Embed Size (px)
Citation preview

Original Paper
The location of pKi67 in the outer dense®brillary compartment of the nucleolus pointsto a role in ribosome biogenesis during thecell division cycle
David E. MacCallum and Peter A. Hall*Department of Molecular and Cellular Pathology, Ninewells Hospital and Medical School, University of Dundee, Dundee DD1 9SY, UK
*Correspondence to:P. A. Hall, King Fahad NationalGuard Hospital, PO Box 22490,Riyadh, 11426, Kingdom ofSaudi Arabia.E-mail: [email protected]
Received: 7 May 1999
Revised: 27 September 1999
Accepted: 26 October 1999
Abstract
Although widely used as a marker of cell proliferation, the biochemical properties and function of
the Ki67 antigen remain poorly understood. Recent data indicate that it can interact with RNA,
DNA, and a number of cellular proteins including elements of the ubiquitin proteolytic pathway
and a novel kinase. The evidence for its expression only in cycling cells is extensive and it is not
regulated by stress, apoptosis or DNA damage. It was reasoned that a detailed characterization of
the localization of pKi67 and analysis of its spatial relationship to other nucleolar proteins may
provide insights into its function. Using high-resolution laser scanning confocal microscopy with
double and triple labelling, pKi67 expression in MCF7 cells has been de®ned in relation to the
distribution of nucleolin, ®brillarin, p130 (human Nopp 140 homologue), p120 (Nol 1), RH-II/Gu
helicase, and topoisomerase II b. All of these molecules are perichromosomal during mitosis and
all but ®brillarin and p130 show extra-nucleolar distribution in early G1. The majority of p120
(Nol 1) and RH-II/Gu helicase co-localize in the diffuse ®brillar centre (DFC) of nucleoli, while
there is only partial overlap with nucleolin and ®brillarin. There is no co-localization between
p130 and pKi67. These data re®ne current understanding of the distribution of pKi67 and its
physical relationship with functional domains of the nucleolus and place pKi67 in a zone of the
DFC associated with late rRNA processing. Taken together with recent biochemical data, these
observations allow the proposal of a model of pKi67 function in which it acts as an `ef®ciency
factor' in ribosome biogenesis during the heavy metabolic demands placed on a cell during the cell
division cycle. Copyright # 2000 John Wiley & Sons, Ltd.
Keywords: Ki67; nucleolus; nucleolin; ®brillarin; p130; p120; helicase; topoisomerase
Introduction
The use of antibodies to the Ki67 protein (pKi67) iswell established in investigative and diagnostic pathol-ogy, since the expression of the Ki67 protein is tightlylinked with cell proliferation [1±4]. Despite its wide-spread use and the cloning of the cDNA encodingKi67 protein [5,6], the function of this large nuclearprotein remains poorly understood. Indeed, biochem-ical and functional data are scarce. Few clues to thefunction of pKi67 come from analysis of its amino acidstructure. It is an enormous protein of y395 kD andcontains a forkhead-associated domain at the Nterminus, a motif seen in a subset of transcriptionfactors [7]. There is a large spliced exon (exon 7) whichhas recently been shown to associate with elements ofthe ubiquitin pathway and with a novel kinasehomologous with the AMP activated family of kinases[8] (and Ross and Hall, in preparation). There is a hugecentral exon (6.8 kB) encoding 16 unusual repeats withruns of both acid and basic residues. The function ofthese repeats is unknown but Ross and Hall haverecently demonstrated that they have the capacity tobind RNA [8] (and Ross and Hall, in preparation). The
amino acid sequence has many potential phosphoryla-tion sites and recent data indicate that profoundalterations in the phosphorylation of pKi67 occurduring the cell cycle [9]. Such changes correlate notonly with alterations in sub-cellular localization, butalso with the ability of pKi67 to interact with DNA [9],a property which is predominantly associated with theC terminus [10]. Other data have shown that pKi67 ispredominantly associated with the nucleolus in cyclingcells, although it associates with the perichromosomalsheath in mitosis, as do other nucleolar proteins.Despite these data, we remain profoundly ignorant ofthe role of pKi67 in cellular homeostasis.
Studies of the localization of pKi67 have beenpreviously reported in cultured cells. During early G1,pKi67 is found in the nucleolus as well as other sitesthat co-localize with telomeric and centromericregions; this pattern has been called type I [11,12]. Ascells progress through G1 into the S phase, pKi67localizes almost exclusively to the nucleolus, calledthe type II pattern. At mitosis, pKi67 localizes to theperichromosomal layer [1,13,14]. The nucleous is theexclusive site of rRNA transcription, processing andmodi®cation of this transcript, and the assembly of
Journal of PathologyJ Pathol 2000; 190: 537±544.
Copyright # 2000 John Wiley & Sons, Ltd.

mature rRNA with ribosomal proteins to formribosomal subunits. Electron microscopic studies(reviewed in ref. [15]) have de®ned three key regionsor domains in the nucleolus, namely the ®brillar centre(FC), the dense ®brillar component (DFC), and thegranular component (GC). These domains containdifferent pro®les of nucleolar proteins and appearspecialized for different aspects of ribosome biogenesis.More recent studies using riboprobes to differentregions of the pre-ribosomal RNA have demonstratedthat the nucleolus is not simply divided into threedomains as de®ned by electron microscopy, but is farmore complex [15,16]. Nevertheless, the basic morpho-logical compartments de®ne early (FC), intermediate(DFC), and late (GC) processing of nascent rRNAtranscripts and ribosome biogenesis. We reasoned thata detailed de®nition of the localization of pKi67 andanalysis of its spatial relationships with other nucleolarproteins might provide new clues as to the function ofthis widely used marker of proliferation. We used arange of different antibodies reported to localize to theelements of the nucleolus for co-localization studiesand have carefully de®ned the localization of pKi67with these other antigens in all phases of the cell cycle.Our data point to pKi67 being an element of the dense®brillar component; it is thus most likely to have a rolein the late stages of rRNA processing and to be anessential factor in the rapid production of ribosomesduring the cell division cycle.
Materials and methods
MCF7 cells were grown on 12 well slides until 80%con¯uent, washed twice in sterile phosphate bufferedsaline (PBS), ®xed in methanol±acetone (50 : 50, v/v)for 10 min, and allowed to air-dry. They were wrappedin foil and stored at x80uC until required. Slides wereallowed to thaw and were rehydrated in PBS andblocked for 1 h in 3% (w/v) BSA in PBS. PolyclonalKi67 antibody was incubated on the slides at 1 : 1000overnight at 4uC, followed by three washes in PBSfor 5 min each. The slides were then incubated with a1 : 50 dilution of FITC conjugated swine anti-rabbit(DAKO) in PBS for half an hour, washed as before,
and mounted using Mowiol either with or withoutpropidium iodide at 0.1 mg/ml. Co-staining was alsoperformed using the antibodies listed in Table 1. As anexample, co-staining for nucleolin and pKi67 wascarried out using a 1 : 250 dilution of the monoclonalantibody 7G2 (a kind gift of G. Dreyfuss) added at thesame time as the polyclonal Ki67 antibody (at 1 : 1000dilution). A 1 : 100 dilution of the rabbit anti-mouseTexas red secondary antibody (Vector Labs Inc.) wasincubated after the FITC secondary antibody had beenincubated and washed 3 times each for 5 min. Theseslides were mounted in the absence of propidiumiodide. For the human auto-immune sera, Texas red-conjugated goat anti-human sera was used at 1 : 100(Vector Labs Inc.) and applied second. Before use, itwas incubated for 30 min in 5% swine serum to preventnon-speci®c cross-reaction. Slides were mounted withan aqueous mountant containing 1.0 mg/ml DAPI.Images were collected using a confocal scanning lasermicroscope (CSLM) (Biorad MRC 600) viewed ateither 60r or 100r objective. For the DAPI co-localization, we used an image capture system (Espritversion 1.2, Life Science Resources Ltd.) attached to alight microscope (Olympus). Concentrations of anti-bodies were adjusted to give roughly similar intensitiesfor a given aperture. The intensities were ®ne-tuned bychanging the gain in each channel. Standard ®lter sets(Biorad MRC 600) were used and relevant single-stained control preparations examined to determine thecontribution of `cross-talk' between channels and levelsof background. By the use of these controls, theseproblems were nulli®ed. All images were assembledusing Adobe Photoshop version 4.0.
Results
The localization of pKi67 in the nucleus
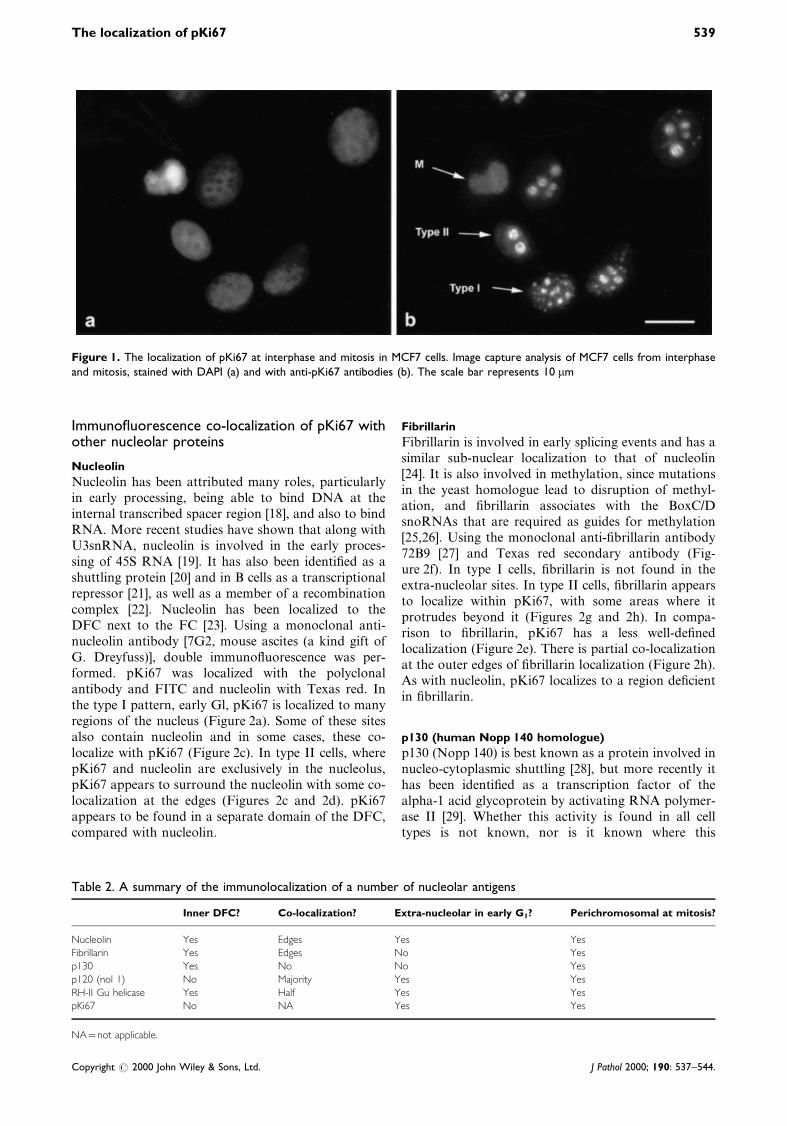
A number of previous studies have demonstrated thatKi67 protein is detectable only in the nucleus ofproliferating cells and that its spatial localizationdiffers in different phases of the cell cycle [1,13,14].An important consideration when performing theselocalization studies is the method of ®xation [17]. Weused both methanol±acetone and paraformaldehyde®xation methods, which yielded the same results. Onlythe ®gures for methanol±acetone ®xation are illu-strated. During early G1, pKi67 is found in the dense®brillar component (DFC) of the nucleolus as well asother sites that co-localize with telomeric and centro-meric regions of chromosomes; this pattern has beencalled type I [11,12]. As cells progress through G1into the S phase, pKi67 localizes almost exclusively tothe nucleolus, called the type II pattern. In MCF7cells, we were able to con®rm these observations andcould easily identify the three patterns of pKi67expression (Figure 1). This provides a basis for ourfurther studies, the results of which are summarized inTable 2.
Table 1. Summary of the primary antibodies used, thetype, and the source of the sera
Molecule (source) Concentration Nature of antibody
Polyclonal Ki67 (DAKO) 5 mg/ml Monoclonal
Nucleolin (G. Dreyfuss) 1 : 250 Concentrated monoclonalFibrillarin (U. Scheer) 1 : 250 Concentrated monoclonal
p130 (N.-H. Yeh) Neat Tissue culture supernatant
p120 (H. Busch) 1 : 10 Tissue culture supernatant
Rh-II/Gu helicase(H. Busch)
1 : 10 000 Human autoimmune sera
Topoisomerase IIb
(I. Hickson)
1 : 100 Concentrated monoclonal
538 D. E. MacCallum and P. A. Hall
Copyright # 2000 John Wiley & Sons, Ltd. J Pathol 2000; 190: 537±544.
Immuno¯uorescence co-localization of pKi67 withother nucleolar proteins
Nucleolin
Nucleolin has been attributed many roles, particularlyin early processing, being able to bind DNA at theinternal transcribed spacer region [18], and also to bindRNA. More recent studies have shown that along withU3snRNA, nucleolin is involved in the early proces-sing of 45S RNA [19]. It has also been identi®ed as ashuttling protein [20] and in B cells as a transcriptionalrepressor [21], as well as a member of a recombinationcomplex [22]. Nucleolin has been localized to theDFC next to the FC [23]. Using a monoclonal anti-nucleolin antibody [7G2, mouse ascites (a kind gift ofG. Dreyfuss)], double immuno¯uorescence was per-formed. pKi67 was localized with the polyclonalantibody and FITC and nucleolin with Texas red. Inthe type I pattern, early Gl, pKi67 is localized to manyregions of the nucleus (Figure 2a). Some of these sitesalso contain nucleolin and in some cases, these co-localize with pKi67 (Figure 2c). In type II cells, wherepKi67 and nucleolin are exclusively in the nucleolus,pKi67 appears to surround the nucleolin with some co-localization at the edges (Figures 2c and 2d). pKi67appears to be found in a separate domain of the DFC,compared with nucleolin.
Fibrillarin
Fibrillarin is involved in early splicing events and has asimilar sub-nuclear localization to that of nucleolin[24]. It is also involved in methylation, since mutationsin the yeast homologue lead to disruption of methyl-ation, and ®brillarin associates with the BoxC/DsnoRNAs that are required as guides for methylation[25,26]. Using the monoclonal anti-®brillarin antibody72B9 [27] and Texas red secondary antibody (Fig-ure 2f). In type I cells, ®brillarin is not found in theextra-nucleolar sites. In type II cells, ®brillarin appearsto localize within pKi67, with some areas where itprotrudes beyond it (Figures 2g and 2h). In compa-rison to ®brillarin, pKi67 has a less well-de®nedlocalization (Figure 2e). There is partial co-localizationat the outer edges of ®brillarin localization (Figure 2h).As with nucleolin, pKi67 localizes to a region de®cientin ®brillarin.
p130 (human Nopp 140 homologue)
p130 (Nopp 140) is best known as a protein involved innucleo-cytoplasmic shuttling [28], but more recently ithas been identi®ed as a transcription factor of thealpha-1 acid glycoprotein by activating RNA polymer-ase II [29]. Whether this activity is found in all celltypes is not known, nor is it known where this
Figure 1. The localization of pKi67 at interphase and mitosis in MCF7 cells. Image capture analysis of MCF7 cells from interphaseand mitosis, stained with DAPI (a) and with anti-pKi67 antibodies (b). The scale bar represents 10 mm
Table 2. A summary of the immunolocalization of a number of nucleolar antigens
Inner DFC? Co-localization? Extra-nucleolar in early G1? Perichromosomal at mitosis?
Nucleolin Yes Edges Yes YesFibrillarin Yes Edges No Yes
p130 Yes No No Yes
p120 (nol 1) No Majority Yes Yes
RH-II Gu helicase Yes Half Yes YespKi67 No NA Yes Yes
NA=not applicable.
The localization of pKi67 539
Copyright # 2000 John Wiley & Sons, Ltd. J Pathol 2000; 190: 537±544.
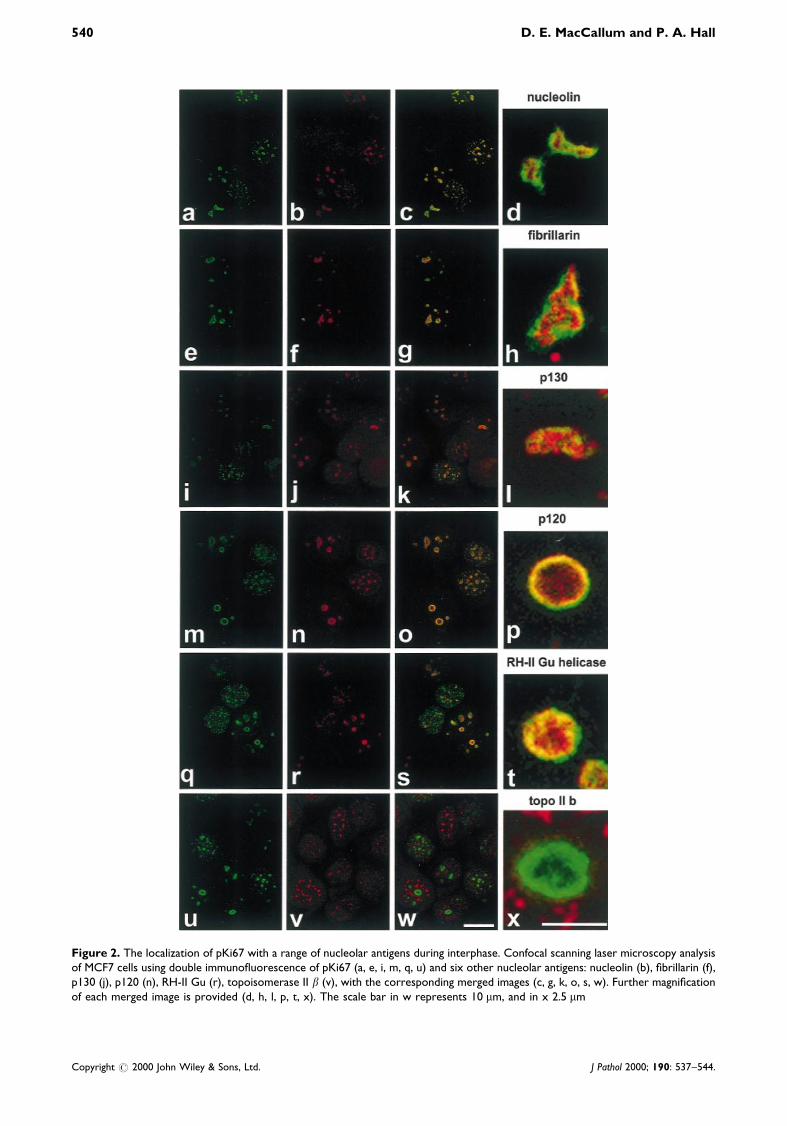
Figure 2. The localization of pKi67 with a range of nucleolar antigens during interphase. Confocal scanning laser microscopy analysisof MCF7 cells using double immuno¯uorescence of pKi67 (a, e, i, m, q, u) and six other nucleolar antigens: nucleolin (b), ®brillarin (f),p130 (j), p120 (n), RH-II Gu (r), topoisomerase II b (v), with the corresponding merged images (c, g, k, o, s, w). Further magni®cationof each merged image is provided (d, h, l, p, t, x). The scale bar in w represents 10 mm, and in x 2.5 mm
540 D. E. MacCallum and P. A. Hall
Copyright # 2000 John Wiley & Sons, Ltd. J Pathol 2000; 190: 537±544.
transcription takes place. p130 is unlikely to be atranscription factor. It is more probable that it plays arole in ribosomal synthesis and, due to its shuttlingcapabilities, may bring proteins into the nucleus forribosomal assembly. Its localization appears verysimilar to ®brillarin and nucleolin, which wouldsuggest that it plays a role in the initial processingevents. Using the CP1 monoclonal antibody, we havecharacterized the localization of p130 within thenucleolus (kind gift of N.-H. Yeh [30]). It localizesspeci®cally to the nucleolus in all stages of interphasein very discrete sites (Figure 2j). p130 sits within pKi67staining and only co-localizes at the outside edges(Figures 2k and 2l).
p120 [proliferation associated antigen (Nol 1)]
We also obtained an anti-p120 antibody whichrecognizes a protein whose yeast homologue(Nopp 2p) is required for methylation of 27S rRNAand ultimately for the processing of 27S RNA into 25SRNA [31]. Mutations in the yeast homologue causedisruption in rRNA processing, speci®cally by prevent-ing methylation [31]. Characterization of the humanp120 has identi®ed that it binds speci®cally to 28SrRNA in vitro via its arginine-rich domain. It alsoassociates with 60S and 80S pre-ribosomal particles.p120 (Nol 1) has previously been reported as a proteinof the DFC. Using a monoclonal antibody (kind gift ofH. Busch), we localized human p120 in relation topKi67. As with pKi67, p120 localized to a number ofextra-nucleolar sites in type I cells and is foundexclusively in the nucleolus of type II cells (Figure 2n).The merged image shows that the majority of p120 co-localizes with pKi67, although pKi67 also surrounds itin some cases (Figures 2o and 2p).
RH-II/Gu helicase
RH-II/Gu helicase is one of only two identi®ed humanDEAH box helicases that primarily associate with thenucleolus, the other being the Werner's syndromehelicase [32]. Initial localization studies performed onRH-II/Gu helicase de®ned it as being associated with
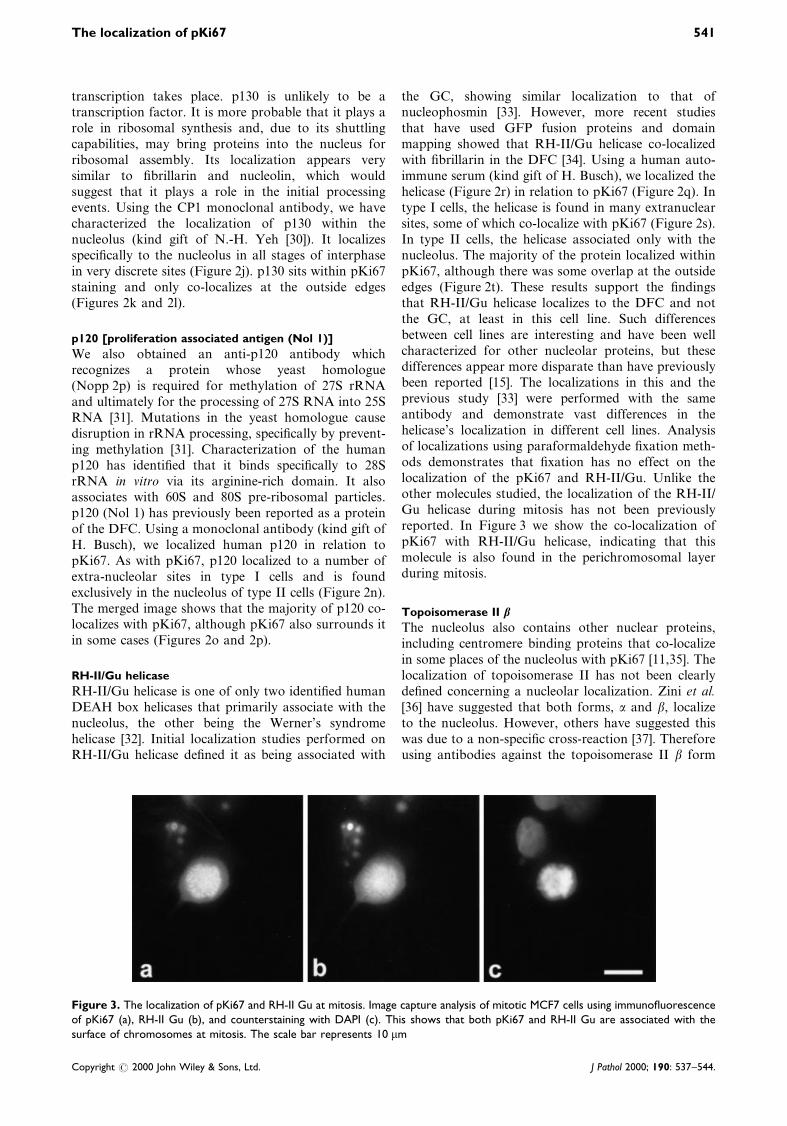
the GC, showing similar localization to that ofnucleophosmin [33]. However, more recent studiesthat have used GFP fusion proteins and domainmapping showed that RH-II/Gu helicase co-localizedwith ®brillarin in the DFC [34]. Using a human auto-immune serum (kind gift of H. Busch), we localized thehelicase (Figure 2r) in relation to pKi67 (Figure 2q). Intype I cells, the helicase is found in many extranuclearsites, some of which co-localize with pKi67 (Figure 2s).In type II cells, the helicase associated only with thenucleolus. The majority of the protein localized withinpKi67, although there was some overlap at the outsideedges (Figure 2t). These results support the ®ndingsthat RH-II/Gu helicase localizes to the DFC and notthe GC, at least in this cell line. Such differencesbetween cell lines are interesting and have been wellcharacterized for other nucleolar proteins, but thesedifferences appear more disparate than have previouslybeen reported [15]. The localizations in this and theprevious study [33] were performed with the sameantibody and demonstrate vast differences in thehelicase's localization in different cell lines. Analysisof localizations using paraformaldehyde ®xation meth-ods demonstrates that ®xation has no effect on thelocalization of the pKi67 and RH-II/Gu. Unlike theother molecules studied, the localization of the RH-II/Gu helicase during mitosis has not been previouslyreported. In Figure 3 we show the co-localization ofpKi67 with RH-II/Gu helicase, indicating that thismolecule is also found in the perichromosomal layerduring mitosis.
Topoisomerase II b
The nucleolus also contains other nuclear proteins,including centromere binding proteins that co-localizein some places of the nucleolus with pKi67 [11,35]. Thelocalization of topoisomerase II has not been clearlyde®ned concerning a nucleolar localization. Zini et al.[36] have suggested that both forms, a and b, localizeto the nucleolus. However, others have suggested thiswas due to a non-speci®c cross-reaction [37]. Thereforeusing antibodies against the topoisomerase II b form
Figure 3. The localization of pKi67 and RH-II Gu at mitosis. Image capture analysis of mitotic MCF7 cells using immuno¯uorescenceof pKi67 (a), RH-II Gu (b), and counterstaining with DAPI (c). This shows that both pKi67 and RH-II Gu are associated with thesurface of chromosomes at mitosis. The scale bar represents 10 mm
The localization of pKi67 541
Copyright # 2000 John Wiley & Sons, Ltd. J Pathol 2000; 190: 537±544.

(kind gift of I. Hickson), its localization was de®ned inthe context of pKi67. Topoisomerase II b localizes tonucleoplasmic dots, which are distributed throughoutthe nucleus (Figure 2v). None of these sites co-localizeswith pKi67 nor appears nucleolar (Figure 2w); parti-cularly not in the DFC as previously described, as itneither co-localizes nor sits within pKi67 localization(Figures 2w and 2x).
Discussion
Despite its extensive use in pathology and cell biologyas a marker of proliferation, we remain ignorant of thefunction of the Ki67 protein. The purpose of thestudies reported here was to characterize the detailedlocalization of pKi67 in relation to other nuclearproteins. Such an analysis is of particular importancein the case of the nucleolar proteins, since thisstructure contains many compartments with complexfunctional sub-divisions. Electron microscopy de®nesthe FC, DFC, and GC, but studies using riboprobeshave shown a more complex picture [16]. Our studiescon®rm this concept, since the nucleolus has quitedistinct antigen pro®les and hence a complex anatomy.In summary (Table 2), we have localized ®ve proteinsof the DFC in relation to pKi67, which also occupiesthis region. Each protein is localized to its own uniquesite, albeit with some overlap. We have also showndifferences between the various nucleolar proteins interms of their distribution in mitotic cells.
The model for pre-RNA processing is that it occursin a radial manner, with early processing occurring atthe inside edge of the DFC and later processing stepsoccurring further out [38]. Co-localization studies withproteins that have been functionally de®ned shouldtherefore help to place pKi67 in amongst these path-ways. Our studies show that ®brillarin, nucleolin, andp130 have a very distinct granular localization that isfound within pKi67 localization in the nucleolus; insome circumstances there are protrusions outside it,but mainly they sit within, con®rming their localizationin the DFC close to the FC. These data suggest thatpKi67 is probably involved in a later processing stepthan ®brillarin, nucleolin, and p130. The localizationof ®brillarin in our studies is consistent with the resultsof Kill [11] and in accordance with this, pKi67 localizesto a ®brillarin-de®cient region of the DFC. We haveextended this by demonstrating that it sits in anucleolin, p130, and ®brillarin-de®cient region of theDFC in MCF7 cells.
The localization of pKi67 to the outer DFC of thenucleolus, with partial co-localization with p120 andRH-II/Gu helicase, suggests a role not in earlyprocessing as de®ned by ®brillarin and nucleolin, norin later processes characterized by nucleophosminlocalization [11]. Its absence from non-proliferatingcells suggests a non-essential function in ribosomalassembly, as it is only required when demand forribosomes is high, such as during replication. The co-
localization of pKi67 with p120 suggests an involve-ment in similar pathways of 28S rRNA maturation.pKi67 may therefore play a role in ef®ciency, increas-ing output in some way, possibly by improving theef®ciency of pre-rRNA processing, which is thought tobe partly responsible for increasing ribosome output[39]. Nucleolin, p120, RH-II/Gu helicase, and pKi67are all found in extra-nucleolar sites in early G1,known as type I cells. Nucleophosmin has also beenreported to localize to these sites [11]. Studies usingDNA probes have demonstrated that these sitescontain centromeric and telomeric satellite DNA [12].The reason for this localization is unclear, although arole in organizing DNA has been proposed [12].
During mitosis, the nucleolus breaks down into itscomponents, which relocate to different regions aroundthe cell. One group of proteins localizes to theperichromosomal layer [40,41]. This list includespKi67, nucleolin, ®brillarin, p120 and p130; we showthat RH-II/Gu helicase is also found in this group. Thesigni®cance of this association is still not known and anumber of possibilities have been put forward [40]. Acompelling (but unproven) model is that the perichro-mosomal localization allows the passive but equaldistribution of these key proteins to the daughter cells.
In conclusion, we have successfully characterizedpKi67 in relation to ®ve further proteins of the DFC,namely nucleolin, ®brillarin, p120, p130 and RH-II/Guhelicase, and to a nuclear protein, topoisomerase II b.We have shown that pKi67 localizes to a speci®cregion of the dense ®brillar component that is de®cientin nucleolin, ®brillarin, and p130, but appears partiallyassociated with the helicase and almost completelyassociated with p120. How do these observationshelp our understanding of Ki67 function? A widelyheld model of nucleolar function infers a centripetalorganization of rRNA transcription, processing, andassembly with ribosomal proteins to allow ribosomebiogenesis. This complex process involves the co-ordinate interaction of many proteins, RNA speciesas well as DNA encoding the ribosome gene clusters.The ef®ciency with which these processes occur willdirectly in¯uence the metabolic and biosyntheticactivity of a cell. Our data place pKi67 in a zone ofthe DFC associated with late rRNA processing. Ourother data [8±10,42] indicate that pKi67 is involved incomplex protein±protein interactions, as well as beingable to bind RNA and DNA species. Taken together,these observations might suggest that the enormousKi67 protein, with its multiple domains of distinctfunction, has some architectural or structural rolewithin the nucleolus. Clearly, cells can be very activeand can synthesize ribosomes without Ki67, sincemany bio-synthetically active cells, such as hepatocytesand pancreatic acinar cells, contain undetectable levelsof Ki67. It is only in cycling cells that Ki67 is expressed± cells which require to be bio-synthetically very activeover short periods and in which the number ofribosomes must be doubled very rapidly, to allow thecell growth events that must precede cytokinesis. We
542 D. E. MacCallum and P. A. Hall
Copyright # 2000 John Wiley & Sons, Ltd. J Pathol 2000; 190: 537±544.

would then postulate that pKi67 acts as an ef®ciencyfactor, enhancing aspects of rRNA metabolism andthus facilitating rapid cell growth prior to cell division.Indirect con®rmation of this model comes from therecent observations of Plaat et al. [43], who show aclose correlation between the rate of protein synthesisand Ki67 expression. The observations reported here,together with our recent biochemical studies [8±10,42],now put us in a position to test this model formally.
Acknowledgements
We thank I. Hickson, G. Dreyfuss, U. Scheer, N.-H. Yeh, and
H. Busch for the gift of antibodies and serum; Frances Fuller-
Pace and David Meek for assistance with microscopy and image
capture; and members of the Hall laboratory, past and present,
for useful discussions. DEM was in receipt of a CRC student-
ship. The Hall laboratory was supported by the AICR, the EU,
the Pathological Society of Great Britain and Ireland, the CRC,
and the Department of Health.
References
1. Gerdes J, Lemke H, Baisch H, Wacker HH, Schwab U, Stein H.
Cell cycle analysis of a cell proliferation-associated human
nuclear antigen de®ned by the monoclonal antibody Ki-67.
J Immunol 1984; 133: 1710±1715.
2. Hall PA, Levison DA. Assessment of cell proliferation in
histological material. J Clin Pathol 1990; 43: 184±192.
3. Scott RJ, Hall PA, Haldane JS, et al. A comparison of
immunohistochemical markers of cell proliferation with experi-
mentally determined growth fraction. J Pathol 1991; 165:
173±178.
4. Ross W, Hall PA. Ki67: from antibody to molecule to
understanding? Clin Mol Pathol 1995; 48: 113±117.
5. Gerdes J, Li L, Schlueter C, et al. Immunobiochemical and
molecular biologic characterization of the cell proliferation-
associated nuclear antigen that is de®ned by monoclonal
antibody Ki-67. Am J Pathol 1991; 138: 867±873.
6. Schluter C, Duchrow M, Wohlenberg C, et al. The cell
proliferation-associated antigen of antibody Ki-67: a very large,
ubiquitous nuclear protein with numerous repeated elements,
representing a new kind of cell cycle-maintaining protein. J Cell
Biol 1993; 123: 513±522.
7. Hofmann K, Bucher P. The FHA domain: a putative nuclear
signalling domain found in protein kinases and transcription
factors. Trends Biochem Sci 1995; 20: 347±349.
8. Ross W. Studies of the human Ki67 protein. PhD Thesis,
University of Dundee, 1998.
9. MacCallum DE, Hall PA. Biochemical characterisation of pKi67
with the identi®cation of a mitotic speci®c form associated with
hyper-phosphorylation and altered DNA binding. Exp Cell Res
1999; 252: 186±198.
10. MacCallum DE, Hall PA. The biochemical characterisation of
the DNA binding activity of pKi67. J Pathol (in press).
11. Kill IR. Localization of the Ki-67 antigen within the nucleolus.
Evidence for a ®brillarin-de®cient region of the dense ®brillar
component. J Cell Sci 1996; 109: 1253±1263.
12. Bridger JM, Kill IR, Lichter P. Association of pKi-67 with
satellite DNA of the human genome in early G1 cells.
Chromosome Res 1998; 6: 13±24.
13. Verheijen R, Kuijpers HJ, Schlingemann RO, et al. Ki-67 detects
a nuclear matrix-associated proliferation-related antigen. I.
Intracellular localization during interphase. J Cell Sci 1989; 92:
123±130.
14. Verheijen R, Kuijpers HJ, van Driel R, et al. Ki-67 detects a
nuclear matrix-associated proliferation-related antigen. II. Loca-
lization in mitotic cells and association with chromosomes. J Cell
Sci 1989; 92: 531±540.
15. Thiry M, Goessens G. The Nucleolus During the Cell Cycle (1st
edn). Chapman & Hall: Austin, TX, 1996.
16. Lazdins IB, Delannoy M, Sollner-Webb B. Analysis of nucleolar
transcription and processing domains and pre-rRNA movements
by in situ hybridization. Chromosoma 1997; 105: 481±495.
17. Melan MA, Sluder G. Redistribution and differential extraction
of soluble proteins in permeabilized cultured cells. Implications
for immuno¯uorescence microscopy. J Cell Sci 1992; 101:
731±743.
18. Olson MOJ, Rivers ZM, Thompson BA, Kao W-Y, Case ST.
Interaction of nucleolar phosphoprotein C23 with cloned
fragments of ribosomal deoxyribonucleic acid. Biochemistry
1983; 22: 3345±3351.
19. Ginisty H, Amalric F, Bouvet P. Nucleolin functions on the ®rst
step of rRNA processing. EMBO J 1998; 17: 1476±1486.
20. Borer RA, Lehner CF, Eppenberger HM, Nigg EA. Major
nucleolar proteins shuttle between nucleus and cytoplasm. Cell
1989; 56: 379±390.
21. Yang TH, Tsai WH, Lee YM, et al. Puri®cation and character-
ization of nucleolin and its identi®cation as a transcription
repressor. Mol Cell Biol 1994; 14: 6068±6074.
22. Borggrefe T, Wabl M, Akhmedov, Jessberger R. A B-cell-speci®c
DNA recombination complex. J Biol Chem 1998; 273: 17025±
17035.
23. Spector DL, Ochs RL, Busch H. Silver staining, immuno¯uor-
escence, and immunoelectron microscopic localization of nucleo-
lar phosphoproteins B23 and C23. Chromosoma 1984; 90: 139±
148.
24. Eichler DC, Craig N. Processing of eukaryotic ribosomal RNA.
In Progress in Nucleic Acid Research and Molecular Biology,
Vol. 49. Academic Press: New York, 1994; 197.
25. Tollervey D, Lehtonen H, Jansen R, Kern H, Hurt EC.
Temperature-sensitive mutations demonstrate roles for yeast
®brillarin in pre-rRNA processing, pre-rRNA methylation, and
ribosome assembly. Cell 1993; 72: 443±457.
26. Kiss-Laszlo Z, Henry Y, Bachellerie JP, Caizergues-Ferrer M,
Kiss T. Site-speci®c ribose methylation of preribosomal RNA:
a novel function for small nucleolar RNAs. Cell 1996; 85:
1077±1088.
27. Reimer G, Pollard KM, Penning CA, et al. Monoclonal
autoantibody from a (New Zealand blackrNew Zealand white)
F1 mouse and some human scleroderma sera target an Mr
34,000 nucleolar protein of the U3 RNP particle. Arthritis
Rheum 1987; 30: 793±800.
28. Meier U, Blobel G. Nopp 140 shuttles on tracks between the
nucleus and cytoplasm. Cell 1992; 70: 127±138.
29. Miau LH, Chang CJ, Tsai WH, Lee SC. Identi®cation and
characterization of a nucleolar phosphoprotein, Nopp 140, as a
transcription factor. Mol Cell Biol 1997; 17: 230±239.
30. Pai CY, Chen HK, Sheu HL, Yeh NH. Cell-cycle-dependent
alterations of a highly phosphorylated nucleolar protein p130 are
associated with nucleologenesis. J Cell Sci 1995; 108: 1911±1920.
31. Hong B, Brockenbrough JS, Wu P, Aris JP. Nop2p is required
for pre-rRNA processing and 60S ribosome subunit synthesis in
yeast. Mol Cell Biol 1997; 17: 378±388.
32. Marciniak RA, Lombard DB, Johnson FB, Guarente L.
Nucleolar localization of the Werner syndrome protein in
human cells. Proc Natl Acad Sci U S A 1998; 95: 6887±6892.
33. Valdez BC, Henning D, Busch RK, et al. A nucleolar RNA
helicase recognized by autoimmune antibodies from a patient
with watermelon stomach disease. Nucleic Acids Res 1996; 24:
1220±1224.
34. Ou Y, Fritzler MJ, Valdez BC, Rattner JB. Mapping and
characterization of the functional domains of the nucleolar
protein RNA helicase II/Gu. Exp Cell Res 1999; 247: 389±398.
35. Ochs RL, Press RI. Centromere autoantigens are associated with
the nucleolus. Exp Cell Res 1992; 200: 339±350.
36. Zini N, Santi S, Ognibene A, et al. Discrete localization of
different DNA topoisomerases in HeLa and K562 cell nuclei and
subnuclear fractions. Exp Cell Res 1994; 210: 336±348.
The localization of pKi67 543
Copyright # 2000 John Wiley & Sons, Ltd. J Pathol 2000; 190: 537±544.

37. Meyer KN, Kjeldsen E, Straub T, et al. Cell cycle-coupled
relocation of types I and II topoisomerases and modulation of
catalytic enzyme activities. J Cell Biol 1997; 136: 775±788.
38. Olson MOJ. The role of proteins in nucleolar structure and
function. In The Eukaryotic Nucleus, Strauss PR (ed.). Telford
Press: Caldwell, 1991.
39. Dudov KP, Dabeva MD. Post-transcriptional regulation of
ribosome formation in the nucleus of regenerating rat liver.
Biochem J 1983; 210: 183±192.
40. Hernandez-Verdun D, Gautier T. The chromosome periphery
during mitosis. Bioessays 1994; 16: 179±185.
41. Medina FJ, Cerdido A, Fernandez-Gomez ME. Components of
the nucleolar processing complex (pre-rRNA, ®brillarin, and
nucleolin) colocalize during mitosis and are incorporated to
daughter cell nucleoli. Exp Cell Res 1995; 221: 111±125.
42. MacCallum DE. The biochemical characterisation of the Ki67
protein. PhD Thesis, University of Dundee, 1999.
43. Plaat B, Kole A, Mastik M, Hoekstra H, Molenaar W, Vaalburg
W. Protein synthesis rate measured with 1-[1±11C] tyrosine
positron emission tomography correlates with mitotic activity
and MIB-1 antibody-detected proliferation in human soft tissue
sarcomas. Eur J Nucl Med 1999; 26: 328±332.
544 D. E. MacCallum and P. A. Hall
Copyright # 2000 John Wiley & Sons, Ltd. J Pathol 2000; 190: 537±544.