Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1994 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 269, No. 26, Issue of July 1, pp. 17440-17447, 1994 Printed in U.S.A.
Endoproteolytic Processing of Proopiomelanocortin and Prohormone Convertases 1 and 2 in Neuroendocrine Cells Overexpressing Prohormone Convertases 1 or 2”
(Received for publication, March 13, 1994)
An Zhou and Richard E. Mains$ From the Department of Neuroscience, The Johns Hopkins University School of Medicine, Baltimore, Maryland 21205
AtT-20 mouse corticotrope tumor cell lines overex- pressing the prohormone convertases PC1 or PC2 were established and used to examine prohormone and pro- hormone convertase biosynthetic processing. On a mo- lar basis, wild-type AtT-20 cells synthesize about 20% as much PC1 as the endogenous prohormone, proopiomela- nocortin (POMC). Kinetic, oligosaccharide, and tem- perature blockade analyses established that proPC1 is converted to PC1 in the endoplasmic reticulum at a rate independent of the level of PC1 or PC2 expression. In contrast, proPC2 is converted to PC2 primarily in a post- trans-Golgi compartment. PC1 is further shortened from its COOH-terminal end in a post-trans-Golgi compart- ment in a step that is accelerated at higher levels of PC1 expression, but unaltered by PC2 overexpression. The initial steps in POMC processing are speeded up by overexpression of PC1, and overexpression of PC1 leads to more extensive cleavage of POMC to smaller prod- ucts. However, even when the rate of PC1 synthesis ex- ceeds that for POMC by 2-fold, PC1 does not cleave the Lys-Lys or Arg-Lys bonds cleaved upon overexpression of PC2.
PC1’ and PC2 are members of a recently identified family of mammalian subtilisin-like prohormone convertases. Selected members of this family are thought to be involved in the endo- proteolytic processing of prohormones at paired basic amino acid sites to generate bioactive peptides (for recent reviews, see Lindberg, 1991; Smeekens, 1993; Bloomquist and Mains, 1993; Seidah et al., 1993).
PC1 and PC2 are expressed primarily in neuroendocrine tis- sues and share several common features including an NH,- terminal prosequence and similar catalytic cores; PC1 has an extended COOH-terminal non-catalytic domain. PC1 and PC2 are distributed differentially among neuroendocrine tissues. For instance, in anterior pituitary corticotropes, PC1 is highly expressed with very low levels of PC2, while in the intermedi- ate lobe melanotropes both enzymes are expressed at high lev-
Grants DA-00097 and DA-00266. The costs of publication of this article * This study was supported by National Institutes of Drug Abuse
were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. $To whom correspondence should be addressed: Dept. of Neuro-
science, The Johns Hopkins University School of Medicine, 725 N. Wolfe St., Baltimore, MD 21205. Tel.: 410-955-6938; Fax: 410-955-0681.
1 and 2; POMC, proopiomelanocortin; Endo H, endoglycosidase H; The abbreviations used are: PC1 and PC2, prohormone convertases
TES, N-tris-[hydroxymethyIlmethyl-2-aminoethane-sulfonic acid; TGN, trans-Golgi-network; ER, endoplasmic reticulum; cpm, counts/ minute; a-MSH, a-melanotropin; p-LPH, P-lipotropin; PAGE, poly- acrylamide gel electrophoresis.
els, with PC2 being more abundant (Bloomquist et al., 1991; Seidah et al., 1990; Day et al., 1992; Hakes et al., 1991).
In parallel with the different distributions of PC1 and PC2 in corticotropes and melanotropes, proopiomelanocortin (POMC), the common precursor of several important peptide hormones including ACTH, a-MSH, and P-endorphin is processed differ- entially in these two cell types. Intermediate lobe melanotropes perform two cleavages not seen with anterior pituitary cortico- tropes; P-endorphin(1-31) is processed into P-endorphin(l-27), and Lys,-y,-MSH is cleaved from the NH,-terminal fragment of POMC (POMC(1-74)) (Mains and Eipper, 1990; Noel et al., 1991). Metabolic labeling studies indicate that these two addi- tional cleavages occur rather late during the biosynthesis of POMC. The differences in POMC processing and PC1 and PC2 distribution in the pituitary have led to several studies on the possible specific involvement of PC1 and PC2 in the various steps in the biosynthetic processing of POMC. Examples of such studies include overexpression of PC2, expression of an- tisense RNA to PC1 in AtT-20 cells, or coexpression of PC1 and PC2 with POMC in neuroendocrine cell lines which do not express endogenous POMC (Benjannet et al., 1991; Thomas et al., 1991; Thorne and Thomas, 1990; Bloomquist et al., 1991; Seidah et al., 1992; Rhodes et al., 1993; Zhou et al., 1993). These studies demonstrate that PC1 and PC2 cleave selectively at certain paired basic amino acid sites of POMC and other propeptides. However, the biosynthesis and endoproteolytic cleavages of PC1 and PC2 have not been studied in detail in parallel with studies on the processing of POMC.
AtT-20 cells are a mouse anterior pituitary corticotrope tu- mor cell line expressing high endogenous levels of POMC and PC1. Previously we established AtT-20 cell lines stably express- ing high levels of PC2 and showed that expression of PC2 in AtT-20 cells resulted in two additional cleavages normally only seen in melanotropes at late times in the biosynthetic pathway of POMC (Zhou et al., 1993). We have also demonstrated that depressing PC1 expression in AtT-20 cells by applying the an- tisense RNA approach blocked POMC processing at early steps (Bloomquist et al., 1991). These results are indicative of se- quential, specific roles for PC1 and PC2 in POMC processing.
In the present study, we have established stable AtT-20 cell lines overexpressing PC1 to use along with our previously es- tablished AtT-20 cell lines overexpressing PC2 to study the role of PC1 in POMC and PC1 processing. We have developed an- tisera and extraction methods which allow us to analyze simul- taneously the kinetics of POMC, PC1 and PC2 biosynthesis, and processing. With these tools we have been able to evaluate the effects of elevated PC expression on homologous and het- erologous PC processing as well as to distinguish better the steps in POMC processing catalyzed by PC1 and PC2.
MATERIALS AND METHODS Construction and Stable Dansfection of AtT-20 Cells with Rat PC1
cDNA (AtT-20 sPC1)”Ib delete the endogenous poly(A) addition signal
17440
Endoproteolysis in Cells Overexpressing PC1 or PC2 1744 1
PC1 protein PC2 mRNA PC2 protein
I PT Northern Western I PT
kDa A sfX2 sEL k D a A m m kDa AIL sf32 &€2
81 76 - 64- *-
- 7( I
PC1 mRNA
Northern
I)
I
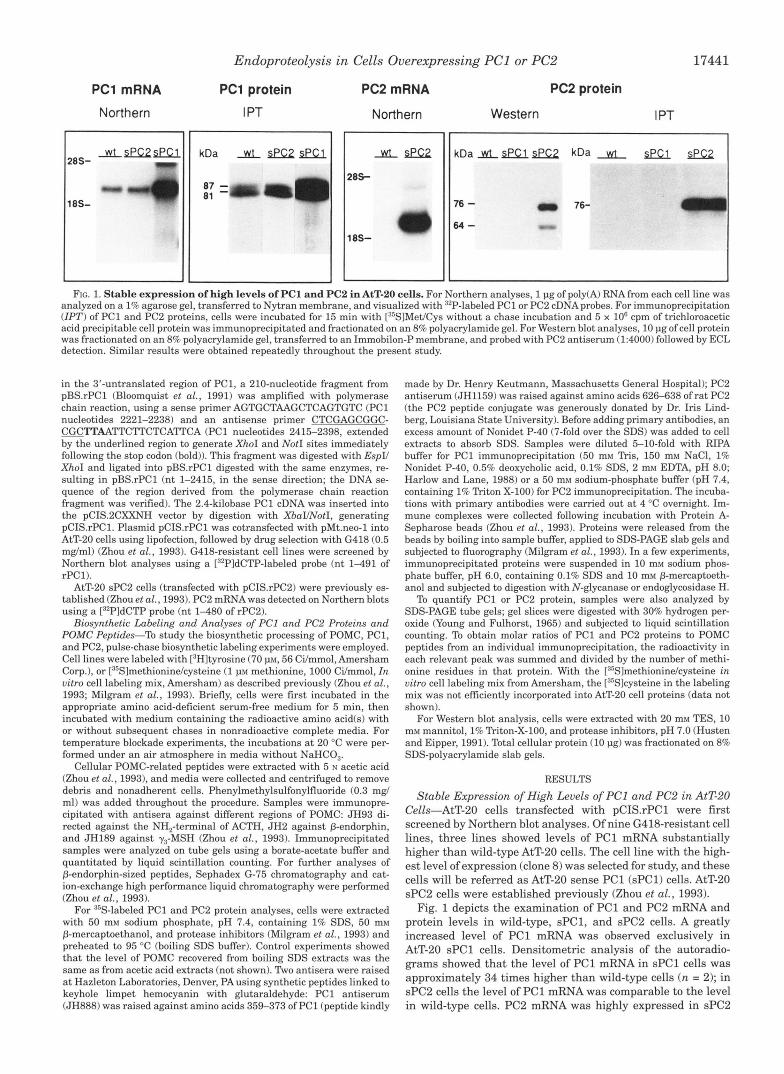
FIG. 1. Stable expression of high levels of PC1 and PC2 in AtT-20 cells. For Northern analyses, 1 pg of poly(A) RNA from each cell line was analyzed on a 1% agarose gel, transferred to Nytran membrane, and visualized with 32P-labeled PC1 or PC2 cDNAprobes. For immunoprecipitation ( IPT) of PC1 and PC2 proteins, cells were incubated for 15 min with ["SlMet/Cys without a chase incubation and 5 x lo6 cpm of trichloroacetic acid precipitable cell protein was immunoprecipitated and fractionated on an 8% polyacrylamide gel. For Western blot analyses, 10 pg of cell protein was fractionated on an 8% polyacrylamide gel, transferred to an Immobilon-P membrane, and probed with PC2 antiserum (1:4000) followed by ECL detection. Similar results were obtained repeatedly throughout the present study.
in the 3"untranslated region of PCI, a 210-nucleotide fragment from pBS.rPC1 (Bloomquist et al., 1991) was amplified with polymerase chain reaction, using a sense primer AGTGCTAAGCTCAGTGTC (PC1 nucleotides 2221-2238) and an antisense primer CTCGAGCGGC- CGCITAA'MCTTCTCATTCA (PC1 nucleotides 2415-2398, extended by the underlined region to generate XhoI and Not1 sites immediately following the stop codon (bold)). This fragment was digested with EspV XhoI and ligated into pBS.rPC1 digested with the same enzymes, re- sulting in pBS.rPC1 (nt 1-2415, in the sense direction; the DNA se- quence of the region derived from the polymerase chain reaction fragment was verified). The 2.4-kilobase PC1 cDNA was inserted into the pCIS.2CXXNH vector by digestion with XballNotI, generating pCIS.rPC1. Plasmid pCIS.rPC1 was cotransfected with pMt.neo-1 into AtT-20 cells using lipofection, followed by drug selection with G418 (0.5 mg/ml) (Zhou et al., 1993). G418-resistant cell lines were screened by Northern blot analyses using a [32PldCTP-labeled probe (nt 1-491 of rPC1).
AtT-20 sPC2 cells (transfected with pCIS.rPC2) were previously es- tablished (Zhouet al., 1993). PC2 mRNA was detected on Northern blots using a [32PldCTP probe (nt 1-480 of rPC2).
Biosynthetic Labeling and Analyses of PC1 and PC2 Proteins and POMC Peptides-% study the biosynthetic processing of POMC, PC1, and PC2, pulse-chase biosynthetic labeling experiments were employed. Cell lines were labeled with ["Hltyrosine (70 PM, 56 Ci/mmol,hnersham Corp.), or 13'Slmethionine/cysteine (1 p~ methionine, 1000 Ci/mmol, In vitro cell labeling mix, Amersham) as described previously (Zhou et al., 1993; Milgram et al., 1993). Briefly, cells were first incubated in the appropriate amino acid-deficient serum-free medium for 5 min, then incubated with medium containing the radioactive amino acid(s) with or without subsequent chases in nonradioactive complete media. For temperature blockade experiments, the incubations at 20 "C were per- formed under an air atmosphere in media without NaHCO,.
Cellular POMC-related peptides were extracted with 5 N acetic acid (Zhou et al., 1993), and media were collected and centrifuged to remove debris and nonadherent cells. Phenylmethylsulfonylfluoride (0.3 mg/ ml) was added throughout the procedure. Samples were immunopre- cipitated with antisera against different regions of POMC: JH93 di- rected against the NH,-terminal of ACTH, JH2 against P-endorphin, and JH189 against y3-MSH (Zhou et al., 1993). Immunoprecipitated samples were analyzed on tube gels using a borate-acetate buffer and quantitated by liquid scintillation counting. For further analyses of P-endorphin-sized peptides, Sephadex G-75 chromatography and cat- ion-exchange high performance liquid chromatography were performed (Zhou et al., 1993).
For 35S-labeled PC1 and PC2 protein analyses, cells were extracted with 50 mM sodium phosphate, pH 7.4, containing 1% SDS, 50 mM P-mercaptoethanol, and protease inhibitors (Milgram et al., 1993) and preheated to 95 "C (boiling SDS buffer). Control experiments showed that the level of POMC recovered from boiling SDS extracts was the same as from acetic acid extracts (not shown). Two antisera were raised a t Hazleton Laboratories, Denver, PAusing synthetic peptides linked to keyhole limpet hemocyanin with glutaraldehyde: PC1 antiserum (JH888) was raised against amino acids 359-373 of PC1 (peptide kindly
made by Dr. Henry Keutmann, Massachusetts General Hospital); PC2 antiserum (JH1159) was raised against amino acids 626-638 of rat PC2 (the PC2 peptide conjugate was generously donated by Dr. Iris Lind- berg, Louisiana State University). Before adding primary antibodies, an excess amount of Nonidet P-40 (7-fold over the SDS) was added to cell extracts to absorb SDS. Samples were diluted 5-10-fold with RIPA buffer for PC1 immunoprecipitation (50 mM Tris, 150 mM NaCI, 1% Nonidet P-40, 0.5% deoxycholic acid, 0.1% SDS, 2 mM EDTA, pH 8.0; Harlow and Lane, 1988) or a 50 mM sodium-phosphate buffer (pH 7.4, containing 1% Triton X-100) for PC2 immunoprecipitation. The incuba- tions with primary antibodies were carried out a t 4 "C overnight. Im- mune complexes were collected following incubation with Protein A- Sepharose beads (Zhou et al., 1993). Proteins were released from the beads by boiling into sample buffer, applied to SDS-PAGE slab gels and subjected to fluorography (Milgram et al., 1993). In a few experiments, immunoprecipitated proteins were suspended in 10 mM sodium phos- phate buffer, pH 6.0, containing 0.1% SDS and 10 mM P-mercaptoeth- anol and subjected to digestion with N-glycanase or endoglycosidase H.
To quantify PC1 or PC2 protein, samples were also analyzed by SDS-PAGE tube gels; gel slices were digested with 30% hydrogen per- oxide (Young and Fulhorst, 1965) and subjected to liquid scintillation counting. To obtain molar ratios of PC1 and PC2 proteins to POMC peptides from an individual immunoprecipitation, the radioactivity in each relevant peak was summed and divided by the number of methi- onine residues in that protein. With the ["Slmethionine/cysteine in vitro cell labeling mix from Amersham, the [3sSlcysteine in the labeling mix was not efficiently incorporated into AtT-20 cell proteins (data not shown).
For Western blot analysis, cells were extracted with 20 mM TES, 10 mM mannitol, 1% Triton-X-100, and protease inhibitors, pH 7.0 (Husten and Eipper, 1991). Total cellular protein (10 pg) was fractionated on 8% SDS-polyacrylamide slab gels.
RESULTS Stable Expression of High Levels of PC1 and PC2 in AtT-20
Cells-AtT-20 cells transfected with pCIS.rPC1 were first screened by Northern blot analyses. Of nine G418-resistant cell lines, three lines showed levels of PC1 mRNA substantially higher than wild-type AtT-20 cells. The cell line with the high- est level of expression (clone 8) was selected for study, and these cells will be referred as AtT-20 sense PC1 (sPC1) cells. AtT-20 sPC2 cells were established previously (Zhou et al., 1993).
Fig. 1 depicts the examination of PC1 and PC2 mRNA and protein levels in wild-type, sPC1, and sPC2 cells. A greatly increased level of PC1 mRNA was observed exclusively in AtT-20 sPCl cells. Densitometric analysis of the autoradio- grams showed that the level of PC1 mRNA in sPCl cells was approximately 34 times higher than wild-type cells ( n = 2); in sPC2 cells the level of PC1 mRNA was comparable to the level in wild-type cells. PC2 mRNA was highly expressed in sPC2
17442
A
7000
6000
5000
4000
3000
PC1 pmlsln
2000
1000 0
0 10 20 30 40 Sllce number
Endoproteolysis in Cells Overexpressing PC1 or PC2
B 2.50 I
wt SPCl sPC2 Cell line
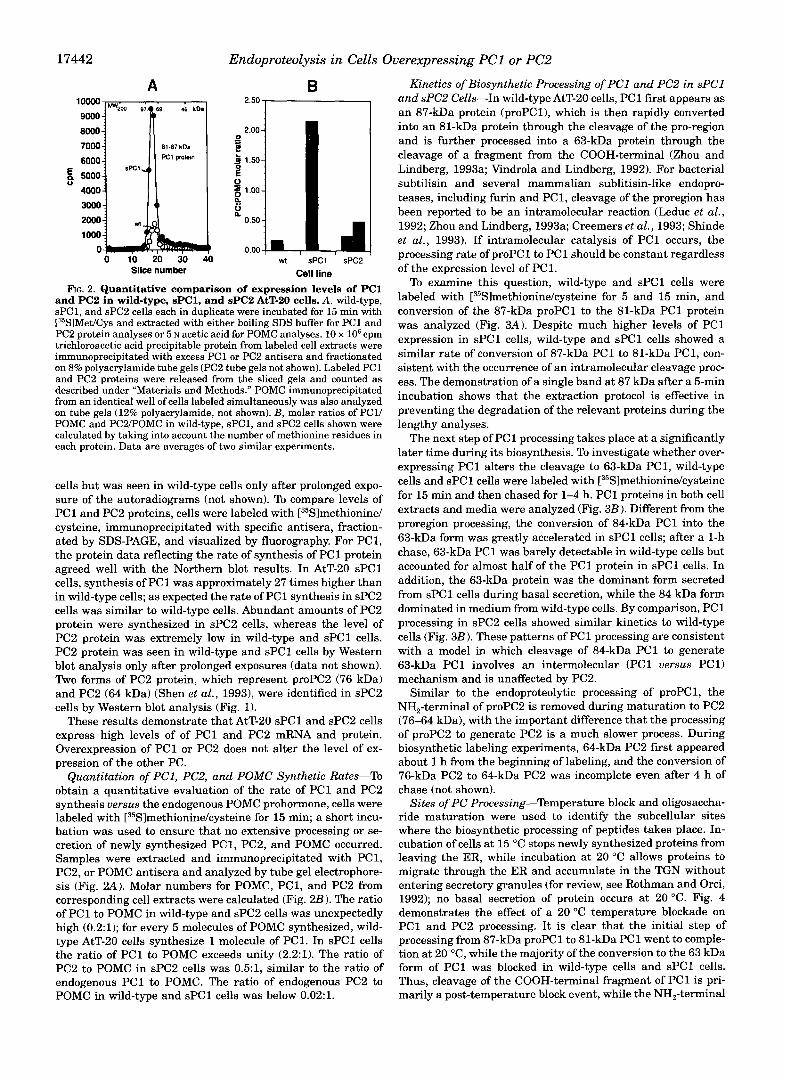
and PC2 in wild-type, sPC1, and sPC2 AtT-20 cells. A. wild-type, FIG. 2. Quantitative comparison of expression levels of PC1
sPC1, and sPC2 cells each in duplicate were incubated for 15 min with [35SlMetfCys and extracted with either boiling SDS buffer for PC1 and PC2 protein analyses or 5 N acetic acid for POMC analyses. 10 x lo6 cpm trichloroacetic acid precipitable protein from labeled cell extracts were immunoprecipitated with excess PC1 or PC2 antisera and fractionated on 8% polyacrylamide tube gels (PC2 tube gels not shown). Labeled PC1 and PC2 proteins were released from the sliced gels and counted as described under "Materials and Methods." POMC immunoprecipitated from an identical well of cells labeled simultaneously was also analyzed
POMC and PC2POMC in wild-type, sPC1, and sPC2 cells shown were on tube gels (12% polyacrylamide, not shown). B, molar ratios of PC1/
calculated by taking into account the number of methionine residues in each protein. Data are averages of two similar experiments.
cells but was seen in wild-type cells only after prolonged expo- sure of the autoradiograms (not shown). To compare levels of PC1 and PC2 proteins, cells were labeled with [35S]methionine/ cysteine, immunoprecipitated with specific antisera, fraction- ated by SDS-PAGE, and visualized by fluorography. For PC1, the protein data reflecting the rate of synthesis of PC1 protein agreed well with the Northern blot results. In AtT-20 sPCl cells, synthesis of PC1 was approximately 27 times higher than in wild-type cells; as expected the rate of PC1 synthesis in sPC2 cells was similar to wild-type cells. Abundant amounts of PC2 protein were synthesized in sPC2 cells, whereas the level of PC2 protein was extremely low in wild-type and sPCl cells. PC2 protein was seen in wild-type and sPCl cells by Western blot analysis only after prolonged exposures (data not shown). Two forms of PC2 protein, which represent proPC2 (76 kDa) and PC2 (64 kDa) (Shen et al., 19931, were identified in sPC2 cells by Western blot analysis (Fig. 1).
These results demonstrate that AtT-20 sPCl and sPC2 cells express high levels of of PC1 and PC2 mRNA and protein. Overexpression of PC1 or PC2 does not alter the level of ex- pression of the other PC.
Quantitation of PC1, PC2, and POMC Synthetic Rates-To obtain a quantitative evaluation of the rate of PC1 and PC2 synthesis uersus the endogenous POMC prohormone, cells were labeled with [35S]methionine/cysteine for 15 min; a short incu- bation was used to ensure that no extensive processing or se- cretion of newly synthesized PC1, PC2, and POMC occurred. Samples were extracted and immunoprecipitated with PC1, PC2, or POMC antisera and analyzed by tube gel electrophore- sis (Fig. 2.4). Molar numbers for POMC, PC1, and PC2 from corresponding cell extracts were calculated (Fig. 2B). The ratio of PC1 to POMC in wild-type and sPC2 cells was unexpectedly high (0.2:l); for every 5 molecules of POMC synthesized, wild- type AtT-20 cells synthesize 1 molecule of PC1. In sPCl cells the ratio of PC1 to POMC exceeds unity (2.2:l). The ratio of PC2 to POMC in sPC2 cells was 0.5:1, similar to the ratio of endogenous PC1 to POMC. The ratio of endogenous PC2 to POMC in wild-type and sPCl cells was below 0.02:l.
Kinetics of Biosynthetic Processing of PC1 and PC2 in sPCl and sPC2 Cells-In wild-type AtT-20 cells, PC1 first appears as an 87-kDa protein (proPCl), which is then rapidly converted into an 81-kDa protein through the cleavage of the pro-region and is further processed into a 63-kDa protein through the cleavage of a fragment from the COOH-terminal (Zhou and Lindberg, 1993a; Vindrola and Lindberg, 1992). For bacterial subtilisin and several mammalian sublitisin-like endopro- teases, including furin and PC1, cleavage of the proregion has been reported to be an intramolecular reaction (Leduc et al., 1992; Zhou and Lindberg, 1993a; Creemers et al., 1993; Shinde et al., 1993). If intramolecular catalysis of PC1 occurs, the processing rate of proPC1 to PC1 should be constant regardless of the expression level of PC1.
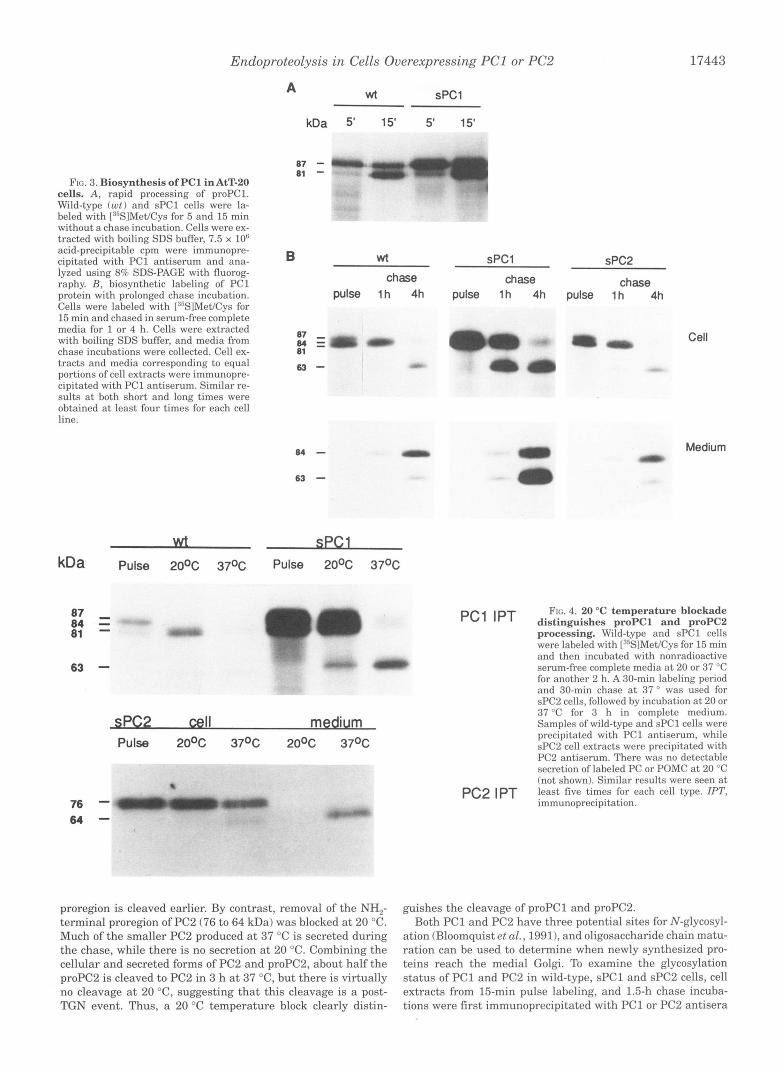
To examine this question, wild-type and sPCl cells were labeled with [35Slmethionine/cysteine for 5 and 15 min, and conversion of the 87-kDa proPC1 to the 81-kDa PC1 protein was analyzed (Fig. 3 A ) . Despite much higher levels of PC1 expression in sPCl cells, wild-type and sPCl cells showed a similar rate of conversion of 87-kDa PC1 to 81-kDa PC1, con- sistent with the occurrence of an intramolecular cleavage proc- ess. The demonstration of a single band at 87 kDa after a 5-min incubation shows that the extraction protocol is effective in preventing the degradation of the relevant proteins during the lengthy analyses.
The next step of PC1 processing takes place at a significantly later time during its biosynthesis. To investigate whether over- expressing PC1 alters the cleavage to 63-kDa PC1, wild-type cells and sPCl cells were labeled with [35S]methionine/cysteine for 15 min and then chased for 1-4 h. PC1 proteins in both cell extracts and media were analyzed (Fig. 3B). Different from the proregion processing, the conversion of 84-kDa PC1 into the 63-kDa form was greatly accelerated in sPCl cells; after a 1-h chase, 63-kDa PC1 was barely detectable in wild-type cells but accounted for almost half of the PC1 protein in sPCl cells. In addition, the 63-kDa protein was the dominant form secreted from sPCl cells during basal secretion, while the 84 kDa form dominated in medium from wild-type cells. By comparison, PC1 processing in sPC2 cells showed similar kinetics to wild-type cells (Fig. 3B). These patterns of PC1 processing are consistent with a model in which cleavage of 84-kDa PC1 to generate 63-kDa PC1 involves an intermolecular (PC1 uersus PC11 mechanism and is unaffected by PC2.
Similar to the endoproteolytic processing of proPC1, the NH,-terminal of proPC2 is removed during maturation to PC2 (76-64 kDa), with the important difference that the processing of proPC2 to generate PC2 is a much slower process. During biosynthetic labeling experiments, 64-kDa PC2 first appeared about 1 h from the beginning of labeling, and the conversion of 76-kDa PC2 to 64-kDa PC2 was incomplete even after 4 h of chase (not shown).
Sites of PC Processing-Temperature block and oligosaccha- ride maturation were used to identify the subcellular sites where the biosynthetic processing of peptides takes place. In- cubation of cells a t 15 "C stops newly synthesized proteins from leaving the ER, while incubation at 20 "C allows proteins to migrate through the ER and accumulate in the TGN without entering secretory granules (for review, see Rothman and Orci, 1992); no basal secretion of protein occurs at 20 "C. Fig. 4 demonstrates the effect of a 20 "C temperature blockade on PC1 and PC2 processing. I t is clear that the initial step of processing from 87-kDa proPC1 to 81-kDa PC1 went to comple- tion a t 20 "C, while the majority of the conversion to the 63 kDa form of PC1 was blocked in wild-type cells and sPCl cells. Thus, cleavage of the COOH-terminal fragment of PC1 is pri- marily a post-temperature block event, while the NH,-terminal
Endoproteolysis i n Cells Overexpressing PC1 or PC2 17443
FIG. 3. Biosynthesis of PC1 in AtT-20 cells. A, rapid processing of proPC1. Wild-type (ut) and sPCl cells were la- beled with [:J5S1Met/Cys for 5 and 15 min without a chase incubation. Cells were ex- tracted with boiling SDS buffer, 7.5 x 10" acid-precipitable cpm were immunopre- cipitated with PC1 antiserum and ana- lyzed using 8% SDS-PAGE with fluorog- raphy. R , biosynthetic labeling of PC1 protein with prolonged chase incubation. Cells were labeled with I:%lMet/Cys for 15 min and chased in serum-free complete media for 1 or 4 h. Cells were extracted with boiling SDS buffer, and media from chase incubations were collected. Cell ex- tracts and media corresponding to equal portions of cell extracts were immunopre- cipitated with PC1 antiserum. Similar re- sults at both short and long times were obtained a t least four times for each cell line.
63 -
76 - 64-
A wt SPCl
87 - 81 -
B wt sPCl sPC2
chase chase chase pulse 1 h 4h pulse 1 h 4h pulse 1 h 4h
84-
63 - 0 Medium
am
pcl IPT distinguishes proPC1 and proPC2 FIG. 4. 20 "C temperature blockade
processing. Wild-type and sPCl cells were labclcd with [:"SIMet/Cys for 15 min and then incubated with nonradioactive
for another 2 h. A 30-min labeling period and 30-min chase at 37 ' was used for sPC2 cells, followed by incubation a t 20 or 37 "C for 3 h in comdete medium.
- serum-free complete media a t 20 or 37 "C
sPC2 cell medium Pulse 20% 37oc 20% 37oc
Samples of wild-type and ;PC1 cells were precipitated with PC1 antiserum, while sPC2 cell extracts were precipitated with PC2 antiserum. There was no detectable secretion of labeled PC or POMC a t 20 "C (not shown). Similar results were seen a t
PC2 IPT !east five times for each cell type. IPT, Immunoprecipitation.
proregion is cleaved earlier. By contrast, removal of the NH,- terminal proregion of PC2 (76 to 64 kDa) was blocked a t 20 "C. Much of the smaller PC2 produced at 37 "C is secreted during the chase, while there is no secretion a t 20 "C. Combining the cellular and secreted forms of PC2 and proPC2, about half the proPC2 is cleaved to PC2 in 3 h a t 37 "C, but there is virtually no cleavage a t 20 "C, suggesting that this cleavage is a post- TGN event. Thus, a 20 "C temperature block clearly distin-
guishes the cleavage of proPC1 and proPC2. Both PC1 and PC2 have three potential sites for N-glycosyl-
ation (Bloomquist et al., 1991), and oligosaccharide chain matu- ration can be used to determine when newly synthesized pro- teins reach the medial Golgi. To examine the glycosylation status of PC1 and PC2 in wild-type, sPCl and sPC2 cells, cell extracts from 15-min pulse labeling, and 1.5-h chase incuba- tions were first immunoprecipitated with PC1 or PC2 antisera
17444 Endoproteolysis i n Cells Overexpressing PC1 or PC2
15 min pulse
kDa Ctr. N-gly. Endo-H Ctr. N-gly. Endo-H
a7
a1 a4
wt
SPCl 63
76 - 64-
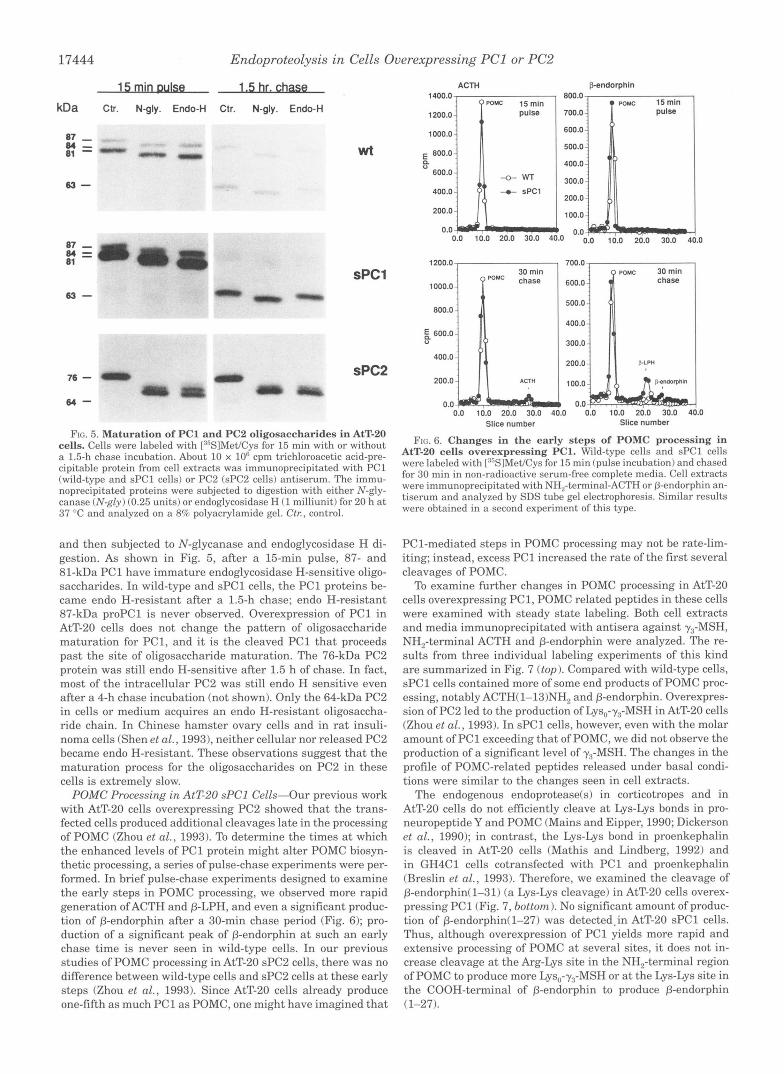
FIG. 5. Maturation of PC1 and PC2 oligosaccharides in AtT-20 cells. Cells were labeled with [:"SIMet/Cys for 15 min with or without a 1.5-h chase incubation. About 10 x lo6 cpm trichloroacetic acid-pre- cipitable protein from cell extracts was immunoprecipitated with PC1 (wild-type and sPCl cells) or PC2 (sPC2 cells) antiserum. The immu- noprecipitated proteins were subjected to digestion with either N-gly- canase (N-gly) (0.25 units) or endoglycosidase H (1 milliunit) for 20 h a t 37 "C and analyzed on a 8% polyacrylamide gel. Ctr, control.
and then subjected to N-glycanase and endoglycosidase H di- gestion. As shown in Fig. 5 , after a 15-min pulse, 87- and 81-kDa PC1 have immature endoglycosidase H-sensitive oligo- saccharides. In wild-type and sPCl cells, the PC1 proteins be- came endo H-resistant after a 1.5-h chase; endo H-resistant 87-kDa proPC1 is never observed. Overexpression of PC1 in AtT-20 cells does not change the pattern of oligosaccharide maturation for PC1, and it is the cleaved PC1 that proceeds past the site of oligosaccharide maturation. The 76-kDa PC2 protein was still endo H-sensitive after 1.5 h of chase. In fact, most of the intracellular PC2 was still endo H sensitive even after a 4-h chase incubation (not shown). Only the 64-kDa PC2 in cells or medium acquires an endo H-resistant oligosaccha- ride chain. In Chinese hamster ovary cells and in rat insuli- noma cells (Shen et al., 19931, neither cellular nor released PC2 became endo H-resistant. These observations suggest that the maturation process for the oligosaccharides on PC2 in these cells is extremely slow.
POMC Processing in AtT-20 sPCl Cells-Our previous work with AtT-20 cells overexpressing PC2 showed that the trans- fected cells produced additional cleavages late in the processing of POMC (Zhou et al., 1993). To determine the times at which the enhanced levels of PC1 protein might alter POMC biosyn- thetic processing, a series of pulse-chase experiments were per- formed. In brief pulse-chase experiments designed to examine the early steps in POMC processing, we observed more rapid generation ofACTH and P-LPH, and even a significant produc- tion of 0-endorphin after a 30-min chase period (Fig. 6); pro- duction of a significant peak of 0-endorphin a t such an early chase time is never seen in wild-type cells. In our previous studies of POMC processing in AtT-20 sPC2 cells, there was no difference between wild-type cells and sPC2 cells a t these early steps (Zhou et al., 1993). Since AtT-20 cells already produce one-fifth as much PC1 as POMC, one might have imagined that
1400.0
1200.0
1000.0
ACTH pendorphln 800.0
600.0
500.0
400.0 E 800.0
600.0
400.0 -0- SPCl
200.0
200.0
100.0
0.0 0.0 0.0 10.0 20.0 30.0 40.0 0.0 10.0 20.0 30.0 40.0
0.0 10.0 20.0 30.0 40.0 0.0 10.0 20.0 30.0 4 Slice number Slice number
.O
AtT-20 cells overexpressing PC1. Wild-type cells and sPCl cells FIG. 6. Changes in the early steps of POMC processing in
were labeled with I:"SIMet/Cys for 15 min (pulse incubation) and chased for 30 min in non-radioactive serum-free complete media. Cell extracts were immunoprecipitated with NH,-terminal-ACTH or P-endorphin an- tiserum and analyzed by SDS tube gel electrophoresis. Similar results were obtained in a second experiment of this type.
PC1-mediated steps in POMC processing may not be rate-lim- iting; instead, excess PC1 increased the rate of the first several cleavages of POMC.
To examine further changes in POMC processing in AtT-20 cells overexpressing PC1, POMC related peptides in these cells were examined with steady state labeling. Both cell extracts and media immunoprecipitated with antisera against y3-MSH, NH,-terminal ACTH and p-endorphin were analyzed. The re- sults from three individual labeling experiments of this kind are summarized in Fig. 7 (top). Compared with wild-type cells, sPCl cells contained more of some end products of POMC proc- essing, notably ACTH(1-13)NH2 and P-endorphin. Overexpres- sion of PC2 led to the production of Lys,-y,-MSH in AtT-20 cells (Zhou et al., 1993). In sPCl cells, however, even with the molar amount of PC1 exceeding that of POMC, we did not observe the production of a significant level of y3-MSH. The changes in the profile of POMC-related peptides released under basal condi- tions were similar to the changes seen in cell extracts.
The endogenous endoprotease(s) in corticotropes and in AtT-20 cells do not efficiently cleave a t Lys-Lys bonds in pro- neuropeptide Y and POMC (Mains and Eipper, 1990; Dickerson et al., 1990); in contrast, the Lys-Lys bond in proenkephalin is cleaved in AtT-20 cells (Mathis and Lindberg, 1992) and in GH4C1 cells cotransfected with PC1 and proenkephalin (Breslin et al., 1993). Therefore, we examined the cleavage of P-endorphin(1-31) (a Lys-Lys cleavage) in AtT-20 cells overex- pressing PC1 (Fig. 7, bottom). No significant amount of produc- tion of 0-endorphin(1-27) was detected.in AtT-20 sPCl cells. Thus, although overexpression of PC1 yields more rapid and extensive processing of POMC a t several sites, it does not in- crease cleavage at the Arg-Lys site in the NH,-terminal region of POMC to produce more Lys,-y,-MSH or at the Lys-Lys site in the COOH-terminal of P-endorphin to produce 0-endorphin (1-27),
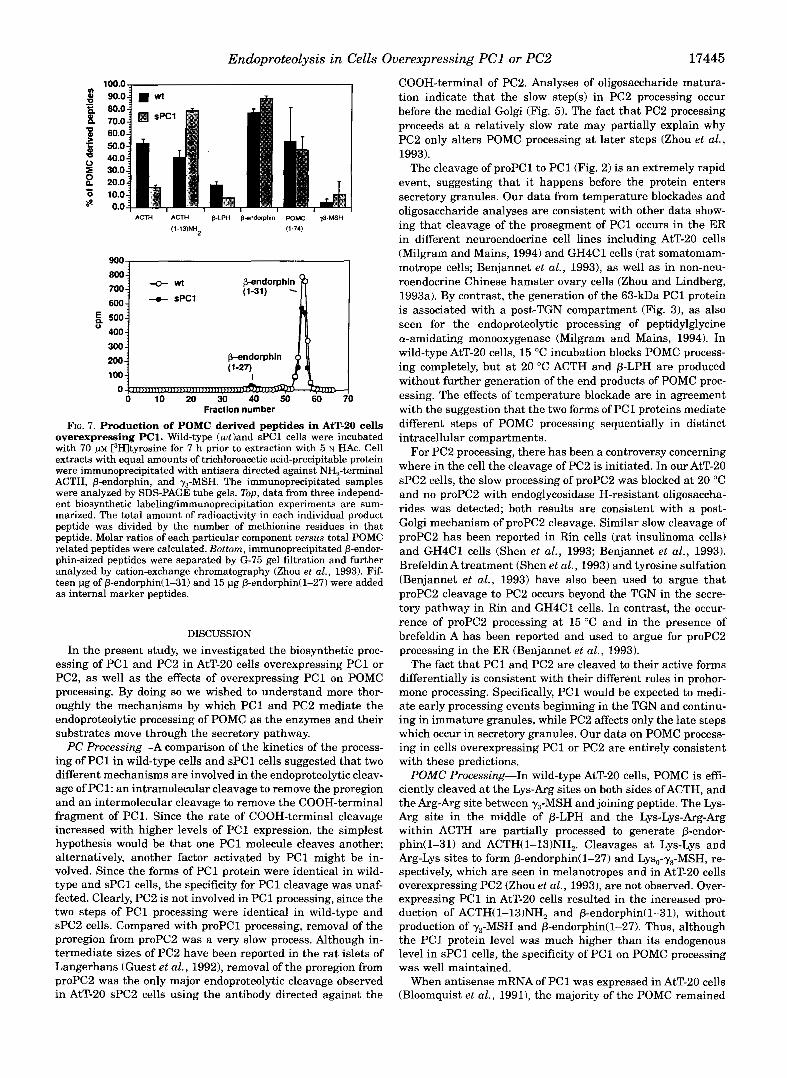
Endoproteolysis in Cells Overexpressing PC1 or PC2 17445
100.0 ~ I
ACTH ACTH B.LPH B-endorphin Pow p-MSH (1-13)NH (1.74)
"_
0 10 20 30 40 50 60 Fractlon number
FIG. 7. Production of POMC derived peptides in AtT-20 cells overexpressing PC1. Wild-type (cotland sPCl cells were incubated with 70 [3Hltyrosine for 7 h prior to extraction with 5 N H A C . Cell extracts with equal amounts of trichloroacetic acid-precipitable protein
ACTH, P-endorphin, and y3-MSH. The immunoprecipitated samples were immunoprecipitated with antisera directed against NH,-terminal
were analyzed by SDS-PAGE tube gels. Top, data from three independ- ent biosynthetic labelinglimmunoprecipitation experiments are sum- marized. The total amount of radioactivity in each individual product peptide was divided by the number of methionine residues in that peptide. Molar ratios of each particular component uersus total POMC related peptides were calculated. Bottom, immunoprecipitated p-endor- phin-sized peptides were separated by G-75 gel filtration and further analyzed by cation-exchange chromatography (Zhou et al., 1993). Fif- teen pg of P-endorphin(1-31) and 15 pg P-endorphin(1-27) were added as internal marker peptides.
DISCUSSION
In the present study, we investigated the biosynthetic proc- essing of PC1 and PC2 in AtT-20 cells overexpressing PC1 or PC2, as well as the effects of overexpressing PC1 on POMC processing. By doing so we wished to understand more thor- oughly the mechanisms by which PC1 and PC2 mediate the endoproteolytic processing of POMC as the enzymes and their substrates move through the secretory pathway.
PC Processing-A comparison of the kinetics of the process- ing of PC1 in wild-type cells and sPCl cells suggested that two different mechanisms are involved in the endoproteolytic cleav- age of PC1: an intramolecular cleavage to remove the proregion and an intermolecular cleavage to remove the COOH-terminal fragment of PC1. Since the rate of COOH-terminal cleavage increased with higher levels of PC1 expression, the simplest hypothesis would be that one PC1 molecule cleaves another; alternatively, another factor activated by PC1 might be in- volved. Since the forms of PC1 protein were identical in wild- type and sPCl cells, the specificity for PC1 cleavage was unaf- fected. Clearly, PC2 is not involved in PC1 processing, since the two steps of PC1 processing were identical in wild-type and sPC2 cells. Compared with proPC1 processing, removal of the proregion from proPC2 was a very slow process. Although in- termediate sizes of PC2 have been reported in the rat islets of Langerhans (Guest et al., 1992), removal of the proregion from proPC2 was the only major endoproteolytic cleavage observed in AtT-20 sPC2 cells using the antibody directed against the
COOH-terminal of PC2. Analyses of oligosaccharide matura- tion indicate that the slow step(s) in PC2 processing occur before the medial Golgi (Fig. 5). The fact that PC2 processing proceeds at a relatively slow rate may partially explain why PC2 only alters POMC processing at later steps (Zhou et al., 1993).
The cleavage of proPC1 to PC1 (Fig. 2) is an extremely rapid event, suggesting that it happens before the protein enters secretory granules. Our data from temperature blockades and oligosaccharide analyses are consistent with other data show- ing that cleavage of the prosegment of PC1 occurs in the ER in different neuroendocrine cell lines including AtT-20 cells (Milgram and Mains, 1994) and GH4C1 cells (rat somatomam- motrope cells; Benjannet et al., 19931, as well as in non-neu- roendocrine Chinese hamster ovary cells (Zhou and Lindberg, 1993a). By contrast, the generation of the 63-kDa PC1 protein is associated with a post-TGN compartment (Fig. 31, as also seen for the endoproteolytic processing of peptidylglycine a-amidating monooxygenase (Milgram and Mains, 1994). In wild-type AtT-20 cells, 15 "C incubation blocks POMC process- ing completely, but at 20 "C ACTH and p-LPH are produced without further generation of the end products of POMC proc- essing. The effects of temperature blockade are in agreement with the suggestion that the two forms of PC1 proteins mediate different steps of POMC processing sequentially in distinct intracellular compartments.
For PC2 processing, there has been a controversy concerning where in the cell the cleavage of PC2 is initiated. In our AtT-20 sPC2 cells, the slow processing of proPC2 was blocked at 20 "C and no proPC2 with endoglycosidase H-resistant oligosaccha- rides was detected; both results are consistent with a post- Golgi mechanism of proPC2 cleavage. Similar slow cleavage of proPC2 has been reported in Rin cells (rat insulinoma cells) and GH4C1 cells (Shen et al., 1993; Benjannet et al., 1993). Brefeldin A treatment (Shen et al., 1993) and tyrosine sulfation (Benjannet et al., 1993) have also been used to argue that proPC2 cleavage to PC2 occurs beyond the TGN in the secre- tory pathway in Rin and GH4C1 cells. In contrast, the occur- rence of proPC2 processing a t 15 "C and in the presence of brefeldin A has been reported and used to argue for proPC2 processing in the ER (Benjannet et al., 1993).
The fact that PC1 and PC2 are cleaved to their active forms differentially is consistent with their different roles in prohor- mone processing. Specifically, PC1 would be expected to medi- ate early processing events beginning in the TGN and continu- ing in immature granules, while PC2 affects only the late steps which occur in secretory granules. Our data on POMC process- ing in cells overexpressing PC1 or PC2 are entirely consistent with these predictions.
POMC Processing-In wild-type AtT-20 cells, POMC is effi- ciently cleaved at the Lys-Arg sites on both sides ofACTH, and the Arg-Arg site between y3-MSH and joining peptide. The Lys- Arg site in the middle of p-LPH and the Lys-Lys-Arg-Arg within ACTH are partially processed to generate p-endor- phin(1-31) and ACTH(1-13)NH2. Cleavages at Lys-Lys and Arg-Lys sites to form p-endorphin(1-27) and Lys,-y,-MSH, re- spectively, which are seen in melanotropes and in AtT-20 cells overexpressing PC2 (Zhou et al., 19931, are not observed. Over- expressing PC1 in AtT-20 cells resulted in the increased pro- duction of ACTH(1-13)NH2 and p-endorphin( 1-31), without production of y3-MSH and p-endorphin( 1-27). Thus, although the PC1 protein level was much higher than its endogenous level in sPCl cells, the specificity of PC1 on POMC processing was well maintained.
When antisense mRNA of PC1 was expressed in AtT-20 cells (Bloomquist et al., 1991), the majority of the POMC remained

17446 Endoproteolysis in Cells Overexpressing PC1 or PC2
u 20° block - 1 W
unknown
A cleavage enhanced by PC1 * cleavage enhanced by PC2
FIG. 8. Biosynthetic processing of PC1 and PC2 and their roles in POMC processing. TFiungles (cleavage enhanced by PC1) and us- terisks (cleavage enhanced by PC2) are located in the sites where the endoproteolytic cleavages initiate. IMG, immature granule; MG, ma- ture granule.
intact, indicating that the initial cleavage step of POMC proc- essing is mediated by PC1. Our pulse-chase experiments on sPCl cells at short times extended these observations, showing increased generation ofACTH, P-LPH, and P-endorphin during the early stages in POMC biosynthetic processing. Such changes at early times of biosynthetic processing were not seen in sPC2 cells. When chase times were extended, increased cleavages ofACTH and 0-LPH to release ACTH(1-13)NH2 and P-endorphin in sPCl cells were observed, as in sPC2 cells. Putting together the kinetic data on PC1 and POMC process- ing, the increased cleavages of ACTH and P-LPH were well correlated with the accelerated conversion of 84-kDa PC1 to 63-kDa PC1 in sPCl cells. Both forms of PC1 protein are active in cleaving synthetic substrate (Zhou and Lindberg, 1993a; Jean et al., 19931, although they have different pH and calcium optima, with the 63-kDa PC1 protein favoring a more acidic environment and a higher calcium concentration (Zhou and Lindberg, 1993b). Thus the two PC1 proteins may mediate sequentially the processing of POMC, as PC1 and POMC mi- grate through secretory granule pathway. PCIPOMC Quantitation-Wild-type AtT-20 cells produce 1
molecule of PC1 for every 5 molecules of POMC. This result was quite unexpected, since the molar ratio of peptidylglycine cw-amidating monooxygenase to POMC was previously estab- lished to be about 1:3000 (Mains and Eipper, 1990). The high molar ratio of endogenous PC1 to POMC has important impli- cations. First, PC1 must exhibit a great deal of specificity, since even a slow rate of aberrant cleavage would result in substan- tial loss of POMC peptides. Second, the turnover number of PC1 need not be very high to perform the required cleavages in the time available. Third, the very high endogenous levels of PC1 raise the possibility that PC1 may have additional roles, perhaps serving a binding function, as suggested for the y-en- doprotease involved in nerve growth factor processing (Brad-
shaw et al., 1990). Fourth, since PC1 expression is normally so high, the expression of antisense RNA to PC1 would be ex- pected to alter POMC processing only if PC1 mRNA were largely eliminated (Bloomquist et al., 1991). By comparison, since PC2 expression in wild-type cells is so low, it was expected that overexpressing PC2 could have dramatic effects on POMC processing, changing the final end products produced (Zhou et al., 1993). Both these predictions were found to be true.
Final Summary-Fig. 8 summarizes our studies on the bio- synthetic processing of PC1 and PC2 in AtT-20 cells and their distinct roles on POMC processing. Starting from ER, proPC1 is first cleaved into the 81-kDa PC1 protein while PC2 and POMC remain intact. As proteins migrate from ER through the Golgi, a process blocked at 15 "C, the first two steps of POMC processing are initiated by PC1, with the consequent produc- tion ofACTH and P-LPH (Milgram and Mains, 1994). Next, as PC1, PC2, and POMC exit the TGN and enter secretory gran- ules, which can be prevented by incubation at 20 "C, the second step of PC1 processing takes place. The cleavage of proPC2 is also initiated. It is not clear at this point which endoprotease(s) might be involved in proPC2 processing. Two cleavages of POMC which release ACTH(1-13)NH2 and P-endorphin(1-31) are mediated through either PC1 or PC21 When secretory gran- ules become mature the last two steps of POMC processing specifically mediated by PC2 are accomplished, resulting in the production of Lys,-7,-MSH and P-endorphin(1-27). Further de- tailed investigation on intracellular sorting and function of PC1 and PC2 in these cells and the abilities of mutant PC1 and PC2 to process POMC will reveal more of the mechanisms by which POMC is processed by PCs.
Acknowledgments-We thank Betty Eipper and Sharon Milgram for many helpful discussions, Iris Lindberg for the PC2 antigen and an initial supply of PC2 antibody, Brian Bloomquist for PC1 and PC2 cDNAs, Rich Johnson for help constructing expression vectors, Carla Berard for help with cell culture, Marie Bell for general laboratory assistance, and Zina Garrett for laboratory administrative management.
REFERENCES
Benjannet, S., Rondeau, N. Day, R., Chrbtien, M., and Seidah, N. G. (1991) Proc.
Benjannet, S., Rondeau, N., Paquet, L., Boudreault, A., Lazure, C., ChrBtien, M.,
Bloomquist, B. T., Eipper, B. A,, and Mains, R. E. (1991) Mol. Endocrinol. 5,
Bloomquist, B. T., and Mains, R. E. (1993) Cell Physiol. Biochem. 3, 197-212 Bradshaw, R. A., Altin, J. G., Blaber, M., Cavanaugh, K. P., Eveleth, D. D., Korn-
blum, H. I., Leslie, F. M., and Raffloni, S. (1990) Prog. Bruin Res. 86, 157-167 Breslin, M. B., Lindberg, I., Benjannet, S., Mathis, J. P., Lazure, C., and Seidah, N.
G. (1993) J. Biol. Chem. 268, 27084-27093 Creemers, J. W. M., Siezen, R. J., Roebroek, A. J. M., Ayoubi, T. A. Y., Huylebroeck,
D., and Van de Ven, W. J. M. (1993) J. Bid. Chem. 268,21826-21834 Day, R., Schafer, M. K.-H., and Watson, S. J. (1992) Mol. Endocrinol. 6, 485-497 Dickerson, I. M., Dixon, J. E., and Mains, R. E. (1990) J. Biol. Chem. 265,2462-
Guest, P. C., Arden, S. D., Bennett, D. L., Clark, A,, Rutherford, N. G., and Hutton
Hakes, D. J., Birch, N. P., Mezey, A,, and Dixon, J. E. (1991) Endocrinology 129,
Harlow, E., and Lane, E. (1988) Antibodies, p. 447, Cold Spring Harbor Laboratory,
Husten, E. J., and Eipper, B. A. (1991) J. Biol. Chem. 266,17004-17010 Jean, F., Basak, A,, Rondeau, N., Benjannet, S., Hendy, G. N., Seidah, N. G.,
Leduc, R., Molloy, S. S., Thorne, B. A,, and Thomas, G. (1992) J. Biol. Chem. 267,
Natl. Acud. Sci. U. S. A. 88, 3564-3568
and Seidah, G. (1993) Biochem. J. 294,735-743
2014-2024
2469
J. C. (1992) J. B i d . Chem. 267,22401-22406
3053-3063
Cold Spring Harbor, NY
ChrBtien, M., and Lazure, C. (1993) Biochem. J. 292,891-900
1 A . ? n b l A908 Lindberg, I. (1991) Mol. Endocrinol. 5, 1361-1365 Mains, R. E., and Eipper, B. A. (1990) Dends Endocrinol. Metub. 1,388394 Mathis. J. P.. and Lindbere. I. (1992) Endocrinolom 131. 2287-2296
"" "I"
Matthews, G:, Shennan, K-I. J.,'Seal, A. J., Taylor,:. A,, dolman, A,, and Docherty,
Milgram, S. L., Mains, R. E., and Eipper, B. A. (1993) J. Cell B i d . 121, 2336 Milgram, S. L., and Mains, R. E. (1994) J. Cell Sci. 107, 737-745 Noel, G., Keutmann, H. T., and Mains, R. E. (1991) Mol. Endocrinol. 5,404413 Rhodes, C. J., Thorne, B. A., Lincoln, B., Nielsen, E., Hutton, J. C., and Thomas, G.
Rothman, J. E., and Orci, L. (1992) Nature 355, 409415
K. (1994) J. B i d . Chem. 269,588-592
(1993) J. B i d . Chem. 268,42674275

Endoproteolysis in Cells Overexpressing PC1 or PC2 17447 Seidah, N. G., Gaspar, L., Mion, P., Marcinkiewicz, M., Mbikay, M., and Chretien,
Seidah, N. G, Fournier, H., Boileau, G., Benjannet, S., Rondeau, N., and Chretien,
Seidah, N. G., Day, R., and Chretien, M. (1993) Biochem. SOC. Duns. 21.685491 Shen, F.-S., Seidah, N. G., and Lindberg, I. (1993)J. Biol. Chem. 268,24910-24915 Shennan, K. I. J., Sea1,A. J., Smeekens, S. P., Steiner, D. F., and Docherty, K. (1991)
Shennan, K. I. J., Smeekens, S. P., Steiner, D. F., and Docherty, K. (1991) FEES
Shinde, U., Li, Y., Chattejee, S., and Inouye, M. (1993) Proc. Natl. Acad. Sci.
M. (1990) DNA Cell Eiol. 9,415-424
M. (1992) FEES Lett 310,235-239
J. Biol. Chem. 266,24011-24017
Lett 284,277-280
U. S . A. Bo, 69244928 Smeekens, S. P. (1993) Bionchnology 11, 182-186 Thomas, L., Leduc, R., Thorne, B. A,, Smeekens, S. P., Steiner, D. F., and Thomas,
Thorne, B. A., and Thomas, G. (1990) J. Eiol. Chem. 265,84368443 Vindrola, O., and Lindberg, I. (1992) Mol. Endocrinol. 6, 1088-1094 Young, R. W. and Fulhorst, H. W. (1965)Anal. Eiochem. 11,389491 Zhou, A,, Bloomquist, B. T. and Mains, R. E. (1993) J. Eiol. Chem. 268,1763-1769 Zhou, Y., and Lindberg, I. (1993a) J. Biol. Chem. 268,5615-5623 Zhou, Y., and Lindberg, I. (1993b)Annual Meeting ofthe Society for Neuroscience,
G. (1991) Proc. Natl. Acad. Sei. U. S. A. 88, 5297-5301
Abstr. 559.11, Nov. 7-12, Washington, D. C.





























