-
Review
Biogenesis and origin of thylakoid membranes
Ute C. Vothknecht a;*, Peter Westho b
a Botanisches Institut der Christian-Albrechts-Universitat Kiel,
Am Botanischen Garten 1^9, D-24118 Kiel, Germanyb Institut fur
Entwicklungs- und Molekularbiologie der Panzen,
Heinrich-Heine-Universitat Dusseldorf, Universitatsstrasse 1,
D-40225 Dusseldorf, Germany
Received 25 July 2001; accepted 1 August 2001
Abstract
Thylakoids are photosynthetically active membranes found in
Cyanobacteria and chloroplasts. It is likely that theyoriginated in
photosynthetic bacteria, probably in close connection to the
occurrence of photosystem II and oxygenicphotosynthesis. In higher
plants, chloroplasts develop from undifferentiated proplastids.
These contain very few internalmembranes and the whole thylakoid
membrane system is built when chloroplast differentiation takes
place. During cell andorganelle division a constant synthesis of
new thylakoid membrane material is required. Also, rapid adaptation
to changes inlight conditions and long term adaptation to a number
of environmental factors are accomplished by changes in the lipid
andprotein content of the thylakoids. Thus regulation of synthesis
and assembly of all these elements is required to ensureoptimal
function of these membranes. 2001 Elsevier Science B.V. All rights
reserved.
Keywords: Chloroplast ; Photosynthesis ; Plastid development;
Organelle evolution; Thylakoid formation
1. Introduction
Evolution of oxygenic photosynthesis with its CO2xation and
oxygen release enabled life on earth aswe experience it today. It
is assumed that oxygenicphotosynthesis developed several billion
years ago inan ancestor of todays cyanobacteria most likelyfrom an
already existing anoxygenic photosynthesisapparatus [1]. The
capacity to perform oxygenic pho-tosynthesis was passed on to algae
and higher plantsby an endosymbiotic event that turned a
cyanobac-terium into a cell organelle, the chloroplast. The
pho-tosynthetic machinery of both, cyanobacteria and
chloroplasts, is located on a special internal mem-brane system,
the thylakoids. Thanks to the uniquearchitecture of this membrane
cyanobacteria andchloroplasts convert solar energy into chemical
en-ergy with an eciency signicantly better than anyman-made
photovoltaic system. Therefore, the abil-ity of the cell to build
and alter this membrane sys-tem is essential for ecient oxygenic
photosynthesis.Resulting from the combination of structural,
bio-chemical, and genetic analysis, we have a wellfounded knowledge
of the ultrastructure and compo-sition of thylakoid membranes, but
despite the im-portance that the thylakoid membrane system hasfor
photosynthesis and the energy metabolism ofplants and
cyanobacteria, the molecular processesconnected to the origin,
synthesis, maintenance andadaptation of the thylakoids remain
elusive. In thisreview we will discuss recent ndings on
thylakoid
0167-4889 / 01 / $ ^ see front matter 2001 Elsevier Science B.V.
All rights reserved.PII: S 0 1 6 7 - 4 8 8 9 ( 0 1 ) 0 0 1 5 3 -
7
* Corresponding author. Fax: +49-431-880-4222.E-mail address:
[email protected] (U.C.
Vothknecht).
BBAMCR 14806 11-12-01
Biochimica et Biophysica Acta 1541 (2001)
91^101www.bba-direct.com
-
biogenesis and evolution and their impact on ourunderstanding.
Since most studies concerning thebiogenesis of thylakoids have been
performed onchloroplasts of higher plants and green algae,
thisreview will focus on these organisms. The last sectionwill deal
with internal membrane systems in bacteria,especially the
thylakoids of cyanobacteria, and theevolution of the thylakoid
membrane in these organ-isms.
2. Form and function in plastids
In 1848 chloroplasts were rst described by Ungersimply as
pigment bound structures. Later in the19th century Schimper [2,3]
characterized these struc-tures, which he called Chlorophyllkorner,
in greaterdetail. With only the resolution of the light micro-scope
available, he described plastids as cell compo-nents containing
chlorophyll or other pigmentswhich develop from colorless
precursors. He alreadyperceived the existence of various types of
plastidsand made the observation that they can pass throughdierent
stages during their development. Shortlyafter the invention of the
electron microscope, therst electron micrographs of chloroplasts
were pub-lished [4], and soon thereafter, this new techniquewas
used for the rst detailed studies of dierentplastid forms and their
development. In the late 50sthe basic structure of thylakoids had
been described[5^7] and the means of thylakoid biogenesis
werediscussed.
Thylakoids are the dominating structure insidefully mature
chloroplasts. The formation and alter-ation of the thylakoid
membrane structure and com-position are closely connected to the
development ofthe chloroplasts from simple, undierentiated
pro-plastids. These are small round shaped organelleshardly
distinguishable from mitochondria (Fig. 1),that contain very few
internal membranes that areoften found as vesicles or small
saccular structures[7^9]. Occasionally these membranes are
observedcontinuous with the inner envelope.
In the presence of light proplastids develop intomature
chloroplasts. This transition has been inten-sively studied in
grasses. The leaves of these mono-cotyledonous plants grow with a
basal meristem andhence form a developmental gradient. Cells found
at
the base of the leaf are youngest and contain mainlyproplastids
while the oldest cells with fully developedchloroplasts are found
close to the tip. Ultra-thinsections revealed that during the
progress of chloro-plast maturation the internal membrane
systembuilds up in consecutive phases. First, long lamellaare
formed which are later complemented by smaller,disc-shaped
structures that associate into so-calledgrana stacks (Fig. 1). At
the same time the typicallens-shape form of the chloroplasts
develops. Finally,mature plant chloroplasts contain a complex and
in-tertwined internal membrane system which wasnamed thylakoids
according to the Greek word3eVKUYASNRf (sack-like) [10]. In fully
mature chlo-roplast no continuation between the inner envelopeand
the thylakoids has been observed.
In the absence of light proplastids turn into etio-plasts which
contain very few internal membranesbut a characteristic prolamellar
body [11,12]. Theprolamellar body is a paracrystalline structure
con-sisting of lipids and essentially a single protein,
theNADPH-dependent protochlorophyllide oxidoreduc-tase [13,14].
Shortly after the onset of illuminationthe prolamellar body is
dispersed and thylakoids be-gin to form [7,12,15]. Since the start
of illuminationcan easily be controlled in experimental setups,
thissystem has often been used to study chloroplast de-velopment.
Prolamellar bodies are mainly consideredin connection with
etioplasts but they are not re-stricted to them. Already after a
short period ofdarkness secondary prolamellar bodies form
insidefully matured chloroplasts [16,17]. This results in
thecoexistence of prolamellar bodies and thylakoids andraises
questions about the function of the prolamellarbody for the mature
chloroplasts.
Proplastids can further develop into chromoplastsor leucoplasts.
These are specialized forms of plastidsused for coloration or
storage [18]. Chromoplasts arecarotenoid-containing plastids found
in many owerpetals, fruits and roots. Coloration of these organs
isoften ascribed to chromoplasts and this might evenbe their main
function. Leucoplasts are characterizedby a lack of coloration and
they can be distinguishedby the substance that is stored, i.e.
amyloplasts, pro-teoplasts or elaioplasts. The nal stage of a
plastidslife is the senescent or gerontoplast. These are plas-tids
that have reached a stage of senescence that isnot reversible. All
plastids, independent of their sta-
BBAMCR 14806 11-12-01
U.C. Vothknecht, P. Westho / Biochimica et Biophysica Acta 1541
(2001) 91^10192
-
tus, retain the ability to develop into each other (Fig.1).
Interconversion of dierent plastid forms requiresdramatic changes
of the ultrastructure, including thebiogenesis, reorganization and
regression of internalmembranes.
3. Structure and composition of thethylakoid membrane
The additional compartment that the thylakoidnetwork creates in
cyanobacterial cells and in chlo-roplasts is an important feature
that distinguishesthese from bacteria performing anoxygenic
photo-synthesis. In these latter organisms, the internal
membranes are invaginations still continuous withthe plasma
membrane [19,20]. In mature chloroplastsand in cyanobacteria it is
assumed that the thyla-koids are no longer connected to the inner
envelopeor the plasma membrane, respectively, because nosuch
continuum can be observed in electron micro-scopic pictures.
How is this unique structural compositionachieved? In
cyanobacteria and many algae, thyla-koids consist mainly of single
layers formed bylong lamellae. The structure of the thylakoid
mem-brane in a fully mature chloroplast is more complex(Fig. 1).
Initiated by earlier electron microscopicstudies a model for the
thylakoid structure as ahuge intertwined network of stroma lamellae
con-
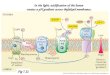
Fig. 1. Overview of the development of chloroplasts. Chloroplast
develop from undierentiated proplastids. During maturation
thecomplex internal thylakoid membrane network is formed.
Proplastids can also develop into other plastids forms, such as
etioplasts,chromoplasts and leucoplasts. Moreover, fully
dierentiated plastids retain the ability to develop into each
other. Gerontoplasts are anal stage in plastid development in which
a level of senescence is reached that is irreversible. The electron
microscopic pictures ofthin sections show several stages in the
development of a chloroplast.
BBAMCR 14806 11-12-01
U.C. Vothknecht, P. Westho / Biochimica et Biophysica Acta 1541
(2001) 91^101 93
-
necting grana stacks was proposed which with littlealterations
is still valid today [21,22]. One can distin-guish two major parts,
the grana and the stromalamellae. Grana are short, disc-shaped
lamellaeclosely packed to form stacks. These stacks are
in-terconnected by stroma lamellae which also formprolonged
extensions into the stroma. Thus, the ar-rangement of the thylakoid
membrane system createsa single huge compartment inside the
chloroplast, thethylakoid lumen. Additionally to creating a
singleinternal space this structure builds a membrane sur-face that
is much larger than a simple invagination ofthe inner envelope
would generate.
To understand the complexity of the task that theformation of
thylakoids presents to the cell, it isimportant to gure the
components that are requiredto build up this special photosynthetic
membrane.Thylakoids are lipid bilayers with a unique glycero-lipid
composition dierent from other cell mem-branes. Thylakoid lipids
have a high content, about70^80%, of galactosyl diglycerides and
both mono-galactosyl diacylglycerol and digalactosyl
diacylgly-cerol are lipids nearly exclusively found in
plastidalmembranes [23]. Notably, these galactolipids containtwo
highly unsaturated fatty acyl chains instead ofone as is common in
membrane lipids and are bothnon-bilayer forming lipids.
Additionally the thyla-koids contain phosphatidylglycerol and
sulfoquino-vosyl diacylglycerol together with other minor
com-ponents [23]. All these lipids are not evenlydistributed along
the thylakoid membrane. Insteadthe lipid distribution diers between
the leaet thatis exposed to the stroma and the inner leaet
thatfaces the thylakoid lumen [23]. It is not clear howthis
asymmetrical arrangement of the lipid distribu-tion is achieved.
Yet it has to be assumed that it isimportant for the function of
the thylakoid mem-brane.
The dominant protein complexes of the thylakoidsare photosystems
I [24] and II [25] and their associ-ated light harvesting antenna,
the cytochrome b6fcomplex [26] and the proton-translocating ATP
syn-thase [27]. These complexes comprise not only manyperipheral
and integral proteins but also a variety ofpigments and co-factors
[28]. Their assembly is,therefore, a complex process and requires a
largernumber of auxiliary and regulatory factors [28,29].These
factors are involved in the membrane integra-
tion, modication and later degradation of the pro-teinaceous
components and are also required for theaddition of the pigments
and co-factors. To compli-cate matters, certain components, like
the two photo-systems, are unevenly distributed in the
thylakoidmembrane network. While photosystem I is mostabundant in
the non-stacked stroma lamellae, photo-system II is the dominating
component of the granastacks [30]. Thus thylakoid biogenesis and
mainte-nance have to assure not only the arrangement of afunctional
but at the same time asymmetric architec-ture of both the lipid and
the protein components ofthis membrane.
4. Thylakoid membrane formation
One of the most elusive aspects of thylakoid for-mation is the
exact mechanism by which the mem-brane itself is formed. In young,
not yet dierenti-ated plastids a continuum can sometimes be
observedbetween the inner envelope and the developing inter-nal
membrane structures [7^9]. Thus the synthesis ofearly thylakoid
membranes might be achieved by in-vagination of the inner envelope.
Even in fully ma-ture chloroplasts the thylakoid membrane is a
verydynamic system. Short-term adaptation to changinglight
conditions is obtained by movement of proteins,especially the light
harvesting complex, within thethylakoid membrane. Long-term
adaptation on theother hand is achieved by a change in the
proteinand lipid content of the thylakoids. Although in ma-ture
chloroplasts a continuum between the inner en-velope and the
thylakoids can no longer be observed,the membrane material required
for synthesis andmaintenance of the thylakoids originates from
thechloroplasts inner envelope [7,31,32] and not fromde novo
synthesis on already existing thylakoids.
How these lipids are transferred from the innerenvelope to the
thylakoids is controversially dis-cussed. One possibility would be
the transfer byvesicles which is a common phenomenon in the
cy-tosol, where vesicle trac is involved in many dier-ent cellular
processes including the secretory path-way, endocytosis, neural
transmission and vacuoleformation [33]. A similar vesicle transfer
fromthe inner envelope to the thylakoids has been impli-cated in
the synthesis of thylakoid membranes
BBAMCR 14806 11-12-01
U.C. Vothknecht, P. Westho / Biochimica et Biophysica Acta 1541
(2001) 91^10194
-
[7,31,32,34,35]. Vesicles inside plastids have been ob-served in
early electron microscopic studies [7^9].They are common in
proplastids and have alsobeen observed on the inner envelope of
etioplastsin dark-grown cells of the Chlamydomonas y-1 mu-tant,
shortly after illumination when chloroplast de-velopment sets in
[36]. On the other hand vesicles arevery rarely detected in mature
chloroplasts. They doaccumulate in the stromal space between the
innerenvelope and the thylakoids after a low temperatureincubation
of leaf tissue [34,35]. A similar phenom-enon is described for
vesicle transport in animal cells,i.e. enodplasmic reticulum to
Golgi and Golgi toplasma membrane, where low temperature blocksthe
fusion of vesicles with their target membrane[37]. Further
indication for vesicle transfer in chlo-roplasts comes from mutant
analysis. In several plantmutants that are aected in thylakoid
biogenesis, anaccumulation of vesicles can be observed. Others,like
the vipp1 mutant of Arabidopsis, no longer ex-hibited low
temperature vesicle accumulation [35].
The possibility of vesicle transfer inside the chlo-roplast
raised the additional question whether solelymembrane lipids would
be transported by thesevesicles. As in vacuole formation the
vesicle trans-port in chloroplasts could be limited to the supplyof
thylakoid lipids that are either synthesized at theinner envelope,
i.e. galactolipids, or imported fromthe cytosol. It is also
possible that non-lipid compo-nents of the thylakoid membrane might
be trans-ported by means of vesicle trac [38,39]. Several ofthe
non-lipid components required for the biogenesisand maintenance of
thylakoids are synthesized onthe envelope, i.e. carotenoids, or in
the cytosol[40,41]. Especially hydrophobic components wouldrequire
a system to travel through the aqueous stro-ma.
During chloroplast maturation an extensive for-mation of
thylakoid membranes occurs in concertwith the accumulation of the
photosynthetic com-plexes. Several of the proteinaceous components
arenuclear encoded and post-translationally importedinto the
chloroplasts. It was suggested that in Chla-mydomonas the nuclear
encoded light harvestingcomplex proteins are inserted into newly
developingmembranes at the inner envelope immediately upontheir
entrance in the organelle [42]. Later on, thedevelopment of the
thylakoid system continues with
the formation of grana stacking. Again, integrationof the light
harvesting complex into the thylakoidmembrane might play an
important role in this struc-tural reconstruction [43]. This early
speculation wassupported recently by Simidjiev and coworkers,
whoshowed that delipidated light harvesting complexeswould
restructure into ordered lamellae by the addi-tion of
monogalactosyl diacylglycerol [44]. They con-cluded that the light
harvesting complex togetherwith monogalactosyl diacylglycerol is
responsiblefor lamellae organization of the thylakoid mem-brane.
Therefore interaction between thylakoid pro-teins and thylakoid
lipids seems important for theformation of the lipid bilayer in a
membrane whosemain components are non-bilayer forming lipids.
5. Regulation of thylakoid biogenesis
How is the formation of the thylakoid lipid bilayercoordinated
with the expression of proteins and thebiosynthesis of pigments and
co-factors? It becameobvious quite early after the identication of
DNAand genome structure that plastid development andthylakoid
formation is controlled by both the ge-nome of the cell (nucleome)
and the organelle (plas-tome). Plastids contain up to several
hundred copiesof a circular chromosome with a size between 120and
220 kb. Encoded on the plastome is an averageof about 100^200
proteins in addition to a full set ofribosomal and transfer RNAs
[45,46]. Chloroplastsare, however, estimated to house about
2000^5000dierent proteins; consequently only 5^10% of theplastidal
proteins are encoded within the plastome[46,47] and the majority of
proteins required for plas-tid development and function are encoded
in the nu-cleus. These nuclear encoded proteins are translatedon
cytoplasmic ribosomes and have to be post-trans-lationally
transported to the chloroplast ([48]; Jarvisand Soll, this
issue).
Many protein complexes and biosynthetic path-ways of the
chloroplast contain components encodedboth in the nucleome and in
the plastome and vir-tually all chloroplast functions require the
concertedaction of nuclear and plastidal encoded factors (Fig.2).
Complex regulatory processes are required to en-sure that gene
expression of proteins encoded in thenucleome is properly
coordinated with the expression
BBAMCR 14806 11-12-01
U.C. Vothknecht, P. Westho / Biochimica et Biophysica Acta 1541
(2001) 91^101 95
-
of plastome encoded proteins. At the same time thecoordinated
development of all plastids in one cellhas to be guaranteed.
Thylakoids become photochemically competentvery early in their
development [49,50]. The level oftranscription, which is quite low
in proplastids, in-creases drastically when the chloroplast begins
tomature [51]. At the same time the translational ap-paratus inside
the plastids is built up [52]. It is be-lieved that the nucleus has
the control over the onsetof chloroplast dierentiation and also
takes the lead-ing part in further developmental stages. To
executethis control most regulatory components have beentransferred
to the nucleus [53]. At the same time theplastids signal back their
development stage and con-dition to the nucleus. These signals,
often called theplastidal factor, inuence the expression of
nuclearencoded plastid proteins [54^56]. The biogenesis andfunction
of the chloroplast are therefore an integralpart of the plant cell
and the development of the celland its organelle are interdependent
[57,58]. This issupported by the fact that plastids cannot easily
beexchanged into a dierent cell background [59^61].
This interdependence of the cell and its organelle isfurther
strengthened by the fact that two dierentRNA polymerases are
required to transcribe plasti-
dal genes [62,63]. This includes a phage-type RNApolymerase of
nuclear origin [64] and an eubacterial,multisubunit enzyme whose
core subunits are en-coded by the plastome [63] while its sigma
factorsubunits are encoded by nuclear genes [65]. The nu-clear
encoded RNA polymerase is primarily respon-sible for transcription
of so-called housekeepinggenes of the chloroplasts, while the
bacterial-typeenzyme preferentially transcribes genes
encodingcomponents of the photosynthetic machinery [66].
Very little is known so far about the regulation ofplastidal
import in relation to plastid development.Most studies on the
regulation of plastidal importhave been done on fully mature,
photosyntheticallyactive chloroplasts (Jarvis and Soll, this
issue). Arecent publication indicates a direct inuence of as-sembly
of the light harvesting complex on the importof the chlorophyll
binding protein into the chloro-plast [42]. A similar regulation
could be envisionedfor other nuclear encoded chloroplast proteins
sincealso in mature chloroplasts the thylakoid composi-tion is very
dynamic and undergoes constant changesin order to adapt to changing
environmental condi-tions. The ability for adaptation is specially
impor-tant since plants are not mobile and can thereforenot escape
unfavorable conditions. Only a constant
Fig. 2. Schematic display of nucleus^chloroplast interaction.
Synthesis of plastid encoded proteins is regulated by nuclear
encoded fac-tors from the point of gene expression and translation
until the nal incorporation into the thylakoid membrane. At the
same timethe chloroplast signals the nucleus about its state of
development. This signal inuences the expression of nuclear encoded
genes. Thisgure is based on a scheme presented by Rochaix
[102].
BBAMCR 14806 11-12-01
U.C. Vothknecht, P. Westho / Biochimica et Biophysica Acta 1541
(2001) 91^10196
-
communication between the organelle and the nu-cleus can ensure
a coordinated supply of all the dif-ferent factors required.
6. Analysis of thylakoid biogenesis through mutants
Mutants are a powerful tool to study the involve-ment of gene
products on specic processes. Manydierent mutants that display
deciencies in plastiddevelopment and thylakoid formation exist in a
widerange of species. Many of these mutants are ran-domly occurring
natural variations, others are man-made. In early work, new mutants
were produced bytreatment of plants or algae with radiation or
chem-ical mutagens [43,67]. Later, the necessary genetictools
became available for random insertional muta-genesis by T-DNA of
Agrobacterium tumefaciens[68^70] or transposable elements. Only a
limited setof these mutants can be discussed within this
section.For a more detailed summary of mutants see[43,71,72].
There are many dierent types of mutations thataect both plastid
development and thylakoid for-mation and the eect that a mutation
has on eitheris often dicult to distinguish. Often these mutantsare
blocked in a step of a biosynthetic pathway lo-cated inside the
chloroplasts. The resulting loss of afunctional component of the
plastid then extends itseect on the macromolecular structures. In
othermutants structural components of the thylakoidmembrane are
missing or defective. Mutations canaect plastids in all stages of
thylakoid formation.In several cases plastids are blocked very
early indevelopment. These mutants include dcl from tomato[73], dag
from Antirrhinum [74], cla1-1 from Arabi-dopsis thaliana [75] and
several albina mutants ofbarley [65,76]. Plastids in dcl, dag and
cla1-1 seemto be arrested in the proplastid stage while plastids
insome of the barley albina mutants can reach the sizeof mature
chloroplasts but remain fully depleted ofinternal membrane
structures except for vesicles thataccumulate in some of them.
A similar phenotype can be observed in vrpoA, Band C1 mutants
that lack the bacterial-type RNApolymerase and consequently the
ability to transcribethe photosynthetic genes which encode subunits
ofthylakoid protein complexes [62,77]. Other mutants
can be found that are blocked in later stages of plas-tid
development, anywhere from the proplastid tomature chloroplasts.
Because of the close connectionbetween plastid development and
thylakoid forma-tion it is often dicult to distinguish pleiotropic
ef-fects of these mutations. In some cases mutants seemto suer from
a secondary destruction of the internalmembrane structure rather
than a defect in thylakoidsynthesis [78,79].
Defects in thylakoid formation are often caused bymutations that
result in a depletion of major protein-aceous components of the
thylakoid membrane, e.g.major components of the photosystems. For
in-stance, the hcf136 mutant of A. thaliana cannot as-semble a
functional photosystem II, and this defect isassociated with a
drastically disturbed thylakoidmembrane system [80]. Mutations of
the protein im-port apparatus of chloroplasts cause similar
defectsin thylakoid formation [81]. Other mutations thathave a
great impact on thylakoid formation are mu-tations that aect the
import pathways by whichproteins are inserted into the thylakoid
membrane[82^84]. Examples for such mutants can be foundin maize in
the form of tha1 and tha5 which inhibitthe SecA-type import pathway
and hcf106 and tha4where the vph or Tat pathway is disrupted
[85,86].Not surprisingly, mutants that aect the synthesis
ofimportant thylakoid lipids display alterations in thechloroplast
ultrastructure. Arabidopsis dgd1 andmdg1 mutants lack the enzymes
monogalactosyl di-acylglycerol synthase or digalactosyl
diacylglycerolsynthase that are required for the formation of
thetwo major thylakoid membrane lipids. These mu-tants show a wide
range of alterations includingchanges in the chloroplast
ultrastructure and proteincomposition [87^89].
Also very common is the connection between de-ciencies in
thylakoid formation and disruption ofpigment biosynthesis
[43,67,90]. While pleiotropic ef-fects of these mutations cannot be
excluded in somecases, many investigations have supported the
poten-tial inuence of chlorophyll production on chloro-plast
development [43,90^93]. This connection is es-pecially interesting
in light of the plastidal factorthat is discussed as a signal from
the chloroplaststo the nucleus (Fig. 2). As described above, the
plas-tidal factor is thought to signal the developmentalstage of
the plastid to the nucleus and aect the
BBAMCR 14806 11-12-01
U.C. Vothknecht, P. Westho / Biochimica et Biophysica Acta 1541
(2001) 91^101 97
-
expression of many dierent nuclear encoded genes[55,56]. So far
the nature of this plastidal factor re-mained elusive and
indication for the existence ofmore than one signaling pathways
exists. A recentpaper by Chory and coworkers on the
Arabidopsismutant uncoupled 5, together with earlier studies
byother groups, provides evidence that one of thesefactors might
have been found ([94] and referencestherein). Their ndings indicate
that a subunit ofMg-chelatase, the enzyme that converts
protopor-phyrin IX into Mg-protoporphyrin, has an addition-al,
distinct function in the plastid^nucleus signalingpathway.
Especially interesting are mutants that aect thy-lakoid
formation in otherwise fully developed chlo-roplasts. One recent
example is the vipp1 mutants ofArabidopsis and Synechocystis.
Mutant analysisshowed that the gene product of vipp1 is involvedin
the biogenesis of thylakoids in Arabidopsis andcyanobacteria
[35,95]. Interruption of the vipp1gene locus results in a complete
loss of thylakoidmembranes. It seems that Vipp1 is directly
involvedin the process of thylakoid biogenesis. Even
more,phylogenetic analysis indicated that the presence ofthis
protein might be a prerequisite to the ability ofcyanobacteria and
chloroplasts to form internalmembranes. Interestingly, the
Arabidopsis vipp1 mu-tant additionally lost the ability for vesicle
forma-tion. A vesicle transport system might thus be impor-tant for
thylakoid formation in mature chloroplasts.
7. Evolution of the thylakoid membrane system
Cyanobacteria are the only phototrophic prokary-otes that carry
out oxygenic photosynthesis with twophotosystems. They very much
resemble chloroplastsand it is assumed that at the time of the
endosym-biotic event they had already invented oxygenic
pho-tosynthesis and developed most of the photosyn-thetic features
found in chloroplasts today. Likechloroplasts, most cyanobacteria
contain an internalmembrane system in which the photosynthetic
appa-ratus is located. Extensive stacking of grana lamellaeis not
found in these organisms. Their thylakoids areorganized in layers
often paralleling the contour ofthe cells. Algae are probably the
organism most sim-ilar to the early endosymbiotic cells. Similar to
cya-
nobacteria, most algae do not contain grana stacks.Chloroplasts
of red algae contain a simple thylakoidstructure similar to
cyanobacteria. In green andbrown algae regions of closely appressed
thylakoidmembranes occur similar to grana stacks in chloro-plasts
of higher plants [15]. Also many algae containonly a single
chloroplast per cell. These structuralsimilarities t well with an
evolutionary position be-tween the cyanobacterial endosymbiont and
higherplants.
It is still a point of debate where photosynthesisdeveloped in
the rst place. Recent results favor anorigin of photosynthesis in
anoxygenic bacteria [1].Phototropic green and purple bacteria carry
out an-oxygenic photosynthesis with a single photosystemstrongly
resembling photosystem I. In green-sulfurbacteria the
photosynthetic machinery is located inthe cytoplasmic membrane and
the antenna com-plexes reside in a special non-membranous
structure,the chlorosomes, closely attached to the
cytoplasmicmembrane [96]. Purple bacteria on the other handoften
display strong invagination of the cytoplasmicmembrane and their
photosystems are concentratedin these intracytoplasmic membrane
regions [97,98].It is believed that these membranes are not
fullyseparated from the cytoplasmic membrane and stillform a
continuum with the latter [20,21]. It is there-fore tempting to
speculate that the development ofoxygenic photosynthesis is
connected to two dier-ent events: the invention of the second
photosystemand the biogenesis of an internal membrane
systemdisconnected from the cell membrane. Support forthis
speculation arose from the identication ofVipp1, a protein
essential for thylakoid formationin higher plant chloroplasts and
cyanobacteria[35,95]. Phylogenetic analysis showed that Vipp1can be
found in organisms that carry out oxygenicphotosynthesis, i.e.
plants, algae and cyanobacteria.No Vipp1 homologue has been found
so far in bac-teria including those that are capable of
anoxygenicphotosynthesis, such as Rhodobacter or Chlorobium.Vipp1
shares sequence homology with a subunit ofthe bacterial phage
shock, pspA, and might haveoriginated from a gene duplication of
the latter inan ancestor of cyanobacteria. It subsequently
ob-tained an additional C-terminal domain that seemsessential for
its function in thylakoid formation. InArabidopsis the vipp1
mutation also interrupts
BBAMCR 14806 11-12-01
U.C. Vothknecht, P. Westho / Biochimica et Biophysica Acta 1541
(2001) 91^10198
-
vesicle trac between the inner envelope and thethylakoids. No
such vesicle transport has yet beenshown in any prokaryotic
organism including cya-nobacteria. Further studies are needed to
showwhether vesicle transport is a feature that developedonly in
chloroplasts.
At least one cyanobacterium performs oxygenicphotosynthesis
without having thylakoids. Gloeo-bacter violaceus was rst isolated
in 1972 from alimestone rock in Switzerland [99]. Electron
micro-scopic studies revealed the complete lack of
internalmembranes. Not even invaginations of the plasmamembrane
were observed. Nevertheless, these cellsperform oxygenic
photosynthesis [100,101]. The pho-tosystems are located on the
plasma membrane and,similar to purple and green-sulfur bacteria,
they formtheir proton gradient along the plasma membrane.This
organism might be a cyanobacterium at a stagebefore biogenesis of
thylakoids was invented or hasresulted from a secondary loss of
thylakoid mem-branes. Compared to cyanobacteria with
thylakoidstheir photosynthetic capacity is very low. Thus, e-cient
oxygenic photosynthesis may require the pres-ence of an internal
membrane system.
While it is easy to envision the evolution of chlo-roplasts from
a cyanobacterium, it is much moredicult to understand the
evolutionary processesthat created the multiple forms of plastids.
There isno indication that the structures found in proplas-tids,
chromoplasts or leucoplasts have been part ofthe genetic plan that
the endosymbiont transferred tothe host cell. It must be assumed
that this develop-ment took place after the endosymbiotic event
andwas imposed on the plastid by the host cell. It will befor
future research to elucidate the evolutionary trueorigin of the
thylakoid membrane and its evolutionfrom simple single membrane
layers to the complexsystem present in plant chloroplasts.
Acknowledgements
The authors would like to thank Prof. Dr. J. Sollfor helpful
suggestions and discussions. We wouldalso like to thank S. Westphal
and C. Glockmanfor the electron microscopic pictures in Fig. 1.
Finan-cial support by the Deutsche Forschungsgemein-schaft SFB TR1
is acknowledged.
References
[1] F. Xiong, W.M. Fischer, K. Inoue, M. Nakahara, C.E. Ba-uer,
Science 289 (2000) 1724^1730.
[2] A.F.W. Schimper, Bot. Z. 7, 8, 9, 10 (1883) 105^112;
121^131; 137^146; 153^160.
[3] A.F.W. Schimper, Jb. Wiss. Bot. 16 (1885) 1^247.[4] G.A.
Kausche, H. Ruska, Naturwissenschaften 28 (1940)
303^304.[5] A. Frey-Wyssling, Protoplasmologia IIA (1955) 2.[6]
S. Granick, Encyclopedia Plant Physiol. 1 (1955) 507.[7] D. von
Wettstein, J. Ultrastruct. Res. 3 (1959) 234^240.[8] S. Strugger,
Naturwissenschaften 37 (1950) 166^167.[9] K. Muhlethaler, A.
Frey-Wyssing, J. Biophys. Biochem. Cy-
tol. 6 (1959) 507^512.[10] W. Menke, Z. Naturforsch. 16b (1961)
334^336.[11] H. Leyon, Exp. Cell Res. 5 (1953) 520^529.[12] B.E.S.
Gunning, Protoplasma 60 (1965) 111^130.[13] M. Ryberg, A.S.
Sandelius, E. Selstram, Physiol. Plant. 57
(1983) 555^560.[14] M. Ryberg, C. Sudqvist, Physiol. Plant. 73
(1988) 218^226.[15] A.R. Wellburn, Int. Rev. Cytol. (1982) Suppl.
17.[16] D. von Wettstein, A. Kahn, in: A.L. Houwink, B.J. Snit
(Eds.) Eur. Reg. Conf. Electron Microscopy Delft, Vol. II,1960,
pp. 1051^1054.
[17] B.E.S. Gunning, M.P. Jagoe, in: T.W. Goodwin (Ed.),
Bio-chemistry of Chloroplasts, Vol. II, Academic Press,
London,1967, pp. 655^676.
[18] J.T.O. Kirk, in: J.T. Kirk, R.A.E. Tilney-Bassett (Eds.),
ThePlastids, W.H. Freeman and Co., London, 1967, pp. 1^91.
[19] R. Dierstein, A. Schumacher, G. Drews, Arch. Microbiol.128
(1981) 376^383.
[20] G. Drews, J.R. Golecki, in: R.E. Blankenship, M.T.
Madi-gan, C.E. Bauer (Eds.), Advances in Photosynthesis, Vol.
2,Kluwer Academic Publishers, Dordrecht, 1995, pp. 231^257.
[21] B.E.S. Gunning, M.W. Steer, Ultrastructure and the
Biologyof Plant Cells, Arnold, London, 1975.
[22] J.M. Anderson, B. Andersson, Trends Biochem. Sci. 13(1988)
351^355.
[23] R. Douce, J. Joyard, in: D.R. Ort, C.F. Yocum (Eds.),
Ad-vances in Photosynthesis, Vol. 4, Kluwer Academic Publish-ers,
Dordrecht, 1996, pp. 69^101.
[24] P.R. Chitnis, Annu. Rev. Plant Physiol. Plant Mol. Biol.
52(2001) 593^626.
[25] B. Hankamer, J. Barber, E.J. Boekema, Annu. Rev.
PlantPhysiol. Plant Mol. Biol. 48 (1997) 641^671.
[26] E.A. Berry, M. Guergova-Kuras, L. Huang, A.R. Crofts,Annu.
Rev. Biochem. 69 (2000) 1005^1075.
[27] G. Groth, H. Strotmann, Physiol. Plant. 106 (1999)
142^148.[28] F.-A. Wollman, L. Minai, R. Nechushtai, Biochim.
Biophys.
Acta 1411 (1999) 21^85.[29] Y. Choquet, O. Vallon, Biochimie 82
(2000) 615^634.[30] P.A . Albertsson, Photosynth. Res. 46 (1995)
141^149.[31] J.P. Carde, J. Joyard, R. Douce, Biol. Cell 44 (1982)
315^
324.[32] J.K. Hoober, Biochem. Plants 10 (1989) 1^74.
BBAMCR 14806 11-12-01
U.C. Vothknecht, P. Westho / Biochimica et Biophysica Acta 1541
(2001) 91^101 99
-
[33] J.B. Bock, H.T. Matern, A.A. Peden, R.H. Scheller,
Nature409 (2001) 839^841.
[34] D.J. Morre, G. Sellden, C. Sundquist, A.S. Sandelius,
PlantPhysiol. 97 (1991) 1558^1564.
[35] D. Kroll, K. Meierho, N. Bechtold, M. Kinoshita, S.
West-phal, U.C. Vothknecht, J. Soll, P. Westho, Proc. Natl.Acad.
Sci. USA 98 (2001) 4238^4242.
[36] J.K. Hoober, C.O. Boyd, L.G. Paavola, Plant Physiol.
96(1991) 1321^1328.
[37] D.J. Morre, N. Minnield, M. Paulik, Biol. Cell 67
(1989)51^60.
[38] J. Joyard, E. Teyssier, C. Miege, D. Berny-Seigneurin,
E.Marechal, M.A. Block, A.-J. Dorne, N. Rolland, G. Ajlani,R.
Douce, Plant Physiol. 118 (1998) 715^723.
[39] D. von Wettstein, Proc. Natl. Acad. Sci. USA 98
(2001)3633^3635.
[40] R. Douce, Science 183 (1974) 852^853.[41] J. Joyard, R.
Douce, H.P. Siebertz, E. Heinz, Eur. J. Bio-
chem. 108 (1980) 171^176.[42] J.K. Hoober, L.L. Eggink, FEBS
Lett. 489 (2001) 1^3.[43] C.R. Somerville, Annu. Rev. Plant
Physiol. 37 (1986) 467^
507.[44] I. Simidjiev, S. Stoylova, H. Amenitsch, R. Javor, L.
Mus-
tardy, P. Laggner, A. Holzenburg, G. Garab, Proc. Natl.Acad.
Sci. USA 97 (2000) 1473^1476.
[45] M. Sugiura, Plant Mol. Biol. 19 (1992) 149^168.[46] W.
Martin, B. Stoebe, V. Goremykin, S. Hansmann, M.
Hasegawa, K.V. Kowallik, Nature 393 (1998) 162^165.[47] F.
Abdallah, F. Salami, D. Leister, Trends Plant Sci. 5
(2000) 141^142.[48] U.C. Vothknecht, J. Soll, Biol. Chem. 381
(2000) 887^897.[49] N.R. Baker, V. Miranda, in: G. Akoyunoglou
(Ed.), Photo-
synthesis, Vol. 5, Chloroplast Development, Balaban
Inter-national Science Services, Philadelphia, PA, 1981, pp.
367^376.
[50] N.R. Baker, A.N. Webber, M. Bradbury, J.P. Markwell,M.G.
Baker, J.P. Thornber, in: L.A. Staehelin, R.B. Hallick,J.P.
Thornber (Eds.), Biosynthesis of the Photosynthetic Ap-paratus:
Molecular Biology, UCLA Symposium Series, AlanR. Liss, New York,
1984, pp. 237-255.
[51] B.J. Baumgartner, J.C. Rapp, J.E. Mullet, Plant Physiol.
89(1989) 1011^1018.
[52] H. Harrak, T. Langrange, C. Bisanz-Seyer, S. Lerbs-Mache,R.
Mache, Plant Physiol. 108 (1995) 685^692.
[53] R.G. Herrmann, in: H.E.A. Schenk, R.G. Herrmann,
K.W.Muller, W. Schwemmler (Eds.), Eukaryotism and
Symbiosis,Springer, Heidelberg, 1997, pp. 73^118.
[54] R. Oelmuller, Photochem. Photobiol. 49 (1989) 229^239.[55]
R. Susek, J. Chory, Aust. J. Plant Physiol. 19 (1992) 387^
399.[56] W.C. Taylor, Annu. Rev. Plant Physiol. Plant Mol. Biol.
40
(1989) 211^233.[57] R.M. Leech, in: N.R. Baker, J. Barber
(Eds.), Topics in
Photosynthesis, Vol. 5, Elsevier, Amsterdam, 1984, pp. 1-21.[58]
R.G. Herrmann, P. Westho, in: B. Anderson, E.M. Aro
(Eds.), Regulatory Aspects of Photosynthesis, Kluwer Aca-demic
Publishers, Dordrecht, 2001, in press.
[59] O. Renner, Ber. Math.-phys. Kl., Sachs. Akad. Wiss.
Leipzig86 (1934) 241^266.
[60] W. Stubbe, Z. Vererbungsl. 90 (1959) 288^298.[61] P.
Medgyesy, in: P.J. Dix (Ed.), Plant Cell Line Selection,
VCH Verlagsgesellschaft, Weinheim, 1990, pp. 287^316.[62] L.
Allison, L. Simon, P. Maliga, EMBO J. 15 (1996) 2802^
2809.[63] W.R. Hess, T. Borner, Int. Rev. Cytol. 190 (1999)
1^59.[64] B. Hedtke, T. Borner, A. Weihe, Science 277 (1997)
809^811.[65] L.A. Allison, Biochimie 82 (2000) 537^548.[66] K.
Liere, P. Maliga, in: B. Anderson, E.M. Aro (Eds.),
Regulatory Aspects of Photosynthesis, Kluwer AcademicPublishers,
Dordrecht, 2001, in press.
[67] K.W. Henningsen, J.E. Boynton, D. von Wettstein, R.
Dan.Acad. Sci. Lett. Biol. Skrifter 42 (1993) 1^349.
[68] V. Sundaresan, Trends Plant Sci. 1 (1996) 184^190.[69] R.
Azpiroz-Leehan, K.A. Feldmann, Trends Genet. 13
(1997) 152^156.[70] V. Walbot, Curr. Opin. Plant Biol. 3 (2000)
103^107.[71] P. Leon, A. Arroyo, S. Mackenzie, Annu. Rev. Plant
Phys-
iol. Plant Mol. Biol. 49 (1998) 453^480.[72] D. von Wettstein,
S. Gough, C.G. Kannangara, Plant Cell 7
(1995) 1039^1057.[73] J.S. Keddie, B. Carroll, J.D.G. Jones, W.
Gruissem, EMBO
J. 15 (1996) 4208^4217.[74] M. Chatterjee, S. Sparvoli, C.
Edmunds, P. Garosi, K. Fin-
dlay, C. Martin, EMBO J. 15 (1996) 4194^4207.[75] M.A. Mandel,
K.A. Feldmann, L. Herrera-Estrella, M. Ro-
cha-Sosa, P. Leon, Plant J. 9 (1996) 649^658.[76] D. von
Wettstein, Can. J. Bot. 39 (1961) 1537^1545.[77] G. De
Santis-Maciossek, W. Kofer, A. Bock, S. Schoch,
R.M. Maier, G. Wanner, W. Rudiger, H.-U. Koop, R.G.Herrmann,
Plant J. 18 (1999) 477^489.
[78] G. Robbelen, Z. Vererbungsl. 90 (1959) 503.[79] W. Stubbe,
D. von Wettstein, Protoplasma 45 (1955) 241^
250.[80] J. Meurer, H. Plucken, K.V. Kowallik, P. Westho,
EMBO
J. 17 (1999) 5286^5297.[81] J. Bauer, K.H. Chen, A. Hiltbunner,
E. Wehrli, M. Eugster,
D. Schnell, F. Kessler, Nature 403 (2000) 203^207.[82] A.
Barkan, R. Voelker, J. Mendel-Hartvig, D. Johnson, M.
Walker, Physiol. Plant. 93 (1995) 163^170.[83] C. Robinson, P.J.
Hynds, D. Robinson, A. Mant, Plant
Mol. Biol. 38 (1998) 209^221.[84] A.M. Settles, R. Martienssen,
Trends Cell Biol. 8 (1998)
494^501.[85] R.A. Martienssen, A. Barkan, M. Freeling, W.C.
Taylor,
EMBO J. 8 (1989) 1633^1639.[86] R. Voelker, A. Barker, EMBO J.
14 (1995) 3905^3914.[87] P. Dormann, S. Homann-Benning, I. Balbo,
C. Benning,
Plant Cell 7 (1995) 1801.[88] P. Dormann, I. Balbo, C. Benning,
Science 284 (1999) 2181^
2184.
BBAMCR 14806 11-12-01
U.C. Vothknecht, P. Westho / Biochimica et Biophysica Acta 1541
(2001) 91^101100
-
[89] P. Jarvis, P. Dormann, C.A. Peto, J. Lutes, C. Benning,
J.Chory, Proc. Natl. Acad. Sci. USA 97 (2000) 8175^8179.
[90] T.G. Falbel, L.A. Staehelin, Plant Physiol. 104 (1994)
639^648.[91] A. Hudson, R. Carpenter, S. Doyle, E.S. Coen, EMBO J.
12
(1993) 3711^3719.[92] P.E. Jensen, R.D. Willows, B.L.
Larsen-Petersen, U.C.
Vothknecht, B.M. Stummann, C.G. Kannangara, D. vonWettstein,
K.W. Henningsen, Mol. Gen. Genet. 250 (1996)383^394.
[93] S. Runge, B. van Cleve, N. Lebedev, G. Armstrong, K.Apel,
Planta 197 (1995) 490^500.
[94] N. Mochizuki, J.A. Brusslan, R. Larkin, A. Nagatani,
J.Chory, Proc. Natl. Acad. Sci. USA 98 (2001) 2053^2058.
[95] S. Westphal, L. Heins, J. Soll, U.C. Vothknecht, Proc.Natl.
Acad. Sci. USA 98 (2001) 4243^4248.
[96] R.E. Blankenship, R.C., in: L.A. Staehelin, C.J.
Arntzen(Eds.), Photosynthesis III, Springer Verlag, Berlin,
1986,pp. 390^399.
[97] N. Pfennig, H.G. Truper, Ann. Microbiol. 134B (1983)
9^20.[98] C.C. Remsen, Int. Rev. Cytol. 76 (1982) 195^223.[99] R.
Rippka, Waterbury, Gg. Cohen-Bazire, Arch. Micro-
biol. 100 (1974) 419^430.[100] G. Guglielmi, G. Cohen-Bazire,
D.A. Bryant, Arch. Micro-
biol. 129 (1981) 181^189.[101] F. Konig, M. Schmitd, Physiol.
Plant. 94 (1995) 621^628.[102] J.-D. Rochaix, Plant Mol. Biol. 32
(1996) 327^341.
BBAMCR 14806 11-12-01
U.C. Vothknecht, P. Westho / Biochimica et Biophysica Acta 1541
(2001) 91^101 101