Embed Size (px)
Citation preview

THE JOURNAL OF BIOL~CICAL CHEMISTRY 0 1994 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 269, No. 7, Issue of February 18. pp. 5440-5445, 1994 Printed in U.S.A.
Processing of Human Acid Sphingomyelinase in Normal and I-cell Fibroblasts*
(Received for publication, September 14, 1993, and in revised form, October 29, 1993)
Robert Hurwitz, Klaus Ferlinz, Gabriele Vlelhaber, Heidi Moczall, and Konrad SandhofM: From the Znstitut fur Organische Chemie und Biochemie, Gerhard-Domagk-Strape 1, 0-53121 Bonn, Germany
The biosynthesis of acid sphingomyelinase in normal and I-cell disease fibroblasts was investigated by meta- bolic labeling with [35Slmethionine and immunoprecipi- tation followed by polyacrylamide gel electrophoresis and fluorography. Two major polypeptides with appar- ent molecular masses of 75 and 72 kDa (peptide chains of 64 and 61 kDa, respectively) and a minor one with mo- lecular mass of 57 kDa (peptide chain of 47 kDa) were found intracellularly soon after pulse labeling. The 75- kDa form is assumed to be the propropolypeptide of sphingomyelinase which is converted into the precursor form of 72 kDa. The precursor is subjected to two dis- tinct processing events. A minor part is already cleaved in the endoplasmic reticulum-Golgi complex yielding the P-endo-N-acetylglucosaminidase H-resistant form of 57 kDa; whereas, the major part of the precursor is proc- essed within 4 h to a 70-kDa mature P-endo-N-acetylglu- cosaminidase H-sensitive form of sphingomyelinase, which is subsequently converted into a polypeptide with molecular mass of 52 kDa within a chase of about 20 h. Both the precursor (72 kDa) as well as its early cleavage product of 57 kDa are secreted into the culture medium to a minor extent. Intracellular transport of sphingomy- elinase into lysosomes depends on the phosphomanno- syl specific receptor by following criteria: (i) about 80% of newly synthesized precursor was secreted in NH4Cl- treated fibroblasts as well as in I-cells, (ii) the matura- tion of sphingomyelinase was inhibited in normal fibro- blasts exposed to NH&l as well as in I-cell fibroblasts, and (iii) the [32Plphosphate associated with oligosaccha- rides was cleavable by P-endo-N-acetylglucosaminidase H. However, endocytosis of radiolabeled extracellular precursor by fibroblasts was not prevented by the addi- tion of mannose &phosphate, whereas uptake of arylsul- fatase A and P-hexosaminidase was almost completely blocked under these conditions. This indicates that en- docytosis of acid sphingomyelinase by cultured fibro- blasts might be mediated by an alternative pathway.
Acid sphingomyelinase (ASM)l (EC 3.1.4.12) is a lysosomal enzyme catalyzing the breakdown of sphingomyelin to ce- ramide and phosphorylcholine. Deficiency of this enzyme leads
* This work was supported by Deutsche Forschungsgemeinschaft Grant DFG 257/12-3. The costs of publication of this article were de- frayed in part by the payment of page charges. This article must there- fore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
hoff, Institut fur Organische Chemie und Biochemie, Gerhard-Domagk f To whom correspondence should be addressed: Prof. Dr. K. Sand-
Strape 1, D-53121 Bonn, Germany. Tel.: 49-228-73-7778; Fax: 49-228- 73-7778.
brefeldin A , Endo H, p-endo-N-acetylglucosaminidase H; MEM, mini- The abbreviations used are: ASM, acid sphingomyelinase; BFA,
mal essential medium; Man-6-P, mannose 6-phosphate; NPD, Ni- emann-Pick disease; PAGE, polyacrylamide gel electrophoresis; PMSF, phenylmethylsulfonyl fluoride; LDL, low density lipoprotein.
to lysosomal accumulation of sphingomyelin in patients suffer- ing from Niemann-Pick disease (NPD) (Brady et al., 1966; Spence and Callahan, 1989).
In 1987, we succeeded in the isolation of ASM from human urine. The purification procedure yielded a homogeneous preparation with an apparent molecular mass of about 70 kDa and with the highest ASM activity reported so far (Quintern et al., 1987). The pure enzyme also cleaved phosphatidylcholine in a phospholipase C-like manner, consistent with previous findings that this activity is deficient in fibroblasts from NPD (Beaudet et al., 1980). Isolation of the cDNA (Quintern et al., 1989) and characterization of genomic sequences encoding ASM (Schuchman et al., 1991,1992) has enabled investigations of the molecular lesions causing NPD (Levran et al., 1991a, 1991b; Ferlinz et al., 1992; Takahashi et al., 1992). To date, however, the known mutations responsible for NPD have not led to identification of the active site nor to an understanding of the cellular events necessary for transport and maturation of this enzyme.
Our interest has focused on processing and targeting of ly- sosomal ASM. For this purpose, we used fibroblasts from pa- tients with I-cell disease (mucolipidosis II), an inherited auto- somal neurometabolic disorder, as a model for studying the function of mannose 6-phosphate residues (Man-6-P) in this sorting process. I-cell fibroblasts fail to phosphorylate mannose residues on newly synthesized lysosomal enzymes, resulting in secretion of large quantities of many acid hydrolases into the culture medium instead of delivery to the lysosomes (Leroy et al., 1972; Hasilik et al., 1981). In these cells, a reduced level of cellular ASM activity was observed (Wenger et al., 1976; Weitz et al., 19831, but the fate and localization of the intracellular retained enzyme in I-cell fibroblasts has not yet been analyzed.
EXPERIMENTAL PROCEDURES Materzals-~-[~~SlMethionine (>lo00 Cilmmol), [3ZPlphosphate
(>go00 Cilmmol), and methionine-deficient minimal essential medium (MEM) were purchased from ICN. 14C-Methylated molecular weight standards were from DuPont NEN; Amplify was fromhersham Corp.; protease inhibitors, endo-6-N-acetylglucosaminidase H, peptide N-gly- canase F, fetal calf serum, and brefeldin A were from Boehringer Mann- heim; protein A-Sepharose CL-4B was from Sigma. Rabbit polyclonal antibody against human arylsulfatase A was a kind gift from Dr. P. Lemansky (Institute of Cell Biology, University of Bonn, Bonn, Ger- many). All other chemicals were of reagent grade and were purchased from various commercial sources.
Cell Culture-Diploid human skin fibroblasts were grown in mono- layers to confluence at 37 “C in 5 % CO, in Eagle’s MEM supplemented with antibiotics, nonessential amino acids, and 10% fetal calf serum, as described previously (Weitz et al., 1983).
Fibroblasts from a patient with mucolipidosis I1 (I-cell disease) were from biopsies submitted to us for diagnosis.
Enzyme Assays-Activities of acid sphingomyelinase and that of phospholipase C were measured with sphingomyelin and phosphatidyl- choline as substrates (Quintern et al., 1987), which were tritium-labeled in the choline moiety, each.
Expression of ASM Protein in Escherichia coli and Immunization- “he full-length cDNA of ASM, except the portion containing the signal
5440

Dansport and Maturation of Human Acid Sphingomyelinase 5441
peptide, was cloned into the prokaryotic expression vector pQE. Expres- sion of the corresponding cDNA construct yielded a fusion protein con- sisting of six N-terminal histidine residues followed by 2 additional amino acids derived from sequences of cloning site and the ASM amino acid sequence, representing the mature protein from residues 47-629. Purification of the fusion protein was achieved by nickel ion chelating chromatography (Hochuli et al., 1988). A rabbit polyclonal anti-ASM antiserum was raised by initial injection of 100 pg of protein in Freund's complete adjuvant followed by three injections of 50 pg of protein in Freund's incomplete adjuvant given in 4-week intervals. Animals were bled 10 days after the final injection.
m n i t y Purification of Antibodies-Immobilization of recombinant ASM protein was performed with a N-hydroxysuccinimide-activated agarose support ("Gel 10, Bio-Rad) according to the manufacturer's instructions.
Antibody purification was accomplished by applying the crude anti- serum over the column, removal of unbound material by rinsing the column with RIPA buffer (10 mM Tris-HC1, pH 8.0, 170 m~ NaCl, 1% Nonidet P-40, 0.5% deoxycholate, 0.1% SDS), and finally eluting the bound antibodies with 3 M KSCN in 10 m~ Tris-HC1, pH 6,O. KSCN was removed by desalting the eluted immunoglobulin fractions on a Sepha- dex G25 column, equilibrated with 10 mM Tris-HC1, pH 7.4, 1 M NaCl, 0.1% Tween 20. Pooled IgG fractions were stored at -20 "C in aliquots. Activity of recovered antibodies was tested by enzyme-linked immuno- sorbent assay and immunoprecipitation methods.
Immunotitration ofAcid Sphingomyelinase-Fibroblasts extracts, as well as purified ASM from human urine (enriched over 10,000-fold by the procedure of Quintern et al., 19871, were diluted in Tris-buffered saline (TBS) containing 1% Nonidet P-40 and mixed with different amounts of either crude antiserum or affinity purified antibodies in a total of 1 ml. Preimmune serum IgG was used as a control. The samples were incubated at 4 "C for 4 h. After addition of 20 pl of a 50% suspen- sion (wh) of protein A-Sepharose in TBS, immunocomplexes were cen- trifuged. Activity of ASM and other hydrolases was measured in the supernatant. Determination of the affinity of anti-ASM immunoglobu- lins (I&) toward native ASM, in terms of the equilibrium constant K, was calculated by reacting various concentrations of ASM with a con- stant concentration of affinity-purified anti-ASM immunoglobulins (IgG). The equilibrium concentration of free ASM was determined by its enzyme activity, assuming a specific molecular activity for ASM of 2.5 mmol/h/mg. A value of K = lo9 M - ~ was estimated.
Metabolic Labeling and Immunoprecipitation-Confluent cell cul- ture flasks (25 cm*) were starved for 2 h in methionine-free medium (MEM) containing 4% dialyzed and heat-inactivated fetal calf serum. Labeling was initiated by the addition of 0.1 mCi/ml of ~-1~~Slmeth io- nine. The chase periods were started by addition of unlabeled methio- nine at a final concentration of 3 m ~ . For labeling with [32Plphosphate, cells were incubated in MEM minus phosphate with 0.3% dialyzed heat-inactivated fetal calf serum and 0.2 mCi/ml [32Plphosphate for 4 h.
For some experiments, cells were treated with either 1 pg/ml of brefeldin A or 10 m~ NH&l beginning 1 h before [35Slmethionine la- beling and continuing through labeling and chase periods.
The labeled fibroblasts were rinsed once with TBS, harvested by scrapping, and lysed in 0.5 ml of TBS containing 1% Nonidet P-40, 10 m~ EDTA, PMSF, pepstatin, and leupeptin (1 m~ each). Sodium fluo- ride and orthovanadate were added in extraction buffers for phosphate- labeled cells a t final concentrations of 5 and 1 m ~ , respectively.
Cell extracts and media were preabsorbed with 20 pl of rabbit pre- immune serum and 40 pl of a 50% suspension (w/v) of protein A-Sepha- rose for 4 h. Immunocomplexes were removed by centrifugation at 520 x g. The preabsorbed homogenates and media were centrifuged at 100,000 x g for 1 h in a Beckman Ti 75 rotor. The supernatants were carefully removed, 2 p1 of crude anti-ASM antiserum was added to the supernatants and incubated for 4 h at 4 "C. Immunoprecipitation was camed out in the presence of 15 pl of protein A-Sepharose (50% sus- pension) for 2 h. Immunocomplexes were treated as described previ- ously (Schnabel et al., 1992).
Digestion with Glycosidases-For p-endo-N-actylglucosaminidase H (Endo H) digestion, the isolated immunocomplexes were solubilized by heating for 3 min at 95 "C in 20 pl of 1% SDS, 10 m~ p-mercaptoethanol, 50 m~ acetate buffer, pH 5.0, and incubated with 2 milliunits of Endo H for 16 h at 37 "C. This incubation mixture was used directly for SDS- PAGE after addition of glycerol. Digestion with peptide-N-glycanase F (50 milliunits) was camed out with SDS-denaturated immunocom- plexes, which had been diluted in 300 pl of 50 mM phosphate buffer, pH 7.2, containing 1% Nonidet P-40. Following incubation, the mixture was precipitated by adding 1 ml of 10% trichloroacetic acid (w/v). The pellet was washed with acetone and treated with sample buffer (65 m~ Tris-
HCI, pH 6.7, 2% SDS, 20 % (v/v) glycerol) for 3 min at 95 "C prior to
Endocytosis-Confluent human skin fibroblasts were labeled in 25- cm2 culture flasks with [35S]methionine in the presence of 10 m~ NH,Cl for 12 h. The radiolabeled culture medium was dialyzed extensively against MEM and added to the medium of unlabeled fibroblasts for 12 h in the presence or absence of 10 m~ mannose 6-phosphate. For im- munoprecipitation, the cell extracts were treated as described above.
RESULTS
SDS-PAGE.
Characterization of Anti-sphingomyelinase Antibodies-The affinity of the polyclonal antibodies toward ASM was assessed using immunotitration experiments. Homogenates of fibro- blasts as well as ASM preparations from human urine, en- riched over 10,000-fold, served as antigen source. Less than 3 pl of the crude immune serum was able to precipitate about 90% of ASM activity in a total of 1 ml of nonionic detergent- containing buffer. Enzyme activities of the main contaminating hydrolases (e.g. acid ceramidase, hexosaminidase, and acid phosphatase) were not precipitated. The affinity of the purified immunoglobulins toward native ASM was calculated to be about lo9 M - ~ , as described under "Experimental Procedures." Besides ASM, the activity of phospholipase C toward phospha- tidylcholine was precipitated from purified ASM to the same extent.
Processing and Glycosylation of A S M in Normal Fibro- blasts-Immunoprecipitates from human skin fibroblasts, la- beled for 1 h with [35S]methionine, contained two major protein forms migrating as a double band in SDS-PAGE, corresponding to apparent molecular masses of 75 and 72 kDa, respectively. A polypeptide of 57 kDa was present in minor amounts account- ing for about 10% of total cross-reactive material (Fig. 1).
Replicate cultures chased with unlabeled methionine from 0.5 to 4 h processed ASM to an intracellular form of 70 kDa, analogous to the mass of the active ASM purified from human urine. This form was subsequently converted into a polypeptide of 52 kDa within 20 h, although small amounts of the 70-kDa form remained visible. No significant changes in molecular mass of the 57-kDa form were observed during these chase times.
During a 4-h chase period, immunoreactive cellular material was significantly reduced and only partially recovered from the culture medium as proteins with molecular masses of 75 and 57 kDa, respectively. After 48 h of chase, only trace amounts of a 52-kDa polypeptide could be visualized by fluorography (not shown). As demonstrated in Fig. 2, this intracellular degrada- tion was prevented by the addition of the fungal metabolite, brefeldin A, which is known to inhibit the regular vesicular transport through the Golgi apparatus (Klausner et al., 1992). Under these circumstances, the secretion is blocked, and ASM remains in its 72-75-kDa precursor form with almost undimin- ished intensity during the chase periods. Even the 57-kDa form was monitored after 8 h of chase, whereas the mature polypep- tides of 70 and 52 kDa could not be detected. These data indi- cate that degradation as well as conversion into the mature forms occur in the acidic compartment, except the proteolytic cleavage of the precursor (75 and/or 72 kDa) into the 57-kDa form.
In order to analyze the type of carbohydrate structures of the various ASM forms in normal fibroblasts, immunoprecipitated ASM, synthesized during a 4-h pulse with [35S]methionine, was treated with either N-glycanase F (peptide-N-glycanase F) or Endo H, as shown in Fig. 3. Peptide-N-glycanase F as well as Endo H treatment resulted in formation of a sharp double band corresponding to molecular masses of 64 and 61 kDa, obviously derived from the 75 , 72-, and 70-kDa forms, respectively. The 57-kDa form was reduced in size to 47 kDa and had acquired oligosaccharides of the complex type as judged by its resistance

5442 Bansport and Maturation of Human Acid Sphingomyelinase
FIG. 1. Biosynthesis of human ASM in normal and I-cell fibroblasts. Cells were pulsed with [""Slmethionine for 1 h and chased for the times indicated. ASM from the cell extracts and the media was precipitated using a polyclonal rabbit anti-ASM antibody. Immunoprecipitates were separated on a 10% SDS-PAGE and analyzed by fluorography. (A faint band at 45 kDa is caused by unspecific contami- nation of immunoprecipitates).
CELLS MEDIA
CONTROL I-CELL CONTROL I-CELL
C h a s e f h l O 4 0 1 2 4 0 1 2 0 1 2 4 8 2 0 1 2 4 8 2 0
kDa
J $ = r , 70
57 - 52 -
r-1 CELLS
lChare[h]l 0 4 8 ) 0 4 81 0 4 81
kDa kDa kDa
97-
69 - -64 " 6 1
57 - -47
45 -
30 - FIG. 2. Effect of brefeldin A and NH,Cl on the processing of
ASM. Normal fibroblasts were pulsed for 1 h with [3sSlmethionine and chased for the times indicated. Labeling was conducted in the presence of 1 pdml brefeldin A (BFA), 10 m~ NH,CI or without (control) addi- tions. A contaminant protein at 45 kDa (0-h chase, + NH,Cl) is visible.
toward Endo H (Fig. 3). The 70-kDa mature ASM, immunopre- cipitated after a chase of 4 h from normal fibroblasts that had been pulse labeled for 1 h with [35S]methionine, was reduced in size to 67 and 64 kDa upon deglycosylation with Endo H and revealed a polypeptide chain of 61 kDa when treated with pep- tide-N-glycanase F, as shown in Fig. 4. Removal of the Endo H-resistant carbohydrate structures of the extracellularly re- leased precursor (75 kDa) led to a peptide chain of 61 kDa (data not shown).
Processing, Glycosylation, and Tkansport of ASM in I-cell Fibroblasts-During a 1-h pulse with [35S]methionine, ASM was synthesized by I-cell fibroblasts as intracellular forms of 75 and 72 kDa, identical to normal cells (Fig. 1). The 57-kDa form was also present. Degradation occurred with a time depen- dence comparable with normal cells, but the loss of cellular immunoreactive material could be recovered from the culture medium, consistent with the finding that intracellular ASM activity in I-cell fibroblasts displays only 15% of the level of normal cells (Wenger et al., 1976; Weitz et al., 1983). More than 70% of total label in ASM was secreted as precursor with a slightly higher molecular mass of 78 kDa compared with the size of the secreted precursor from normal fibroblasts (75 kDa).
and IceU fibroblasts. Cells were pulsed for 4 h with [35Slmethionine. FIG. 3. N-Glycosylation of newly synthesized ASM in normal
Immunoprecipitated material was treated with either peptide-N-gly- canase F (PNGase F ) or with Endo H and analyzed by fluorography.
Chase
1Endo H I - - + I kDa kDa
70 - 61
4 67 - 64
FIG. 4. N-Glycosylation of mature ASM in normal fibroblasts. Fibroblasts were pulsed for 1 h with [3"S]methionine, and ASM was immunoprecipitated aRer a chase of 4 h and subjected to either peptide- N-glycanase F (PNGase F) or Endo H as described under"Experimenta1 Procedures." ER, endoplasmic reticulum.
The amount of extracellularly released 57-kDa form was not significantly increased over that from normal fibroblasts.
In contrast to normal cells, conversion into the mature form of 70 kDa as well as cleavage into the 52 kDa form was not observed, indicating that lysosomal sorting of ASM depends on the Man-6-P-mediated pathway.

Dansport and Maturation of Human Acid Sphingomyelinase 5443
100 4 1
h Control
v -
0 a 1 6 24 32 40 48
Chase Ihl
0 a 1 6 24 32 40 48
Chase Ihl
FIG. 5. Quantitation of ASM synthesis and secretion. Normal fibroblasts were pulse-labeled with [:'sSlmethionine for 1 h in the ab- sence (A) or presence of 10 mM NH,Cl ( B ) and chased for the indicated times. Aliquots of immunoprecipitated material from cell extracts and media were counted by liquid scintillation. Values are expressed as percent of totally immunoprecipitated countdmin from 1-h pulse (0-h chase = 100%) in cell extracts and media together ( 0 , in cells alone (W), and media alone (A).
The intracellularly retained forms ofASM revealed the pres- ence of oligosaccharide residues, predominantly of the complex type, as confirmed by their resistance toward Endo H (Fig. 3). Furthermore, deglycosylation with peptide-N-glycanase F re- sulted in the same band pattern (64 and 61 kDa) as in the case of normal cells. This indicates that the differences between the early forms of 75 and 72 kDa are due to proteolysis.
Effect of NH&l on Processing and Secretion of ASM- Similar to the experiments with I-cells, normal fibroblasts sub- jected to [35S]methionine labeling in the presence of NH4CI also failed to process the ASM precursor to the mature forms (Fig. 2). Under these conditions, secretion of ASM precursor re- sembled closely to that of I-cells (not shown). Weak bases are known to induce secretion of lysosomal precursor proteins tar- geted via the Man-6-P receptor system and prevent their matu- ration (Hasilik and Neufeld, 1980).
Fig. 5 shows the quantitation of immunoreactive material obtained from radiolabeled cell extracts and media during chase times. Curve A, representing the changes of the relative amount of totally immunoprecipitated ASM from 1-h pulse (chase: 0 h) during chase periods, demonstrates that a rapid loss of more than 60% of ASM during a 4-h chase period is
kDa
97 -
69 -
45 -
30 - FIG. 6. Phosphorylation of ASM in normal fibroblasts. Fibro-
blasts were metabolically labeled with ["Plphosphate for 4 h. Cell ex- tracts were subjected to immunoprecipitation using anti-ASM antibod- ies. One-half of the resulting precipitate was treated with Endo H, as described under "Experimental Procedures." Samples were applied to 10% SDS-PAGE and visualized by fluorography.
caused by intracellular degradation rather than by secretion. In normal cells treated with NH4CI (curve B ) , as well as in I-cell fibroblasts (not shown), the mean levels of extracellular ASM generated during 1-h pulse were at least %fold higher than in untreated fibroblasts. In this case, the loss of newly synthesized ASM can be explained by enhanced secretion into the medium.
Phosphorylation of ASM-A common feature of most soluble lysosomal enzymes is the presence of oligomannosyl linked Man-6-P residues. To define the phosphorylation status ofASM forms, fibroblasts were metabolically labeled with [32Plphos- phate (Fig. 6). A pulse of 4 h resulted in formation of labeled intracellular form of 70 kDa, whereas the 57-kDa form could not be monitored. The absence of the 52-kDa late processed form might be due to the short pulse time, insufficient for the formation of this species. The incorporated [32P]phosphate was mainly associated with carbohydrate. Endo H digestion re- moved about 80% of the phosphate label and reduced the size of the 70-kDa form by about 10 kDa.
Endocytosis of ASM-The dialyzed culture medium from fi- broblasts that had been labeled with [35S]methionine in the presence of NH4CI was added to unlabeled fibroblasts. After incubation for 12 h in the presence or absence of Man-6-P, the recipient cells were analyzed for endocytosed ASM (Fig. 7). About 5% of radiolabeled ASM was internalized by the cells and processed to the mature form of 70 and 52 kDa, respectively. Surprisingly, endocytosis was not affected by Man-6-P. The 57- kDa form of ASM could not be monitored in the recipient cells. This might be due to its low abundance and its rapid intracel- lular degradation. Nonetheless, it underlines the fact that its formation occurs in the lumen of the endoplasmic reticulum- Golgi complex, which endocytosed proteins cannot reach.
To ascertain the observed findings, same extracts were ana- lyzed for endocytosis of lysosomal arylsulfase A and p-hexos- aminidase which are known to be internalized by Man-6-P receptor-mediated endocytosis (Stein et al., 1987; Gupta et al., 1984). As expected, the addition of Man-6-P completely blocked endocytosis of both enzymes, whereas in the absence of this drug about 10% of the secreted arylsulfase A (Fig. 6) as well as hexosaminidase (not shown) were taken up and processed to the mature enzymes.

5444 Dansport and Maturation of Human Acid Sphingomyelinase
ASM ASA
Control 1 Cells 1 Medium I Cells I Medium
M-6-P - + - + - + - +
kDa
97 -
*
4s -
FIG. 7. Endocytosis of ASM by cultured human fibroblasts is not affected by mannose &phosphate. NH,C1-induced secretions of [3sSlmethionine-labeled fibroblasts were dialyzed against MEM and added to the culture media of unlabeled fibroblasts in the absence or presence of 10 m M mannose 6-phosphate (M-6-P). After 12 h cells were harvested and analyzed for internalized immunoprecipitable sphingo- myelinase (ASM) and arylsulfatase A (ASA).
DISCUSSION The first report on the biosynthesis of human acid sphingo-
myelinase in fibroblasts by Jobb and Callahan (Jobb and Cal- lahan, 1989) described the occurrence of a 110-kDa polypeptide, which is processed into a lower form of 84 kDa. This study clearly differs from the results communicated in the present study. Considering the problems of producing specific anti-ASM antisera during the last decade (Weitz et al., 1985; Freemann et al., 1984; Rousson et al., 1987), it remains questionable whether the antibodies used by these groups were able to rec- ognize specifically the enzyme deficient in Niemann-Pick dis- ease. The antibodies used in the present study had been raised against recombinant ASM protein derived from bacterial ex- pression of the human cDNA. The obtained antiserum was assessed to recognize native human ASM with high affinity. The antibody also precipitates the activity of phospholipase C toward phosphatidylcholine from purified preparations of ASM, confirming earlier discussions regarding the identity be- tween lysosomal phospholipase C and ASM (Beaudet et al., 1980; Wherrett and Huterer, 1983; Quintern et al., 1987).
Previous [35S]methionine labeling studies with COS-1 cells, which were transiently transfected with the wild-type cDNA of human ASM, established the relationship between the expres- sion product of the cDNA and immunoprecipitated molecular forms with molecular masses of 75-70 kDa, as well as 57 kDa, respectively.2
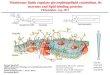
Processing-The processing events are illustrated schemati- cally in Fig. 8. First detectable forms of human ASM in fibro- blasts are polypeptides with apparent molecular masses of 75 and 72 kDa. The difference between their molecular masses is obviously due to proteolysis, since this difference is preserved after deglycosylation. Therefore, the 75-kDa form is believed to be the propropolypeptide ofASM. The precursor form of 72 kDa is processed within 4 h to a protein with a molecular mass of 70 kDa. Since this form corresponds to the molecular size of the native ASM purified from human urine, it is suggested to be the mature form of ASM. There is currently no direct evidence for
* K. Ferlinz, R. Hurwitz, G. Vielhaber, K. Suzuki, and K. Sandhoff, manuscript in preparation.
ER EWGolgi Endoromel Lv0000me
, 'IS (61)
57 (47)
70 (61) malure ASM
62 - mature ASM
FIG. 8. Processing of acid sphingomyelinase in human fibro- blasts. The numbers in brackets refer to the molecular masses (kDa) of the corresponding peptide chains.
attributing this reduction in molecular mass to proteolysis. Both the 72-kDa precursor as well as the 70-kDa mature form have a protein core of the same size (61 kDa), suggesting that maturation depends on posttranslational modifications rather than proteolysis, occurring in a compartment distal to the site of brefeldin A action.
After 8 h of chase, a further peptide of 52 kDa appeared, whereas the intensity of the 70-kDa form was diminished. Con- version of the latter was accomplished only partially after a further period of 16 h. Taken these data together, the question arises as to how the processing ofASM reflects the distribution of mature ASM forms at the steady state level. One would expect the 52-kDa form to be the major component in fibro- blasts and presumably in other tissues. Hence, it is noteworthy that Rousson et al. (1993) reported on the isolation ofASM from human placenta, yielding two protein forms with molecular mass of 70 kDa and approximately 54-57 kDa, respectively. In their final purification step, the latter was the prevailing form. Monoclonal antibodies raised against this preparation were shown to recognize both forms in Western analysis of placental extracts and in fibroblasts extracts enriched by immunoaftinity chromatography. On the other hand, it cannot be ruled out that the the lower molecular mass form detected by Rousson corre- sponds to the 57-kDa form reported here. Concerning this early cleavage product, there is only little information available, mainly due to its low abundance. Studies are in progress to elucidate its enzymatic properties.
Dunsport ofASM-The failure of I-cell fibroblasts to process the precursor to the mature form of 70 kDa, as well as to the 52-kDa polypeptide, indicates that correct intracellular target- ing ofASM to the lysosomes depends on the Man-6-P-mediated pathway. The observation that almost 80% of the newly syn- thesized ASM is recovered from the medium confirms this point of view. It further implies that conversion into mature ASM regularly occurs in the acidic compartment.
Interestingly, [32P]phosphate labeling gave evidence that mannose 6-phosphate residues in the 57-kDa form are missing, consistent with its oligosaccharide chains being predominantly of the complex type. These data are in accordance with the observed secretion rate of the newly synthesized 57-kDa form, which appeared to be identical in normal and I-cell fibroblasts. Therefore, it is questionable whether this protein reaches the lysosomes independently of a Man-6-P receptor mechanism.
Degradation of ASM-Compared with other lysosomal en- zymes, the rapid degradation of ASM is exceptional. Most ly- sosomal enzymes persist within the lysosomes with half-lifes ranging from a few days to several weeks. The present results clearly demonstrate that the rapid loss of radiolabeled ASM is primarily caused by intracellular degradation of the early forms of ASM, i.e. the 72-kDa precursor and the 57-kDa early cleavage product. This pronounced degradation is probably re- flected in the low abundance of ASM in most tissues. Degrada-

Dansport and Maturation of Human Acid Sphingomyelinase 5445
tion is prevented by the addition of brefeldin A, suggesting that this process takes place in the acidic environment. The mature forms of ASM seem to escape this rapid degradative process as judged by their prolonged half-life evaluated by pulse-chase experiments. Therefore, the successive events of maturation might be required to obtain an enzyme configuration, which allows optimal resistance toward proteases and most likely to achieve catalytic activity.
Endocytosis-The presence of Man-6-P-containing culture medium was expected to result in impaired uptake of exog- enously added radiolabeled ASM from fibroblast cultures. Al- most normal rates of endocytosis in the presence of Man-6-P may point to an alternative pathway of receptor-mediated en- docytosis. In human fibroblasts, LDL and other lipoproteins are internalized by receptor-mediated uptake (Anderson et al., 1977). Sphingomyelin and cholesterol are both transported through the bloodstream as integral components of lipopro- teins. Binding of ASM to sphingomyelin, phosphatidylcholine or even to cholesterol may provide an efficient instrument for endocytosis into fibroblasts by LDL- or LDL-related receptors. Further experiments on endocytosis of ASM in the absence of lipoproteins as well as with LDL receptor-deficient fibroblasts should permit elucidation of this theory.
Acknowledgment-We thank Prof. Kunihiko Suzuki for valuable dis- cussion and comments on the manuscript.
REFERENCES
Anderson, R. G. W., Brown, M. S., and Goldstein, J. L. (1977) Cell 10, 351-365 Beaudet, A. L., Hampton, M. S., Patel, K., and Sparrow, J. W. (1980) Clin. Chim.
Brady, R. 0.. Kanfer, J . N., Mock, M. B., and Frederickson, D. S. (1966) Proc. Natl.
Ferlinz, K., Hunvitz, R., and Sandhoff, K. (1991) Biochem. Biophys. Res. Commun.
Freeman, S. J., Davidson, D. J., Shankaran, P., and Callahan, J. W. (1984) Anal.
Acta 108,403414
Acad. Sci. U. S. A. 55, 367-371
179, 1187-1191
Gupta, D. K., Schmidt,,A., von Figura, K., and Hasilik, A. (1984) Hoppe-Seyler’s Z.
Hasilik, A,, and Neufeld, E. F. (1980) J. Biol. Chem. 255, 49374945 Hasilik, A,, Waheed, A., and von Figura, K. (1981) Biochem. Biophys. Res. Com-
Hochuli, E., Bannwarth, W., Dobeli, H., Gentz, R., and Stiiber, D. (1988) Biol
Jobb, E. A,, and Callahan, J. W. (1989) Biochem. Cell. Bid. 67, 801407 Klausner, R. D., Donaldson, J. G., and Lippinscott-Schwartz, J. (1992) J. Cell Biol.
Leroy, J. G., Ho, M. W., MacBrinn, M. C., Zeilke, K., Jacob, J., and O’Brien, J. S. 116, 1071-1080
Levran, O., Desnick, R. J., and Schuchman, E. H. (1991a) Proc. Natl. Acad. Sci. (1972) Pediatr. Res. 6, 752-757
U. S. A. 88,3742-3748 Levran, O., Desnick, R. J., and Schuchman, E. H. (1991bj J. Clin. Znuesf. 88,
80W310 Quintern, L. E., Weitz, G., Nehrkorn, H., Tager, J. M.,, Schram, A. W., and Sand-
hoff, K. (1987) Biochim. Biophys. Acta 922, 323-336 Quintern, L. E., Schuchman, E. H., Levran, 0.. Suchi, M., Ferlinz, K., Reinke, H.,
Sandhoff, K., and Desnick, R. J. (1989) EMBO J. 8,2469-2473 Rousson, R., Bounet, J . , Louisot, P., and Vanier, M. T. (1987) Biochim. Biophys.
Rousson, R., Parvaz, P., Bonnet, J., Rodriguez-Lafrasse, C., Louisot, P., and Vanier, Acta 924,522-528
Schnabel, D., Schroder, M., Fiirst, W., Klein, A., Hunvitz, R., Zenk, T., Weber, J . , M. T. (1993) J. Zmmunol. Methods 160, 199-206
Harzer, K., Paton, B. C., Poulos, A,, Suzuki, K., and Sandhoff, K. (1992) J. Bid.
Biochem. 141,248-252
Physiol. Chem. 366,667476
mun. 98,761-767
Technology, 6, 1321-1325
Schuchman, E. H., Suchi, M., Takahashi, T., Sandhoff, K., and Desnick, R. J. (1991)
Schuchman, E. H., Levran, O., Pereira, L., and Desnick, R. J. (1992) Genomics 12,
Chem. 267, 3312-3315
J. Bid. C h m . 266,85314539
197-205 Spence, M. W., and Callahan, J. W. (1989) in The Metabolic Basis of Inherited
Disease (Scriver, C. R., Beaudet, A. L., Sly, W. S., and Valle, D., edsj 8th Ed., pp. 1655-1676, McGraw-Hill, New York
Stein, M., Zijderhand-Bleekemolen, J. E., Geuze, H., Hasilik, A,, and von Figura, K. (1987) EMBO J. 6,2677-2681
Takahashi, T., Suchi, M., Desnick, R. J., Takada, G., and Schuchman, E. H. (1992) J. Biol. Chem. 267, 12552-12558
Weitz, G., Lindl, T., Hinrichs, U., and Sandhoff, K. (1983) Hoppe-Seyler’s 2. Physiol. Chem. 364, 863-871
Weitz, G., Driessen, M., Brouwer-Kelder, E. M., Sandhoff, IC, Barranger, J. A., Tager, J. M., and Schram, A. W. (1985) Biochim. Biophys. Acta 838, 92-97
Wenger, D. A., Sattler, M., Clark, C., and Wharton, C. (1976) Life Sci. 19,413420 Wherrett, J. R., and Huterer, S. (1983) Neorol. Res. 8, 89-98
~~ ~~~