD19 gene expression by S1 nuclease mapping(Fig. 1). Si probes that map an internal portion of CD19mRNA were generated by subcloning small fragments ofmurine and human CD19 cDNAs (see Materials and Meth-ods). CD19 signals were normalized by reference to glycer-aldehyde phosphate dehydrogenase (gapdh) mRNA. Asshown in Fig. 1, the CD19 gene is expressed in all pro-B-,pre-B-, and B-cell lines but not in plasma cells, with twoexceptions. The human plasma cell line HS-Sultan expressesCD19 mRNA, while this transcript is absent from the murinecell line LyD9, which was originally described as a pro-B-cell line (25) and was subsequently identified as a multipo-tential bone marrow stem cell clone (18). There is, however,complete correlation between BSAP and CD19 gene expres-sion in both the mouse and human systems, as all CD19-positive cell lines contain BSAP-binding activity, whereas allCD19-negative cell lines lack this activity (3). For all mouseB-lymphoid cells, this correlation extends even to the levelof gene expression, since the CD19-expressing cell linescontain not only the same amount of CD19 mRNA (Fig. 1A)but also similar levels of BSAP-binding activity (3). The
VOL. 12, 1992
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

2664 KOZMIK ET AL.
pro-B pre-B B-cells plasma cells
"-rn
M.i-O r- c- () C
CD D
L CL CD r c
Mn< <> 0 Q RlB3C 0 EL0 O L500-404- ,_353--
242 -
input
gapdh
190- *4
B
500 -
plasmapro-B pre-B B-cells cells
-tJCo J
Xn< u << < D)3 CLXMLu z c m zm a E I D IesSflamflinS
404 -
353 -
a--242 -
147- _f 500- _ _404- - _ .M
.....
40.4- _ _ __
353 - _t _-uI_ _
89- _ _-1I
0CD19=
242-
+ + + + + + + + + + BSAP67-7
- + + + + + + + + BSAP
FIG. 1. CDi9 gene expression in murine and human B-lymphoid cell lines. (A) Murine B-lymphoid cell lines. Total RNA (15 ,ug) of eachcell line was simultaneously analyzed by S1 nuclease mapping with a 5'-end-labeled mouse CDi9 and a rat gapdh S1 probe (see Materials andMethods). The specific radioactivity of the gapdh probe was 10-fold lower than that of the CDi9 probe. Si-resistant DNA fragments wereseparated on an 8% polyacrylamide sequencing gel together with an end-labeled DNA size marker (lane M; pUC18 digested with MspI). Inputdenotes the position of the two intact Si DNA probes. Correct protection by the CD19 mRNA results in an 88-nucleotide-long Si-resistantDNA fragment. The triplet below it is generated by Si digestion into an A+T-rich sequence of the RNA-DNA hybrid. All cell lines werepreviously analyzed for the presence (+) or absence (-) of BSAP-binding activity, as indicated at the bottom (3). (B) Human B-lymphoid celllines. Total RNA (20 ,ug) of each cell line was separately analyzed with a human gapdh and a human CD19 Si probe. Bandshift analysis ofnuclear extracts with a sea urchin TSAP probe revealed the same amount (+) of BSAP activity in the cell lines Ramos, MC/CAR, IM-9, andHS-Sultan as in the other previously analyzed cell lines (3), while RPMI 8226 cells lack BSAP activity (-) (unpublished data). Theautoradiographic exposure time for both CD19 Si gels was 7 days at -70°C. Sizes are indicated in nucleotides.
strict correlation between CD19 and BSAP expression sup-ports our hypothesis that the CD19 gene is a target for BSAPregulation and predicts that a BSAP-binding site(s) should bepart of the regulatory region of the CD19 gene. An alterna-tive explanation is that another transcription factor with thesame expression profile regulates both the CD19 and BSAPgenes. To distinguish between these two possibilities, wecloned and analyzed the human CD19 gene.
Cloning and characterization of the human CD19 gene. The
CD19 gene was isolated from a human genomic cosmidlibrary by screening with a homologous cDNA probe (33).One of several positive cosmid clones was further charac-terized by subcloning and DNA sequencing. Comparisonwith published cDNA sequences (33, 34) revealed that thehuman CD19 gene is 7.37 kb long and consists of 15 exons(Fig. 2). The first four exons encode the N-terminal extra-cellular domain, exon 5 contains the transmembrane region,and exons 6 to 14 code for the intracellular portion of the
Go
1 2 3 4
m
TM
5 6 7 8 9 101112 13 14 15
HA
1 kb
I- XS
AX
FIG. 2. Exon-intron structure of the human CD19 gene. Coding and noncoding exons are indicated by black and open boxes, respectively.A thick line denotes the regions which have been sequenced. Vector sequences are indicated by a wavy line. Only the restriction enzyme sitesused for generating the three subclones are shown. TM, transmembrane domain.
- input
- gapdh
input
- CD19
El m
Alm 1: .: 1-: ,:-_---;,zi-
111.111 'I ..-!:;
MOL. CELL. BIOL.
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

REGULATION OF THE CD19 GENE BY BSAP 2665
ACDcm
Ita:co
<c
94-
79-
CD195ends
BCD
ca
4y:. 5.
110- P
84- 4i
67-~*.........
_- _U __ m4 gapdh
C1- published cDNA
RACE CCTCCAGGCCCCTGCCTGCCCCAGCATCCCCTGCGCGAAGCTGGGTGCCCCGGAGAGTCTGACCACCATG
vv v TTV V TVVYTTT VT VvY V VCC TCCAGGCCCCTGCCTGCCCCAGCATCCCCTGCGCGAAGCTGGGTGCCCCGGAGAGTCTGACCACCATGSi GGAGGTCCGGGGACCGACGGGGTCGTAGGGGACGCGCTTCGACCCACGGGGCCTCTCAGACTGGTGG
FIG. 3. Mapping of the initiation sites of CD19 gene transcription. (A) Si nuclease analysis. Poly(A)+ RNA (-5 pg) of each cell line wasanalyzed by S1 mapping with a DNA probe that was end labeled at anAvaI site in the first exon of the CD19 gene and that extends 300 bpup to the next XbaI sites in the promoter. Si-resistant DNA fragments were electrophoresed on a denaturing 8% polyacrylamide gel togetherwith the cleavage products of the G+A sequencing reaction of the same DNA probe. One-tenth of each poly(A)+ RNA (-0.5 ,ug) wasstandardized in a separate Si experiment with a human gapdh DNA probe. The nucleotide positions relative to the labeled AvaI end areindicated at the left. (B) RNase protection experiment. Total RNA (20 pg) of BJA-B and RPMI 8226 cells was hybridized to a riboprobecontaining the CD19 sequences from the AvaI site (exon 1) to the XbaI site (promoter) and then subjected to RNase digestion and analysison an 8% sequencing gel as described previously (36). An end-labeled MspI digest of pUC19 DNA was used as size marker (lane M). (C)Summary of 5'-end mapping. The RACE method was used to clone the 5'-terminal sequences of CD19 mRNA from BJA-B cells. Twenty-onecDNA clones were sequenced, and their starting nucleotides are indicated by arrows. The published cDNA sequence (33) starts 15 nucleotidesupstream of the translation start codon, which is underlined. The transcription initiation sites as mapped by Si nuclease analysis are indicatedbelow by filled and open arrowheads corresponding to intense and weak Si signals, respectively. The number of transcription start sites mayhave been overestimated by Si nuclease mapping in comparison with the RNase protection experiment, possibly because of "nibbling" ofthe Si nuclease at the 5' end of the protected DNA fragments.
CD19 protein. The CD19 cosmid clone analyzed contains 1.3kb and -30 kb of 5' and 3' flanking sequences, respectively.
Heterogeneous initiation ofCD19 gene transcription. The 5'ends of the relatively rare CD19 mRNA were mapped bythree different, highly sensitive methods (Fig. 3). Poly(A)+RNA was used for Si nuclease analysis with a DNA probethat was 5' end labeled at the AvaI site in the first exon (Fig.3A). An identical pattern of heterogeneous start sites wasobserved for three CD19-positive cell lines (MC/CAR, REH,BJA-B). Alignment with the control sequence ladder (G+A)mapped these start sites within a 40-bp-long, highly G+C-rich DNA sequence immediately adjacent to the translationinitiation codon (Fig. 3C). A similar pattern of heteroge-neous start sites was obtained by RNase protection analysisof BJA-B RNA with an antisense RNA probe that spans thesame sequences used for Si mapping (Fig. 3B). Finally, wehave identified the 5' ends of CD19 transcripts in BJA-BRNA by the RACE procedure (10; see Materials and Meth-ods). Twenty-one independent cDNA clones were isolated,
and their starting nucleotides were determined by DNAsequencing (Fig. 3C). The shortest of these cDNA clonescorresponded to the 5' end of the published CD19 cDNAsequence (33). In conclusion, all three methods revealedheterogeneous initiation of CD19 gene transcription within a40-bp G+C-rich DNA sequence. In agreement with thisfinding, there is no TATA-like sequence in the immediateupstream region.Mapping of BSAP-binding sites in the CD19 gene. An
important prerequisite for the rapid identification of BSAP-binding sites on the CD19 cosmid clone was the availabilityof partially pure BSAP protein, which was isolated fromBJA-B cells by a two-step procedure (1). Typically, BSAPrepresented -10% of the total protein in the final fraction,corresponding to a 5,000-fold enrichment. Such a BSAPpreparation was used for multiple bandshift assay (16). Anend-labeled MspI digest of the CD19 cosmid DNA wasincubated with increasing amounts of BSAP prior to separa-tion on a native polyacrylamide gel (Fig. 4A). The prediction
VOL. 12, 1992
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

2666 KOZMIK ET AL.
A B
BSAP (pI) - 1 2 2 2competitor 1 50
(ng)
H .. ,. ....-.S.A:_
H *
G *-o_
F > - `
D >-
B 10
4-1A *1 ,#S
1~~ ~ ~ ~ ~~~~~~~~~2 3 4 5
3 21 4 5 6
M MH L M M
- C -A- B
FIG. 4. Identification of BSAP-binding sites in the 5' region of the CD19 gene (A) Multiple bandshift assay. An end-labeled MspI digestof CD19 cosmid DNA was incubated with (lanes 2 to 5) or without (lanes 1 and 6) purified BSAP protein prior to electrophoretic separationon a native 4% polyacrylamide gel. An oligonucleotide containing the TSAP-binding site of the sea urchin H2A-2.2 gene was used as a specificcompetitor in lanes 4 and 5. Arrowheads indicate the position of MspI fragments with BSAP-binding sites. (B) DNase I footprint analysis.MspI fragments A to C were cloned into the AccI site of plasmid pSP64, and end-labeled DNA fragments containing these inserts were usedfor DNase I footprint analysis in the presence (+) or absence (-) of purified BSAP protein. A map of the 5' region of the human CD19 geneis shown below; the positions of MspI fragments A to C and the six DNase I-protected BSAP-binding sites are indicated. H, high-affinityBSAP-binding site; M, BSAP site with an affinity 4-fold lower than that of the sea urchin TSAP-binding sites; L, BSAP site with an affinity16-fold lower than that of the TSAP-binding sites.
was that any DNA fragment containing one or more BSAP-binding sites should form a complex upon BSAP binding(Fig. 4A, lanes 2 and 3) and thus be retarded in its migrationrelative to the unbound DNA fragment (Fig. 4A, lanes 1 and6). The MspI fragments labeled A to H (except for C) clearlyshowed this behavior, and furthermore, their retardationwas prevented by the addition of increasing amounts ofspecific competitor DNA to the binding reaction mixture(Fig. 4A, lanes 4 and 5). Comparison of the MspI digestionpattern of the entire cosmid clone with that of three differentsubclones (Fig. 2) allowed us to map fragments A and C to
the 5' region (clone HA), fragment B to the central part(clone AX), and fragment H to the 3' part (clone XS) of theCD19 gene (1Sa). Fragments D to G must therefore originatefrom the 3' flanking sequences. No BSAP-binding sites weremapped to MspI fragments of the cosmid vector.The MspI fragments A and B were subcloned, and their
precise positions on the CD19 map were determined byDNA sequencing. BSAP recognition sequences within theseDNA fragments were then identified by DNase I footprinting(Fig. 4B). DNase I footprints 1 to 6 could all be competed forwith an excess of a wild-type but not a mutant BSAP-binding
BSAP*c+C, - +
s,t$ .-<..Z".. ..44.:: BSAP[..
....UI_
*flw
*.4--!
t .
....a:.
2 at-
3: ,iC
BSAP
I.._
a..'.enw
__
o 413
A
_..C..
C0
MOL. CELL. BIOL.
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from
REGULATION OF THE CD19 GENE BY BSAP 2667
A0-
<) BSAP BSAPprobes -- site 1 site 2
B site 1
F B
'UF B
m _
site 2
F B F B
_W_ -_wu
BSAP _ _M_ i..._
M :4
competitor 41
TSAP (H2A-2.2) -
Pu (SV40) -
_.... . m...<
mm>- _4~~~~~~Wl* -4~~~~~~~~~~~~~~~1j4:
-- - - -
- - - -
- .4
"- %M . Q4
-_ .4
C
3 'vsite 1 CCCCCGCAGACACCCATGGTTGAGTGCCCTCCAGGCCCCTGCCTG
GGGGGCGTCTGTGGGTACCA ACTCACGGGAG GTCCGGGGACGGACI A ~~~~AA vL
I-- w--
site 2 ccTGGAGAATGGGGCCTGAGGCGTGACACCGCCTTCCTCTCTGGGGACCTCTTACCCCGACTCCGCACTGG TGGCGGAAGGAGAGACC
A Aupper lower upper lower
strand strand
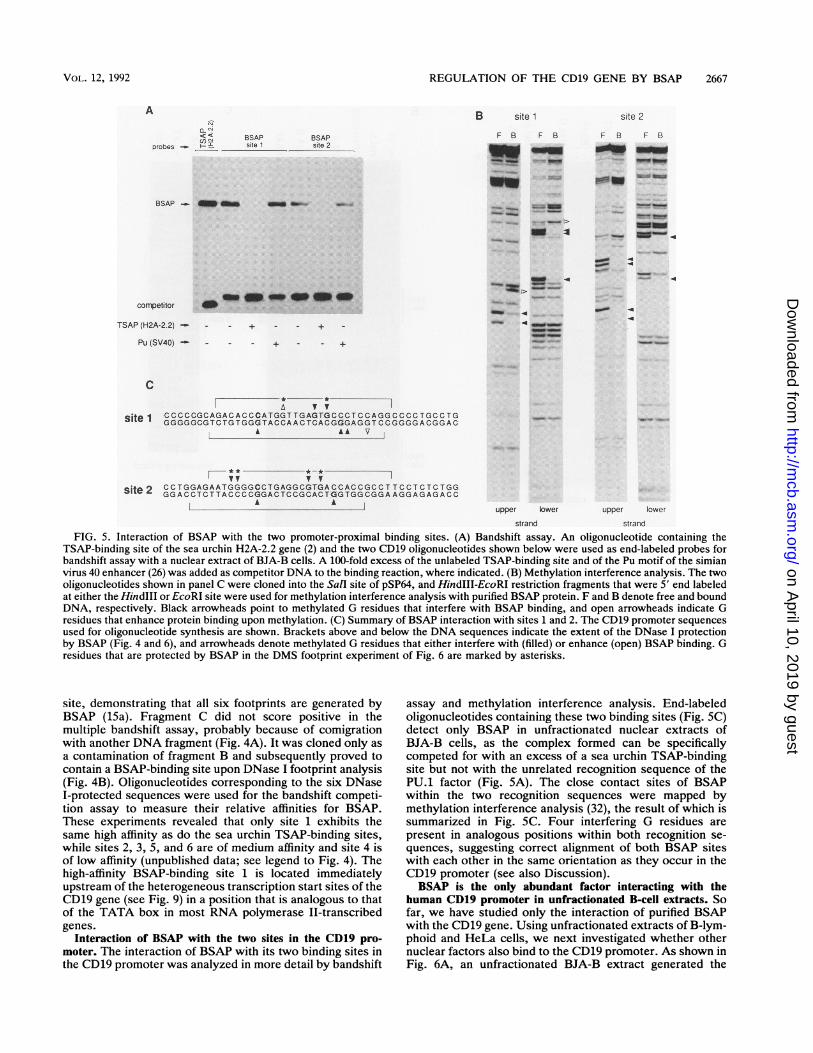
FIG. 5. Interaction of BSAP with the two promoter-proximal binding sites. (A) Bandshift assay. An oligonucleotide containing theTSAP-binding site of the sea urchin H2A-2.2 gene (2) and the two CD19 oligonucleotides shown below were used as end-labeled probes forbandshift assay with a nuclear extract of BJA-B cells. A 100-fold excess of the unlabeled TSAP-binding site and of the Pu motif of the simianvirus 40 enhancer (26) was added as competitor DNA to the binding reaction, where indicated. (B) Methylation interference analysis. The twooligonucleotides shown in panel C were cloned into the Sall site of pSP64, and HindIII-EcoRI restriction fragments that were 5' end labeledat either the HindIII or EcoRI site were used for methylation interference analysis with purified BSAP protein. F and B denote free and boundDNA, respectively. Black arrowheads point to methylated G residues that interfere with BSAP binding, and open arrowheads indicate Gresidues that enhance protein binding upon methylation. (C) Summary of BSAP interaction with sites 1 and 2. The CDl9 promoter sequencesused for oligonucleotide synthesis are shown. Brackets above and below the DNA sequences indicate the extent of the DNase I protectionby BSAP (Fig. 4 and 6), and arrowheads denote methylated G residues that either interfere with (filled) or enhance (open) BSAP binding. Gresidues that are protected by BSAP in the DMS footprint experiment of Fig. 6 are marked by asterisks.
site, demonstrating that all six footprints are generated byBSAP (15a). Fragment C did not score positive in themultiple bandshift assay, probably because of comigrationwith another DNA fragment (Fig. 4A). It was cloned only asa contamination of fragment B and subsequently proved tocontain a BSAP-binding site upon DNase I footprint analysis(Fig. 4B). Oligonucleotides corresponding to the six DNaseI-protected sequences were used for the bandshift competi-tion assay to measure their relative affinities for BSAP.These experiments revealed that only site 1 exhibits thesame high affinity as do the sea urchin TSAP-binding sites,while sites 2, 3, 5, and 6 are of medium affinity and site 4 isof low affinity (unpublished data; see legend to Fig. 4). Thehigh-affinity BSAP-binding site 1 is located immediatelyupstream of the heterogeneous transcription start sites of theCD19 gene (see Fig. 9) in a position that is analogous to thatof the TATA box in most RNA polymerase II-transcribedgenes.
Interaction of BSAP with the two sites in the CD19 pro-moter. The interaction of BSAP with its two binding sites inthe CD19 promoter was analyzed in more detail by bandshift
assay and methylation interference analysis. End-labeledoligonucleotides containing these two binding sites (Fig. SC)detect only BSAP in unfractionated nuclear extracts ofBJA-B cells, as the complex formed can be specificallycompeted for with an excess of a sea urchin TSAP-bindingsite but not with the unrelated recognition sequence of thePU.1 factor (Fig. 5A). The close contact sites of BSAPwithin the two recognition sequences were mapped bymethylation interference analysis (32), the result of which issummarized in Fig. SC. Four interfering G residues arepresent in analogous positions within both recognition se-quences, suggesting correct alignment of both BSAP siteswith each other in the same orientation as they occur in theCD19 promoter (see also Discussion).BSAP is the only abundant factor interacting with the
human CD19 promoter in unfractionated B-cell extracts. Sofar, we have studied only the interaction of purified BSAPwith the CD19 gene. Using unfractionated extracts of B-lym-phoid and HeLa cells, we next investigated whether othernuclear factors also bind to the CD19 promoter. As shown inFig. 6A, an unfractionated BJA-B extract generated the
VOL. 12, 1992
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

2668 KOZMIK ET AL.
A
o () -5Cm mn
BSAPsite 1
BSAPsile 2
ca) +
-.- comp.
B .
coJCO-
0 a) CL < LU -C: m ACc Er C CD
C
EL
U)aD
WI.
BSA[site
BSAP ; - _ _ ..i
coding strand
cI X <+ oU)
m!00 a:
BSAP.site21
BSAP_site '' ...,_, B>:
E.
q,
non-codingsstrn
* -:f. ...'i.
non-coding strand
FIG. 6. In vitro footprint of the CD19 promoter in unfractionated nuclear extracts. (A) DNase I footprint of the human CD19 promoter.The coding strand of the CD19 gene was 3' end labeled at a Sau3A site in the promoter, and this DNA probe was incubated with eitherpartially purified BSAP protein or unfractionated nuclear extracts of BJA-B and HeLa cells prior to DNase I digestion and electrophoresison an 8% sequencing gel. An oligonucleotide containing the TSAP-binding site of the H2A-2.2 gene was used as a specific competitor. Theextent of DNase I protection is indicated by brackets. Lane no XT, control DNase I digestion of naked DNA; lane G+A, Maxam-Gilbert (21)sequencing ladder. (B) DMS protection of the human CD19 promoter. The DNA probe was 3' end labeled at a HpaII site in the transcriptioninitiation region of the human CD19 promoter, incubated in unfractionated nuclear extracts, briefly exposed to DMS, and then subjected tochemical cleavage at methylated G residues and separation on a denaturing 8% polyacrylamide gel. Black arrowheads point to protected Gresidues; open arrowheads indicate hyperreactive guanosines. (C) DNase I footprint of the mouse CD19 promoter. The DNA probe was 3'end labeled at a BamHI site which was introduced into the first exon by PCR cloning (see Materials and Methods). The putative positionsof BSAP site 2 and the transcription initiation region in analogy to the human CD19 promoter are indicated by dashed lines.
same DNase I footprint pattern on the CD19 promoter as didpurified BSAP protein. This result suggests that BSAP is theonly abundant DNA-binding protein capable of interactingwith the proximal promoter of the CD19 gene in BJA-B cells.This conclusion is supported by the methylation protectionexperiment shown in Fig. 6B. An end-labeled CD19 pro-moter fragment was incubated in different nuclear extracts,briefly exposed to DMS, and then subjected to cleavage atmethylated G residues and to electrophoretic analysis. Spe-cific G residues within BSAP sites 1 and 2 were protectedfrom methylation only by three BSAP-positive extracts(Raji, REH, and BJA-B) and by the purified BSAP protein.In fact, the same G residues prevented BSAP binding in themethylation interference experiment shown in Fig. 5. No Gresidues outside of the two BSAP sites were protected, whiletwo guanines were hyperreactive in BSAP-positive nuclearextracts for unknown reasons. The control HeLa cell nu-clear extract contains an activity that also bound to the
BSAP site 2 sequence but generated a DNase I footprintdistinct from that of BSAP (Fig. 6A). This activity was notfurther characterized and could not even be detected in theDMS protection experiment (Fig. 6B), possibly because ofdifferences in the binding condition of the two assays. ADNA fragment spanning the first 200 bp of the CD19promoter was used as a probe for bandshift experiments withBJA-B nuclear extracts. Again no specific DNA-bindingactivity other than BSAP was identified by this method(1Sa). We therefore conclude that BSAP is the only abundantDNA-binding protein that interacts in vitro with the proxi-mal promoter of the human CD19 gene.
Evolutionary conservation ofBSAP site 1. The mouse CD19promoter was cloned by a genomic walking technique (9)which is based on the ligation-mediated PCR method previ-ously developed for in vivo footprinting (22). The onlyinformation required for this cloning procedure was themouse CD19 full-length cDNA sequence (38), allowing the
MOL. CELL. BIOL.
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

REGULATION OF THE CD19 GENE BY BSAP 2669
< (Dz N
C14~~~~~~~~~~~~'m 0
coaS]IC: cotocm
a)~~~~~~~~a -,-
__- D
cn
U)~~~~~~~~~~~~~~~~~~~c
LO
non- coding strand
FIG. 7. In vivo footprint of the CD19 promoter. Intact BJA-B,
HeLa, and RPMI 8226 cells or naked DNA isolated from BJA-B
cells were incubated with DMS, and DNA was isolated and sub-
jected to chemical cleavage at methylated G residues, linker liga-
tion, and PCR amplification as described by Mueller and Wold (22).The CD19 gene-specific primers used for PCR are described in
Materials and Methods. Control sequence ladders were generated
by genomic sequencing of naked BJA-B DNA following depurina-tion with formic acid (lane G+A) or modification with hydrazine
(lane C).
BJA-B J558L SP2/0, I /
CO Gg~ c C)Gt co G<)p M
5'end - _ W - 190
reference _Lumm .. amm~~~~~~,..
_ - 147
SV40TATA p-globin enhancer
OVEC-S - - /i
BSAPsitel
OVEC-CD19 -
OVEC-Retf -
delFIG. 8. Evidence that the BSAP-binding site 1 is a functional
promoter element in B cells. The test plasmids OVEC-S andOVEC-CD19 were transiently transfected into BJA-B, J558L, andSP2/0 cells together with OVEC-Ref, and expression of the 3-globinreporter gene was analyzed by SP6 mapping as described in Mate-rials and Methods. BJA-B is a human B-cell line; J558L and SP2/0are two mouse plasma cell lines. OVEC-S and OVEC-Ref weredescribed by Westin et al. (37). OVEC-CD19 contains the BSAP site1 oligonucleotide (flanked by SalI ends; Fig. SC) in the SalI site ofOVEC-S. 5' end indicates the correctly initiated ,B-globin transcriptsof the test plasmids; reference indicates transcripts of plasmidOVEC-Ref. 5'-end-labeled pUC18 DNA digested with MspI wasused as a size marker (lane M; sizes given in nucleotides).
design of 5'-terminal, CD19-specific primers. The mousepromoter was isolated as a 670-bp PCR fragment (seeMaterials and Methods). DNA sequence analysis revealedthat the BSAP-binding site 1 has been relatively well con-served between the human and mouse CD19 promoters, asonly 4 of 26 nucleotide positions differ by point mutation (seeFig. 10). In contrast, an insertion of six nucleotides wasfound in the putative BSAP site 2 of the mouse promoter. Inagreement with this finding, purified BSAP bound only tosite 1 of the mouse promoter, as shown by the DNase Ifootprint experiment of Fig. 6C. Bandshift competition ex-periments furthermore demonstrated that BSAP binds toboth the mouse and human BSAP site 1 with the same highaffinity (1Sa).BSAP binds in vivo to its high-affinity binding site in the
CD19 promoter. The interaction of BSAP with the CD19promoter was next investigated by in vivo footprinting usingthe ligation-mediated PCR method described by Mueller andWold (22). BJA-B, RPMI 8226, and HeLa cells were brieflyexposed to DMS, and genomic DNA was isolated andcleaved at methylated G residues. After complementarystrand synthesis, the DNA was subjected to linker ligationand exponential amplification of CD19 promoter sequenceswith specific oligonucleotides (see Materials and Methods).Figure 7 demonstrates that exactly the same methylationprotection pattern is observed for BSAP site 1 in BJA-B cellsin vivo as is seen with nuclear extracts in vitro (Fig. 6). Inaddition, two G residues in the middle of the transcriptioninitiation region are partially protected. There is, however,no evidence for an in vivo interaction of BSAP with thedistal site 2 in BJA-B cells (compare Fig. 6B and 7).Moreover, no protection of site 1 is seen in the BSAP-
negative RPMI 8226 and HeLa cells. In summary, we havedemonstrated that the proximal high-affinity site of the CD19promoter is fully occupied in vivo by BSAP in a CD19-expressing B-cell line.BSAP site 1 is a functional promoter element in B cells. The
evolutionary conservation and the in vivo footprint experi-ment strongly suggest that the high-affinity BSAP site 1 is afunctional promoter element. We have tested this supposi-tion by the transient transfection experiment shown in Fig.8. An oligonucleotide containing the BSAP site 1 was clonedupstream of the TATA box of the rabbit ,B-globin gene in thevector OVEC-S (37). This test plasmid and the parentalvector were transfected into the B-cell line BJA-B and thetwo plasma cell lines J558L and SP2/0 together with areference gene containing a deletion in the ,3-globin leadersequence (OVEC-Ref [37]). Cytoplasmic RNA was isolated48 h later, and transcripts of both the test and referencegenes were mapped by RNase protection assay. The BSAPsite 1 of the human CD19 gene clearly functions as anefficient promoter element to stimulate P-globin gene tran-scription in B cells but not in plasma cells. A similarexperiment previously demonstrated that the TSAP-bindingsite of the sea urchin H2A-2.2 gene confers B-cell specificityto the same ,B-globin reporter gene (3). We therefore con-clude that the BSAP site 1 of the CD19 gene is not onlybiochemically equivalent but also functionally similar to thesea urchin TSAP-binding site.
DISCUSSION
The B-cell-specific transcription factor BSAP was previ-ously identified as a mammalian homolog of the sea urchin
VOL. 12, 1992
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

2670 KOZMIK ET AL.
BSAP site 2in vitro binding (M)
' Ty A TVhuman CAGCGTGGCAGGGAGGAGGCAGTGTTGTGAGTCTGGAGGGTTCCTGGAGATGGGGCCTGAGGCGTGACCACCGCCTTCCTCTCTGGGGGGACTGCCTGCmouse -T-G -A G- GA-T A-AG-G-CT-CA A-| A T C-T A-AC A-
'A GCCTM
BSAP site 1in vivo and in vitro binding (H)
5' ends of CD19 mRNA
i -
t;I ~~~~~~~~~~~MP P P R
CGCCCCCGCAGACACCCATGGTTGAGTGCCCTCCAGGCCCCTGCCTGCCCCAGCATCCCCTGCGCGAGCTGGGTGCCCCGGAGAGTCTGACCACCATGCCACCTCCTCGC-A-T-A C-T A-T T TC T C A-G-C-G-T T T-
AGCCACGCAFIG. 9. Promoter sequences of the human and mouse CD19 genes and summary of protein-DNA interactions. Only the nucleotides that
differ from the human CD19 sequence are shown for the mouse promoter. The data of the DNase I footprint and methylation interferenceanalyses are indicated by the symbols described in the legend to Fig. 5. Dots denote the G residues that are either protected or hyperreactivein the in vivo footprint experiment (Fig. 7). Transcription initiation sites (mapped by Si nuclease analysis; Fig. 3) are shown by short verticallines. M and H denote medium- and high-affinity binding sites, respectively.
protein TSAP. Competition experiments did not reveal anyhigh-affinity BSAP-binding site in known regulatory regionsof immunoglobulin and class II major histocompatibilitycomplex genes, suggesting that BSAP may regulate a differ-ent set of B-lymphoid-specific genes (3). In this report, wedescribe the cloning and characterization of the human CD19gene. Five lines of evidence indicate that BSAP is involvedin the regulation of this gene. First, complete correlation wasobserved between CD19 gene expression and BSAP activityin a large panel of murine and human B-lymphoid cell lines.Second, a high-affinity BSAP-binding site was found imme-diately upstream of the transcription start sites of the CD19gene. Third, this high-affinity binding site has been evolu-tionarily conserved between the human and mouse CD19promoters. Fourth, in vivo footprinting demonstrated thatthis site is fully occupied by BSAP in a CD19-expressingB-cell line but not in nonexpressing cells. Fifth, this high-affinity BSAP site was shown to function as an efficientpromoter element in transiently transfected B cells. Figure 9summarizes the results of our CD19 promoter analyses.
Transcription of the human CD19 gene starts at multiplesites within a 40-bp G+C-rich sequence, as shown by threedifferent methods. A very similar situation has been de-scribed for another B-cell-specific gene coding for the B29protein (15). Such heterogeneity of mRNA 5' ends is nor-mally observed with genes that lack a canonical TATA boxin their -30 promoter regions (4, 12). In agreement with thisfinding, there is no TATA-like sequence in the proximalpromoter of the CD19 gene. The high-affinity site 1 which isbound by BSAP in vivo is instead located in the -30 region(Fig. 9). We have previously shown that a BSAP-binding sitecooperates with a TATA element to bring about efficienttranscription of a reporter gene in transiently transfected Bcells (3). The specific location of BSAP site 1 in the CD19promoter now suggests that BSAP, like Spl (28) and GCN4(6), is capable of interacting with the transcription machineryeven in the absence of a TATA box.BSAP is the only abundant DNA-binding activity present
in B-cell nuclear extracts that could be shown to interactwith the first 200 bp of the CD19 promoter by in vitroprotein-DNA binding studies. The human CD19 promotercontains a second BSAP-binding site of lower affinity whichis located 35 bp upstream of site 1. However, no evidence for
an interaction of BSAP with this site in vivo was obtained.Moreover, this second binding site has been eliminated fromthe mouse CD19 promoter by insertion of six nucleotidesinto the corresponding sequence (Fig. 6C and 9). It istherefore likely that the second BSAP-binding site of thehuman CD19 promoter plays only a minor role, if any, inCD19 gene transcription.We have previously shown that the DNA sequence rec-
ognition of BSAP is identical to that of the sea urchintranscription factor TSAP (3). Only a degenerate consensussequence for TSAP could be deduced from four high-affinitybinding sites of sea urchin histone genes, and comparison ofthree different parameters (primary sequence, DNase I foot-print, and methylation interference pattern) was required forcorrect alignment of all four sequences (2). Analysis of thesame three parameters for BSAP sites 1 and 2 of the humanCD19 promoter resulted in the best alignment with the TSAPconsensus sequence, as shown in Fig. 10. A gap of onenucleotide had to be introduced into site 2, which mayexplain its lower affinity for BSAP. Preliminary mutationalanalysis is in agreement with the alignment shown in Fig. 10.A C-to-A mutation at the position corresponding to the onlystrictly invariant C residue of the TSAP consensus sequence(Fig. 10) reduces the affinity of BSAP site 1 by a factor of 5,while insertion of an A residue eliminating the gap in site 2creates a high-affinity binding site (15a).The high-affinity site 1 of the CD19 gene functions as an
TSAP consensus ---GACGCA- TGAAGGA - - - A G ----
BSAPsite1 (m) AGAC C T GTaCATTGC CACTA
BSAPsite1 (h) AGAC C T GI T ATGC CTCCA
BSAP site 2 (h) AAT GG MC GMTGA CACCG
FIG. 10. Comparison of the BSAP recognition sequences of theCD19 promoter BSAP-binding sites 1 and 2 of the human (h) CD19promoters and site 1 of the mouse (m) promoter were aligned withthe previously deduced consensus sequence of the sea urchinTSAP-binding sites (2). Nucleotides that fit to the consensus se-quence are shaded, and the position of the invariant C residue ismarked by an arrow. A black box indicates the gap that had to beintroduced into site 2 for optimal alignment.
MOL. CELL. BIOL.
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

REGULATION OF THE CD19 GENE BY BSAP 2671
efficient promoter element upstream of the TATA box of a3-globin reporter gene in transiently transfected B cells but
not in plasma cells (Fig. 8). The BSAP site 1 is thereforefunctionally similar to the TSAP-binding site of the H2A-2.2gene, which was previously analyzed in the same manner inB-lymphoid cells (3). These data suggest, but do not prove,that the high-affinity BSAP site 1 is important for B-cell-specific regulation of the CD19 gene. A rigorous examinationof this question critically depends on an efficient expressionsystem for the CD19 gene, that allows a mutational analysisof BSAP site 1 in its normal promoter context. We havefused 1,300 or 300 bp of CD19 5' flanking sequences in thefirst exon to a rabbit 1-globin gene which either lacked orcontained a downstream simian virus 40, IgK, or IgH en-hancer or additional multimerized BSAP-binding sites. Withall of these constructs we failed, however, to detect tran-scription from the human CD19 promoter in transientlytransfected B cells (36a). It is still possible that far upstreamor downstream sequences of the CD19 gene which areabsent from our constructs are absolutely required forexpression. We note, however, that the CD19 mRNA is notat all abundant in expressing B-lymphoid cells (Fig. 1),suggesting low transcriptional activity of the endogenousCD19 promoter. This observation combined with the poortransfection efficiency of B-cell lines may explain our failureto detect CD19 promoter activity. The human BSAP genehas recently been cloned (1), and hence loss-of-functionexperiments eliminating BSAP activity from B lymphocytescan now be used as alternative approaches to determine therole of BSAP in CD19 gene regulation.The CD19 gene is most likely just one of several BSAP
target genes in B-lymphoid cells. Analysis of two othercandidate genes, mb-i (30) and Lyb-2 (24), demonstratedthat only the expression pattern of the mb-i gene strictlycorrelates with the presence of BSAP activity in the samepanel of cell lines analyzed in Fig. 1 (7a). However, therecent characterization of the mb-i gene promoter did notreveal any BSAP-binding site but instead led to the identifi-cation of a novel transcription factor. This DNA-bindingactivity, termed EBF, is required for efficient transcriptionof the mb-i gene and shows an expression profile similar tothat of BSAP (13). So far we have not detected any EBF-binding site in the 5' flanking sequences of the CD19 gene. Itwill be of special interest to determine whether BSAP isinvolved in the regulation of the EBF gene or vice versa. Asthe BSAP gene has been isolated, the tools should now beavailable to address this question and more generally tostudy the functional significance of BSAP for B-cell differ-entiation.
ACKNOWLEDGMENTSWe are grateful to B. Seed for providing the human CD19 cDNA
clone, to A.-M. Frischauf for supplying the human genomic cosmidlibrary, to G. Schaffner for oligonucleotide synthesis, to A. Schnat-tinger and R. Kurzbauer for DNA sequencing, to H. Tkadletz forartwork, and to C. Wrighton and M. Zenke for critical reading of themanuscript.
This work was supported by the Institute of Molecular Pathologyand by a grant from the Austrian Industrial Research PromotionFund.
REFERENCES1. Adams, B., P. Dorfler, A. Aguzzi, Z. Kozmik, P. Urbanek, L.
Maurer-Fogy, and M. Busslinger. Submitted for publication.la.Barberis, A., G. Superti-Furga, and M. Busslinger. 1987. Mutu-
ally exclusive interaction of the CCAAT-binding factor and of adisplacement protein with overlapping sequences of a histone
gene promoter. Cell 50:347-359.2. Barberis, A., G. Superti-Furga, L. Vitelli, I. Kemler, and M.
Busslinger. 1989. Developmental and tissue-specific regulationof a novel transcription factor of the sea urchin. Genes Dev.3:663-675.
3. Barberis, A., K. Widenhorn, L. Vitelli, and M. Busslinger. 1990.A novel B-cell lineage-specific transcription factor present atearly but not late stages of differentiation. Genes Dev. 4:849-859.
4. Breathnach, R., and P. Chambon. 1981. Organization andexpression of eukaryotic split genes coding for proteins. Annu.Rev. Biochem. 50:349-383.
5. Busslinger, M., N. Moschonas, and R. A. Flavell. 1981. Ithalassemia: aberrant splicing results from a single point muta-tion in an intron. Cell 27:289-298.
6. Chen, W., and K. Struhl. 1989. Yeast upstream activator proteinGCN4 can stimulate transcription when its binding site replacesthe TATA element. EMBO J. 8:261-268.
7. de Rie, M. A., T. N. M. Schumacher, G. M. W. van SchUndel,R. A. W. van Lier, and F. Miedema. 1989. Regulatory role ofCD19 molecules in B-cell activation and differentiation. Cell.Immunol. 118:369-381.
7a.Dfirfler, P. Unpublished data.8. Ehrich, E., A. Craig, A. Poustka, A.-M. Frischauf, and H.
Lehrach. 1987. A family of cosmid vectors with the multi-copyR6K replication origin. Gene 57:229-237.
9. Fors, L., R. A. Saavedra, and L. Hood. 1990. Cloning of theshark Po promoter using a genomic walking technique based onthe polymerase chain reaction. Nucleic Acids Res. 18:2793-2799.
10. Frohman, M. A., M. K. Dush, and G. R. Martin. 1988. Rapidproduction of full-length cDNAs from rare transcripts: amplifi-cation using a single gene-specific oligonucleotide primer. Proc.Natl. Acad. Sci. USA 85:8998-9002.
11. Golay, J. T., and D. H. Crawford. 1987. Pathways of humanB-lymphocyte activation blocked by B-cell specific monoclonalantibodies. Immunology 62:279-284.
12. Grosschedl, R., and M. L. Birnstiel. 1980. Identification ofregulatory sequences in the prelude sequences of an H2Ahistone gene by the study of specific deletion mutants in vivo.Proc. Natl. Acad. Sci. USA 77:1432-1436.
13. Hagman, J., A. Travis, and R. Grosschedl. 1991. A novellineage-specific nuclear factor regulates mb-1 gene transcriptionat the early stages of B-cell differentiaton. EMBO J. 10:3409-3417.
14. Hebell, T., J. M. Ahearn, and D. T. Fearon. 1991. Suppressionof the immune response by a soluble complement receptor of Blymphocytes. Science 254:102-105.
15. Hermanson, G. G., M. Briskin, S. Sigman, and R. Wall. 1989.Immunoglobulin enhancer and promoter motifs 5' of the B29B-cell-specific gene. Proc. Natl. Acad. Sci. USA 86:7341-7345.
15a.Kozmik, Z. Unpublished data.16. Kozmik, Z., and V. Paces. 1990. Multiple bandshift assay: rapid
identification and cloning of DNA fragments containing specificprotein-binding sites. Gene 90:287-291.
17. Ledbetter, J. A., P. S. Rabinovitch, C. H. June, C. W. Song,E. A. Clark, and F. M. Uckun. 1988. Antigen-independentregulation of cytoplasmic calcium in B-cells with a 12-kDaB-cell growth factor and anti-CD19. Proc. Natl. Acad. Sci. USA85:1897-1901.
18. Lee, K. H., T. Kinashi, K. Tohyama, K. Tashiro, N. Funato, K.Hama, and T. Honijo. 1991. Different stromal cell lines supportlineage-selective differentiation of the multipotential bone mar-row stem cell clone LyD9. J. Exp. Med. 173:1257-1266.
19. Maniatis, F., E. F. Fritsch, and J. SambrooL 1982. Molecularcloning: a laboratory manual, p. 312-328. Cold Spring HarborLaboratory Press, Cold Spring Harbor, N.Y.
20. Matsumoto, A. K., J. Kopicky-Burd, R. H. Carter, D. A.Tuveson, T. F. Tedder, and D. F. Fearon. Intersection of thecomplement and immune systems: a signal transduction com-plex of the B lymphocyte-containing complement receptor type2 and CD19. J. Exp. Med. 173:55-64.
21. Maxam, A. M., and W. Gilbert. 1980. Sequencing end-labeled
VOL. 12, 1992
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from

2672 KOZMIK ET AL.
DNA with base-specific chemical cleavages. Methods Enzymol.65:499-560.
22. Mueller, P. R., and B. Wold. 1989. In vivo footprinting of amuscle specific enhancer by ligation mediated PCR. Science246:780-786.
23. Nadler, L. M., K. C. Anderson, G. Marti, M. Bates, E. Park,J. F. Daley, and S. F. Schlossman. 1983. B4, a human Blymphocyte-associated antigen expressed on normal, mitogen-activated, and malignant B lymphocytes. J. Immunol. 131:244-250.
24. Nakayama, E., I. von Hoegen, and J. R. Parnes. 1989. Sequenceof the Lyb-2 B-cell differentiation antigen defines a gene super-family of receptors with inverted membrane orientation. Proc.Natl. Acad. Sci. USA 86:1352-1356.
25. Palacios, R., H. Karasuyama, and A. Rolink. 1987. Lyl+ Pro-Blymphocyte clones: phenotype, growth requirements and differ-entiation in vitro and in vivo. EMBO J. 6:3687-3693.
26. Pettersson, M., and W. Schaffner. 1987. A purine-rich DNAsequence motif present in SV40 and lymphotropic papovavirusbinds a lymphoid-specific factor and contributes to enhanceractivity in lymphoid cells. Genes Dev. 1:962-972.
27. Pezzutto, A., B. Dorken, P. S. Rabinovitch, J. A. Ledbetter, G.Moldenhauer, and E. A. Clark. 1987. CD19 monoclonal anti-body HD37 inhibits anti-immunoglobulin-induced B cell activa-tion and proliferation. J. Immunol. 138:2793-2799.
28. Pugh, B. F., and R. Tjian. 1990. Mechanism of transcriptionalactivation by Spl: evidence for coactivators. Cell 61:1187-1197.
29. Rolink, A., and F. Melchers. 1991. Molecular and cellular originsof B lymphocyte diversity. Cell 66:1081-1094.
30. Sakaguchi, N., S.-I. Kashiwamura, M. Kimoto, P. Thalmann,and F. Melchers. 1988. B lymphocyte-restricted expression ofmb-1, a gene with CD3-like structural properties. EMBO J.7:3457-3464.
31. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequenc-ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci.USA 74:5463-5467.
32. Siebenlist, U., R. Simpson, and W. Gilbert. 1988. E. coli RNApolymerase interacts homologously with two different promot-ers. Cell 20:269-281.
33. Stamenkovic, I., and B. Seed. 1988. CD19, the earliest differen-tiation antigen of the B cell lineage, bears three extracellularimmunoglobulin-like domains and an Epstein-Barr virus-relatedcytoplasmic tail. J. Exp. Med. 168:1205-1210.
34. Tedder, T. F., and C. M. Isaacs. 1989. Isolation of cDNAsencoding the CD19 antigen of human and mouse B lympho-cytes: a new member of the immunoglobulin superfamily. J.Immunol. 143:712-717.
35. Uckun, F. M., W. Jaszcz, J. L. Ambrus, A. S. Fauci, K.Gajl-Peczalska, C. W. Song, M. R. Wick, D. E. Myers, K.Waddick, and J. A. Ledbetter. 1988. Detailed studies on expres-sion and function of CD19 surface determinant by using B43monoclonal antibody and the clinical potential of anti-CD19immunotoxins. Blood 71:13-29.
36. Vitelli, L., I. Kemler, B. Lauber, M. L. Birnstiel, and M.Busslinger. 1988. Developmental regulation of micro-injectedhistone genes in sea urchin embryos. Dev. Biol. 127:54-63.
36a.Wang, S., and P. Dorfler. Unpublished data.37. Westin, G., T. Gerster, M. M. Muller, G. Schaffner, and W.
Schaffner. 1987. OVEC, a versatile system to study transcrip-tion in mammalian cells and cell-free extracts. Nucleic AcidsRes. 15:6787-6798.
38. Zhou, L.-J., D. C. Ord, A. L. Hughes, and T. F. Tedder. 1991.Structure and domain organization of the CD19 antigen ofhuman, mouse, and guinea pig B lymphocytes. J. Immunol.147:1424-1432.
MOL. CELL. BIOL.
on April 10, 2019 by guest
http://mcb.asm
.org/D
ownloaded from





























