Embed Size (px)
Citation preview
LETTERS
The molecular basis for water taste in DrosophilaPeter Cameron1, Makoto Hiroi1, John Ngai2 & Kristin Scott1,3
The detection of water and the regulation of water intake areessential for animals to maintain proper osmotic homeostasis1.Drosophila and other insects have gustatory sensory neurons thatmediate the recognition of external water sources2–4, but little isknown about the underlying molecular mechanism for water tastedetection. Here we identify a member of the degenerin/epithelialsodium channel family5, PPK28, as an osmosensitive ion channelthat mediates the cellular and behavioural response to water. Weuse molecular, cellular, calcium imaging and electrophysiologicalapproaches to show that ppk28 is expressed in water-sensing neu-rons, and that loss of ppk28 abolishes water sensitivity. Moreover,ectopic expression of ppk28 confers water sensitivity to bitter-sensing gustatory neurons in the fly and sensitivity to hypo-osmotic solutions when expressed in heterologous cells. Thesestudies link an osmosensitive ion channel to water taste detec-tion and drinking behaviour, providing the framework for
examining the molecular basis for water detection in otheranimals.
To uncover molecules involved in taste detection, we performed amicroarray-based screen for genes expressed in taste neurons.Proboscis RNA from flies homozygous for a recessive poxn nullmutation was compared to RNA from heterozygous controls. poxnmutants have a transformation of labellar gustatory chemosensorybristles into mechanosensory bristles, and therefore lack all tasteneurons6,7. Whole-genome microarray comparisons showed that256 of ,18,500 transcripts were significantly decreased in poxnmutants (.2-fold enrichment in control relative to poxn, P , 0.05,moderated t-test). These included 18 gustatory receptors (represent-ing a 21-fold enrichment in the gene set) and eight odorant-bindingproteins (13-fold enrichment) (Supplementary Fig. 1 and Sup-plementary Table 1; Gene Expression Omnibus (GEO) accessionnumber GSE19984).
1Department of Molecular and Cell Biology and Helen Wills Neuroscience Institute, 16 Barker Hall, 2Department of Molecular and Cell Biology, Helen Wills Neuroscience Institute andFunctional Genomics Laboratory, 142 Life Sciences Addition, 3Howard Hughes Medical Institute, University of California-Berkeley, Berkeley, California 94720, USA.
16
12
8
4Max
imum
ΔF/
F (%
)
a b
dc
e
Proboscis Brain
PPK28 PPK28GR5A GR66A
NaCl Sucrose Ribose NMDG PEG0 0.2 0.5 1 M 0 0.2 0.5 1 M 0 0.2 0.5 1 M 0 0.2 0.5 1 M 0 8 20%
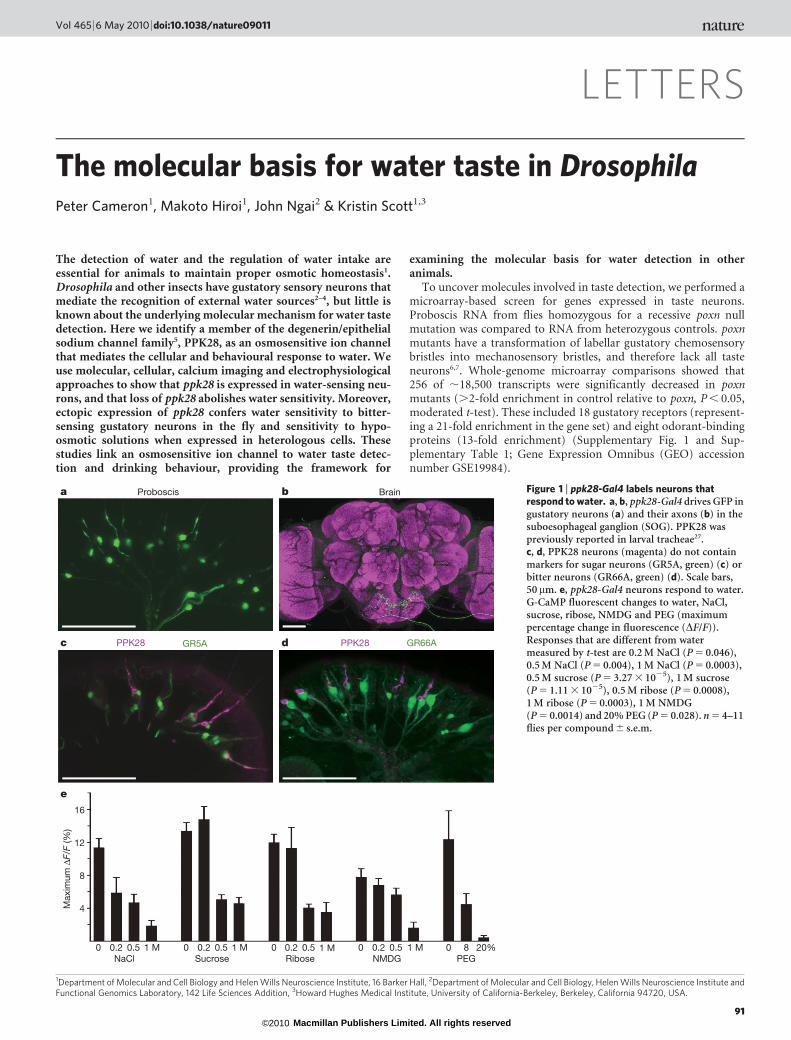
Figure 1 | ppk28-Gal4 labels neurons thatrespond to water. a, b, ppk28-Gal4 drives GFP ingustatory neurons (a) and their axons (b) in thesuboesophageal ganglion (SOG). PPK28 waspreviously reported in larval tracheae27.c, d, PPK28 neurons (magenta) do not containmarkers for sugar neurons (GR5A, green) (c) orbitter neurons (GR66A, green) (d). Scale bars,50mm. e, ppk28-Gal4 neurons respond to water.G-CaMP fluorescent changes to water, NaCl,sucrose, ribose, NMDG and PEG (maximumpercentage change in fluorescence (DF/F)).Responses that are different from watermeasured by t-test are 0.2 M NaCl (P 5 0.046),0.5 M NaCl (P 5 0.004), 1 M NaCl (P 5 0.0003),0.5 M sucrose (P 5 3.27 3 1025), 1 M sucrose(P 5 1.11 3 1025), 0.5 M ribose (P 5 0.0008),1 M ribose (P 5 0.0003), 1 M NMDG(P 5 0.0014) and 20% PEG (P 5 0.028). n 5 4–11flies per compound 6 s.e.m.
Vol 465 | 6 May 2010 | doi:10.1038/nature09011
91Macmillan Publishers Limited. All rights reserved©2010

In the mammalian gustatory system, ion channels mediate thedetection of sour and salt tastes8, suggesting that ion channel genesmay also participate in Drosophila taste detection. We thereforeexamined the expression pattern of candidate taste-enriched ionchannels. The putative promoter of one gene, pickpocket 28(ppk28), directed robust reporter expression in taste neurons onthe proboscis (Fig. 1a). PPK28 belongs to the degenerin/epithelialsodium channel family (Deg/ENaC) and these channels are involvedin the detection of diverse stimuli, including mechanosensory stimuli,acids and sodium ions5. In the brain, ppk28-Gal4 drives expression ofgreen fluorescent protein (GFP) in gustatory sensory axons thatproject to the primary taste region, the suboesophageal ganglion(Fig. 1b and Supplementary Fig. 2). In situ hybridization experimentsconfirmed that transgenic expression recapitulates that of the endo-genous gene, as 48 out of 52 of ppk28-Gal4 neurons expressed endo-genous ppk28.
Previous studies have identified different taste cell populations inthe proboscis, including cells labelled by the gustatory receptor GR5A(encoded by Tre) that respond to sugars9–12 and cells marked byGR66A that respond to bitter compounds10–13. To determine whetherthese taste neurons express ppk28-Gal4, we performed co-labellingexperiments with reporters for GR5A and GR66A. These experimentsshowed that PPK28 did not co-label GR5A cells or GR66A cells, and isthus unlikely to participate in sweet or bitter taste detection (Fig. 1c,d). An enhancer-trap Gal4 line, NP1017-Gal4, labels water-sensingneurons in taste bristles on the proboscis4 and carbonation-sensingneurons in taste pegs14 (Supplementary Fig. 3). PPK28 is expressed intaste bristles but not in taste pegs. Notably, ppk28 showed partial co-expression with NP1017-Gal4 (Supplementary Fig. 3), with most
PPK28-positive cells containing NP1017-Gal4 (22 out of 30). Thiscorrelation suggested the intriguing possibility that PPK28 partici-pates in water taste detection.
To investigate directly the response-specificity of ppk28-expressingneurons, we expressed the genetically encoded calcium sensorG-CaMP in ppk28-Gal4 cells, stimulated the proboscis with tastesubstances and monitored activation of ppk28-Gal4 projections inthe living fly by confocal microscopy12. We tested ppk28-Gal4 neu-rons with a panel of taste solutions, including sugars, bitter com-pounds, salts, acids and water. ppk28-Gal4 neurons showed robustactivity after water stimulation (Fig. 1e). In addition, PPK28-positivecells responded to other aqueous solutions even in the presence ofa wide range of chemically distinct compounds. This response dimi-nished as a function of concentration. Taste compounds such asNaCl, sucrose and citric acid significantly decreased the response(Fig. 1e and Supplementary Fig. 4). In addition, compounds unlikelyto elicit taste cell activity such as ribose, a sugar that does not activateGR5A cells, N-methyl-D-glucamine (NMDG), an impermeantorganic cation, and the non-ionic high molecular mass polymerpolyethylene glycol (PEG, 3,350 g mol21), all blunted the responsein a concentration-dependent manner (Fig. 1e and Supplemen-tary Fig. 4). These data demonstrate that ppk28-expressing neuronsrespond to hypo-osmotic solutions. This response profile is consist-ent with previous electrophysiological studies that identified a classof labellar taste neurons activated by water and inhibited by salts,sugars and amino acids4,15.
To determine the function of PPK28 in the water response, wegenerated a ppk28-null mutant by piggyBac-transposon-mediated genedeletion, removing 1.769 kilobases (kb) surrounding the ppk28 gene16.
a
b
c
d
e
60
40
20
ppk28 control ppk28 mutant ppk28 rescueWater Sucrose
Sp
ikes
s–1
Inge
stio
n tim
e (s
)
8
4
50
Control Mutant Rescue Control Mutant Rescue
100
10
20
30
100 ms
3 m
V
ΔF/F
(%)
2
4
6
12
Max
imum
ΔF/
F (%
)
ppk28 controlppk28 mutantppk28 rescue
Controlppk28ppk28; ppk28-Gal4ppk28; UAS-ppk28ppk28; ppk28-Gal4,UAS-ppk28
Control
Mutant
Rescue
0
4
8
12
16
Water
Water Sucrose
0.1 M NaCl 1 M NaCl 1 M sucrose
***
*** ***
*
*
*
***
**
Figure 2 | The ppk28 gene is necessary for cellular and behavioural waterresponses. a, Extracellular bristle recordings of ppk28 control, mutant andrescue flies after water (left) or 100 mM sucrose (right) stimulation, showingaction potentials. Stimulation begins at recording. b, Scatter plot of waterand sugar responses (mean 6 s.e.m. in bars; data points as dots). Waterresponses are ***P 5 0.001 by Dunn’s multiple comparison. c, G-CaMPfluorescence increase in ppk28 control, mutant and rescue projections towater (SOG, scale bar, 50 mm). d, Fluorescence change summary after water,0.1 M NaCl, 1 M NaCl and 1 M sucrose (n 5 8–11 trials per
concentration 6 s.e.m.; t-test, ppk28 control versus mutant, water:***P 5 0.0008, 1 M NaCl: *P 5 0.03). e, Behavioural assays measuring wateror 500 mM sucrose consumption time. Control flies drink more water thanppk28 mutants (*P 5 0.017), ppk28 mutants 1 ppk28-Gal4 (*P 5 0.037) orppk28 mutants 1 UAS-ppk28 (**P 5 0.008). Water consumption of controland rescue is not different (P 5 0.53). Sucrose consumption is not different(versus control, mutant: P 5 0.63; rescue: P 5 0.53). n 5 3 6 s.e.m. trials,18–25 flies per trial per genotype, t-test.
LETTERS NATURE | Vol 465 | 6 May 2010
92Macmillan Publishers Limited. All rights reserved©2010

We examined the water responses of ppk28 control, mutant and rescueflies by extracellular bristle recordings of l-type labellar taste sensilla.These recordings monitor the responses of the four gustatory neuronsin a bristle, including water cells and sugar cells3. Control flies showed12.0 6 0.9 spikes s21 (mean 6 s.e.m.) when stimulated with water(Fig. 2a, b). Remarkably, ppk28 mutant cells showed a complete lossof the response to water (spikes s21 5 0.8 6 0.1). This response waspartially rescued by reintroduction of ppk28 into the mutant back-ground (spikes s21 5 6.4 6 1.0), demonstrating that defects weredue to the loss of ppk28 (Fig. 2a, b). The responses to sucrose werenot significantly different among the three genotypes (58.9 6 3.3spikes s21, 46.9 6 2.6 spikes s21 and 49.0 6 1.8 spikes s21, for control,mutant and rescue flies, respectively) (Fig. 2a, b), arguing that the lossof ppk28 specifically eliminates the water response. These results wereconfirmed by G-CaMP imaging experiments that monitor the res-ponse of the entire ppk28 population. As expected, ppk28-Gal4 neu-rons in the mutant did not show fluorescent increases to water andtransgenic re-introduction of ppk28 rescued the water response(Fig. 2c, d). Taken together, the electrophysiological and imaging datademonstrate that PPK28 is required for the cellular response to water.
The detection of water in the environment and the internal state ofthe animal may both contribute to drive water consumption1. Toevaluate the degree to which water taste detection contributes toconsumption, we examined the behavioural responses of ppk28control, mutant and rescue flies to water. Drinking time rather thandrinking volume was used to monitor consumption due to difficultyin reliably detecting small volume changes. When presented with awater stimulus, control flies drank on average for 10.3 6 1.1 s(mean 6 s.e.m.), mutants drank 3.0 6 0.3 s and rescue flies drank
11.5 6 1.5 s (Fig. 2e). Furthermore, control, mutant and rescue fliesingested sucrose equally, showing that ppk28 mutants do not havegeneral drinking defects. Similar defects in water detection were seenwhen control, mutant and rescue flies were tested on the proboscisextension reflex to water (Supplementary Fig. 5a), or when genetic-ally ablating ppk28-Gal4 neurons (Supplementary Fig. 5b). Althoughppk28 mutants lack water taste cell responses and drink less, they stilldo consume water, arguing that other mechanisms must exist toensure water uptake. These experiments show that water taste neuronsare necessary for normal water consumption. Moreover, they establisha link between water taste detection in the periphery and the drive todrink water.
We next examined whether PPK28 is directly involved in waterdetection. If PPK28 is the water sensor, then its expression in non-water sensing cells should bestow responsiveness to water. To testthis, we used the Gal4/UAS system to ectopically express ppk28 inGr66a-expressing, bitter-sensing neurons and monitored taste-induced responses by extracellular bristle recordings and G-CaMPimaging experiments (Fig. 3). For extracellular bristle recordings, res-ponses were recorded from i-type sensilla that contain bitter-sensing,GR66A-positive neurons but lack water cells17. Expression of ppk28 inGr66a-Gal4 neurons did not significantly affect the response to dena-tonium (G-CaMP imaging: maximum percentage change in fluor-escence (%DF/F) 5 11.9 6 1.2; misexpression %DF/F 5 13.8 6 0.7) orcaffeine (control 18.8 6 3.0 spikes s21; misexpression 20.6 6 2.9spikes s21; Fig. 3a, b), endogenous ligands for Gr66a-Gal4 neurons12.In response to water, Gr66a-Gal4 neurons showed no significant activ-ity, consistent with previous studies (Fig. 3)12. Notably, misexpression ofppk28 in Gr66a-Gal4 neurons conferred sensitivity to water, as seen by
a
b
c
d
e
Water
0.5 M NMDG
1 M NMDG
Caffeine
Water NMDG Caffeine NMDG Sucrose0 0.5 1 10.01 M 2 M10.50.5 00
3040
30
20
10
Sp
ikes
s–1
10
Max
imum
ΔF/
F (%
)
16
ΔF/F
(%)
3 s 4% Δ
F/F
100 ms
3 m
V
Gr66a-Gal4
Gr66a-Gal4; UAS-ppk28
8
0
8
6
4
2
20
10
– ppk28 + ppk28
– ppk28 GR66A GR66A + ppk28+ ppk28
******
**
**
*
*
Figure 3 | Ectopic expression of ppk28 confers water sensitivity.a, Extracellular bristle recordings of i-type sensilla (non-water responsive)from Gr66a-Gal4 flies lacking (2) or containing (1) UAS-ppk28 after water,0.5 M NMDG, 1 M NMDG or 0.01 M caffeine stimulation (at recording).b, Scatter plot of water responses (mean 6 s.e.m. in bars; data points aredots) and summary of all responses (mean 6 s.e.m.). Responses are differentto water (***P 5 2.52 3 10216) and 0.5 M NMDG (*P 5 0.016) (t-test;n 5 7–27). c, G-CaMP fluorescence increase in GR66A bitter-sensing
projections (left) and GR66A projections expressing ppk28 (right), afterwater stimulation (maximumDF/F) (SOG, scale bar, 50 mm). d, Responses ofGR66A cells (left) and GR66A cells expressing ppk28 (right) to water (atarrow). e, Summary of fluorescence changes in GR66A cells without (grey)or with (green) ppk28 tested with water, 0.5 and 1 M NMDG and 0.5, 1 and2 M sucrose. n 5 4–5 trials per concentration 6 s.e.m.; t-test, versus GR66Acontrol, water: **P 5 0.013, 0.5 M NMDG: *P 5 0.03; water: **P 5 0.002,0.5 M sucrose: ***P 5 0.0002.
NATURE | Vol 465 | 6 May 2010 LETTERS
93Macmillan Publishers Limited. All rights reserved©2010
extracellular bristle recordings (Fig. 3a, b) and G-CaMP imaging(Fig. 3c–e). Moreover, the response was blunted as solute concentrationwas increased. Both NMDG and sucrose (substances that do not acti-vate Gr66a-Gal4 neurons) produced dose-sensitive response decreases.The finding that both activation by water and inhibition by othercompounds are conferred by PPK28 strongly suggests that PPK28 senseslow osmolarity.
To determine whether PPK28 requires a taste cell environment tofunction or confers responsiveness to other cell-types, ppk28 wasexpressed in HEK293 heterologous cells. A Flag-tagged PPK28 (insertedafter amino acid 222 in the extracellular domain) was expressed inHEK293 cells, confirming that the protein was made and trafficked tothe cell surface (Supplementary Fig. 6). For calcium-imaging experi-ments, an untagged version of PPK28 was co-transfected with the fluor-escent marker dsRed. Cells expressing the mammalian TRPV4osmosensitive ion channel18 were used as a positive control, and cellstransfected with the vector alone as a negative control. Cells were grownin a modified Ringer’s solution at 303 mmol kg21, loaded with Fluo-4 tovisualize calcium changes and challenged with Ringer’s solution of dif-ferent osmolalities (236, 216 and 174 mmol kg21; 80%, 70% and 60%osmotic strength to the isotonic solution, respectively). Cells transfected
with vector alone showed a modest increase at 60% osmotic strength,whereas cells transfected with mammalian Trpv4 showed fluorescenceincreases to all hypo-osmotic solutions, as expected (Fig. 4b–d).Notably, cells transfected with ppk28 significantly responded todecreased osmolality, with dose-sensitive responses elicited by osmolal-ities of 216 and 174 mmol kg21 (Fig. 4a, d). These experiments show thatppk28 bestows sensitivity to hypo-osmotic solutions in a variety of non-native environments and argue that the channel itself senses low osmo-larity. This work provides a foundation for future studies of the bio-physical properties of channel activation. Moreover, the ability toexpress PPK28 in heterologous cells and study its function creates theopportunity to compare its mechanism of gating with other Deg/ENaCfamily members involved in mechanosensation or sodium sensing.
Overall, these studies examined the molecular basis for water tastedetection in Drosophila and identified an ion channel belonging tothe Deg/ENaC family, PPK28, as the water gustatory sensor. Ourwork demonstrates that an ion channel responding to low osmolaritymediates cellular and behavioural responses to water. Although thetaste of water has received relatively little attention as a classic tastemodality, water-responsive taste neurons have been described inmany other insects, such as the blowfly and mosquitoes2,19, as wellas in mammals, such as cats and rats20,21. The identification of PPK28as a water taste receptor provides a framework for examining watertaste detection in other animals, including humans.
Osmosensation is important not only for the detection of externalwater sources by peripheral neurons but also for monitoring theplasma osmolality by central neurons1. Several studies have identifiedmembers of the transient receptor potential family as candidate peri-pheral and central osmosensors18,22–24, but the role of members of theDeg/ENaC family in osmosensation has received little attention. Ourfinding that PPK28 is an osmosensitive ion channel raises the pos-sibility that Deg/ENaC ion channels may participate broadly in peri-pheral and central osmosensation.
METHODS SUMMARYTransgenic flies and ppk28 mutants. The ppk28 promoter-Gal4 construct was
generated with a 1.004-kb upstream fragment (16699333–16700336, GenBank
accession number NC_004354.3). Full-length ppk28 (transcript variant a,
NM_132941) was subcloned into pUAST. ppk28 mutants were generated by
FLP-FRT-mediated recombination between piggyBac transposons f05788 and
e02329.
Immunohistochemistry and in situ hybridization. Labelling was performed as
described11. In Fig. 1b, the brain was counterstained with nc82 (ref. 25). In Fig. 1c,
d, CD2 and GFP were detected by immunohistochemistry on flies containing
ppk28-Gal4, UAS-CD2, Gr66a-GFP-IRES-GFP-IRES-GFP or Gr5a-GFP-IRES-GFP-IRES-GFP transgenes11,12.
G-CaMP imaging experiments. Imaging studies were performed as described12.
Further details are in the Methods.
Behavioural assays. Control flies were isogenic w1118 fly strain (Exelixis strain
A5001, BL-6326) and transgenes were backcrossed seven times to the control
strain. 2–5-day-old flies were starved for ,18–24 h with water, kept in a humid
chamber for 2–3 h and then stimulated on the proboscis with a taste substance.
Flies were allowed to ingest freely until they did not ingest after five consecutive
stimulations. For water ingestion, flies were stimulated on the proboscis with
1 M sucrose afterward, and only responders were kept for data tally.
Electrophysiology. Electrophysiology was performed as described26. Further
details are in the Methods.
HEK293 calcium-imaging experiments. Measurements in cells were made byusing calcium indicator Fluo-4 and a confocal laser scanning microscope (Zeiss
LSM510, Carl Zeiss). Further details are in the Methods.
Full Methods and any associated references are available in the online version ofthe paper at www.nature.com/nature.
Received 16 November 2009; accepted 16 March 2010.Published online 4 April 2010.
1. Bourque, C. W. Central mechanisms of osmosensation and systemicosmoregulation. Nature Rev. Neurosci. 9, 519–531 (2008).
2. Evans, D. R. & Mellon, D. Jr. Electrophysiological studies of a water receptorassociated with the taste sensilla of the blow-fly. J. Gen. Physiol. 45, 487–500(1962).
PPK28
TRPV4
Vector
Max
imum
ΔF
Osmotic strength (%) to 303 mmol kg–1
40 PPK28TRPV4Vector
50 s
5 ΔF
80%70%60%
80%70%60%
80%70%60%
a
b
c
d
30
20
10
7080 60
303 174
*****
**
**
**
Figure 4 | Heterologous cells expressing PPK28 respond to hypo-osmolarity. a–c, Pseudocolour images of maximum fluorescence increasesto isotonic (303 mmol kg21) and reduced (174 mmol kg21) osmolality forHEK293 cells expressing PPK28, TRPV4 or vector. Colour bar indicatesmaximum DF ranging from 210 to 80. On the right, plots of fluorescencechange per frame over the stimulation (bar) at 80%, 70% and 60% ofisotonic osmolality (236, 216 and 174 mmol kg21). d, Concentration curveof responses to osmolalities. n 5 4–5 trials per concentration 6 s.e.m.; t-test,versus vector, PPK28: 70% **P 5 0.00583; 60% **P 5 0.00632; TRPV4:80% **P 5 0.00384; 70% ***P 5 0.000120; 60% **P 5 0.00615.
LETTERS NATURE | Vol 465 | 6 May 2010
94Macmillan Publishers Limited. All rights reserved©2010

3. Meunier, N., Ferveur, J. F. & Marion-Poll, F. Sex-specific non-pheromonal tastereceptors in Drosophila. Curr. Biol. 10, 1583–1586 (2000).
4. Inoshita, T. & Tanimura, T. Cellular identification of water gustatory receptorneurons and their central projection pattern in Drosophila. Proc. Natl Acad. Sci. USA103, 1094–1099 (2006).
5. Kellenberger, S. & Schild, L. Epithelial sodium channel/degenerin family of ionchannels: a variety of functions for a shared structure. Physiol. Rev. 82, 735–767(2002).
6. Awasaki, T. & Kimura, K. pox-neuro is required for development of chemosensorybristles in Drosophila. J. Neurobiol. 32, 707–721 (1997).
7. Boll, W. & Noll, M. The Drosophila Pox neuro gene: control of male courtshipbehavior and fertility as revealed by a complete dissection of all enhancers.Development 129, 5667–5681 (2002).
8. Yarmolinsky, D. A., Zuker, C. S. & Ryba, N. J. Common sense about taste: frommammals to insects. Cell 139, 234–244 (2009).
9. Chyb, S., Dahanukar, A., Wickens, A. & Carlson, J. R. Drosophila Gr5a encodes ataste receptor tuned to trehalose. Proc. Natl Acad. Sci. USA 100 (suppl. 2),14526–14530 (2003).
10. Thorne, N., Chromey, C., Bray, S. & Amrein, H. Taste perception and coding inDrosophila. Curr. Biol. 14, 1065–1079 (2004).
11. Wang, Z., Singhvi, A., Kong, P. & Scott, K. Taste representations in the Drosophilabrain. Cell 117, 981–991 (2004).
12. Marella, S. et al. Imaging taste responses in the fly brain reveals a functional mapof taste category and behavior. Neuron 49, 285–295 (2006).
13. Moon, S. J., Kottgen, M., Jiao, Y., Xu, H. & Montell, C. A taste receptor required forthe caffeine response in vivo. Curr. Biol. 16, 1812–1817 (2006).
14. Fischler, W., Kong, P., Marella, S. & Scott, K. The detection of carbonation by theDrosophila gustatory system. Nature 448, 1054–1057 (2007).
15. Meunier, N., Marion-Poll, F. & Lucas, P. Water taste transduction pathway iscalcium dependent in Drosophila. Chem. Senses 34, 441–449 (2009).
16. Parks, A. L. et al. Systematic generation of high-resolution deletion coverage of theDrosophila melanogaster genome. Nature Genet. 36, 288–292 (2004).
17. Hiroi, M., Meunier, N., Marion-Poll, F. & Tanimura, T. Two antagonistic gustatoryreceptor neurons responding to sweet-salty and bitter taste in Drosophila. J.Neurobiol. 61, 333–342 (2004).
18. Liedtke, W. et al. Vanilloid receptor-related osmotically activated channel (VR-OAC), a candidate vertebrate osmoreceptor. Cell 103, 525–535 (2000).
19. Werner-Reiss, U., Galun, R., Crnjar, R. & Liscia, A. Sensitivity of the mosquitoAedes aegypti (Culicidae) labral apical chemoreceptors to blood plasmacomponents. J. Insect Physiol. 45, 485–491 (1999).
20. Lindemann, B. Taste reception. Physiol. Rev. 76, 718–766 (1996).21. Gilbertson, T. A. Hypoosmotic stimuli activate a chloride conductance in rat taste
cells. Chem. Senses 27, 383–394 (2002).22. Colbert, H. A., Smith, T. L. & Bargmann, C. I. OSM-9, a novel protein with
structural similarity to channels, is required for olfaction, mechanosensation, andolfactory adaptation in Caenorhabditis elegans. J. Neurosci. 17, 8259–8269 (1997).
23. Muraki, K. et al. TRPV2 is a component of osmotically sensitive cation channels inmurine aortic myocytes. Circ. Res. 93, 829–838 (2003).
24. Liu, L. et al. Drosophila hygrosensation requires the TRP channels water witch andnanchung. Nature 450, 294–298 (2007).
25. Hummel, T., Krukkert, K., Roos, J., Davis, G. & Klambt, C. Drosophila Futsch/22C10is a MAP1B-like protein required for dendritic and axonal development. Neuron 26,357–370 (2000).
26. Hiroi, M., Marion-Poll, F. & Tanimura, T. Differentiated response to sugars amonglabellar chemosensilla in Drosophila. Zoolog. Sci. 19, 1009–1018 (2002).
27. Liu, L., Johnson, W. A. & Welsh, M. J. Drosophila DEG/ENaC pickpocket genes areexpressed in the tracheal system, where they may be involved in liquid clearance.Proc. Natl Acad. Sci. USA 100, 2128–2133 (2003).
Supplementary Information is linked to the online version of the paper atwww.nature.com/nature.
Acknowledgements We thank K. Vranizan for assistance with microarrayanalyses. K. Gerhold and D. Bautista provided the TRPV4 construct, protocols andadvice for HEK293 experiments; the Roelink laboratory provided tissue culturefacilities and advice. G. Agarwaal generated heat map images in Matlab for datapresentation. W. Fischler generated the NP1017 G-CaMP data in SupplementaryInformation. We are grateful to C. Zuker and members of the Scott laboratory forcomments on the manuscript. This work was supported by a grant from the NIH(NIDCD), a Burroughs-Wellcome CAREER Award and a John Merck Award to K.S.and a NIH predoctoral fellowship to P.C. K.S. is an HHMI Early Career Scientist.
Author Contributions P.C. performed most experiments and co-wrote themanuscript. M.H. performed the electrophysiological recordings and the HEK293heterologous experiments. J.N. provided expertise on the microarray experiments.K.S. co-wrote the manuscript and supervised the project.
Author Information Reprints and permissions information is available atwww.nature.com/reprints. The authors declare no competing financial interests.Correspondence and requests for materials should be addressed to K.S.([email protected]).
NATURE | Vol 465 | 6 May 2010 LETTERS
95Macmillan Publishers Limited. All rights reserved©2010

METHODSTransgenic flies and ppk28 mutants. The ppk28 promoter-Gal4 construct was
generated with a 1.004-kb upstream fragment (16699333–16700336, GenBank
accession number NC_004354.3). Full-length ppk28 (transcript variant a,
NM_132941) was subcloned into pUAST. ppk28 mutants were generated by
FLP-FRT-mediated recombination between piggyBac transposons f05788 and
e02329.
Immunohistochemistry and in situ hybridization. Labelling was performed as
described11. In Fig. 1b, the brain was counterstained with nc82 (ref. 25). In Fig. 1c,
d, CD2 and GFP were detected by immunohistochemistry on flies containingppk28-Gal4, UAS-CD2, Gr66a-GFP-IRES-GFP-IRES-GFP or Gr5a-GFP-IRES-
GFP-IRES-GFP transgenes11,12.
G-CaMP imaging experiments. Imaging studies were performed as described12.
Flies were aged ,2–5 weeks. For Fig. 1e, flies were UAS-G-CaMP; ppk28-Gal4;
UAS-G-CaMP. For NaCl, sucrose and ribose, flies were stimulated 2–3 times,
ending with a positive control (.8% DF/F). For NMDG (pH 7.4 with HCl),
and PEG (molecular mass 3,350 g mol21), concentrations were presented ran-
domly, ending with a positive control (.7% DF/F). For Fig. 2c, d, genotypes were
as follows. Control: UAS-G-CaMP;ppk28-Gal4;UAS-G-CaMP. Mutant: Dppk28,
UAS-G-CaMP;ppk28-Gal4;UAS-G-CaMP. Rescue:Dppk28, UAS-G-CaMP;ppk28-
Gal4;UAS-G-CaMP, UAS-ppk28. Compounds were presented randomly and
experiments were performed blind to genotype. For Fig. 3, genotypes were as
follows. Gr66a: UAS-G-CaMP;Gr66a-Gal4;TM2/TM6b. Gr66a 1 ppk28: UAS-G-
CaMP;Gr66a-Gal4;UAS-ppk28. Compounds were presented randomly followed
by 10 mM denatonium (.8% DF/F).
Behavioural assays. Control flies were isogenic w1118 fly strain (Exelixis strain
A5001, BL-6326) and transgenes were backcrossed seven times to the control
strain. Two-to-five-day-old flies were starved for ,18–24 h with water, kept in ahumid chamber for 2–3 h and then stimulated on the proboscis with a taste
substance. Flies were allowed to ingest freely until they did not ingest after five
consecutive stimulations. For water ingestion, flies were stimulated on the pro-
boscis with 1 M sucrose afterward and only responders were kept for data tally.
Electrophysiology. Electrophysiology was performed as described26. Two-to-
three-day-old flies were transferred on fresh medium 1 day before the experi-
ment. For recording activity from labellar taste neurons, a reference glass
electrode filled with AHL solution12 was placed in the proboscis base and a
recording electrode filled with testing taste solution covered the tip of a single
taste bristle. All test solutions contain 1 mM KCl as an electrolyte. The signal was
amplified (1003 total), filtered (low-pass: ,2,800 Hz) by amplifiers (DTP-2,
Syntech; CyberAmp 320, Molecular Devices) and stored on a PC. Action poten-
tials were counted for the first 1 s. For Fig. 3, only i-type sensilla were recorded, as
they contain the bitter cell but lack the water cell. Statistical analyses were done
using a two-tailed Student’s t-test or Kruskal–Wallis analysis of variance
(ANOVA) (for comparisons among more than two groups) unless otherwise
noted. Significant differences were analysed using Dunn’s multiple comparison
test as the post-hoc test (significance level 5 0.001).
HEK293 calcium-imaging experiments and immunohistochemistry. Measure-
ments in cells were made by using calcium indicator Fluo-4 (Invitrogen) and a
confocal laser scanning microscope (Zeiss LSM510, Carl Zeiss). Cells were seeded
on poly-D-lysine-coated glass 1 day before transfection (Lipofectamine 2000,
Invitrogen), and incubated for 24–48 h before imaging. Cells were then loaded
with 10mM Fluo-4 for 45 min at 37 uC in isotonic calcium-imaging buffer (76 mM
NaCl, 5 mM KCl, 2 mM MgCl2, 2 mM CaCl2, 10 mM glucose, 10 mM HEPES,
mannitol, pH 7.4) in dark conditions. Solutions of varying osmolalities (303, 236,
216 and 174 mmol kg21) were prepared by adjusting the mannitol concentration.
Osmolality of test solutions was measured using a vapour pressure osmometer
(Vapro 5520, Wescor Inc.).
Cells were set in a perfusion chamber with isotonic solution for 3 min before
stimulating with osmotic test solutions. Solution flow was kept constant at
3.3 ml min21. Fluorescence emission at 480 nm was filtered by a 505–530 band-
pass filter. Images were analysed using automated routines written in Matlab.
Total fluorescence change for the dsRed-positive cells in the field was calculated
and divided by cell area to normalize for cell density. Responses were averaged
from 3 to 5 independent experiments per stimulation per transfected cell line.
doi:10.1038/nature09011
Macmillan Publishers Limited. All rights reserved©2010





























