Embed Size (px)
Citation preview

426
/ 2b2c 2509 Mp 426 Tuesday Jun 03 10:14 PM EL–FRB 2509
Free Radical Biology & Medicine, Vol. 23, No. 3, pp. 426–434, 1997Published by Elsevier Science Inc.
Printed in the USA. All rights reserved0891-5849/97 $17.00 / .00
PII S0891-5849(97)00113-5
Original Contribution
THE IMPORTANCE OF SODIUM PYRUVATE IN ASSESSING DAMAGEPRODUCED BY HYDROGEN PEROXIDE
ALBERT R. GIANDOMENICO, GEORGE E. CERNIGLIA, JOHN E. BIAGLOW,CRAIG W. STEVENS, and CAMERON J. KOCH
Department of Radiation Oncology, University of Pennsylvania, Philadelphia, PA, USA
(Received 13 May 1996; Revised 3 September 1996; Re-revised 22 December 1996; Accepted 6 January 1997)
Abstract—Instability of hydrogen peroxide solutions was noted during the experimental exposure of human cellsin culture to hydrogen peroxide in experiments designed to study the production and repair of DNA single-strandbreaks. A hydrogen peroxide concentrate was diluted into culture medium, which was then added to experimentalcell cultures at various times, with all cultures assessed for DNA damage at 2 h. Only cells treated by the firstaddition had observable DNA damage. This result was unexpected since these cells had had the maximum repairtime. It was determined that the hydrogen peroxide had been eliminated by the culture medium. To determine themechanism of this elimination, 200 mM hydrogen peroxide was added to various cell culture components, and thesolutions were assayed for hydrogen peroxide after 1 h at 377C. Although most components (except the balancedsalts) showed some hydrogen peroxide degradation, it was found that sodium pyruvate was most effective, by awide margin, in eliminating hydrogen peroxide and its toxic effects. This was confirmed by addition of pyruvate tobalanced salt solutions or buffers, and observing the same elimination of hydrogen peroxide. We subsequently founda few earlier reports describing the decarboxylation reaction between hydrogen peroxide and pyruvate, but no kineticmeasurements have been published and there seems to be no general appreciation for the very high efficiency of thisreaction. The present work presents a preliminary assessment of the importance of pyruvate in the study of hydrogenperoxide and other reactive oxygen species in mammalian cell culture. Published by Elsevier Science Inc.
Keywords—Hydrogen peroxide, Hydrogen peroxide assays or measurement, Pyruvate, Protection from oxidativedamage, DNA damage, DNA single strand breaks, Reactive oxygen species, Toxicity, Decarboxylation
INTRODUCTION
Hydrogen peroxide and other reactive oxygen species(ROS) such as superoxide and hydroxyl radical are aninteresting class of cytotoxic agents that tend to arisenaturally from a wide variety of chemical and biochem-ical oxidations. ROS are also implicated in tissue dam-age found in a number of serious diseases and patho-logic conditions such as ischemia and reperfusioninjury (see ref. 1 for review). Reactive oxygen speciescan interact with macromolecules (e.g., DNA), cellsand tissues to produce damage, which is as yet poorlyunderstood. A key problem is the interconversion of
Address correspondence to: Dr. Cameron J. Koch, University ofPennsylvania, Radiation Oncology, 195 John Morgan Bldg., Phila-delphia, PA 19104-6072.
Work Supported by grant CA49498 (CJK) and CA44982 (JEB)from the National Institutes of Health.
the various molecules and radicals through intermedi-ate reactions with enzymes, metals, and natural reduc-ing agents such as thiols. Thus, it is difficult to establishwhich damage is coming from which reactive species.Often, however, hydrogen peroxide is an essential com-ponent for toxicity.2–7
The molecular mechanism of toxicity from hydro-gen peroxide has been particularly refractory to de-tailed understanding. In mammalian cells, hydrogenperoxide is thought to be primarily eliminated by theenzymes catalase and glutathione peroxidase, whereasit is both produced and eliminated by oxidations of re-ducing agents such as thiols and ascorbate, often ac-companied by the simultaneous production of super-oxide.2,3 In reacting with reduced metals to produce thehydroxyl radical, hydrogen peroxide can produce spe-cific damage at the site of the metal with possible sub-

427Assessing hydrogen peroxide damage using sodium pyruvate
/ 2b2c 2509 Mp 427 Tuesday Jun 03 10:14 PM EL–FRB 2509
sequent redox cycling of the metal requiring reductionby reducing agents or superoxide. The action of hydro-gen peroxide in damaging DNA has been suggested toinvolve primarily the production of isolated single-strand breaks,8 but the details of the damage kineticshave varied substantially in different laboratories. Theperoxide concentration has seldom been monitored inthese studies, and typical protocols have simply addeda given concentration for a ‘‘reasonable’’ time (e.g., 1h). The original studies by Bradley and Erickson9
found similar amounts of DNA single-strand breakdamage (SSB) when the exposure was at either 07 or377. Repair after peroxide treatment proceeded some-what more rapidly than after ionizing radiation. In con-trast, Ward and Blakely found SSB damage at 07 butnot at 377, and attributed the latter to an extremely rapidrepair.8,10 Prise et al.11 found damage at both tempera-tures but did not study repair. Baker and He12 dem-onstrated rather similar amounts of damage from anumber of different peroxides, at both low and hightemperatures. Byrnes and Petering13 found very littledamage from peroxide in isolated DNA but Baker andHe12 showed extensive damage to isolated DNA.
In investigating these differences we observed whatappeared to be quite anomalous kinetics for the induc-tion and repair of DNA single strand breaks by hydro-gen peroxide; the break frequency appeared to be highimmediately after peroxide exposure and moderate af-ter a 2 h repair time, but negligible at intermediatetimes. This result turned out to be an artifact of theexperimental timing and the method of mixing the per-oxide stock solution, which had been diluted into freshmedium and held at 377 to minimize temperature andpH fluctuations on addition to the cells. Thus, if thediluted peroxide was added immediately to the cells,for either immediate assay or for the longest repairtimes, the actual peroxide concentration was as ex-pected. All other conditions involved a loss of peroxideby reaction with medium component(s) . Although pre-vious experiments had suggested that peroxide was sta-ble when diluted into medium, we realized that the me-dium formulation was different in the present work.After considerable investigation, the peroxide-consum-ing additive was found to be pyruvate. Although wesubsequently found a limited number of references tothe use of pyruvate in reducing certain types of oxi-dative injury, because of the ‘‘well known’’ decarbox-ylation reaction14–18 it seemed as though there was infact almost no available information on the kinetics orsignificance of this reaction in vitro or in chemical sys-tems.
This article provides some initial characterization ofthe reaction between hydrogen peroxide and pyruvate.
We show that cells incubated with pyruvate are verysubstantially protected against hydrogen peroxide in-duced damage, in both survival and DNA strand breakassays. Two new assays for hydrogen peroxide arecompared, and our results may be helpful to those wish-ing to monitor peroxide in biologically complex media.Pyruvate may be useful as an efficient, readily diffus-ible and specific scavenger of hydrogen peroxide infuture studies determining the mechanism of action ofROS. Our results suggest that the chemical eliminationof hydrogen peroxide by pyruvate can compete effec-tively with cellular enzyme systems at low to moderatecell densities.
MATERIALS AND METHODS
Cell culture
Cell culture solutions were from GIBCO (Grand Is-land, NY, USA). Chemicals and biochemicals wereobtained in the best quality available from Sigma(Sigma Chemical Co., St. Louis, MO, USA) and usedas supplied. A549 human lung carcinoma cells (ob-tained from ATTC) were maintained in exponentialgrowth phase in a 1/1 mixture of Dulbecco’s Mini-mum Essential and Ham’s F-12 media containing stan-dard antibiotics (DMEMTF12) and supplemented with10% fetal calf serum (DMEMTF12TS) at 377 in a hu-midified atmosphere of 95% air plus 5% carbon diox-ide. The two components of this medium formulationcontained 0.0 mM and 1.0 mM pyruvate, respectively.
DNA damage assay
DNA single-strand breaks were measured by a mod-ification of the alkaline elution method.19 Briefly, 10–15 million exponentially growing cells in a T75 flaskwere incubated with 14C-thymidine (0.05 mCi/ml) for20 h; the cells were trypsinized, counted, and platedonto 60 mm diameter glass dishes (1.0 1 106 cells in3 ml of medium). After 2–3 h incubation at 377 toallow firm attachment, exposure to hydrogen peroxideor radiation was initiated, in some cases followed by arepair interval at 377 in the absence of damaging agent.Then the cells were removed from the glass dishes withtrypsin and subjected to the alkaline elution assay. Inthis assay, cells are layered onto a filter and lysed inalkaline detergent; subsequently the relatively nakedDNA is eluted at pH 12.2. The percentage of radioac-tive DNA remaining on the polycarbonate filter is mea-sured vs. time by collected eluted fractions and mea-suring the radioactive counts. The slopes of these‘‘elution profiles’’ have been shown to be linear withradiation dose.19,20

428 A. R. GIANDOMENICO et al.
/ 2b2c 2509 Mp 428 Tuesday Jun 03 10:14 PM EL–FRB 2509
Peroxide assays
Hydrogen peroxide was measured by two methods,a commercial colorimetric method (Pierce PeroXO-quant, product number 23280, Pierce Chemical Com-pany, Chicago, IL, USA) and an enzyme assay modi-fied from that described by Flohe et al.21 In thecommercial assay, hydrogen peroxide in the test solu-tion oxidizes complexed ferrous ion, and the resultingferric ion forms a colored complex with xylenol orange.The enzyme based assay involved an initial precipita-tion of macromolecules in 5% trichloroacetic acid(TCA) followed by high-speed centrifugation in a mi-crofuge to remove any endogenous proteins or othermacromolecules. After neutralizing the acid superna-tant, a sample was added to a solution containing 100mM potassium phosphate, with 1 mM ethylenediami-netetraacetic acid, 0.2 mM diethylenetriaminepenta-acetic acid (DTPA), 1 mM sodium azide, 200 mMglutathione (GSH) and 1 unit /ml of glutathione per-oxidase, pH 7.5. The following reaction takes place:
2GSH / H O r GSSG / 2 H O2 2 2
and after completion (typically 10–20 min at 377) theremaining GSH was determined by reaction with added5,5 *-dithio-bis(2-nitrobenzoic acid) (DTNB). Be-cause two GSH molecules are consumed for each per-oxide, the effective molar absorption change is (-2) 113600 (i.e., twice the molar absorption coefficient forreduced DTNB). Flohe et al.’s method did not measurethe remaining glutathione with DTNB; instead, gluta-thione was regenerated using a second enzyme (glu-tathione reductase) and this reaction was followed bymonitoring the loss of the required substrate, NADPH.Since the molar absorption coefficient of NADPH isabout 6400, our method has a four-fold higher sensi-tivity, uses a non-UV wavelength (412 nm vs. 340 nm)and does not require the extra components. As withmost peroxide assays, the enzyme method is not spe-cific for hydrogen peroxide because of the general reac-tivity of glutathione peroxidase.
The concentration of stock peroxide solutions (typ-ically 30% or Ç 10 M) was determined after dilutionsmade by mass of 1 part peroxide plus 999 parts water.One part of this solution was added to 99 parts of 1mM ferrous sulfate in 1 N sulfuric acid and the increasein absorbance at 224 noted. This is the ‘‘Fricke’’ do-simeter solution, which has been very accurately char-acterized to have a molar absorbance change in acidfor ferrous r ferric of 4565 @ 224nm.22 Two ferrousions are oxidized for each hydrogen peroxide molecule,since ferrous ion plus peroxide produces ferric ion plushydroxyl radical; the hydroxyl radical then oxidizes a
second ferrous ion. Thus, the effective molar absor-bance change is 9130. The Fricke and enzyme assaysgave indistinguishable results for pure solutions of per-oxide but the Fricke solution did not work with com-plex media or at nonacid pH. Both assays are based onabsolute chemical determinations. Although this is nottrue for the Pierce assay, it appeared to be equally re-liable when calibrated by a suitable standard. For boththe Pierce and enzyme assays, a relatively small vol-ume of experimental solution was added to the assayreagents ( typically less than 10% by volume).
Cellular consumption of peroxide
In experiments to determine the cellular componentof hydrogen peroxide consumption, A549 cells wereeither plated onto glass dishes (330,000 or 33,000 cellsper ml, holding the medium volume constant at 3 ml)or trypsinized and incubated in stirred suspensions at33,000 cells /ml. Dishes containing attached cells wererinsed free of medium and allowed to equilibrate in 3ml of buffered saline. Hydrogen peroxide was addedto fresh, warm, DMEMTF12 (serum free) or saline afew seconds before addition to the cells. Samples werewithdrawn periodically to assay peroxide. Toxicity ofhydrogen peroxide was assayed with the suspensionculture in serum-free medium with or without 1 mMpyruvate. Aliquots were removed and diluted with cat-alase-containing medium (25 U/ml) , then the cellswere plated at known numbers in 100 mm plastic disheswith fresh DMEMTF12TS. Surviving cells were as-sessed by colony formation after appropriate growthtimes.23
Chemical consumption of peroxide
The chemical consumption of peroxide in varioussolutions was measured by adding peroxide concen-trates, mixing rapidly, and then taking samples for as-say at various times following peroxide addition. Sincethe stock peroxide concentration (30%) was roughly10 M, initial peroxide dilutions were made in deionizedwater. The chemicals or enzymes responsible for per-oxide depletion did not interfere with the peroxide as-say since ‘‘zero’’ time values, with one exception,agreed with standard peroxide solutions made up inwater or simple buffers, diluted to the same concentra-tions. The one exception was ferrous ion, which ap-peared to react with peroxide as quickly as the solutionscould be mixed (see Results section).
RESULTS
When hydrogen peroxide was premixed withDMEMTF12TS at 377 to a final concentration of 200
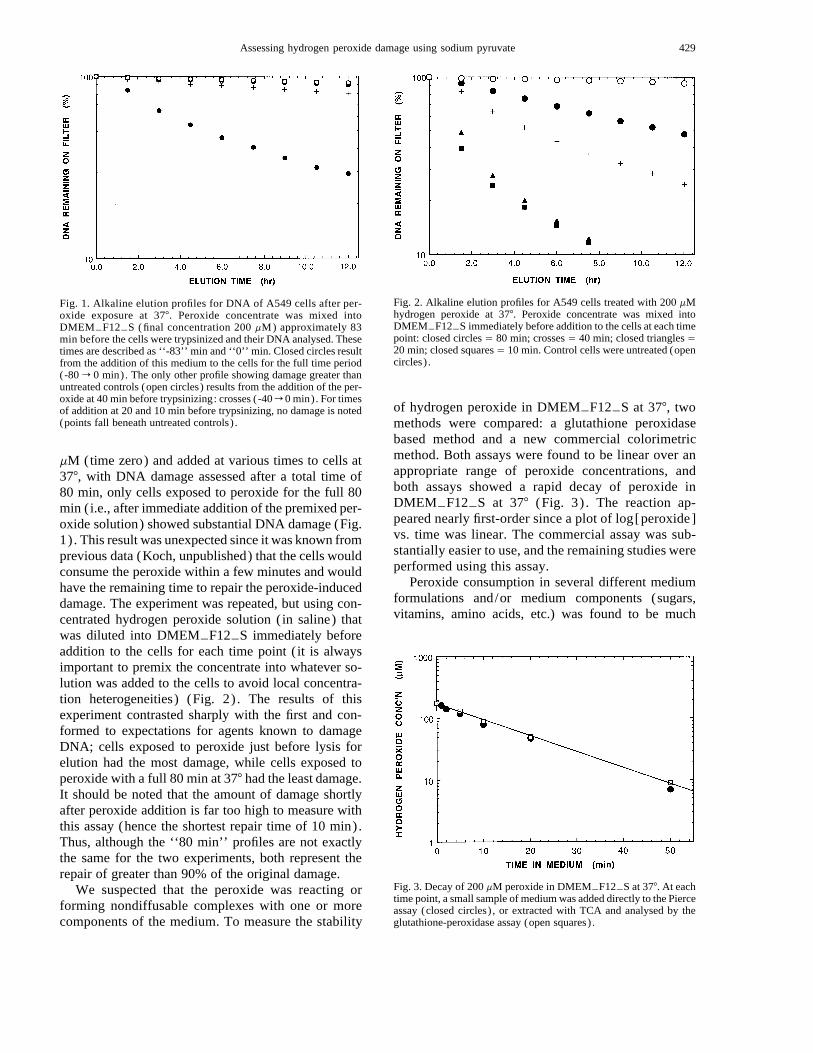
429Assessing hydrogen peroxide damage using sodium pyruvate
/ 2b2c 2509 Mp 429 Tuesday Jun 03 10:14 PM EL–FRB 2509
Fig. 1. Alkaline elution profiles for DNA of A549 cells after per-oxide exposure at 377. Peroxide concentrate was mixed intoDMEMTF12TS (final concentration 200 mM) approximately 83min before the cells were trypsinized and their DNA analysed. Thesetimes are described as ‘‘-83’’ min and ‘‘0’’ min. Closed circles resultfrom the addition of this medium to the cells for the full time period(-80 r 0 min). The only other profile showing damage greater thanuntreated controls (open circles) results from the addition of the per-oxide at 40 min before trypsinizing: crosses (-40 r 0 min). For timesof addition at 20 and 10 min before trypsinizing, no damage is noted(points fall beneath untreated controls) .
Fig. 2. Alkaline elution profiles for A549 cells treated with 200 mMhydrogen peroxide at 377. Peroxide concentrate was mixed intoDMEMTF12TS immediately before addition to the cells at each timepoint: closed circles Å 80 min; crosses Å 40 min; closed triangles Å20 min; closed squares Å 10 min. Control cells were untreated (opencircles) .
Fig. 3. Decay of 200 mM peroxide in DMEMTF12TS at 377. At eachtime point, a small sample of medium was added directly to the Pierceassay (closed circles) , or extracted with TCA and analysed by theglutathione-peroxidase assay (open squares) .
mM (time zero) and added at various times to cells at377, with DNA damage assessed after a total time of80 min, only cells exposed to peroxide for the full 80min (i.e., after immediate addition of the premixed per-oxide solution) showed substantial DNA damage (Fig.1) . This result was unexpected since it was known fromprevious data (Koch, unpublished) that the cells wouldconsume the peroxide within a few minutes and wouldhave the remaining time to repair the peroxide-induceddamage. The experiment was repeated, but using con-centrated hydrogen peroxide solution (in saline) thatwas diluted into DMEMTF12TS immediately beforeaddition to the cells for each time point ( it is alwaysimportant to premix the concentrate into whatever so-lution was added to the cells to avoid local concentra-tion heterogeneities) (Fig. 2) . The results of thisexperiment contrasted sharply with the first and con-formed to expectations for agents known to damageDNA; cells exposed to peroxide just before lysis forelution had the most damage, while cells exposed toperoxide with a full 80 min at 377 had the least damage.It should be noted that the amount of damage shortlyafter peroxide addition is far too high to measure withthis assay (hence the shortest repair time of 10 min).Thus, although the ‘‘80 min’’ profiles are not exactlythe same for the two experiments, both represent therepair of greater than 90% of the original damage.
We suspected that the peroxide was reacting orforming nondiffusable complexes with one or morecomponents of the medium. To measure the stability
of hydrogen peroxide in DMEMTF12TS at 377, twomethods were compared: a glutathione peroxidasebased method and a new commercial colorimetricmethod. Both assays were found to be linear over anappropriate range of peroxide concentrations, andboth assays showed a rapid decay of peroxide inDMEMTF12TS at 377 (Fig. 3) . The reaction ap-peared nearly first-order since a plot of log[peroxide]vs. time was linear. The commercial assay was sub-stantially easier to use, and the remaining studies wereperformed using this assay.
Peroxide consumption in several different mediumformulations and/or medium components (sugars,vitamins, amino acids, etc.) was found to be much
430 A. R. GIANDOMENICO et al.
/ 2b2c 2509 Mp 430 Tuesday Jun 03 10:14 PM EL–FRB 2509
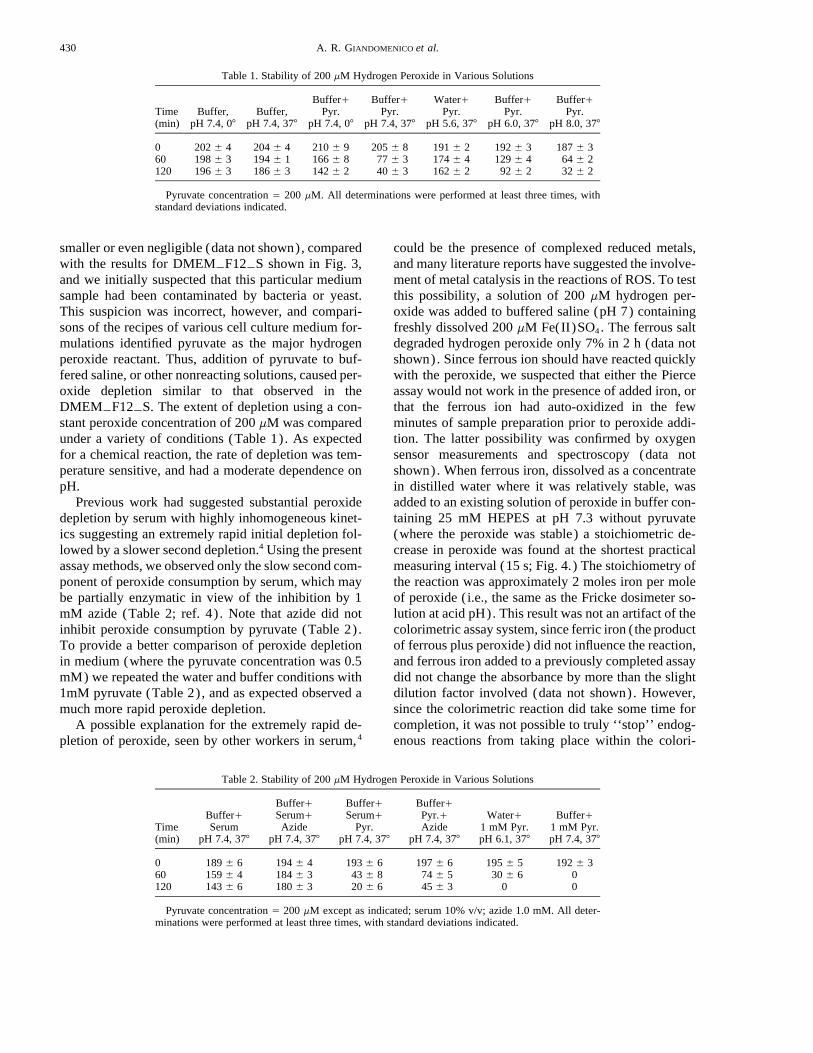
Table 1. Stability of 200 mM Hydrogen Peroxide in Various Solutions
Time(min)
Buffer,pH 7.4, 07
Buffer,pH 7.4, 377
Buffer/Pyr.
pH 7.4, 07
Buffer/Pyr.
pH 7.4, 377
Water/Pyr.
pH 5.6, 377
Buffer/Pyr.
pH 6.0, 377
Buffer/Pyr.
pH 8.0, 377
0 202 { 4 204 { 4 210 { 9 205 { 8 191 { 2 192 { 3 187 { 360 198 { 3 194 { 1 166 { 8 77 { 3 174 { 4 129 { 4 64 { 2120 196 { 3 186 { 3 142 { 2 40 { 3 162 { 2 92 { 2 32 { 2
Pyruvate concentration Å 200 mM. All determinations were performed at least three times, withstandard deviations indicated.
Table 2. Stability of 200 mM Hydrogen Peroxide in Various Solutions
Time(min)
Buffer/Serum
pH 7.4, 377
Buffer/Serum/Azide
pH 7.4, 377
Buffer/Serum/
Pyr.pH 7.4, 377
Buffer/Pyr./Azide
pH 7.4, 377
Water/1 mM Pyr.pH 6.1, 377
Buffer/1 mM Pyr.pH 7.4, 377
0 189 { 6 194 { 4 193 { 6 197 { 6 195 { 5 192 { 360 159 { 4 184 { 3 43 { 8 74 { 5 30 { 6 0120 143 { 6 180 { 3 20 { 6 45 { 3 0 0
Pyruvate concentration Å 200 mM except as indicated; serum 10% v/v; azide 1.0 mM. All deter-minations were performed at least three times, with standard deviations indicated.
smaller or even negligible (data not shown), comparedwith the results for DMEMTF12TS shown in Fig. 3,and we initially suspected that this particular mediumsample had been contaminated by bacteria or yeast.This suspicion was incorrect, however, and compari-sons of the recipes of various cell culture medium for-mulations identified pyruvate as the major hydrogenperoxide reactant. Thus, addition of pyruvate to buf-fered saline, or other nonreacting solutions, caused per-oxide depletion similar to that observed in theDMEMTF12TS. The extent of depletion using a con-stant peroxide concentration of 200 mM was comparedunder a variety of conditions (Table 1). As expectedfor a chemical reaction, the rate of depletion was tem-perature sensitive, and had a moderate dependence onpH.
Previous work had suggested substantial peroxidedepletion by serum with highly inhomogeneous kinet-ics suggesting an extremely rapid initial depletion fol-lowed by a slower second depletion.4 Using the presentassay methods, we observed only the slow second com-ponent of peroxide consumption by serum, which maybe partially enzymatic in view of the inhibition by 1mM azide (Table 2; ref. 4) . Note that azide did notinhibit peroxide consumption by pyruvate (Table 2).To provide a better comparison of peroxide depletionin medium (where the pyruvate concentration was 0.5mM) we repeated the water and buffer conditions with1mM pyruvate (Table 2), and as expected observed amuch more rapid peroxide depletion.
A possible explanation for the extremely rapid de-pletion of peroxide, seen by other workers in serum,4
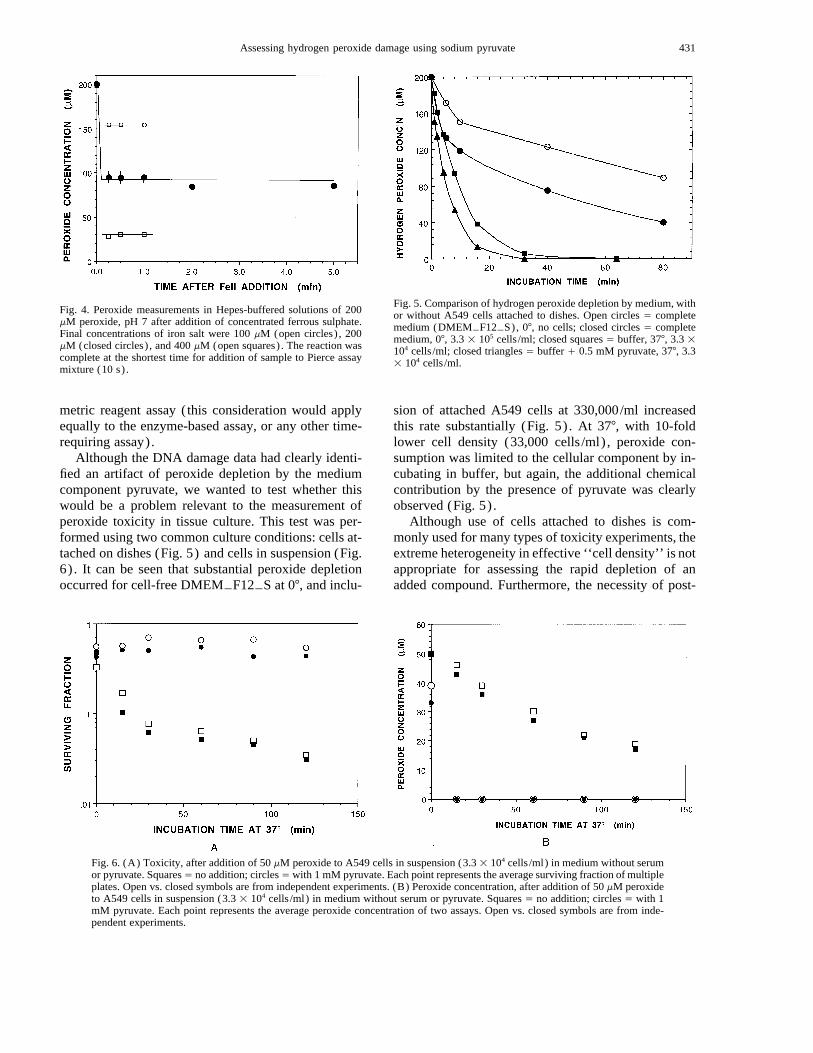
could be the presence of complexed reduced metals,and many literature reports have suggested the involve-ment of metal catalysis in the reactions of ROS. To testthis possibility, a solution of 200 mM hydrogen per-oxide was added to buffered saline (pH 7) containingfreshly dissolved 200 mM Fe(II)SO4. The ferrous saltdegraded hydrogen peroxide only 7% in 2 h (data notshown). Since ferrous ion should have reacted quicklywith the peroxide, we suspected that either the Pierceassay would not work in the presence of added iron, orthat the ferrous ion had auto-oxidized in the fewminutes of sample preparation prior to peroxide addi-tion. The latter possibility was confirmed by oxygensensor measurements and spectroscopy (data notshown). When ferrous iron, dissolved as a concentratein distilled water where it was relatively stable, wasadded to an existing solution of peroxide in buffer con-taining 25 mM HEPES at pH 7.3 without pyruvate(where the peroxide was stable) a stoichiometric de-crease in peroxide was found at the shortest practicalmeasuring interval (15 s; Fig. 4.) The stoichiometry ofthe reaction was approximately 2 moles iron per moleof peroxide (i.e., the same as the Fricke dosimeter so-lution at acid pH). This result was not an artifact of thecolorimetric assay system, since ferric iron (the productof ferrous plus peroxide) did not influence the reaction,and ferrous iron added to a previously completed assaydid not change the absorbance by more than the slightdilution factor involved (data not shown). However,since the colorimetric reaction did take some time forcompletion, it was not possible to truly ‘‘stop’’ endog-enous reactions from taking place within the colori-
431Assessing hydrogen peroxide damage using sodium pyruvate
/ 2b2c 2509 Mp 431 Tuesday Jun 03 10:14 PM EL–FRB 2509
Fig. 4. Peroxide measurements in Hepes-buffered solutions of 200mM peroxide, pH 7 after addition of concentrated ferrous sulphate.Final concentrations of iron salt were 100 mM (open circles) , 200mM (closed circles) , and 400 mM (open squares) . The reaction wascomplete at the shortest time for addition of sample to Pierce assaymixture (10 s) .
Fig. 5. Comparison of hydrogen peroxide depletion by medium, withor without A549 cells attached to dishes. Open circles Å completemedium (DMEMTF12TS), 07, no cells; closed circles Å completemedium, 07, 3.3 1 105 cells /ml; closed squares Å buffer, 377, 3.3 1104 cells /ml; closed triangles Å buffer / 0.5 mM pyruvate, 377, 3.31 104 cells /ml.
Fig. 6. (A) Toxicity, after addition of 50 mM peroxide to A549 cells in suspension (3.3 1 104 cells /ml) in medium without serumor pyruvate. SquaresÅ no addition; circlesÅ with 1 mM pyruvate. Each point represents the average surviving fraction of multipleplates. Open vs. closed symbols are from independent experiments. (B) Peroxide concentration, after addition of 50 mM peroxideto A549 cells in suspension (3.3 1 104 cells /ml) in medium without serum or pyruvate. Squares Å no addition; circles Å with 1mM pyruvate. Each point represents the average peroxide concentration of two assays. Open vs. closed symbols are from inde-pendent experiments.
metric reagent assay (this consideration would applyequally to the enzyme-based assay, or any other time-requiring assay).
Although the DNA damage data had clearly identi-fied an artifact of peroxide depletion by the mediumcomponent pyruvate, we wanted to test whether thiswould be a problem relevant to the measurement ofperoxide toxicity in tissue culture. This test was per-formed using two common culture conditions: cells at-tached on dishes (Fig. 5) and cells in suspension (Fig.6) . It can be seen that substantial peroxide depletionoccurred for cell-free DMEMTF12TS at 07, and inclu-
sion of attached A549 cells at 330,000/ml increasedthis rate substantially (Fig. 5) . At 377, with 10-foldlower cell density (33,000 cells /ml) , peroxide con-sumption was limited to the cellular component by in-cubating in buffer, but again, the additional chemicalcontribution by the presence of pyruvate was clearlyobserved (Fig. 5) .
Although use of cells attached to dishes is com-monly used for many types of toxicity experiments, theextreme heterogeneity in effective ‘‘cell density’’ is notappropriate for assessing the rapid depletion of anadded compound. Furthermore, the necessity of post-

432 A. R. GIANDOMENICO et al.
/ 2b2c 2509 Mp 432 Tuesday Jun 03 10:14 PM EL–FRB 2509
treatment trypsinization to assess plating efficiency cancause problems of interpretation with agents such ashydrogen peroxide, which may damage cell mem-branes. Thus, we used cells in suspension to assess tox-icity and peroxide depletion under relatively homoge-neous conditions. A549 cells were stirred (33,000cells /ml) in serum free medium, with or without 1 mMpyruvate, and treated with 50 mM hydrogen peroxide(Fig. 6) . The pyruvate provided complete protectionagainst peroxide’s toxicity (Fig. 6A) and dramaticallyenhanced clearance of peroxide from the suspension(Fig 6B). The time course for peroxide depletion andcausation of toxicity was very similar for the non-py-ruvate treated cell suspensions (Figs. 6A and 6B; openand closed squares) .
DISCUSSION
Our results show that physiological concentrationsof pyruvate, present in various medium preparationsand presumably also in serum, can dramatically alterthe effective lifetime of added hydrogen peroxide, in arapid chemical reaction. The temperature and pH de-pendence of the reaction, and the highly variableamount of pyruvate found in biological media, indeedin living cells, may help to explain some of the contro-versy surrounding the toxicity and metabolic effects ofhydrogen peroxide. It certainly has aided in the inter-pretation of discrepancies in our own work.
Although it was not our intention to provide a de-tailed analysis of the toxic effects of hydrogen perox-ide, the present studies on DNA damage and repair mayhelp to consolidate the substantial variations in effectsreported in the literature. Because of various modifi-cations to the alkaline elution procedure19 we are ableto measure damage in the 0.25 to 5 Gy range, corre-sponding to relative plating efficiencies of 1.0 to 0.2,respectively. At concentrations of peroxide well belowthe toxic range, at either 07 or 377, we observe the pro-duction of very large numbers of alkaline-elution de-tectable single strand breaks (Figs. 1 and 2; data notshown for 07) . Our results at zero degrees are some-what consistent with several literature reports,8,9,12,24
except that we observe significant damage at evenlower peroxide concentrations than have previouslybeen studied. Some reports have suggested little or nodamage at 37725 and this has been attributed to rapidrepair of lesions with little consequence. We tested thisspecifically in the experiments shown in Fig. 2. Clearly,the breaks are repaired quite slowly at 377 and similarrepair kinetics are observed when the peroxide expo-sure is at 07 with repair at 377 (data not shown). Thus,our data agrees most closely with that of Baker andHe12 except that the conditions of our assay suggest
even greater amounts of initial damage. The reasonsfor the failure of some investigators to observe damageat 377 are as yet unclear, but our own similar initialfindings (Fig. 1) were clearly caused by an artifact ofperoxide loss by pyruvate.
In studies performed several years ago, we foundthat the toxicity of peroxide in Chinese hamster fibro-blasts in culture was inversely proportional to cell den-sity over a wide range of cell densities; equivalent tox-icity was found if the peroxide concentration wasvaried to match cell density. This suggested that thetoxicity was related to the total amount of peroxideadded per cell, since it was found, using the enzymeassay described in this paper, that all of the peroxidewas consumed by the cell culture. These initial studieswere preempted from publication by similar work doneby other investigators at that time5; nevertheless, it isvery worth emphasizing that although hydrogen per-oxide is moderately toxic in the range of 50 mM or soat cell densities such as the 33,000 per ml, as shown inFigs. 5 and 6, the actual quantity of peroxide metabo-lized by each cell in such conditions is closer to themolar range, since all peroxide is consumed and theratio of medium volume to cell volume is roughly10,000:1. Our more recent studies had not confirmedthe cell density effects found previously; thus, the tox-icity seen at low cell densities and correspondingly lowperoxide concentrations was not confirmed. This resulthas been traced directly to the addition of pyruvate inthe current medium formulation. If one scans the lit-erature, it is possible to observe substantially differentlevels of toxicity produced by hydrogen peroxide; in-variably, low toxicity is found for high cell densities ormedium formulations known to contain pyruvate.
The mechanism of pyruvate’s elimination of hydro-gen peroxide has been under limited investigation foralmost a century. A 1904 reference,26 describes the re-action of 30% hydrogen peroxide and solid pyruvicacid as producing an ‘‘efflorescence.’’ In 1929, Battieet al.27 indicated that carbon dioxide was produced incopper catalyzed reactions of organic acids with hy-drogen peroxide, and a 1949 paper suggested a possiblemechanism for base catalyzed oxidation of a-keto acidsby hydrogen peroxide.14 By 1977, a college biochem-istry textbook referred to the ‘‘well known’’ nonen-zymatic decarboxylation of a-keto acids by hydrogenperoxide.28 However, this information is certainly notgenerally acknowledged in more current texts, or in-deed most of the biological /biochemical literature.Some exceptions do exist however. Pyruvate has beenshown to protect E. coli from peroxide damage causedby visible light during culture of sea water,29 and drink-ing water.30 The addition of either catalase or pyruvateto culture media has been shown to reverse the affects

433Assessing hydrogen peroxide damage using sodium pyruvate
/ 2b2c 2509 Mp 433 Tuesday Jun 03 10:14 PM EL–FRB 2509
of heat stress or air drying on cultures of both Staph-ylococcus aureus , 31 and Salmonella senftenberg .32 Thebacterial results might be anticipated since such organ-isms can have a pyruvate decarboxylase enzyme. Per-haps less expected is the demonstration of pyruvate’sattenuation of renal reperfusion injury15 and cardiac re-perfusion injury.16 Also, the cytotoxicity during isch-emia (by metal catalyzed autooxidation of cysteine andgeneration of peroxide) has been shown by Nath andco-workers17 to be attenuated by pyruvate. These in-vestigators have also shown protection by pyruvate andother a-keto acids against menadione-induced oxida-tive injury (both cell viability and DNA double-strandbreaks) , and furthermore have directly measured 14CO2
release from labeled pyruvate.18 The only paper wehave found, demonstrating protection by pyruvateagainst hydrogen peroxide toxicity for cells in tissueculture at low cell density, is by Andrae et al.33 Lookingat the other side of the coin, ‘‘contaminants’’ that no-nenzymatically decarboxylate pyruvate in liver homo-ginates have been shown to be peroxide derivativesfrom laboratory chemicals.34 A general conclusionmight be that although the protective action of pyruvateagainst hydrogen peroxide damage has been studied ina variety of systems, an application of this knowledgehas not generally been found in the mammalian cellliterature, and we have been unable to find informationon the kinetics of the process.
The very substantial protection observed for cells inculture medium with only 1 mM added pyruvate (Fig.6) is certainly higher than one might expect consideringthe overall kinetics of chemical and cellular peroxidedepletion (Figs. 3 and 5 and Tables 1 and 2). A pos-sible explanation for this could be transport of pyruvateinto cells; thus, the intracellular protection, whether be-cause of the concentration or distribution of pyruvatecould be advantageous compared with, for example, thecompartmentalized distribution of catalase. It is alsopossible that this cell line has a peroxide-requiring py-ruvate decarboxylase enzyme, and we are presently in-vestigating this possibility.
The data presented here neither confirm nor refutethe role of metals in the toxic action of peroxide. Un-chelated ferrous ion is clearly unstable at pH 7. Evenallowing for this instability, peroxide and ferrous ioncannot co-exist within practical standard chemical mea-suring times (Fig. 4) . If the initial product of the re-action between peroxide and ferrous had been a freehydroxyl radical, in the often discussed ‘‘Fenton’’ re-action, then it might have been expected that one per-oxide molecule would be consumed per ferrous ion,since there was a large excess of hydroxyl radical scav-enger present ( the 25 mM HEPES buffer) .35 Our datado not support this sequence of events, since the ob-
served stoichiometery was two ferrous ions per per-oxide consumed, although we cannot rule out a rapidreaction between the HEPES radical, resulting from hy-droxyl attack, and a second ferrous ion. Although agreat deal more work would be required to elucidatethe precise chemical mechanism, the data resultingfrom this simple experiment would support the modelof Masarwa et al., 36 who suggested that the reactionbetween peroxide and ferrous ion proceeds in a com-plex with two reduced metal ions and a single peroxidemolecule.
Perhaps our most important result is the demonstra-tion that pyruvate represents an important third com-ponent of protection against hydrogen peroxide andother ROS in cells, supplementing the well-recognizedroles of catalase and glutathione peroxidase. Nath etal.18 came to a similar conclusion resulting from themenadione-toxicity studies. A potential advantage forprotection by pyruvate is that it is specific, and pro-duces no toxic byproducts (carbon dioxide and ace-tate) . In contrast, peroxide’s reaction with catalase mayproduce high local oxygen concentrations or the per-oxidation of other substrates; peroxide’s reaction withglutathione peroxidase and GSH produces GSSG,which must then be reduced by glutathione reductaseand NADPH. If one considers that during a peroxide-induced toxic event, a cell may metabolize 0.1 to 0.5molar-equivalent of peroxide, the generation of halfthis quantity of oxygen or glutathione disulfide (prod-ucts of catalase or glutathione peroxidase, respectively)may be in itself toxic. Surprisingly, the consequencesof production of high oxygen or glutathione disulfideconcentrations have seldom been considered in the tox-icity of hydrogen peroxide.
Finally, the introduction of two peroxide assays thatcan be used in complex media may be helpful in futurework. There are many literature reports of difficultiesin the quantitation of peroxide in serum and media,with horseradish-peroxidase based techniques predom-inating.4 We have traced most of these to nonspecific-ities in the reactivity of horseradish-peroxidase orunusable wavelengths (data not shown). The enzyme-based assay described herein is particularly immune tosuch interferences, and the new Pierce assay may offersimilar advantages. It is hoped that the new data andperoxide quantification methods introduced in this pa-per will help in the further elucidation of some of thebaffling properties of hydrogen peroxide damage inmammalian cell systems.
REFERENCES
1. Halliwell, B.; Gutteridge, J. M. C., eds. Free radicals in biologyand medicine. London: Oxford University Press; 1989.

434 A. R. GIANDOMENICO et al.
/ 2b2c 2509 Mp 434 Tuesday Jun 03 10:14 PM EL–FRB 2509
2. Takagi, Y.; Shikita, M.; Terasima, T.; Akaboshi, S. Specificityof radioprotective and cytotoxic effects of cysteamine in HeLaS3 cells: generation of peroxide as the mechanism of paradoxicaltoxicity. Radiat. Res. 60:292–301; 1974.
3. Koch, C. J.; Biaglow, J. E. Toxicity, radiation sensitivity modi-fication and metabolic effects of dehydroascorbate and ascorbatein mammalian cells. J. Cell Physiol. 94:299–306; 1978.
4. Link, E. M.; Riley, P. A. Role of hydrogen peroxide in the cy-totoxicity of the xanthine/xanthine oxidase system. Biochem. J.249:391–399; 1988.
5. Spitz, D. R.; Dewey, W. C.; Li, G. C. Hydrogen peroxide or heatshock induces resistance to hydrogen peroxide in Chinese ham-ster fibroblasts. J. Cell. Psysiol. 131:364–373; 1987.
6. Peak, M. J.; Jones, C. A.; Sedita, B. A.; Dudek, E. J.; Spitz,D. R.; Peak, J. G. Evidence that hydrogen peroxide generated by365-nm UVA radiation is not important in mammalian cell kill-ing. Radiat. Res. 123:220–223; 1990.
7. Biaglow, J. E.; Mitchell, J. B.; Held, K. The importance of per-oxide and superoxide in the X-ray response. Int. J. Radiat. Oncol.Biol. Phys. 22:665–669; 1992.
8. Ward, J. F.; Blakely, W. F.; Joner, E. I. Mammalian cells are notkilled by DNA single strand breaks caused by hydroxyl radicalsfrom hydrogen peroxide. Radiat. Res. 103:383; 1985.
9. Bradley, M. O.; Erickson, L. C. Comparison of the effects ofhydrogen peroxide and x-ray irradiation on toxicity, mutation,and DNA damage/repair in mammalian cells (V-79). Biochim.Biophys. Acta 654:135–141; 1981.
10. Ward, J. F.; Blakely, W. F.; Moberly, J. B. A comparison of theenzymatic repair kinetics of ionizing radiation and of hydrogenperoxide induced DNA strand breaks in V79 cells. Radiat. Res.94:629–630; 1983.
11. Prise, K. M.; Davies, S.; Michael, B. D. Cell killing and DNAdamage in Chinese hamster V79 cells treated with hydrogen per-oxide. Int. J. Radiat. Biol. 55:583–592; 1989.
12. Baker, M. A.; He, S. Elaboration of cellular DNA breaks byhydroperoxides. Free Radic. Biol. Med. 11:563–572; 1991.
13. Byrnes, R. W.; Petering, D. H. DNA strand breakage in isolatednuclei subjected to bleomycin or hydrogen peroxide. Biochem.Pharmacol. 48:575–582; 1994.
14. Bunton, C. A. Oxidation of a-diketones and a-diacids by hydro-gen peroxide. Nature 163:444; 1949.
15. Salahudeen, A. K.; Clark, E. C.; Nath, K. A. Hydrogen peroxide-induced renal injury. A protective role for pyruvate in vivo andin vitro. J. Clin. Invest. 88:1886–1893; 1991.
16. Deboer, L. W. V.; Bekx, P. A.; Han, L.; Steinke, L. Pyruvateenhances recovery of rat hearts after ischemia and reperfusionby preventing free radical generation. Am. J. Physiol. (HeartCirc. Physiol. 34) 265:H1571–H1576; 1993.
17. Nath, K. A.; Salahudeen, A. K. Autoxidation of cysteine gener-ates hydrogen peroxide: cytotoxicity and attenuation by pyru-vate. Am. J. Physiol. (Renal Fluid Electrolyte Physiol. 33)264:F306–F314; 1993.
18. Nath, K. A.; Ngo, E. O.; Hebbel, R. P.; Croatt, A. J.; Zhou, B.;Nutter, L. M. a-Ketoacids scavenge H2O2 in vitro and in vivoand reduce menadione-induced DNA injury and cytotoxicity.Am. J. Physiol. 268:C227–C236; 1995.
19. Koch, C. J.; Giandomenico, A. R. The alkaline elution techique
for measuring DNA single strand breaks: increased sensitivityand reliability. Analyt. Biochem. 220:58–65; 1994.
20. Kohn, K. W.; Friedman, C. A.; Ewig, R. A. G.; Iqbal, Z. M.DNA chain growth during replication of asynchronous L1210cells. Alkaline elution of large DNA segments from cells lysedon filters. Biochem. 13:4134–4139; 1974.
21. Flohe, L.; Gunzler, W. A. Assays of glutathione peroxidase.Meth. Enzymol. 105:114–121; 1984.
22. Matthews, R. W.; Barker, N. T.; Sangster, D. F. A comparisonof some aqueous chemical dosimeters for absorbed doses of lessthan 1000 rads. Int. J. Appl. Radiat. Isot. 29:1–8; 1978.
23. Koch, C. J.; Stobbe, C. C.; Bump, E. A. The effect on the Kmfor radiosensitization at 07C of thiol depletion by diethylmaleatepretreatment: quantitative differences found using the radiationsensitizing agent misonidazole or oxygen. Radiat. Res. 98:141–153; 1984.
24. Iliakis, G. E.; Pantelias, G. E.; Okayasu, R.; Blakely, W. F. In-duction by H2O2 of DNA and interphase chromosome damagein plateau-phase Chinese hamster ovary cells. Radiat. Res.131:192–203; 1992.
25. Ward, J. F.; Evans, J. W.; Limoli, C. L.; Calabro-Jones, P. M.Radiation and hydrogen peroxide induced free radical damageto DNA. Br. J. Cancer 55:105–112; 1987.
26. Holleman, A. F. Notice sur l’Action de l’eau oxygenee sur lesacides a-cetoniques et sur les Dicetones 1.2. Recueil TravauxChimiques Pays-Bas Belgique 23:169–171;1904.
27. Battie, M. A.; Smedley-Maclean, I. The catalytic action of cupricsalts in promoting the oxidation of fatty acids by hydrogen per-oxide. Biochem. J. 23:593–599; 1929.
28. Metzler, D. E. In: Biochemistry. The chemical reactions of livingthings. New York: Academic Press; 1977:491.
29. Kapuscinski, R. B.; Mitchell, R. Solar radiation induces sublethalinjury in escherichia coli in seawater. Appl. Environ. Microbiol.41:670–674; 1981.
30. Arana, I.; Muela, A.; Iriberri, J.; Egea, L.; Barcina, I. Role ofhydrogen peroxide in loss of culturability mediated by visiblelight in Escherichia Coli in a freshwater ecosystem. Appl. En-viron. Microbiol. 58:3903–3907; 1992.
31. Martin, S. E.; Flowers, R. S.; Ordal, Z. J. Catalase: its effect on mi-crobial enumeration. Appl. Environ. Microbiol. 32:731–734; 1976.
32. Rayman, M. K.; Aris, B.; El Derea, H. B. The effect of com-pounds which degrade hydrogen peroxide on the enumeration ofheat-stressed cells of Salmonella senftenberg. Can. J. Microbiol.24:883–885; 1978.
33. Andrae, U.; Singh, J.; Ziegler-Skylakakis, K. Pyruvate and re-lated a-ketoacids protect mammalian cells in culture against hy-drogen peroxide-induced cytotoxcity. Toxicol. Lett. 28:93–98;1985.
34. Constantopoulos, G.; Barranger, J. A. Nonenzymatic decarbox-ylation of pyruvate. Analyt. Biochem. 139:353–358; 1984.
35. von Sonntag, C., ed. The chemical basis of radiation biology.New York: Taylor and Francis; 1987.
36. Masarwa, M.; Cohen, H.; Meyerstein, D.; Hickman, D. L.;Bakac, A.; Espenson, J. H. Reactions of low-valent transition-metal complexes with hydrogen peroxide. Are they ‘‘Fenton-like’’ or not? 1. The case of Cu/aq and Cr2/aq. J. Am. Chem.Soc. 110:4293–4297; 1988.






















![유독물질(제3조 관련)1)_180330_NIER_notice...- 1 - [별표] 유독물질(제3조 관련) 고유번호 화학물질의 명칭 97-1-1 과산화 나트륨[Sodium peroxide; 1313-60-6]](https://img.pdfslide.us/doc/110x75/5f10497c7e708231d4485c87/oeeeoe3-ee-1180330niernotice-1-eoe-oeeeoe3.jpg)





