Embed Size (px)
Citation preview

The Primate Cranial Base:Ontogeny, Function, and Integration
DANIEL E. LIEBERMAN,1 CALLUM F. ROSS,2 AND MATTHEW J. RAVOSA3
1Department of Anthropology, George Washington University,Washington, DC 20052, and Human Origins Program, National Museumof Natural History, Smithsonian Institution, Washington, DC 205602Department of Anatomical Sciences, Health Sciences Center, SUNY atStony Brook, Stony Brook, NY 11794-80813Department of Cell and Molecular Biology, Northwestern UniversityMedical School, Chicago, Illinois 60611-3008, and Mammals Division,Department of Zoology, Field Museum of Natural History,Chicago, Illinois 60605-2496
KEY WORDS cranial base; basicranium; chondrocranium; pri-mates; humans
ABSTRACT Understanding the complexities of cranial base develop-ment, function, and architecture is important for testing hypotheses aboutmany aspects of craniofacial variation and evolution. We summarize keyaspects of cranial base growth and development in primates that are usefulfor formulating and testing hypotheses about the roles of the chondrocraniumand basicranium in cranial growth, integration, and function in primate andhuman evolution. We review interspecific, experimental, and ontogeneticevidence for interactions between the cranial base and brain, and betweenthe cranial base and the face. These interactions indicate that the cranialbase plays a key role in craniofacial growth, helping to integrate, spatiallyand functionally, different patterns of growth in various adjoining regions ofthe skull such as components of the brain, the eyes, the nasal cavity, the oralcavity, and the pharynx. Brain size relative to cranial base length appears tobe the dominant influence on many aspects of basicranial variation, espe-cially the angle of the cranial base in the midsagittal plane, but other factorssuch as facial size, facial orientation, and posture may also be important.Major changes in cranial base shape appear to have played crucial roles inthe evolution of early primates, the origin of anthropoids, and the origin ofHomo sapiens. Yrbk Phys Anthropol 43:117–169, 2000. © 2000 Wiley-Liss, Inc.
TABLE OF CONTENTS
Glossary, Landmark, and Measurement Definitions ....................................................... 118Introduction ......................................................................................................................... 120Anatomy and Development ................................................................................................ 120
Development of the chondrocranium ............................................................................. 121Patterns and processes of basicranial growth .............................................................. 122
Antero-posterior growth .............................................................................................. 123Medio-lateral growth ................................................................................................... 125Supero-inferior growth ................................................................................................ 126Angulation .................................................................................................................... 126
Associations Between Cranial Base and Brain ................................................................ 131Brain size and cranial base angle .................................................................................. 131Brain shape and cranial base angle .............................................................................. 134
YEARBOOK OF PHYSICAL ANTHROPOLOGY 43:117–169 (2000)
© 2000 WILEY-LISS, INC.

Brain volume and posterior cranial fossa shape .......................................................... 138Associations Between Cranial Base and Face .................................................................. 138
Anterior cranial fossa shape and upper facial growth ................................................. 138Middle cranial fossa shape and midfacial growth ........................................................ 140Basicranial width and overall facial shape in humans ................................................ 144
The Cranial Base and Posture ........................................................................................... 148Major Unresolved Issues of Cranial Base Variation in Primate Evolution ................... 151
Primate origins, encephalization, and circumorbital form .......................................... 151Anthropoid origins and cranial base-face interactions ................................................. 152Variation in hominin cranial base angle ....................................................................... 153Cranial base flexion and vocal tract shape in hominins .............................................. 154Cranial base shape and facial projection in Homo ....................................................... 156Cranial base characters in phylogenetic analyses ........................................................ 158
Conclusions .......................................................................................................................... 159What major factors generate variation in the cranial base? ....................................... 160What role does the cranial base play in craniofacial integration? .............................. 160
Correlation ................................................................................................................... 161Constraint .................................................................................................................... 162Ontogenetic sequence .................................................................................................. 162
Future research ............................................................................................................... 163Acknowledgments ............................................................................................................... 163Literature Cited .................................................................................................................. 163
GLOSSARY
Basioccipital clivus: midline “plane” of theposterior cranial base formed by the supe-rior (endocranial) aspects of the basioccipi-tal and the posterior sphenoid.Brain stem: the ventral parts of the brain,excluding the telencephalon. Specifically, inthis paper, the brain stem consists of themedulla oblongata and mesencephalon (5optic tectum and tegmentum) of Stephan etal. (1981; see also Butler and Hodos, 1996).Chondrocranium: cartilaginous precursorsto the basicranium.Constraint: a limitation or bias on processesand/or patterns of evolution, growth, form,and function.Cranial base angulation: a series of eventsby which bone or cartilage deposition in themidline cranial base changes the angle be-tween intersecting prechordal (see below)and postchordal (see below) lines. This causesthe inferior cranial base angle to become moreacute (flexion) or more obtuse (extension).Displacement: a series of events by whichan osseous region “moves” relative to an-other osseous region through bone deposi-tion (primary displacement), or throughbone deposition in an adjoining bone (sec-ondary displacement).
Drift: a series of events by which an osseouswall “moves” relative to another anatomicalregion through bone deposition on one sur-face and bone resorption on its opposingsurface.Ethmomaxillary complex: the upper part ofthe face, mostly comprising the ethmoid, thenasal capsule, and the maxilla.Facial projection: degree to which faceprojects in front of cranial base; measuredhere by nasion-foramen caecum.Integration: the genetic, epigenetic, or func-tional association among elements via “a setcausal mechanisms so that change in oneelement is reflected by change in another”(Smith, 1996). The results of integration aremost often recognized as a pattern of signif-icant, hierarchical covariation among thecomponents of a system.Kyphosis: angle of some aspect of facial ori-entation relative to the neuro- and/or basi-cranium, measured here using angle of fa-cial kyphosis (AFK) for the orientation ofthe palate, and angle of orbit axis orienta-tion (AOA) for orientation of the orbital axis.Planum sphenoideum: midline “plane” ofthe anterior cranial base from the sphenoi-dale (Sp) to the planum sphenoideum (PS)point (see below).
118 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000

Postchordal cranial base: portion of the cra-nial base posterior to the sella; frequentlycalled the posterior cranial base.Prechordal cranial base: portion of the cra-nial base anterior to sella; frequently calledthe anterior cranial base.Telencephalon: forebrain, consisting ofpaired olfactory lobes, the basal ganglia,and the neocortex.
LANDMARK DEFINITIONS
Ba, basion: midsagittal point on anteriormargin of foramen magnum.CP, clival point: midline point on basioccip-ital clivus inferior to point at which dorsumsellae curves posteriorly.FC, foramen caecum: pit on cribriform platebetween crista galli and endocranial wall offrontal bone.H, hormion: most posterior midline point onvomer.OA: supero-inferior midpoint between supe-rior orbital fissures and inferior rims of op-tic canals; for mammals without completelyenclosed orbits, OA is defined as inferior rimof optic foramen.OM: supero-inferior midpoint betweenlower and upper orbital rims.Op, opisthion: most posterior point in fora-men magnum.PMp, PM point: average of projected mid-line points of most anterior point on laminaof greater wings of sphenoid.PP, pituitary point: “the anterior edge ofthe groove for the optic chiasma, just infront of the pituitary fossa” (Zuckerman,1955).PS, planum sphenoideum point: most supe-rior midline point on sloping surface inwhich cribriform plate is set.Ptm, pterygomaxillare: average of projectedmidline points of most inferior and posteriorpoints on maxillary tuberosities.S, sella: center of sella turcica, independentof contours of clinoid processes.Sb, sphenobasion: midline point on spheno-occipital synchondrosis on external aspect ofclivus.Sp, sphenoidale: most posterior, superiormidline point of planum sphenoideum.
ANGLE, LINE, AND PLANE DEFINITIONS
AOA: orbital axis orientation relative to CO(Ross and Ravosa, 1993).BL1: Ba-PP 1 PP-Sp (Ross and Ravosa,1993; Ross and Henneberg, 1995).BL2: Ba-S 1 S-FC (Spoor, 1997).CBA1: Ba-S relative to S-FC (Liebermanand McCarthy, 1999).CBA2: Ba-S relative to Sp-PS (Liebermanand McCarthy, 1999).CBA3: Ba-CP relative to S-FC (Liebermanand McCarthy, 1999).CBA4: Ba-CP relative to Sp-PS (Liebermanand McCarthy, 1999).CO, clivus ossis occipitalis: endocranial linefrom Ba to spheno-occipital synchondrosis(Ross and Ravosa, 1993).External CBA (CBA5): angle between basion-sphenobasion-hormion (Lieberman and Mc-Carthy, 1999).FM, foramen magnum: Ba-Op.Forel’s axis: from most antero-inferior pointon frontal lobe to most postero-inferior pointon occipital lobe (Hofer, 1969).Head-neck angle: orientation of head rela-tive to neck in locomoting animals, calcu-lated as neck inclination 2 orbit inclination(Strait and Ross, 1999).IRE1: cube root of endocranial volume/BL 1(Ross and Ravosa, 1993).IRE2: cube root of neocortical volume/BL 1(Ross and Ravosa, 1993).IRE3: cube root of telencephalon volume/BL1 (Ross and Ravosa, 1993).IRE4: cube root of neocortical volume/palatelength (Ross and Ravosa, 1993).IRE5: cube root of endocranial volume/BL 2(McCarthy, 2001).Meynert’s axis: from ventral edge of junc-tion between pons and medulla to caudalrecess of interpeduncular fossa (Hofer,1969).Neck inclination: orientation of surface ofneck relative to substrate (Strait and Ross,1999).NHA: neutral horizontal axis of orbits; fromOM to OA (Enlow and Azuma, 1975).Orbital axis orientation: line from optic fo-ramen through superoinferior midpoint oforbital aperture (Ravosa, 1988).Orbit inclination: orientation relative tosubstrate of a line joining superior and in-
PRIMATE CRANIAL BASE 119D.E. Lieberman et al.]

ferior margins of orbits (Strait and Ross,1999).PM plane, posterior maxillary plane: fromPtm to PMp (Enlow and Azuma, 1975).
INTRODUCTION
The cranial base has important integra-tive and functional roles in the skull, manyof which reflect its phylogenetic history asthe oldest component of the vertebrate skull(de Beer, 1937). Architecturally, the cranialbase provides the platform upon which thebrain grows and around which the facegrows. In addition, the cranial base con-nects the cranium with the rest of the body:it articulates with the vertebral column andthe mandible, provides conduits for all thevital neural and circulatory connections be-tween the brain and the face and neck,houses and connects the sense organs in theskull, and forms the roof of the nasophar-ynx. The shape of the cranial base is there-fore a multifactorial product of numerousphylogenetic, developmental, and func-tional interactions.
The importance of the cranial base ismatched by several challenges that make itdifficult to study. Because the cranial baseis difficult to access surgically, there havebeen few experimental studies of cranialbase growth and function. Also, a large pro-portion of the cranial base is not only com-plex anatomically, but is also difficult tomeasure and/or see externally. In addition,the cranial base in many fossils is missing,damaged, or unobservable without specialtechnology. However, new developmentalstudies, and new techniques for imaging,have led to a modest renaissance of researchon cranial base morphology (reviewed inSpoor et al., 2000). In addition, new analyt-ical techniques which quantitatively com-pare three-dimensional differences in formhave opened up new possibilities for study-ing growth and variation in complex regionssuch as the cranial base (Cheverud andRichtsmeier, 1986; Bookstein, 1991; Lele,
1993; O’Higgins, 2000). Ultimately, betterinformation about the relationships be-tween cranial base morphology and the restof the skull may help to resolve a number ofimportant phylogenetic and behavioral is-sues throughout primate evolution.
The goals of this review are to provide abackground on key aspects of cranial basegrowth and development necessary to for-mulate or test hypotheses about the role ofthe cranial base in cranial growth, integra-tion, and function. Therefore, we review re-cent research on cranial base variation, de-velopment, and evolution in primates,focusing on the major dimensions of the cra-nial base (especially width, length, and an-gulation in the sagittal plane). Other, moredetailed aspects of cranial base anatomyand morphology, most notably the inner ear,were recently reviewed by Spoor and Zon-neveld (1998) and will not be covered in thisreview (see also Braga et al., 1999). Whererelevant, we have made an effort to includedata from the few experimental studies oncranial base growth and function. Most ex-perimental research on the mammalianskull has focused on the face and neurocra-nium; however, some of these studies pro-vide indirect clues on interrelationshipsamong the brain, cranial base, and face(e.g., Sarnat, 1988, 1999). Finally, we con-clude with a short discussion of two mainissues which we believe require further re-search to address: what factors determinemost of the variation in cranial base shapeamong primates, and to what extent doesvariation in cranial base form influence on-togenetic and interspecific patterns of vari-ation in craniofacial morphology?
ANATOMY AND DEVELOPMENT
A detailed understanding of the series ofevents and underlying mechanisms thatgenerate patterns1 of morphological varia-tion in the basicranium is vital for develop-ing and testing hypotheses about the cra-nial base’s role in craniofacial integration
Please address all correspondence to: Daniel E. Lieberman,Department of Anthropology, George Washington University,2110 G St, Washington DC 20052.E-mail: danliebgwu.edu; phone: 202-994-0873; fax: 202-994-6097.
1The term “pattern” here refers to a static description of aconfiguration or relationship among things, whereas “process”refers to a series of events that occur during something’s forma-tion. Note that we do not define process here as a causal mech-anism. Most of the processes described here have multiple andhierarchical levels of causation which merit further study.
120 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000
and function. So we begin with a brief sum-mary of cranial base embryology, fetalgrowth, and postnatal growth. Most of theinformation summarized below derives fromstudies of human basicranial growth anddevelopment; the majority of these patternsand processes are generally applicable to allprimates, but we tried to distinguish thosethat are unique to humans or other species.Further information is available in Bjork(1955), Ford (1958), Scott (1958), Moore andLavelle (1974), Starck (1975), Bosma (1976),Moss et al. (1982), Slavkin (1989), Sperber(1989), Enlow (1990), and Jeffery (1999), aswell as the many references cited below.
Development of the chondrocranium
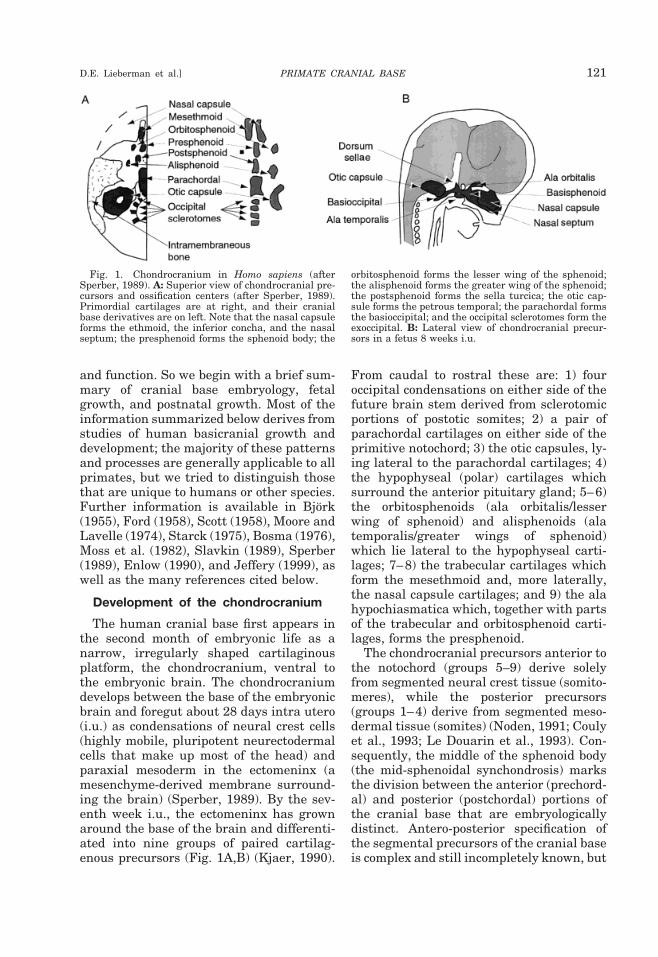
The human cranial base first appears inthe second month of embryonic life as anarrow, irregularly shaped cartilaginousplatform, the chondrocranium, ventral tothe embryonic brain. The chondrocraniumdevelops between the base of the embryonicbrain and foregut about 28 days intra utero(i.u.) as condensations of neural crest cells(highly mobile, pluripotent neurectodermalcells that make up most of the head) andparaxial mesoderm in the ectomeninx (amesenchyme-derived membrane surround-ing the brain) (Sperber, 1989). By the sev-enth week i.u., the ectomeninx has grownaround the base of the brain and differenti-ated into nine groups of paired cartilag-enous precursors (Fig. 1A,B) (Kjaer, 1990).
From caudal to rostral these are: 1) fouroccipital condensations on either side of thefuture brain stem derived from sclerotomicportions of postotic somites; 2) a pair ofparachordal cartilages on either side of theprimitive notochord; 3) the otic capsules, ly-ing lateral to the parachordal cartilages; 4)the hypophyseal (polar) cartilages whichsurround the anterior pituitary gland; 5–6)the orbitosphenoids (ala orbitalis/lesserwing of sphenoid) and alisphenoids (alatemporalis/greater wings of sphenoid)which lie lateral to the hypophyseal carti-lages; 7–8) the trabecular cartilages whichform the mesethmoid and, more laterally,the nasal capsule cartilages; and 9) the alahypochiasmatica which, together with partsof the trabecular and orbitosphenoid carti-lages, forms the presphenoid.
The chondrocranial precursors anterior tothe notochord (groups 5–9) derive solelyfrom segmented neural crest tissue (somito-meres), while the posterior precursors(groups 1–4) derive from segmented meso-dermal tissue (somites) (Noden, 1991; Coulyet al., 1993; Le Douarin et al., 1993). Con-sequently, the middle of the sphenoid body(the mid-sphenoidal synchondrosis) marksthe division between the anterior (prechord-al) and posterior (postchordal) portions ofthe cranial base that are embryologicallydistinct. Antero-posterior specification ofthe segmental precursors of the cranial baseis complex and still incompletely known, but
Fig. 1. Chondrocranium in Homo sapiens (afterSperber, 1989). A: Superior view of chondrocranial pre-cursors and ossification centers (after Sperber, 1989).Primordial cartilages are at right, and their cranialbase derivatives are on left. Note that the nasal capsuleforms the ethmoid, the inferior concha, and the nasalseptum; the presphenoid forms the sphenoid body; the
orbitosphenoid forms the lesser wing of the sphenoid;the alisphenoid forms the greater wing of the sphenoid;the postsphenoid forms the sella turcica; the otic cap-sule forms the petrous temporal; the parachordal formsthe basioccipital; and the occipital sclerotomes form theexoccipital. B: Lateral view of chondrocranial precur-sors in a fetus 8 weeks i.u.
PRIMATE CRANIAL BASE 121D.E. Lieberman et al.]

appears to mostly involve the expression ofthe Hox and Dlx gene clusters (Lufkin et al.,1992; Robinson and Mahon, 1994; Vielle-Grosjean et al., 1997). For recent summa-ries of pattern formation and gene expres-sion in the vertebrate cranial base, seeLangille and Hall (1993) and Schilling andThorogood (2000).
At least 41 ossification centers, which be-gin to appear in the chondrocranium about8 weeks i.u., are responsible for the trans-formation of the chondrocranium into thebasicranium (Sperber, 1989; Kjaer, 1990).These centers (Fig. 1B) form within a perfo-rated and highly irregularly shaped plat-form known as the basal plate. In general,ossification begins with the mesodermallyderived cartilages toward the caudal end ofthe chondrocranium, and proceeds rostrallyand laterally, eventually forming the fourmajor bones that comprise the primate ba-sicranium: the ethmoid, most of the sphe-noid, and parts of the occipital and temporalbones (which also include some intramem-branous elements). The sequence in whichthe four bones of the cranial base ossify fromthe chondrocranium is complex, and stillnot entirely resolved (reviewed in Sperber,1989; Williams et al., 1995; Jeffery, 1999),but we highlight here the major steps, pro-ceeding from caudal to rostral. The occipitalcomprises four bones surrounding the fora-men magnum. The squamous portion is pri-marily intramembranous bone of the cra-nial vault, except for the nuchal region,which ossifies endochondrally from two sep-arate centers (Srivastava, 1992) and fuseswith the lateral exoccipitals on either side ofthe foramen magnum that fuse with thebasioccipital. The sphenoid body forms fromfusion of the presphenoids and basisphenoidaround the pituitary, forming the sella tur-cica (“Turkish saddle”). The greater andlesser wings of the sphenoid develop fromthe fusion of the alisphenoid and orbito-sphenoid cartilages to the body (Kodama,1976a–c; Sasaki and Kodama, 1976). Later,the medial and lateral pterygoid plates andportions of the greater wings ossify in-tramembraneously. The temporals, whichform much of the lateral aspect of the basi-cranium, develop from approximately 21 os-sification centers, several of which are in-
tramembranous, including the squamous,tympanic, and zygomatic regions (Shapiroand Robinson, 1980; Sperber, 1989). The pe-trous and mastoid parts of the temporalform the inner ear from the otic capsule, andthe styloid process of the temporal ossifiesfrom cartilage in the second branchial arch.The ethmoid, which is entirely endochon-dral in origin, forms the center of the ante-rior cranial floor, and most of the nasal cav-ity from three ossification centers in themesethmoid and nasal capsule cartilages(Hoyte, 1991). An additional cartilaginousossification center detaches from the ecteth-moid to form a separate scrolled bone, theinferior nasal concha, inside the nasal cav-ity.
Patterns and processes of basicranialgrowth
In order to understand how the basicra-nium grows and functions during the fetaland postnatal periods, it is useful to keep inmind three important principles of basicra-nial development. First, the center of thebasicranium (an oval-shaped region aroundthe sphenoid body) attains adult size andshape more rapidly than the anterior, pos-terior, and lateral portions, presumably be-cause almost all the vital cranial nerves andmajor vessels perforate the cranial base inthis region (Figs. 1A, 2) (Sperber, 1989).Second, the prechordal (anterior) and post-chordal (posterior) cranial base grow some-what independently, perhaps reflectingtheir distinct embryonic origins and theirdifferent spatial and functional roles (out-lined above). Third, most basicranial growthin the three cranial fossae occurs indepen-dently (Fig. 2). The posterior cranial fossa,which houses the occipital lobes and thebrain stem (the cerebellum and the medullaoblongata), is bounded laterally by the pe-trous and mastoid portions of the temporalbone, and anteriorly by the dorsum sellae ofthe sphenoid. The butterfly-shaped middlecranial fossa, which supports the temporallobes and the pituitary gland, is boundedposteriorly by the dorsum sellae and thepetrous portions of the temporal, and ante-riorly by the posterior borders of the lesserwings of the sphenoid, and by the anteriorclinoid processes of the sphenoid. The ante-
122 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000
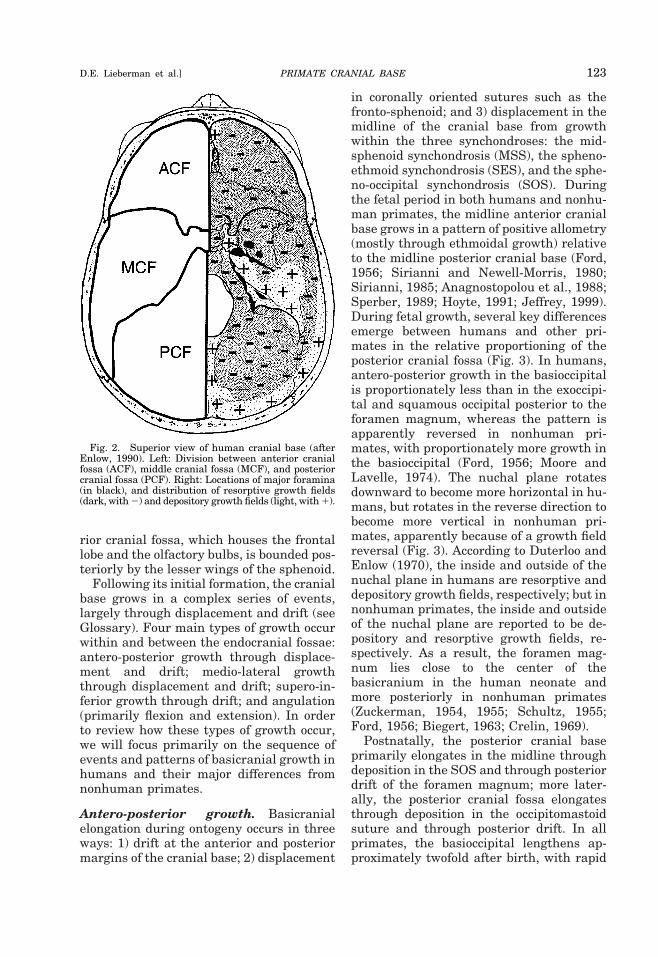
rior cranial fossa, which houses the frontallobe and the olfactory bulbs, is bounded pos-teriorly by the lesser wings of the sphenoid.
Following its initial formation, the cranialbase grows in a complex series of events,largely through displacement and drift (seeGlossary). Four main types of growth occurwithin and between the endocranial fossae:antero-posterior growth through displace-ment and drift; medio-lateral growththrough displacement and drift; supero-in-ferior growth through drift; and angulation(primarily flexion and extension). In orderto review how these types of growth occur,we will focus primarily on the sequence ofevents and patterns of basicranial growth inhumans and their major differences fromnonhuman primates.
Antero-posterior growth. Basicranialelongation during ontogeny occurs in threeways: 1) drift at the anterior and posteriormargins of the cranial base; 2) displacement
in coronally oriented sutures such as thefronto-sphenoid; and 3) displacement in themidline of the cranial base from growthwithin the three synchondroses: the mid-sphenoid synchondrosis (MSS), the spheno-ethmoid synchondrosis (SES), and the sphe-no-occipital synchondrosis (SOS). Duringthe fetal period in both humans and nonhu-man primates, the midline anterior cranialbase grows in a pattern of positive allometry(mostly through ethmoidal growth) relativeto the midline posterior cranial base (Ford,1956; Sirianni and Newell-Morris, 1980;Sirianni, 1985; Anagnostopolou et al., 1988;Sperber, 1989; Hoyte, 1991; Jeffrey, 1999).During fetal growth, several key differencesemerge between humans and other pri-mates in the relative proportioning of theposterior cranial fossa (Fig. 3). In humans,antero-posterior growth in the basioccipitalis proportionately less than in the exoccipi-tal and squamous occipital posterior to theforamen magnum, whereas the pattern isapparently reversed in nonhuman pri-mates, with proportionately more growth inthe basioccipital (Ford, 1956; Moore andLavelle, 1974). The nuchal plane rotatesdownward to become more horizontal in hu-mans, but rotates in the reverse direction tobecome more vertical in nonhuman pri-mates, apparently because of a growth fieldreversal (Fig. 3). According to Duterloo andEnlow (1970), the inside and outside of thenuchal plane in humans are resorptive anddepository growth fields, respectively; but innonhuman primates, the inside and outsideof the nuchal plane are reported to be de-pository and resorptive growth fields, re-spectively. As a result, the foramen mag-num lies close to the center of thebasicranium in the human neonate andmore posteriorly in nonhuman primates(Zuckerman, 1954, 1955; Schultz, 1955;Ford, 1956; Biegert, 1963; Crelin, 1969).
Postnatally, the posterior cranial baseprimarily elongates in the midline throughdeposition in the SOS and through posteriordrift of the foramen magnum; more later-ally, the posterior cranial fossa elongatesthrough deposition in the occipitomastoidsuture and through posterior drift. In allprimates, the basioccipital lengthens ap-proximately twofold after birth, with rapid
Fig. 2. Superior view of human cranial base (afterEnlow, 1990). Left: Division between anterior cranialfossa (ACF), middle cranial fossa (MCF), and posteriorcranial fossa (PCF). Right: Locations of major foramina(in black), and distribution of resorptive growth fields(dark, with 2) and depository growth fields (light, with 1).
PRIMATE CRANIAL BASE 123D.E. Lieberman et al.]
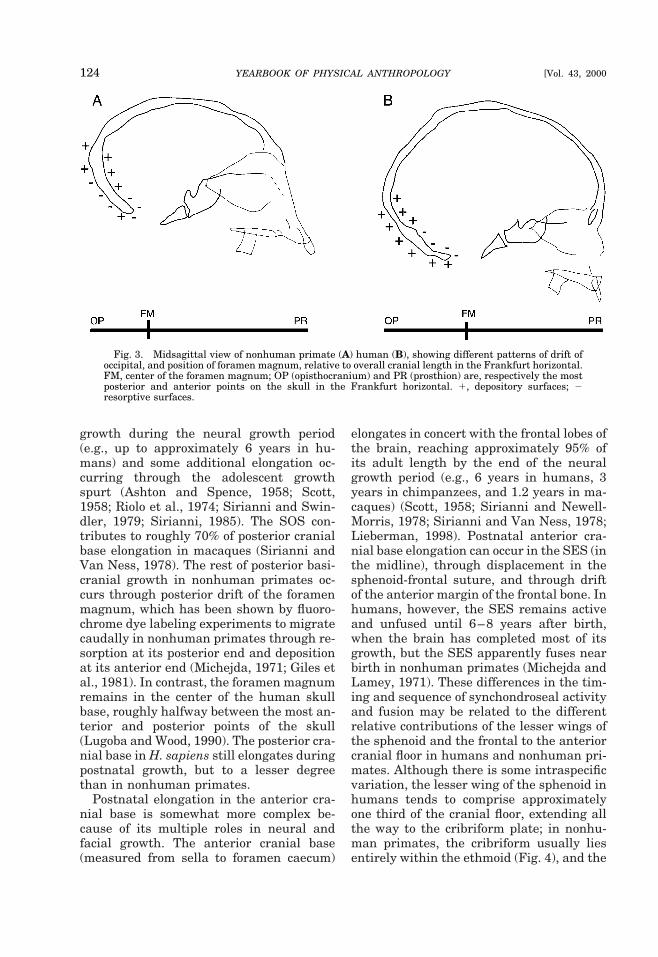
growth during the neural growth period(e.g., up to approximately 6 years in hu-mans) and some additional elongation oc-curring through the adolescent growthspurt (Ashton and Spence, 1958; Scott,1958; Riolo et al., 1974; Sirianni and Swin-dler, 1979; Sirianni, 1985). The SOS con-tributes to roughly 70% of posterior cranialbase elongation in macaques (Sirianni andVan Ness, 1978). The rest of posterior basi-cranial growth in nonhuman primates oc-curs through posterior drift of the foramenmagnum, which has been shown by fluoro-chrome dye labeling experiments to migratecaudally in nonhuman primates through re-sorption at its posterior end and depositionat its anterior end (Michejda, 1971; Giles etal., 1981). In contrast, the foramen magnumremains in the center of the human skullbase, roughly halfway between the most an-terior and posterior points of the skull(Lugoba and Wood, 1990). The posterior cra-nial base in H. sapiens still elongates duringpostnatal growth, but to a lesser degreethan in nonhuman primates.
Postnatal elongation in the anterior cra-nial base is somewhat more complex be-cause of its multiple roles in neural andfacial growth. The anterior cranial base(measured from sella to foramen caecum)
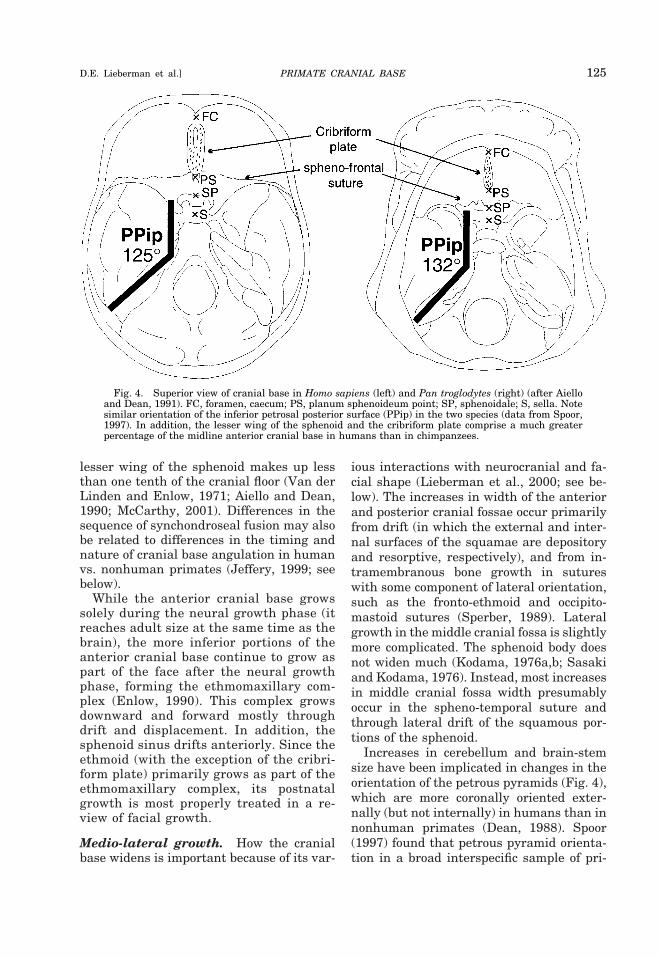
elongates in concert with the frontal lobes ofthe brain, reaching approximately 95% ofits adult length by the end of the neuralgrowth period (e.g., 6 years in humans, 3years in chimpanzees, and 1.2 years in ma-caques) (Scott, 1958; Sirianni and Newell-Morris, 1978; Sirianni and Van Ness, 1978;Lieberman, 1998). Postnatal anterior cra-nial base elongation can occur in the SES (inthe midline), through displacement in thesphenoid-frontal suture, and through driftof the anterior margin of the frontal bone. Inhumans, however, the SES remains activeand unfused until 6–8 years after birth,when the brain has completed most of itsgrowth, but the SES apparently fuses nearbirth in nonhuman primates (Michejda andLamey, 1971). These differences in the tim-ing and sequence of synchondroseal activityand fusion may be related to the differentrelative contributions of the lesser wings ofthe sphenoid and the frontal to the anteriorcranial floor in humans and nonhuman pri-mates. Although there is some intraspecificvariation, the lesser wing of the sphenoid inhumans tends to comprise approximatelyone third of the cranial floor, extending allthe way to the cribriform plate; in nonhu-man primates, the cribriform usually liesentirely within the ethmoid (Fig. 4), and the
Fig. 3. Midsagittal view of nonhuman primate (A) human (B), showing different patterns of drift ofoccipital, and position of foramen magnum, relative to overall cranial length in the Frankfurt horizontal.FM, center of the foramen magnum; OP (opisthocranium) and PR (prosthion) are, respectively the mostposterior and anterior points on the skull in the Frankfurt horizontal. 1, depository surfaces; 2resorptive surfaces.
124 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000
lesser wing of the sphenoid makes up lessthan one tenth of the cranial floor (Van derLinden and Enlow, 1971; Aiello and Dean,1990; McCarthy, 2001). Differences in thesequence of synchondroseal fusion may alsobe related to differences in the timing andnature of cranial base angulation in humanvs. nonhuman primates (Jeffery, 1999; seebelow).
While the anterior cranial base growssolely during the neural growth phase (itreaches adult size at the same time as thebrain), the more inferior portions of theanterior cranial base continue to grow aspart of the face after the neural growthphase, forming the ethmomaxillary com-plex (Enlow, 1990). This complex growsdownward and forward mostly throughdrift and displacement. In addition, thesphenoid sinus drifts anteriorly. Since theethmoid (with the exception of the cribri-form plate) primarily grows as part of theethmomaxillary complex, its postnatalgrowth is most properly treated in a re-view of facial growth.
Medio-lateral growth. How the cranialbase widens is important because of its var-
ious interactions with neurocranial and fa-cial shape (Lieberman et al., 2000; see be-low). The increases in width of the anteriorand posterior cranial fossae occur primarilyfrom drift (in which the external and inter-nal surfaces of the squamae are depositoryand resorptive, respectively), and from in-tramembranous bone growth in sutureswith some component of lateral orientation,such as the fronto-ethmoid and occipito-mastoid sutures (Sperber, 1989). Lateralgrowth in the middle cranial fossa is slightlymore complicated. The sphenoid body doesnot widen much (Kodama, 1976a,b; Sasakiand Kodama, 1976). Instead, most increasesin middle cranial fossa width presumablyoccur in the spheno-temporal suture andthrough lateral drift of the squamous por-tions of the sphenoid.
Increases in cerebellum and brain-stemsize have been implicated in changes in theorientation of the petrous pyramids (Fig. 4),which are more coronally oriented exter-nally (but not internally) in humans than innonhuman primates (Dean, 1988). Spoor(1997) found that petrous pyramid orienta-tion in a broad interspecific sample of pri-
Fig. 4. Superior view of cranial base in Homo sapiens (left) and Pan troglodytes (right) (after Aielloand Dean, 1991). FC, foramen, caecum; PS, planum sphenoideum point; SP, sphenoidale; S, sella. Notesimilar orientation of the inferior petrosal posterior surface (PPip) in the two species (data from Spoor,1997). In addition, the lesser wing of the sphenoid and the cribriform plate comprise a much greaterpercentage of the midline anterior cranial base in humans than in chimpanzees.
PRIMATE CRANIAL BASE 125D.E. Lieberman et al.]

mates was significantly negatively corre-lated with relative brain size (r 5 20.85,P , 0.001) but not with the cranial baseangle. However, Jeffery (1999) found thatpetrous pyramid orientation is independentof relative brain size in fetal humans (dur-ing the second trimester).
Supero-inferior growth. Most braingrowth apparently causes the neurocra-nium and parts of the basicranium to growsuperiorly, anteriorly, and laterally (deBeer, 1937). However, the endocranial fos-sae also become slightly deeper throughdrift because most of the endocranial floor isresorptive, while the inferior side of the ba-sicranium is depository (Fig. 2) (Duterlooand Enlow, 1970; Enlow, 1990). The en-docranial margins between the fossae thatseparate the different portions of the brain(the petrous portion of the temporal and thelesser wing of the sphenoid) do not driftinferiorly because they remain depositorysurfaces (Enlow, 1976). Differences in driftmost likely reflect variation in the relativesize of the components of the brain in con-junction with other spatial relationshipsamong components of the skull. In particu-lar, inferior drift of the anterior cranialfossa is presumably minimal because itwould impinge upon the orbits and nasalcavity that lie immediately below. The onlyexception is the cribriform plate whichdrifts inferiorly, slightly in humans (Moss,1963), but sometimes forming a “deep olfac-tory pit” in many species of nonhuman pri-mates (Cameron, 1930; Aiello and Dean,1990). Inferior drift of the middle cranialfossa presumably reflects inferiorly directedgrowth of the temporal lobes, but this hy-pothesis has not been tested. Likewise, in-ferior drift in the posterior cranial fossa,which is shallow in most nonhuman pri-mates, is hypothesized to be a function ofthe size of the occipital lobes, the cerebel-lum, and the brain stem below the tento-rium cerebelli. Note that cranial base flex-ion during growth, which occurs uniquely inhumans (see below), complements inferiordrift in the posterior cranial fossa by movingthe floor of the posterior cranial fossa morebelow the middle cranial fossa.
Angulation. Angulation of the cranialbase occurs when the prechordal and post-chordal portions of the basicranium flex orextend relative to each other in the midsag-ittal plane (technically, flexion and exten-sion describe a series of events in which theangle between the inferior or ventral sur-faces of the cranial base decrease or in-crease, respectively). Angulation has beenthe subject of much research because flexionand extension of the cranial base affect therelative positions of the three endocranialfossae, thereby influencing a wide range ofspatial relationships among the cranialbase, brain, face, and pharynx (see below).
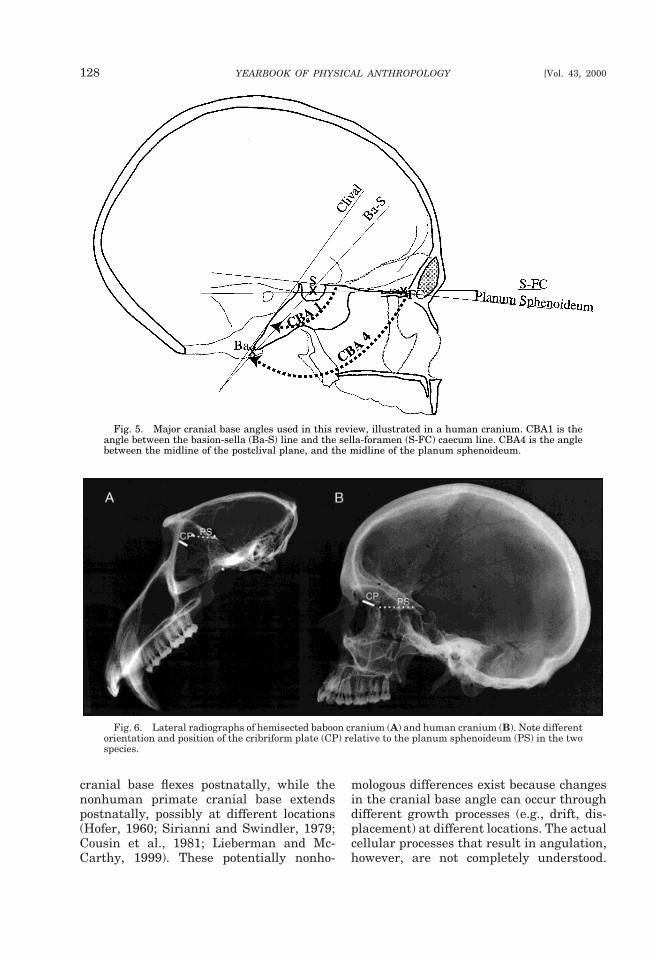
Although all measures of cranial base an-gle are similar in that they attempt to quan-tify the overall degree of angulation in themidsagittal plane between the prechordaland postchordal portions of the cranial base,there have been at least 17 different mea-surements used since Huxley (1867) firstattempted to quantify the angle (reviewedin Lieberman and McCarthy, 1999, andsummarized in Table 1). Many of these an-gles differ considerably in how they mea-sure the prechordal and postchordal planesand, consequently, the point of intersectionbetween them. Figure 5 illustrates some ofthese angles. The postchordal plane is mostcommonly defined using two landmarks,usually basion and sella, or using the linecreated by the dorsal surface of the basioc-cipital clivus (the clival line). The pre-chordal plane has been measured in morediverse ways. Historically, the most com-mon plane is defined by two landmarks,sella and nasion. The sella-nasion line isproblematic, however, because nasion is ac-tually part of the face and moves anteriorlyand inferiorly relative to the cranial basethroughout the period of facial growth(Scott, 1958; Enlow, 1990). Recently, mostresearchers have defined the prechordalplane either from sella to the foramen cae-cum (a pit on the anterior end of the cribri-form plate between the crista galli betweenfrontal squama), or using the planum sphe-noideum which extends from sphenoidale(the most postero-superior point on the tu-berculum sellae) to the planum sphenoi-deum point (defined as the most anteriorpoint on the surface of the midline anterior
126 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000

cranial base posterior to the cribriformplate).
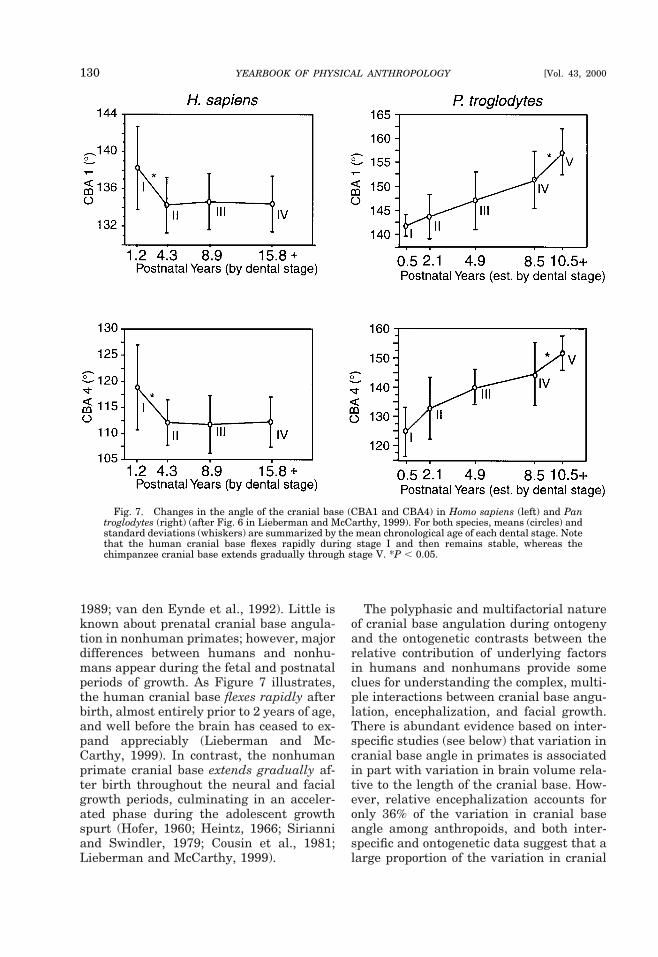
Since different lines emphasize differentaspects of cranial base anatomy, the choiceof which cranial base angle to use is largelydependent on the question under study.Both postchordal lines tend to yield roughlysimilar results (George, 1978; Liebermanand McCarthy, 1999), but the prechordallines can be substantially different. In par-ticular, S-FC spans the entire length of theanterior cranial base, including the cribri-form plate, whereas the planum sphenoi-deum does not measure the portion of thecranial base that includes the cribriformplate. Because of variation in the growthand position of the cribriform plate, thesedifferences affect comparisons of anthro-poids with strepsirrhines, or comparisons of
primates with other mammals (McCarthy,in press). In humans and some anthropoids,the cribriform plate lies in approximatelythe same plane as the planum sphenoi-deum, but in other anthropoids the cribri-form plate lies in a deep olfactory pit withinthe ethmoidal notch of the frontal bone (Fig.6) (Aiello and Dean, 1990; Ravosa and Shea,1994). Moreover, in strepsirrhines andother mammals with more divergent orbitsand projecting snouts, the cribriform platetypically lies at a steep angle relative to theplanum sphenoideum (Cartmill, 1970).
Variations in cranial base angulationneed to be considered in both comparativeand ontogenetic studies. For example, it iswell known that humans have a much moreflexed cranial base than other primates, butit is not well appreciated that the human
TABLE 1. Commonly used measures of midsagittal cranial base angle
AnglePosterior (P) and anterior (A)
planes used References
External cranial base angle P: basion-sella Bjork, 1951, 1955; Stamrud, 1959Nasion-sella-basion A: sella-nasion Melsen, 1969; George, 1978
Landzert’s sphenoidal angleClivus/clival angleCBA4, planum angle
P: clival planeA: ethmoidal plane (planum
sphenoideum/ale)
Landzert, 1866; Biegert, 1957;Moss, 1958; Hofer, 1957;Angst, 1967; Cartmill, 1970;Dmoch, 1975; Ross andRavosa, 1993; Ross andHenneberg, 1995
Clivus angle P: clival plane George, 1978A: sphenoidale-fronton
Ethmoidal angle P: basion-sella Stamrud, 1959Internal cranial base angle A: sella-ethmoidale
Spheno-ethmoidal angle P: basion-prosphenion Huxley, 1867; Topinard, 1890;Duckworth, 1904; Cameron,1924; Zuckerman, 1955
Cameron’s cranio-facial axis P: basion-pituitary pointA: pituitary point-nasion
Cameron, 1924, 1925, 1927a,b,1930
Basioccipito-septal angle P: basion-pituitary point Ford, 1956A: pituitary point-septal point
Bolton’s external cranial base angle P: Bolton point-sellaA: sella-nasion
Brodie, 1941, 1953
Anterior cranial base angle P: clival plane Scott, 1958; Cramer, 1977A: prosphenion-anterior cribriform
point (ACP)Internal cranial base angle, basion-
sphenoidale-frontonP: basion-sphenoidaleA: sphenoidale-fronton
George, 1978
Internal cranial base angle, basion-sella-fronton
P: basion-sellaA: sella-fronton
George, 1978
Internal cranial base angle, basion-sella-foramen caecum CBA1
P: basion-sellaA: sella-foramen caecum
Cousin et al., 1981; Spoor, 1997Lieberman and McCarthy,1999
External cranial base angle, nasion-sphenoidale-basion
P: basion-sphenoidaleA: sphenoidale-nasion
George, 1978
Orbital angle P: clival plane Moss, 1958A: plane of superior orbital roof
Planum angle (PANG) P: basion-sella Anton, 1989A: planum sphenoidale
Orbital angle (OANG) P: basion-sella Anton, 1989A: plane of superior orbital roof
PRIMATE CRANIAL BASE 127D.E. Lieberman et al.]
cranial base flexes postnatally, while thenonhuman primate cranial base extendspostnatally, possibly at different locations(Hofer, 1960; Sirianni and Swindler, 1979;Cousin et al., 1981; Lieberman and Mc-Carthy, 1999). These potentially nonho-
mologous differences exist because changesin the cranial base angle can occur throughdifferent growth processes (e.g., drift, dis-placement) at different locations. The actualcellular processes that result in angulation,however, are not completely understood.
Fig. 5. Major cranial base angles used in this review, illustrated in a human cranium. CBA1 is theangle between the basion-sella (Ba-S) line and the sella-foramen (S-FC) caecum line. CBA4 is the anglebetween the midline of the postclival plane, and the midline of the planum sphenoideum.
Fig. 6. Lateral radiographs of hemisected baboon cranium (A) and human cranium (B). Note differentorientation and position of the cribriform plate (CP) relative to the planum sphenoideum (PS) in the twospecies.
128 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000

Some researchers (Scott, 1958; Giles et al.,1981; Enlow, 1990) suggest that changes incranial base angulation occur interstitiallywithin synchondroses through a hinge-likeaction. If so, flexion would result from in-creased chrondrogenic activity in the supe-rior vs. inferior aspect of the synchondrosis,while extension would result from increasedchrondrogenic activity in the inferior vs. su-perior aspect of the synchondrosis. Experi-mental growth studies in macaques, whichlabeled growth using flurochrome dyes,show that angulation also occurs throughdrift in which depository and resorptivegrowth fields differ on either side of a syn-chondrosis, causing rotations around anaxis through the synchondrosis (Michejda,1971, 1972a; Michejda and Lamey, 1971;Giles et al., 1981). All three synchondrosesare involved in prenatal angulation (Hofer,1960; Hofer and Spatz, 1963; Sirianni andNewell-Morris, 1980; Diewert, 1985; Ana-gastopolou et al., 1988; Sperber, 1989; vanden Eynde et al., 1992); however, the extentto which each synchondrosis participates inpostnatal flexion and extension is poorlyknown, and probably differs between hu-mans and nonhuman primates. The SOS,which remains active until after the erup-tion of the second permanent molars, isprobably the most active synchondrosis ingenerating angulation in primates (Bjork,1955; Scott, 1958; Melsen, 1969). The MSSfuses prior to birth in humans (Ford, 1958),but may also be important in nonhumanprimates (Scott, 1958; Hofer and Spatz,1963; Michejda, 1971, 1972a; but see Lager,1958; Melsen, 1971; Giles et al., 1981). Fi-nally, the SES fuses near birth in nonhu-man primates, and remains active only as asite of cranial base elongation in humansduring the neural growth period (Scott,1958; Michejda and Lamey, 1971). Otherontogenetic changes in the cranial base an-gle (not necessarily involved in angulationitself) include posterior drift of the foramenmagnum (see above), inferior drift of thecribriform plate relative to the anterior cra-nial base (Moss, 1963), and remodeling ofthe sella turcica, which causes posteriormovement of the sella (Baume, 1957; Sha-piro, 1960; Latham, 1972).
A few experimental studies provide evi-dence for the presence of complex interac-tions between the brain and cranial basesynchondroses that influence variation inthe cranial base angle. DuBrul and Laskin(1961), Moss (1976), Butow (1990), and Rei-denberg and Laitman (1991) all inhibitedgrowth in the SOS in various animals(mostly rats), causing a more flexed cranialbase, presumably through inhibition of cra-nial base extension. In most of these stud-ies, experimentally induced kyphosis of thebasicranium was also associated with ashorter posterior portion of the cranial base,and a more rounded neurocranium (see be-low). Artificial deformation of the cranialvault also causes slight but significant in-creases in cranial base angulation (Anton,1989; Kohn et al., 1993). However, no con-trolled experimental studies have yet exam-ined disruptions to the other cranial basesynchondroses. In addition, there have beenfew controlled studies of the effect of in-creasing brain size on cranial base angula-tion. In one classic experiment, Young(1959) added sclerosing fluid into the cra-nial cavity in growing rats, which causedenlargement of the neurocranium with littleeffect on angular relationships in the cra-nial base. Additional evidence for some de-gree of independence between the brain andcranial base during development is providedby microcephaly and hydrocephaly, inwhich cranial base angles tend to be close tothose of humans with normal encephaliza-tion (Moore and Lavelle, 1974; Sperber,1989).
Important differences in cranial base an-gulation among primates exist in terms ofthe ontogenetic pattern of flexion and/or ex-tension, which presumably result from dif-ferences in the rate, timing, duration, andsequence of the growth processes outlinedabove. Jeffery (1999) suggested that, prena-tally, the basicranium in humans initiallyflexes rapidly during the period of rapidhindbrain growth in the first trimester, re-mains fairly stable during the second tri-mester, and then extends during the thirdtrimester in conjunction with facial exten-sion, even while the brain is rapidly increas-ing in size relative to the rest of the cranium(see also Bjork, 1955; Ford, 1956; Sperber,
PRIMATE CRANIAL BASE 129D.E. Lieberman et al.]
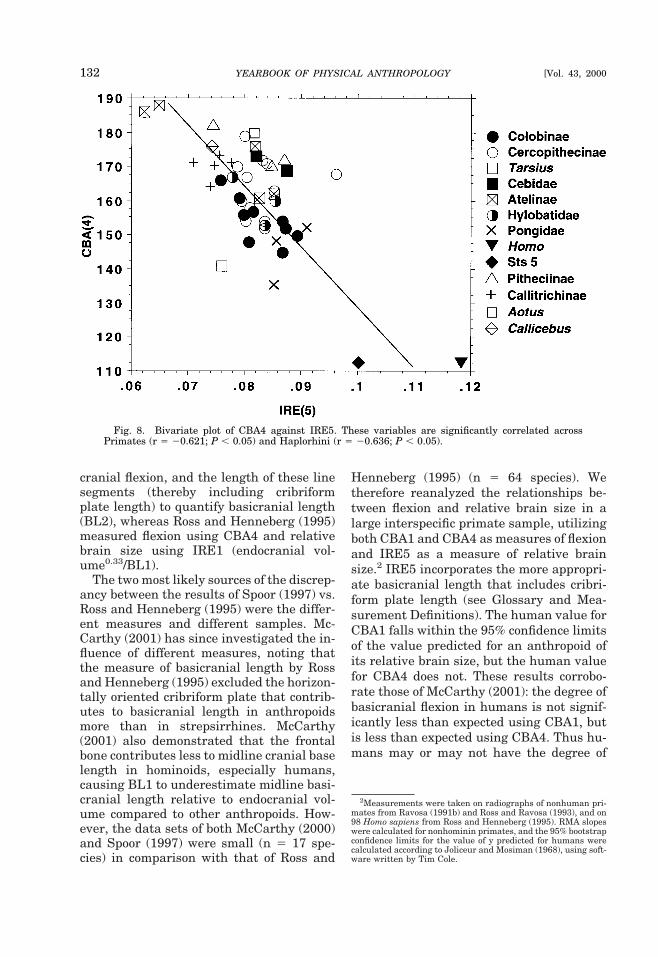
1989; van den Eynde et al., 1992). Little isknown about prenatal cranial base angula-tion in nonhuman primates; however, majordifferences between humans and nonhu-mans appear during the fetal and postnatalperiods of growth. As Figure 7 illustrates,the human cranial base flexes rapidly afterbirth, almost entirely prior to 2 years of age,and well before the brain has ceased to ex-pand appreciably (Lieberman and Mc-Carthy, 1999). In contrast, the nonhumanprimate cranial base extends gradually af-ter birth throughout the neural and facialgrowth periods, culminating in an acceler-ated phase during the adolescent growthspurt (Hofer, 1960; Heintz, 1966; Sirianniand Swindler, 1979; Cousin et al., 1981;Lieberman and McCarthy, 1999).
The polyphasic and multifactorial natureof cranial base angulation during ontogenyand the ontogenetic contrasts between therelative contribution of underlying factorsin humans and nonhumans provide someclues for understanding the complex, multi-ple interactions between cranial base angu-lation, encephalization, and facial growth.There is abundant evidence based on inter-specific studies (see below) that variation incranial base angle in primates is associatedin part with variation in brain volume rela-tive to the length of the cranial base. How-ever, relative encephalization accounts foronly 36% of the variation in cranial baseangle among anthropoids, and both inter-specific and ontogenetic data suggest that alarge proportion of the variation in cranial
Fig. 7. Changes in the angle of the cranial base (CBA1 and CBA4) in Homo sapiens (left) and Pantroglodytes (right) (after Fig. 6 in Lieberman and McCarthy, 1999). For both species, means (circles) andstandard deviations (whiskers) are summarized by the mean chronological age of each dental stage. Notethat the human cranial base flexes rapidly during stage I and then remains stable, whereas thechimpanzee cranial base extends gradually through stage V. *P , 0.05.
130 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000

base angle among primates must also berelated to variation in facial growth, orbitorientation, and relative orbit size (Ross andRavosa, 1993; Ravosa et al., 2000a). Asnoted above, the ontogenetic pattern of pre-natal cranial base angulation in humans islargely unrelated to the rate at which thebrain expands (Jeffery, 1999). In addition,the nonhuman primate cranial base angle(regardless of whether the cribriform plateis included in the measurement) mostly ex-tends during the period of facial growth,after the brain has ceased to expand(Lieberman and McCarthy, 1999). There-fore, we will next explore in greater depththe relationship between cranial base angle,brain size, relative orbit size and position,facial orientation, and other factors such aspharyngeal shape and facial projection.
ASSOCIATIONS BETWEEN CRANIALBASE AND BRAIN
Because of the close relationship betweenthe brain and the cranial base during devel-opment (see above), the hypothesis thatbrain size and shape influence basicranialmorphology is an old and persistent one.The bones of the cranial cavity, includingthe cranial base, are generally known toconform to the shape of the brain, but thespecifics of this relationship and any recip-rocal effects of cranial base size and shapeon brain morphology remain unclear. Forexample, the human basicranium is flexedwhen it first appears in weeks 5 and 6 be-cause in the fourth week, the neural tubebends ventrally at the cephalic flexure(O’Rahilly and Muller, 1994). The para-chordal condensations caudal to the ce-phalic flexure are therefore in a differentanatomical plane than the more rostral pre-chordal condensations (which develop byweek 7). However, as noted above, it is dif-ficult to attribute many of the subsequentchanges in prenatal chondrocranial or basi-cranial angulation (or other measures of thebase) as responses solely to changes in brainmorphology.
Here we review several key aspects of theassociation between brain and cranial basemorphology, as derived from interspecificanalyses of adult specimens. Structural re-
lationships between the cranial base andthe face are discussed below.
Brain size and cranial base angle
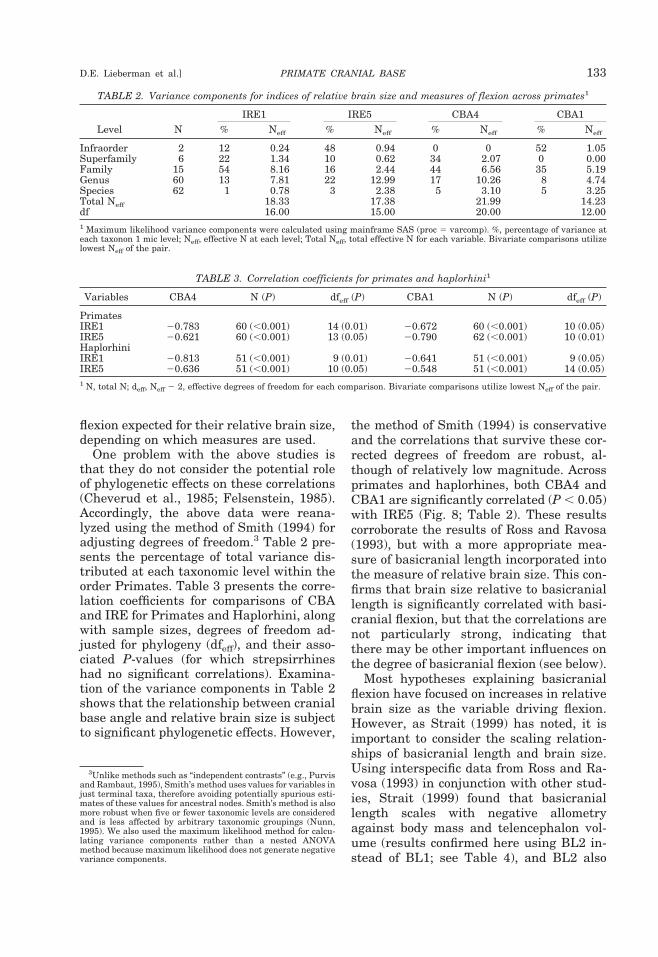
Numerous anatomists have posited a re-lationship between brain size and basicra-nial angle (e.g., Virchow, 1857; Ranke, 1892;Cameron, 1924; Bolk, 1926; Dabelow, 1929,1931; Biegert, 1957, 1963; Delattre and Fe-nart, 1963; Hofer, 1969; Gould, 1977; Rossand Ravosa, 1993; Ross and Henneberg,1995; Spoor, 1997; Strait, 1999; Strait andRoss, 1999; McCarthy, 2001). The mostwidely accepted of these hypotheses is thatthe angle of the midline cranial base in thesagittal plane correlates with the volume ofthe brain relative to basicranial length(DuBrul and Laskin, 1961; Vogel, 1964;Riesenfeld, 1969; Gould, 1977). This hypoth-esis is supported by independent analyses ofdifferent measures of basicranial flexionacross several interspecific samples of pri-mates (Ross and Ravosa, 1993; Spoor, 1997;McCarthy, 2001) (Fig. 8): the adult midlinecranial base is significantly and predictablymore flexed in species with larger endocra-nial volumes relative to basicranial length.In particular, the analysis by Ross and Ra-vosa (1993) of a broad interspecific sampleof primates found that the correlation coef-ficient between relative encephalization(IRE1, see below) and cranial base angle(CBA4, see below) was 0.645 (P , 0.001),explaining approximately 40% of the varia-tion in cranial base angle.
Attempts to extend this relationship tohominins have proved controversial. Rossand Henneberg (1995) reported that Homosapiens have less flexed basicrania thanpredicted by either haplorhine or primateregressions. They posited that spatial con-straints limit the degree of flexion possible,and that humans accommodate furtherbrain expansion relative to cranial baselength through means other than flexion,such as superior, posterior, and lateral neu-rocranial expansion. In contrast, Spoor(1997), using different measures of flexionand relative brain size taken on a differentsample, found H. sapiens to have the degreeof flexion expected for its relative brain size.Spoor (1997) used the angle basion-sella-foramen caecum (CBA1) to quantify basi-
PRIMATE CRANIAL BASE 131D.E. Lieberman et al.]
cranial flexion, and the length of these linesegments (thereby including cribriformplate length) to quantify basicranial length(BL2), whereas Ross and Henneberg (1995)measured flexion using CBA4 and relativebrain size using IRE1 (endocranial vol-ume0.33/BL1).
The two most likely sources of the discrep-ancy between the results of Spoor (1997) vs.Ross and Henneberg (1995) were the differ-ent measures and different samples. Mc-Carthy (2001) has since investigated the in-fluence of different measures, noting thatthe measure of basicranial length by Rossand Henneberg (1995) excluded the horizon-tally oriented cribriform plate that contrib-utes to basicranial length in anthropoidsmore than in strepsirrhines. McCarthy(2001) also demonstrated that the frontalbone contributes less to midline cranial baselength in hominoids, especially humans,causing BL1 to underestimate midline basi-cranial length relative to endocranial vol-ume compared to other anthropoids. How-ever, the data sets of both McCarthy (2000)and Spoor (1997) were small (n 5 17 spe-cies) in comparison with that of Ross and
Henneberg (1995) (n 5 64 species). Wetherefore reanalyzed the relationships be-tween flexion and relative brain size in alarge interspecific primate sample, utilizingboth CBA1 and CBA4 as measures of flexionand IRE5 as a measure of relative brainsize.2 IRE5 incorporates the more appropri-ate basicranial length that includes cribri-form plate length (see Glossary and Mea-surement Definitions). The human value forCBA1 falls within the 95% confidence limitsof the value predicted for an anthropoid ofits relative brain size, but the human valuefor CBA4 does not. These results corrobo-rate those of McCarthy (2001): the degree ofbasicranial flexion in humans is not signif-icantly less than expected using CBA1, butis less than expected using CBA4. Thus hu-mans may or may not have the degree of
2Measurements were taken on radiographs of nonhuman pri-mates from Ravosa (1991b) and Ross and Ravosa (1993), and on98 Homo sapiens from Ross and Henneberg (1995). RMA slopeswere calculated for nonhominin primates, and the 95% bootstrapconfidence limits for the value of y predicted for humans werecalculated according to Joliceur and Mosiman (1968), using soft-ware written by Tim Cole.
Fig. 8. Bivariate plot of CBA4 against IRE5. These variables are significantly correlated acrossPrimates (r 5 20.621; P , 0.05) and Haplorhini (r 5 20.636; P , 0.05).
132 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000
flexion expected for their relative brain size,depending on which measures are used.
One problem with the above studies isthat they do not consider the potential roleof phylogenetic effects on these correlations(Cheverud et al., 1985; Felsenstein, 1985).Accordingly, the above data were reana-lyzed using the method of Smith (1994) foradjusting degrees of freedom.3 Table 2 pre-sents the percentage of total variance dis-tributed at each taxonomic level within theorder Primates. Table 3 presents the corre-lation coefficients for comparisons of CBAand IRE for Primates and Haplorhini, alongwith sample sizes, degrees of freedom ad-justed for phylogeny (dfeff), and their asso-ciated P-values (for which strepsirrhineshad no significant correlations). Examina-tion of the variance components in Table 2shows that the relationship between cranialbase angle and relative brain size is subjectto significant phylogenetic effects. However,
the method of Smith (1994) is conservativeand the correlations that survive these cor-rected degrees of freedom are robust, al-though of relatively low magnitude. Acrossprimates and haplorhines, both CBA4 andCBA1 are significantly correlated (P , 0.05)with IRE5 (Fig. 8; Table 2). These resultscorroborate the results of Ross and Ravosa(1993), but with a more appropriate mea-sure of basicranial length incorporated intothe measure of relative brain size. This con-firms that brain size relative to basicraniallength is significantly correlated with basi-cranial flexion, but that the correlations arenot particularly strong, indicating thatthere may be other important influences onthe degree of basicranial flexion (see below).
Most hypotheses explaining basicranialflexion have focused on increases in relativebrain size as the variable driving flexion.However, as Strait (1999) has noted, it isimportant to consider the scaling relation-ships of basicranial length and brain size.Using interspecific data from Ross and Ra-vosa (1993) in conjunction with other stud-ies, Strait (1999) found that basicraniallength scales with negative allometryagainst body mass and telencephalon vol-ume (results confirmed here using BL2 in-stead of BL1; see Table 4), and BL2 also
3Unlike methods such as “independent contrasts” (e.g., Purvisand Rambaut, 1995), Smith’s method uses values for variables injust terminal taxa, therefore avoiding potentially spurious esti-mates of these values for ancestral nodes. Smith’s method is alsomore robust when five or fewer taxonomic levels are consideredand is less affected by arbitrary taxonomic groupings (Nunn,1995). We also used the maximum likelihood method for calcu-lating variance components rather than a nested ANOVAmethod because maximum likelihood does not generate negativevariance components.
TABLE 2. Variance components for indices of relative brain size and measures of flexion across primates1
Level NIRE1 IRE5 CBA4 CBA1
% Neff % Neff % Neff % Neff
Infraorder 2 12 0.24 48 0.94 0 0 52 1.05Superfamily 6 22 1.34 10 0.62 34 2.07 0 0.00Family 15 54 8.16 16 2.44 44 6.56 35 5.19Genus 60 13 7.81 22 12.99 17 10.26 8 4.74Species 62 1 0.78 3 2.38 5 3.10 5 3.25Total Neff 18.33 17.38 21.99 14.23df 16.00 15.00 20.00 12.001 Maximum likelihood variance components were calculated using mainframe SAS (proc 5 varcomp). %, percentage of variance ateach taxonon 1 mic level; Neff, effective N at each level; Total Neff, total effective N for each variable. Bivariate comparisons utilizelowest Neff of the pair.
TABLE 3. Correlation coefficients for primates and haplorhini1
Variables CBA4 N (P) dfeff (P) CBA1 N (P) dfeff (P)
PrimatesIRE1 20.783 60 (,0.001) 14 (0.01) 20.672 60 (,0.001) 10 (0.05)IRE5 20.621 60 (,0.001) 13 (0.05) 20.790 62 (,0.001) 10 (0.01)HaplorhiniIRE1 20.813 51 (,0.001) 9 (0.01) 20.641 51 (,0.001) 9 (0.05)IRE5 20.636 51 (,0.001) 10 (0.05) 20.548 51 (,0.001) 14 (0.05)1 N, total N; deff, Neff 2 2, effective degrees of freedom for each comparison. Bivariate comparisons utilize lowest Neff of the pair.
PRIMATE CRANIAL BASE 133D.E. Lieberman et al.]

scales with negative allometry against mea-sures of facial size (Table 4). The only vari-able that scales close to isometry with basi-cranial length is brain-stem volume, whichis isometric with BL1 and scales close toisometry with BL2 (Table 4). Intuitively thisisometry makes sense, because the brainstem rests on the basicranium. However,the posterior basicranium (Ba-S), whichpredominantly underlies the brain stem,scales with positive allometry to brain-stemvolume, whereas the anterior cranial baselength scales isometrically with brain-stemvolume. Notably, the anterior cranial basealways shows lower slopes than the poste-rior cranial base, suggesting that the strongnegative allometry of cranial base lengthrelative to neural variables, and possiblyflexion, is disproportionately attributable to
relative shortening of the anterior cranialbase.
Brain shape and cranial base angle
Given the close anatomical relationshipbetween the brain and basicranium, itseems intuitive that the shapes of the twoshould be related, but testing this hypothe-sis in a controlled fashion has been difficult.The best evidence for the presence of inter-actions between brain shape and the cranialbase came from studies of artificial cranialvault deformation and from the effects ofclosing various cranial vault sutures on thecranial base. The effects of head-binding aredifficult to interpret without precise data onthe timing and forces used to deform theskull. Nevertheless, antero-posterior head-binding tends to cause lateral expansion of
TABLE 4. Reduced major axis regression equations for scaling of basicranial dimensions againstneural variables1
Comparison Intercept Slope95% CIof slope Scaling3 N r
BL2 vs.Body mass 0.908 0.654 0.035 2ve 61 0.978Telencephalon 0.541 0.742 0.091 2ve 33 0.960Brain stem 0.404 1.123 0.100 1ve/Iso 33 0.975Neurocranial volume 1.270 0.734 0.056 2ve 62 0.962Palate (Pros-PNS) 1.394 0.642 0.032 2ve 58 0.929Anterior face (Pros-Nas) 1.623 0.577 0.028 2ve 58 0.933Upper toothroow 1.425 0.666 0.035 2ve 58 0.921Geometric mean2 0.951 0.750 0.023 2ve 58 0.972
Ba-S vs.Body mass 0.380 0.759 0.056 2ve 63 0.958Brain stem 20.226 1.354 0.161 1ve 33 0.946Telencephalon 0.313 0.908 0.135 Iso 33 0.924Cerebellum 0.105 0.953 0.111 Iso 33 0.948Infratentorial brain 20.064 1.044 0.123 Iso 33 0.947Palate (Pros-PNS) 0.595 0.757 0.040 2ve 58 0.917Anterior face (Pros-Nas) 0.862 0.681 0.037 2ve 58 0.914Upper toothroow 0.626 0.787 0.043 2ve 58 0.924Geometric mean2 0.067 0.886 0.034 2ve 58 0.957
S-FC vs.Ba-S 0.473 0.759 0.063 2ve 33 0.949Body mass 0.748 0.587 0.040 2ve 56 0.965Brain stem 0.249 1.070 0.101 Iso 33 0.971Telencephalon 0.378 0.715 0.082 2ve 33 0.963Cerebellum 0.516 0.749 0.079 2ve 33 0.964Infratentorial brain 0.381 0.822 0.085 2ve 33 0.965Palate (Pros-PNS) 1.382 0.581 0.032 2ve 58 0.912Anterior face (Pros-Nas) 1.586 0.523 0.027 2ve 58 0.922Upper toothroow 1.406 0.604 0.035 2ve 58 0.900Geometric mean2 0.977 0.680 0.026 2ve 58 0.9581 All volumetric and mass variables converted to cube roots. All calculations in log-log space (base 10). Calculations with facialdimensions do not include humans; all other calculations include humans.2 Geometric mean of the following measures: palate length, palate breadth, anterior face length, maxillary postcanine toothrowlength, outer biorbital breadth, bizygomatic breadth, basicranial length, and lower skull length. Measures of brain volume fromStephan et al. (1981), measures of body mass from Smith and Jungers (1994), and measures of facial size derive from the same skullsfrom which the radiographs were taken.3 2ve, negative; 1ve, positive; Iso, isometric.
134 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000
the cranial base along with a slight increasein CBA; conversely, annular head-bindingtends to cause medio-lateral narrowing andantero-posterior elongation of the cranialbase, also with a slight increase in CBA(Anton, 1989; Cheverud et al., 1992; Kohn etal., 1993). Natural or experimentally in-duced premature closure of sutures (synos-toses) in the cranial vault have similarlypredictable effects. For example, bilateralcoronal synostoses cause antero-posteriorshortening of the cranial base (Babler, 1989;David et al., 1989), and unilateral coronalsynostoses (plagiocephaly) cause markedasymmetry in the cranial vault, cranialbase, and face.
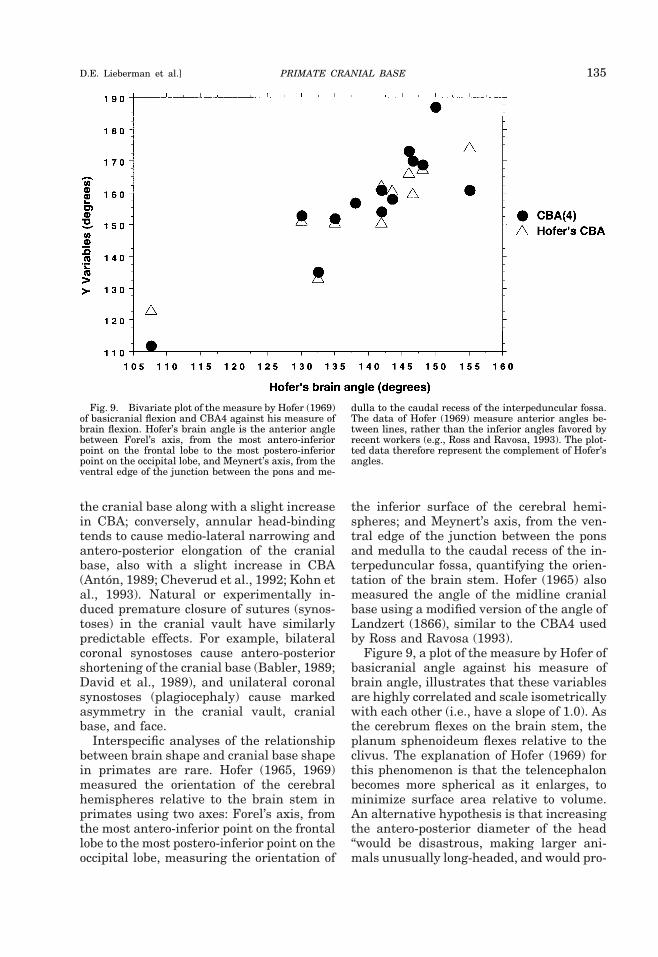
Interspecific analyses of the relationshipbetween brain shape and cranial base shapein primates are rare. Hofer (1965, 1969)measured the orientation of the cerebralhemispheres relative to the brain stem inprimates using two axes: Forel’s axis, fromthe most antero-inferior point on the frontallobe to the most postero-inferior point on theoccipital lobe, measuring the orientation of
the inferior surface of the cerebral hemi-spheres; and Meynert’s axis, from the ven-tral edge of the junction between the ponsand medulla to the caudal recess of the in-terpeduncular fossa, quantifying the orien-tation of the brain stem. Hofer (1965) alsomeasured the angle of the midline cranialbase using a modified version of the angle ofLandzert (1866), similar to the CBA4 usedby Ross and Ravosa (1993).
Figure 9, a plot of the measure by Hofer ofbasicranial angle against his measure ofbrain angle, illustrates that these variablesare highly correlated and scale isometricallywith each other (i.e., have a slope of 1.0). Asthe cerebrum flexes on the brain stem, theplanum sphenoideum flexes relative to theclivus. The explanation of Hofer (1969) forthis phenomenon is that the telencephalonbecomes more spherical as it enlarges, tominimize surface area relative to volume.An alternative hypothesis is that increasingthe antero-posterior diameter of the head“would be disastrous, making larger ani-mals unusually long-headed, and would pro-
Fig. 9. Bivariate plot of the measure by Hofer (1969)of basicranial flexion and CBA4 against his measure ofbrain flexion. Hofer’s brain angle is the anterior anglebetween Forel’s axis, from the most antero-inferiorpoint on the frontal lobe to the most postero-inferiorpoint on the occipital lobe, and Meynert’s axis, from theventral edge of the junction between the pons and me-
dulla to the caudal recess of the interpeduncular fossa.The data of Hofer (1969) measure anterior angles be-tween lines, rather than the inferior angles favored byrecent workers (e.g., Ross and Ravosa, 1993). The plot-ted data therefore represent the complement of Hofer’sangles.
PRIMATE CRANIAL BASE 135D.E. Lieberman et al.]

duce serious problems for balancing theskull on the skeleton” (Jerison, 1982, p. 82).In the context of such spatial constraints(i.e., limited cerebral diameter and tendencyto sphericity), increased cerebrum size canonly be accommodated by expanding thecranial base inferiorly, posteriorly, or ante-riorly, thereby necessarily flexing the brainand the basicranium. The hypothesis ofJerison (1982) is refuted by animals such ascamels, llamas, and giraffes, which havelong heads on relatively orthograde necks;and as discussed below, there is no convinc-ing evidence that head and neck posture aresignificant influences on basicranial anglein primates (Strait and Ross, 1999).
Biegert (1963) made a claim similar tothat of Hofer (1969), in arguing that in-creases in primate brain size relative to cra-nial base size, as well as increases in “neo-pallium” (i.e., neocortex) size relative toother parts of the brain, produced a rounderbrain such that “adaptations in the struc-ture of the cranium accompanied thesechanges in the size and shape of the brain”(Biegert, 1963, p. 120). The predictedchanges in skull shape include increasedvaulting of the frontal and occipital bonesand increased basicranial flexion. Ross andRavosa (1993) evaluated the hypothesis ofBiegert (1963) by calculating correlation co-efficients between cranial base angle(CBA4) and the ratio of neocortical vol-ume0.33/basicranial length. Although theyfound a significant relationship across pri-mates and haplorhines, the correlation coef-ficients were low (around 0.5). Recalculationof these correlation coefficients using BL2as a measure of basicranial length producessignificant correlations across primates,haplorhines, and strepsirrhines, but onlythe primate level correlations survive ad-justed degrees of freedom (Table 5).
Strait (1999) proposed a hypothesis simi-lar to those of Hofer (1969) and Biegert(1963). Noting that total basicranial lengthscales close to isometry with noncorticalbrain volume, Strait (1999) suggested thatvariation in the midline basicranial anglemight be due to increases in the size of thetelencephalon relative to the noncorticalpart of the brain,4 rather than the size of thebrain relative to the cranial base. Analysisof our data set confirms this hypothesis:there are significant correlations betweenflexion and the size of the telencephalonrelative to the brain stem across primates,using CBA4 (Table 5). Moreover, the onlysignificant correlation between cranial baseangle and a neural variable among strepsir-rhines is the comparison of CBA4 with theratio of telencephalon to brain-stem volume(Table 5). However, despite relatively highcorrelation coefficients for haplorhines, thesecorrelations do not remain significant withphylogenetically adjusted degrees of free-dom. This suggests that brain size relativeto basicranial length may be a better expla-nation for flexion, because it appears to bemore independent of phylogenetic effects.
Whether basicranial flexion accommo-dates increases in telencephalon volume rel-ative to brain-stem volume, and/or in-creases in overall endocranial volumerelative to cranial base length, the end re-sult is a change in brain shape. The en-larged telencephalon of primates is an out-growth of the rostral end of the brain stemthat communicates with the rest of thebrain through the diencephalon at its root.
4Strait (1999) refers to this as the “noncortical scaling hypoth-esis;” however, the telencephlon consists of more than cortex: italso includes the white matter and the basal ganglia. Here weevaluate the role of relative telencephalon volume in producingflexion.
TABLE 5. Correlation coefficients for flexion vs. measures of relative brain size
N Neff (df) CBA4 CBA1
Neocortex volume/BL2Primates 29 12 (10) 20.613 (0.05) 20.759 (0.01)Haplorhines 22 9 (7) 20.636 (ns) 20.660 (ns)Strepsirhines 7 6 (4) 20.282 (ns) 0.771 (ns)Telencephalon/brain-stem volumePrimates 29 9 (7) 20.744 (0.05) 20.663 (ns)Haplorhines 19 6 (4) 20.726 (ns) 20.643 (ns)Strepsirhines 9 7 (5) 20.760 (0.05) 20.286 (ns)
136 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000

Increasing the size of the outer cortex of thetelencephalon (the neocortex) while stillconnecting to the rest of the brain throughthe diencephalon might be expected to gen-erate a more spheroid shape, regardless ofany functional constraints on skull or brainshape. In other words, the telencephalonmay be spheroidal because of the geometryof its connections and the way it develops,rather than for any functional or adaptivereason. Alternately, a spheroidal cerebrummay minimize “wiring length” in the brain,a potentially important principle of designin neural architecture (Allman and Kaas,1974; Barlow, 1986; Mitchison, 1991; Cher-niak, 1995; Van Essen, 1997). Accordingly, aspheroid telencephalon may optimize neo-cortical wiring lengths as well as minimizethe distance from all points in the cerebrumto the diencephalon, a structure throughwhich all connections to the rest of the brainmust pass (Ross and Henneberg, 1995). An-other possible advantage of a flexed basicra-nium derives from the in vitro experimentsof Demes (1985), showing that the angula-tion of the cranial base in combination witha spherical neurocranium helps distributeapplied stresses efficiently over a large areaand decreases stresses in the anterior cra-nial base during loading of the temporoman-dibular joint. This interesting model, how-ever, requires further testing.
Whether the spheroid shape of the tel-encephalon is a functional adaptation or astructural consequence of geometry anddevelopmental processes remains to be de-termined. Nevertheless, the presence ofthe cerebellum, and ultimately of thebrain stem, prevents caudal expansion ofthe telencephalon, making rostral expan-sion of the telencephalon the easiest route.This would cause the especially large hu-man brain to develop a kink of the kindmeasured by Hofer, which in turn maycause flexion of the basicranium. If thishypothesis is correct, then some propor-tion of the variation in basicranial angleamong primates is caused by intrinsicchanges in brain shape, and not the rela-tionship between the size of the brain andthe base on which it sits.
One caution (noted above) is that ontoge-netic data suggest that the interspecific
variation in cranial base angle and shapepresented above is partially a consequenceof variables other than relative encephaliza-tion or intrinsic brain shape. Ontogeneticdata are useful because they allow one toexamine temporal relationships among pre-dicted causal factors. The human ontoge-netic data provide mixed support for thehypothesis that cranial base angulation re-flects relative encephalization. Jeffery(1999) found no significant relationship be-tween CBA1 and IRE1 during the secondfetal trimester in humans, when braingrowth is especially rapid; but Liebermanand McCarthy (1999) found that the humancranial base flexes rapidly during the first 2postnatal years, when most brain growthoccurs. Why relative brain size in humanscorrelates with cranial base angle afterbirth but not before remains to be ex-plained. In addition, and in contrast to hu-mans, the cranial base in all nonhuman pri-mates so far analyzed extends rather thanflexes during the period of postnatal braingrowth, and continues to extend throughoutthe period of facial growth, after braingrowth has ceased. In Pan, for example, ap-proximately 88% of cranial base extension(CBA1) occurs after the brain has reached95% adult size (Lieberman and McCarthy,1999). Similar results characterize othergenera (e.g., Macaca; Sirianni and Swin-dler, 1985; Schneiderman, 1992).
Ontogenetic data do not disprove the hy-pothesis that variation in cranial base angleis related to brain size, but instead highlightthe likelihood that the processes which gen-erate variation in cranial base angle arepolyphasic and multifactorial. Notably, theontogenetic data suggest that the tightstructural relationship between the faceand the anterior cranial base (discussed be-low) is also an important influence on cra-nial base angle. This suggests that a largeproportion of the interspecific variation inCBA, IRE, and other aspects of neural sizeand shape reported above is explained byinteractions between the brain and the cra-nial base prior to the end of the neuralgrowth phase. Thereafter, other factors (es-pecially those related to the face) influencethe shape of the cranial base. One obviousway to test this hypothesis is to compare the
PRIMATE CRANIAL BASE 137D.E. Lieberman et al.]

above interspecific analyses of adults withcomparable analyses of infants at the periodwhen the brain has ceased growing, but be-fore much of the face has grown.
Brain volume and posterior cranialfossa shape
Dean and Wood (1981, 1982) and Aielloand Dean (1990) hypothesized that in-creases in cerebellum size correlate with in-creases in the size of the posterior cranialfossa. This correlation is purportedly a re-sult of increases in basicranial flexion; andby lateral and anterior displacement of thelateral aspects of the petrous pyramids,which cause the petrous pyramids to bemore coronally oriented in humans than ingreat apes. However, Ross and Ravosa(1993) found little support for a link be-tween absolute cerebellum volume andCBA4; in addition, Spoor (1997) did not finda correlation between cerebellum volumeand petrous orientation. Rather, Spoor(1997) showed that more coronally orientedpetrous pyramids in adult primates corre-late better with increases in brain volumerelative to basicranial length. In addition,the petrous pyramids, when viewed fromthe internal aspect of the cranial base, arenot more coronally oriented in humans vs.other apes (Spoor, 1997).
The probable explanation for these re-sults may be that the posterior cranial base(Ba-S) scales with isometry against bothcerebellum volume and the volume of infra-tentorial neural structures (cerebellum, me-dulla oblongata, mesencephalon) (Table 4).If this scaling pattern characterizes thosedimensions of the posterior cranial fossa notin the mid-sagittal plane, then increasedinfratentorial neural volumes would not ne-cessitate changes in the proportions of theentire posterior cranial fossa. This wouldimply that variation in the orientation ofthe petrous pyramids may be linked tochanges in other cranial systems and is notsolely a structural response to changes incerebellar volume.
ASSOCIATIONS BETWEEN CRANIALBASE AND FACE
It has long been known that the cranialbase plays an important role in facial
growth, but many details of how these re-gions interact remain poorly understood.While the face has some influence on cranialbase growth (see below), there are two ma-jor reasons to believe that the cranial baseexerts a greater influence on the face thanvice versa during growth by setting up cer-tain key spatial relationships. First, the ma-jority of the cranial base (with the exceptionof the ethmoidal portions of the ethmomax-illary complex) attains adult size long beforethe face (Moore and Lavelle, 1974). Second,as noted above, most of the face grows an-teriorly, laterally, inferiorly, and around thecranial base. In all mammals, the upperportion of the face (the orbital and uppernasal regions) grows antero inferiorly rela-tive to the anterior cranial base and floor;and the middle face (mostly the nasal re-gion) grows anteriorly relative to the middlecranial fossa. The lower portion of the face(the mandibular and maxillary arches andtheir supporting structures) interacts onlyindirectly with the cranial base, since themaxillary arch grows inferiorly from themiddle face and anteriorly relative to thepterygoid processes of the sphenoid.
These spatial and developmental associa-tions raise an important question: to whatextent does the cranial base influence facialgrowth and form? In order to address thisissue, we first discuss the relationships be-tween two regions of the face and cranialbase that are contiguous across functionalor developmental boundaries (the so-calledgrowth counterparts of Enlow, 1990): 1) theanterior cranial fossa and the upper, orbital,and nasal portions of the face, and 2) themiddle cranial fossa and the middle, ethmo-maxillary portion of the face. We concludewith a brief discussion of the possible rela-tionships between cranial base shape andoverall facial shape.
Anterior cranial fossa shape and upperfacial growth
The upper face comprises the orbital cav-ities, the orbital superstructures, and theupper portion of the nasal cavity. The upperface therefore incorporates elements of theanterior cranial base, including the eth-moid, parts of the sphenoid, and significantportions of the frontal bone. The upper face
138 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000

grows away from the rest of the cranial basein three ways. Initially, as the eyeballs ex-pand, the orbital cavity expands anteriorly,inferiorly, and laterally through drift anddisplacement (Moss and Young, 1960; En-low, 1990). Animals enucleated during theperiod of eyeball growth consequently havedeficient anterior and lateral growth of theupper face (see Sarnat, 1982). In addition,since the roof of the orbit also contributes tothe floor of the anterior cranial fossa (i.e.,the orbital plates of the frontal bone andlesser wings of the sphenoid), the position,orientation, and shape of the orbital roofmust inevitably be affected by growth of thefrontal lobes and anterior cranial fossa. Fi-nally, the orbital cavities and superstruc-tures grow anteriorly and laterally awayfrom the anterior cranial fossa, but only af-ter some period of postnatal development.In humans, for example, the upper face doesnot emerge from under the anterior cranialbase until after the eruption of the secondmolars (Riolo et al., 1974; Lieberman, 2000).In contrast, the front of the upper face inmost nonhuman primates projects anteri-orly relative to the front of the anterior cra-nial fossa prior to the eruption of the firstmolars (Krogman, 1969; Lieberman, 1998,2000). Variation in this spatial separation,termed neurocranial-orbital or neuro-or-bital disjunction, has been analyzed by var-ious researchers to explain ontogenetic andinterspecific patterns of supraorbital torusmorphology (Weidenreich, 1941; Moss andYoung, 1960; Radinsky, 1968, 1970, 1977,1979; Shea, 1985a, 1986, 1988; Ravosa,1988, 1991a,b; Hylander and Ravosa, 1992;Vinyard and Smith, 1997; Lieberman, 1998,2000; Ravosa et al., 2000b).
These developmental and architecturalrelationships between the anterior cranialbase and upper face have several conse-quences for interspecific variation in pri-mates. First, the orientation of the orbitsand the upper face is intimately related tothe orientation of the anterior cranial basein haplorhines (Ravosa, 1991a,b; Ross andRavosa, 1993; Ross and Henneberg, 1995),confirming the hypothesis of Dabelow (1929,1931) that the orientation of the orbits andthe anterior cranial base should be tightlycorrelated in animals like haplorhines,
whose orbits are closely approximated be-low the olfactory tract (see also Cartmill,1970, 1972; Ravosa et al., 2000a,2000b).Several recent studies have found supportfor this hypothesis. First, brow-ridge size inanthropoids (but not strepsirhines) is highlycorrelated with variation in the position ofthe orbits relative to the anterior cranialbase (Ravosa, 1988, 1991a,b; Hylander andRavosa, 1992; Vinyard and Smith, 1997;Lieberman, 2000; Ravosa et al., 2000b). Fi-nally, there is evidence that the upper facetends to be rotated dorsally relative to theposterior cranial base in primates with ahighly extended cranial base, but more ven-trally rotated relative to the posterior cra-nial base in primates such as humans andbonobos with a more flexed cranial base (De-lattre and Fenart, 1956; Moss and Young,1960; Heintz, 1966; Cramer, 1977; Shea,1985a, 1986, 1988; Ravosa, 1988, 1991a,b;Lieberman, 2000).
While the structural boundaries sharedby the anterior cranial base and the upperface result in a high degree of integrationbetween these two regions, the extent towhich anterior cranial base shape influ-ences other aspects of facial shape is lessclear. In an interspecific study of 68 species,Ross and Ravosa (1993) found that both or-bit and palate orientation are significantlycorrelated with anterior cranial base orien-tation, but that palate orientation accountsfor none of the variation in cranial baseangle independent of orbital axis orienta-tion. McCarthy and Lieberman (2001) havealso shown that the orientation of the orbitsand the anterior cranial base are correlatedwith each other (r 5 0.617, P , 0.05) inhaplorhines but not in strepsirhines. Thesestudies suggest that the orientation of theanterior cranial base affects the orientationof the upper face directly, but that it onlyindirectly influences palate orientationthrough the integration of palate and orbits(see also Ravosa, 1988; Ravosa and Shea,1994). Consequently, it seems likely thatthe anterior cranial base exerts a slight in-fluence on facial orientation as a whole, butthat only the orbital region of the face isdirectly integrated with the anterior cranialbase. Higher levels of integration appear tocharacterize those organisms with greater
PRIMATE CRANIAL BASE 139D.E. Lieberman et al.]
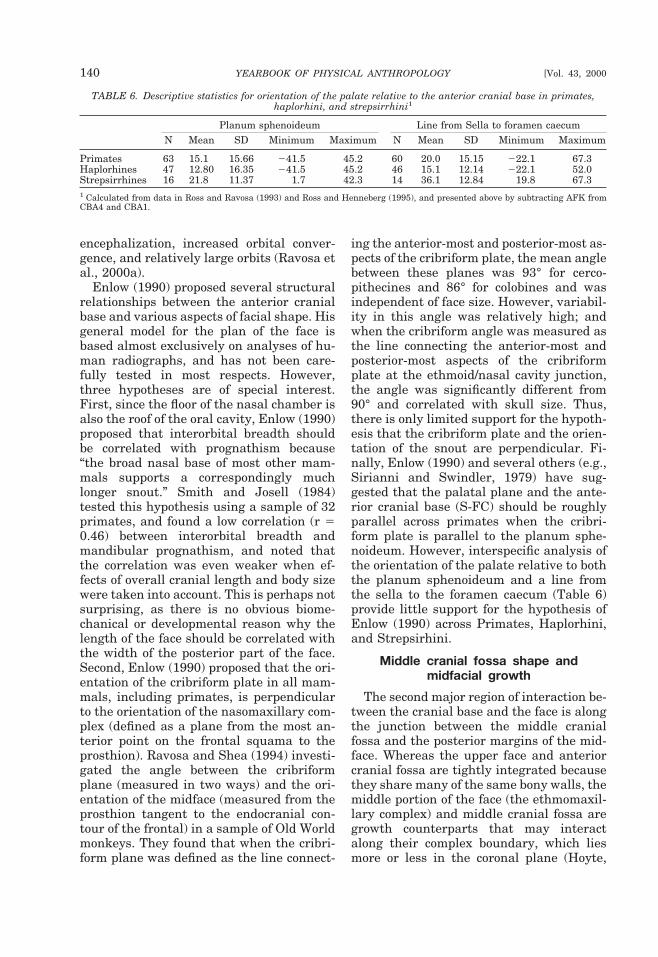
encephalization, increased orbital conver-gence, and relatively large orbits (Ravosa etal., 2000a).
Enlow (1990) proposed several structuralrelationships between the anterior cranialbase and various aspects of facial shape. Hisgeneral model for the plan of the face isbased almost exclusively on analyses of hu-man radiographs, and has not been care-fully tested in most respects. However,three hypotheses are of special interest.First, since the floor of the nasal chamber isalso the roof of the oral cavity, Enlow (1990)proposed that interorbital breadth shouldbe correlated with prognathism because“the broad nasal base of most other mam-mals supports a correspondingly muchlonger snout.” Smith and Josell (1984)tested this hypothesis using a sample of 32primates, and found a low correlation (r 50.46) between interorbital breadth andmandibular prognathism, and noted thatthe correlation was even weaker when ef-fects of overall cranial length and body sizewere taken into account. This is perhaps notsurprising, as there is no obvious biome-chanical or developmental reason why thelength of the face should be correlated withthe width of the posterior part of the face.Second, Enlow (1990) proposed that the ori-entation of the cribriform plate in all mam-mals, including primates, is perpendicularto the orientation of the nasomaxillary com-plex (defined as a plane from the most an-terior point on the frontal squama to theprosthion). Ravosa and Shea (1994) investi-gated the angle between the cribriformplane (measured in two ways) and the ori-entation of the midface (measured from theprosthion tangent to the endocranial con-tour of the frontal) in a sample of Old Worldmonkeys. They found that when the cribri-form plane was defined as the line connect-
ing the anterior-most and posterior-most as-pects of the cribriform plate, the mean anglebetween these planes was 93° for cerco-pithecines and 86° for colobines and wasindependent of face size. However, variabil-ity in this angle was relatively high; andwhen the cribriform angle was measured asthe line connecting the anterior-most andposterior-most aspects of the cribriformplate at the ethmoid/nasal cavity junction,the angle was significantly different from90° and correlated with skull size. Thus,there is only limited support for the hypoth-esis that the cribriform plate and the orien-tation of the snout are perpendicular. Fi-nally, Enlow (1990) and several others (e.g.,Sirianni and Swindler, 1979) have sug-gested that the palatal plane and the ante-rior cranial base (S-FC) should be roughlyparallel across primates when the cribri-form plate is parallel to the planum sphe-noideum. However, interspecific analysis ofthe orientation of the palate relative to boththe planum sphenoideum and a line fromthe sella to the foramen caecum (Table 6)provide little support for the hypothesis ofEnlow (1990) across Primates, Haplorhini,and Strepsirhini.
Middle cranial fossa shape andmidfacial growth
The second major region of interaction be-tween the cranial base and the face is alongthe junction between the middle cranialfossa and the posterior margins of the mid-face. Whereas the upper face and anteriorcranial fossa are tightly integrated becausethey share many of the same bony walls, themiddle portion of the face (the ethmomaxil-lary complex) and middle cranial fossa aregrowth counterparts that may interactalong their complex boundary, which liesmore or less in the coronal plane (Hoyte,
TABLE 6. Descriptive statistics for orientation of the palate relative to the anterior cranial base in primates,haplorhini, and strepsirrhini1
Planum sphenoideum Line from Sella to foramen caecumN Mean SD Minimum Maximum N Mean SD Minimum Maximum
Primates 63 15.1 15.66 241.5 45.2 60 20.0 15.15 222.1 67.3Haplorhines 47 12.80 16.35 241.5 45.2 46 15.1 12.14 222.1 52.0Strepsirrhines 16 21.8 11.37 1.7 42.3 14 36.1 12.84 19.8 67.31 Calculated from data in Ross and Ravosa (1993) and Ross and Henneberg (1995), and presented above by subtracting AFK fromCBA4 and CBA1.
140 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000
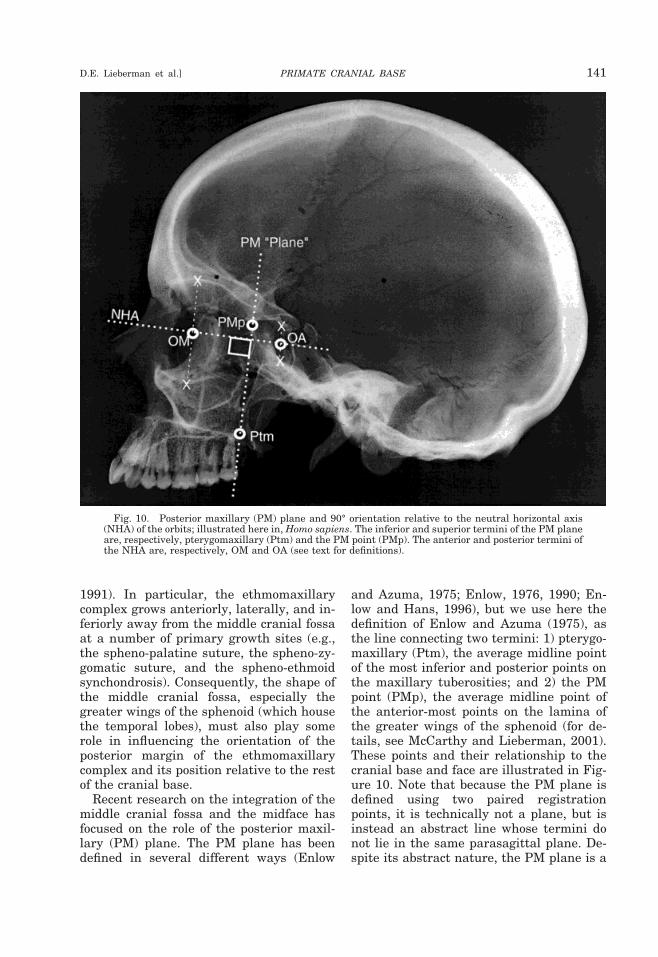
1991). In particular, the ethmomaxillarycomplex grows anteriorly, laterally, and in-feriorly away from the middle cranial fossaat a number of primary growth sites (e.g.,the spheno-palatine suture, the spheno-zy-gomatic suture, and the spheno-ethmoidsynchondrosis). Consequently, the shape ofthe middle cranial fossa, especially thegreater wings of the sphenoid (which housethe temporal lobes), must also play somerole in influencing the orientation of theposterior margin of the ethmomaxillarycomplex and its position relative to the restof the cranial base.
Recent research on the integration of themiddle cranial fossa and the midface hasfocused on the role of the posterior maxil-lary (PM) plane. The PM plane has beendefined in several different ways (Enlow
and Azuma, 1975; Enlow, 1976, 1990; En-low and Hans, 1996), but we use here thedefinition of Enlow and Azuma (1975), asthe line connecting two termini: 1) pterygo-maxillary (Ptm), the average midline pointof the most inferior and posterior points onthe maxillary tuberosities; and 2) the PMpoint (PMp), the average midline point ofthe anterior-most points on the lamina ofthe greater wings of the sphenoid (for de-tails, see McCarthy and Lieberman, 2001).These points and their relationship to thecranial base and face are illustrated in Fig-ure 10. Note that because the PM plane isdefined using two paired registrationpoints, it is technically not a plane, but isinstead an abstract line whose termini donot lie in the same parasagittal plane. De-spite its abstract nature, the PM plane is a
Fig. 10. Posterior maxillary (PM) plane and 90° orientation relative to the neutral horizontal axis(NHA) of the orbits; illustrated here in, Homo sapiens. The inferior and superior termini of the PM planeare, respectively, pterygomaxillary (Ptm) and the PM point (PMp). The anterior and posterior termini ofthe NHA are, respectively, OM and OA (see text for definitions).
PRIMATE CRANIAL BASE 141D.E. Lieberman et al.]
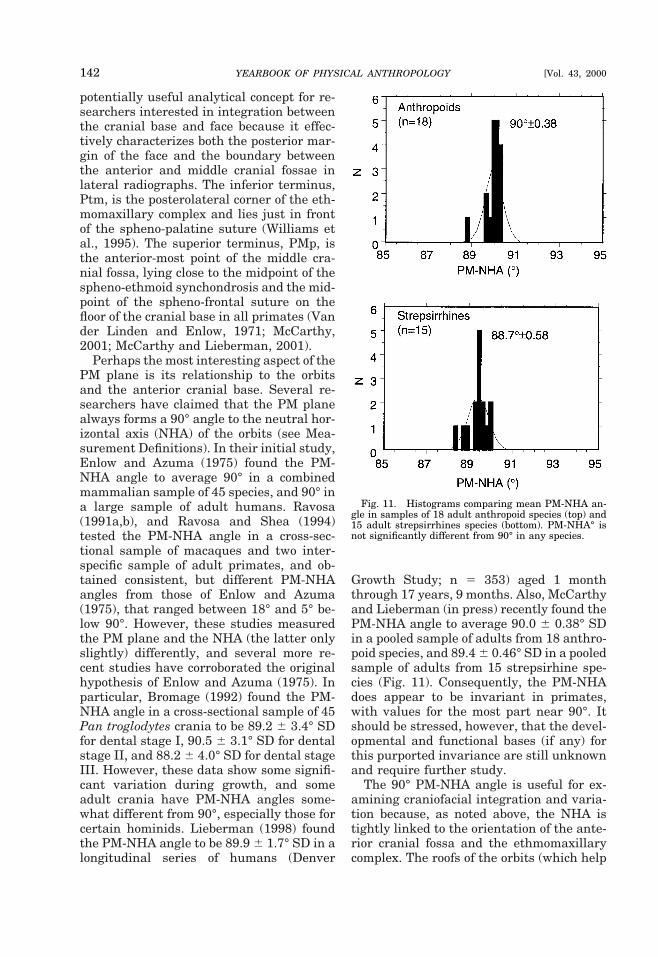
potentially useful analytical concept for re-searchers interested in integration betweenthe cranial base and face because it effec-tively characterizes both the posterior mar-gin of the face and the boundary betweenthe anterior and middle cranial fossae inlateral radiographs. The inferior terminus,Ptm, is the posterolateral corner of the eth-momaxillary complex and lies just in frontof the spheno-palatine suture (Williams etal., 1995). The superior terminus, PMp, isthe anterior-most point of the middle cra-nial fossa, lying close to the midpoint of thespheno-ethmoid synchondrosis and the mid-point of the spheno-frontal suture on thefloor of the cranial base in all primates (Vander Linden and Enlow, 1971; McCarthy,2001; McCarthy and Lieberman, 2001).
Perhaps the most interesting aspect of thePM plane is its relationship to the orbitsand the anterior cranial base. Several re-searchers have claimed that the PM planealways forms a 90° angle to the neutral hor-izontal axis (NHA) of the orbits (see Mea-surement Definitions). In their initial study,Enlow and Azuma (1975) found the PM-NHA angle to average 90° in a combinedmammalian sample of 45 species, and 90° ina large sample of adult humans. Ravosa(1991a,b), and Ravosa and Shea (1994)tested the PM-NHA angle in a cross-sec-tional sample of macaques and two inter-specific sample of adult primates, and ob-tained consistent, but different PM-NHAangles from those of Enlow and Azuma(1975), that ranged between 18° and 5° be-low 90°. However, these studies measuredthe PM plane and the NHA (the latter onlyslightly) differently, and several more re-cent studies have corroborated the originalhypothesis of Enlow and Azuma (1975). Inparticular, Bromage (1992) found the PM-NHA angle in a cross-sectional sample of 45Pan troglodytes crania to be 89.2 6 3.4° SDfor dental stage I, 90.5 6 3.1° SD for dentalstage II, and 88.2 6 4.0° SD for dental stageIII. However, these data show some signifi-cant variation during growth, and someadult crania have PM-NHA angles some-what different from 90°, especially those forcertain hominids. Lieberman (1998) foundthe PM-NHA angle to be 89.9 6 1.7° SD in alongitudinal series of humans (Denver
Growth Study; n 5 353) aged 1 monththrough 17 years, 9 months. Also, McCarthyand Lieberman (in press) recently found thePM-NHA angle to average 90.0 6 0.38° SDin a pooled sample of adults from 18 anthro-poid species, and 89.4 6 0.46° SD in a pooledsample of adults from 15 strepsirhine spe-cies (Fig. 11). Consequently, the PM-NHAdoes appear to be invariant in primates,with values for the most part near 90°. Itshould be stressed, however, that the devel-opmental and functional bases (if any) forthis purported invariance are still unknownand require further study.
The 90° PM-NHA angle is useful for ex-amining craniofacial integration and varia-tion because, as noted above, the NHA istightly linked to the orientation of the ante-rior cranial fossa and the ethmomaxillarycomplex. The roofs of the orbits (which help
Fig. 11. Histograms comparing mean PM-NHA an-gle in samples of 18 adult anthropoid species (top) and15 adult strepsirrhines species (bottom). PM-NHA° isnot significantly different from 90° in any species.
142 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000

define the NHA) comprise much of the floorof the anterior cranial fossa. Therefore, itfollows that the PM plane and the anteriorcranial base should also form an approxi-mately 90° angle in primates whose orbitsare approximated to the midline. This hy-pothesis was tested by McCarthy andLieberman (2001), who found that the anglebetween the PM plane and the planumsphenoideum averaged 95.2 6 7.6° SD (n 518) in anthropoids and 82.8 6 9.5° SD (n 514) in strepsirhines. McCarthy and Lieber-man (2001) also found that the angle be-tween the PM plane and the midline ante-rior cranial base (from the sella to theforamen caecum) averages 89.2 6 9.97° SD(n 5 18) in anthropoids, but 70.6 6 10.5° SD(n 5 15) in strepsirrhines. The high stan-dard deviations of these angles indicate thatthe integration between the back of the faceand the anterior cranial base is not verystrong. Ross and Ravosa (1993) also came tosimilar conclusions by comparing the orbitalaxis orientation relative to the posterior cra-nial base, against the orientation of the pla-num sphenoideum relative to the posteriorcranial base. The differences between an-thropoids and strepsirhines in the relation-ship of the orbits to the cranial base can beexplained by the fact that the roof of theorbits does not contribute to the midlinecranial base in strepsirhines, and becausethe cribriform plate tends to be orientedmore vertically relative to the planum sphe-noideum in strepsirrhines than in anthro-poids (Cartmill, 1970).
The potential integration of the middleand anterior cranial fossae with the face (asmeasured via the PM plane) and the ante-rior cranial base raises several interestingissues. Most importantly, the top and backof the face appear to form an integratedunit, the “facial block” which rotates duringontogeny around an axis through the inter-section of the anterior and middle cranialfossae at the front of the greater wings ofthe sphenoid (McCarthy and Lieberman,2001). This facial block is characteristic ofanthropoids but not strepsirhines, and man-ifests itself through correlations betweencranial base angle and upper facial orienta-tion in primates (Weidenriech, 1941; Mossand Young, 1960; Biegert, 1963; Shea,
1985a, 1986, 1988; Ravosa, 1988, 1991a,b;Ross and Ravosa, 1993; Ross, 1995a,b; Mayand Sheffer, 1999; Lieberman, 2000; Ravosaet al., 2000a, 2000b). In particular, as theanterior cranial base flexes relative to theposterior cranial base, the PM plane alsomust flex relative to the posterior cranialbase, rotating the posterior and upper por-tions of the face underneath the anteriorcranial fossa (klinorhynchy). In contrast, ex-tension of the anterior cranial base relativeto the posterior cranial base will rotate theposterior and upper portions of the face dor-sally relative to the posterior cranial base(airorhynchy) (Fig. 12).
The relationship of the orientation of theback of the face (as measured for example bythe PM plane) to the anterior cranial basealso influences nasopharynx shape. As Fig-ure 12 shows, flexion of the anterior cranialbase and/or face relative to the posteriorcranial base not only rotates the face underthe anterior cranial fossa, but it also short-ens (absolutely and relatively) the length ofthe pharyngeal space between the back ofthe palate and the front of the vertebralcolumn (Laitman and Heimbuch, 1982;Spoor et al., 1999; McCarthy and Lieber-man, 2001). While flexion of the cranial baseduring ontogeny is completely independentof the descent of the hyoid and larynx(Lieberman and McCarthy, 1999), variationin cranial base angle does influence someaspects of pharyngeal shape (Laitman andHeimbuch, 1982; see below).
Ross and Henneberg (1995) suggestedthat there must be functional constraints onhow far back the hard palate can be posi-tioned without occluding the airway. Theintegration of the anterior cranial base withthe upper and posterior margins of the facemeans that these constraints on pharynxposition might determine the maximumpossible degree of basicranial angle, partic-ularly in genera such as Pongo and Alouattawith relatively large pharyngeal structures(Biegert, 1957, 1963). Ross and Henneberg(1995) suggested that hominoids mighthave found a way to circumvent these “con-straints.” Hominoids have more airorhynch(dorsally rotated and less frontated) orbitsand palates than nonhominoid primateswith comparably flexed basicrania (Shea,
PRIMATE CRANIAL BASE 143D.E. Lieberman et al.]

1985a, 1988; Ross and Henneberg, 1995).This airorhynchy has yet to be explaineddevelopmentally and functionally; however,Ross and Henneberg (1995) suggested thatit evolved in hominoids in response to in-creased flexion of the cranial base producingposterior displacement of the palate.
More research is needed on the integra-tion of the midface and cranial base. In par-ticular, why is the PM plane oriented at 90°relative to the NHA during postnatal ontog-eny and thus across taxa? Another questionof interest is, what aspects of cranial baseand facial shape are responsible for most ofthe variation in PM plane position, andhence facial orientation? This problem hasnot been well studied, but the orientation ofthe PM plane is probably most affected bythe size of the middle cranial fossa, espe-cially the length of the temporal lobes, byflexion of the sphenoid, and by the length ofthe anterior sphenoid in the midline cranialbase. Three-dimensional studies of the in-terface between the PM plane and the cra-nial base are needed to resolve these andother questions about cranial base-face in-terrelations and interactions.
Basicranial width and overall facialshape in humans
Although it is clear that the cranial baseplays a major role in influencing facial ori-
entation relative to the neurocranium, thereis less information about the potential influ-ence of the cranial base on other aspects offacial shape such as height, length, andwidth. To what extent is overall facial shapeindependent of the cranial base? It is com-monly assumed that the majority of facialgrowth is independent of cranial basegrowth, largely because much of the facegrows in a skeletal growth trajectory afterthe end of the neural growth phase. In hu-mans, for example, the face attains 95%adult size by 16–18 years, at least 10 yearsafter the cranial base reaches adult size(Stamrud, 1959; Moore and Lavelle, 1974).In addition, most facial and basicranial di-mensions appear to be genetically indepen-dent in adults (Cheverud, 1996). However,there is some evidence to suggest thatchanges in the proportions of the cranialbase can influence facial shape. This inter-action is predicted to be especially impor-tant, and perhaps exclusive to humans, inwhich the upper face lies almost completelyunderneath the anterior cranial fossa (Wei-denreich, 1941; Howells, 1973; Enlow andBhatt, 1984; Enlow, 1990; Lieberman et al.,2000).
The most explicit of these hypotheses isthat of Enlow (1990), who suggested thathumans with absolutely narrow cranialbases (primarily dolichocephalics) tend to
Fig. 12. Proposed “facial block,” showing the effects of angular invariance between the back of the face(summarized by the PM plane) and the top of the face, which is also the bottom of the anterior cranialbase (S-FC). Changes in cranial base angle cause the top and back of the face to rotate together aroundan imaginary axis through the PM point. See text for details.
144 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000
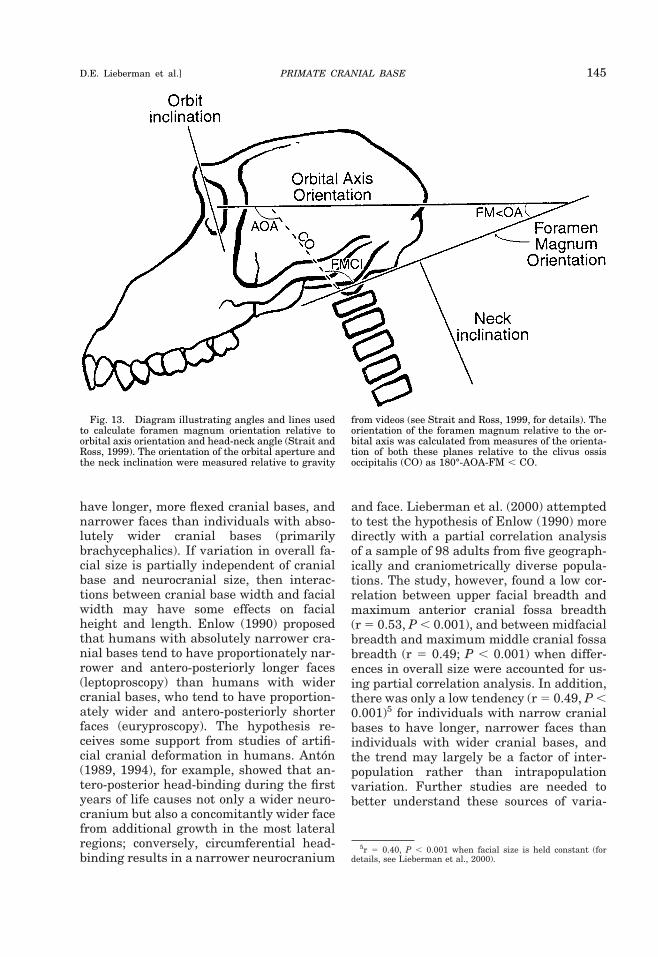
have longer, more flexed cranial bases, andnarrower faces than individuals with abso-lutely wider cranial bases (primarilybrachycephalics). If variation in overall fa-cial size is partially independent of cranialbase and neurocranial size, then interac-tions between cranial base width and facialwidth may have some effects on facialheight and length. Enlow (1990) proposedthat humans with absolutely narrower cra-nial bases tend to have proportionately nar-rower and antero-posteriorly longer faces(leptoproscopy) than humans with widercranial bases, who tend to have proportion-ately wider and antero-posteriorly shorterfaces (euryproscopy). The hypothesis re-ceives some support from studies of artifi-cial cranial deformation in humans. Anton(1989, 1994), for example, showed that an-tero-posterior head-binding during the firstyears of life causes not only a wider neuro-cranium but also a concomitantly wider facefrom additional growth in the most lateralregions; conversely, circumferential head-binding results in a narrower neurocranium
and face. Lieberman et al. (2000) attemptedto test the hypothesis of Enlow (1990) moredirectly with a partial correlation analysisof a sample of 98 adults from five geograph-ically and craniometrically diverse popula-tions. The study, however, found a low cor-relation between upper facial breadth andmaximum anterior cranial fossa breadth(r 5 0.53, P , 0.001), and between midfacialbreadth and maximum middle cranial fossabreadth (r 5 0.49; P , 0.001) when differ-ences in overall size were accounted for us-ing partial correlation analysis. In addition,there was only a low tendency (r 5 0.49, P ,0.001)5 for individuals with narrow cranialbases to have longer, narrower faces thanindividuals with wider cranial bases, andthe trend may largely be a factor of inter-population rather than intrapopulationvariation. Further studies are needed tobetter understand these sources of varia-
5r 5 0.40, P , 0.001 when facial size is held constant (fordetails, see Lieberman et al., 2000).
Fig. 13. Diagram illustrating angles and lines usedto calculate foramen magnum orientation relative toorbital axis orientation and head-neck angle (Strait andRoss, 1999). The orientation of the orbital aperture andthe neck inclination were measured relative to gravity
from videos (see Strait and Ross, 1999, for details). Theorientation of the foramen magnum relative to the or-bital axis was calculated from measures of the orienta-tion of both these planes relative to the clivus ossisoccipitalis (CO) as 180°-AOA-FM , CO.
PRIMATE CRANIAL BASE 145D.E. Lieberman et al.]
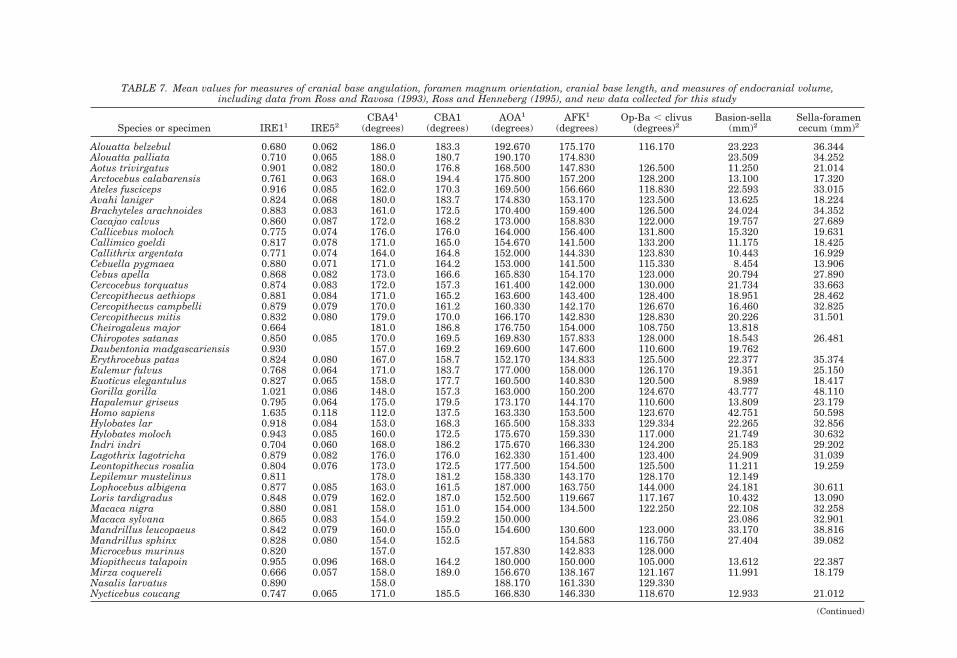
TABLE 7. Mean values for measures of cranial base angulation, foramen magnum orientation, cranial base length, and measures of endocranial volume,including data from Ross and Ravosa (1993), Ross and Henneberg (1995), and new data collected for this study
Species or specimen IRE11 IRE52CBA41
(degrees)CBA1
(degrees)AOA1
(degrees)AFK1
(degrees)Op-Ba , clivus
(degrees)2Basion-sella
(mm)2Sella-foramencecum (mm)2
Alouatta belzebul 0.680 0.062 186.0 183.3 192.670 175.170 116.170 23.223 36.344Alouatta palliata 0.710 0.065 188.0 180.7 190.170 174.830 23.509 34.252Aotus trivirgatus 0.901 0.082 180.0 176.8 168.500 147.830 126.500 11.250 21.014Arctocebus calabarensis 0.761 0.063 168.0 194.4 175.800 157.200 128.200 13.100 17.320Ateles fusciceps 0.916 0.085 162.0 170.3 169.500 156.660 118.830 22.593 33.015Avahi laniger 0.824 0.068 180.0 183.7 174.830 153.170 123.500 13.625 18.224Brachyteles arachnoides 0.883 0.083 161.0 172.5 170.400 159.400 126.500 24.024 34.352Cacajao calvus 0.860 0.087 172.0 168.2 173.000 158.830 122.000 19.757 27.689Callicebus moloch 0.775 0.074 176.0 176.0 164.000 156.400 131.800 15.320 19.631Callimico goeldi 0.817 0.078 171.0 165.0 154.670 141.500 133.200 11.175 18.425Callithrix argentata 0.771 0.074 164.0 164.8 152.000 144.330 123.830 10.443 16.929Cebuella pygmaea 0.880 0.071 171.0 164.2 153.000 141.500 115.330 8.454 13.906Cebus apella 0.868 0.082 173.0 166.6 165.830 154.170 123.000 20.794 27.890Cercocebus torquatus 0.874 0.083 172.0 157.3 161.400 142.000 130.000 21.734 33.663Cercopithecus aethiops 0.881 0.084 171.0 165.2 163.600 143.400 128.400 18.951 28.462Cercopithecus campbelli 0.879 0.079 170.0 161.2 160.330 142.170 126.670 16.460 32.825Cercopithecus mitis 0.832 0.080 179.0 170.0 166.170 142.830 128.830 20.226 31.501Cheirogaleus major 0.664 181.0 186.8 176.750 154.000 108.750 13.818Chiropotes satanas 0.850 0.085 170.0 169.5 169.830 157.833 128.000 18.543 26.481Daubentonia madgascariensis 0.930 157.0 169.2 169.600 147.600 110.600 19.762Erythrocebus patas 0.824 0.080 167.0 158.7 152.170 134.833 125.500 22.377 35.374Eulemur fulvus 0.768 0.064 171.0 183.7 177.000 158.000 126.170 19.351 25.150Euoticus elegantulus 0.827 0.065 158.0 177.7 160.500 140.830 120.500 8.989 18.417Gorilla gorilla 1.021 0.086 148.0 157.3 163.000 150.200 124.670 43.777 48.110Hapalemur griseus 0.795 0.064 175.0 179.5 173.170 144.170 110.600 13.809 23.179Homo sapiens 1.635 0.118 112.0 137.5 163.330 153.500 123.670 42.751 50.598Hylobates lar 0.918 0.084 153.0 168.3 165.500 158.333 129.334 22.265 32.856Hylobates moloch 0.943 0.085 160.0 172.5 175.670 159.330 117.000 21.749 30.632Indri indri 0.704 0.060 168.0 186.2 175.670 166.330 124.200 25.183 29.202Lagothrix lagotricha 0.879 0.082 176.0 176.0 162.330 151.400 123.400 24.909 31.039Leontopithecus rosalia 0.804 0.076 173.0 172.5 177.500 154.500 125.500 11.211 19.259Lepilemur mustelinus 0.811 178.0 181.2 158.330 143.170 128.170 12.149Lophocebus albigena 0.877 0.085 163.0 161.5 187.000 163.750 144.000 24.181 30.611Loris tardigradus 0.848 0.079 162.0 187.0 152.500 119.667 117.167 10.432 13.090Macaca nigra 0.880 0.081 158.0 151.0 154.000 134.500 122.250 22.108 32.258Macaca sylvana 0.865 0.083 154.0 159.2 150.000 23.086 32.901Mandrillus leucopaeus 0.842 0.079 160.0 155.0 154.600 130.600 123.000 33.170 38.816Mandrillus sphinx 0.828 0.080 154.0 152.5 154.583 116.750 27.404 39.082Microcebus murinus 0.820 157.0 157.830 142.833 128.000Miopithecus talapoin 0.955 0.096 168.0 164.2 180.000 150.000 105.000 13.612 22.387Mirza coquereli 0.666 0.057 158.0 189.0 156.670 138.167 121.167 11.991 18.179Nasalis larvatus 0.890 158.0 188.170 161.330 129.330Nycticebus coucang 0.747 0.065 171.0 185.5 166.830 146.330 118.670 12.933 21.012
(Continued)
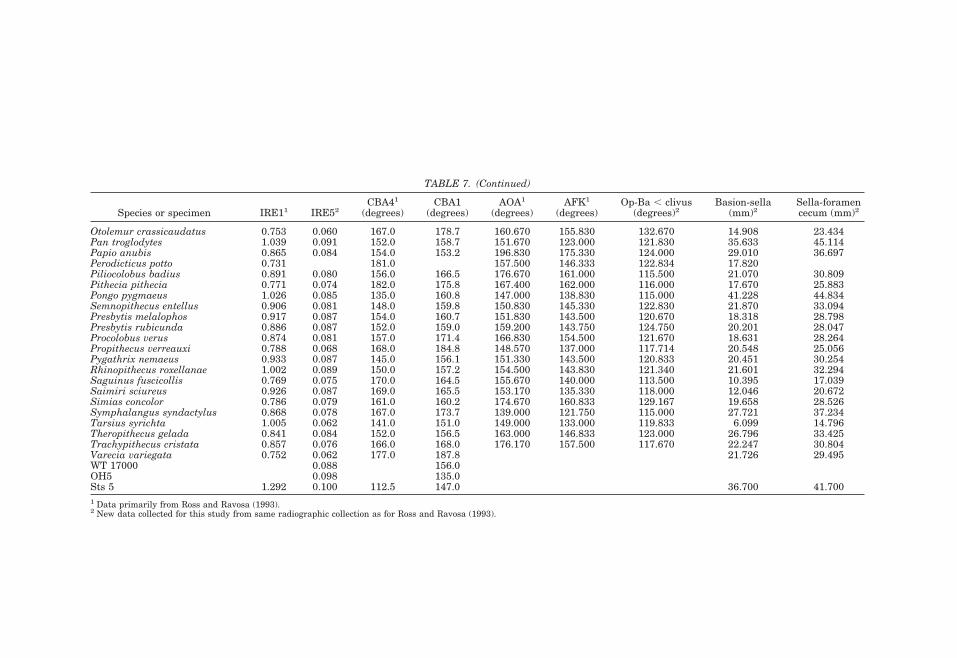
TABLE 7. (Continued)
Species or specimen IRE11 IRE52CBA41
(degrees)CBA1
(degrees)AOA1
(degrees)AFK1
(degrees)Op-Ba , clivus
(degrees)2Basion-sella
(mm)2Sella-foramencecum (mm)2
Otolemur crassicaudatus 0.753 0.060 167.0 178.7 160.670 155.830 132.670 14.908 23.434Pan troglodytes 1.039 0.091 152.0 158.7 151.670 123.000 121.830 35.633 45.114Papio anubis 0.865 0.084 154.0 153.2 196.830 175.330 124.000 29.010 36.697Perodicticus potto 0.731 181.0 157.500 146.333 122.834 17.820Piliocolobus badius 0.891 0.080 156.0 166.5 176.670 161.000 115.500 21.070 30.809Pithecia pithecia 0.771 0.074 182.0 175.8 167.400 162.000 116.000 17.670 25.883Pongo pygmaeus 1.026 0.085 135.0 160.8 147.000 138.830 115.000 41.228 44.834Semnopithecus entellus 0.906 0.081 148.0 159.8 150.830 145.330 122.830 21.870 33.094Presbytis melalophos 0.917 0.087 154.0 160.7 151.830 143.500 120.670 18.318 28.798Presbytis rubicunda 0.886 0.087 152.0 159.0 159.200 143.750 124.750 20.201 28.047Procolobus verus 0.874 0.081 157.0 171.4 166.830 154.500 121.670 18.631 28.264Propithecus verreauxi 0.788 0.068 168.0 184.8 148.570 137.000 117.714 20.548 25.056Pygathrix nemaeus 0.933 0.087 145.0 156.1 151.330 143.500 120.833 20.451 30.254Rhinopithecus roxellanae 1.002 0.089 150.0 157.2 154.500 143.830 121.340 21.601 32.294Saguinus fuscicollis 0.769 0.075 170.0 164.5 155.670 140.000 113.500 10.395 17.039Saimiri sciureus 0.926 0.087 169.0 165.5 153.170 135.330 118.000 12.046 20.672Simias concolor 0.786 0.079 161.0 160.2 174.670 160.833 129.167 19.658 28.526Symphalangus syndactylus 0.868 0.078 167.0 173.7 139.000 121.750 115.000 27.721 37.234Tarsius syrichta 1.005 0.062 141.0 151.0 149.000 133.000 119.833 6.099 14.796Theropithecus gelada 0.841 0.084 152.0 156.5 163.000 146.833 123.000 26.796 33.425Trachypithecus cristata 0.857 0.076 166.0 168.0 176.170 157.500 117.670 22.247 30.804Varecia variegata 0.752 0.062 177.0 187.8 21.726 29.495WT 17000 0.088 156.0OH5 0.098 135.0Sts 5 1.292 0.100 112.5 147.0 36.700 41.7001 Data primarily from Ross and Ravosa (1993).2 New data collected for this study from same radiographic collection as for Ross and Ravosa (1993).

tion, and also to study potential interactionsbetween cranial base shape and facial shapein primates and other mammals.
THE CRANIAL BASE AND POSTURE
Apart from a few recent studies reviewedhere, little is known about the relationshipbetween basicranial morphology, head andneck posture, and other aspects of headmorphology related to locomotion. One per-sistent hypothesis that is especially rele-vant to hominin evolution is that flexion ofthe cranial base is an adaptation for ortho-grade posture in hominins because it causesthe foramen magnum to have a relativelymore anterior position and ventral orienta-tion (Bolk, 1909, 1910; Duckworth, 1915;Weidenreich, 1941; Schultz, 1942, 1955;Ashton and Zuckerman, 1952, 1956; Ash-ton, 1952; Moore et al., 1973; Adams andMoore, 1975; DuBrul, 1977, 1979; Dean andWood, 1981, 1982). Indeed, these featuresare often invoked in attempts to reconstructhead posture in fossil hominins (e.g., Whiteet al., 1994). All primates have the center ofmass of the head located anterior to theoccipital condyles, such that more anteriorlypositioned occipital condyles relative tohead length reduce the lever arm betweenthe center of mass and the atlanto-occipitaljoint. This balancing, in turn, reduces torqueabout this joint, thereby reducing the mag-nitude of the force required from the nuchalmuscles to hold up the head (Schultz, 1942).The occipital condyles can be moved ros-trally relative to overall head length by flex-ing the basicranium and/or shortening theposterior cranial base. In vitro experimentsby Demes (1985) demonstrated that themore ventral orientation of rostrally placedoccipital condyles orients the articular sur-faces closer to perpendicular to the compres-sive force acting through the center of massof the head, potentially reducing shearingforces acting across the joint that need to beresisted by muscles or ligaments.
There are some experimental data thatrodents forced to walk bipedally (Moss,1961; Fenart, 1966; Riesenfeld, 1966) de-velop more flexed cranial bases. However,the hypothesis that variations in cranialbase angle are adaptations for head posture,however, is not well supported by compara-
tive data. Among primates, basicranial flex-ion has been shown to be uncorrelated witheither qualitative estimates of body posture(Ross and Ravosa, 1993) or quantitativemeasures of head and neck orientation(Strait and Ross, 1999). The partial correla-tion analysis of Strait and Ross (1999) con-firmed relative brain size as a more impor-tant determinant of variation in basicranialangle, even when facial orientation andhead and neck posture were taken into ac-count. Although foramen magnum orienta-tion relative to anterior cranial base orien-tation (S-FC) has been shown to be relatedto relative brain size (Biegert, 1963; Spoor,1997), the relationship between head andneck posture and foramen magnum orienta-tion has not yet been evaluated.
We gathered data on foramen magnumorientation (FM) from the same radiographsused by Ross and Ravosa (1993) and Ravosa(1991b), and combined these data with mea-sures of hominids reported by Spoor (1993)and with measures of head and neck orien-tation reported by Strait and Ross (1999)(Fig. 13). The head-neck angle is the anglebetween neck inclination and orbit inclina-tion, both relative to the substrate (Straitand Ross, 1999). The values for FM orienta-tion relative to the clivus (FM , CO) are inTable 7 and are summarized in Figure 14.The values for FM orientation relative tothe orbital axis were calculated from mea-sures of the orientation of both these planesrelative to the clivus ossis occipitalis (CO) as180°-AOA-FM , CO. These data show thatFM , CO is not significantly correlatedwith basicranial flexion, orbital axis orien-tation, the orientation of the head relativeto the neck, or the size of the cerebellumrelative to the posterior basicranium (Table8). Nor is foramen magnum orientation rel-ative to the orbital axis (FM , AOA) corre-lated with any of these variables, exceptAOA (Table 8). Of particular interest is thelack of correlation between the head-neckangle of Strait and Ross (1999) (Fig. 14),suggesting that foramen magnum orienta-tion is not a good indicator of the orientationof the neck during habitual locomotion. Thiscalls into question attempts to estimatehead and neck posture from data on fora-men magnum orientation in fossils.
148 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000
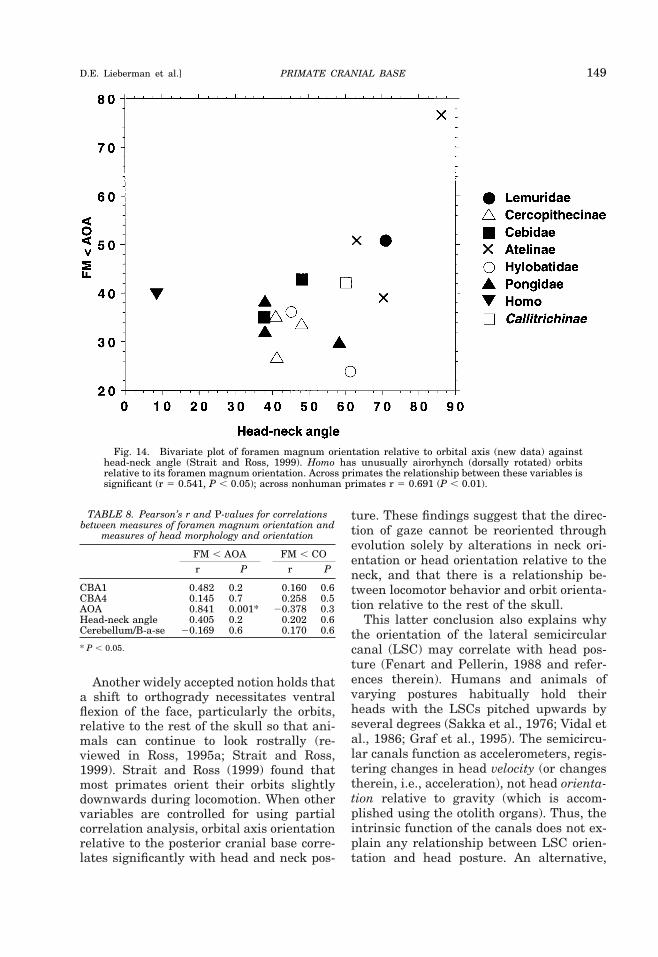
Another widely accepted notion holds thata shift to orthogrady necessitates ventralflexion of the face, particularly the orbits,relative to the rest of the skull so that ani-mals can continue to look rostrally (re-viewed in Ross, 1995a; Strait and Ross,1999). Strait and Ross (1999) found thatmost primates orient their orbits slightlydownwards during locomotion. When othervariables are controlled for using partialcorrelation analysis, orbital axis orientationrelative to the posterior cranial base corre-lates significantly with head and neck pos-
ture. These findings suggest that the direc-tion of gaze cannot be reoriented throughevolution solely by alterations in neck ori-entation or head orientation relative to theneck, and that there is a relationship be-tween locomotor behavior and orbit orienta-tion relative to the rest of the skull.
This latter conclusion also explains whythe orientation of the lateral semicircularcanal (LSC) may correlate with head pos-ture (Fenart and Pellerin, 1988 and refer-ences therein). Humans and animals ofvarying postures habitually hold theirheads with the LSCs pitched upwards byseveral degrees (Sakka et al., 1976; Vidal etal., 1986; Graf et al., 1995). The semicircu-lar canals function as accelerometers, regis-tering changes in head velocity (or changestherein, i.e., acceleration), not head orienta-tion relative to gravity (which is accom-plished using the otolith organs). Thus, theintrinsic function of the canals does not ex-plain any relationship between LSC orien-tation and head posture. An alternative,
Fig. 14. Bivariate plot of foramen magnum orientation relative to orbital axis (new data) againsthead-neck angle (Strait and Ross, 1999). Homo has unusually airorhynch (dorsally rotated) orbitsrelative to its foramen magnum orientation. Across primates the relationship between these variables issignificant (r 5 0.541, P , 0.05); across nonhuman primates r 5 0.691 (P , 0.01).
TABLE 8. Pearson’s r and P-values for correlationsbetween measures of foramen magnum orientation and
measures of head morphology and orientation
FM , AOA FM , COr P r P
CBA1 0.482 0.2 0.160 0.6CBA4 0.145 0.7 0.258 0.5AOA 0.841 0.001* 20.378 0.3Head-neck angle 0.405 0.2 0.202 0.6Cerebellum/B-a-se 20.169 0.6 0.170 0.6
* P , 0.05.
PRIMATE CRANIAL BASE 149D.E. Lieberman et al.]
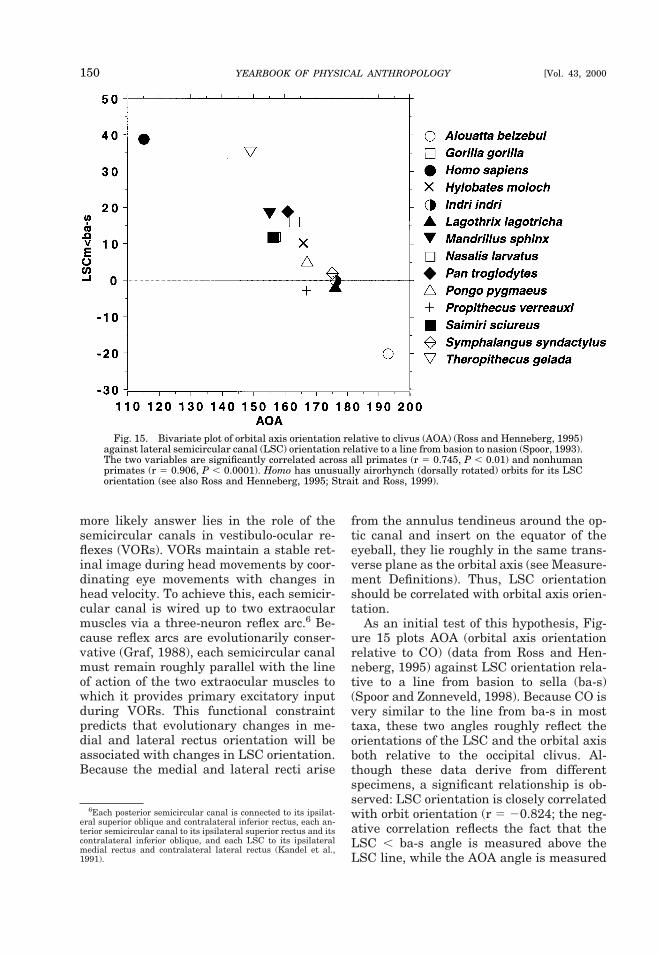
more likely answer lies in the role of thesemicircular canals in vestibulo-ocular re-flexes (VORs). VORs maintain a stable ret-inal image during head movements by coor-dinating eye movements with changes inhead velocity. To achieve this, each semicir-cular canal is wired up to two extraocularmuscles via a three-neuron reflex arc.6 Be-cause reflex arcs are evolutionarily conser-vative (Graf, 1988), each semicircular canalmust remain roughly parallel with the lineof action of the two extraocular muscles towhich it provides primary excitatory inputduring VORs. This functional constraintpredicts that evolutionary changes in me-dial and lateral rectus orientation will beassociated with changes in LSC orientation.Because the medial and lateral recti arise
from the annulus tendineus around the op-tic canal and insert on the equator of theeyeball, they lie roughly in the same trans-verse plane as the orbital axis (see Measure-ment Definitions). Thus, LSC orientationshould be correlated with orbital axis orien-tation.
As an initial test of this hypothesis, Fig-ure 15 plots AOA (orbital axis orientationrelative to CO) (data from Ross and Hen-neberg, 1995) against LSC orientation rela-tive to a line from basion to sella (ba-s)(Spoor and Zonneveld, 1998). Because CO isvery similar to the line from ba-s in mosttaxa, these two angles roughly reflect theorientations of the LSC and the orbital axisboth relative to the occipital clivus. Al-though these data derive from differentspecimens, a significant relationship is ob-served: LSC orientation is closely correlatedwith orbit orientation (r 5 20.824; the neg-ative correlation reflects the fact that theLSC , ba-s angle is measured above theLSC line, while the AOA angle is measured
6Each posterior semicircular canal is connected to its ipsilat-eral superior oblique and contralateral inferior rectus, each an-terior semicircular canal to its ipsilateral superior rectus and itscontralateral inferior oblique, and each LSC to its ipsilateralmedial rectus and contralateral lateral rectus (Kandel et al.,1991).
Fig. 15. Bivariate plot of orbital axis orientation relative to clivus (AOA) (Ross and Henneberg, 1995)against lateral semicircular canal (LSC) orientation relative to a line from basion to nasion (Spoor, 1993).The two variables are significantly correlated across all primates (r 5 0.745, P , 0.01) and nonhumanprimates (r 5 0.906, P , 0.0001). Homo has unusually airorhynch (dorsally rotated) orbits for its LSCorientation (see also Ross and Henneberg, 1995; Strait and Ross, 1999).
150 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000

below the orbital axis line). These datatherefore lend support to the hypothesisthat animals orient their heads at rest (andduring locomotion) primarily to orient theireyes towards the horizon rather than to ori-ent their semicircular canals in any partic-ular way. In sum, orbital axis orientationrelative to the clivus and foramen magnumorientation relative to the orbital axis arehypothesized to be evolutionary adaptationsto head posture during habitual locomotion,and appear necessary because of the limitedrange of motion at the atlanto-occipital joint(Graf et al., 1995).
MAJOR UNRESOLVED ISSUES OFCRANIAL BASE VARIATION IN PRIMATE
EVOLUTION
Studies of cranial base variation in fossilprimates and hominins have been rare be-cause this region of the skull is usuallypoorly preserved or destroyed in most fos-sils, and because it is hard to visualize ormeasure without radiographs or computedtomography (CT) scans. The coming years,however, are likely to see a renaissance ofresearch on the role of the cranial base inprimate cranial evolution as CT scans offossils become more readily available. Herewe review six major topics where future re-search on the cranial base in both fossil andextant primates promises to provide impor-tant insights: 1) the relationship betweenencephalization, circumorbital form, andthe origin of primates; 2) the evolution ofthe integrated “facial block” in haplorhines;3) the determinants of basicranial flexion inhominins; 4) the relationship between basi-cranial flexion and the shape of the vocaltract; 5) the role of the cranial base in facialretraction in Homo sapiens; and 6) the reli-ability of basicranial characters as indica-tors of primate phylogeny.
Primate origins, encephalization, andcircumorbital form
The basicranium likely played a key rolein the evolution of the unique configurationof the primate skull. Over the past threedecades, the visual predation hypothesis(VPH) has become a well-accepted model ofprimate origins (Martin, 1990; Fleagle,1999). The VPH argues that the first pri-
mates were nocturnal visual predators ofsmall invertebrates and vertebrates, andthis required more anteriorly facing andclosely approximated orbital apertures(Cartmill, 1970, 1972, 1974, 1992). In-creased orbital convergence enlarges thebinocular field for greater stereoscopic vi-sion and a clear retinal image for depthperception and prey location (Allman, 1977,1982). The VPH further posits that rela-tively larger orbits and grasping append-ages with nails are adaptations to beingnocturnal in an arboreal, terminal-branchsetting (Cartmill, 1970, 1972, 1974, 1992;Kay and Cartmill, 1974, 1977; Dagosto,1988; Covert and Hamrick, 1993; Hamrick,1998, 1999; Lemelin, 1999). These adapta-tions differ significantly from the cranialand locomotor specializations of putativesister taxa such as plesiadapiforms and der-mopterans (Cartmill, 1972, 1974, 1992; Kayand Cartmill, 1974, 1977; Beard, 1993; Ra-vosa et al., 2000a).
According to the VPH, increased orbitalconvergence moves the orbital apertures outof the plane of the temporal fossa, a condi-tion entailing greater ocular disruption dur-ing mastication (Cartmill, 1970, 1972, 1974,1992). A rigid postorbital bar may functionto stiffen the lateral orbital margins andthus counter ocular deformation during bit-ing and chewing to ensure a high level ofstereoscopic acuity in an organism that pro-cesses food while hunting and foraging(Cartmill, 1970, 1972). This appears to beparticularly important, given that strep-sirhines with unfused symphyses have beenshown to recruit relatively less balancing-side than working-side adductor muscleforce during unilateral mastication (Hy-lander et al., 1998, 2000). This differentialmuscle recruitment results in a pattern oflower strains along the balancing-side pos-torbital bar than the working-side postor-bital bar (Ravosa et al., 2000a). Thus, anorganism with a postorbital ligament and astepsirhine-like adductor pattern (the latterof which is inferred for basal primates basedon the presence of unfused symphyses; Ra-vosa, 1996, 1999) would experience anasymmetrical circumorbital and, in turn, anocular loading pattern that is hypothesized
PRIMATE CRANIAL BASE 151D.E. Lieberman et al.]

to compromise effective stereoscopic visualacuity.
Analyses of felids (perhaps the best ana-log for the skull of basal primates), herpes-tids, and pteropodids demonstrate thatpostorbital bar formation characterizes taxawith increased orbital convergence and/orgreater orbital frontation (Noble et al.,2000; Ravosa et al., 2000a). As both orbitalcharacteristics became more developed dur-ing early primate evolution (Simons, 1962;Cartmill, 1970, 1972, 1974, 1992; Szalay etal., 1987; Martin, 1990; Fleagle, 1999), it islikely that primate postorbital bar develop-ment is correlated with both evolutionarytransformations in the orbital complex (Ra-vosa et al., 2000a).
The role of orbital frontation in postor-bital bar formation is especially significantbecause it apparently reflects interactionsamong several factors unique to the earlyevolution of small primates (Ravosa et al.,2000a). First among these factors is enceph-alization. Both early primates and their pu-tative ancestors were tiny, weighing be-tween 100–300 g (Kay and Cartmill, 1977;Dagosto, 1988; Martin, 1990; Beard, 1993;Covert and Hamrick, 1993; Fleagle, 1999).Thus, the greater encephalization of basalprimates (Martin, 1990) did not result fromphyletic size decreases7 (Gould, 1975; Shea,1987). Instead, increased relative brain sizeand orbital frontation appear linked to noc-turnal visual predation (Ravosa et al.,2000a), which is related to increases in therelative size of the visual cortex (Cartmill,1974, 1992) and greater encephalization(Barton, 1998). This hypothesis comple-ments prior claims that increased enceph-alization in basal primates is related to theunique combination of arboreality, precoci-ality, and small body size (Shea, 1987).
Apart from being more encephalized thannonprimate archontans, the first primates(and felids) possessed relatively larger eyesand more convergent orbits (Kay and Cart-mill, 1977; Martin, 1990; Covert and Ham-rick, 1993; Cartmill, 1992; Ravosa et al.,
2000a). This combination of derived fea-tures creates a spatial packing problem inwhich the position of the frontal lobes andanterior cranial fossa have a major influ-ence on orbital aperture orientation, inte-grating morphological variation in these ad-jacent cranial regions (see above). Becauseneural and ocular size scale with negativeallometry (Schultz, 1940; Gould, 1975), thisstructural constraint would be particularlymarked in smaller, and thus more fron-tated, taxa—the morphospace in whichbasal primates and felids happen to developpostorbital bars.
Anthropoid origins and cranialbase-face interactions
Another interesting problem is the role ofthe cranial base in the fundamental rear-rangement of the face along the stem lin-eage leading to haplorhines and anthro-poids. These changes include: increasedorbital convergence and frontation; in-creased relative brain size, especially in thefrontal and temporal lobes; a more acutelyangled (flexed) cranial base; partial retrac-tion of the face below the braincase; re-duction of the olfactory apparatus andinterorbital region with concomitantapproximation of the orbits; relativelysmaller orbits associated with diurnality;and the presence of a postorbital septum(Cartmill, 1970, 1980; Radinsky, 1968,1979; Rosenberger, 1985, 1986; Ravosa,1991b; Ross and Ravosa, 1993; Ross,1995a,b). Not surprisingly, changes in cra-nial base shape have been key features of allthe models proposed to explain the order inwhich these features arose, their putativefunctions, and their interactions. Accordingto Cartmill (1970), the fundamental differ-ence between haplorhines and strepsirhinescenters on whether orbital convergence oc-curs towards the top of the skull or at thefront. If, as in haplorhines, the orbits con-verge anteriorly (associated with increasedorbital frontation), then the orbits not onlycompress the olfactory region, reducing itsdimensions, but also become closely approx-imated to the midline anterior cranial base.This necessarily reduces the size of the ol-factory apparatus and forms an interorbitalseptum, facilitating a subsequent increase
7As phyletic dwarfs are more encephalized than like-sizedsister taxa (Gould, 1975; Shea, 1987), presumably they are alsomore flexed, highlighting the complex relationships betweenbrain evolution and changes in basicranial and facial form.
152 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000

in relative encephalization by expansion ofthe brain forward over the top of the orbits.It has been argued that, as predicted byDabelow (1931), by bringing the orbits andanterior cranial base into close continuity,they became structurally integrated suchthat changes in the orientation of one nec-essarily affect the other (Ross and Ravosa,1993). Small nocturnal strepsirhines, incontrast, tend to converge their orbits to-wards the top of their head, not compressingthe olfactory apparatus, and preventingbrain expansion over the orbits concomitantwith a shift to diurnality. The orbits andanterior cranial base in all strepsirhines arenot as unified structurally, and thus basi-cranial flexion and orbit orientation do notcovary as greatly.
The evidence that all omomyiforms thatare well enough preserved have an interor-bital septum below the olfactory tract, likethat of Tarsius and small anthropoids (Ross,1994), suggests that this integration of an-terior cranial base and upper face may be anearly unique derived feature of the haplo-rhine stem lineage. This integration set thestage for the subsequent evolution of thehaplorhine postorbital septum. Increasedorbital frontation (klinorhynchy) and con-vergence in early anthropoids caused theanterior temporal muscles and orbit to comeinto close proximity, leading to the evolutionof a postorbital septum (Ross, 1995b, 1996).Exactly why increased frontation occurredin haplorhines remains unexplained, al-though it has been argued that increasedrelative brain size might have caused in-creased basicranial flexion, producing in-creased orbital frontation as a consequenceof the integration of the anterior cranialbase and orbits (Ross, 1996; see also Ravosaet al., 2000a on the effects of small size onskull form in basal primates). This hypoth-esis remains to be evaluated in fossil stemanthropoids for want of well-preserved spec-imens, but is not supported by the fact thatearly anthropoids had a postorbital septumin conjunction with relatively small brains(Ross, 2000). It remains to be determinedwhy the haplorhine stem lineage evolvedmore frontated orbits, but the findings ofStrait and Ross (1999) suggest that the roleof posture should be considered.
Variation in hominin cranial base angle
How to account for variation in basicra-nial flexion in hominins has been controver-sial and remains unresolved. Several recentstudies (e.g., Ross and Henneberg, 1995;Spoor, 1997) have shown that the cranialbase in Australopithecus and Homo is gen-erally more flexed than in Pan or other non-human primates. This difference raises twoquestions. First, how much is cranial baseflexion among early hominins related to up-right posture, facial orientation, or brainsize? Second, what factors account for theobserved variation in cranial base angleamong hominins?
Although cranial base angles vary sub-stantially among hominin species, australo-pithecines have CBAs generally intermedi-ate between humans and chimpanzees, butwith more flexed cranial bases among A.boisei and less flexed cranial bases for A.aethiopicus (KNM-WT 17000) (F. Spoor,personal communication). Until now, thehypothesis that cranial base flexion in homi-nins is an adaptation for increased brainsize relative to basicranial length (Gould,1977) has received the most support (Rossand Ravosa, 1993; Ross and Henneberg,1995; Spoor, 1997; McCarthy, 2001). In par-ticular, if one measures IRE5 vs. CBA1(thereby including the cribriform plate inmeasures of both basicranial length and theangle of the cranial base), then H. sapiensand other hominin taxa such as A. africanushave exactly the degree of flexion expectedby basicranial length.
However, not all of the variation in homi-nin CBA can be explained by relative brainsize. For example, Neanderthals and ar-chaic Homo fossils such as Kabwe have con-siderably more extended CBA1s than H. sa-piens (about 15°), even though they arebipedal and similarly encephalized (Ruff etal., 1997). In addition, other measures ofcranial base angle and relative encephaliza-tion, which do not include the cribriformplate (CBA4 and IRE5), indicate that H.sapiens have less flexed cranial bases thenexpected for anthropoids of their size (Rossand Henneberg, 1995). This finding high-lights the likelihood that no single explana-tion will account for interspecific differences
PRIMATE CRANIAL BASE 153D.E. Lieberman et al.]

in CBA among hominins. Other factors thathave been argued to contribute to variationin CBA, including facial kyphosis and headand neck posture, cannot be shown to beimportant correlates of basicranial flexion.Most importantly, the comparative study byStrait and Ross (1999) of CBA, head-neckangle, and orbital axis orientation amongextant primates found that basicranial flex-ion is primarily influenced by relative brainsize and not head and neck posture. Thetraditional view that human basicranialflexion is somehow an adaptation for ortho-grade posture is no longer tenable.
Cranial base flexion and vocal tractshape in hominins
Flexion of the cranial base has been ar-gued to be an important correlate of theshape of the vocal tract. In particular, sev-eral researchers (Lieberman et al., 1972,1992; Laitman and Crelin, 1976; Laitman etal., 1978, 1979; Laitman and Heimbuch,1982) claimed that the degree of flexion ofthe external cranial base (between the infe-rior aspect of the basioccipital clivus and thepalate) influences the position of the hyo-laryngeal complex. In most mammals, thehyoid and larynx lie close to the soft palateso that the epiglottis and soft palate canengage, permitting simultaneous breathingand swallowing (Laitman and Reidenberg,1993; German et al., 1996; Larson and Her-ring, 1996; Crompton et al., 1997). In con-trast, humans have a uniquely shaped phar-ynx in which the hyo-laryngeal complexdescends inferiorly relative to the oral cav-ity, causing the soft palate and epiglottis tobecome disengaged so that the trachea andesophagus share a common passageway(Negus, 1949; Crelin, 1973). Hyo-laryngealdescent is an important physiological basisfor many aspects of human speech. Flexionof the external cranial base in combinationwith a low position of the larynx relative tothe palate divide the human vocal tract (VT)into separate horizontal and vertical “tubes”of approximately equal length, whose cross-sectional areas can be modified indepen-dently at least tenfold by the tongue(Stevens and House, 1955; Fant, 1960;Stevens, 1972; Baer et al., 1991). Dynamicfiltering in the two-tube human VT func-
tions to produce a wide range of vowelswhose formant frequencies are acousticallydistinct regardless of vocal tract length(Fant, 1960; Nearey, 1978; Lieberman,1984; Carre et al., 1994; Beckman et al.,1995).
Although the speech-related acousticalproperties of the unique human vocal tractare not in dispute, the role of cranial baseflexion in hyo-laryngeal descent is contro-versial, yet potentially important for recon-structing the anatomy of the vocal tract infossil hominins Two questions are of specialimportance. First, what is the relationshipof external cranial base flexion to the de-scent and/or positioning of the hyoid rela-tive to the oral cavity? Second, how does theangle of the external cranial base changeduring development, and what is its rela-tionship to internal cranial base angle?
The original hypothesis that flexion of theexternal cranial base in humans contributesto laryngeal descent, and thus can be usedto reconstruct the vocal tract of fossil homi-nins (Lieberman and Crelin, 1971; Lieber-man et al., 1972; Laitman and Crelin, 1976;Laitman and Heimbuch, 1982; Lieberman,1984), was based on three observations.First, these researchers inferred a relation-ship between cranial base flexion and laryn-geal descent on the basis of comparisons ofneonatal and adult humans vs. nonhumanprimates. In particular, human neonates,like nonhuman primate adults, have a rela-tively extended cranial base and a high hy-oid, whereas only human adults have both aflexed cranial base and a descended larynx.Second, internal and external basicranialflexion were believed to covary with the de-scent of the larynx (George, 1978). Third,external cranial base flexion was believed toreorient the suprahyoid muscles and liga-ments more vertically, and to shorten theantero-posterior length of the oropharynxbetween the palate and the cervical verte-brae, thus requiring the larynx and poste-rior tongue to be positioned lower relative tothe hard palate.
Lieberman and McCarthy (1999) testedthe relationship between cranial base flex-ion and hyo-laryngeal position using a lon-gitudinal series of radiographs in humans(the Denver Growth Study), which pre-
154 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000
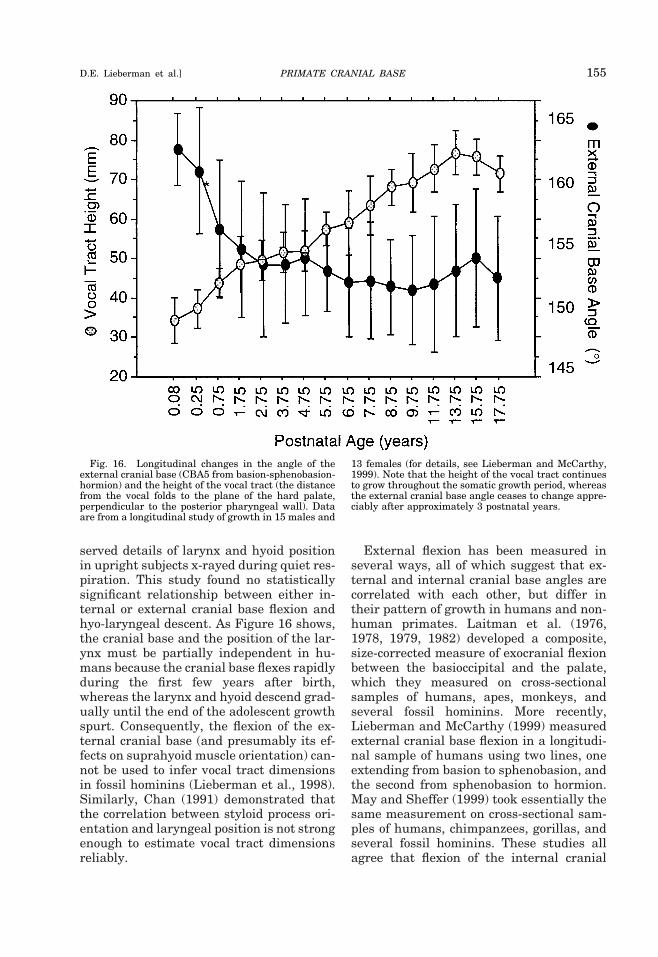
served details of larynx and hyoid positionin upright subjects x-rayed during quiet res-piration. This study found no statisticallysignificant relationship between either in-ternal or external cranial base flexion andhyo-laryngeal descent. As Figure 16 shows,the cranial base and the position of the lar-ynx must be partially independent in hu-mans because the cranial base flexes rapidlyduring the first few years after birth,whereas the larynx and hyoid descend grad-ually until the end of the adolescent growthspurt. Consequently, the flexion of the ex-ternal cranial base (and presumably its ef-fects on suprahyoid muscle orientation) can-not be used to infer vocal tract dimensionsin fossil hominins (Lieberman et al., 1998).Similarly, Chan (1991) demonstrated thatthe correlation between styloid process ori-entation and laryngeal position is not strongenough to estimate vocal tract dimensionsreliably.
External flexion has been measured inseveral ways, all of which suggest that ex-ternal and internal cranial base angles arecorrelated with each other, but differ intheir pattern of growth in humans and non-human primates. Laitman et al. (1976,1978, 1979, 1982) developed a composite,size-corrected measure of exocranial flexionbetween the basioccipital and the palate,which they measured on cross-sectionalsamples of humans, apes, monkeys, andseveral fossil hominins. More recently,Lieberman and McCarthy (1999) measuredexternal cranial base flexion in a longitudi-nal sample of humans using two lines, oneextending from basion to sphenobasion, andthe second from sphenobasion to hormion.May and Sheffer (1999) took essentially thesame measurement on cross-sectional sam-ples of humans, chimpanzees, gorillas, andseveral fossil hominins. These studies allagree that flexion of the internal cranial
Fig. 16. Longitudinal changes in the angle of theexternal cranial base (CBA5 from basion-sphenobasion-hormion) and the height of the vocal tract (the distancefrom the vocal folds to the plane of the hard palate,perpendicular to the posterior pharyngeal wall). Dataare from a longitudinal study of growth in 15 males and
13 females (for details, see Lieberman and McCarthy,1999). Note that the height of the vocal tract continuesto grow throughout the somatic growth period, whereasthe external cranial base angle ceases to change appre-ciably after approximately 3 postnatal years.
PRIMATE CRANIAL BASE 155D.E. Lieberman et al.]

base in humans occurs prior to the eruptionof the first permanent molars and then re-mains stable, but that the external cranialbase extends gradually in all nonhuman pri-mates throughout the period of facialgrowth.8 Moreover, the patterns of externaland internal cranial base angulation mirroreach other. The internal cranial base (mea-sured using both Ba-S-FC and Landzert’sangle) flexes in humans rapidly prior to 2postnatal years and then remains stable,but extends in nonhuman primates gradu-ally throughout the period of facial growth(Fig. 7) (Lieberman and McCarthy, 1999).
Cranial base shape and facialprojection in Homo
Although the effects of cranial base angu-lation on the angle of the face relative to therest of the skull (facial kyphosis) have longbeen the subject of much research (seeabove), there has been recent interest in therole of the cranial base on facial projection.Facial projection (which is a more generalterm for neuro-orbital disjunction) is de-fined here as the extent to which the nonros-tral portion of the face is positioned anteri-orly relative to the foramen caecum, themost antero-inferior point on the cranialbase (note that facial projection and prog-nathism are different). Variation in facialprojection, along with an understanding oftheir developmental bases, may be impor-tant for testing hypotheses about recenthominin evolution. In particular, Lieber-man (1995, 1998, 2000) has argued thatvariation in facial projection accounts formany of the major differences in overallcraniofacial form between H. sapiens andother closely-related “archaic” Homo taxa,including the Neanderthals. Whereas allnonextant hominins have projecting faces,“anatomically modern” H. sapiens isuniquely characterized by a retracted facialprofile in which the majority of the face liesbeneath the braincase (Weidenreich, 1941;Moss and Young, 1960; Vinyard, 1994;Lieberman, 1995, 1998; Vinyard and Smith,
1997; May and Sheffer, 1999; Ravosa et al.,2000b). As a consequence, H. sapiens alsohas a more vertical frontal profile, less pro-jecting browridges, a rounder overall cra-nial shape, and a relatively shorter oropha-ryngeal space between the back of the hardpalate and the foramen magnum—virtuallyall of the supposed autapomorphies of “an-atomically modern” H. sapiens.
What is the role of the cranial base infacial projection? Lieberman (1998, 2000)proposed that four independent factors ac-count for variation in facial projection: 1)antero-posterior facial length, 2) anteriorcranial base length, 3) cranial base angle,and 4) the antero-posterior length of themiddle cranial fossa from sella to PMplane.9 Each of these variables has a differ-ent growth pattern, but combine to influ-ence the position of the face relative to thebasicranium and neurocranium. For exam-ple, facial projection can occur through hav-ing a long face relative to a short anteriorcranial fossa, a long middle cranial fossarelative to the length of the anterior cranialfossa, and/or a more extended cranial base.Partial correlation analyses of cross-sec-tional samples of Homo sapiens and Pantroglodytes indicate that each contributessignificantly to the ontogeny of facial projec-tion in humans and apes when the associa-tions between these variables and withoverall cranial length and endocranial vol-ume as well as other cranial dimensions areheld constant (Lieberman, 2000). In otherwords, chimpanzees and humans with rela-tively longer faces, shorter anterior cranialbases, less flexed cranial bases, and/orlonger middle cranial fossae tend to haverelatively more projecting faces.
In an analysis of radiographs of fossilhominins, Lieberman (1998) argued thatthe major cause for facial retraction and itsresulting effects on modern human cranialshape was a change in the cranial baserather than the face itself. Specifically, mid-dle cranial fossa length (termed ASL) wasestimated to be approximately 25% shorterin anatomically modern humans, both re-
8May and Sheffer (1999) did not find evidence for postnatalextension of the cranial base in Pan, largely because of insuffi-cient sample sizes that were divided into overly large ontoge-netic stages.
9This dimension was originally termed anterior sphenoidlength (ASL), but it is really a measure of the midline prechordallength of the middle cranial fossa (Lieberman, 2000).
156 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000
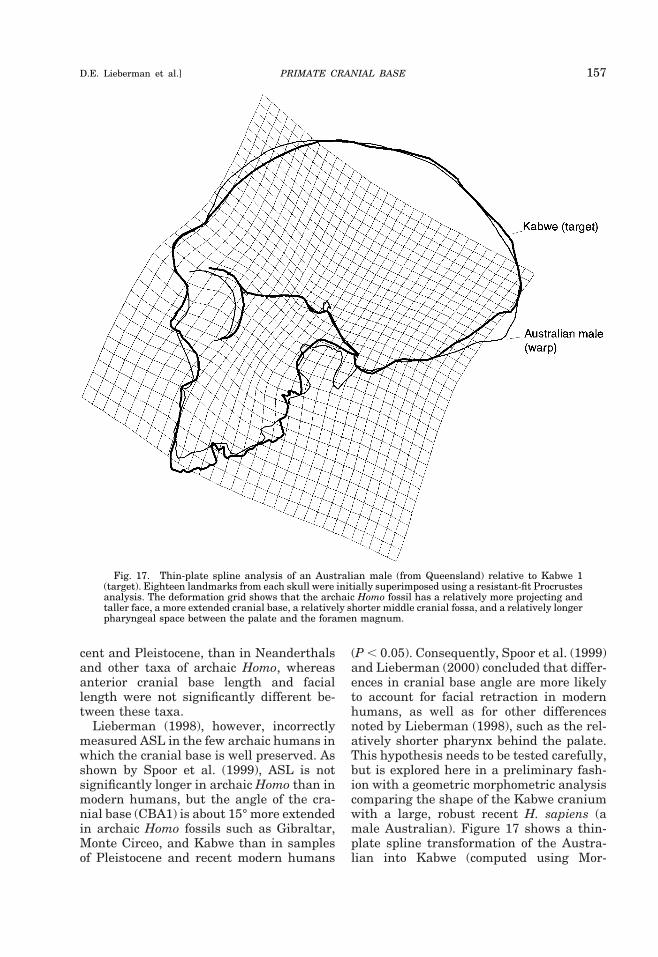
cent and Pleistocene, than in Neanderthalsand other taxa of archaic Homo, whereasanterior cranial base length and faciallength were not significantly different be-tween these taxa.
Lieberman (1998), however, incorrectlymeasured ASL in the few archaic humans inwhich the cranial base is well preserved. Asshown by Spoor et al. (1999), ASL is notsignificantly longer in archaic Homo than inmodern humans, but the angle of the cra-nial base (CBA1) is about 15° more extendedin archaic Homo fossils such as Gibraltar,Monte Circeo, and Kabwe than in samplesof Pleistocene and recent modern humans
(P , 0.05). Consequently, Spoor et al. (1999)and Lieberman (2000) concluded that differ-ences in cranial base angle are more likelyto account for facial retraction in modernhumans, as well as for other differencesnoted by Lieberman (1998), such as the rel-atively shorter pharynx behind the palate.This hypothesis needs to be tested carefully,but is explored here in a preliminary fash-ion with a geometric morphometric analysiscomparing the shape of the Kabwe craniumwith a large, robust recent H. sapiens (amale Australian). Figure 17 shows a thin-plate spline transformation of the Austra-lian into Kabwe (computed using Mor-
Fig. 17. Thin-plate spline analysis of an Australian male (from Queensland) relative to Kabwe 1(target). Eighteen landmarks from each skull were initially superimposed using a resistant-fit Procrustesanalysis. The deformation grid shows that the archaic Homo fossil has a relatively more projecting andtaller face, a more extended cranial base, a relatively shorter middle cranial fossa, and a relatively longerpharyngeal space between the palate and the foramen magnum.
PRIMATE CRANIAL BASE 157D.E. Lieberman et al.]

phometrika, version 0.007, Jeff Walker,Chicago, IL), based on an initial resistant-fitsuperimposition Procrustes analysis of sixcranial base landmarks, seven facial land-marks, and five neurocranial landmarks.10
The thin-plate spline, which is based on aProcrustes analysis that geometrically cor-rects for most effects of size differences,shows that Kabwe has a considerably moreprojecting face (i.e., between nasion and theforamen caecum) in conjunction with a rel-atively more extended cranial base (14o), adorsally rotated PM plane relative to theposterior cranial base, and a relativelylonger pharyngeal space between the max-illa and the foramen magnum. In addition,when one examines specific, size-correcteddimensions (standardized by the geometricmean of all the interlandmark distances),midfacial length (PM-Na) is 19% longer inKabwe than in the Australian modern indi-vidual, but the length of the anterior cranialbase (S-FC) is only 4% shorter. In otherwords, Kabwe exhibits a considerably moreprojecting face than the H. sapiens speci-men because it has a more extended cranialbase in conjunction with a relatively longerface. Changes in middle cranial fossa shape,therefore, appear to have had major effectson facial retraction in human evolution, butneed to be further examined using addi-tional specimens.
Additional, intriguing evidence that thecranial base can play an important role infacial retraction may be come from studiesof craniofacial growth abnormalities andfrom laboratory studies of mice. Mice thatare homozygous-recessive for the retrog-nathic Brachyrrhine (Br) allele are charac-terized by a primary growth defect in theanterior cranial base (Beechey et al., 1998;Ma and Lozanoff, 1999) that leads to a se-verely retrognathic midfacial profile but amorphologically normal nasal septum andface (Lozanoff, 1993; Lozanoff et al., 1994;
Ma and Lozanoff, 1999). The extent towhich the morphological differences be-tween the retrognathic Br mouse and con-trols is at all similar to the differences evi-dent between H. sapiens and archaic Homohas yet to be determined. However, thesestudies, in conjunction with facial growthdefects caused by chondrogenic growth dis-orders such as Crouzon syndrome, Pierre-Robin syndrome, and Down syndrome, high-light the important role the cranial baseplays in facial growth and integration (Dav-id et al., 1989).
Cranial base characters in phylogeneticanalyses
One final consideration of interest iswhether it might be profitable to focus oncharacters from the cranial base in taxo-nomic and phylogenetic analyses of pri-mates. This possibility has been raised by anumber of authors (e.g., Olson, 1985; Shea,1985a, 1988; Lieberman et al., 1996; Lieber-man, 1997; Strait et al., 1997; Strait, 1998)for three reasons. First, the cranial basederives from endochrondral precursorsrather than though intramembranous ossi-fication processes (endochondral bones arethought to have more direct genetic influ-ence in terms of initial shape); second, thecranial base is the first part of the skull toreach adult size and shape (Moore andLavelle, 1974); and third, the cranial basemay play a greater role in influencing facialshape than vice versa (see above). Conse-quently, one might expect cranial base char-acters to preserve more phylogenetic signalthan facial characters by virtue of beingmore heritable and less influenced by theepigenetic responses to external influencesduring postnatal ontogeny rampant in thefacial skeleton (Herring, 1993).
Expectations aside, there are currently nodata suggesting that the basicranium is ac-tually a better source of characters thanother regions of the skull for phylogeneticanalyses. In terms of narrow-sense herita-bility (h2), basicranial, neurocranial, and fa-cial characters have similar levels of herita-bility among primates (Sjøvold, 1984;Cheverud and Buikstra, 1982; Cheverud,1995). In addition, basicranial variables ap-pear to be equally well (or poorly) correlated
10These landmarks are: basion, sella, pituitary point, sphenoi-dale, PMp (the anteriormost point on the greater wings of thesphenoid), foramen caecum, nasion, ANS, PNS, prosthion, max-illary tuberosities, fronton, orbitale, opisthocranion, the superi-or-most point on the vault, fronton, the inferior-most point onthe supratoral sulcus, glabella. Resistant fit superimpositionwas used in order to minimize effects of the very different shapeof the neurocranium in archaic Homo vs. modern H. sapiens.
158 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000

with other dimensions or features in homi-noids (Strait, 1998), and in the human skull(Lieberman et al., 2000). Finally, those fewstudies that have focused on cranial basecharacters do not yield results substantiallydifferent from analyses that incorporateother craniodental characters. Lieberman etal. (1996) found that basicranial and vaultcharacters tend to yield similar cladogramsthat differ only slightly from cladogramsbased on facial characters. Strait et al.(1997) did not specifically examine charac-ters from the cranial base, but found nomajor difference between trees based onmasticatory characters vs. those that werepredominantly neurocranial and basicra-nial. In their study of primate higher taxo-nomic relationships, Ross et al. (1998) foundthe cranial data, consisting primarily of ba-sicranial traits, to yield trees similar tothose produced by the dental data, althoughwith better resolution at older nodes. Thus,whether the basicranium is a better sourceof phylogenetic data remains open to ques-tion. The cranial base may be a good place tolook for reliable characters if researchersare to focus on characters that describe de-velopmental processes or events (e.g., flex-ion vs. extension at the spheno-occipitalsynchondrosis) rather than using charac-ters that solely describe morphological vari-ation (e.g., the angle of the whole cranialbase) (Hall, 1994; Lieberman, 1999). Thishypothesis, however, has yet to be tested.
The above studies, however, raise anotherkey issue relevant to phylogenetic studies:the problem of independence. Cladisticanalyses explicitly require one to use inde-pendent characters to avoid problems ofconvergent and correlated characters incor-rectly biasing the outcome of any parsimonyanalysis. Yet the above studies demonstratethe existence of multiple and complicatedinteractions between the cranial base andthe neurocranium and between the cranialbase and the face. For example, variationsin orbit size and orientation are linked tovariations in the angle of the cranial base,brain size, and the orientation of the facerelative to the rest of the skull. Many ofthese features have been treated as inde-pendent characters in recent cladistic anal-yses (e.g., Strait et al, 1997), but their effect
on the results has yet to be determined.Further study of these interactions isneeded to improve the reliability of phyloge-netic analyses, and again highlight the needto consider morphological characters interms of their generative processes (e.g.,Gould, 1977; Cheverud, 1982; Shea, 1985b;Hall, 1994; Lieberman, 1999).
CONCLUSIONS
Since the last major reviews of cranialbase anatomy in primates (Scott, 1958;Moore and Lavelle, 1974; Sirianni andSwindler, 1979), there has been a tremen-dous increase in our knowledge of chondro-cranial embryology, the patterns and pro-cesses of basicranial growth, and the natureof basicranial variation across primates.Major advances include details of the mor-phogenetic independence between the pre-chordal and postchordal portions of thechondrocranium; comparative data on therelative importance of brain size, orbital ori-entation, facial orientation, and posture asfactors that account for variation in cranialbase angulation; and ontogenetic and com-parative data on the structural relation-ships between the anterior cranial base andupper face in haplorhines vs. strepsirhines,and their influences on facial form. Despitethese advances, many aspects of cranialbase growth, variation, and function remainpoorly understood. For example, we do notknow what ontogenetic processes governflexion and extension of the cranial base, orwhich synchondroses are active in cranialbase elongation vs. angulation in humansand other primates. Future research needsto be aimed at studying these processes asthey relate to cranial shape, function, andevolution.
To conclude, we highlight two importantpractical and theoretical issues which webelieve merit special consideration, andwhich promise to further our understandingof craniofacial growth and variation. First,what are the major factors that generatevariation in the cranial base among pri-mates? Second, to what extent does the cra-nial base function to coordinate these fac-tors within the craniofacial complex duringgrowth and development? As noted above,these questions need to be addressed using
PRIMATE CRANIAL BASE 159D.E. Lieberman et al.]

two approaches. Ontogenetic studies arecrucial for testing hypotheses about the gen-eration of morphological covariation pat-terns, and comparative studies are impor-tant for probing the extent to whichinterspecific patterns of morphological evo-lution are epiphenomena of developmentalprocesses.
What major factors generate variationin the cranial base?
The studies summarized above suggestthat relative brain size, particularly relativeto basicranial length, is an important deter-minant of the degree of basicranial angula-tion. However, the effects of (especiallyprenatal) brain shape on basicranialmorphology have yet to be thoroughly inves-tigated. Further experimental and compar-ative morphological research is needed torelate soft-tissue and bony morphology inthe primate head. Despite a long traditionsuggesting links between basicranial mor-phology and head, neck, or body posture,there is currently little empirical supportfor the hypothesis that these factors are di-rectly related to variation in the cranialbase, especially angulation in the midsagit-tal plane. Better data are also needed onhead and neck posture during locomotionbefore locomotor-related cranial adapta-tions can be definitively identified. How-ever, connections between facial orientationand basicranial morphology on the one handand head posture on the other leave openthe possibility of an indirect link betweenbasicranial angle and head posture.
What role does the cranial base play incraniofacial integration?
Because many variables influence cranialbase shape, it follows that these variablesalso influence other aspects of cranial shapevia the cranial base. Consequently, a keyissue that emerges repeatedly in discus-sions of the role of the basicranium in cra-nial development, growth, and evolution isintegration. Integration, which is definedhere as “the association of elements througha set of causal mechanisms so that changein one element is reflected by change inanother” (Smith, 1996, p. 70), thereby gen-erating a pattern of significant covariation
(see also Olson and Miller, 1958; Cheverud,1982; Zelditch, 1988),11 is an important is-sue because of the many genetic, develop-mental, and functional interactions that oc-cur between the basicranium and itsneighboring anatomical components. As de-scribed above, the cranial base is likely todirectly interact, developmentally and func-tionally, with various adjoining skeletal,muscular, and neurosensory complexes,most notably the brain, the orbits, the eth-momaxillary complex, and the neck. These“units,” in turn, have direct and indirectinteractions with other putative units suchas the oropharynx, nasopharynx, mandible,and maxillary arches.
But does the cranial base play an active ora passive role in integrating cranial shapeamong these disparate units, and how muchintegration actually occurs? On an intuitivelevel, there are several reasons to suggestthat the cranial base acts in part as a struc-tural “interface” during growth between thebrain and the face, and between the headand the neck. In many regions, the basicra-nium serves as the actual structural bound-ary between disparate components of theskull: the floor of the anterior cranial fossais the roof of the orbits; the back of themidface is the front of the middle cranialfossa; and the posterior cranial base is theposterior roof of the oropharynx. Yet, inspite of these obvious relationships, it isdifficult to define or assess quantitative hy-potheses about craniofacial integration be-cause we know so little about the extent andnature of the numerous interactions thatpresumably occur between and among re-gions of the cranial base and other parts ofthe skull. What are the actual units in thecranial base and skull that interact, andwhat regulates their interrelations and, es-pecially, interactions? In other words, theappropriate null hypothesis to be tested isthat the cranial base, while associated withvariation in other parts of the skull, plays
11Note that some researchers (most notably Olson and Miller,1958) define morphological integration solely as a multivariatepattern of covariation with regard to some a priori biologicalhypothesis (thus recognizing the importance of generative pro-cesses), whereas others (e.g., Cheverud, 1996) define integrationmore generally as either a pattern or a process that refers to“connections or relationships among morphological elements.”
160 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000

no more of an integrative role than anyother part of the skull (e.g., the mandible,the brain).
In order to explore the basicranium’s rolein craniofacial integration, it is first neces-sary to decide how to test hypotheses aboutthe many different processes through whichintegration occurs, and the many structuraland functional levels at which integration isevident. Readers interested in this huge andcomplex topic (which is too large to reviewthoroughly here) should consult the recentreview by Chernoff and Magwene (1999). Asa first-order analysis, it is useful to testhypotheses about developmental integra-tion by considering the processes by whichdifferent components of a system influenceone another. Thus, in the context of cranialbase growth within the skull, integrationcan occur through the direct inductiveand/or mechanical effects of neighboring tis-sue-tissue interactions (see Moss, 1997);through secondary effects of growth in oneregion causing changes in the positional re-lationships among bones in other regions;and genetically through single genes whichhave effects on multiple regions, or throughthe coordinated action of multiple genes viapleiotropy and linkage (Atchley and Hall,1991; Cheverud, 1982, 1995, 1996; Zelditchand Fink, 1995, Zelditch and Fink, 1996;Chernoff and Magwene, 1999). Of course,the strength of such ontogenetic interac-tions can vary due to changes in the timingof developmental events, something of greatimportance for the analysis of heterochrony,as well as due to allometric (size-correlated,size-required) factors. Note that we considerhere integration solely in terms of develop-mental events and processes (e.g., epige-netic responses to mechanical loading, dis-placement due to cell division), but it isuseful to recognize that developmental inte-gration leads to, and is sometimes causedby, structural and functional integration.
Another problem with assessing any inte-grative role of the cranial base within theskull is the lack of widely accepted criteriafor testing hypotheses of integration. Fol-lowing Olson and Miller (1958) andChernoff and Magwene (1999), we applythree nonexclusive criteria (correlation, con-straint, and ontogenetic sequence) in a pre-
liminary fashion to solely phenotypic varia-tion in the cranial base, using the datapresented above.
Correlation. Integration is most basi-cally revealed by complex patterns of corre-lation and covariation which indicate a lackof independence among variables (see Chev-erud, 1995, 1996), and which can be recog-nized a posteriori by comparing theoreti-cally and empirically derived correlationmatrices (Cheverud, 1982; Shea, 1985b;Zelditch, 1987, 1988; Cheverud et al., 1989;Wagner, 1989). Such studies have yet to becarried out for models that explicitly focuson the primate cranial base (but see Chev-erud, 1995). However, as noted above, ahigh degree of covariation is frequently ev-ident in interspecific analyses of variationin the cranial base and other parts of theskull. Perhaps the most obvious example ofthis phenomenon is the percentage of vari-ation in cranial base angle accounted for byfactors such as brain volume, basicraniallength, facial angle, and posture. IRE1,which appears to be the dominant factorthat influences cranial base angle in pri-mates, explains 58% of the variation inCBA1 and 38% of the variation in CBA4 (seeTable 5). Furthermore, according to Rossand Ravosa (1993, their Table 2), orbitalaxis orientation explains 41% of the varia-tion in CBA4 among primates, while facialorientation explains 22% of the variation inCBA4 among primates. The partial correla-tion analysis of Strait and Ross (1999) (dis-cussed above) found that IRE explained 36%of the variation in CBA1 when the effects oforbital axis and head-neck angle were fac-tored out. Further evidence for this sort ofcomplex pattern of covariation is docu-mented among humans between CBA, cra-nial base length, cranial base width, andbrain volume (Lieberman et al., 2000).These results are indicative (but not proof)of a pattern whereby multiple factors com-bine to influence CBA in such a way thatvariation in CBA itself may play some rolein modulating the interactions among dif-ferent, spatially separated components ofthe cranium. This hypothesis, however, re-quires further testing with comparative on-togenetic data.
PRIMATE CRANIAL BASE 161D.E. Lieberman et al.]

Constraint. Another expected outcome ofintegration is constraint, which is definedmost generally as a restriction or limitationon variation. Regardless of their causes,which can be phylogenetic, functional, de-velopmental, or structural (Alberch, 1985;Maynard Smith et al., 1985), constraints aremost basically evident in patterns of invari-ance. There has not been much work onphenotypic constraint in the cranial base,but several examples suggest it deservesfurther research. One source of evidence forconstraint is revealed by allometry, whichmeasures size-related conservation of shape(keeping in mind that size-correlated pat-terns due to epigenetic and perhaps geneticfactors differ from size-required patternsthat have a primarily functional basis). AsTable 4 (see also Strait, 1999; McCarthy,2001) illustrates, there are numerous,strongly correlated ontogenetic scaling rela-tionships across primates between variouscomponents of the cranial base, the volumeof neural regions, and facial dimensions. In-tegration between the brain and face via thecranial base is implied by the fact that manyscaling relationships between contiguousanatomical units (e.g., the noncortical brainand the posterior cranial base) result in ad-ditional strongly correlated scaling relation-ships between noncontiguous anatomicalunits, such as those between brain volumeand facial size. These scaling relationshipsrequire more study.
Another potential source of evidence re-garding the presence of a constraint are pat-terns of angular invariance. One importantexample is the apparently invariant 90° an-gle between the PM plane and the NHA,and the limited variability that results fromthis relationship on the angle between thePM plane and both the planum sphenoi-deum and the anterior cranial base (S-FC)in anthropoids (McCarthy and Lieberman,2001). Other less secure examples of invari-ance may include the relationship betweencribriform plate orientation and facial ori-entation (Ravosa and Shea, 1994), and thenear 45° angle between the external audi-tory meatus, the maxillary tuberosities, andthe midpoint of the orbital aperture (Bro-mage, 1992). Further research, however, isneeded on the extent to which invariant an-
gles and spatial relationships occur in theskull, and additional research is needed toassess the developmental, structural, andfunctional bases of these relationships, andwhether they reflect constraints that resultfrom integration.
Ontogenetic sequence. Finally, hypothe-ses of phenotypic integration may some-times be inferred or tested by examiningontogenetic sequences during normalgrowth and in the context of controlled ex-periments. Ontogenetic data allow one totest hypotheses of integration by examiningthe structural relationships between onevariable and another during growth (e.g.,heterochrony, heterotopy), and to comparethe pattern and timing of developmentalevents. So far, there have been few attemptsto examine hypotheses of integration in thecranial base using ontogenetic data (espe-cially from embryonic stages), but a few ex-amples indicate that the sequence of inter-actions between the cranial base, the face,and the brain are complex and multiphasic,with the cranial base mediating various in-teractions between the face and the brain.For example, the prenatal human cranialbase initially flexes, then remains stable,and then extends, all during periods of rapidneural growth (Jeffery, 1999). Postnatally,the nonhuman primate cranial base (beststudied in Pan and Macaca) appears to ex-tend slightly during the period of neuralgrowth, and subsequently extends morerapidly and for a long time as the face con-tinues to grow. In contrast, the human cra-nial base flexes rapidly during the first fewyears of brain growth, and subsequently re-mains stable. These contrasting sequencesimply multiple interactions between thebrain and the face via the cranial base, buthave yet to be resolved in terms of the actualprocesses that cause flexion and extensionat specific locations during different periodsof growth. One hypothesis, which remainsto be tested, is that the cranial base func-tions to accommodate and perhaps coordi-nate these different aspects of growth. Con-trolled experimental studies, which canpotentially isolate local and regional effectsof specific growth stimuli on the cranial baseand the rest of the skull, are a promising
162 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000

avenue for future research on this problem(see Sarnat, 1982; Butow, 1990; Reidenbergand Laitman, 1991).
Future research
Although the cranial base does appear toplay some integrative role in the cranium,we have only a vague, incomplete picture ofhow this integration occurs, and how muchof a role it plays in influencing various as-pects of craniofacial form and function, andhow these processes relate to evolutionaryshifts in the primate cranial base. More re-search is needed to isolate and define theactual morphogenetic units which interact,to identify and quantify their direct and in-direct interactions, and to understand theprocesses by which they interact. Thesegoals may be accomplished by combining atleast four approaches. First, we need togather more three-dimensional data on cra-nial base variation in ontogenetic samplesat all stages of growth and development (fe-tal to adult) among different species. Thecranial base and the rest of the skull com-prise a complex three-dimensional structurewhose internal and external structures dif-fer substantially, yet most studies of cranialbase variation so far have used externallandmarks and/or two-dimensional radio-graphic analysis of midsagittal landmarks(or nonmidsagittal landmarks projected intothe midsagittal plane). Second, more dataare needed on the developmental mecha-nisms which generate variation in the cra-nial base, and which regulate interactionsamong components of the skull. Experimen-tal, histological, and other kinds of develop-mental information will be useful in thisregard, because observed morphologicalpatterns are potentially generated by differ-ent genetic and epigenetic processes. Third,more data are needed on the genetic basesfor variation in cranial base growth andform. Lastly, more detailed interspecificanalyses are needed to extend the evolution-ary implications of ontogenetic tests of hy-potheses about interactions and relation-ships among morphogenetic units of theskull. Such future research on the cranialbase should provide interesting and valu-able insights on other aspects of craniofacial
growth, function, and evolution in primatesand other mammals.
ACKNOWLEDGMENTS
We thank N. Jeffery, W. Jungers, R. Mc-Carthy, C. Ruff, B. Shea, F. Spoor, and twoanonymous reviewers for their commentsand assistance.
LITERATURE CITED
Adams LM, Moore WJ. 1975. A biomechanical appraisalof some skeletal features associated with head bal-ance and posture in the Hominoidea. Acta Anat(Basel) 92:580–594.
Aiello L, Dean C. 1990. An introduction to human evo-lutionary anatomy. San Diego: Academic Press.
Alberch P. 1985. Developmental constraints: why St.Bernards often have an extra digit and poodles neverdo. Am Nat 126:430–433.
Allman J. 1977. Evolution of the visual system in earlyprimates. In: Sprague JM, Epstein JM, editors.Progress in psychobiology, physiology, and psychol-ogy. Volume 7. New York: Academic Press. p 1–53.
Allman J. 1982. Reconstructing the evolution of thebrain in primates through the use of comparativeneurophysiological and neuroanatomical data. In:Armstrong E, Falk D, editors. Primate brain evolu-tion. New York: Plenum Press. p 13–28.
Allman JM, Kaas JH. 1974. The organization of thesecond visual area (V II) in the owl monkey: a secondorder transformation of the visual hemifield. BrainRes 76:247–265.
Anagastopolou S, Karamaliki DD, Spyropoulos MN.1988. Observations on the growth and orientation ofthe anterior cranial base in the human foetus. EurJ Orthod 10:143–148.
Angst R. 1967. Beitrag zum formwandel des craniumsder ponginen. Z Morphol Anthropol 58:109–151.
Anton SC. 1989. Intentional cranial vault deformationand induced changes in the cranial base and face.Am J Phys Anthropol 79:253–267.
Anton SC. 1994. Mechanical and other perspectives onNeandertal craniofacial morphology. In: CorrucciniRS, Ciochon RL, editors. Integrative paths to thepast: palaeoanthropological advances in honor of F.Clark Howell. Englewood Cliffs, NJ: Prentice Hall. p677–695.
Ashton EH. 1952. Age changes in the basicranial axis ofthe anthropoidea. Proc Zool Soc Lond [Biol] 129:61–74.
Ashton EH, Spence TF. 1958. Age changes in the cra-nial capacity and foramen magnum of hominoids.Proc Zool Soc Lond [Biol] 130:169–181.
Ashton EH, Zuckerman S. 1952. Age changes in theposition of the occipital condyles in the chimpanzeeand gorilla. Am J Phys Anthropol 10:277–288.
Ashton EH, Zuckerman S. 1956. Age changes in theposition of the foramen magnum in hominoids. ProcZool Soc Lond [Biol] 126:315–325.
Atchley WR, Hall BK. 1991. A model for developmentand evolution of complex morphological structures.Biol Rev 66:101–157.
Babler WJ. 1989. Relationship of altered cranial suturegrowth to the cranial base and midface. In: PersingJA, Edgerton MT, Jane JA, editors. Scientific founda-tions and surgical treatment of craniosynostosis. Bal-timore: Williams and Wilkins. p 87–95.
Baer T, Gore JC, Gracco LC, Nye PW. 1991. Analysis ofvocal tract shape and dimensions using magnetic res-
PRIMATE CRANIAL BASE 163D.E. Lieberman et al.]

onance imaging: vowels. J Acoust Soc Am 90:799–828.
Barlow HB. 1986. Why have multiple cortical areas?Vision Res 26:81–90.
Barton RA. 1998. Visual specialization and brain evo-lution in primates. Proc R Soc Lond [Biol] 265:1933–1937.
Baume JL. 1957. A biologist looks at the sella point. EurOrthod Soc Trans 33:150–159.
Beard KC. 1993. Phylogenetic systematics of the Pri-matomorpha, with special reference to Dermoptera.In: Szalay FS, Novacek MJ, McKenna MC, editors.Mammal phylogeny: placentals. New York: Springer-Verlag. p 129–150.
Beckman ME, Jung T-P, Lee S-H, de Jong K, Krish-namurthy AK, Ahalt SC, Cohen KB, Collins MJ.1995. Variability in the production of quantal vowelsrevisited. J Acoust Soc Am 97:471–489.
Beechey C, Boyd Y, Searle AG. 1998. Brachyrrhine, Br,a mouse craniofacial mutant maps to distal mousechromosome 17 and is a candidate model for midlinecleft syndrome. Mouse Genome 95:692–694.
Biegert J. 1957. Der Formwandel der Primatenschadelsund seine Beziehungen zur ontogenetischen Entwick-lung und den phylogenetischen. Gegenbaurs MorpholJahrb 98:77–199.
Biegert J. 1963. The evaluation of characteristics of theskull, hands and feet for primate taxonomy. In: Wash-burn SL, editor. Classification and human evolution.Chicago: Aldine. p 116–145.
Bjork A. 1955. Cranial base development. Am J Orthod41:198–225.
Bolk L. 1909. On the position and displacement of theforamen magnum in the Primates. Verh Akad WetAmst 12:362–377.
Bolk L. 1910. On the slope of the foramen magnum inPrimates. Verh Akad Wet Amst 12:525–534.
Bolk L. 1926. Das Problem der Menschwerdung. Jena:Gustav Fischer.
Bookstein F. 1991. Morphometric tools for landmarkdata: geometry and biology. New York: John Wileyand Sons.
Bosma JF, editor. 1976. Development of the basicra-nium. Bethesda, MD: National Institutes of Health.
Braga J, Crubezy E, Elyaqtine M. 1999. The posteriorborder of the sphenoid greater wing and its phyloge-netic usefulness in human evolution. Am J Phys An-thropol 107:387–399.
Brodie AG. 1941. On the growth pattern of the humanhead from the third month to the eighth year of life.Am J Anat 68:208–262.
Brodie AG. 1953. Late growth changes in the humanface. Angle Orthod 23:146–157.
Bromage TG. 1992. The ontogeny of Pan troglodytescraniofacial architectural relationships and implica-tions for early hominids. J Hum Evol 23:235–251.
Butler AB, Hodos W. 1996. Comparative vertebrateneuroanatomy, evolution and adaptation. New York:Wiley-Liss.
Butow K-W. 1990. Craniofacial growth disturbance af-ter skull base and associated suture synostoses in thenewborn Chacma baboon: a preliminary report. CleftPalate J 27:241–252.
Cameron J. 1924. The cranio-facial axis of Huxley. PartI. Embryological considerations. Trans R Soc Can 18:115–123.
Cameron J. 1927. The main angle of cranial flexion (thenasion-pituitary-basion angle). Am J Phys Anthropol10:275–279.
Cameron J. 1930. Craniometric memoirs. No. II. Thehuman and comparative anatomy of Cameron’scraniofacial axis. J Anat 64:324–336.
Carre R, Lindblom B, MacNeilage P. 1994. Acousticfactors in the evolution of the human vocal tract. JAcoust Soc Am 95:2924.
Cartmill M. 1970. The orbits of arboreal mammals.Ph.D. dissertation, University of Chicago.
Cartmill M. 1972. Arboreal adaptations and the originof the order Primates. In: Tuttle RH, editor. The func-tional and evolutionary biology of primates. Chicago:Aldine. p 97–122.
Cartmill M. 1974. Rethinking primate origins. Science184:436–443.
Cartmill M. 1980. Morphology, function, and evolutionof the anthropoid postorbital septum. In: Ciochon RLChiarelli AB, editors. Evolutionary biology of the NewWorld monkeys and continental drift. New York: Ple-num Press. p 243–274.
Cartmill M. 1992. New views on primate origins. EvolAnthropol 1:105–111.
Chan L-K. 1991. The deduction of the level of the hu-man larynx from bony landmarks: its relevance to theevolution of speech. Hum Evol 6:249–261.
Cherniak C. 1995. Neural component placement.Trends Neurosci 18:522–527.
Chernoff B, Magwene P. 1999. Morphological integra-tion: forty years later. In: Olson EC, Miller RL, edi-tors. Morphological integration. Chicago: Universityof Chicago Press. p 319–348.
Cheverud JM. 1982. Phenotypic, genetic, and environ-mental morphological integration in the cranium.Evolution 36:499–516.
Cheverud JM. 1995. Morphological integration in thesaddle back tamarin (Saguinas fuscicollis) cranium.Am Nat 145:63–89.
Cheverud JM. 1996. Developmental integration and theevolution of pleiotropy. Am Zool 36:44–50.
Cheverud JM, Buikstra J. 1982. Quantitative geneticsof skeletal non-metric traits in the rhesus macaqueson Cayo Santiago III. Relative heritability of skeletalmetric and non-metric traits. Am J Phys Anthropol59:151–155.
Cheverud JM, Richtsmeier J. 1986. Finite element scal-ing applied to sexual dimorphism in rhesus macaque(Macaca mulatta) facial growth. Syst Zool 35:381–399.
Cheverud JM, Dow MM, Leutenegger W. 1985. Thequantitative assessment of phylogenetic constraintsin comparative analyses: sexual dimorphism in bodyweight among primates. Evolution 39:1335–1351.
Cheverud JM, Wagner GP, Dow MM. 1989. Methods forthe comparative analysis of variation patterns. SystZool 38:201–213.
Cheverud JM, Kohn LAP, Konigsberg LW, Leigh SR.1992. The effects of fronto-occipital artifical cranialvault modification on the cranial base and face. Am JPhys Anthropol 88:323–345.
Couly GF, Coltey PM, Le Douarin NM. 1993. The tripleorigin of skull in higher vertebrates. Development117:409–429.
Cousin RP, Fenart R, Deblock R. 1981. Variations on-togenetiques des angles basicraniens et faciaux. BullMem Soc Anthropol Paris 8:189–212.
Covert HH, Hamrick MW. 1993. Description of newskeletal remains of the early Eocene anaptomorphineprimate Absarokius (Omomyidae) and a discussionabout its adaptive profile. J Hum Evol 25:351–362.
Cramer DL. 1977. Craniofacial morphology of Pan pa-niscus: a morphometric and evolutionary appraisal.Contrib Primatol 10:1–64.
Crelin E. 1969. Anatomy of the newborn: an atlas. Phil-adelphia: Lea and Febiger.
Crelin ES. 1973. Functional anatomy of the newborn:an atlas. New Haven: Yale University Press.
164 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000

Crompton AW, German RZ, Thexton AJ. 1997. Mecha-nisms of swallowing and airway protection in infantmammals (Sus domesticus and Macaca fascicularis).J Zool Lond 241:89–102.
Dabelow A. 1929. Uber Korrelationenin der phyloge-netischen Entwicklung der Schadelform. GegenbaursMorphol Jahrb 63:1–49.
Dabelow A. 1931. Uber Korrelationenin der phyloge-netischen Entwicklung der Schadelform. II. Die Bez-iehungen zwischen Gehirn und Schadelbasisform beiden Mammaliern. Gegenbaurs Morphol Jahrb 67:84–133.
Dagosto M. 1988. Implications of postcranial evidencefor the origin of euprimates. J Hum Evol 17:35–56.
David DJ, Hemmy DC, Cooter RD. 1989. Craniofacialdeformities. New York: Springer Verlag.
Dean MC. 1988. Growth processes in the cranial base ofhominoids and their bearing on the morphologicalsimilarities that exist in the cranial base of Homo andParanthropus. In: Grine FE, editor. Evolutionary his-tory of the “robust” Australopithecines. New York:Aldine. p 107–112.
Dean MC, Wood BA. 1981. Metrical analysis of thebasicranium of extant hominoids and Australopithe-cus. Am J Phys Anthropol 59:53–71.
Dean MC, Wood BA. 1982. Basicranial anatomy of Plio-Pleistocene hominids from East and South Africa.Am J Phys Anthropol 59:157–174.
de Beer GR. 1937. The development of the vertebrateskull. Oxford: Oxford University Press.
Delattre A, Fenart R. 1956. Le developpement du cranedu gorille et du chimpanze compare au developpe-ment du crane humain. Bull Mem Soc AnthropolParis 6:159–173.
Delattre A, Fenart R. 1963. Etudes des projections hori-zontale et vertico-frontales du crane au cours del’hominisation. Anthropologie 67:525–561.
Demes AB. 1985. Biomechanics of the primate skullbase. Advances in anatomy, embryology and cell biol-ogy. Volume 94. Berlin: Springer-Verlag.
Diewert VM. 1985. Development of human craniofacialmorphology during the late embryonic and early fetalperiods. Am J Orthod 88:64–76.
Dmoch R. 1975. Beitrage zum formenwandel des prima-tencraniums mit bemerkungen zu den sagittalenknickungsverhaltnissen. II Streckenanalyse, I, Neu-rocranium. Gegenbaurs Morphol Jahrb 121:521–601.
DuBrul EL. 1950. Posture, locomotion and the skull inLagomorpha. Am J Anat 87:277–313.
DuBrul EL. 1977. Early hominid feeding mechanisms.Am J Phys Anthropol 47:305–320.
DuBrul EL. 1979. Origin and adaptations of the homi-nid jaw joint. In: Sarnat BG, Laskin DM, editors. Thetemporomandibular jaw joint. 3rd ed. Springfield, IL:Thomas. p 5–34.
DuBrul EL, Laskin DM. 1961. Preadaptive potentiali-ties of the mammalian skull: an experiment in growthand form. Am J Anat 109:117–132.
Duckworth WLH. 1904. Studies from the anthropologi-cal laboratory. Cambridge: Cambridge UniversityPress.
Duckworth WLH. 1915. Morphology and anthropology.2nd ed. Cambridge: Cambridge University Press.
Duterloo HS, Enlow DH. 1970. A comparative study ofcranial growth in Homo and Macaca. Am J Anat127:357–368.
Enlow DH. 1976. Postnatal growth and development ofthe face and cranium. In: Cohen B, Kramer IRV,editors. Scientific foundations of dentistry. London:Heinemann. p 29–46.
Enlow DH. 1990. Facial growth. 3rd ed. Philadelphia:Saunders.
Enlow DH, Azuma M. 1975. Functional growth bound-aries in the human and mammalian face. In: BergsmaD, editor. Morphogenesis and malformation of faceand brain. New York: A.R. Liss. p 217–230.
Enlow DH, Bhatt MK. 1984. Facial morphology varia-tions associated with head form variations. J CharlesTweed Found 12:21–23.
Enlow DH, Hans MG. 1996. Essentials of facial growth.Philadelphia: W.B. Saunders.
Fant G. 1960. Acoustic theory of speech production. TheHague: Mouton.
Felsenstein J. 1985. Phylogenies and the comparativemethod. Am Nat 125:1–15.
Fenart R. 1966. Changements morphologiques del’encephale, chez le rat ampute des membres anter-ieurs. J Hirnforsch 8:493–501.
Fenart R, Pellerin C. 1988. The vestibular orientationmethod: its application in the determination of anaverage human skull type. Int J Anthropol 3:223–239.
Fleagle JG. 1999. Primate adaptation and evolution.2nd ed. New York: Academic Press.
Ford EH. 1956. The growth of the foetal skull. J Anat90:63–72.
Ford EH. 1958. Growth of the human cranial base. Am JOrthod 44:498–506.
George SL. 1978. A longitudinal and cross-sectionalanalysis of the growth of the postnatal cranial baseangle. Am J Phys Anthropol 49:171–178.
German RZ, Crompton AW, McCluskey C, Thexton AJ.1996. Coordination between respiration and degluti-tion in a preterm infant mammal, Sus scrofa. ArchOral Biol 41:619–622.
Giles WB, Philips CL, Joondeph DR. 1981. Growth inthe basicranial synchondroses of adolescent Macacamullata. Anat Rec 19:259–266.
Gould SJ. 1975. Allometry in primates, with emphasison scaling and the evolution of the brain. ContribPrimatol 5:244–292.
Gould SJ. 1977. Ontogeny and phylogeny. Cambridge:Belknap Press.
Graf W. 1988. Motion detection in physical space and itsperipheral and central representation. Ann NY AcadSci 545:154–169.
Graf W, deWaele C, Vidal PP, Wang DH, Evinger C.1995. The orientation of the cervical vertebral columnin unrestrained awake animals. II. Movement strat-egies. Brain Behav Evol 45:209–231.
Hall BK, editor. 1994. Homology: the hierarchical basisof comparative biology. New York: Academic Press.
Hamrick MW. 1998. Functional and adaptive signifi-cance of primate pads and claws: evidence from NewWorld anthropoids. Am J Phys Anthropol 106:113–127.
Hamrick MW. 1999. Pattern and process in the evolu-tion of primate nails and claws. J Hum Evol 37:293–297.
Heintz N. 1966. Le crane des anthropomorphes: crois-sance relative, variabilite, evolution. Musee Royale del’Afrique Centrale-Tervuren. Ann Sci Zool 6:1–122.
Herring SW. 1993. Epigenetic and functional influenceson skull growth. In: Hanken J, Hall BK, editors. Theskull. Volume 1. Chicago: University of ChicagoPress. p 153–206.
Hofer H. 1960. Studien zum Problem des Gestaltwan-dels des Schadels der Saugetiere, insbesondere derPrimaten. I. Die medianen Kruemmundgen desSchadels und ihr Ehrfassung nach Landzert. Z Mor-phol Anthropol 50:299–316.
Hofer H. 1965. Die morphologische Analyse desSchadels des Menschen. In: Hereberer G, editor.
PRIMATE CRANIAL BASE 165D.E. Lieberman et al.]

Menschliche Abstammungslehre. Stuttgart: G. Fi-scher. p 145–226.
Hofer H. 1969. On the evolution of the craniocerebraltopography in primates. Ann NY Acad Sci 162:341–356.
Hofer H, Spatz W. 1963. Studien zum Problem desGestaltwandels des Schadels der Saugetiere, insbe-sondere der Primaten. II. Uber die kyphosen fetalerund neonater Primatenschadel. Z Morphol Anthropol53:29–52.
Hooton EA. 1946. Up from the ape. 3rd ed. New York:Macmillan.
Howells WW. 1973. Cranial variation in man. Cam-bridge: Peabody Museum Papers, no. 67.
Hoyte DA. 1991. The cranial base in normal and abnor-mal growth. Neurosurg Clin North Am 2:515–537.
Huxley TH. 1867. On two widely contrasted forms of thehuman cranium. J Anat Physiol 1:60–77.
Hylander WL, Ravosa MJ. 1992. An analysis of thesupraorbital region of primates: a morphometric andexperimental approach. In: Smith P, Tchernov E, ed-itors. Structure, function and evolution of teeth. Lon-don: Freund. p 223–255.
Hylander WL, Ravosa MJ, Ross CF, Johnson KR. 1998.Mandibular corpus strain in primates: further evi-dence for a functional link between symphyseal fu-sion and jaw-adductor muscle force. Am J Phys An-thropol 107:257–271.
Hylander WL, Ravosa MJ, Ross CF, Wall CE, JohnsonKR. 2000. Symphyseal fusion and jaw-adductor mus-cle force: an EMG study. Am J Phys Anthropol 112:469–492.
Jeffery N. 1999. Fetal development and evolution of thehuman cranial base. Ph.D. dissertation, UniversityCollege, London.
Jerison HJ. 1982. Allometry, brain size and convoluted-ness. In: Armstrong E, Falk D, editors. Primate brainevolution. Methods and concepts. New York: PlenumPress. p 77–84.
Jolicoeur P, Mossiman J. 1968. Intervalles de confi-dance pour la peute de l’axe majeur d’une distributionnormale bidimensionelle. Biometrie Praximetrie9:121–140.
Kandel ER, Schwartz JH, Jessell TM. 1991. Principlesof neural science. 3rd ed. Norwalk, CT: Apple andLang.
Kay RF, Cartmill M. 1974. Skull of Palaechthon nacimi-enti. Nature 252:37–38.
Kay RF, Cartmill M. 1977. Cranial morphology andadaptations of Palaechthon nacimienti and otherParomomyidae (Plesiadapoidea, ?Primates), with adescription of a new genus and species. J Hum Evol6:19–35.
Kjaer I. 1990. Ossification of the human basicranium. JCraniofac Genet Dev Biol 10:29–38.
Kodama G. 1976a. Developmental studies of the pres-phenoid of the human sphenoid bone. In: Bosma JF,editor. Development of the basicranium. Bethesda,MD: National Institutes of Health. p 141–155.
Kodama G. 1976b. Developmental studies of the body ofthe human sphenoid bone. In: Bosma JF, editor. De-velopment of the basicranium. Bethesda, MD: Na-tional Institutes of Health. p. 156–165.
Kodama G. 1976c. Developmental studies of the orbito-sphenoid of the human sphenoid bone. In: Bosma JF,editor. Development of the basicranium. Bethesda,MD: National Institutes of Health. p 166–176.
Kohn LAP, Leigh SR, Jacobs SC, Cheverud JM. 1993.Effects of annular cranial vault modification on thecranial base and face. Am J Phys Anthropol 90:147–168.
Krogman WM. 1969. Growth changes in the skull, face,jaws and teeth of the chimpanzee. In: Bourne GH,editor. The chimpanzee. Volume 1. Basel: S. Karger. p104–164.
Lager H. 1958. A histological description of the cranialbase in Macaca rhesus. Eur Orthod Soc Trans 34:147–156.
Laitman JT, Heimbuch RC. 1982. The basicranium ofPlio-Pleistocene hominids as an indicator of their up-per respiratory systems. Am J Phys Anthropol 59:323–343.
Laitman JT, Reidenberg JS. 1993. Specializations of thehuman upper respiratory and upper digestive sys-tems as seen through comparative and developmentalanatomy. Dysphagia 8:318–325.
Laitman JT, Heimbuch RC, Crelin ES. 1978. Develop-mental change in a basicranial line and its relation-ship to the upper respiratory system in living pri-mates. Am J Anat 152:467–483.
Laitman JT, Heimbuch RC, Crelin ES. 1979. The basi-cranium of fossil hominids as an indicator of theirupper respiratory systems. Am J Phys Anthropol 51:15–34.
Landzert T. 1866. Der Sattelwinkel und sein Verhaelt-nis zur Pro- und Orthognathie. Abh SenckenbergNaturforsch Ges 6:19–165.
Langille RM, Hall BK. 1993. Pattern formation and theneural crest. In: Hanken J, Hall BK, editors. Theskull. Volume 1. Chicago: University of ChicagoPress. p 77–111.
Larson JE, Herring SW. 1996. Movement of the epiglot-tis in mammals. Am J Phys Anthropol 100:71–82.
Latham RA. 1972. The sella point and postnatal growthof the human cranial base. Am J Orthod 61:156–162.
Le Douarin NM, Ziller C, Couly GF. 1993. Patterning ofneural crest derivatives in the avian embryo: in vivoand in vitro studies. Dev Biol 159:24–49.
Lele S. 1993. Euclidean distance matrix analysis(EDMA): estimation of mean form and mean formdifference. Math Geol 25:573–602.
Lemelin P. 1999. Morphological correlates of substrateuse in didelphid marsupials: implications for primateorigins. J Zool Lond 247:165–175.
Lieberman DE. 1995. Testing hypotheses about recenthuman evolution using skulls: integrating morphol-ogy, function, development, and phylogeny. Curr An-thropol 36:159–197.
Lieberman DE. 1997. Making behavioral and phyloge-netic inferences from fossils: considering the develop-mental influence of mechanical forces. Annu Rev An-thropol 26:185–210.
Lieberman DE. 1998. Sphenoid shortening and the evo-lution of modern human cranial shape. Nature 393:158–162.
Lieberman DE. 1999. Homology and hominid phylog-eny: problems and potential solution. Evol Anthropol7:142–151.
Lieberman DE. 2000. Ontogeny, homology, and phylog-eny in the hominid craniofacial skeleton: the problemof the browridge. In: O’Higgins P, Cohn M, editors.Development, growth and evolution: implications forthe study of hominid skeletal evolution. London: Ac-ademic Press. p 85–122.
Lieberman DE, McCarthy RC. 1999. The ontogeny ofcranial base angulation in humans and chimpanzeesand its implications for reconstructing pharyngealdimensions. J Hum Evol 36:487–517.
Lieberman DE, Pilbeam DR, Wood BA. 1996. Ho-moplasy and early Homo: an analysis of the evolu-tionary relationships of H. habilis sensu stricto andH. rudolfensis. J Hum Evol 30:97–120.
166 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000

Lieberman DE, McCarthy RC, Hiiemae K, LiebermanP, Palmer JB. 1998. New estimates of fossil hominidvocal tract dimension. J Hum Evol 34:12–13.
Lieberman DE, Pearson OM, Mowbray KM. 2000. Ba-sicranial influence on overall cranial shape. J HumEvol 38:291–315.
Lieberman P. 1984. The biology and evolution of lan-guage. Cambridge, MA: Harvard University Press.
Lieberman P, Crelin ES. 1971. On the speech of Nean-derthal man. Ling Inq 1:203–223.
Lieberman P, Crelin ES, Klatt DH. 1972. Phonetic abil-ity and related anatomy of the newborn and adulthuman, Neanderthal man, and the chimpanzee. AmAnthropol 74:287–307.
Lieberman P, Laitman JT, Reidenberg JS, Gannon PJ.1992. The anatomy, physiology, acoustics and percep-tion of speech: essential elements in analysis of theevolution of human speech. J Hum Evol 23:447–467.
Lozanoff S. 1993. Midfacial retrusion in adult Brachyr-rhine mice. Acta Anat (Basel) 147:125–132.
Lozanoff S, Jureczek S, Feng T, Padwal R. 1994. Ante-rior cranial base morphology in mice with midfacialretrusion. Cleft Palate Craniofac J 31:417–428.
Lufkin T, Mark M, Hart CP, Dolle P, LeMeur M, Cham-bon P. 1992. Homeotic transformation of the occipitalbones by ectopic expression of a homeobox gene. Na-ture 359:835–841.
Lugoba SA, Wood BA. 1990. Position and orientation ofthe foramen magnum in higher primates. Am J PhysAnthropol 81:67–76.
Ma W, Lozanoff S. 1999. Spatial and temporal distribu-tion of cellular proliferation in the anterior cranialbase of normal midfacially retrusive mice. Clin Anat12:315–325.
Martin RD. 1990. Primate origins and evolution.Princeton: Princeton University Press.
May R, Sheffer DB. 1999. Growth changes in measure-ments of upper facial positioning. Am J Phys An-thropol 108:269–280.
Maynard Smith J, Burian R, Kauffman S, Alberch P,Campbell J, Goodwin B, Lande R, Raup D, Wolpert L.1985. Developmental constraints and evolution. QRev Biol 60:265–287.
McCarthy R. 2001. Anthropoid cranial base architec-ture and scaling relationships. J Hum Evol (in press).
McCarthy RC, Lieberman DE. 2001. The posterior max-illary (PM) plane and anterior cranial architecture inprimates. Anat Rec (in press).
Melsen B. 1969. Time of closure of the spheno-occipitalsynchondrosis determined on dry skulls. A radio-graphic craniometric study. Acta Odontol Scand 27:73–90.
Melsen B. 1971. The postnatal growth of the cranialbase in Macaca rhesus analyzed by the implantmethod. Tandlaegebladet 75:1320–1329.
Michejda M. 1971. Ontogenetic changes of the cranialbase in Macaca mulatta. In: Biegert J, LeuteneggerW, editors. Proceedings of the Third InternationalCongress of Primatology Zurich 1970. Basel: Karger.p 215–225.
Michejda M. 1972a. The role of basicranial synchondro-ses in flexure processes and ontogenetic developmentof the skull base. Am J Phys Anthropol 37:143–150.
Michejda M. 1972b. Significance of basicranial synchon-droses in nonhuman primates and man. Proc 3rdConf Exp Med Surg Primates, Lyon 1:372–378.
Michejda M, Lamey D. 1971. Flexion and metric agechanges of the cranial base in Macaca mulatta. In-fants and juveniles. Folia Primatol (Basel) 34:133–141.
Mitchison G. 1991. Neuronal branching patterns andthe economy of cortical wiring. Proc R Soc Lond [Biol]245:151–158.
Moore WJ, Lavelle CJB. 1974. Growth of the facialskeleton in the Hominoidea. London: Academic Press.
Moore WJ, Adams LM, Lavelle CLB. 1973. Head pos-ture in the Hominoidea. J Zool Lond 169:409–416.
Moss ML. 1958. The pathogenesis of artificial cranialdeformation. Am J Phys Anthropol 16:269–286.
Moss ML. 1961. Rotation of the otic capsule in bipedalrats. Am J Phys Anthropol 19:301–307.
Moss ML. 1963. Morphological variations of the cristagalli and medial orbital margin. Am J Phys Anthropol21:259–264.
Moss ML. 1976. Experimental alteration of basi-syn-chondroseal cartilage growth in rat and mouse. In:Bosma JF, editor. Development of the basicranium.Bethesda, MD: National Institutes of Health. p 541–569.
Moss ML. 1997. The functional matrix hypothesis revis-ited. 1. The role of mechanotransduction. Am JOrthod Dentofacial Orthop 112:8–11.
Moss ML, Young RW. 1960. A functional approach tocraniology. Am J Phys Anthropol 18:281–292.
Moss ML, Moss-Salentjin L, Vilmann H, Newell-MorrisL. 1982. Neuro-skeletal topology of the primate basi-cranium: its implications for the “fetalization hypoth-esis.” Gegenbaurs Morphol Jahrb 128:58–67.
Nearey T. 1978. Phonetic features for vowels. Blooming-ton: Indiana University Linguistics Club.
Negus V. 1949. The comparative anatomy and physiol-ogy of the larynx. New York: Hafner.
Noble VE, Kowalski EM, Ravosa MJ. 2000. Orbital ori-entation and the function of the mammalian postor-bital bar. J Zool Lond 250:405–418.
Noden DM. 1991. Cell movements and control of pat-terned tissue assembly during craniofacial develop-ment. J Craniofac Genet Dev Biol 11:192–213.
Nunn CL. 1995. A simulation test of Smith’s “degress offreedom” correction for comparative studies. Am JPhys Anthropol 98:355–367.
O’Higgins P. 2000. Quantitative approaches to thestudy of craniofacial growth and evolution: advancesin morphometric techniques. In: O’Higgins P, CohnM, editors. Development, growth and evolution: im-plications for the study of hominid skeletal evolution.London: Academic Press. p 163–185.
Olson E, Miller R. 1958. Morphological integration. Chi-cago: University of Chicago Press.
Olson TR. 1985. Cranial morphology and systematics ofthe Hadar hominids and “Australopithecus” africa-nus. In: Delson E, editor. Ancestors: the hard evi-dence. New York: A.R. Liss. p 102–119.
O’Rahilly R, Muller F. 1999. The embryonic humanbrain: an atlas of developmental stages. 2nd ed. NewYork: Wiley-Liss.
Purvis A, Rambaut A. 1995. Comparative analysis byindependent contrasts (CAIC): an Apple Macintoshapplication for analyzing comparative data. CompAppl Biosci 11:247–251.
Radinsky LB. 1968. A new approach to mammaliancranial analysis, illustrated by examples of prosimianprimates. J Morphol 124:167–179.
Radinsky LB. 1970. The fossil record of prosimian brainevolution. In: Noback C, Montagna W, editors. Theprimate brain. Advances in primatology. Volume 1.New York: Appleton-Century-Crofts. p 209–224.
Radinsky LB. 1977. Early primate brains: facts andfiction. J Hum Evol 6:79–86.
Radinsky LB. 1979. The fossil evidence of primate brainevolution (James Arthur Lecture). New York: Amer-ican Museum of Natural History.
PRIMATE CRANIAL BASE 167D.E. Lieberman et al.]

Ranke J. 1892. Uber einige gesetzmassige Beziehungenzwischen Schadelgrund, Gehirn- und Gesichts-schadel. Beitrage Anthropol Urgesch Bayerns 12:1–132.
Ravosa MJ. 1988. Browridge development in Cerco-pithecidae: a test of two models. Am J Phys Anthropol76:535—555.
Ravosa MJ. 1991a. Ontogenetic perspective on mechan-ical and nonmechanical models of primate circumor-bital morphology. Am J Phys Anthropol 85:95–112.
Ravosa MJ. 1991b. Interspecific perspective on mechan-ical and nonmechanical models of primate circumor-bital morphology. Am J Phys Anthropol 86:369–396.
Ravosa MJ. 1996. Mandibular form and function inNorth American and European Adapidae and Omo-myidae. J Morphol 229:171–190.
Ravosa MJ. 1999. Anthropoid origins and the modernsymphysis. Folia Primatol (Basel) 70:65–78.
Ravosa MJ, Shea BT. 1994. Pattern in craniofacial bi-ology: evidence from the Old World monkeys (Cerco-pithecidae). Int J Primatol 15:801–822.
Ravosa MJ, Noble VE, Hylander WL, Johnson KR, Ko-walski EM. 2000a. Masticatory stress, orbital orien-tation and the evolution of the primate postorbitalbar. J Hum Evol 38:667–693.
Ravosa MJ, Vinyard CJ, Hylander WL. 2000b. Stressedout: masticatory forces and primate circumorbitalform. Anat Rec (New Anat) 261:(in press).
Reidenberg JS, Laitman JT. 1991. Effect of basicranialflexion on larynx and hyoid position in rats: an exper-imental study of skull and soft tissue interactions.Anat Rec 230:557–569.
Riesenfeld A. 1966. The effects of experimental bipedal-ism and upright posture in the rat and their significancefor human evolution. Acta Anat (Basel) 65:449–521.
Riesenfeld A. 1969. The adaptive mandible: an experi-mental study. Acta Anat (Basel) 72:246–262.
Riolo ML, Moyers RE, McNamara JA Jr, Hunter WS.1974. An atlas of craniofacial growth: cephalometricstandards from the University School Growth Study,the University of Michigan. Ann Arbor: Center forHuman Growth and Development, monograph no. 2.
Robinson GW, Mahon KA. 1994. Differential and over-lapping expression domains of DLx2 and Dlx3 suggestdistinct roles for distal-less genes in craniofacial de-velopment. Mech Dev 48:199–215.
Rosenberger AL. 1985. In favor of the Necrolemur-Tarsier hypothesis. Folia Primatol (Basel) 45:179–194.
Rosenberger AL. 1986. Platyrrhines, catarrhines andthe anthropoid transition. In: Wood BA, Martin L,Andrews P, editors. Major topics in primate and hu-man evolution. Cambridge: Cambridge UniversityPress. p 66–88.
Ross CF. 1994. Craniofacial evidence for anthropoid andtarsier relationships. In: Fleagle JG, Kay RF, editors.Anthropoid origins. New York: Plenum Press. p 469–547.
Ross CF. 1995a. Allometric and functional influences onprimate orbit orientation and the origins of the An-thropoidea. J Hum Evol 29:201–227.
Ross CF. 1995b. Muscular and osseous anatomy of theprimate anterior temporal fossa and the functions ofthe postorbital septum. Am J Phys Anthropol 98:275–306.
Ross CF. 1996. An adaptive explanation for the origin ofthe Anthropoidea (Primates). Am J Primatol 40:205–230.
Ross CF. 2000. Into the light: the origin of Anthro-poidea. Annu Rev Anthropol 29:147–194.
Ross CF, Henneberg M. 1995. Basicranial flexion, rela-tive brain size and facial kyphosis in Homo sapiensand some fossil hominids. Am J Phys Anthropol 98:575–593.
Ross CF, Ravosa MJ. 1993. Basicranial flexion, relativebrain size and facial kyphosis in nonhuman primates.Am J Phys Anthropol 91:305–324.
Ross CF, Williams BA, Kay RF. 1998. Phylogeneticanalysis of anthropoid relationships. J Hum Evol 35:221–306.
Ruff CB, Trinkaus E, Holliday TW. 1997. Body massand encephalization in Pleistocene Homo. Nature387:173–176.
Sakka M, Aaron C, Herve D. 1976. Etude anatomique,fonctionelle et radiologique de l’ensemble cervico-cephalique dorsal du Chien. Applications aux prob-lemes du port de tete en station erigree chez les Homi-nides. Zbl Vet Med C 5:1420.
Sarnat BG. 1982. Eye and orbital size in the young andadult. Some postnatal experimental and clinical rela-tionships. Ophthalmology 185:74–89.
Sarnat BG. 1988. Craniofacial change and non-changeafter experimental surgery in young and adult ani-mals. Angle Orthod 58:321–342.
Sarnat BG. 1999. Basic science and clinical experimen-tal primate studies in craniofaciodental biology: apersonal historical review. J Oral Maxillofac Surg57:714–724.
Sasaki H, Kodama G. 1976. Developmental studies ofthe postsphenoid of the human sphenoid bone. In:Bosma JF, editor. Development of the basicranium.Bethesda, MD: National Institutes of Health. p. 177–190.
Schilling TF, Thorogood PV. 2000. Development andevolution of the vertebrate skull. In: O’Higgins P,Cohn M, editors. Development, growth and evolution:implications for the study of hominid skeletal evolu-tion. London: Academic Press. p 57–83.
Schneiderman ED. 1992. Facial growth in the rhesusmonkey: a longitudinal and cephalometric study.Princeton: Princeton University Press.
Schultz AH. 1940. The size of the orbit and of the eye inprimates. Am J Phys Anthropol 26:398–408.
Schultz AH. 1942. Conditions for balancing the head inprimates. Am J Phys Anthropol 29:483–497.
Schultz AH. 1955. The position of the occipital condylesand of the face relative to the skull base in primates.Am J Phys Anthropol 13:97–120.
Scott JH. 1958. The cranial base. Am J Phys Anthropol16:319–348.
Shapiro R. 1960. The normal skull: a Roentgen study.New York: Hoeber.
Shapiro R, Robinson F. 1980. The embryogenesis of thehuman skull. Cambridge: Harvard University Press.
Shea BT. 1985a. On aspects of skull form in Africanapes and orangutans, with implications for hominoidevolution. Am J Phys Anthropol 68:329–342.
Shea BT. 1985b. Ontogenetic allometry and scaling: adiscussion based on the growth and form of the skullin African apes. In: Jungers WL, editor. Size andscaling in primate biology. New York: Plenum Press.p 175–205.
Shea BT. 1986. On skull form and the supraorbitaltorus in primates. Curr Anthropol 27:257–259.
Shea BT. 1987. Reproductive strategies, body size, andencephalization in primate evolution. Int J Primatol8:139–156.
Shea BT. 1988. Phylogeny and skull form in the homi-noid primates. In: Schwartz JH, editor. The biology ofthe orangutan. New York: Oxford University Press. p233–246.
Simons EL. 1962. Fossil evidence relating to the earlyevolution of primate behavior. Ann NY Acad Sci 102:282–294.
Sirianni J. 1985. Nonhuman primates as models forhuman craniofacial growth. In: Watt ES, editor. Non-
168 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 43, 2000

human primate models for human growth and devel-opment. New York: A.R. Liss. p 95–124.
Sirianni J, Newell-Morris L. 1980. Craniofacial growthof fetal Macaca nemestrina: a cephalometric roent-genographic study. Am J Phys Anthropol 53:407–421.
Sirianni JE, Swindler DR. 1979. A review of postnatalcraniofacial growth in Old World monkeys and apes.Yrbk Phys Anthropol 22:80–104.
Sirianni JE, Swindler DR. 1985. Growth and developmentof the pigtailed macaque. Boca Raton: CRC Press.
Sirianni JE, Van Ness AL. 1978. Postnatal growth ofthe cranial base in Macaca nemestrina. Am J PhysAnthropol 49:329–340.
Sjøvold T. 1984. A report on the heritability of somecranial measurements and non-metric traits. In: VanVark GN, Howells WW, editors. Multivariate statis-tics in physical anthropology. Dordrecht: D. Reidel. p223–246.
Slavkin HC. 1989. Developmental craniofacial biology.Philadelphia: Lea and Febiger.
Smith KK. 1996. Integration of craniofacial structuresduring development in mammals. Am Zool 26:70–79.
Smith RJ. 1994. Degrees of freedom in interspecificallometry: an adjustment for the effects of phyloge-netic constraint. Am J Phys Anthropol 93:95–107.
Smith RJ, Josell SD. 1984. The plan of the human face: atest of three general concepts. Am J Orthod 85:103–108.
Sokal RF, Rohlf FJ. 1981. Biometry. 2nd ed. New York:W.H. Freeman and Co.
Sperber GH. 1989. Craniofacial embryology. 4th ed.London: Wright.
Spoor CF. 1993. The comparative morphology and phy-logeny of the human bony labyrinth. Ph.D. disserta-tion, University of Utrecht.
Spoor CF. 1997. Basicranial architecture and relativebrain size of Sts 5 (Australopithecus africanus) andother Plio-Pleistocene hominids. S Afr J Sci 93:182–186.
Spoor CF, Zonneveld F. 1998. A comparative review ofthe human bony labyrinth. Yrbk Phys Anthropol 41:211–251.
Spoor CF, O’Higgins P, Dean MC, Lieberman DE. 1999.Anterior sphenoid in modern humans. Nature 397:572.
Spoor CF, Jeffery N, Zonneveld F. 2000. Imaging skel-etal growth and evolution. In: O’Higgins P, Cohn M,editors. Development, growth and evolution: implica-tions for the study of hominid skeletal evolution. Lon-don: Academic Press. p 123–161.
Srivastava HC. 1992. Ossification of the membranousportion of the squamous part of the occipital bone inman. J Anat 180:219–224.
Stamrud L. 1959. External and internal cranial base.Acta Odontol Scand 17:239–266.
Starck D. 1975. The skull of the fetal chimpanzee. In:Bourne GH, editor. The chimpanzee. Volume 6. Basel:S. Karger. p 1–33.
Stephan H, Frahm H, Baron G. 1981. New and reviseddata on volumes of brain structure in insectivores andprimates. Comp Primate Biol 4:1–38.
Stevens KN. 1972. Quantal nature of speech. In: DavidEE Jr, Denes PB, editors. Human communication: aunified view. New York: McGraw Hill.
Stevens KN, House AS. 1955. Development of a quan-titative description of vowel articulation. J Acoust SocAm 27:484–493.
Strait DS. 1998. Evolutionary integration in the homi-nid cranial base. Ph.D. dissertation, State Universityof New York, Stony Brook.
Strait DS. 1999. The scaling of basicranial flexion andlength. J Hum Evol 37:701–719.
Strait DS, Ross CF. 1999. Kinematic data on primatehead and neck posture: implications for the evolution
of basicranial flexion and an evaluation of registra-tion planes. Am J Phys Anthropol 108:205–222.
Strait DS, Grine FE, Moniz MA. 1997. A reappraisal ofhominid phylogeny. J Hum Evol 32:17–82.
Szalay FS, Rosenberger AL, Dagosto M. 1987. Diagno-sis and differentiation of the order Primates. YbkPhys Anthropol 30:75–105.
Topinard P. 1890. Anthropology. London: Chapman andHall.
van den Eynde B, Kjaer I, Solo B, Graem N, Kjaer TW,Mathiesen M. 1992. Cranial base angulation andprognathism related to cranial and general skeletalmaturation in human fetuses. J Craniofac Genet DevBiol 12:22–32.
Van der Linden FPG, Enlow DH. 1971. A study of theanterior cranial base. Am J Orthod 41:119–124.
Van Essen DC. 1997. A tension-based theory of morpho-genesis and compact wiring in the central nervoussystem. Nature 385:313–318.
Vidal PP, Graf W, Berthoz A. 1986. The orientation of thecervical vertebral column in unrestrained awake ani-mals. I. Resting position. Exp Brain Res 61:549–559.
Vielle-Grosjean I, Hunt P, Gulisano M, Boncinelli E,Thorogood PV. 1997. Branchial HOX gene expressionand human craniofacial development. Dev Biol 183:49–60.
Vinyard CI. 1994. A quantitative assessment of thesupraorbital region in moden Melanesians. MA The-sis Anthropology, Northern Illinois University.
Vinyard CJ, Smith FH. 1997. Morphometric relation-ships between the supraorbital region and frontalsinus in Melanesian crania. Homo 48:1–21.
Virchow R. 1857. Untersuchungen uber die Entwick-lung des Schadelgrundes im gesunden undFrankhaften Zustande. Berlin: Gesichissiblung undGehirnbru.
Vogel C. 1964. Uber eine Schadelbasisanomalie beieinem freier Wildbahn geschossen Cercopithecus tor-quatus atys. Z Morphol Anthropol 55:262–276.
Wagner GP. 1989. The biological homology concept.Annu Rev Ecol Syst 20:51–69.
Weidenreich F. 1941. The brain and its role in thephylogenetic transformation of the human skull.Trans Am Philos Soc 31:321–442.
White TD, Suwa G, Berhane A. 1994. Australopithecusramidus, a new species of early hominid from Aramis,Ethiopia. Nature 371:306–312.
Williams PL, Bannister LH, Berry MM, Collins P,Dyson M, Dussek JE, Ferguson MWJ. 1995. Gray’sanatomy. 38th ed. Edinburgh: Churchill Livingstone.
Young RW. 1959. The influence of cranial contents onpostnatal growth of the skull in the rat. Am J Anat105:383–390.
Zelditch ML. 1987. Evaluating models of developmentalintegration in the laboratory rat using confirmatoryfactor analysis. Syst Zool 36:368–380.
Zelditch ML. 1988. Ontogenetic variation in patterns ofphenotypic integration in the laboratory rat. Evolu-tion 42:28–41.
Zelditch ML, Fink WL. 1995. Allometry and develop-mental integration of body growth in a pirhana, Py-gocentrus natterieri (Teleostei: Ostariophysi). J Mor-phol 223:341–355.
Zelditch ML, Fink WL. 1996. Heterochrony and heter-otopy: stability and innovation in the evolution ofform. Paleobiology 22:241–254.
Zuckerman S. 1954. Correlation of change in the evolu-tion of higher primates. In: Huxley JS, Hardy A, FordEB, editors. Evolution as process. London: Allen andUnwin. p 347–401.
Zuckerman S. 1955. Age changes in the basicranial axisof the human skull. Am J Phys Anthropol 13:521–539.
PRIMATE CRANIAL BASE 169D.E. Lieberman et al.]





























