Embed Size (px)
Citation preview

The phylogenetic position of Peucedanum sensu lato and allied
genera and their placement in tribe Selineae (Apiaceae, subfamily
Apioideae)
K. Spalik1, J.-P. Reduron
2, and S. R. Downie
3
1Department of Plant Systematics and Geography, Warsaw University, Warszawa, Poland2Conservatoire botanique de la Ville de Mulhouse, Mulhouse, France3Department of Plant Biology, University of Illinois at Urbana-Champaign, Urbana, Illinois, USA
Received March 3, 2003; accepted July 29, 2003Published online: January 15, 2004� Springer-Verlag 2004
Abstract. ITS sequences of members of Peuceda-num and its segregates (Cervaria, Holandrea,Imperatoria, Oreoselinum, Pteroselinum, Thysseli-num, Tommasinia, and Xanthoselinum), as well asrepresentatives of other members of the previouslyidentified ‘Angelica’ and ‘Arracacia’ clades, wereanalysed using distance, maximum likelihood, andmaximum parsimony methods in order to establishtheir phylogenetic relationships. The ‘Angelica’ and‘Arracacia’ clades comprise a strongly supportedmonophyletic group that is formally recognised astribe Selineae. The genus Holandrea, expanded toinclude Cervaria caucasica and Peucedanum acha-icum, is distantly related to the other peucedanoidgenera. The remaining segregates are closely relatedto one another and to Peucedanum sensu stricto.The genera Seseli and Selinum are polyphyletic.Members of the genus Angelica form a single cladethat also includes representatives of Selinum andSphenosciadium. New combinations in Holandrea(H. caucasica, H. achaica) and Angelica (A. capi-tellata) are proposed, and Selinum pyrenaeum isrecognised in Angelica (A. pyrenaea).
Key words: Peucedanum, Apiaceae, Umbelliferae,Selineae, phylogeny, evolution, comb. nov.
Apiaceae (Umbelliferae) include about 450genera and 3700 species (Pimenov and Leonov1993). However, these species are not evenlydistributed among the genera: 41% of thegenera are monotypic and 26% comprise onlytwo or three species each. Sixty percent of thetotal number of species are placed in arelatively small number of large genera thatcomprise more than 20 species each (Spaliket al. 2001). These genera include Ferula (170spp.), Ligusticum (40–50 spp.), Seseli (100–120spp.), and Peucedanum (100–120 spp.), all ofwhich are polyphyletic (Katz-Downie et al.1999, Downie et al. 2000b). Consequently, theyare poorly defined which makes their identifi-cation difficult.
The genus Peucedanum is distributed inEurasia and Africa (Pimenov and Leonov1993). Its members are characterised by flat-tened fruits with more or less developed lateralwings. It has long been regarded as heteroge-neous and several authors have postulated itsdivision into smaller, presumably more naturalunits (Calestani 1905, Leute 1966, Pimenov1987, Frey 1989). Pimenov and Leonov (1993)
Plant Syst. Evol. 243: 189–210 (2004)DOI 10.1007/s00606-003-0066-2

even suggested that it should be restricted tothe 8–10 species comprising sect. Peucedanum.Indeed, many species once recognised inPeucedanum have found other affinities, some-times very distant ones. For instance, P. tere-binthaceum (Fisch. ex Trevir.) Fisch. ex Turcz.,along with several other species, were trans-ferred to Kitagawia (Pimenov 1986). Based onmolecular data, P. terebinthaceum—thenomenclatural type of Kitagawia—is sister toPleurospermum hookeri (Downie et al. 2000b)that at present is recognised in tribe Pleuro-spermeae (Downie et al. 2001). Two Socotranspecies of Peucedanum have appeared to beclosely related to another Socotran endemic,Nirarathamnos asarifolius, the entire cladebeing a sister group to tribe Echinophoreae(Downie et al. 2000a, b). Moreover, Peuceda-num sensu Drude (1897–1898) encompassedmany North American umbellifers that arenow placed in Lomatium.
Even the European members of Peuceda-num are suspected of being distantly related toone another and several species have beentreated within segregate genera. For example,in the Flora Europaea account (Tutin et al.1968), a broad concept of Peucedanum wasadopted, with the exclusion of those speciesplaced in segregate genera Imperatoria andTommasinia. In contrast, Reduron et al. (1997)recognised a narrow definition of Peucedanumand confirmed restitution of Cervaria, Imper-atoria, Oreoselinum, Pteroselinum, Thysseli-num, and Xanthoselinum, as well as describedthe new genus Holandrea, with three includedspecies (Table 1). They based their treatmenton morphological and phytochemical data;however, they did not perform a formalphylogenetic analysis of these data.
Peucedanum is a member of the ‘Angelica’clade sensu stricto, belonging to the largeapioid superclade (Downie et al. 2001).Throughout this paper, names in single quo-tation marks refer to informal groups,whereas those without quotation marksdenote genera or postulated genera. Closelyrelated to the ‘Angelica’ clade is the ‘Arracacia’clade, the latter comprising some ten genera of
meso-American distribution. In some analyses,the ‘Arracacia’ clade arises within the ‘Angel-ica’ clade (Plunkett et al. 1996, Downie et al.1998). Downie et al. (2001) suggested that theformer may eventually be subsumed within thelatter. Both groups comprise many genera thatwere traditionally placed in Drude’s (1897–1898) tribe Peucedaneae subtribes Angelicinaeand Ferulinae, or tribes Peucedaneae andAngeliceae in the classification system ofPimenov and Leonov (1993).
The aim of this paper is to verify thetaxonomic division of Peucedanum sensu latoagainst a phylogeny inferred from nuclearribosomal DNA internal transcribed spacer(rDNA ITS) sequences. We include the typespecies of these segregate genera and a broadrepresentation of the ‘Angelica’ and ‘Arraca-cia’ clades in order to find the closest relativesof these segregates.
Material and methods
Taxon sampling. The total number of accessionsexamined was 107. We included almost all availableITS sequences of members of the ‘Arracacia’ cladeand Old World members of the ‘Angelica’ clade(Table 2). Since the phylogenetic affinities of theNorth American members of the ‘Angelica’ cladewere the subject of two recent studies (Downie et al.2002, Sun et al. in press), we considered only arepresentative sample of these taxa. To these, weadded 30 new accessions including the type speciesof Peucedanum and its segregates, as well as severalmembers of the putatively related genera Angelica,Seseli, and Selinum including their type species. Theaccessions denoted as Peucedanum sp. and Thys-selinum sp. represent two taxa from France whoseaffinities are unclear.
Since Thysselinum palustre was excluded fromthe ‘Angelica’ clade in an earlier molecular analysis(Katz-Downie et al. 1999), we included a broadrepresentation of outgroup genera from the apioidsuperclade, such as Azilia, Cnidium, Coriandrum,Ferula, Opopanax, Prangos, Smyrniopsis, andSphaenolobium. The trees were rooted with Kruberaperegrina (¼Capnophyllum dichotomum in Downieet al. 1998) that in one of the earlier analyses wasbasal to this part of the apioid superclade (Katz-Downie et al. 1999).
190 K. Spalik et al.: The phylogenetic position of Peucedanum sensu lato and allied genera

Laboratory procedures. Total DNA was ex-tracted from herbarium specimens using the PlantDNeasy Extraction Kit (Qiagen, Inc., Valencia,CA). Details of PCR amplifications are provided inDownie et al. (2000a). Each PCR product waselectrophoresed in a 1% agarose gel, stainedwith ethidium bromide, then excised and elutedusing the QIAEX II Gel Extraction Kit (Qiagen).Cycle sequencing reactions were performed usingthe purified PCR product, AmpliTaq DNA poly-merase, and fluorescent Big Dye terminators(Perkin-Elmer Corp., Norwalk, CT). The productswere resolved by electrophoresis using ABI Prism310 or 377 DNA sequencers. Scans were edited andcorrected when necessary.
Data analysis. The DNA sequences werealigned using the program CLUSTAL X (Thomp-son et al. 1997), and manually adjusted. The aligneddata matrix is available from the authors uponrequest. Phylogenetic analyses included neighbour-joining (NJ), maximum parsimony (MP), and max-imum likelihood (ML) methods; all were performedusing PAUP* 4.0b10 (Swofford 1998). For the NJanalysis, we used several substitution models avail-able in PAUP*. Bootstrap analysis was performedfor 1000 pseudoreplicates. For the MP analysis,the ‘inverse constraint’ approach was employed(Catalan et al. 1997). One thousand heuristic
searches were initiated with random addition andtree-bisection-reconnection (TBR) branch swap-ping, with no more than ten shortest trees savedper replicate. These trees were then used as startingtrees for another search, saving 10 000 shortest trees.The consensus of these trees was used as a phyloge-netic constraint in another round of 10 000 searches.This time, only those trees that did not match theconstraint were saved. Since all of these saved treeswere longer than those obtained from the initialsearches, one may suppose that the consensus treesummarises all possible shortest tree topologies.Gaps were treated as missing data; however, theabsence/presence of indels was scored as separatebinary characters, combined with sequence data,and the MP analyses repeated. Bootstrap support(Felsenstein 1985) was estimated using 1000 resam-pled data sets, saving no more than 10 trees perreplicate. Bremer (1984) support values wereobtained using AutoDecay 4.0 (Eriksson 1998).
For the ML analysis, the model of nucleotidesubstitution was selected using Modeltest 3.0(Posada and Crandall 1998) and the likelihoodratio test statistic. The values of the modelparameters estimated with Modeltest were used insubsequent heuristic searches. Due to time limita-tion, bootstrap support values were not calculatedfor the ML trees.
Table 1. Genera segregated from Peucedanum sensu lato analysed in this paper and their type species
Genus Number ofspecies
Type species
Peucedanum L. (sensu stricto) 8–10 Peucedanum officinale L.Cervaria Wolf 4 Cervaria rivini Gaertn. (”Peucedanum cervaria
(L.) Lapeyr.)Holandrea Reduron, Charpinet Pimenov
3 Holandrea carvifolia (Vill.) Reduron, Charpin etPimenov (”Peucedanum carvifolia Vill.)
Imperatoria L. 3 Imperatoria ostruthium L. (” Peucedanumostruthium (L.) W. D. J. Koch)
Oreoselinum Mill. 1 Oreoselinum nigrum Delarbre (”Peucedanumoreoselinum (L.) Moench
Pteroselinum Rchb. 2 Pteroselinum austriacum (Jacq.) Rchb.(”Peucedanum austriacum (Jacq.)W. D. J. Koch)
Thysselinum Adans. 2 Thysselinum palustre (L.) Hoffm.(”Peucedanum palustre (L.) Moench)
Tommasinia Bertol. 1 Tommasinia verticillaris (L.) Bertol.(”Peucedanum verticillare (L.) W. D. J. Koch ex DC.)
Xanthoselinum Schur 1 Xanthoselinum alsaticum (L.) Schur(”Peucedanum alsaticum L.)
K. Spalik et al.: The phylogenetic position of Peucedanum sensu lato and allied genera 191
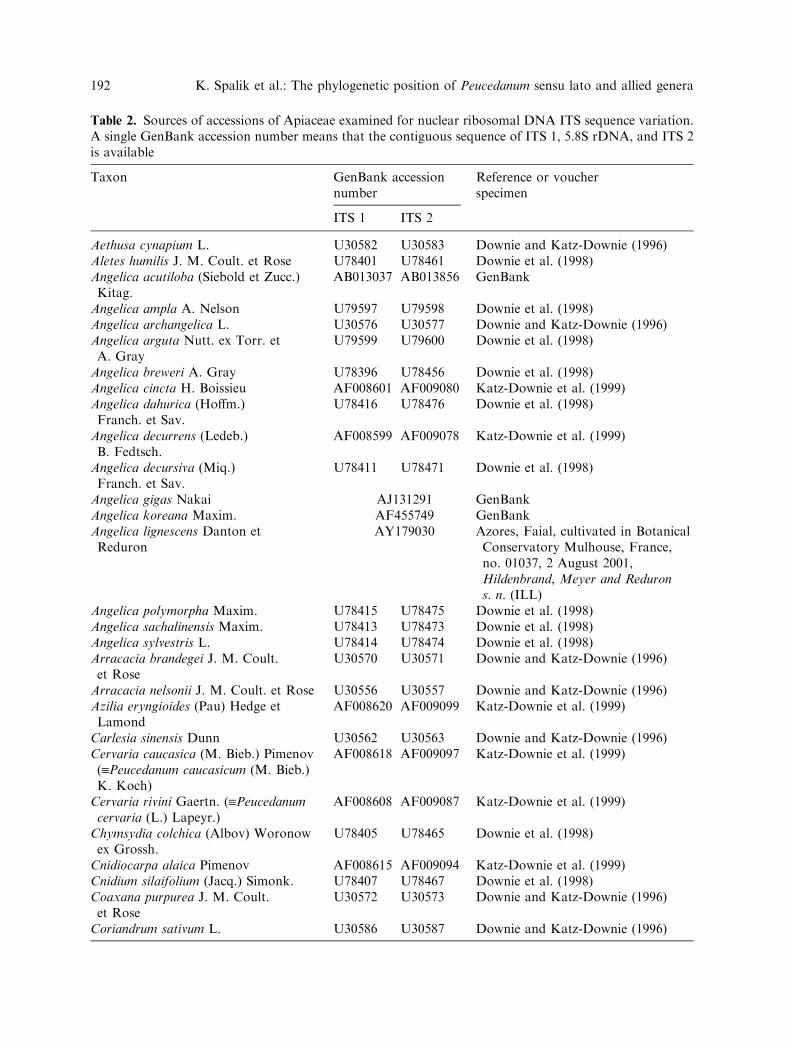
Table 2. Sources of accessions of Apiaceae examined for nuclear ribosomal DNA ITS sequence variation.A single GenBank accession number means that the contiguous sequence of ITS 1, 5.8S rDNA, and ITS 2is available
Taxon GenBank accessionnumber
Reference or voucherspecimen
ITS 1 ITS 2
Aethusa cynapium L. U30582 U30583 Downie and Katz-Downie (1996)Aletes humilis J. M. Coult. et Rose U78401 U78461 Downie et al. (1998)Angelica acutiloba (Siebold et Zucc.)Kitag.
AB013037 AB013856 GenBank
Angelica ampla A. Nelson U79597 U79598 Downie et al. (1998)Angelica archangelica L. U30576 U30577 Downie and Katz-Downie (1996)Angelica arguta Nutt. ex Torr. etA. Gray
U79599 U79600 Downie et al. (1998)
Angelica breweri A. Gray U78396 U78456 Downie et al. (1998)Angelica cincta H. Boissieu AF008601 AF009080 Katz-Downie et al. (1999)Angelica dahurica (Hoffm.)Franch. et Sav.
U78416 U78476 Downie et al. (1998)
Angelica decurrens (Ledeb.)B. Fedtsch.
AF008599 AF009078 Katz-Downie et al. (1999)
Angelica decursiva (Miq.)Franch. et Sav.
U78411 U78471 Downie et al. (1998)
Angelica gigas Nakai AJ131291 GenBankAngelica koreana Maxim. AF455749 GenBankAngelica lignescens Danton etReduron
AY179030 Azores, Faial, cultivated in BotanicalConservatory Mulhouse, France,no. 01037, 2 August 2001,Hildenbrand, Meyer and Redurons. n. (ILL)
Angelica polymorpha Maxim. U78415 U78475 Downie et al. (1998)Angelica sachalinensis Maxim. U78413 U78473 Downie et al. (1998)Angelica sylvestris L. U78414 U78474 Downie et al. (1998)Arracacia brandegei J. M. Coult.et Rose
U30570 U30571 Downie and Katz-Downie (1996)
Arracacia nelsonii J. M. Coult. et Rose U30556 U30557 Downie and Katz-Downie (1996)Azilia eryngioides (Pau) Hedge etLamond
AF008620 AF009099 Katz-Downie et al. (1999)
Carlesia sinensis Dunn U30562 U30563 Downie and Katz-Downie (1996)Cervaria caucasica (M. Bieb.) Pimenov(”Peucedanum caucasicum (M. Bieb.)K. Koch)
AF008618 AF009097 Katz-Downie et al. (1999)
Cervaria rivini Gaertn. (”Peucedanumcervaria (L.) Lapeyr.)
AF008608 AF009087 Katz-Downie et al. (1999)
Chymsydia colchica (Albov) Woronowex Grossh.
U78405 U78465 Downie et al. (1998)
Cnidiocarpa alaica Pimenov AF008615 AF009094 Katz-Downie et al. (1999)Cnidium silaifolium (Jacq.) Simonk. U78407 U78467 Downie et al. (1998)Coaxana purpurea J. M. Coult.et Rose
U30572 U30573 Downie and Katz-Downie (1996)
Coriandrum sativum L. U30586 U30587 Downie and Katz-Downie (1996)
192 K. Spalik et al.: The phylogenetic position of Peucedanum sensu lato and allied genera
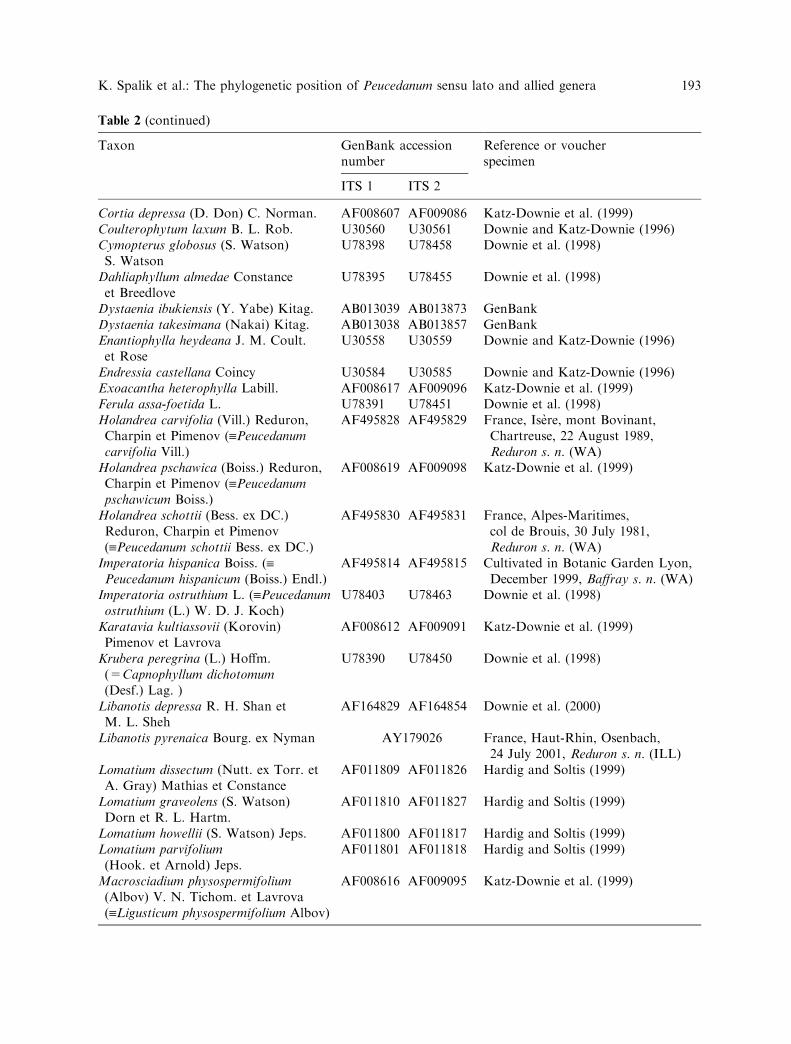
Table 2 (continued)
Taxon GenBank accessionnumber
Reference or voucherspecimen
ITS 1 ITS 2
Cortia depressa (D. Don) C. Norman. AF008607 AF009086 Katz-Downie et al. (1999)Coulterophytum laxum B. L. Rob. U30560 U30561 Downie and Katz-Downie (1996)Cymopterus globosus (S. Watson)S. Watson
U78398 U78458 Downie et al. (1998)
Dahliaphyllum almedae Constanceet Breedlove
U78395 U78455 Downie et al. (1998)
Dystaenia ibukiensis (Y. Yabe) Kitag. AB013039 AB013873 GenBankDystaenia takesimana (Nakai) Kitag. AB013038 AB013857 GenBankEnantiophylla heydeana J. M. Coult.et Rose
U30558 U30559 Downie and Katz-Downie (1996)
Endressia castellana Coincy U30584 U30585 Downie and Katz-Downie (1996)Exoacantha heterophylla Labill. AF008617 AF009096 Katz-Downie et al. (1999)Ferula assa-foetida L. U78391 U78451 Downie et al. (1998)Holandrea carvifolia (Vill.) Reduron,Charpin et Pimenov (”Peucedanumcarvifolia Vill.)
AF495828 AF495829 France, Isere, mont Bovinant,Chartreuse, 22 August 1989,Reduron s. n. (WA)
Holandrea pschawica (Boiss.) Reduron,Charpin et Pimenov (”Peucedanumpschawicum Boiss.)
AF008619 AF009098 Katz-Downie et al. (1999)
Holandrea schottii (Bess. ex DC.)Reduron, Charpin et Pimenov(”Peucedanum schottii Bess. ex DC.)
AF495830 AF495831 France, Alpes-Maritimes,col de Brouis, 30 July 1981,Reduron s. n. (WA)
Imperatoria hispanica Boiss. (”Peucedanum hispanicum (Boiss.) Endl.)
AF495814 AF495815 Cultivated in Botanic Garden Lyon,December 1999, Baffray s. n. (WA)
Imperatoria ostruthium L. (”Peucedanumostruthium (L.) W. D. J. Koch)
U78403 U78463 Downie et al. (1998)
Karatavia kultiassovii (Korovin)Pimenov et Lavrova
AF008612 AF009091 Katz-Downie et al. (1999)
Krubera peregrina (L.) Hoffm.(=Capnophyllum dichotomum(Desf.) Lag. )
U78390 U78450 Downie et al. (1998)
Libanotis depressa R. H. Shan etM. L. Sheh
AF164829 AF164854 Downie et al. (2000)
Libanotis pyrenaica Bourg. ex Nyman AY179026 France, Haut-Rhin, Osenbach,24 July 2001, Reduron s. n. (ILL)
Lomatium dissectum (Nutt. ex Torr. etA. Gray) Mathias et Constance
AF011809 AF011826 Hardig and Soltis (1999)
Lomatium graveolens (S. Watson)Dorn et R. L. Hartm.
AF011810 AF011827 Hardig and Soltis (1999)
Lomatium howellii (S. Watson) Jeps. AF011800 AF011817 Hardig and Soltis (1999)Lomatium parvifolium(Hook. et Arnold) Jeps.
AF011801 AF011818 Hardig and Soltis (1999)
Macrosciadium physospermifolium(Albov) V. N. Tichom. et Lavrova(”Ligusticum physospermifolium Albov)
AF008616 AF009095 Katz-Downie et al. (1999)
K. Spalik et al.: The phylogenetic position of Peucedanum sensu lato and allied genera 193
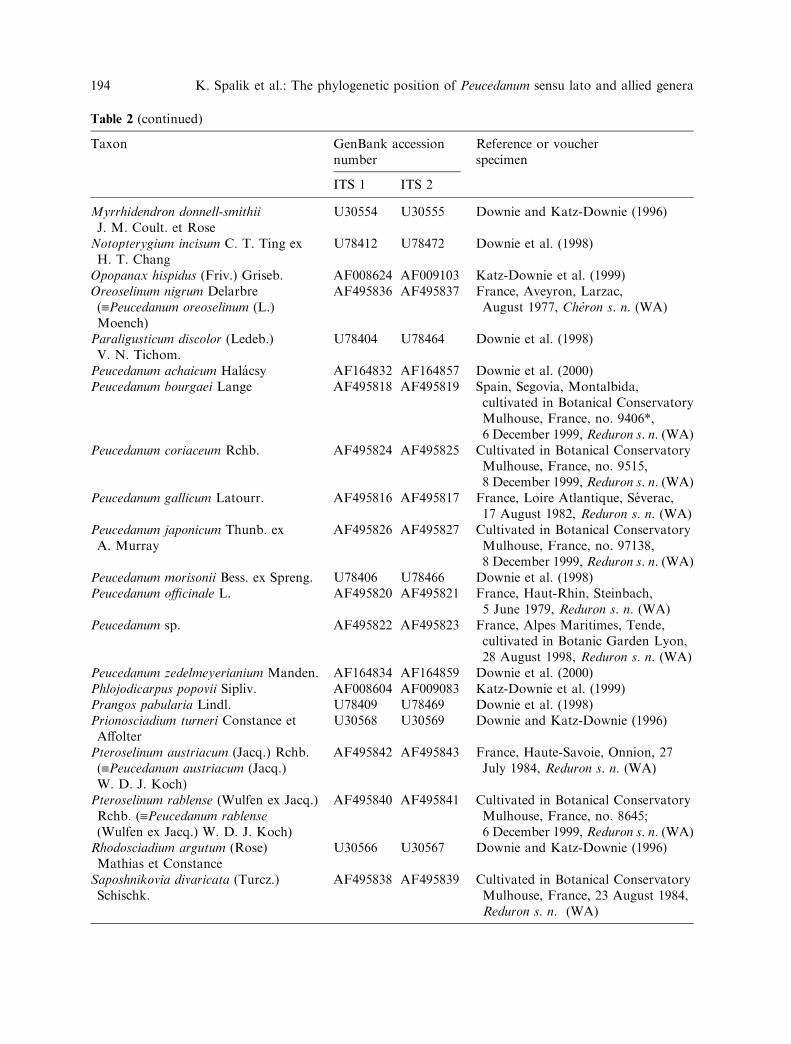
Table 2 (continued)
Taxon GenBank accessionnumber
Reference or voucherspecimen
ITS 1 ITS 2
Myrrhidendron donnell-smithiiJ. M. Coult. et Rose
U30554 U30555 Downie and Katz-Downie (1996)
Notopterygium incisum C. T. Ting exH. T. Chang
U78412 U78472 Downie et al. (1998)
Opopanax hispidus (Friv.) Griseb. AF008624 AF009103 Katz-Downie et al. (1999)Oreoselinum nigrum Delarbre(”Peucedanum oreoselinum (L.)Moench)
AF495836 AF495837 France, Aveyron, Larzac,August 1977, Cheron s. n. (WA)
Paraligusticum discolor (Ledeb.)V. N. Tichom.
U78404 U78464 Downie et al. (1998)
Peucedanum achaicum Halacsy AF164832 AF164857 Downie et al. (2000)Peucedanum bourgaei Lange AF495818 AF495819 Spain, Segovia, Montalbida,
cultivated in Botanical ConservatoryMulhouse, France, no. 9406*,6 December 1999, Reduron s. n. (WA)
Peucedanum coriaceum Rchb. AF495824 AF495825 Cultivated in Botanical ConservatoryMulhouse, France, no. 9515,8 December 1999, Reduron s. n. (WA)
Peucedanum gallicum Latourr. AF495816 AF495817 France, Loire Atlantique, Severac,17 August 1982, Reduron s. n. (WA)
Peucedanum japonicum Thunb. exA. Murray
AF495826 AF495827 Cultivated in Botanical ConservatoryMulhouse, France, no. 97138,8 December 1999, Reduron s. n. (WA)
Peucedanum morisonii Bess. ex Spreng. U78406 U78466 Downie et al. (1998)Peucedanum officinale L. AF495820 AF495821 France, Haut-Rhin, Steinbach,
5 June 1979, Reduron s. n. (WA)Peucedanum sp. AF495822 AF495823 France, Alpes Maritimes, Tende,
cultivated in Botanic Garden Lyon,28 August 1998, Reduron s. n. (WA)
Peucedanum zedelmeyerianium Manden. AF164834 AF164859 Downie et al. (2000)Phlojodicarpus popovii Sipliv. AF008604 AF009083 Katz-Downie et al. (1999)Prangos pabularia Lindl. U78409 U78469 Downie et al. (1998)Prionosciadium turneri Constance etAffolter
U30568 U30569 Downie and Katz-Downie (1996)
Pteroselinum austriacum (Jacq.) Rchb.(”Peucedanum austriacum (Jacq.)W. D. J. Koch)
AF495842 AF495843 France, Haute-Savoie, Onnion, 27July 1984, Reduron s. n. (WA)
Pteroselinum rablense (Wulfen ex Jacq.)Rchb. (”Peucedanum rablense(Wulfen ex Jacq.) W. D. J. Koch)
AF495840 AF495841 Cultivated in Botanical ConservatoryMulhouse, France, no. 8645;6 December 1999, Reduron s. n. (WA)
Rhodosciadium argutum (Rose)Mathias et Constance
U30566 U30567 Downie and Katz-Downie (1996)
Saposhnikovia divaricata (Turcz.)Schischk.
AF495838 AF495839 Cultivated in Botanical ConservatoryMulhouse, France, 23 August 1984,Reduron s. n. (WA)
194 K. Spalik et al.: The phylogenetic position of Peucedanum sensu lato and allied genera
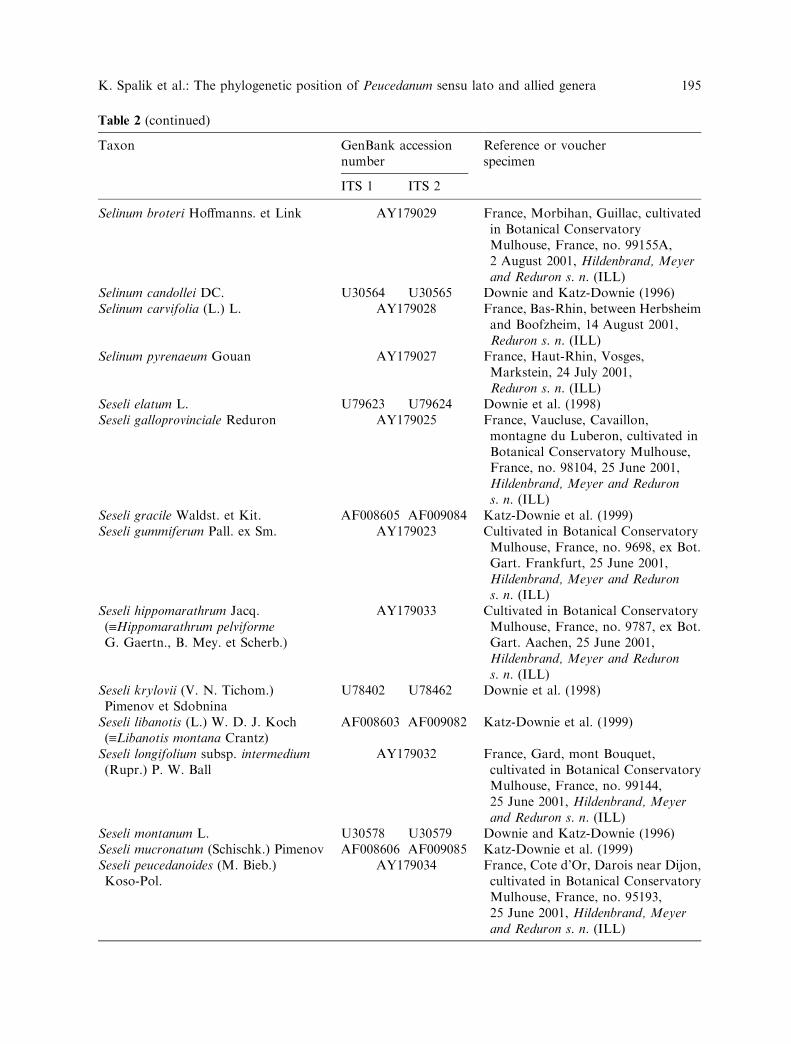
Table 2 (continued)
Taxon GenBank accessionnumber
Reference or voucherspecimen
ITS 1 ITS 2
Selinum broteri Hoffmanns. et Link AY179029 France, Morbihan, Guillac, cultivatedin Botanical ConservatoryMulhouse, France, no. 99155A,2 August 2001, Hildenbrand, Meyerand Reduron s. n. (ILL)
Selinum candollei DC. U30564 U30565 Downie and Katz-Downie (1996)Selinum carvifolia (L.) L. AY179028 France, Bas-Rhin, between Herbsheim
and Boofzheim, 14 August 2001,Reduron s. n. (ILL)
Selinum pyrenaeum Gouan AY179027 France, Haut-Rhin, Vosges,Markstein, 24 July 2001,Reduron s. n. (ILL)
Seseli elatum L. U79623 U79624 Downie et al. (1998)Seseli galloprovinciale Reduron AY179025 France, Vaucluse, Cavaillon,
montagne du Luberon, cultivated inBotanical Conservatory Mulhouse,France, no. 98104, 25 June 2001,Hildenbrand, Meyer and Redurons. n. (ILL)
Seseli gracile Waldst. et Kit. AF008605 AF009084 Katz-Downie et al. (1999)Seseli gummiferum Pall. ex Sm. AY179023 Cultivated in Botanical Conservatory
Mulhouse, France, no. 9698, ex Bot.Gart. Frankfurt, 25 June 2001,Hildenbrand, Meyer and Redurons. n. (ILL)
Seseli hippomarathrum Jacq.(”Hippomarathrum pelviformeG. Gaertn., B. Mey. et Scherb.)
AY179033 Cultivated in Botanical ConservatoryMulhouse, France, no. 9787, ex Bot.Gart. Aachen, 25 June 2001,Hildenbrand, Meyer and Redurons. n. (ILL)
Seseli krylovii (V. N. Tichom.)Pimenov et Sdobnina
U78402 U78462 Downie et al. (1998)
Seseli libanotis (L.) W. D. J. Koch(”Libanotis montana Crantz)
AF008603 AF009082 Katz-Downie et al. (1999)
Seseli longifolium subsp. intermedium(Rupr.) P. W. Ball
AY179032 France, Gard, mont Bouquet,cultivated in Botanical ConservatoryMulhouse, France, no. 99144,25 June 2001, Hildenbrand, Meyerand Reduron s. n. (ILL)
Seseli montanum L. U30578 U30579 Downie and Katz-Downie (1996)Seseli mucronatum (Schischk.) Pimenov AF008606 AF009085 Katz-Downie et al. (1999)Seseli peucedanoides (M. Bieb.)Koso-Pol.
AY179034 France, Cote d’Or, Darois near Dijon,cultivated in Botanical ConservatoryMulhouse, France, no. 95193,25 June 2001, Hildenbrand, Meyerand Reduron s. n. (ILL)
K. Spalik et al.: The phylogenetic position of Peucedanum sensu lato and allied genera 195
Table 2 (continued)
Taxon GenBank accessionnumber
Reference or voucherspecimen
ITS 1 ITS 2
Seseli praecox (Gamisans) Gamisans AY179024 France, Corse, Bussaglia, cultivated inBotanical Conservatory Mulhouse,France, no. 9528, 25 June 2001,Hildenbrand, Meyer and Redurons. n. (ILL)
Seseli tortuosum L. AY179031 Portugal, Lisboa, Sintra Praja dasMacas, cultivated in BotanicalConservatory Mulhouse, France, no.98042, 2 August 2001, Hildenbrand,Meyer and Reduron s. n. (ILL)
Seseli webbii Coss. AY179037 Canaries, Tenerife, cultivated inBotanical Conservatory Mulhouse,France, no. 20117D, 25 June 2001,Hildenbrand, Meyer and Redurons. n. (ILL)
Shoshonea pulvinata Evert et Constance U78400 U78460 Downie et al. (1998)Smyrniopsis aucheri Boiss. U78393 U78453 Downie et al. (1998)Spermolepis inermis (Nutt. ex DC.)Mathias et Constance
AF008602 AF009081 Katz-Downie et al. (1999)
Sphaenolobium tianschanicum(Korovin) Pimenov
AF008622 AF009101 Katz-Downie et al. (1999)
Sphenosciadium capitellatum A. Gray AF008600 AF009079 Katz-Downie et al. (1999)Taenidia integerrima (DC.) Drude U78399 U78459 Downie et al. (1998)Thaspium pinnatifidum (Buckley)A. Gray
U78410 U78470 Downie et al. (1998)
Thysselinum lancifolium (Lange)Calest.
AY179036 France, Loire Atlantique, St Lyphard,10 August 1982, Reduron s. n. (WA)
Thysselinum palustre (L.) Hoffm.(”Peucedanum palustre (L.) Moench)
AF008621 AF009100 Katz-Downie et al. (1999)
Thysselinum palustre (L.) Hoffm.(”Peucedanum palustre (L.) Moench)
AY179035 Poland, Warszawa, Bemowo,23 August 1985, Kirpluk s. n. (WA)
Thysselinum sp. AF495832 AF495833 France, Morbihan, Thehillac,Reduron and Riviere s. n. (WA)
Tommasinia verticillaris (L.) Bertol.(”Peucedanum verticillare (L.)W. D. J. Koch ex DC.)
AF008609 AF009088 Katz-Downie et al. (1999)
Xanthogalum purpurascens Ave-Lall.(”Angelica purpurascens(Ave-Lall.) Gilli)
AF008611 AF009090 Katz-Downie et al. (1999)
Xanthogalum tatianae (Bordz.)Schischk. (”Angelica tatianae Bordz.)
AF008610 AF009089 Katz-Downie et al. (1999)
Xanthoselinum alsaticum (L.)Schur (”Peucedanum alsaticum L.)
AF495834 AF495835 France, Haut-Rhin, Westhalten,21 August 1979, Reduron s. n. (WA)
Zizia aurea (L.) W. D. J. Koch U30574 U30575 Downie and Katz-Downie (1996)
196 K. Spalik et al.: The phylogenetic position of Peucedanum sensu lato and allied genera

Results
Sequence analysis. The intervening 5.8S rDNAregion was not available for most previouslypublished ITS sequences, therefore only the ITS1 and ITS 2 loci were considered. Complete ITS1 and ITS 2 sequences for 107 accessions rangedfrom 424 to 446 base pairs (bp). The alignedmatrix included 462 positions, with ITS 1 andITS2 each accounting for 226 and236positions,respectively. All positions were unambiguous:210 positions were parsimony informative, 83variable positions were parsimony uninforma-tive, and 169 positions were constant. Thirty-nine unambiguous gaps were introduced toalign these positions. Thirty-three of these were1-bp in size; single indels ranging from2 to 14 bpeach were also identified. Twenty indels wereparsimony informative.Meanuncorrectedpair-wise distance was 0.075; overall, distance valuesranged from identity to 0.219.
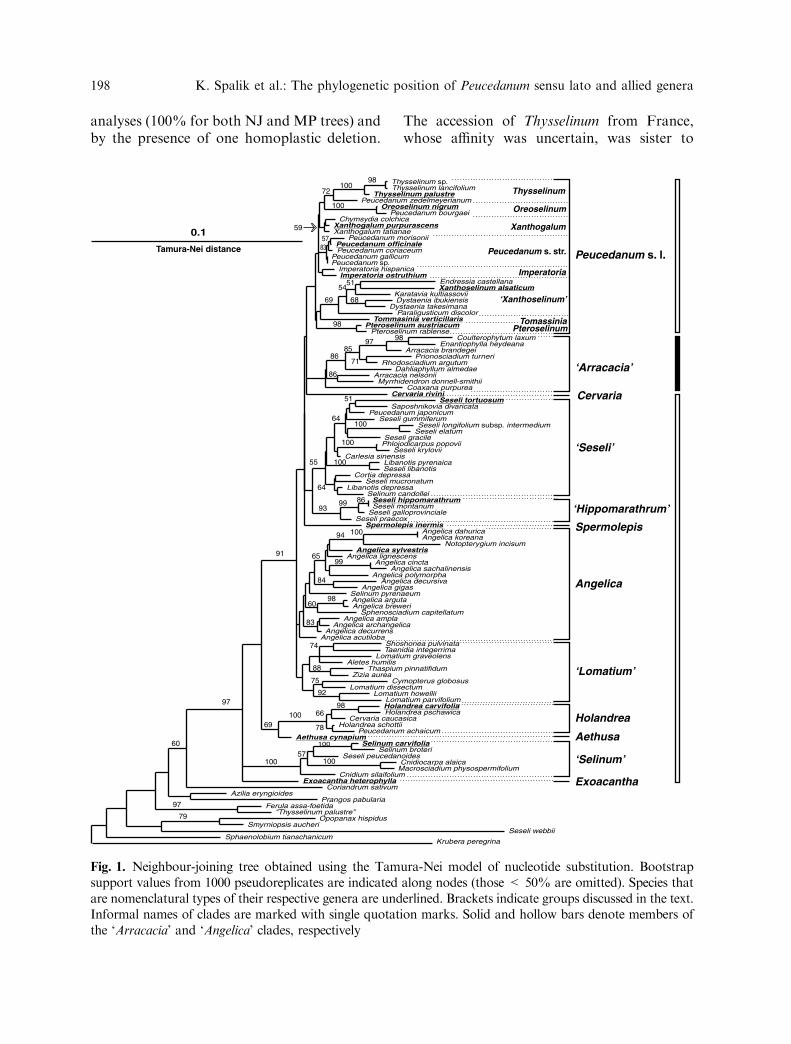
Phylogenetic analysis. NJ trees obtainedusing Jukes-Cantor, Kimura 2-parameter,Tamura-Nei, and GTR models of nucleotidesubstitutions were generally similar. The treeusing the Tamura-Nei model is shown inFig. 1. In all NJ trees, the formerly identified‘Arracacia’ clade is monophyletic (althoughwith bootstrap support < 50%) and nestedwithin the ‘Angelica’ clade sensu Downie et al.(2001). Several subclades were present in all NJtrees; these subclades are identified in Fig. 1and are discussed below.
Maximum parsimony searches resulted inthe preset limit of 10 000 shortest trees, each989-steps long, with a retention index of 0.666and a consistency index of 0.468 (includinguninformative characters). The strict consensusof these trees (not shown) yielded a largepolytomy, and the relationship between theformerly identified ‘Arracacia’ and ‘Angelica’clades could not be resolved. MP searches ofcombined ITS sequences and scored indelsresulted in 10 000 shortest trees of 1064 stepseach, a retention index of 0.672 and a consis-tency index of 0.483 (including uninformativecharacters). The strict consensus of these trees(Fig. 2) showed less polytomy and stronger
internal support than in the MP analysis ofsequence positions alone. Therefore, from hereon, we only refer to the results of MP analysesof combined data. Of the 20 phylogeneticallyinformative indels, 10 were apomorphic and 10were homoplastic.
Modeltest’s likelihood ratio test statisticselected the Tamura-Nei (1993) model ofnucleotide substitution, with gamma distribu-tion of substitution rates and a shape param-eter of 0.727, for use in the ML analysis. Thetree obtained from ML searches (ln L ¼)6086.77, Fig. 3) had a similar topology tothose obtained from the NJ analyses. All majorclades and subclades (identified on the Tamura-Nei tree, Fig. 1) had the same composition,with the differences between them includingminor rearrangements within these clades orsomewhat different affinities among them.
In all analyses, the monophyly of the‘Angelica’ clade, with the ‘Arracacia’ cladeincluded within, was strongly supported (97%and 98% for NJ and MP analyses, respec-tively). In both the NJ and ML trees, as well assome MP trees, nearly all members of Peuce-danum and its segregates – i.e., Peucedanumsensu stricto, Imperatoria, Thysselinum,Oreoselinum, Tommasinia, Pteroselinum, andXanthoselinum – occurred in a single clade,denoted as Peucedanum sensu lato. This cladealso included several taxa that were nothitherto placed in Peucedanum, for exampleEndressia and Xanthogalum. However, in theNJ tree, this clade received weak bootstrapsupport (<50%). Within this clade, severalsubclades were apparent. These mostly corre-sponded to the narrowly defined Peucedanumsegregates, with the notable exception of amore heterogenic ‘Xanthoselinum’ subclade.Many of these same subclades were alsoapparent in trees resulting from MP analysisof combined data (Fig. 2).
The Thysselinum subclade included threenewly sequenced accessions of Thysselinumand the accession of Peucedanum zedelmeye-rianum. The monophyly of Thysselinummembers was strongly supported by bootstrap
K. Spalik et al.: The phylogenetic position of Peucedanum sensu lato and allied genera 197
analyses (100% for both NJ and MP trees) andby the presence of one homoplastic deletion.
The accession of Thysselinum from France,whose affinity was uncertain, was sister to
Fig. 1. Neighbour-joining tree obtained using the Tamura-Nei model of nucleotide substitution. Bootstrapsupport values from 1000 pseudoreplicates are indicated along nodes (those < 50% are omitted). Species thatare nomenclatural types of their respective genera are underlined. Brackets indicate groups discussed in the text.Informal names of clades are marked with single quotation marks. Solid and hollow bars denote members ofthe ‘Arracacia’ and ‘Angelica’ clades, respectively
198 K. Spalik et al.: The phylogenetic position of Peucedanum sensu lato and allied genera

Fig. 2. Strict consensus tree of 10 000 shortest 1064-step trees obtained from maximum parsimony analyses ofcombined ITS sequences and scored indels (consistency index with uninformative characters ¼ 0.483, retentionindex ¼ 0.672). Decay indices/bootstrap values from 1000 pseudoreplicates are indicated along nodes (onlythose consistent with the 50% majority-rule consensus tree are indicated). Group identifiers are the same as inFig. 1
K. Spalik et al.: The phylogenetic position of Peucedanum sensu lato and allied genera 199
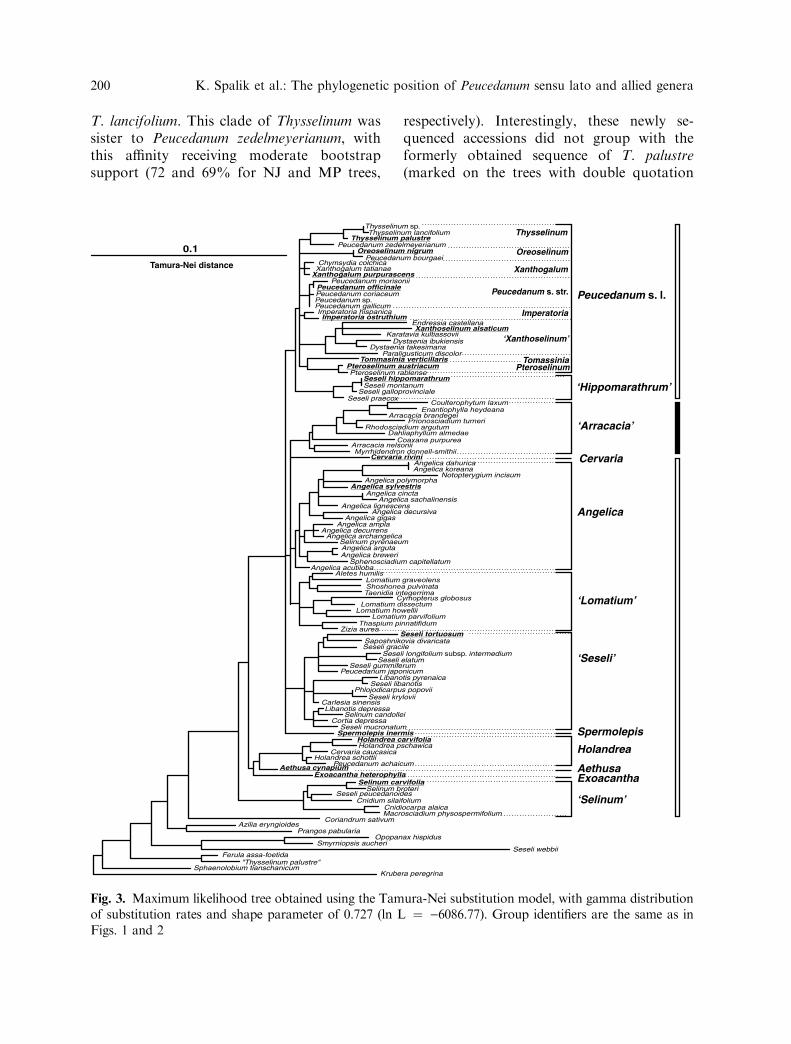
T. lancifolium. This clade of Thysselinum wassister to Peucedanum zedelmeyerianum, withthis affinity receiving moderate bootstrapsupport (72 and 69% for NJ and MP trees,
respectively). Interestingly, these newly se-quenced accessions did not group with theformerly obtained sequence of T. palustre(marked on the trees with double quotation
Fig. 3. Maximum likelihood tree obtained using the Tamura-Nei substitution model, with gamma distributionof substitution rates and shape parameter of 0.727 (ln L ¼ )6086.77). Group identifiers are the same as inFigs. 1 and 2
200 K. Spalik et al.: The phylogenetic position of Peucedanum sensu lato and allied genera

marks) that was sister to Ferula assa-foetida,one of several outgroup representatives. Theaffinity of these two taxa was supported bythe presence of three indels, two of themsynapomorphic. One of these synapomorphieswas also shared by Sphaenolobium tianschan-icum.
The monotypic genus Oreoselinum groupedwith Peucedanum bourgaei (100% bootstrapsupport for both MP and NJ trees). TheXanthogalum subclade encompassed two spe-cies of this genus, as well as the monotypicChymsydia; all three of these species aredistributed in the Caucasus. This subcladeoccurred in all analyses, although its mono-phyly was weakly supported (bootstrap values59 and 62% for NJ and MP trees, respec-tively).
The Peucedanum sensu stricto subcladeencompassed five accessions that are tradition-ally placed in sect. Peucedanum (bootstrapsupport 83% and 70% for NJ and MP trees,respectively). Included here is P. officinale, thenomenclatural type of the genus. This subcladealso included the Peucedanum accession (Peu-cedanum sp.) of uncertain affinity from France.Its sequence was identical to that ofP. gallicum.
Both species of Imperatoria grouped to-gether in the NJ and ML trees (Figs. 1 and 3,respectively), although this affinity was poorlysupported in the NJ analysis (<50%). Thesetwo species did not always form a monophy-letic group in the MP analysis (Fig. 2).
The ‘Xanthoselinum’ subclade encompassedthe type species of this genus, as well asrepresentatives of Endressia, Karatavia, Dys-taenia, and Paraligusticum. This clade receivedmoderate bootstrap support (69 and 70% forNJ and MP trees, respectively), albeit therelationships within were poorly supported. Inthe NJ trees, this subclade was allied weaklywith two members of Pteroselinum and themonotypic Tommasinia. The monophylyof Pteroselinum was supported by highbootstrap values (98 and 99% for NJ andMP, respectively) and by one synapomorphicinsertion.
The genus Cervaria was polyphyletic. Thephylogenetic position of Cervaria rivini, thenomenclatural type of the genus, was equivo-cal, although it commonly grouped with themembers of Peucedanum sensu lato and the‘Arracacia’ clade. The other species, C. cauca-sica, was placed among the members ofHolandrea. Holandrea was distantly related tothe other peucedanoid genera. The Holandreaclade included the three species hitherto recog-nised in the genus, plus C. caucasica andPeucedanum achaicum. This clade was wellsupported, with 100% bootstrap values in bothNJ and MP trees, and was sister to Aethusacynapium. Monophyly of the Holandrea cladewas also supported by two synapomorphicinsertions. These were 1 and 5-bp long andoccurred at the beginning of the ITS 1 regionand close to the 3’ end of ITS 2, respectively.
Most members of Seseli and its segregateLibanotis Haller ex Zinn, including their typespecies (S. tortuosum for Seseli and S. libanotis¼ Libanotis montana for Libanotis), wereplaced in a single clade (‘Seseli’), which how-ever received only moderate bootstrap support(55 and 67% for NJ and MP trees, respec-tively). This branch also contained severalAsiatic umbellifers that are placed in thegenera Saposhnikovia, Phlojodicarpus, Carle-sia, and Cortia. In the NJ trees, the nomen-clatural type of Saposhnikovia, S. divaricata,was sister to S. tortuosum, the nomenclaturaltype of Seseli, and both were found to beclosely related to Peucedanum japonicum. Inthe MP trees, S. divaricata was sister toP. japonicum, and this affinity was supportedby the presence of one homoplastic insertion.In the ML tree (Fig. 3), S. tortuosum andS. divaricata were sister taxa, whereasP. japonicum formed a branch of a polytomytwo nodes away.
Four European members of Seseli(S. hippomarathrum, S. galloprovinciale,S. montanum, and S. praecox) formed astrongly supported clade (93–95% bootstrapvalues) that was sister to the ‘Seseli’ clade inthe NJ analysis (Fig. 1). In the ML tree(Fig. 3), this clade was sister to the Peuceda-
K. Spalik et al.: The phylogenetic position of Peucedanum sensu lato and allied genera 201

num sensu lato clade. In the MP trees, itsplacement was equivocal, but closer to thePeucedanum sensu lato group than to the‘Seseli’ clade. Since this group included Seselihippomarathrum, the type species of Hippo-marathrum G. Gaertn., B. Mey. et Scherb., wetherefore refer to it as the ‘Hippomarathrum’clade.
Two members of Seseli were placedfar away from its other congeners. Seselipeucedanoides was included in the ‘Selinum’clade, whereas S. webbii was clustered with theoutgroup species. Searches in GenBank usingBLAST identified ITS sequences of Ammi
majus and Petroselinum crispum as being mostsimilar to S. webbii, and this affinity wasconfirmed through additional phylogeneticanalyses using a larger data set (not shown).Both Spermolepis inermis and Exoacanthaheterophylla appeared to have no closerelatives among the taxa included in theanalysis.
In all trees, the species of Angelica formed amonophyletic group, albeit one that was onlypoorly supported (<50% bootstrap values).This clade also included the Asiatic Notoptery-gium and North American Sphenosciadium, aswell as a species of Selinum from Spain and
Table 3. Generic composition of tribe Selineae Spreng. in Roem. et Schult.
Aethusa L. Meum Mill.Aletes J. M. Coult. et Rose Musineon Raf.Angelica L. (inc. Sphenosciadium A. Gray) Myrrhidendron J. M. Coult. et RoseArracacia Bancroft Neoparrya MathiasCarlesia Dunn. Oreonana Jeps.Cervaria Wolf Oreoselinum Mill.Chamaele Miq. Oreoxis Raf.Chymsydia Albov Orogenia S. Wats.Cnidiocarpa Pimenov Paraligusticum V. N. Tikhom.Cnidium Cusson in Juss. pro parte Peucedanum L.(=Katapsuxis Raf.) Phlojodicarpus Turcz. ex Ledeb.
Coaxana J. M. Coult. et Rose Podistera S. Wats.Coelopleurum Ledeb. Polytaenia DC.Cortia DC. Prionosciadium S. Wats.Coulterophytum B. L. Rob. Pseudocymopterus J. M. Coult. et RoseCymopterus Raf. Pteroselinum Rchb.Dahliaphyllum Constance et Breedlove Pteryxia (Nutt. ex Torr. et A. Gray)Donnellsmithia J. M. Coult. et Rose J. M. Coult. et RoseDystaenia Kitag. Rhodosciadium S. Wats.Enantiophylla J. M. Coult. et Rose Saposhnikovia Schischk.Endressia J. Gay Selinum L.Exoacantha Labill. Seseli L.Glehnia F. Schmidt ex Miq. Shoshonea Evert et ConstanceGrafia Rchb. Spermolepis Raf.Harbouria J. M. Coult. et Rose Taenidia (Torr. et A. Gray) DrudeHolandrea Reduron, Charpin et Pimenov Tauschia Schltdl.Imperatoria L. Thaspium Nutt.Karatavia Pimenov et Lavrova Thysselinum Adans.Libanotis Haller ex Zinn Tommasinia Bertol.Lomatium Raf. Xanthogalum Lallem.Macrosciadium V. N. Tichom. et Lavrova Xanthoselinum SchurMathiasella Constance et C. Hitch. Zizia W. D. J. Koch
202 K. Spalik et al.: The phylogenetic position of Peucedanum sensu lato and allied genera

France. Sphenosciadium capitellatum groupedwith the two North American representativesof Angelica included in this study, whereasNotopterygium incisum from China groupedwith two Angelica species from the Far East(A. koreana and A. dahurica).
The ‘Lomatium’ clade included severalspecies endemic to central and western NorthAmerica. This group was monophyletic in theNJ and ML trees but formed two clades in theMP strict consensus tree.
Discussion
Tribe Selineae. Most of the genera consideredin this study are traditionally classified inDrude’s (1897–1898) tribe Peucedaneae sub-tribes Angelicinae and Ferulinae, or tribesPeucedaneae and Angeliceae in the classifica-tion system of Pimenov and Leonov (1993). Inan outline of a new classification of umbellifersbased on molecular data, Downie et al. (2001)placed all of these taxa into two informalgroups, named the ‘Angelica’ clade and the‘Arracacia’ clade. However, the phylogeneticposition of the ‘Arracacia’ clade was not clear,for in several previous studies it arose fromwithin the ‘Angelica’ clade (Plunkett et al. 1996,Downie et al. 1998). Downie et al. (2001)suggested that the ‘Arracacia’ clade mighteventually be subsumed within the ‘Angelica’clade. Our study confirms those earlier resultsindicating that the weakly monophyletic ‘Arra-cacia’ clade is indeed nested within the ‘Angel-ica’ clade. The monophyly of this expanded‘Angelica’ clade is, however, very stronglysupported (with 97-98% bootstrap values).Herein, we recognise this expanded clade as atribe. It includes several nomenclatural types ofvalidly published tribal names of which theearliest is Selineae Spreng. in Roem. et Schult.(Syst. Veg. 6: 46 1820). Table 3 contains thegeneric composition of tribe Selineae. Thistable includes 18 additional genera not exam-ined herein; their placement within the tribe isbased on Downie et al. (2001).
Peucedanum sensu lato vs. Peucedanumsensu stricto. The delimitation of Peucedanum
as found in most taxonomic monographs isambiguous. Traditionally, members of thisgenus are characterised by dorsally com-pressed orthospermous fruits without promi-nent dorsal ribs and with a broad commissure.This last character distinguishes Peucedanumfrom Angelica, which has a narrow commis-sure (Ostroumova and Pimenov 1997). Manyspecies of Peucedanum have winged fruits.Such a fruit structure is an evident adaptationfor wind dispersal and likely evolved indepen-dently in several distant lineages of umbellifers(Theobald 1971). Therefore, not surprisingly,Peucedanum encompassed taxa that were sub-sequently found to be distantly related to thetype of the genus. For example, Steganotaenia,which was synonymised with Peucedanum byDrude (1897-1898), is placed close to thesubfamily Saniculoideae on the basis of molec-ular evidence (Downie and Katz-Downie1999). Apart from these foreign elements,Peucedanum sensu amplissimo (i.e. sensuDrude 1897–1898) includes many diverse taxathat are indeed closely related, such as theNorth American Lomatium and Orogenia, andthe Peucedanum segregates from the OldWorld (i.e. Cervaria, Holandrea, Imperatoria,Oreoselinum, Pteroselinum, Thysselinum, andXanthoselinum). Based on the analysis pre-sented herein, these taxa are scattered amongmembers of tribe Selineae. This finding sug-gests that the similarity of the former membersof Peucedanum sensu amplissimo is homoplas-tic and supports the splitting of the genus intosmaller units that may be defined on the basisof apomorphic characters. In spite of theexclusion of the North American taxa thatare now placed in Lomatium and relatedgenera, Peucedanum sensu lato still includes100-120 species (Pimenov and Leonov 1993).
Based on the results of the NJ and MLanalyses, and with the exception of Holandreaand Cervaria, the segregates of Peucedanumsensu lato form a monophyletic group. Asubset of the 10 000 MP trees also revealed thesame clade. From the point of view ofnomenclatural stability, retaining such a delim-itation of the genus (but with the exclusion of
K. Spalik et al.: The phylogenetic position of Peucedanum sensu lato and allied genera 203

Holandrea and Cervaria) would be advanta-geous. However, the monophyly of this groupis weakly supported. Moreover, it also includesseveral taxa that do not conform to thetraditional delimitation of Peucedanum. Forexample, Xanthoselinum alsaticum is sistertaxon to Endressia castellana, which is arepresentative of an Iberian endemic genusencompassing only two species. Contrary toPeucedanum, the fruits of Endressia are later-ally compressed and do not have any wings,and both dorsal and commissural ribs are ofthe same size (Arenas Posada and GarcıaMartın 1993). The ‘Xanthoselinum’ group, withincluded genera Dystaenia, Karatavia, andParaligusticum, is the most heterogeneousamong the subclades identified within Peuce-danum sensu lato, and it is very difficult to findany obvious morphological character thatwould unite these taxa. Another member ofthe Peucedanum sensu lato clade, Xantho-galum, was recognised in Angelica by Vasil’evaand Pimenov (1991). While its members aregenerally similar to those of Angelica, theydiffer in having conspicuous calyx teeth, whichare persistent and erect in fruit, and theabsence of bracts and bracteoles (Chamberlain1972). The similarity between Xanthogalumand Angelica is homoplastic, as these generaare not immediately related (Downie et al.2000b). In summary, Peucedanum sensu lato istoo difficult to delimit based on morphology.
The advantage of Linnaean binomials isthat the generic name permits the identifica-tion of the closest relatives of a specieswithout referring to any external classifica-tion system. This is particularly useful fornon-taxonomists who deal with species iden-tification, such as in floristic or ecologicalstudies. Therefore, any study that clarifiesgeneric boundaries in the Apiaceae would bevery useful. Ideal genera should be mono-phyletic, clearly delimited based on morphol-ogy, and manageable with respect to size(Spalik et al. 2001). Both monotypic and verylarge genera should be avoided, if possible.However, it seems that if we reject a broaddefinition of Peucedanum, we end up with
numerous small genera, each containing only1–3 species. Sometimes, these species areproblematic and often reduced to infraspecificranks. For instance, Pteroselinum rablense isoften recognised as a subspecies of Pterosel-inum austriacum, whereas Peucedanum bour-gaei is frequently regarded as a subspecies ofPeucedanum oreoselinum (”Oreoselinum ni-grum). Imperatoria includes three parapatricspecies that are morphologically similar:European I. ostruthium, Iberian I. hispanica,and I. lowei from Madeira. Xanthoselinumand Tommasinia are each monotypic. ThePeucedanum sensu stricto clade includes taxathat are very similar with respect to their ITSsequences, which suggests that the radiationof this group was recent and thus, includedtaxa may deserve infraspecific rank ratherthan that of species. They are very similar inhabit, sharing not only fruit characters butalso vegetative features, like linear-filiformleaf lobes. The accession denoted as Peuce-danum sp. represents a population whosemembers are morphologically very similar toP. gallicum. However, this population occurswithin the range of P. coriaceum rather thanof P. gallicum. The sequences of Peucedanumsp. and P. gallicum are identical and some-what different from P. coriaceum, suggestingthat Peucedanum sp. and P. gallicum areconspecific.
The results of the NJ analysis suggests thatPteroselinum, Tommasinia, and the ‘Xanthosel-inum’ clade are closely related. The affinityamong Pteroselinum austriacum, Tommasiniaverticillaris, and Xanthoselinum alsaticum waspreviously revealed by phytochemical data(Hadacek 1989). The relationships amongmembers of Peucedanum sensu lato, Angelica,and several related genera were recently inves-tigated by Shneyer et al. (2003), using immu-nological methods. Their study confirmed thatspecies classified in segregate genera Xantho-selinum, Pteroselinum, Tommasinia, and Oreo-selinum are closely related to one another andto P. morisonii, a member of Peucedanumsensu stricto. Apparently, more data fromDNA sequences, as well as morphology and
204 K. Spalik et al.: The phylogenetic position of Peucedanum sensu lato and allied genera

phytochemistry, are needed to resolve thetaxonomy of Peucedanum sensu lato. Wetherefore refrain from taxonomic changes inthis group.
Cervaria and Holandrea. The genus Cer-varia was restored by Pimenov (1987) for theFlora Iranica. Pimenov listed several differ-ences between Peucedanum sensu stricto andCervaria, including fruit, flower, and leafcharacteristics. In contrast to Peucedanumsensu stricto, the fruits of Cervaria arecharacterised by the presence of lignifiedparenchymatic cells in the mesocarp. Calyxteeth are absent and ultimate leaf segmentsare broad in Cervaria, while in Peucedanumsensu stricto the calyx is present and the leafsegments are linear-filiform. However, itseems that such a delimitation of Cervaria isincorrect.
From among the four species included inCervaria, two – C. rivini and C. caucasica –were considered in our study. Molecular anal-yses suggested that these two species wereneither closely related to members of Peuce-danum sensu stricto nor to one another. Theplacement of the nomenclatural type of thegenus, C. rivini, was ambiguous and dependedon the phylogenetic method used, whereasC. caucasica was allied with those formermembers of Peucedanum that we recognise inHolandrea.
The genus Holandrea was described byReduron et al. (1997) as comprising threespecies, all of which have been included in ourstudy. Apart from their general morphologicalsimilarity (in habit, leaves, and fruits), theyhave similar stomata (Guyot 1984). However,they differ from members of Peucedanum withrespect to their chemical constituents. They alllack coumarins and all have C13-acetyleniccompounds (Hadacek 1989). Two members ofthe Holandrea clade, H. pschawica and C.caucasica, were also included in immunologicalstudies of seed proteins (Shneyer et al. 2003),which confirmed that these species were moreclosely related to one another than to the otherrepresentatives of Peucedanum sensu lato. Ourdata confirm this distant position of Holandrea
with respect to the Peucedanum segregates.Upon the inclusion of C. caucasica andP. achaicum, the genus Holandrea is mono-phyletic. Two new combinations are thereforenecessary.
Holandrea caucasica (M. Bieb.) Spalik,Reduron et S. R. Downie, comb. nov. ;basionym: Selinum caucasicum M. Bieb., Fl.Taur.-Cauc. 1: 213 1808. Type: ‘Ex Iberia: insylvis circa Ananur lectum. a. 1802’, Marschallvon Bieberstein (lectotype, LE, not seen; fideMenitsky 1991).
Holandrea achaica (Halacsy) Spalik, Redu-ron et S. R. Downie, comb. nov. ; basionym:Peucedanum achaicum Halacsy, Suppl. Consp.Fl. Graec. 42 1908. Type: ‘in rupibus vallisfluminis Voreikos infra Zachloru Achaiae,’July 1896, Saint Lager (WU-Hal-Graec, notseen; G, isotype!).
Thysselinum. The previously obtained ITSsequence of Thysselinum palustre allied withCoenolophium denudatum and some membersof Ferula, such as F. assa-foetida (Katz-Dow-nie et al. 1999, Downie et al. 2000b). Incontrast, the present study indicates that allnewly sequenced accessions of Thysselinumform a monophyletic group within the Peuce-danum sensu lato clade. It appears that thenewly sequenced Thysselinum palustre acces-sion is distantly related to the old accession ofthe same species; apparently, these two taxaare not conspecific. The results of our analysesconfirm the close relationship betweenT. palustre and T. lancifolium that was inferredpreviously from morphology and phytochem-istry (Reduron et al. 1997). These species arevery similar and differ only in flower colourand shape of the leaf lobes. Thysselinumlancifolium is characterised by having greenishflowers and long linear lobes, whereas theflowers of T. palustre are white and its leaflobes are shorter. Therefore, we have no doubtthat the correct placement of Thysselinum is inthe Peucedanum sensu lato group of tribeSelineae. The accession denoted as Thysseli-num sp. comes from a population that occursat the contact zone of T. palustre and T.lancifolium, and its members show intermedi-
K. Spalik et al.: The phylogenetic position of Peucedanum sensu lato and allied genera 205

ate characters between the two species. ITSsequences suggest that this taxon is moreclosely related to T. lancifolium than to T.palustre.
Angelica. Phylogenetic analyses using NJ,ML and MP methods confirm the monophylyof the genus Angelica. However, internalsupport for this clade is low. In contrast toour results, Shneyer et al. (2003) found signif-icant differences among the species of Angelicaincluded in their immunological study. Partic-ularly, those species that are sometimes placedin the segregate genus Ostericum differ consid-erably from the type of Angelica (A. sylvestris)and its relatives, as well as from the membersof Peucedanum sensu lato. Therefore, Shneyeret al. (2003) concluded that Ostericum deservedthe status of an independent genus. None ofthe putative members ofOstericum has hithertobeen included in molecular analyses.
The Angelica subclade outlined hereinincludes three taxa that are usually placedin other genera: Notopterygium incisum, Sphe-nosciadium capitellatum, and Selinum pyrena-eum. Notopterygium H. Boissieu includes fivespecies that occur in Asia, and mostly inChina (Pu et al. 2000). The only speciesincluded in our analysis, N. incisum, groupedwith two species of Angelica from the FarEast (A. dahurica and A. koreana). Apartfrom a similar geographic distribution, thisrelationship obtained strong support frombootstrap and decay analyses. This groupbelongs to a larger branch that also includesAngelica sylvestris, the nomenclatural type ofthe genus. The position of Notopterygium wasrecently investigated by Valiejo-Roman et al.(2002) using ITS data from the nomenclatur-al type of the genus, N. forbesii, and anotheraccession of N. incisum (as N. weberbauer-ianum). Their analyses rejected the closerelationship between Notopterygium and An-gelica and suggested that the former wassister to Hansenia. Our preliminary analysesof ITS a data from two other accessions ofthese species confirm the affinity betweenNotopterygium and Hansenia (unpublisheddata).
The placement of Selinum pyrenaeum hasbeen the subject of some controversy, as it wasalso placed in Seseli, Peucedanum, and Angel-ica. Reduron et al. (1997) regarded this speciesas related to Ligusticum sensu lato and rein-stated it in the monotypic genus Epikeros Raf.However, they noted that it shared somefeatures with members of Angelica, includingsimilar pathogens (Durrieu 1982) and thepresence of coumarins (Muckensturm andDiyani 1995). Based on the drawings in ArenasPosada and Garcıa Martın (1993), the fruitanatomy of S. pyrenaeum is more similar tothose winged fruits of Angelica than it is to itscongener, S. carvifolia. Nevertheless, its habitis different from most members of Angelica.Our data indicate that the placement of thisspecies is rather in Angelica than in Ligusticumsensu lato. Since it is nested within the former,it does not deserve the status of a separategenus. The name of this species in Angelica isA. pyrenaea (L.) Spreng.
Sphenosciadium capitellatum, the onlymember of Sphenosciadium A. Gray, is astriking umbellifer due to its woolly indumen-tum covering the umbellets. Otherwise, thisspecies is very similar in habit to its closestrelatives based on ITS sequences, which are A.arguta and A. breweri. These three taxa occurin North America, whereas the other membersof the Angelica clade are from the Old World.Therefore, retaining the monotypic Spheno-sciadium is not justified. The close relationshipbetween Sphenosciadium and Angelica wasalready indicated based on matK sequences(Plunkett et al. 1996), and the transfer of theformer into Angelica was suggested by Katz-Downie et al. (1999). Therefore, we formallysynonymise Sphenosciadium with Angelica:
Angelica capitellata (A. Gray) Spalik, Re-duron et S. R. Downie, comb. nov.; basionym:Sphenosciadium capitellatum A. Gray, Proc.Am. Acad. 6: 537 1865. Holotype: ‘In theSierra Nevada, near Ebbett’s Pass, AlpineCounty, California,’’ USA, Brewer 2026(GH, not seen).
Seseli, Libanotis, Cortia and Hippomara-thrum. With 100–120 included species, the
206 K. Spalik et al.: The phylogenetic position of Peucedanum sensu lato and allied genera

genus Seseli is one of the largest in the family(Pimenov and Leonov 1993), and similar toother large genera in Apiaceae, it is polyphy-letic (Katz-Downie et al. 1999, Downie et al.2000b). Several attempts to divide it intosmaller units have already been made. Forexample, Pimenov and Leonov (1993) listed 16generic names in synonymy. Of these segre-gates, only Libanotis has gained some accep-tance. The type of this genus is Libanotismontana, a nomenclatural synonym of Seselilibanotis. This species was placed in the ‘Seseli’clade, thus confirming that Libanotis andSeseli are closely related. Seseli libanotis issister to Libanotis pyrenaica, a taxon that isoften included in the former.
Another species described under Libanotis,L. depressa, is not related to the type of thegenus, but rather is sister to Selinum candollei,an Asiatic umbellifer, which is also recognisedin Cortia DC. (as C. candollei (DC.) Leute).The type of Cortia is C. lindleyi DC., anomenclatural synonym of C. depressa, whichwas grouped in the same subclade. The fourthmember of this subclade, Seseli mucronatum, isalternatively placed in Ligusticum (Flora ofChina, Umbelliferae, Draft 1: World Umbel-lifer Database, Royal Botanic Garden Edin-burgh, http://rbg-web2.rbge.org.uk/URC/Nomenclature/). Such a placement is notsupported by our data. Instead, the results ofthe phylogenetic analyses suggest that thesefour species may be either included in a broadgenus Seseli or in a more narrow genus Cortia.Such a decision requires, however, confirma-tion from independent data, both morpholog-ical and molecular. The relationship of thissubclade to the other subclades of the ‘Seseli’group is also unclear.
Most species recognised in Seseli belong tothe ‘Seseli’ clade. Based on additional unpub-lished ITS data, one may add three otherspecies to this clade: S. rigidum Waldst. et Kit.,S. farreynii Molero et A. Pujadas, andS. rhodopeum Velen. Surprisingly, the closestrelatives of Seseli tortuosum, the nomencla-tural type of the genus, are not the othercongeners but two species from the Far East:
Saposhnikovia divaricata and Peucedanum ja-ponicum. However, the phylogenetic affinitieswithin this clade received very low internalsupport. Therefore, additional sampling isneeded before any definite conclusions aremade and taxonomic changes introduced.
The taxonomic history of Hippomarathrumis complex. This namewas once used for a groupof species related to Cachrys sicula L., thenomenclatural type of Hippomarathrum Link,1821, Enum. Horti Berol. Alt. 1: 271. However,it is a later homonym of Hippomarathrum G.Gaertn., B. Mey. et Scherb. 1799, Oecon.Techn. Fl. Wetterau, 1: 249, 413 typified bySeseli hippomarathrum. The proposition toconserve Hippomarathrum Link against its ear-lier homonym was rejected by the Committeefor Spermatophyta (cf. Taxon 27:385–314,1978). The group of Seseli species denoted onthe tree as the ‘Hippomarathrum’ clade seems tobe distinct from its congeners. Although the NJanalysis suggests its sister relationship with the‘Seseli’ clade, such an affinity is not supportedby the results of theML andMP analyses. Withadditional sampling, it is entirely possible thatthese two clades of Seseli may form a singlelineage in all analyses. Therefore, further sam-pling of members of Seseli is required before therestoration of Hippomarathrum is considered.
Two species of Seseli, S. peucedanoides andS. webbii, have been placed far away from theircongeners. This separation is also supportedby morphological characters: those two specieshave yellow flowers, while in all other Seselispecies the flowers are white.
Selinum. Only few species are recognised inSelinum at present (Pimenov and Leonov1993), although nearly 300 binomials usingthis generic name exist (World UmbelliferDatabase, Royal Botanic Garden Edinburgh,http://rbg-web2.rbge.org.uk/URC/Nomencla-ture/). These include S. carvifolia, S. pyrenae-um, and S. candollei; the fourth member of thisgenus included in our study – S. broteri – isusually treated as a synonym of S. carvifolia orregarded as its subspecies since the differencesbetween them are minor (Arenas Posada andGarcıa Martın 1993). With S. pyrenaeum and
K. Spalik et al.: The phylogenetic position of Peucedanum sensu lato and allied genera 207

S. candollei excluded, the genus is practicallymonotypic. Interestingly, the ‘Selinum’ cladealso includes several taxa with incorrect genericaffiliation. The sister to Selinum is a member ofSeseli, and Cnidium silaifolium is sometimesrecognised in the monotypic genus KatapsuxisRaf. (Reduron et al. 1997). Since the nomen-clatural type of Cnidium, C. monnieri (L.)Cusson, has not been so far included inmolecular studies, we are reluctant to acceptthis placement. The fruits of Seseli peucedano-ides are generally similar to those of Cnidium,supporting the affinity of these species.
There is little doubt that Ligusticum physo-spermifolium is not related to L. scoticum L.,the nomenclatural type of the genus. There-fore, we recognise this taxon in MacrosciadiumV.N. Tichom. et Lavrova. Nonetheless, thisplacement may be provisional since thenomenclatural type of Macrosciadium, whichis M. alatum (M. Bieb.) V. N. Tikhom. etLavrova, has not been hitherto considered inmolecular analyses either. Macrosciadiumphysospermifolium seems to be very closelyrelated to Cnidiocarpa alaica, the nomencla-tural type of Cnidiocarpa Pimenov. Therefore,Macrosciadium may eventually appear to be ataxonomic synonym of Cniodiocarpa.
Lomatium. This clade was weakly mono-phyletic in the results of the NJ and MLanalyses and formed two clades in the MPtrees. Its members are endemic to central andwestern North America. They are all perennialtaxa and are characterised by several non-molecular characters, such as a protogynousbreeding system and the lack of a prominentconical-shaped stylopodium. The phylogeny ofthis group was recently investigated by Downieet al. (2002) and Sun et al. (in press). Theydemonstrated that upon comprehensive sam-pling, this group forms a clade, albeit it is stillweakly supported.
Conclusions
With increased sampling, resolution withinthe previously delimited ‘Angelica’ and ‘Ar-racacia’ clades, recognised herein as tribe
Selineae, increases and several subcladesbecome apparent. Some of these subcladesare more or less equivalent to currentlyrecognised genera, while others may eventu-ally be formally recognised as such. Rela-tively low ITS sequence variation – ascompared, for instance, with the high vari-ability exhibited by Apiaceae tribe Scandi-ceae and its subtribes (Downie et al. 2000a) –coupled with a large number of included taxamakes the reclassification of this branch ofumbellifers particularly difficult. Polyphyly ofits included genera and low internal supportfor most hitherto identified clades indicatethat this task would require a broadersampling, both with respect to the numberof taxa and the sources of data providinginformation on relatedness.
The authors thank Fengjie Sun, AnetaWojewodzka, and Deborah Katz-Downie forlaboratory assistance and Alexei Oskolski andWalter Till for checking the types. This researchwas supported by grants from the Polish Commit-tee for Scientific Research (6 P04C 039 21), WarsawUniversity (BW 1455/12/99), and the NationalScience Foundation (DEB 0089452).
References
Arenas Posada J. A., Garcıa Martın F. (1993) Atlascarpologico y corologico de la subfamilia Apioi-deae Drude (Umbelliferae) en Espana peninsulary Baleares. Ruizia 12: 1–245.
Bremer K. (1994) Branch support and tree stability.Cladistics 10: 295–304.
Calestani V. (1905) Conspectus specierum euro-paearum generis Peucedani. Bull. Soc. Bot. Ital.ann. 1905: 193–201.
Catalan P., Kellogg E. A., Olmstead R. G. (1997)Phylogeny of Poaceae subfamily Pooideae basedon chloroplast ndhF gene sequences. Mol. Phyl.Evol. 8: 150–166.
Chamberlain D. F. (1972). Xanthogalum Lallem.In: Davis P. H. (ed.) Flora of Turkey and theEast Aegean Islands, vol. 4. Edinburgh Univer-sity Press, Edinburgh, pp. 432–434.
Downie S. R., Katz-Downie D. S. (1996) A molec-ular phylogeny of Apiaceae subfamily Apioi-
208 K. Spalik et al.: The phylogenetic position of Peucedanum sensu lato and allied genera

deae: evidence from nuclear ribosomal DNAinternal transcribed spacer sequences. Amer.J. Bot. 83: 234–251.
Downie S. R., Katz-Downie D. S. (1999) Phyloge-netic analysis of chloroplast rps16 intronsequences reveals relationships within the woodysouthern African Apiaceae subfamily Apioideae.Canad. J. Bot. 77: 1120–1135.
Downie S. R., Katz-Downie D. S., Spalik K.(2000a) A phylogeny of Apiaceae tribe Scandi-ceae: evidence from nuclear ribosomal DNAinternal transcribed spacer sequences. Amer.J. Bot. 87: 76–95.
Downie S. R., Ramanath S., Katz-Downie D. S.,Llanas E. (1998) Molecular systematics of Api-aceae subfamily Apioideae: phylogenetic analy-ses of nuclear ribosomal DNA internaltranscribed spacer and plastid rpoC1 intronsequences. Amer. J. Bot. 85: 563–591.
Downie S. R., Watson M. F., Spalik K., Katz-Downie D. S. (2000b) Molecular systematics ofOld World Apioideae (Apiaceae): relationshipsamong some members of tribe Peucedaneaesensu lato, the placement of several islandendemics, and resolution within the apioidsuperclade. Canad. J. Bot. 78: 506–528.
Downie S. R., Plunkett G. M., Watson M. F.,Spalik K., Katz-Downie D. S., Valiejo-RomanK. M., Terentieva E. I., Troitsky A. V., LeeB.-Y., Lahham J., El-Oqlah A. (2001) Tribesand clades within Apiaceae subfamily Apioi-deae: the contribution of molecular data.Edinburgh J. Bot. 58: 301–330.
Downie S. R. Hartman R. L., Sun F.-J.,Katz-Downie D. S. (2002) Polyphyly of thespring-parsleys (Cymopterus): molecular andmorphological evidence suggests complexrelationships among the perennial endemicgenera of western North American Apiaceae.Canad. J. Bot. 80: 1295–1324.
Drude C. G. O. (1897–1898) Umbelliferae. In:Engler A., Prantl K. (eds.) Die naturlichenPflanzenfamilien, vol. 3. Wilhelm Engelmann,Leipzig, pp. 63–250.
Durrieu G. (1982) Les champignons parasiteset leur apport a la systematique des Ombelliferes.In: Cauwet-Marc A.-M., Carbonnier J. (eds.)Les Ombelliferes ‘‘Contributions pluridiscipli-naires a la systematique.’’ Actes du 2eme Sym-posium International sur les Ombelliferes.Monographs in Systematic Botany from the
Missouri Botanical Garden, vol. 6. Braun-Brum-field, Ann Arbor, Michigan, pp. 525–533.
Eriksson T. (1998) AutoDecay ver. 4.0 (programdistributed by the author). Department ofBotany, Stockholm University, Stockholm.
Felsenstein J. (1985) Confidence limits on phylog-enies: an approach using the bootstrap. Evolu-tion 39: 783–791.
Frey R. (1989) Taxonomische Revision der Gat-tung Peucedanum: Sektion Peucedanum undSektion Palimbioidea (Umbelliferae). Candollea44: 257–327.
Guyot M. (1984) Les types stomatiques dans lesespeces francaises du genre Peucedanum (Om-belliferes). Revue Cytol. Biol. Veg. Bot. 7: 17–30.
Hadacek F. (1989) Vergleichende phytochemischeUntersuchungen in der Gattung Peucedanum(Apiaceae - Apioideae). Stapfia 18: 1–186.
Hardig T. M., Soltis P. S. (1999) An ITS-basedphylogenetic analysis of the Euryptera speciesgroup in Lomatium (Apiaceae). Plant Syst. Evol.219: 65–78.
Katz-Downie D. S., Valiejo-Roman C. M.,Terentieva E. I., Troitsky A. V., Pimenov M.G., Lee B.-Y., Downie S. R. (1999) Towards amolecular phylogeny of Apiaceae subfamilyApioideae: additional information from nuclearribosomal DNA ITS sequences. Plant Syst. Evol.216: 167–195.
Leute G.-H. (1966) Die Gattungen Imperatoria L.und Tommasinia Bertol. (Apiaceae). Ann. Na-turhist. Mus. Wien 69: 69–79.
Menitsky G. L. (1991) Konspekt vidov semejstvaApiaceae flory Kakvkaza. Bot. Zhurn. 76:1749–1764.
Muckensturm B., Diyani F. (1995) Rapportpreliminaire sur les constituants volatils deSelinum pyrenaeum, Selinum carvifolia. Univ.Haute-Alsace, Mulhouse.
Ostroumova T. A., Pimenov M. G. (1997) Carpo-logical diversity of African Peucedanum s.l.(Umbelliferae). I. The species of southern Africa.Feddes Repert. 108: 299–318.
Pimenov M. G. (1986) Kitagawia – a new Asiaticgenus of the family Umbelliferae. Bot. Zhurn.71: 942–949.
Pimenov M. G. (1987) Cervaria. In: Hedge I. C.,Lamond J. M., Rechinger K. H., et al. Umbel-liferae. In: Rechinger K. H. (ed.) Flora iranica,vol. 162. Akademische Druck- und Verlagsan-stalt, Graz, pp. 83–110.
K. Spalik et al.: The phylogenetic position of Peucedanum sensu lato and allied genera 209

Pimenov M. G., Leonov M. V. (1993) The generaof Umbelliferae. Royal Botanical Gardens, Kew.
Plunkett G. M., Soltis D. E., Soltis P. S. (1996)Evolutionary patterns in Apiaceae: inferencesbased on matK sequence data. Syst. Bot. 21:477–495.
Posada D., Crandall K. A. (1998) Modeltest:testing the model of DNA substitution. Bioin-formatics 14: 817–818.
Pu F. T., Wang P. L., Zheng Z. H., Wang Y. P.(2000) A reclassification of Notopterygium Bos-sieu (Umbelliferae). [In Chinese]. Acta Phytotax.Sin. 38: 430–436.
Reduron J.-P., Charpin A., Pimenov M. G. (1997)Contribution a la nomenclature generique desApiaceae (Ombelliferes). J. Bot. Soc. bot. Fr.1: 91–104.
Shneyer V. S., Kutyavina N. G., Pimenov M. G.(2003) Systematic relationships within and be-tween Peucedanum and Angelica (Umbelliferae-Peucedaneae) inferred from immunological stud-ies of seed protein. Plant Syst. Evol. 236: 175–194.
Spalik K., Wojewodzka A., Downie S. R. (2001)The delimitation of genera in Apiaceae withexamples from Scandiceae subtribe Scandicinae.Edinburgh J. Bot. 58: 331–346.
Sun F.-J, Downie S. R., Hartman R. L. (2004) AnITS-based phylogenetic analysis of the perennial,endemic Apiaceae subfamily Apioideae ofwestern North America. Syst. Bot. (in press).
Swofford D. L. (1998) PAUP*. Phylogenetic Anal-ysis Using Parsimony (*and other methods).Version 4. Sinauer Associates, Sunderland.
Tamura K., Nei M. (1993) Estimation ofthe number of nucleotide substitution inthe control region of mitochondrial DNA inhumans and chimpanzees. Mol. Biol. Evol. 10:512–526.
Theobald W. L. (1971) Comparative anatomicaland developmental studies in the Umbelliferae.In: Heywood V. H. (ed.) The biology andchemistry of the Umbelliferae. Academic Press,London, pp. 177–197.
Thompson J. D., Gibson T. J., Plewniak F.,Jeanmougin F., Higgins D. G. (1997) TheClustalX windows interface: flexible strategiesfor multiple sequence alignment aided byquality analysis tools. Nucleic Acids Res. 24:4876–4882.
Tutin T. G., Heywood V. H., Burges N. A., MooreD. M., Valentine D. H., Walters S. M., Webb D.A. (eds.) (1968) Flora Europaea. Vol. 2. Cam-bridge University Press, Cambridge.
Valiejo-Roman C. M., Terentieva E. I., SamigullinT. H., Pimenov M. G. (2002) nrDNA ITSsequences and affinities of Sino-HimalayanApioideae (Umbelliferae). Taxon 51: 685–701.
Vasil’eva M. G., Pimenov M. G. (1991)Karyotaxonomical analysis in the genus Angel-ica (Umbelliferae). Plant Syst. Evol. 177: 117–138.
Addresses of the authors: Krzysztof Spalik(e-mail: [email protected]), Zakład Systemat-yki i Geografii Roslin, Uniwersytet Warszawski,Aleje Ujazdowskie 4, PL-00-478 Warszawa,Poland. Jean-Pierre Reduron, Communaute del’Agglomeration mulhousienne 1, rue des Orphe-lins, 68200 Mulhouse, France. Stephen R. Downie,Department of Plant Biology, University ofIllinois, 505 S Goodwin Ave., Urbana, IL 61801,USA.
210 K. Spalik et al.: The phylogenetic position of Peucedanum sensu lato and allied genera





![Production and quality of fennel - Acta Universitatis Sapientiae · Fennel belongs to the family Apiaceae (Umbelliferae). It was taxonomically ranged by Keller et al. [in 5]. The](https://img.pdfslide.us/doc/110x75/5feca14016f52b650c3a4b36/production-and-quality-of-fennel-acta-universitatis-fennel-belongs-to-the-family.jpg)























