Embed Size (px)
Citation preview


The physiological and molecular response of StCDF
antisense lncRNA to drought stress in Solanum tuberosum
Li Shi
Reg. Nr: 941129759050
July 2018
MSc Thesis
Plant Sciences Group
Laboratory of Plant Breeding
Breeding for Growth and Development, Potato Group
WAGENINGEN UNIVERSITY
Supervisor:
dr. C.W.B. Bachem
Examiners:
dr. C.W.B. Bachem
dr. CG (Gerard) van der Linden

Acknowledgements
Writing this thesis would not have been possible without the help of many others. Therefore, I
would like to take this chance to thank these lovely people. First of all, I would like to thank
Christian Bachem, for the opportunity to work on this subject and his guidance during my thesis.
Furthermore, I would like to thank Lorena Ramirez for her daily supervision during this thesis. She
was always very patient in every discussion and always willing to explain the difficult parts to me. I
would also like to thank Marian Oortwijn for her help in the lab, especially in the beginning. Next, I
would like to thank the other people in the lab that were always very helpful whenever I was
looking for something or needed assistance. And finally, I would like to thank the other students.
Sharing the problems that we had during our research and discussing possible solutions really
helped the process of writing a thesis.

Table of Contents
Table of Contents ................................................................................................................................................ 1
List of figures ....................................................................................................................................................... 5
Abbreviations ...................................................................................................................................................... 6
Abstract ............................................................................................................................................................... 7
1. Introduction ..................................................................................................................................................... 8
2. Materials and Methods ................................................................................................................................. 12
2.1 Available plant lines ................................................................................................................................. 12
2.2 Experiment 1: non-drought comparison ................................................................................................. 13
2.2.1 Stomatal size and density. ................................................................................................................ 13
2.2.2 ABA response in stomates by using calculated stomatal conductance. .......................................... 13
2.3 Experiment 2: drought comparison......................................................................................................... 15
2.4 Experiment 3: Gene expression of StFLORE1 under ABA treatment. ..................................................... 15
2.5 Statistical analyses ................................................................................................................................... 16
3. Results ........................................................................................................................................................... 17
3.1 Stomatal ABA responsiveness ................................................................................................................. 17
3.2 In vitro, StFLORE has a function in response to ABA treatment. ............................................................. 19
3.3 Stomatal phenotype under non-drought conditions. ............................................................................. 20
3.3.1 Stomata size and density. ................................................................................................................. 20
3.3.2 Stomata index and pavement cell enlargement .............................................................................. 22
3.4 Calculated stomatal conductance (gs) .................................................................................................... 24
3.5 Phenotypic response to drought ............................................................................................................. 25
4. Discussion ...................................................................................................................................................... 26
4.1 Novel functions of StCDFs in regulating plant drought tolerance ........................................................... 27
4.2. StCDF1 has an antagonistic effect on stomatal ABA responsiveness ..................................................... 28
4.3 The role of StFLORE in drought response ................................................................................................ 29
4.4 StCDFs affect stomatal features in multiple ways ................................................................................... 31
4.4.1 Stomatal initiation ............................................................................................................................ 31
4.4.2 Epidermal cells expansion ................................................................................................................ 31
4.5 In potato, StCDF1-StFLORE connects tuberization and transpiration. .................................................... 33
5.Conclusion ...................................................................................................................................................... 35
6. Ref .................................................................................................................................................................. 36
7. Supporting information ................................................................................................................................. 39

List of figures
Figure1. Simplified visualization of the tuberisation pathway ..................................................................................... 9
Figure2. The drought response in different plant line with different combination of StCDF1 and StFLORE. ............. 10
Figure 3. Structure of the three different alleles of the StCDF1 gene. ....................................................................... 11
Table 1: Non-GMO lines analysed for non-drought treatment and drought tolerance ............................................. 12
Table 2: GMO lines analysed for non-drought treatment and drought tolerance ..................................................... 12
Figure 4. Measurement parameters of stomata......................................................................................................... 13
Figure 5. Stomatal conductance measurement. ......................................................................................................... 14
Table 3: plant lines analyzed for gene expression under ABA treatment ................................................................... 15
Table 4: Primers used for making cDNA for the StFLORE1 ......................................................................................... 15
Table 5: Primers used for qPCR analysis of StFLORE1 ................................................................................................. 16
Figure 6. Pore aperture ratio in different plant lines with different combination of StCDF1 and StFLORE. ............... 18
Figure 7. Relative gene expression of StFLORE1 of in vitro......................................................................................... 19
Figure 8. Leaf abaxial surface of transgenic lines and the non-transgenic lines under a magnification of 40X. ........ 20
Figure 9. stomatal size and density in different plant lines with different combinations of StCDF1 and StFLORE. ... 21
Figure 10. Stomatal index in different plant lines with different combination of StCDF1 and StFLORE. .................... 22
Figure 11. enlargement of stomata and pavement cell in different plant lines with different combination of StCDF1
and StFLORE. ....................................................................................................................................................... 23
Figure 12. Calculated stomatal conductance in different plant lines with different combination of StCDF1 and
StFLORE. .............................................................................................................................................................. 24
Figure 13. drought response in non-transgenic plant lines. ....................................................................................... 25
Figure 14. Drought response in transgenic plant lines and their background plants. ................................................ 26
Figure 15. Overview of H+-ATPase-mediated stomatal movement. .......................................................................... 28
Figure 16. Different roles of antisense lncRNAs in stress response in plants. ............................................................ 29
Figure 17. The control of stomatal development. ...................................................................................................... 31
Figure 18. epidermal cells expansion in Chinese cabbage and potato ....................................................................... 32
Figure 19. Proposed model on StCDF1 cooperates with its long non-coding partner StFLORE in regulating drought
response related stomatal characteristic. .......................................................................................................... 33
Figure 20. Drought response strategy in potato. ........................................................................................................ 34

Abbreviations
AA Amino acids ABA Abscisic acid Bp Base pair CDF Cycling Dof Factor CK Cytokinin CO Constans DOF DNA binding with one finger ES Epidermal cell size FKF1 Flavin-Binding Kelch Repeat F-Box FT Flowering locus T GA Gibberellin GI Gigantea
Gs Stomatal conductance LD Long days LncRNA Long non-coding RNA MES 2-(N-morpholino) ethanesulfonicacid RNAi RNA interference SD Short days SD Stomata density SI Stomatal index SS Stomatal size WT Wild type

Abstract
Potato (Solanum tuberosum) is one of the most important crops in the word. Potato originates from
the Andean regions of South America and evolved short-day-dependent tuber formation as a
reproductive strategy. The transition from vegetative to reproductive growth is controlled by
photoperiodic information via the circadian rhythm. In previous studies, we have identified the
central regulator called StCDF1. This gene belongs to the DOF (DNA-binding with one finger)
transcription factors family and it controls tuberization and plant life cycle. Apart from the wild type
(WT) allele (StCDF1.1) that regulates tuberization under short day conditions, there are two
mutated alleles which induce tuberization under long day conditions; StCDF1.2 with a 7bp insertion
and StCDF1.3 with an 865bp insertion. Both StCDF1.2 and StCDF1.3 have a damaged regulatory
domain and increased stability during the day. In addition, we have detected the presence of a
200bp lncRNA we have named StFLORE, transcribed in an opposite orientation covering the second
exon of the StCDF1 coding region. However, in StCDF1.3, StFLORE is unlikely to be functional since
the large insertion displaced StFLORE.
In previous observations, the StCDF1 knock down plants were highly tolerant to drought and the
StFLORE expression was enhanced under drought condition in these plants. These preliminary
results show a potential link between StCDF1-StFLORE locus and drought tolerance. In potato
breeding, reducing water loss by having lower value of transpiration is considered to be a beneficial
trait in responding to drought. In this study, we further look into how different combinations of
StCDF1 alleles and different expression levels of StCDFs affect transpiration related stomatal
characteristics, such as stomatal ABA responsiveness, stomatal size, stomatal density, etc. By
calculating stomatal conductance (gs) to estimate transpiration level, we found that StCDF1 knock
down plants have a very low calculated gs value and this result is in agreement with the drought
tolerance phenotype. Moreover, overexpressing StCDF1 leads to an abscisic acid (ABA) insensitive
stomata phenotype and this result provides a possible explanation for the negative effect of StCDF1
in drought response. In this study, we also examined the StFLORE expression level in both control
and ABA treatment conditions. As expected, the ABA treatment enhanced StFLORE expression and
this result delivered the message that StFLORE has a positive effect on drought response under the
influence of ABA.
Key words: Solanum tuberosum, Potato, CDF1, StFLORE, Drought Tolerance, Tuberization,
Transpiration.

1. Introduction
Potato (Solanum tuberosum) is the fourth largest food crop, following rice, wheat, and corn (Alva,
Fan, Qing, Rosen, & Ren, 2011). The total world potato production had achieved 375 million metric
tons (MMT) at 2014 (FAOSTAT, 2014), and this number has not changed much today. The previous
studies indicate the increasing aridity has become one of the most threatening factors to crops
productivity (Godfray et al. 2010), especially for potato. Despite the fact that potato stands out for
its high water-use efficiency, it is also highly sensitive to the drought stress. In 2010, the severe
summer drought resulted in 30% (around 7 million metric tons) loss in potato production in Russia
(GAIN, 2010). With the increasing uncertainty as a result of climate change, this situation could
reappear in many other countries in the future. Potato’s drought sensitivity is thought to result
from the regulation of transpiration from leaves and shallow root system. The preliminary data in
our research group indicates that the factors regulating tuber formation are closely connected to
the regulation of drought susceptibility and tolerance.
Transpiration is the essential process by which water is moved through plant from roots to leaves,
where water will be released to the atmosphere via stomata (Lloyd, 1908; Fanourakis et al., 2014).
In plants, stomata control the water evaporation and the gas exchange, that is essential for
photosynthesis. Under non-drought conditions, high light intensity and high humidity lead to
negative cell’s electrical potential, increasing in the stomatal aperture (Christodoulakis, Menti &
Galatis, 2002). However, under drought condition, to prevent water loss by transpiration, plants
accumulate a stress-response hormone called abscisic acid (ABA) that induces the rapid closing of
stomata by regulating Ca+ and K+ channels (Cai et al., 2017).
The variation in stomatal anatomical features and stomatal closing ability influences transpiration in
a diverse way. Recent research indicates that lower stomatal density and larger stomatal size had
reduced transpiration, and increased tolerance to limited water availability (Doheny-Adams et
al.,2012). For stomatal closing ability, the genotype was recognized as the major role in stomatal
response to drought (Skirycz and Inze, 2010), but the genetic variation of stomatal closing ability
hasn’t been explored much yet (Fanourakis et al., 2014). In potato, several genes have been
reported to be modulated under drought stress (Obidiegwu, Jones, & Prashar, 2015). The recently
unpublished studies by J. Abelenda et al. demonstrated that silencing the gene StCDF1 increase
drought stress (personal communication) by modulating stomatal anatomical features and stomatal
closing ability.
The stomatal anatomical features are highly depending on the development of stomata which
involved complicated genes and several plant hormones. The initiation of stomata has been known
that multiple plant hormones involved in this regulation. Both auxin and ABA has been reported
that they can negatively effect on stomatal density in Arabidopsis (Tanaka, Nose, Jikumaru &
Kamiya, 2013; Saibo, et al., 2003). However, gibberellin (GA) treatment in Arabidopsis and

enhanced cytokinin (CK) signalling in tomato increase the stomatal density (Saibo, et al., 2003;
Farber, Attia & Weiss, 2016). The knowledge about stomatal size regulation is still quite limited. By
studying the stomatal formation in Arabidopsis thaliana, this process involves at least one
asymmetric division as well as a single symmetric division (Qi, & Torii, 2018). Some researchers
suggest the stomatal and epidermal cells enlargement can be result from endoreduplication
(Kondorosi, Roudier, & Gendreau, 2000; Tanaka, Nose, Jikumaru & Kamiya, 2013).
In higher plants, DNA-binding with one finger (DOF) type transcription factors are involved in many
regulatory processes, such as flowering time and seed maturation (Corrales et al., 2017). Among all
DOF factors, there is a family of DOF factors in Arabidopsis whose transcription fluctuated under
the constant light condition and they are known as CYCLING DOF FACTORS (CDF1-5) (Fornara et al.
2009). In potato, a CDF-homologue called StCDF1 is located on Chromosome 5 that was identified
as an indirect inducer of tuberization (Kloosterman et al. 2013). Under short-day conditions, StCDF1
is not degraded because of the lack of StGI-StFKF1 complex which normally binds StCDF1 third
domain tagging it for ubiquitination and degradation by proteasome. Therefore, under inductive
short-day conditions, StCDF1 remains available to repress StCO (CONSTANS) gene expression,
promoting an indirectly increasing of StSP6A expression giving to initiation of tuberization (Navarro
et al. 2011; Abelenda, Navarro, & Prat, 2014; Navarro, Cruz-Oró, & Prat, 2015) (figure1).
Allelic variation at the StCDF1 locus also results in natural variation in the time point of tuber
formation. The wild-type allele which results in late tuberisation under long-day conditions has
been called StCDF1.1. Additional early alleles (StCDF1.2 and StCDF1.3) code for truncated variants
of the StCDF1 protein that prevent the binding of the FKF1 protein, thereby evading the normal
degradation of the protein resulting in constitutive repression of StCO gene expression leading to
day-length independent tuberization. Due to their early phenotype, these two alleles have been
Figure1. Simplified visualization of the tuberisation pathway. Under short days, StGI and StFKF1 do not
bind to StCDF1, resulting in transport of StSP6A from the stolon to the tubers.

widely selected and applied in the modern potato breeding and cultivation. However, the current
results from our research group, show that the homozygous diploid potatoes for StCDF1.3 allele
grow unhealthily and weakly even under normal condition. Furthermore, the heterozygous diploid
potatoes with single copy of StCDF1.3 allele show significant less tolerance to drought (figure 2.).
In Arabidopsis, the CDF genes have been shown coding transcription factor for the photoperiodic
flowering-time control (Imaizumi et al, 2005). Recently, a long non-coding transcript (lncRNA)
covering the 3’- half of the CDF5 gene was mapped and named as FLORE. In potato, we have also
detected the presence of a 200bp lncRNA we have called StFLORE1, transcribed in an opposite
orientation from the 3’-UTR of StCDF1 gene extending into the first exon. Moreover, experiments
from our laboratory indicate that plants with silenced StCDF1 have not only enhanced drought
tolerance but also remarkable increased expression of StFLORE1 (Bachem et al. unpublished data).
As mentioned above, the earliness phenotypes of StCDF1.2 and StCDF1.3 are result from mutations.
In the case of StCDF1.2 there is a premature stop codon in the second exon due to a 7-base-pair
(bp) insertion, whereas StCDF1.3 has a larger insertion of 865bp at the same location that results in
an additional 22 amino acids (AAs) extension of the StCDF1 protein (Kloosterman et al., 2013)
(figure 3). The insertion in the StCDF1.3 allele displaces the StFLORE1 transcript, which is likely to
render it non-functional, while in the StCDF1.2 allele the 7bp insertion is less likely to have a
significant effect on the StFLORE1 transcript. In a comparison of heterozygous 1.1/1.3 with 1.1/1.2
diploid potatoes, the latter allelic configuration was shown to have larger size of stomata with
lower density and these stomatal phenotypes seems related with its lower susceptibility to drought
stress. That delivered important message about it is possible for StFLORE involved in the regulation
of drought response. However, without more genetic modified plants, the unique gene structure of
this locus makes it very hard to analysis the individual effect of StFLORE.
Figure2. The drought response in different plant line with different combination of StCDF1 and StFLORE. A) 1;
Heterozygous diploid potato carrying StCDF1.2/1.3 alleles (drought sensitive), 2, 3; two independent StCDF1:
RNAi knockdown plant with elevated StFLORE1 expression (drought tolerant). B) 1; diploid homozygous
StCDF1.1, wild type (drought tolerant) 2; diploid StCDF1.1/1.3 heterozygote (moderately drought sensitive).3;
StCDF1.3 homozygote (very drought sensitive).
1 2 3 1 2 3

In Arabidopsis CDF proteins appear to be largely functionally redundant (Fornara et al. 2009). In
potato, StCDF1 appears to be a master transcription factor that not only regulates CONSTANS genes
but also supresses the expression of StCDF2 and 3 while not effecting expression of StCDF4 & 5. At
the same time, over expression of StCDF3 was shown to also lead to early tuberization indicating a
degree of functional overlap. Moreover, only StCDF1 and StCDF4 genes appear to have a
corresponding lncRNA. Together, these data indicate that there are a complex set of regulatory
networks between CDF genes and their associated lncRNA transcripts.
Base on this background information, the formulated research questions are:
1. How does StCDF1-StFLORE affect drought response in potato?
2. How different combinations of different StCDF1 allelic variations’ affect stomata phenotypes?
3. Which gene give the main influence on drought response, StCDF1 or StFLORE?
Figure 3. Structure of the three different alleles of the StCDF1 gene. Blue box: StCDF1 coding
region, grey triangles: insertions, red box: StFLORE1 promoter, green: intron, arrows: transcripts
(dotted indicates non-functional transcript)

2. Materials and Methods
In this study, three experiments have been done. The first experiment was done under the non-
drought condition, as a baseline experiment to observe the phenotypic differences of stomata in
genotypes with different variations of StCDF1 alleles and transgenic genotypes with over-expression
or knock down of CDFs genes listed below. Then, both transgenic plants and non-transgenic plants
leave samples were exposed to ABA treatment. The third experiment is designed to test the StCDF1
and related genes expression level response to ABA treatment in vitro. In experiment 4, the plants
we used for experiment 1 will be put under drought condition to determine the macroscopic and
microscopic response to drought stress.
2.1 Available plant lines
In this experiment, plant lines 1-8 were grown. From each plant line, 6 replicates were prepared.
Thus, 60 plants will be used in this experiment in total. First, plants were grown on MS medium for 2
weeks and then were transferred to the greenhouse conditions which was conducted at 25 °C and
20% relative humidity.
Table 1: Non-GMO lines analysed for non-drought treatment and drought tolerance
Number Name Ploidy CDF1 alleles StFLORE expected
1 CE3027 Diploid 1.1/1.1 Two copies
2 CE605 Diploid 1.1/1.2 Two copies
3 CE630 Diploid 1.1/1.3 One copy
4 RHXE_30 Diploid 1.3/1.3 No
5 CE3130 Diploid 1.2/1.3 No
Table 2: GMO lines analysed for non-drought treatment and drought tolerance
Number Name Ploidy Background
Genotype
StFLORE expected
6 CE3130 RNAi CDF1 Diploid 1.2/1.3 One copy
7 35S CDF1.2 CE3027_4 Diploid 1.1/1.1 Two copies
8 35S CDF3 CE3130_5 Diploid 1.2/1.3 One copy

2.2 Experiment 1: non-drought comparison
2.2.1 Stomatal size and density.
Stomatal density and size were measured in 9 potato plants lines after 4 weeks of being grown in
the greenhouse by using the terminal leaf leaflet of the mature leaf closest to the apical zone.
The sampling preparation was obtain according to Pei,
Kuchitsu,Ward, Schwarz, and Schroeder (1997), briefly,
cleared epidermal peels from transparent nail polish
was applied in the abaxial leaflet to obtain an imprint
of the epidermal surface that was examined under the
microscope using 40X magnification (Olympus Provis
AX70). The lecture was made in 10 rectangular fields of
view for 5 replicates by selecting randomly 40 stomates
per line (40 stomatasx9 lines, N=360).
The stomata density (SD) and stomatal index (SI) was
calculate as follow:
SD(𝑚𝑚−2) =𝑁° of 𝑠𝑡𝑜𝑚𝑎𝑡𝑎𝑠
𝑓𝑖𝑒𝑙𝑑 𝑎𝑟𝑒𝑎 (0.0314 𝑚𝑚2)
SI (%) =𝑠𝑡𝑜𝑚𝑎𝑡𝑎𝑙 𝑑𝑒𝑛𝑠𝑖𝑡𝑦
𝑆𝑡𝑜𝑚𝑎𝑡𝑎𝑙 𝑑𝑒𝑛𝑠𝑖𝑡𝑦 + 𝑒𝑝𝑖𝑑𝑒𝑟𝑚𝑎𝑙 𝑐𝑒𝑙𝑙 𝑑𝑒𝑛𝑠𝑖𝑡𝑦
× 100%
Whereas for the stomatal size (SS) and epidermal cell size (ES) we calculated:
SS(𝑚𝑚2) = 𝜋 ×𝑠𝑡𝑜𝑚𝑎𝑡𝑎𝑙 𝑙𝑒𝑛𝑔𝑡ℎ
2×
𝑠𝑡𝑜𝑚𝑎𝑡𝑎𝑙 𝑤𝑖𝑑𝑡ℎ
2
ES(𝑚𝑚2) =𝑓𝑖𝑒𝑙𝑑 𝑎𝑟𝑒𝑎 (0.0314 𝑚𝑚2) − (SS̅̅ ̅ × SD̅̅̅̅ )
1 − 𝑆𝐼
It is important to mention that with the purpose of measuring the stomata size properly we decided to force stomatal closure for all the 9 lines evaluated by using a buffer 50Mm ABA MES/KOH buffer (50Mm ABA 5mM KCl,10mM MES(2-(N-morpholino) ethanesulfonicacid), 50μM CaCl2, pH 6.15) reported by Desikan et al,.2008 and Eisele et al, 2016.
2.2.2 ABA response in stomates by using calculated stomatal conductance.
To estimate the ABA response in stomates, at least 6 leaf samples from each plant line were collected
in the morning. As figure 5 shown, first each leaf sample was incubated in Petri dishes with MES/KOH
buffer (5mM KCl,10mM MES(2-(N-morpholino) ethanesulfonicacid), 50μM CaCl2, pH 6.15; (Desikan
et al,.2008; Eisele, Fäßler, Bürgel, & Chaban, 2016) for two hours to force stomata opening, and after
that it was cut along the midrib. Half of one leaflet was treated for 30 minutes with ABA MES/KOH
buffer (10μM ABA, MES/KOH buffer) to induce stomata closure and the other half was used as a
control (only treat with MES/KOH buffer, Mock). At least 30 stomata were obtained from each line
Figure 4. Measurement parameters of stomata.

(30 stomates x 8 lines, N=240). The length and width of the stomata aperture was examined under
the micro-scope using 40X magnification and measured by image J program.
Figure 5. Stomatal conductance measurement.
The pore aperture ratio between ABA treatment and MOCK treatment indicate the sensitivity of stomata to ABA. Therefore, ∆pore aperture, was calculated by following equation:
∆𝑝𝑜𝑟𝑒 𝑎𝑝𝑒𝑟𝑡𝑢𝑟𝑒 𝑟𝑎𝑡𝑖𝑜 =𝑝𝑜𝑟𝑒 𝑎𝑝𝑒𝑟𝑡𝑢𝑟𝑒 𝑎𝑟𝑒𝑎 𝑖𝑛 𝐴𝐵𝐴 𝑡𝑟𝑒𝑎𝑡𝑚𝑒𝑛𝑡
𝑝𝑜𝑟𝑒 𝑎𝑝𝑒𝑟𝑡𝑢𝑟𝑒 𝑎𝑟𝑒𝑎 𝑖𝑛 𝑀𝑂𝐶𝐾 𝑡𝑟𝑒𝑎𝑡𝑚𝑒𝑛𝑡× 100%
In order to have general view of gas change and water vapor, the gs was calculated for abaxial leaf surface under both MOCK and ABA treatment conditions, based on the following equation (Franks and Farquhar, 2001).
gs =(𝑑𝑖𝑓𝑓𝑢𝑠𝑖𝑜𝑛 𝑐𝑜𝑒𝑓𝑓𝑖𝑐𝑖𝑒𝑛 𝑡) × (𝑠𝑡𝑜𝑚𝑎𝑡𝑎𝑙 𝑑𝑒𝑛𝑠𝑖𝑡𝑦) × (𝑝𝑜𝑟𝑒 𝑎𝑟𝑒𝑎)
(𝑚𝑜𝑙𝑎𝑟 𝑣𝑜𝑙𝑢𝑚𝑒 𝑜𝑓 𝑎𝑖𝑟) × [(𝑝𝑜𝑟𝑒 𝑑𝑒𝑝𝑡ℎ) + 𝜋/2 × √(𝑝𝑜𝑟𝑒 𝑎𝑟𝑒𝑎/𝜋)2 ]
The mean molar volume of air was considered 0.024 m3mol-1, and the effective diffusion coefficient
for water vapor in air was 2.43x10−5 m2 s–1 at 21 °C, according to Jones (1992). Stomatal pore depth
was considered to be equal to the guard cell width (i.e. stomatal width/2), assuming that guard cells
inflate to a circular cross-section (Franks & Beerling 2009; Franks & Farquhar 2007).

2.3 Experiment 2: drought comparison
After experiment 1, 8 plant lines (40 plants in total) grown for another 3 weeks under non-drought
condition before drought treatment. The drought treatment continued 3 weeks. After 3 weeks
drought treatment, the IR camera (FLIR one Pro IOS) used to measuring leaves temperature for each
plant line and the temperature difference recorded in photos.
2.4 Experiment 3: Gene expression of StFLORE1 under ABA treatment.
Because RNAi CDF1 transgenic plants were the candidates to be more drought tolerant, we decided
to test the StFLORE1 expression under ABA treatment for RNAi CDF1 3027, RNAi CDF1 CE3130 and
include CE3027 and CE3130 as controls. Plants were grown under the same conditions in climate
chamber and after 2 weeks, 6 apexes per each genotype were cut and planted in plates containing
MS medium (0.5×MS, 1% sucrose, and 0.8% agar) for 7 days. Thereafter, the ABA treatment was
imposed, in which 3 replicates per each genotype were transfer to MS with ABA (50μM) and others
3 were transfer to new MS media without ABA for 4 hours at 22°C with a 16-h light/8-h dark cycle.
After treatment, the roots and apex were collected and store at -80 °C.
Table 3: plant lines analyzed for gene expression under ABA treatment
Name Ploidy CDF1 alleles StFLORE expected
CE3027 Diploid 1.1/1.1 Yes
CE3130 Diploid 1.2/1.3 No
CE3130 RNAi Diploid 1.2/1.3 Yes
CE3027 RNAi Diploid 1.1/1.1 Yes
Briefly, total RNA was extracted using the RNeasy Plant Mini Kit from Qiagen, with the corresponding
protocol. RNA was treated with DNAse I (Invitrogen) to eliminate residual DNA. After DNAse
treatment, cDNA was performed from 1ug RNA template using Invitrogen SuperScript III (Appendix
X) and diluted 1:10. Quantitative real-time PCR was conducted using iQ SYBR Green supermix (Bio-
rad) in a Bio-Rad iCycleriQ machine.
Table 4: Primers used for making cDNA for the StFLORE1
target Name of the primer Sequence
CDF1 cDNAr1CDF GACGTGATTGTAGTTACTAACC
CDF1 cDNAr2CDF TGGGACGTAAAGACGTTCTC
EF3 RT_REV_349 REV CGTTGGTGAATGCGGCAGTAGG
NAC NAC_reverse TCCATGATAGCAGAGACTA
For Real Time PCR, a master mix contained 5uL 2X iQ SYBR GREEN, 1uL forward primer (3uM), 1uL
reverse primer (3uM) and 3 uL cDNA template into a final volume of 10uL was prepared.
Thermocycling conditions were 95C for 3 minutes,40 cycles of 95C for 30 seconds and 60C for 30

seconds. The primers used for the expression analysis are listed in Table X. We tested NAC and ELF3
as housekeeping, but finally we kept ELF3 as a housekeeping control gene because the expression
was more homogenous.
Table 5: Primers used for qPCR analysis of StFLORE1
target Name of the primer Sequence
StFLORE1 qPCRrFCDF qPCRrRCDF
TGCAGACTCGTCGATTGAAC GAGTGCCTTTTCCTCACTCG
NAC NAC_forward NAC_reverse
ATATAGAGCTGGTGATGACT TCCATGATAGCAGAGACTA
ERF3 ERF3_forward ERF3_reverse
For the data analysis an RGE expression was calculated by the following formula according to Livak
and Schmittgen, 2001.
𝑅𝐺𝐸 = 2𝐶𝑞𝑁𝐴𝐶−𝐶𝑞𝐺𝑂𝐼
2.5 Statistical analyses
Data analysis was performed using the IBM SPSS software (version 22;
https://www.ibm.com/analytics/spss-statistics-software). The stomatal size, stomatal density, stomatal
index, stomatal ABA responsiveness and calculated stomatal conductance (gs) data were subjected to an
analysis of variance homogeneity to determine if its distribution was normal. Since the data presented non-
normal distribution we applied non-parametric test by using Kruskal-Wallis Test. The differences among
the genotypes were found through Fisher's LSD test. The differences were considered significant for a
value of P ≤ 0.05.

3. Results
3.1 Stomatal ABA responsiveness
We have shown that overexpressing StCDF3 and StCDF1.2 in potato induces earliness phenotype.
While knocking down StCDF1 gene expression by RNAi delays tuberisation, these transgenics also
show an enhanced tolerance to drought. However, no clear link has been established between
drought responsiveness and StCDF proteins. Since stomatal opening is linked to transpiration and
stomatal opening is regulated by the plant hormone ABA, we examined ABA responsiveness in
three transgenic lines (RNAi CDF1 CE3130, 35S CDF3 CE3130 and 35S CDF1.2 CE3027) and their
background lines (CE3130 and CE3027). These plants all have either different allelic variants of
StCDF1 or have transgenes with altered StCDF1 or StCDF3 expression.
In this study, the ABA responsiveness is represented by the pore aperture ratio between ABA
treatment and control treatments (see Material & Methods). A lower value of aperture ratio
indicates a more rapid stomatal response to ABA treatment. The StCDF1 knock-down in CE3130
(1.2/1.3) plants had reduced pore aperture ratio compared to CE3130 (1.2/1.3) plants (figure 6 A).
This result suggests the more rapid drought response in RNAi CDF1 CE3130. However,
overexpressing StCDF3 in CE3130 did not seem to affect ABA responsiveness, since the pore
aperture ratio of 35S CDF3 CE3130 did not show a significant difference with CE3130. We conclude
that, StCDF3 may not be directly involved in regulating stomatal movements.
Overexpressing StCDF1.2 in CE3027 resulted in a very high pore aperture ratio (>1; figure 6 B). This
indicates the stomata in this line seem to barely respond to the stomatal closing signal and may
need a longer time or more ABA to trigger stomatal closure.
Although both StCDF1 and StCDF3 show tuberization inducing ability, they work differently in
regulating stomatal ABA response. Unlike StCDF3, overexpressing StCDF1.2 seems to have a strong
negative effect on stomatal ABA responsiveness. In a previous study, the StCDF1 had been
proposed to have general negative effect on drought response in potato (Bachem et al.,
unpublished data). Based on this study, the negative effect of StCDF1 may relate to stomatal
insensitivity to ABA.
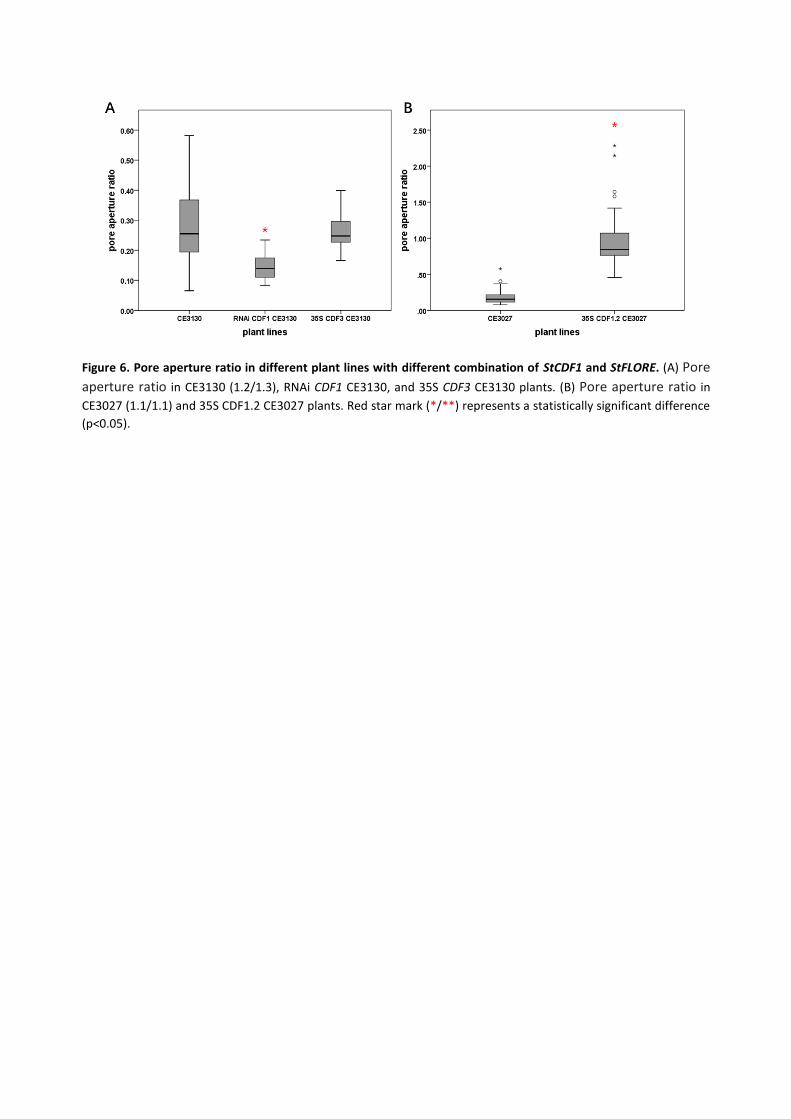
Figure 6. Pore aperture ratio in different plant lines with different combination of StCDF1 and StFLORE. (A) Pore
aperture ratio in CE3130 (1.2/1.3), RNAi CDF1 CE3130, and 35S CDF3 CE3130 plants. (B) Pore aperture ratio in
CE3027 (1.1/1.1) and 35S CDF1.2 CE3027 plants. Red star mark (*/**) represents a statistically significant difference
(p<0.05).
3.2 In vitro, StFLORE has a function in response to ABA treatment.
In potato the StCDF1 locus encodes a long non-coding RNA transcribed in an antisense orientation
called StFLORE. We have shown that multiple DOF binding sites present in the StFLORE promoter
make it likely that this transcript is regulated by StCDF1. The upregulation of StFLORE in RNAi CDF1
CE3130 is a confirmation of this hypothesis (Bachem et al, unpublished data). As described above,
the stronger stomatal ABA response was only observed in RNAi CDF1 CE3130. Therefore, to gain an
insight into whether the stronger stomatal responsiveness to ABA in RNAi CDF1 CE3130 relates to
StFLORE, the expression level of StFLORE under control and ABA treatment (50 µM) for 4 hours
exposition was examined in plants of 1 weeks old in MS medium in apex and root samples.
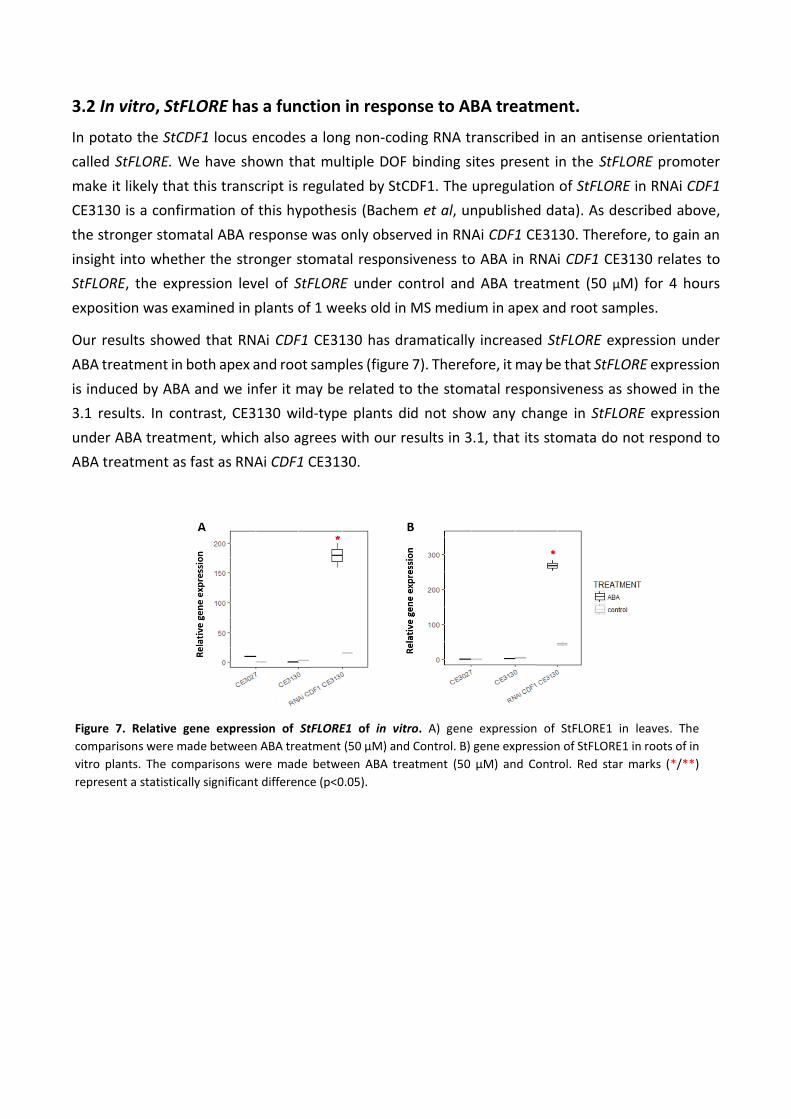
Our results showed that RNAi CDF1 CE3130 has dramatically increased StFLORE expression under
ABA treatment in both apex and root samples (figure 7). Therefore, it may be that StFLORE expression
is induced by ABA and we infer it may be related to the stomatal responsiveness as showed in the
3.1 results. In contrast, CE3130 wild-type plants did not show any change in StFLORE expression
under ABA treatment, which also agrees with our results in 3.1, that its stomata do not respond to
ABA treatment as fast as RNAi CDF1 CE3130.
Figure 7. Relative gene expression of StFLORE1 of in vitro. A) gene expression of StFLORE1 in leaves. The
comparisons were made between ABA treatment (50 µM) and Control. B) gene expression of StFLORE1 in roots of in
vitro plants. The comparisons were made between ABA treatment (50 µM) and Control. Red star marks (*/**)
represent a statistically significant difference (p<0.05).

3.3 Stomatal phenotype under non-drought conditions.
3.3.1 Stomata size and density.
Stomatal size and density have always been recognized as two important values for estimating
stomatal conductance and plant performance under drought. Plants with larger stomata and lower
stomatal density have been considered to have a “water saving” stomatal phenotype (Doheny-
Adams et al.,2012). So far, no link has been found between StCDF1 and stomatal phenotype. In this
study, we did a further investigation on stomatal phenotype among 8 different genotypes to
discover if StCDF1-StFLORE locus has an effect on these stomatal characters (see figure 8).
Unfortunately, no significant difference was found between the early varieties and late varieties.
Only CE605 (harboring StCDF1.1/1.2 alleles) stands out among all non-transgenic plant lines for its
larger stomatal size and lower density (figure 9, A-B).
The stomatal characteristics of RNAi CDF1 CE3130 fit with previous expectations. Knocking down
StCDF1 in CE3130 background showed increased stomatal size and reduced stomatal density (figure
9, C-D). However, the most dramatic stomatal enlargement and reduced density were found in the
two overexpression lines (figure 9, C-F). Both of these two plant lines increase their stomata size to
almost twice that of their background plants and have half of their background plants’ stomatal
density. There was no significant difference in stomatal size and density between 35S CDF3 CE3130
and 35S CDF1.2 CE3027.
Figure 8. Leaf abaxial surface of transgenic lines and the non-transgenic lines under a magnification of 40X. (A) From
right to left are CE3027 (1.1/1.1), CE3130 (1.2/1.3), CE605 (1.1/1.2), CE630 (1.1/1.3) and RHXE30 (1.3/1.3). (B) From right
to left are 35S CDF1.2 CE3027, 35S CDF3 CE3130, RNAi CDF1 CE3130. All the photos were taken with a scale bar = 50 µm

The transgenic plant lines delivered a clear message that StCDFs are involved in stomatal phenotypes
regulation. Unlike stomatal ABA response, overexpressing StCDF1.2 and StCDF3 seem to act similarly
in inducing stomatal enlargement and reducing stomatal density. However, the increased stomatal
size and reduced stomatal density has also been found in RNAi CDF1 CE3130. That suggests there
may be more than one way to obtain the “water saving” stomatal features. For CE605, more sublines
are required in future research to exclude the additional effects of genetic background.
Figure 9. stomatal size and
density in different plant
lines with different
combinations of StCDF1
and StFLORE. (A-B)
stomatal size and density
in non-transgenic plants
lines: CE605 (1.1/1.2),
RHxE30(1.3/1.3), CE3130
(1.2/1.3), CE630(1.1/1.3),
and CE3027 (1.1/1.1); C-D)
stomatal size and density
CE3130 background plants;
red star mark (*/**)
represents the significant
difference level.
3.3.2 Stomata index and pavement cell enlargement
In chapter 3.3.1, lower stomatal density was observed in several plant lines. The similar pattern of
increasing stomatal size and decreasing density in 35S CDF3 CE3130 and RNAi CDF1 CE3130 lead us
to look into additional stomatal characteristics. Considering the decrease of stomatal density could
result from less stomatal formation, epidermal cell expansion or other reasons, we further measured
stomatal index (formula can be found in Material & Method) in three transgenic lines (35S CDF3
CE3130, RNAi CDF1 CE3130 and 35S CDF1.2 CE3027) and their background plant lines (CE3130 and
CE3027).
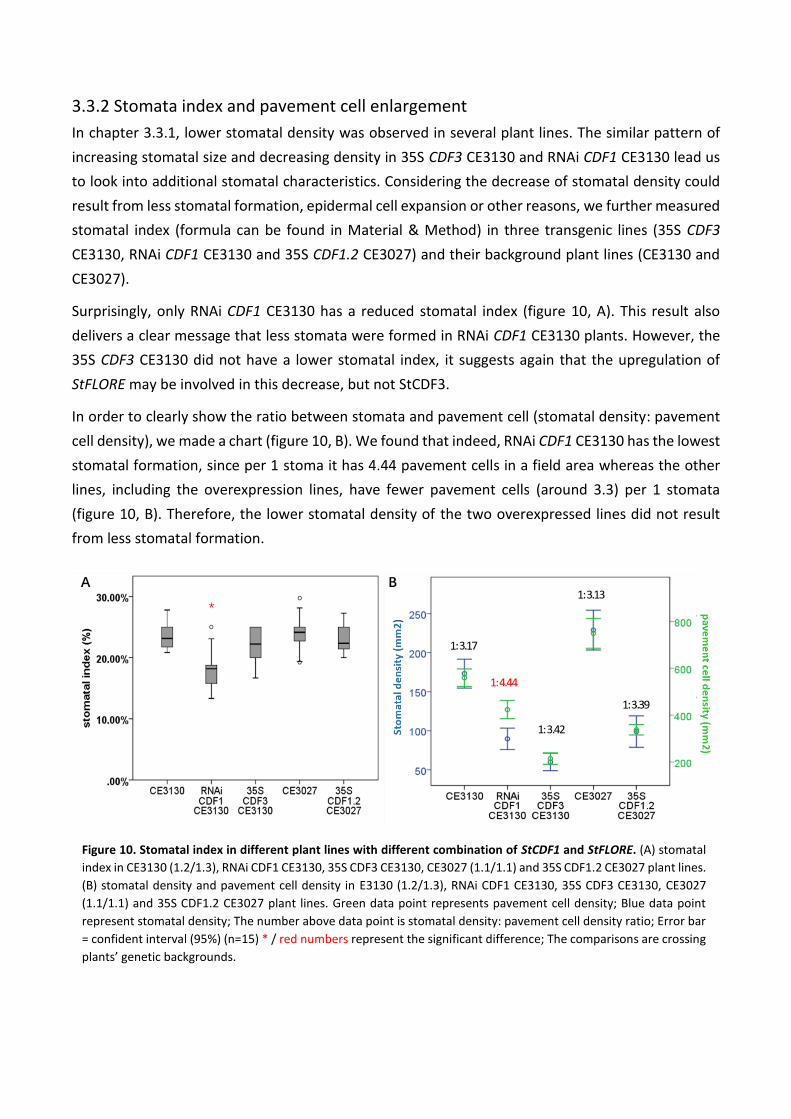
Surprisingly, only RNAi CDF1 CE3130 has a reduced stomatal index (figure 10, A). This result also
delivers a clear message that less stomata were formed in RNAi CDF1 CE3130 plants. However, the
35S CDF3 CE3130 did not have a lower stomatal index, it suggests again that the upregulation of
StFLORE may be involved in this decrease, but not StCDF3.
In order to clearly show the ratio between stomata and pavement cell (stomatal density: pavement
cell density), we made a chart (figure 10, B). We found that indeed, RNAi CDF1 CE3130 has the lowest
stomatal formation, since per 1 stoma it has 4.44 pavement cells in a field area whereas the other
lines, including the overexpression lines, have fewer pavement cells (around 3.3) per 1 stomata
(figure 10, B). Therefore, the lower stomatal density of the two overexpressed lines did not result
from less stomatal formation.
Figure 10. Stomatal index in different plant lines with different combination of StCDF1 and StFLORE. (A) stomatal
index in CE3130 (1.2/1.3), RNAi CDF1 CE3130, 35S CDF3 CE3130, CE3027 (1.1/1.1) and 35S CDF1.2 CE3027 plant lines.
(B) stomatal density and pavement cell density in E3130 (1.2/1.3), RNAi CDF1 CE3130, 35S CDF3 CE3130, CE3027
(1.1/1.1) and 35S CDF1.2 CE3027 plant lines. Green data point represents pavement cell density; Blue data point
represent stomatal density; The number above data point is stomatal density: pavement cell density ratio; Error bar
= confident interval (95%) (n=15) * / red numbers represent the significant difference; The comparisons are crossing
plants’ genetic backgrounds.
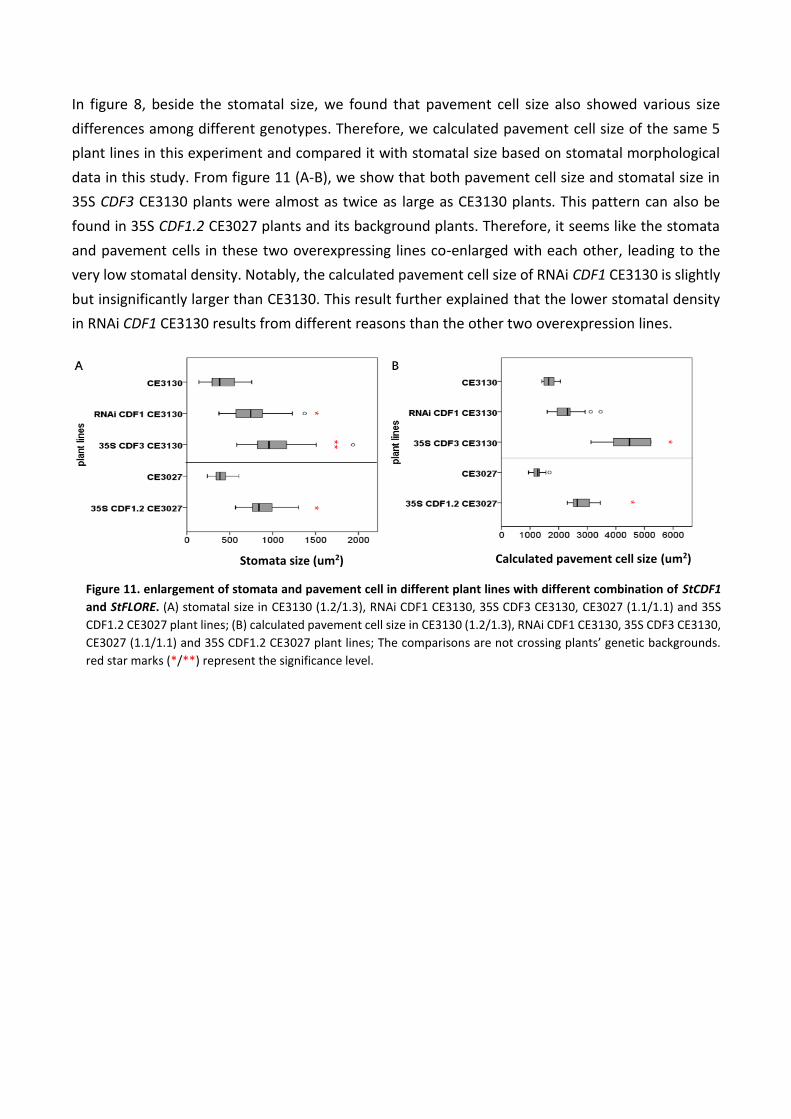
In figure 8, beside the stomatal size, we found that pavement cell size also showed various size
differences among different genotypes. Therefore, we calculated pavement cell size of the same 5
plant lines in this experiment and compared it with stomatal size based on stomatal morphological
data in this study. From figure 11 (A-B), we show that both pavement cell size and stomatal size in
35S CDF3 CE3130 plants were almost as twice as large as CE3130 plants. This pattern can also be
found in 35S CDF1.2 CE3027 plants and its background plants. Therefore, it seems like the stomata
and pavement cells in these two overexpressing lines co-enlarged with each other, leading to the
very low stomatal density. Notably, the calculated pavement cell size of RNAi CDF1 CE3130 is slightly
but insignificantly larger than CE3130. This result further explained that the lower stomatal density
in RNAi CDF1 CE3130 results from different reasons than the other two overexpression lines.
Stomata size (um2) Calculated pavement cell size (um2)
Figure 11. enlargement of stomata and pavement cell in different plant lines with different combination of StCDF1
and StFLORE. (A) stomatal size in CE3130 (1.2/1.3), RNAi CDF1 CE3130, 35S CDF3 CE3130, CE3027 (1.1/1.1) and 35S
CDF1.2 CE3027 plant lines; (B) calculated pavement cell size in CE3130 (1.2/1.3), RNAi CDF1 CE3130, 35S CDF3 CE3130,
CE3027 (1.1/1.1) and 35S CDF1.2 CE3027 plant lines; The comparisons are not crossing plants’ genetic backgrounds.
red star marks (*/**) represent the significance level.
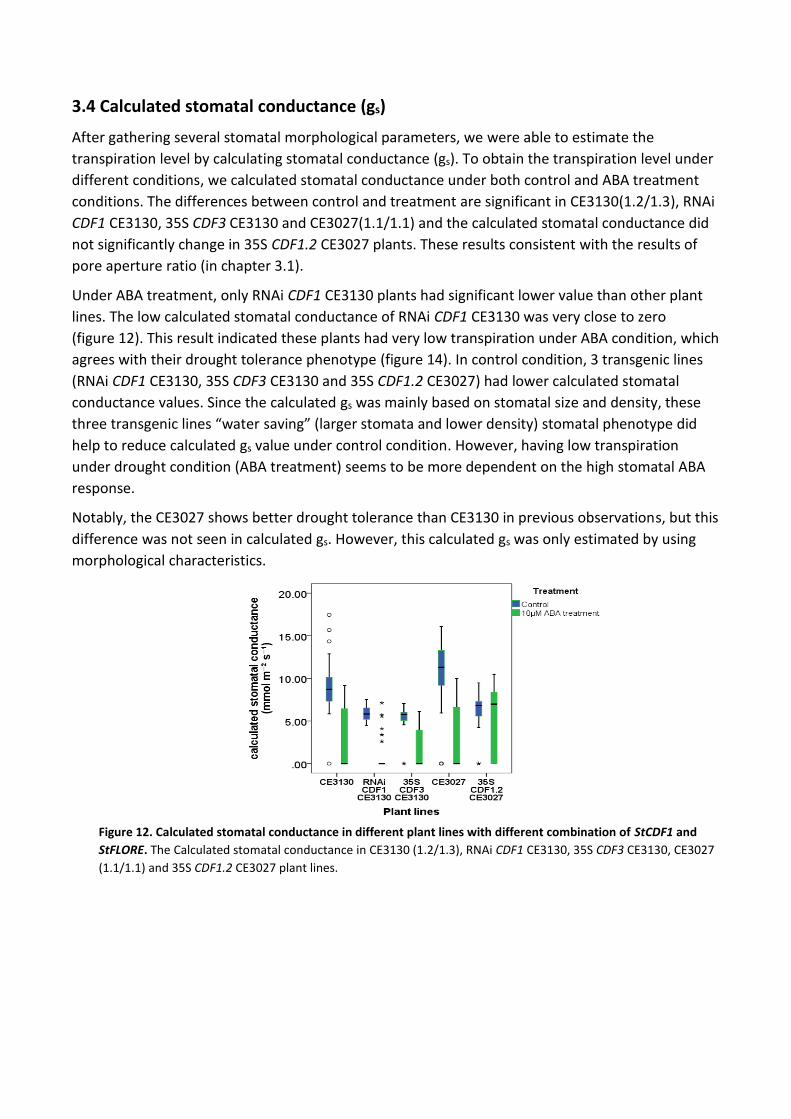
3.4 Calculated stomatal conductance (gs)
After gathering several stomatal morphological parameters, we were able to estimate the
transpiration level by calculating stomatal conductance (gs). To obtain the transpiration level under
different conditions, we calculated stomatal conductance under both control and ABA treatment
conditions. The differences between control and treatment are significant in CE3130(1.2/1.3), RNAi
CDF1 CE3130, 35S CDF3 CE3130 and CE3027(1.1/1.1) and the calculated stomatal conductance did
not significantly change in 35S CDF1.2 CE3027 plants. These results consistent with the results of
pore aperture ratio (in chapter 3.1).
Under ABA treatment, only RNAi CDF1 CE3130 plants had significant lower value than other plant
lines. The low calculated stomatal conductance of RNAi CDF1 CE3130 was very close to zero
(figure 12). This result indicated these plants had very low transpiration under ABA condition, which
agrees with their drought tolerance phenotype (figure 14). In control condition, 3 transgenic lines
(RNAi CDF1 CE3130, 35S CDF3 CE3130 and 35S CDF1.2 CE3027) had lower calculated stomatal
conductance values. Since the calculated gs was mainly based on stomatal size and density, these
three transgenic lines “water saving” (larger stomata and lower density) stomatal phenotype did
help to reduce calculated gs value under control condition. However, having low transpiration
under drought condition (ABA treatment) seems to be more dependent on the high stomatal ABA
response.
Notably, the CE3027 shows better drought tolerance than CE3130 in previous observations, but this
difference was not seen in calculated gs. However, this calculated gs was only estimated by using
morphological characteristics.
Figure 12. Calculated stomatal conductance in different plant lines with different combination of StCDF1 and
StFLORE. The Calculated stomatal conductance in CE3130 (1.2/1.3), RNAi CDF1 CE3130, 35S CDF3 CE3130, CE3027
(1.1/1.1) and 35S CDF1.2 CE3027 plant lines.

3.5 Phenotypic response to drought
To have a better view of the performance of selected genotypes under drought condition, we
examined 8 genotypes (table 1-2) under 3 weeks drought treatment. The plants’ performance under
drought and control treatments were tested and recorded in photos. Among the non-transgenic
plant lines, the CE3027(1.1/1.1) plants and CE605(1.1/1.2) plants stand out for their better drought
tolerance. Both of these two lines had no significant difference between control treatment and
drought treatment (figure 13, A-B). It seems like the 7bp insertion in StCDF1.2 did not affect drought
sensitivity much. Unlike StCDF1.2, the heavily truncated allele StCDF1.3 seems to have a more
negative effect in general. Although the RHXE 30 (1.3/1.3) plants also seem less affected by drought,
the plants growth was largely repressed (figure 14, A-B). Even after almost 10 weeks growing in the
glasshouse, the leaf size and plant height of RHXE-30 were far smaller than in other plant lines. The
drought sensitivity of CE3130 (1.2/1.3) plants was very similar to CE630 (1.1/1.3) plants’. Both of
these two StCDF1.3 allele carriers’ (CE3130 and CE630) plants died very quickly under drought
condition.
Figure 13. drought response in non-transgenic plant lines. (A) from left to right, the about 7 weeks old diploid potato
plants CE3027(1.1/1.1), CE605(1.1/1.2), CE3130(1.2/1.3), CE630(1.1/1.3) and RHXE-30(1.3/1.3) was treated as control.
The CE605(1.1/1.2) plants were 5 days younger than the other plants. The plant growth of RHXE-30 was strongly
repressed and adaxial leaf side turns to purple even at young age. (B) from lift to right, the about 7 weeks diploid potato
CE3027(1.1/1.1), CE605(1.1/1.2), CE3130(1.2/1.3), CE630(1.1/1.3) and RHXE-30(1.3/1.3) was treated under drought for
3 weeks.

Knocking down StCDF1 in CE3130 plants improved the drought tolerance significantly (figure 14). The
RNAi CDF1 CE3130 plants also became taller than CE3130 (1.2/1.3) plants. However, overexpressing
StCDF3 in CE3130 results in the dwarf phenotype. Apart from that, the 35S CDF3 CE3130 plants also
seem to be more tolerant to drought than CE3130 plants. Unlike 35S CDF3 CE3130 plants,
overexpressing StCDF1.2 in CE3027 did not affect plants growth. Moreover, the 35S CDF1.2 CE3027
plants had very similar drought response to CE3027 plants.
In conclusion, the results indicate that the StCDF1.3 can induce a negative effect on plant growth and
drought tolerance. The StCDF1.1 and StCDF1.2 may have comparably slightly more positive effects
on drought tolerance than StCDF1.3. Moreover, the high drought sensitivity in CE630 and CE3130
suggests the StCDF1.3 allele is dominant over the StCDF1.1 and StCDF1.2 allele. Overexpressing
StCDF3 seems to be able to affect plant growth but also induce positive effect on drought response.
Considering the previous results in this study, the improvement of drought response in 35S CDF3
CE3130 may result from very low stomatal density.
Unfortunately, due to timing issues, plants were quite old (7 weeks old) before drought treatment.
Therefore, some early genotype (CE3130, CE630) control plants shows less vigor and are unhealthy
after another 3 weeks control treatment. The CE605 (1.1/1.2) plants in figure 14 were 9 weeks old (1
week younger than other plant), because we had to regrow them after roof leakage in the glasshouse
destroyed the original CE605 plants.
Figure 14. Drought response in transgenic plant lines and their background plants. (A) from lift to right, the about
7 weeks old diploid potato plants CE3130(1.2/1.3), RNAi CDF1 CE3130, 35S CDF3 CE3130, CE3027(1.1/1.1) and 35S
CDF1.2 CE3027 were treated as control. (B) from lift to right, the about 7 weeks diploid potato CE3130(1.2/1.3),
RNAi CDF1 CE3130, 35S CDF3 CE3130, CE3027(1.1/1.1) and 35S CDF1.2 CE3027 were treated under drought for 3
weeks.

4. Discussion
4.1 Novel functions of StCDFs in regulating plant drought tolerance
In previous study, we have found that knocking down StCDF1 enhanced drought tolerance (Bachem
et al. unpublished data). Therefore, we further look into how drought tolerance has been improved
in this study. We examined 8 genotypes with different StCDF1 alleles combination or different
StCDFs expression levels. Our results showed transpiration related stomatal characters, such as
stomatal size, stomatal initiation, and stomatal ABA responsiveness are significantly affected by the
combinations of different StCDF1 allelic variants or the expression level of StCDFs. These results
suggest a clear participation of StCDFs in regulating transpiration in several ways to affect drought
tolerance.
The well-documented function of CDF1 is regulating photoperiodic flowering-time in Arabidopsis,
by affecting the diurnal rhythm of CONATANS (CO) expression and consequently the expression of
FLOWERING LOCUS T (FT) (Fornara et al. 2009; Corrales et al, 2017). In potato, the homologs of FT
did more than inducing flowering. It was shown that an FT homolog called StSP6A acts as a
tuberization signal in potato, which regulated by two light responded proteins called StGI and
StFKF1 via StCDFs and StCO (Abelenda, Navarro, & Prat, 2014).
Beside the reproduction, the circadian clock also responsible for regulating many other processes
including stomatal movements (Hubbard, & Webb, 2011). In the day time, photosynthesis requires
stomatal opening to assimilate carbon dioxide. In Arabidopsis, FT has been reported as a stomatal
opening signal which regulated by CO (Kinoshita et al. 2011). CDF also involved in this regulation by
mediating CO and FT expression. However, it is also possible that CDF more directly mediate
stomatal movements. Several Dof binding sites have been found in the promoter of guard cell
specific genes and these Dof binding motifs have been shown to be essential for their expression
(Cominelli et al.,2011). Although Cominelli et al. suggested in their analysis that Arabidopsis CDFs
were not involved in regulation of this stomatal specific MYB transcription factor, it should be
noted that the quadruple CDF mutant line that this work was based on, remains considerably leaky
with respect to StCDF4 & 5 expression (Dr. Fabio Fornara, personal communication). This finding
suggest Dof protein may have direct effect on stomatal movements and CDFs still could be good
candidates.
To adapt to drought, decreasing water loss while maintaining water uptake is necessary for plants.
For potato, canopy temperature (transpiration) has been used as selection factor in breeding high
drought tolerance variety (Prashar et al., 2013; Obidiegwu, Jones, & Prashar, 2015). Our results
show that the combinations of different StCDF1 allelic variations result in diverse transpiration
levels under ABA treatment. Furthermore, the low transpiration level plant lines are mostly in
agreement with the plant lines which has better performance under drought treatment. Therefore,

here, we propose a novel function of StCDFs in regulating drought response by affecting
transpiration.
4.2. StCDF1 has an antagonistic effect on stomatal ABA responsiveness
ABA is a stress-response hormone, which induces rapid stomatal closure. Under stress conditions,
the stomatal ABA sensitivity has a crucial impact on minimizing water loss by transpiration (Lawson
& Blatt, 2014). Our study demonstrates that StCDF1 has a general repressive effect on stomatal ABA
responsiveness, and this effect was consistent in CE3130, RNAi CDF1 CE3130, CE3027 and 35S
CDF1.2 CE3027. Moreover, the strong negative effect on stomatal ABA responsiveness induced by
overexpressing StCDF1.2 could not be induced by overexpressing StCDF3 as same.
Stomatal opening and closure induced by different signals. As figure 15 shows, light induce stomatal
opening and the stomatal opening could be repressed by ABA. However, overexpressing StCDF1.2
prevent stomata closure by ABA. A previous study shows a novel function in regulating stomatal
movement by FT. This is a function that is independent of the regulating flowering and tuberization.
Researchers nominated FT as a cell autonomously regulated timekeeper for regulating opening and
closure of stomatal guard cells (Kinoshita et al. 2011). Their results suggest that FT provides a
positive effect on stomatal opening via the activity of H+-ATPase in guard cells (figure 15). Based on
our results, this model possibly also be applied to potato. However, we suggest that the central
regulator is more likely to be StCDF1. In potato, the StCDF3 has a redundant gene function for
StCDF1. The StCDF3 also represses StCO and indirectly regulates expression of FT homologs’ such as
the tuberization signal StSP6A. However, overexpressing StCDF3 in CE3130 (1.2/1.3) did not reduce
the stomatal response to ABA as overexpressing StCDF1.2 in CE3027 did. Therefore, the stomatal
Figure 15. Overview of H+-ATPase-mediated stomatal movement. Transcriptional regulation of flowering is possibly
involved in activation of the H+-ATPase to open stomata (orange arrows); ABA inhibition of the H+-ATPase to indcuce
stomatal closure (red arrows). (Kinoshita et al. 2011; Wang, Y., Shimazaki, K. I., & Kinoshita, T. 2014)

opening signal could only be induced by StCDF1. Thus, we propose that StCDF1 play a non-
redundant role in regulating stomatal movement.
Moreover, both late genotypes ---- RNAi CDF1 CE3130 and CE3027 had stomata that are very
sensitive to ABA. In these plants, StCDF1 protein was expected to absent, due to RNAi or protein
degradation. Our results suggest having less StCDF1 protein enhanced stomatal ABA sensitivity and
is consistent with the discussion outlined above. It is interesting to note, that expression of the long
non-coding RNA, StFLORE1 that coincides with the StCDF1 locus, is increased in StCDF1 RNAi
knockdown in CE3130. This opens up the possibility that this lncRNA transcript may also have a role
in stomatal ABA responsiveness regulation.
4.3 The role of StFLORE in drought response
Several natural antisense lncRNAs (NATs) have been reported to be involved in phytohormone-
mediated response, via regulating gene expression at multiple levels (Wang et al., 2017). Such
regulation mechanisms include i) antisense lncRNAs that complex with other target transcripts
(Csorba et al., 2014). ii) act as a linker between proteins. iii) change chromatin configuration
(Helliwell et al., 2011) or modify miRNA expression (Pai et al. 2017) (see Figure 17). Another recent
finding was an Arabidopsis lncRNA called DRIR which is also induced by ABA treatment allowing
decreasing transpiration rate and the ability to up-regulate other genes related to drought stress
such as aquaporins, ARSK1 (root-specific kinase) and others (Qin et al., 2017). Moreover, a study
from Cagirici in 2017, showed that in wheat, lncRNAs can mimic the role of a miRNA1439 to
increase aquaporin translation under drought stress. Since it is known that aquaporin is able to
induce stomata closing under drought stress (Cagirici et al., 2017). These findings indicate firstly
that lncRNAs activities may be complex and secondly that lncRNA regulations is frequenctly
accociated with regulation of stress responses, light signaling and regulation of flowering (Wang et
al., 2014)
In chapter 3.2, we described the major finding that StFLORE expression response to ABA treatment
in vitro plants. In RNAi CDF1 CE3130, ABA induced StFLORE expression in both root and apex, while
root tissues had generally higher expression in both control and ABA conditions. Thus, it is possible
that StFLORE transcript in both apex and root or mainly transcript in root and then being transport
Figure 16. Different roles of antisense lncRNAs in stress response in plants.

to apex tissue. In previous study, we have found that StCDF1 seems able to repress StFLORE
expression, probably by binding to the multiple [A]3G binding sites present in the promoter of this
transcript (data not shown). In chapter 4.2, we discussed that the negative role played by StCDF1 in
regulating stomatal response to ABA. Moreover, the ABA can enhance the StFLORE expression. It is
possible that StFLORE was involved in inducing stomatal closure under influence of ABA, with
StCDF1 protein acting as a repressor in this process.
Several models can be used to predict how StFLORE enhance stomatal ABA sensitivity. For example,
it is possible that StFLORE act as a linker for recruiting other ABA response proteins to trigger
stomatal closure etc. However, the most urgent research question is whether StFLORE actually
inducing stomatal closure or other drought tolerance phenotypes. Therefore, building up the
construct that overexpressing StFLORE in CE3130 background is necessary for providing consistent
evidence.
Notably, the ABA response has been found only in RNAi CDF1 CE3130 in vitro plants. We expected
CE3027 should has similar response to ABA, since it supposes to have two copies of functional
StFLORE. However, we did not observe the ABA response in CE3027. It is possible that due to the
circadian regulation of all the genes and transcripts involved in this complex process, only a detailed
time-course expression analysis will reveal the actual regulation mechanisms in all genotypes
tested.

4.4 StCDFs affect stomatal features in multiple ways
4.4.1 Stomatal initiation
In this study, the reduced stomatal index has been found only in RNAi CDF1 CE3130 and it suggest
that, in addition to its roles in diurnal stomatal dynamics, StCDF1 may affect stomatal formation via
regulating StFLORE. Based on the current knowledge of stomatal development, the reduced
stomatal index is very likely to be a result of fewer protodermal cells transforming into meristemoid
mother cells (MMC) (figure 18). In Arabidopsis, SPEECHLESS (SPCH) induces this transformation. In
SPCH loss-of-function mutants, stomata initiation will be inhibited, and only epidermal pavement
cells will develop (Lampard, MacAlister, & Bergmann, 2008; Bergmann, Lukowitz, & Somerville,
2004; Qi, & Torii, 2018). ABA has been shown to prevent stomatal initiation via prolonged
expression of SPCH and MUTE (Tanaka et al., 2013; Le, et al., 2014). Since StFLORE response to ABA
and this response has been found in leaves, it is possible that StFLORE participates in the
prevention stomatal initiation. In future study, overexpressing StFLORE in CE3130 background
would provide more insight of this potential regulation.
4.4.2 Epidermal cells expansion
Besides reducing stomatal index, cell expansion is another way to obtain lower stomatal density.
We have found enlarged stomata and pavement cells in 35S CDF1.2 CE3027 and 35S CDF3 CE3130
plants. In both of the two overexpressing lines, epidermal cells are almost as twice as big in the
wild-type backgrounds. Moreover, the stomatal densities in these lines is around half of their
background plant lines. As mentioned in the introduction, larger stomatal size and lower stomatal
density have the positive effect on reducing transpiration. Although overexpressing StCDF3 and
StCDF1.2 can result in epidermal cell expansion (larger stomata, lower density) and indeed both
Figure 17. The control of stomatal development. Stomatal initiation and differentiation are determined by three
essential transcription factors, SPCH, MUTE, and FAMA (Kalve, & Beemster, 2014).

genotypes had a drought tolerance phenotype, the latter genotype lead to ABA insensitivity
stomatal phenotype, while overexpressing StCDF3 lead to dwarf phenotype. Thus, overexpressing
StCDFs seems induce epidermal cell expansion and it has positive effects in reducing transpiration,
but the negative effects on plant fitness should also be noted.
However, the cell expansion in this study also showed that StCDFs may have more complicated
functions other than regulating tuberization and transpiration. So far, not much single gene
overexpression plant or single gene mutant were reported to have similar stomatal pattern like
this. The most similar stomatal pattern we found is in polyploidy (figure 19) (Gu et al., 2016).
Moreover, Tai and Gardi has also report similar pattern that Polyploid tomato had fewer and larger
stomata and ordinary epidermal cells per leaf unit area than diploid tomato (Tal & Gardi, 1976). The
polyploid epidermal cells expansion also has been linked with lower transpiration (Chen & Tang,
1945). Thus, it is like that StCDFs has diverse effects on plant development.
Figure 18. Epidermal cells expansion in Chinese cabbage and potato (Gu et al., 2016).

4.5 In potato, StCDF1-StFLORE connects tuberization and transpiration.
Potato has always been recognized as a drought sensitive crop (Porter et al., 1999). The most
common explanation for this characteristic has been its shallow and sparse root system, negatively
affecting water uptake (Iwama and Yamaguchi, 2006; Jefferies, 1993). Apart from access to water
resources in the soil, reducing water-loss though transpiration has been considered as another
possible way to improve water stress resilience in potato (Wang and Clarke, 1993; Prashar et al.,
2013). In this study, the better phenotypic response to drought in different genotypes are mostly in
agreement with the low calculated stomata conductance (gs) genotypes. These results show that
reducing transpiration is a promising way to improve drought tolerance in potato.
The StCDFs have been known for being an important mediator between the circadian clock and the
tuberization signal (StSP6A) in recent years. Tuberization requires photosynthesis products and
photosynthesis requires stomatal opening to obtain CO2. Here, we have found the circadian clock
genes StCDFs also mediate transpiration regulation. Based on data present in this work, we
proposed a model of how the circadian clock is connected to transpiration related characteristics.
(see figure 19).
As figure 15 shows, light drives StCDF1 to induce stomatal opening, while the stress hormone ABA
represses the stomatal opening to reduce the water loss. The competition between these two
pathways determines plant performance under drought condition. In our study, we found the
StCDF1-StFLORE locus determines which pathway takes the priority under drought condition. These
findings shed new light on the key role played by StCDFs in optimizing CO2 gain against water loss in
stressful environments.
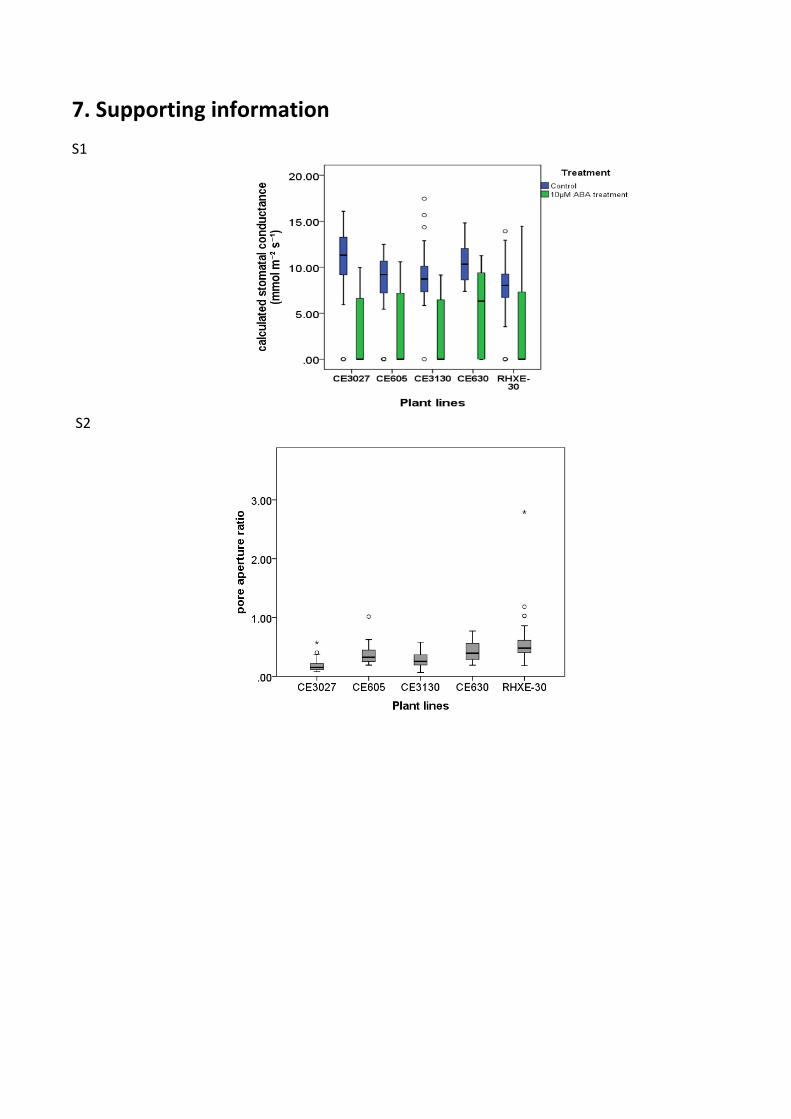
Our results show the CE630 (1.1/1.3) and RHXE30 (1.3/1.3) plants were more sensitive to drought
than others. Their stomatal also seems less sensitive to ABA than other genotypes (S2). As
mentioned in the introduction, the 200bp insertion in StCDF1.3 helps the protein escape from
degradation to induce earliness. Considering StCDF1 can induce stomatal opening, the decreased
Figure 19. Proposed model on StCDF1 cooperates with its long non-coding partner StFLORE in regulating drought
response related stomatal characteristic. When StGI and StFKF1 do not bind to StCDF1, StCDF1 leads to stomatal
opening (figure 6) and repress StFLORE expression (data not shown). The StFLORE expression could be induced by
stress hormone ABA (figure 7) and it may have a beneficial effect on drought response related stomatal
characteristics.

degradation of StCDF1.3 may lead to a higher transpiration rate. Moreover, the 200bp insertion in
StCDF1.3 also very likely results in non-functional StFLORE. Although the function of StFLORE is
unclear, StFLORE is very likely to have positive effect on drought response which is regulated by the
stress hormone ABA.
Whereas knocking down StCDF1 in CE3130(1.2/1.3) and the CE3027(1.1/1.1) background plants
resulted in a delayed tuberization, they also had better drought tolerance. In these two plants,
StCDF1 protein levels were expected to be lower, due to RNAi or protein degradation. The delayed
tuberization mechanism has been well explained by Kloosterman et al., 2013 and Navarro et al.,
2011. Our results indicated StCDF1 also acts as a stomatal opening signal and fewer StCDF1 proteins
lead to better stomatal sensitivity to ABA for reducing transpiration under stress condition. Beside
the reduced stomatal opening signal, the upregulation of StFLORE in RNAi CDF1 CE3130 may
contribute to beneficial characteristics to drought response. Therefore, these genotypes are more
tolerant to drought due to the higher priority of water conservation.
In conclusion, this study provides new information about the function StCDF1 as a key factor that
displays multiple roles related to plant responses to drought stress and the developmental program
underlying the transition from vegetative to reproductive phase.
Figure 20. Drought response strategy in potato.

5.Conclusion
In this research, we proposed a model of how the circadian clock gene StCDF1 is connected to
transpiration related characteristics to influence drought response. In this model, StCDF1 has a
negative effect on drought response by inducing an ABA insensitive stomata phenotype. The
upregulated StFLORE expression under ABA treatment shows that StFLORE is likely to have a
positive effect on drought tolerance under ABA influence. However, more research is needed to
substantiate this model. For future research overexpressing StFLORE in CE3130 as well as CE3027
plants without a functional StFLORE promoter would be more than useful.

6. References
Abelenda, J. A., Navarro, C., & Prat, S. (2014). Flowering and tuberization: a tale of two nightshades. Trends
in plant science, 19(2), 115-122.
Alva, A., Fan, M., Qing, C., Rosen, C., & Ren, H. (2011). Improving nutrient-use efficiency in Chinese potato production: experiences from the United States. Journal of crop improvement, 25(1), 46-85.
Cagirici, H. B., Alptekin, B., & Budak, H. (2017). RNA Sequencing and Co-expressed Long Non-coding RNA
in Modern and Wild Wheats. Scientific reports, 7(1), 10670.
Cai, S., Chen, G., Wang, Y., Huang, Y., Marchant, D. B., Wang, Y., ... & Nevo, E. (2017). Evolutionary conservation of ABA signaling for stomatal closure. Plant physiology, 174(2), 732-747.
Chen, S. L., & Tang, P. S. (1945). Studies on colchicine-induced autotetraploid barley. III. Physiological studies. American
Journal of Botany, 177-179.
Christodoulakis, N. S., Menti, J., & Galatis, B. (2002). Structure and Development of Stomata on the Primary Root of Ceratonia siliqua L. Annals of Botany, 89(1), 23-29.
Corrales, A. R., Carrillo, L., Lasierra, P., Nebauer, S. G., Dominguez‐Figueroa, J., Renau‐Morata, B., ... &
Medina, J. (2017). Multifaceted role of cycling Dof Factor 3 (CDF3) in the regulation of flowering time and
abiotic stress responses in Arabidopsis. Plant, cell & environment, 40(5), 748-764.
Csorba, T., Questa, J. I., Sun, Q., & Dean, C. (2014). Antisense COOLAIR mediates the coordinated
switching of chromatin states at FLC during vernalization. Proceedings of the National Academy of Sciences,
111(45), 16160-16165.
Desikan, R., Horák, J., Chaban, C., Mira-Rodado, V., Witthöft, J., Elgass, K., ... & Neill, S. J. (2008). The histidine
kinase AHK5 integrates endogenous and environmental signals in Arabidopsis guard cells. PLoS One, 3(6),
e2491.
Doheny-Adams, T., Hunt, L., Franks, P. J., Beerling, D. J., & Gray, J. E. (2012). Genetic manipulation of
stomatal density influences stomatal size, plant growth and tolerance to restricted water supply across a
growth carbon dioxide gradient. Philosophical Transactions of the Royal Society B: Biological
Sciences, 367(1588), 547-555.
Eisele, J. F., Fäßler, F., Bürgel, P. F., & Chaban, C. (2016). A Rapid and Simple Method for Microscopy-Based
Stomata Analyses. PloS one, 11(10), e0164576.
Fanourakis, D., Giday, H., Milla, R., Pieruschka, R., Kjaer, K. H., Bolger, M., ... & Ottosen, C. O. (2014). Pore size regulates operating stomatal conductance, while stomatal densities drive the partitioning of conductance between leaf sides. Annals of botany, 115(4), 555-565.
FAOSTAT (2014). From: http://www.fao.org/faostat/en/?#data/QC
Farber, M., Attia, Z., & Weiss, D. (2016). Cytokinin activity increases stomatal density and transpiration rate in tomato. Journal of experimental botany, erw398.
Fornara F., Panigrahi K.C.S., Gissot L., Sauerbrunn N., Rühl M., Jarillo J.A. & Coupland G. (2009) Arabidopsis DOF transcription factors act redundantly to reduce CONSTANS expression and are essential for a photoperiodic flowering response. Developmental Cell 17, 75–86.
Franks, P. J., & Farquhar, G. D. (2001). The effect of exogenous abscisic acid on stomatal development, stomatal mechanics, and leaf gas exchange in Tradescantia virginiana. Plant Physiology, 125(2), 935-942.GAIN (2010). Global Agricultural Information Network of the USDA Foreign Agricultural Service. GAIN Report No RS1060.

Franks, P. J., & Farquhar, G. D. (2007). The mechanical diversity of stomata and its significance in gas-exchange control. Plant physiology, 143(1), 78-87.
Godfray, H. C. J., Beddington, J. R., Crute, I. R., Haddad, L., Lawrence, D., Muir, J. F., ... & Toulmin, C. (2010). Food security: the challenge of feeding 9 billion people. science, 327(5967), 812-818.
Gu, A. X., Zhao, J. J., Li, L. M., Wang, Y. H., Zhao, Y. J., Hua, F., ... & Shen, S. X. (2016). Analyses of
phenotype and ARGOS and ASY1 expression in a ploidy Chinese cabbage series derived from one
haploid. Breeding science, 66(2), 161-168.
Helliwell, C. A., Robertson, M., Finnegan, E. J., Buzas, D. M., & Dennis, E. S. (2011). Vernalization-repression
of Arabidopsis FLC requires promoter sequences but not antisense transcripts. PloS one, 6(6), e21513
Hubbard, K. E., & Webb, A. A. (2011). Circadian rhythms: FLOWERING LOCUS T extends opening
hours. Current Biology, 21(16), R636-R638.
Imaizumi, T., Schultz, T. F., Harmon, F. G., Ho, L. A., & Kay, S. A. (2005). FKF1 F-box protein mediates cyclic degradation of a repressor of CONSTANS in Arabidopsis. Science, 309(5732), 293-297.
Iwama K., Yamaguchi J. (2006). Abiotic stresses, in Handbook of Potato Production, Improvement and
Postharvest Management, eds Gopal J., Khurana S. M., editors. (New York, NY: Food Product Press; ), 231–
278.
Jefferies R. A. (1993a). Cultivar responses to water-stress in potato: effects of shoot and roots. New
Phytol. 123, 491–498.
Jones, H. G. (1992). Plant and microclimate: a quantitative approach to environmental plant physiology (No. 581.5222 J6 1992).
Jones, H. G. (1998). Stomatal control of photosynthesis and transpiration. Journal of Experimental Botany,
387-398.
Kalve, S., De Vos, D., & Beemster, G. T. (2014). Leaf development: a cellular perspective. Frontiers in plant
science, 5, 362.
Kondorosi, E., Roudier, F., & Gendreau, E. (2000). Plant cell-size control: growing by ploidy?. Current opinion
in plant biology, 3(6), 488-492.
Lawson, T., & Blatt, M. (2014). Stomatal size, speed and responsiveness impact on photosynthesis and water
use efficiency. Plant Physiology, pp-114.
Le, J., Zou, J., Yang, K., & Wang, M. (2014). Signaling to stomatal initiation and cell division. Frontiers in plant
science, 5, 297.
Lloyd, F. E. (1908). The physiology of stomata (No. 82). Carnegie Institution of Washington.
Obidiegwu, J. E., Bryan, G. J., Jones, H. G., & Prashar, A. (2015). Coping with drought: stress and adaptive
responses in potato and perspectives for improvement. Frontiers in plant science, 6, 542.
Pai, A. A., Henriques, T., McCue, K., Burkholder, A., Adelman, K., & Burge, C. B. (2017). The kinetics of pre-
mRNA splicing in the Drosophila genome and the influence of gene architecture. Elife, 6
Porter G. A., Opena G. B., Bradbury W. B., McBurnie J. C., Sisson J. A. (1999). Soil management and
supplemental irrigation effects on potato: I. Soil properties, tuber yield, and quality. Agron. J. 91, 416–425.
Prashar, A., Yildiz, J., McNicol, J. W., Bryan, G. J., & Jones, H. G. (2013). Infra-red thermography for high
throughput field phenotyping in Solanum tuberosum. PLoS One, 8(6), e65816.
Qi, X., & Torii, K. U. (2018). Hormonal and environmental signals guiding stomatal development. BMC biology, 16(1), 21.
Qin, T., Zhao, H., Cui, P., Albesher, N., & Xiong, L. (2017). A Nucleus-localized Long Non-Coding RNA
Enhances Drought and Salt Stress Tolerance. Plant physiology, pp-00574.

Riboni, M., Galbiati, M., Tonelli, C., & Conti, L. (2013). GIGANTEA enables drought escape response via
abscisic acid-dependent activation of the florigens and SUPPRESSOR OF OVEREXPRESSION OF
CONSTANS1. Plant physiology, 162(3), 1706-1719.
Saibo, N. J., Vriezen, W. H., Beemster, G. T., & Van Der Straeten, D. (2003). Growth and stomata development of Arabidopsis hypocotyls are controlled by gibberellins and modulated by ethylene and auxins. The Plant Journal, 33(6), 989-1000.
Skirycz A, Inze D. 2010. More from less: plant growth under limited water. Current Opinionin Biotechnology21: 197–203.
Tal, M., and I. Gardi. "Physiology of polyploid plants: water balance in autotetraploid and diploid tomato under low and
high salinity." Physiologia plantarum 38.4 (1976): 257-261.
Wang H., Clarke J. M. (1993). Relationship of excised-leaf water-loss and stomatal frequency in wheat. Can. J. Plant
Sci. 73, 93–99.
Wang, J., Meng, X., Dobrovolskaya, O. B., Orlov, Y. L., & Chen, M. (2017). Non-coding RNAs and their roles in stress
response in plants. Genomics, proteomics & bioinformatics.
Wang, Y., Shimazaki, K. I., & Kinoshita, T. (2014). Multiple roles of the plasma membrane H+-ATPase and its regulation.
In The Enzymes (Vol. 35, pp. 191-211). Academic Press.
Wang, H., Chung, P. J., Liu, J., Jang, I. C., Kean, M. J., Xu, J., & Chua, N. H. (2014). Genome-wide identification of long
noncoding natural antisense transcripts and their responses to light in Arabidopsis. Genome research.
7. Supporting information
S1
S2





























