Embed Size (px)
Citation preview

Hydrobidogia 206: 133-138, 1990. 0 1990 Kluwer Academic Publishers. Printed in Belgium. 133
The influence of humic substances on the aerobic decomposition of submerged leaf litter
Robert G. Qualls ‘7 ’ & Bruce L. Haines ’ ‘Institute of Ecology and Botany Department, University of Georgia, Athens, GA 30602, USA; 2Present address: School of Forestry and Environmental Studies, Duke University, Durham, NC 27706, USA
Received 5 April 1989; in revised form 15 November 1989; accepted 12 December 1989
Key words: fulvic acid, litter decomposition, swamps, wetlands, acidity, humic
Abstract
Leaf material was incubated in flasks containing streamwater in which the pH and the concentration of isolated fulvic acid were varied independently of one another. Decomposition of the leaf material was slower at pH 4 than at pH 5 or 7, but the concentration of fulvic acid had no effect when the pH was held constant. At pH $20 mg C I- ’ humic acid also had no effect on decomposition. High concentra- tions of dissolved fulvic acids may contribute to the slow decomposition of plant litter characteristic of many wetlands through their contribution to hydrogen ion activity, but we could find no evidence for other properties of fulvic acid which inhibit leaf litter decomposition.
Introduction
The decomposition of plant litter is slow in many types of wetlands compared with other freshwater habitats (Brinson et al., 1981). The resulting accumulation of organic matter has a profound effect on the structure and nutrient cycling of the ecosystem, exemplified in the extreme case by peat bogs. Several hypotheses may account for the slow decomposition of submerged plant litter in these habitats: (1) anaerobic conditions, (2) acidity of the water, and (3) inhibition of decomposition by dissolved
humic substances. Many other factors can regulate decomposition, but the factors listed above are distinctive of many wetlands. Anaerobic conditions have long been
known to inhibit decomposition (Acharya, 1935; Godshalk & Wetzel, 1978). However, some flow- ing water wetlands maintain aerobic conditions during much of the year yet exhibit slow decom- position (Kuenzler et al., 1977).
Many freshwater wetlands, particularly swamps and bogs, are acidic. This natural acidity is caused by high concentrations of dissolved (defined here as passing a 0.2 pm filter) humic substances containing carboxylic acid groups (Thurman, 1985). The acid, highly humic water draining from the Okefenokee Swamp typically has a pH ranging from 3.6 to 4.5 (Blood, 1982), and in one case, contained 22 mg C 1 - l fulvic acid and 8 mg C l- r humic acid (Thurman & Malcolm, 1981). High concentrations of humic substances can also be found in the water of wet- lands that are circum-neutral, particularly where

134
there is an input of groundwater high in alkalinity (Gorham et al., 1985).
There was also reason to suspect that high concentrations of dissolved humic substances may be inhibitory to decomposers for reasons other than their effect on hydrogen ion activity. Wetzel (1983, p. 678) attributed antibacterial, inhibitory properties to humic substances, in addi- tion to acidity, which contribute to the accumula- tion of plant remains in bogs. The superficial resemblance of humic substances in water to tan- nins might also suggest inhibitory properties. Tannins are known to inhibit microbial activity on certain substrates by their precipitation of pro- teins (Basaraba & Starkey, 1966). Janzen (1974), for example, associated the poverty of biological activity in the Amazonian blackwater rivers with inhibitory, tannin-like substances in the water. Polyphenols also may be an intrinsic factor within the leaf itself inhibiting initial leaf litter decom- position. Although tannins and polyphenols may be involved in the genesis of some humic acids (Thurman, 1985), the persistence of the inhibitory properties has not been demonstrated. Humic and fulvic acids in solution have been shown to inhibit enzyme activity (Ladd & Butler, 1975 ; Mato & Mendez, 1970). Concentrations in the ranges found in natural waters, 2 to 20 mg L-r, inhibited invertase and protease activity. Humic acids were more effective, in at least one case, than fulvic acids (Ladd & Butler, 1975).
The recent acidification of streams and lakes by mineral acids makes it important to distinguish between the effects of organic and mineral acidity on decomposition (Havas et al., 1984; Gorham et al., 1986; Brakke et al., 1987). Parts of Scandinavia, the Northeastern USA, and Southeastern Canada, in particular, are areas where highly humic waters occur in regions sub- ject to acid precipitation (Gorham et al., 1986; Brakke et al., 1987).
The objective of this experiment was to dis- tinguish between the effects of hydrogen ion activ- ity and dissolved fulvic or humic acid on the decomposition of submerged leaf litter under aerobic conditions.
Methods
The effects of pH, fulvic acid concentration, and their possible interaction, were tested in a randomized two-way factorial design with pH (4.0, 5.0, and 7.0) and fulvic acid concentration (0, 10, 20, and 40 mg C 1 - ’ added) as indepen- dent variables. Several considerations were ad- dressed in our techniques: (a) pH should be ma- nipulated with ions that would not act as limiting nutrients to decomposers (HCl and NaOH), (b) streamwater and fulvic acid should have low concentrations of potentially inhibitory metals, such as Al, to avoid confounding effects and com- plexing interactions, and (c) leaves should be ex- posed to a wide variety of colonizing decomposers which might be adopted to the range (acid to neutral) of the treatments.
Leaf material
Discs were cut from recently abscised autumn leaves on an Acer rubrum L. tree. Discs were air dried and weighed to the nearest 0.01 mg. Twelve were oven dried at 70 “C for an an-to-oven dried weight conversion. To allow discs to be colonized with a diverse microbial assemblage adapted to a range of conditions, submerged litter was col- lected from 3 sources: (1) the Okefenokee Swamp, Ga. with acid (pH 3.9), highly humic (dissolved organic carbon (DOC) = 32 mg 1-l) water, (2) a stream near Tifton, Ga. with moderately acid (pH 5.0), moderately humic (DOC = 9.2 mg l- ‘) water, and (3) a stream at the Coweeta Hydrologic Laboratory, N.C. with neutral (pH 6.9) water, very low in dissolved organic carbon (DOC = 0.92 mg 1-l). For colo- nization, leaf discs were incubated in contact with a mixture of litter from the 3 sources for 18 d. During colonization, half of the discs and litter were incubated in Okefenokee Swamp water and half in Coweeta streamwater. One disc from each of the 2 groups was later included in each flask to avoid bias. For the purpose of measuring weight loss during colonization, 24 discs were harvested after the 18 d period.

135
Preparation of solutions
Fulvic and humic acids were isolated as part of another study (Qualls & Johnson, 1983) from Black Lake, N.C., USA, an acid, highly humic lake-wetlands complex. The fulvic and humic acids were isolated using a slight modification of the procedure of Thurman & Malcolm (1981) detailed in Christman et al. (198 1). Filtered water was acidified, humic substances adsorbed on Amberlite XAD-8 resin (Rohm and Hass, Inc.), eluted with 0.1 N NaOH, reaciditied, and cen- trifuged to remove humic acid. The fulvic and humic acids were then separately readsorbed to the XAD-8 resin, eluted with NaOH, desalted with a cation exchange resin, and lyophilized. To eliminate any low molecular size labile hydro- phobic acids, the fulvic acid was dissolved and dialyzed for 48 hr at 5 ‘C in a Spectrapor dialysis bag (1000 nominal molecular weight cutoff). The nominal molecular weight cutoff is unlikely to be valid for humic substances, but the treatment was meant only to eliminate a small portion of the lowest molecular size material. About 80% of the DOC was retained. The high molecular size fulvic acid concentrate was mixed with Coweeta streamwater, which contained 0.9 mg DOC l- i, to produce DOC concentrations of 0.9, 10.9,20.9, and 40.9 mg C l- ‘. After being acidified and desalted, the fulvic acid was very low in metal ions such as Al, as was the Coweeta streamwater. Metals in the stock solutions were determined by inductively coupled plasma emission spec- troscopy. The pH of stocks of streamwater and fulvic were adjusted to 4.0, 5.0, or 7.0 using HCl or NaOH. Stock solutions were filtered (0.2 pm) into sterile bottles and stored at 3 “C to preserve them. One stock of 20 mg C l- ’ humic acid solu- tion at pH 5 was also prepared.
Incubations
Two leaf discs were incubated in 100 ml solution in each of the 36 flasks (3 replicate flasks x 4 DOC levels x 3 pH levels). Three other smaller groups of flasks were included to check for var-
ious effects. A set of 3 flasks contained disks incubated in the 20 mg C l- ‘, pH 5, humic acid solution to determine whether the effects of humic acid differed from the effects of fulvic acid at the same pH. Another set of 3 flasks contained 0.17 mM NaCl in pH 7 streamwater, which was approximately the maximum amount of Na or Cl ions added to any flask. Nylon fabric discs, inocu- lated by contact with leaves, were placed in other flasks containing 40 mg C 1 - ’ fulvic acid to check for flocculation or decay of the fulvic acid itself.
Each flask was aerated with clean, humid air in a way which produced a gentle water circulation. Dissolved 0, remained near saturation. To minimize the accumulation or depletion of any substances in the solutions in the flasks, solutions were replaced every 5 days with fresh stock. DOC concentrations were measured in the solutions that were removed. The pH in each flask was checked every 1 or 2 days and adjusted if neces- sary. Temperature varied only between 22 and 23 “C.
The leaf discs were harvested after 38 d, dried at 70 ‘C, and weighed. The percent carbon in the discs was determined using a Perkin-Elmer C-H-N Analyzer. All the incubation solutions were filtered with glass fiber filters and analyzed for DOC content by persulfate oxidation and infra-red CO, analysis using a Model 700 Total Organic Carbon Analyzer (01 Corp.). Carbon content of suspended particles filtered from the solutions, and material possibly adhering to the walls of the flasks at the end of the experiment, was estimated by acid-dichromate oxidation (Strickland & Parsons, 1968).
The loss of carbon from the leaf discs was divided into respiration, leaching, and fragmen- tation. Total loss of C during incubations was expressed as a percentage of the initial weight before colonization:
[(initial wt.)( %CJ(P)-(final wt)( %Cf)]/[(initial wt.)( %C,)]
where P, the average proportion remaining after colonization, was calculated from the weight and% C of the 24 discs harvested after the 18 d colonization period. The C leached from the discs
136
during the 38 d incubation was calculated from the final minus initial DOC for each 5 d water change. Fragmentation was the sum of all particu- late C filtered from each flask during the incu- bations. No measurable flocculation or decom- position of the fulvic acid occurred in flasks containing only solution and nylon discs during the short 5 d periods between solution changes. Respiration was presumed to be C loss from discs minus the sum of leaching and fragmentation.
Another estimate of respiration was made with a Gilson Differential Respirometer on days 15 and 30 of the incubations. Only 4 treatments were compared: 0 and 40 mg C l- ’ fulvic acid added, at both pH 4 and pH 7. Discs from each of the 3 replicate flasks for each treatment were placed in fresh incubation solutions in the respirometer. Respiration in samples of the old incubation solu- tions were also measured separately. Results were expressed in terms of carbon respired by assum- ing a respiration quotient of 0.85.
Results and discussion
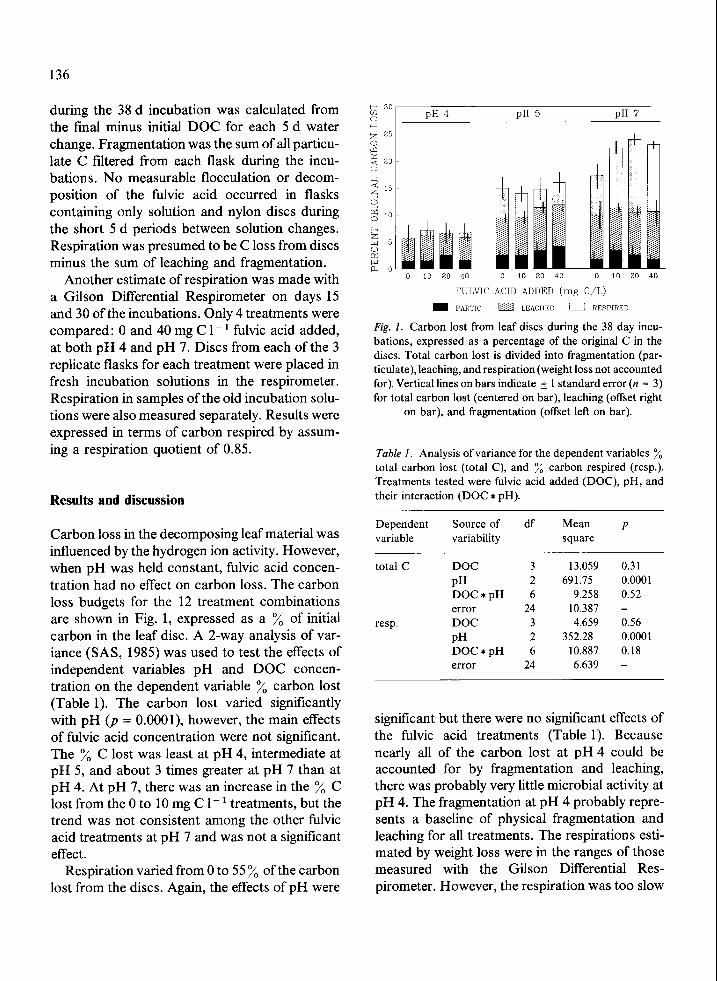
Carbon loss in the decomposing leaf material was influenced by the hydrogen ion activity. However, when pH was held constant, fulvic acid concen- tration had no effect on carbon loss. The carbon loss budgets for the 12 treatment combinations are shown in Fig. 1, expressed as a y0 of initial carbon in the leaf disc. A 2-way analysis of var- iance (SAS, 1985) was used to test the effects of independent variables pH and DOC concen- tration on the dependent variable y0 carbon lost (Table 1). The carbon lost varied significantly with pH (p = O.OOOl), however, the main effects of fulvic acid concentration were not significant. The % C lost was least at pH 4, intermediate at pH 5, and about 3 times greater at pH 7 than at pH 4. At pH 7, there was an increase in the % C lost from the 0 to 10 mg C l- ’ treatments, but the trend was not consistent among the other fulvic acid treatments at pH 7 and was not a significant effect.
Respiration varied from 0 to 55 y0 of the carbon lost from the discs. Again, the effects of pH were
$ 30
s PH 4 PH 5 PH 7
0 10 20 40 0 10 20 40 0 10 20 40
FULVIC ACID ADDED (mg C/L) m PARTIC Ea LEACHED 0 RESPIRED
Fig. 1. Carbon lost from leaf discs during the 38 day incu- bations, expressed as a percentage of the original C in the discs. Total carbon lost is divided into fragmentation (par- ticulate), leaching, and respiration (weight loss not accounted for). Vertical lines on bars indicate & 1 standard error (n = 3) for total carbon lost (centered on bar), leaching (offset right
on bar), and fragmentation (offset left on bar).
Table 1. Analysis of variance for the dependent variables % total carbon lost (total C), and % carbon respired (resp.). Treatments tested were fulvic acid added (DOC), pH, and their interaction (DOC * pH).
Dependent Source of df Mean P variable variability square
total C
resp.
DOC 3 13.059 PH 2 691.75 DOC*pH 6 9.258 error 24 10.387 DOC 3 4.659 PH 2 352.28 DOC*pH 6 10.887 error 24 6.639
0.31 0.000 1 0.52
0.56 0.000 1 0.18 -
significant but there were no significant effects of the fulvic acid treatments (Table 1). Because nearly all of the carbon lost at pH 4 could be accounted for by fragmentation and leaching, there was probably very little microbial activity at pH 4. The fragmentation at pH 4 probably repre- sents a baseline of physical fragmentation and leaching for all treatments. The respirations esti- mated by weight loss were in the ranges of those measured with the Gilson Differential Res- pirometer. However, the respiration was too slow

137
to be measured precisely with the respirometer and the results were not useful for statistical com- parisons. The respirometer measurements for the pH 7 treatments averaged 1.7( of: 0.8)pg C hr - ‘. The average respiration rate for the same discs estimated by weight loss over the entire incubation period was equivalent to 1.4( f 0.4)pg C hr- ‘.
A major mode of carbon loss was by leaching, ranging from 32 to 74% of the carbon lost during the 38 d incubation. The rate of leaching was cal- culated from the small, but precisely ( k 0.1 mg C l- ’ ) measurable, extra accumulation of DOC during the 5 d between changes of solu- tion. This leaching amounted to 2 mg l- ’ DOC at the most, above that added to the solutions, so its possible influence on decomposition was minim- ized by frequent renewal of the solution. The pos- sible decay of the leachate itself in the solution was also minimized by the short period between changes.
The leaf discs incubated with 20 mg C l- ’ humic acid at pH 5 lost 14.8 (k 1.5)x of their initial carbon. This loss was not significantly dif- ferent (p > 0.05) from the other treatments at pH 5, including the one to which no fulvic acid had been added (Duncan’s Multiple Range Test). While the full range of interactions with pH were not tested as they were with fulvic acid, we con- sidered the lack of effects of fulvic acid and the one humic acid treatment to suggest that humic acid would cause no response at the other levels of pH. Most of the humic substances in natural water are classified in the fulvic acid fraction (Thurman, 1985). While Ladd & Butler (1975) found humic acids to be more inhibitory to en- zyme activity than fulvic acids on a weight basis, neither fraction appeared to inhibit leaf litter de- composition in this experiment.
The role of humic substances in complexing heavy metals was a potentially confounding factor. The high concentration of free Al found in non-humic acid lakes is an example of an indirect effect of hydrogen ion concentration. Havas et al. (1984) have pointed out that high concentrations of organic acids which complex Al may ameliorate the toxic effects of Al; an impor-
tant contrast between organic and mineral acidity in lakes and streams. The method of preparation of the fulvic acid and the very low metal concen- tration in the Coweeta streamwater minimized the possible effects of heavy metals. The maximum concentrations of several metals in the incubation solutions were: Al, 33 pg 1 - ’ ; Cu, 43 pg 1 - ’ ; Cr, 20 pg 1 - *. The decay in incubations to which 0.17 mM NaCl was added was not significantly different from the other treatments at pH 7, indi- cating that the varying concentrations (although low) of Na or Cl ions was not a factor in the experiment.
At least under aerobic conditions, we found no support for the hypothesis that a property of humic substances other than their contribution of H + can cause the slower decay of plant litter often observed in humic water environments. This conclusion is consistent with the findings of Benner et al. (1985) who found that raising the pH of a sample of acid, highly humic, Okefenokee Swamp water resulted in accelerated respiration of the cellulose fraction of a radiolabelled ligno- cellulose preparation incubated in the water. However, respiration of the lignin fraction varied little with pH. Nevertheless, humic substances appear capable of inhibiting litter decay by their contribution of H’ ion from carboxylic acid groups. The inhibition of leaflitter decomposition by mineral acidity in natural waters subject to acid deposition has been noted by Francis et al. (1984) and Traaen (1980). For example, Traaen (1980) found leaflitter weight loss to be 45 %, 48x, and 52% of initial weight at pH 4, 4.5-5.2, and 6, respectively; all significantly different.
The results of this experiment suggest that the direct effects of pH on decomposition in acid waters differ little between naturally acid humic waters and waters subjected to acidification from anthropogenic mineral sources. However, at the level of the ecosystem, indirect effects such as metal complexation (Havas et al., 1984) or microenvironmental pH changes may well con- tribute to more subtle differences.

138
Acknowledgements
This research was supported by a grant from the U.S. National Science Foundation (no. BSR- 8501424) and from the Botany Department, UGA.
References
Acharya, C. N., 1935. Studies on the anaerobic decomposi- tion of plant materials. III. Comparisons of the course of decomposition of rice straw under anaerobic, aerobic, and partially aerobic conditions. Biochem. J. 29: 1116-l 120.
Basaraba, J. & R. L. Starkey, 1966. Effects of plant tannins on decomposition of organic substances. Soil Sci. 101: 17-23.
Benner, R., M. A. Moran & R. E. Hodson, 1985. Effect of pH and plant source on lignocellulose biodegradation rates in two wetland ecosystems, the Okefenokee Swamp and a Georgia salt marsh. Limnol. Oceanogr. 30: 489-499.
Blood, E. R., 1982. Surface water hydrology and biogeo- chemistry of the Okefenokee Swamp watershed. Ph.D. Dissertation, University of Georgia, Athens, GA, USA, 194 pp.
Brakke, D. F., A. Henriksen & S. A. Norton, 1987. The relative importance of acidity sources for humic lakes in Norway. Nature 329: 432-434.
Brinson, M. M., A. E. Lugo & S. Brown, 1981. Primary pro- ductivity, decomposition and consumer activity in fresh- water wetlands. Ann. Rev. Ecol. Syst. 12: 123-161.
Christman, R. F., W. T. Liao, D. S. Millington & J. D. Johnson, 1981. Oxidative degradation of aquatic humic material. In L. H. Keith (ed.), Advances in the Identifi- cation and Analysis of Organic Pollutants in Water, ~012. Ann Arbor Science, Ann Arbor, MI, USA: 979-999.
Francis, A. J., H. L. Quinby & G. R. Hendrey, 1984. Effect of lake pH on microbial decomposition of allochthonous litter. In G. R. Hendrey (ed.) Early biotic responses to advancing lake acidification. Ann Arbor Science, Ann Arbor, MI, USA: l-21.
Godshalk, G. L. & R. G. Wetzel, 1978. Decomposition in the littoral zone of lakes. In R. E. Good, et al. (eds) Fresh-
water Wetlands: Ecological Processes and Management Potential. Academic Press, NY: 39-51.
Gorham, E., S. J. Eisenreich, J. Ford & M. V. Santelmann, 1985. The chemistry of bog waters. In W. Stumm (ed.) Chemical Processes in Lakes. John Wiley & Sons, New York: 339-362.
Gorham, E., J. K. Underwood, F. B. Martin & J. G. Ogden, 1986. Natural and anthropogenic causes of lake acidifi- cation in Nova Scotia. Nature 324: 451-453.
Havas, M., T. C. Hutchison & G. E. Likens, 1984. Red herrings in acid rain research. Envir. Sci. Technol. 18: 176A-184A.
Janzen, D. H., 1974. Tropical blackwater rivers, animals, and mast fruiting by the Dipterocarpaceae. Biotropica 6: 69-103.
Kuenzler, E. J., P. J. Mulholland, L. A. Ruley & R. P. Sniffen, 1977. Water quality in North Carolina Coastal Plain streams and effects of channelization. Water Resources Research Inst., Rpt. 127, University of NC., Raleigh, NC, USA, 73 pp.
Ladd, J. N. & J. H. A. Butler, 1975. Humus-enzyme deriva- tives in soil. In E. A. Paul & A. D. McLaren, (eds) Soil Biochemistry, vol. 4. Marcel Dekker, New York: 143-186.
Mato, M. C. & J. Mendez, 1970. Inhibition of indoleacetic acid-oxidase by sodium humate. Geoderma 3: 255-258.
Qualls, R. G. & J. D. Johnson, 1983. Kinetics of the short- term consumption of chlorine by fulvic acid. Envir. Sci. Technol. 17: 692-698.
Strickland, J. D. H. & T. R. Parsons, 1968. A practical handbook of seawater analysis. Bull. Fish Res. Bd Can., 167: 310 pp.
SAS Institute Inc., 1985. SAS user’s guide: statistics. Version 5 ed., Cary, NC., USA, 958 pp.
Thurman, E. M. & R. L. Malcolm, 1981. Preparative iso- lation of aquatic humic substances. Envir. Sci. Technol. 15: 463-466.
Thurman, E. M., 1985. Organic geochemistry of natural waters. Martinus Nijhoff/Junk, Boston, MA, USA, 493 pp.
Traaen, T. S., 1980. Effects of acidity on decomposition of organic matter in aquatic environments. In D. Drablss & A. Tollan (eds) Ecological Impact of Acid Precipitation. Oslo-As, Oslo: 340-341.
Wetzel, R. G., 1983. Limnology. 2nd ed. Saunders College Pub., New York, 743 pp.