Embed Size (px)
Citation preview

Signaling and Regulation
The Id3/E47 Axis Mediates Cell-Cycle Control inHuman Pancreatic Ducts and Adenocarcinoma
Seung-Hee Lee1,7, Ergeng Hao1,7, Alice Kiselyuk2,3, James Shapiro9, David J. Shields4,8,Andrew Lowy5, Fred Levine7, and Pamela Itkin-Ansari1,6
AbstractPancreatic ductal adenocarcinoma (PDA) has a 5-year survival rate of less than 5%, and therapeutic advances
have been hampered by gaps in our understanding of cell-cycle control in the adult pancreas. Previously, wereported that basic Helix-Loop-Helix (bHLH) transcription factors regulate cell fate specification in the pancreas.In the present study, we found that a repressor of bHLH activity, Id3, was profoundly upregulated in ductal cellsin murine models of pancreatitis and pancreatic intraepithelial neoplasia (PanIN). Id3 was also pervasivelyexpressed in neoplastic lesions in human PDA in situ. We hypothesized that an imbalance in bHLH versus Idactivity controlled cell growth in PDA. Consistent with this model, cell-cycle progression in PDA cells wasimpeded by siRNA-mediated depletion of Id3 or overexpression of the bHLH protein E47. The precursors ofhuman PDA are normally quiescent duct cells which do not proliferate in response to high serum or growthfactors. The finding that Id3 was expressed in pancreatitis, as well as PDA, suggested that Id3 might induce cell-cycle entry in ducts. To test this hypothesis, primary human pancreatic duct cells were transduced with anadenovirus-expressing Id3. Remarkably, Id3 expression alone was sufficient to trigger efficient cell-cycle entry, asmanifested by expression of the proliferation markers Ki67, phospho-cyclin E, and phospho-histone H3.Collectively, the data establish dysregulation of the Id/bHLH axis as an early and sustained feature of ductalpathogenesis and mark this axis as a potential therapeutic target for intervention in pancreatitis and PDA.Mol Cancer Res; 9(6); 782–90. �2011 AACR.
Introduction
Pancreatic cancer is the fourth leading cause of cancer-related death in the United States (1–3). At the time ofdiagnosis, more than 80% of patients have locally advancedor metastatic disease. The late diagnosis, disease aggression,and scarcity of treatment options combine to give a mediansurvival time of 6 months, and a 5-year survival rate of lessthan 5% (2, 4). Cancer of the exocrine duct cells [pancreaticductal adenocarcinoma (PDA)] constitutes 90% of pan-creatic cancers (5).
Mutations and/or deletions in oncogenes and tumorsuppressor genes are common in PDA. As for many cancers,the genes most affected lie in pathways that control the cellcycle; they include Kras, p16/CDKN2A, p53, SMAD4/DPC4, and BRCA2 (6, 7). Our knowledge of the eventsthat initiate and promote tumor growth in pancreatic cancerhas increased, but the precise molecular signature remainsincomplete. A better understanding of the earliest carcino-genetic events is required to develop tools for earlier diag-nosis and to identify novel candidate molecules for targetedtherapies.The inhibitor of differentiation (Id) proteins are tran-
scriptional regulators with critical roles in normal cellgrowth and differentiation (8, 9). The primary functionof the 4 Id proteins (Id1–Id4) is to bind to and inhibit theactivity of basic helix-loop-helix (bHLH) transcriptionfactors (e.g., E47; ref. 10). The bHLH proteins activatetranscription by forming homodimers or heterodimers thatbind to regulatory "E-box" DNA sequences in target genes.Id proteins lack the basic DNA-binding domains and formnonfunctional Id/bHLH heterodimers (10) thus, seques-tering bHLH proteins.Because Id proteins antagonize the activity of many
bHLH transcription factors involved in cell-cycle control(8, 11), fluctuations in Id protein expression correlate withproliferation in many cell types. Overexpression of Id
Authors' Affiliations: Departments of 1Pediatrics and 2Bioengineering,and 3Skaggs School of Pharmacy and Pharmaceutical Sciences, Univer-sity of California San Diego; 4Departments of Pathology and 5Surgery,Division of Surgical Oncology, Moores UCSD Cancer Center; 6Develop-ment and Aging Program, 7Sanford Children's Health Research Center,Sanford-Burnham Medical Research Institute, and 8Pfizer, La Jolla, CA,and 9Department of Surgery, Faculty of Medicine and Dentistry, Universityof Alberta, Edmonton, Alberta, Canada
Note: Supplementary data for this article are available at Molecular CancerResearch Online (http://mcr.aacrjournals.org/).
Corresponding Author: Pamela Itkin-Ansari, UCSD, 9500 Gilman Drive,La Jolla, CA 92093. Phone: (858)-646-3100; Fax: (858)-795-5298; E-mail:[email protected]
doi: 10.1158/1541-7786.MCR-10-0535
�2011 American Association for Cancer Research.
MolecularCancer
Research
Mol Cancer Res; 9(6) June 2011782
Research. on February 20, 2020. © 2011 American Association for Cancermcr.aacrjournals.org Downloaded from
Published OnlineFirst April 15, 2011; DOI: 10.1158/1541-7786.MCR-10-0535

proteins may thus contribute to tumorigenesis (12, 13).Consistent with this possibility, abnormal expression levelsof Id mRNA and protein have been reported in numeroushuman cancers including those of the colon (14) and liver(15). Furthermore, ectopic Id expression in transgenic miceis associated with development of tumors including lym-phomas (16, 17) and intestinal adenomas (18). Impor-tantly, changes in Id expression have been proposed asprognostic factors for some cancers, providing furtherimpetus to understand the role played by Id proteins intumorigenesis and tumor growth (19).In the present study, we have investigated the role of Id3
in cell-cycle control of normal and pathogenic ductal cells.We identify high Id3 expression as an early event inpancreatitis and pancreatic intraepithelial neoplasia (PanIN)formation and a prominent characteristic of human PDA.In primary cells, we show that Id3 is sufficient to inducecell-cycle entry in quiescent human pancreatic ductal cells.We further show a functional requirement for Id3 inproliferation of PDA cells. Growth arrest can be inducedin PDA cells by altering the balance of Id/bHLH proteinseither by knockdown of Id3 or by overexpression of E47.The results support the hypothesis that Id3 expression playsa key role in pancreatic tumorigenesis and suggest thatrestoring the balance of the Id3/E47 axis is a promisingtherapeutic approach to PDA.
Materials and Methods
MiceMale C57BL/6J or ICR mice were purchased from
Harlan Sprague Dawley, Inc. The animals were obtainedat 5 weeks of age and housed 4 per cage on a 12:12 hourlight–dark schedule. Pancreatic duct ligation (PDL) wascarried out by ligation of the main pancreatic duct aspreviously described (20, 21). This study was approvedby IACUC at the Sanford-Burnham Institute for MedicalResearch. Pancreata were also obtained from 2- to 8-month-old Pdx-1-cre;LSL-KrasG12D mice and control Pdx-1-cremice (22) as approved by IACUC at UCSD.
Human cells and tissueThe human pancreatic cancer cell line, Panc-1 (Amer-
ican Type Culture Collection) was cultured withDulbecco's modified Eagle's media (DMEM)/10%FBS/1% Pen/Strep (Invitrogen). Panc-1/E47MER cellswere generated by transduction of Panc-1 with a retroviralvector encoding E47MER and CD25. E47MER-positivecells were selected by FACS (fluorescence-activated cellsorting) sorting for CD25 (23, 24). To induce E47activity, 4 mmol/L tamoxifen (Sigma) or vehicle (ethanol)was added to cultures of Panc-1/E47MER cells for 48hours. For siRNA studies, Id3 siRNA or scrambled siRNA(Silencer Pre-Designed & Validated siRNAs; AppliedBiosystems) was transfected with Lipofectamine RNAi-MAX (Invitrogen) for 96 hours in Panc-1 cells. Cells weretreated with BrdU (bromodeoxyuridine; 1:1,000; GEHealthcare) for 1 hour prior to harvest.
Primary adult human pancreatic exocrine tissue wasobtained from the NIH Islet Cell Resources-Administrativeand Bioinformatics Coordinating Center (ICR-ABCC),JDRF (Juvenile Diabetes Research Foundation), and Uni-versity of Alberta Islet Isolation Center. Primary cells werecultured in 5.5 mmol/L glucose RPMI/10% FBS/1% Pen/Strep (Invitrogen) on HTB9 matrix plates or collagen plates(Becton Dickinson) as previously described (24). Humanpancreatic cancer samples were obtained under IRB (Insti-tutional Review Board) approval from UCSD.
Id3 and LacZ adenovirus transductionPrimary adult human pancreatic exocrine cells were
infected with Adeno-Id3 (25) and Adeno-LacZ viruses atMOI (multiplicity of infection) ¼ 50 to 100 resulting inapproximately 50% infected cells. After 16 hours, cells werewashed with media and at 48 hours, cells were fixed foranalysis.
Histology and immunohistochemistryFor analysis of PDL, tissue proximal and distal to the
ligature was harvested 7 days following surgery and fixed in4% paraformaldehyde (USB Corp.) for 16 to 18 hours at4�C followed by freezing in OCT (optimum cutting tem-perature)-embedding media (Sakura Finetek). Pancreatafrom Pdx-1-cre;LSL-KrasG12D mice and control Pdx-1-cre mice were harvested and paraffin embedded usingstandard procedures as previously described (22). All tissuesamples were sectioned to a mean thickness of 5 mm.Cultured Panc-1 and Panc-1/E47MER cells and primaryhuman exocrine cells were fixed in 4% paraformaldehyde(USB Corp.) for 15 minutes at 4�C.All samples were permeabilized with 0.3% Triton X-100
in PBS for 15 minutes and blocked for 1 hour at roomtemperature. Cells were incubated with the following pri-mary antibodies overnight at 4�C: Id1, Id2, Id3, and Id4(Santa Cruz, Abcam, US Bio), CK19 and panCK (DAKO),E47 (Santa Cruz), mucin-1 (Muc-1; Neomarkers), Ki67(Abcam, DAKO), BrdU (GE Healthcare), phospho-cyclinE, phospho-Histone H3 (pHH3; Cell Signaling). Id3 anti-body specificity was confirmed using competing Id3 pep-tide. For fluorescent imaging, samples were incubated withAlexa 488 (Molecular Probes), Rhodamine, or Alexa 596(Jackson Immuno Research) fluor-labeled anti-mouse/rab-bit/rat/hamster and nuclear counterstained with DAPI(40,6-diamidino-2-phenylindole; Molecular Probes). Digi-tal images of fluorescently labeled and stained sections werecaptured using a fluorescence microscope with a digitalcamera (Nikon) or with a confocal microscope (Bio-RadLaboratories, Inc.) equipped with krypton/argon laser.Hematoxylin and eosin (H&E) was analyzed by brightfieldwith a conventional inverted microscope (Olympus, PlanFl40x/0.60). Image J (NIH) was employed for intensitymeasurements.
Quantitative RT-PCRTotal RNA was extracted after 4 days with RNeasy Mini
kit (Qiagen). One microgram of total RNA was prepared for
Id3/E47 Pancreatic Adenocarcinoma
www.aacrjournals.org Mol Cancer Res; 9(6) June 2011 783
Research. on February 20, 2020. © 2011 American Association for Cancermcr.aacrjournals.org Downloaded from
Published OnlineFirst April 15, 2011; DOI: 10.1158/1541-7786.MCR-10-0535
cDNA and 2 mL of cDNA was used as template for qPCR.Quantitative reverse transcriptase (RT)-PCR used theOpticon Real-Time System (BioRad), with SYBR Green(BioPioneer) for human Id3 (forward: ACTCAGCTTAG-CCAGGTGGA, reverse: AAGCTCCTTTTGTCGTT-GG) and GAPDH (glyceraldehydes-3-phosphatedehydrogenase; ref. 24), for normalization.
Statistical analysesData are presented as means � SEM. The statistical
significance of the differences between groups was analyzedby Student's t test.
Results
Id3 is highly expressed in hyperplastic ducts inpancreatitisTo determine whether the Id family of proteins plays an
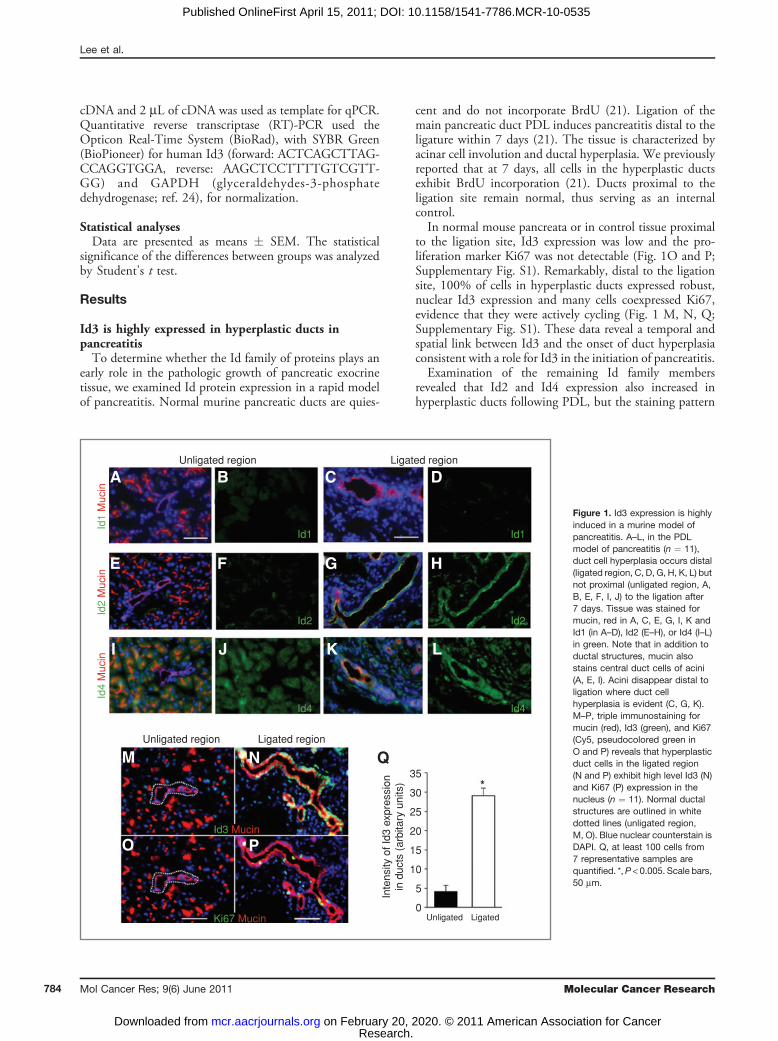
early role in the pathologic growth of pancreatic exocrinetissue, we examined Id protein expression in a rapid modelof pancreatitis. Normal murine pancreatic ducts are quies-
cent and do not incorporate BrdU (21). Ligation of themain pancreatic duct PDL induces pancreatitis distal to theligature within 7 days (21). The tissue is characterized byacinar cell involution and ductal hyperplasia. We previouslyreported that at 7 days, all cells in the hyperplastic ductsexhibit BrdU incorporation (21). Ducts proximal to theligation site remain normal, thus serving as an internalcontrol.In normal mouse pancreata or in control tissue proximal
to the ligation site, Id3 expression was low and the pro-liferation marker Ki67 was not detectable (Fig. 1O and P;Supplementary Fig. S1). Remarkably, distal to the ligationsite, 100% of cells in hyperplastic ducts expressed robust,nuclear Id3 expression and many cells coexpressed Ki67,evidence that they were actively cycling (Fig. 1 M, N, Q;Supplementary Fig. S1). These data reveal a temporal andspatial link between Id3 and the onset of duct hyperplasiaconsistent with a role for Id3 in the initiation of pancreatitis.Examination of the remaining Id family members
revealed that Id2 and Id4 expression also increased inhyperplastic ducts following PDL, but the staining pattern
Unligated region
Id1
Muc
in
Id1
Id2
Id4
Id1
Id2
Id4
Id2
Muc
inId
4 M
ucin
Id3 Mucin
Ki67 Mucin
Ligated region
Unligated region Ligated region
35
30
25
20
15
10
5
0Unligated Ligated
Inte
nsity
of I
d3 e
xpre
ssio
nin
duc
ts (
arbi
tary
uni
ts)
M
A
E
I
B
F
J
C
G
K
D
H
L
N Q
O P
Figure 1. Id3 expression is highlyinduced in a murine model ofpancreatitis. A–L, in the PDLmodel of pancreatitis (n ¼ 11),duct cell hyperplasia occurs distal(ligated region, C, D, G, H, K, L) butnot proximal (unligated region, A,B, E, F, I, J) to the ligation after7 days. Tissue was stained formucin, red in A, C, E, G, I, K andId1 (in A–D), Id2 (E–H), or Id4 (I–L)in green. Note that in addition toductal structures, mucin alsostains central duct cells of acini(A, E, I). Acini disappear distal toligation where duct cellhyperplasia is evident (C, G, K).M–P, triple immunostaining formucin (red), Id3 (green), and Ki67(Cy5, pseudocolored green inO and P) reveals that hyperplasticduct cells in the ligated region(N and P) exhibit high level Id3 (N)and Ki67 (P) expression in thenucleus (n ¼ 11). Normal ductalstructures are outlined in whitedotted lines (unligated region,M, O). Blue nuclear counterstain isDAPI. Q, at least 100 cells from7 representative samples arequantified. *,P < 0.005. Scale bars,50 mm.
Lee et al.
Mol Cancer Res; 9(6) June 2011 Molecular Cancer Research784
Research. on February 20, 2020. © 2011 American Association for Cancermcr.aacrjournals.org Downloaded from
Published OnlineFirst April 15, 2011; DOI: 10.1158/1541-7786.MCR-10-0535
suggested a diffuse subcellular distribution, rather thannuclear expression. PDL did not affect the expression ofId1 (Fig. 1A–L).
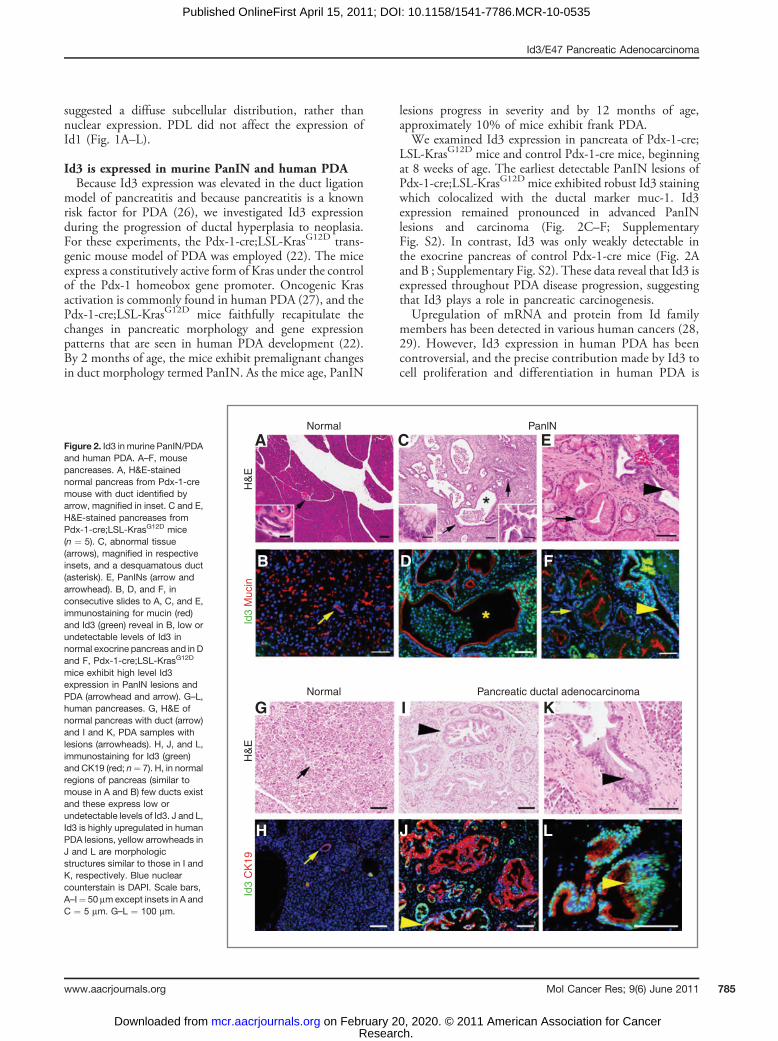
Id3 is expressed in murine PanIN and human PDABecause Id3 expression was elevated in the duct ligation
model of pancreatitis and because pancreatitis is a knownrisk factor for PDA (26), we investigated Id3 expressionduring the progression of ductal hyperplasia to neoplasia.For these experiments, the Pdx-1-cre;LSL-KrasG12D trans-genic mouse model of PDA was employed (22). The miceexpress a constitutively active form of Kras under the controlof the Pdx-1 homeobox gene promoter. Oncogenic Krasactivation is commonly found in human PDA (27), and thePdx-1-cre;LSL-KrasG12D mice faithfully recapitulate thechanges in pancreatic morphology and gene expressionpatterns that are seen in human PDA development (22).By 2 months of age, the mice exhibit premalignant changesin duct morphology termed PanIN. As the mice age, PanIN
lesions progress in severity and by 12 months of age,approximately 10% of mice exhibit frank PDA.We examined Id3 expression in pancreata of Pdx-1-cre;
LSL-KrasG12D mice and control Pdx-1-cre mice, beginningat 8 weeks of age. The earliest detectable PanIN lesions ofPdx-1-cre;LSL-KrasG12D mice exhibited robust Id3 stainingwhich colocalized with the ductal marker muc-1. Id3expression remained pronounced in advanced PanINlesions and carcinoma (Fig. 2C–F; SupplementaryFig. S2). In contrast, Id3 was only weakly detectable inthe exocrine pancreas of control Pdx-1-cre mice (Fig. 2Aand B ; Supplementary Fig. S2). These data reveal that Id3 isexpressed throughout PDA disease progression, suggestingthat Id3 plays a role in pancreatic carcinogenesis.Upregulation of mRNA and protein from Id family
members has been detected in various human cancers (28,29). However, Id3 expression in human PDA has beencontroversial, and the precise contribution made by Id3 tocell proliferation and differentiation in human PDA is
Figure 2. Id3 in murine PanIN/PDAand human PDA. A–F, mousepancreases. A, H&E-stainednormal pancreas from Pdx-1-cremouse with duct identified byarrow, magnified in inset. C and E,H&E-stained pancreases fromPdx-1-cre;LSL-KrasG12D mice(n ¼ 5). C, abnormal tissue(arrows), magnified in respectiveinsets, and a desquamatous duct(asterisk). E, PanINs (arrow andarrowhead). B, D, and F, inconsecutive slides to A, C, and E,immunostaining for mucin (red)and Id3 (green) reveal in B, low orundetectable levels of Id3 innormal exocrine pancreas and in Dand F, Pdx-1-cre;LSL-KrasG12D
mice exhibit high level Id3expression in PanIN lesions andPDA (arrowhead and arrow). G–L,human pancreases. G, H&E ofnormal pancreas with duct (arrow)and I and K, PDA samples withlesions (arrowheads). H, J, and L,immunostaining for Id3 (green)and CK19 (red; n¼ 7). H, in normalregions of pancreas (similar tomouse in A and B) few ducts existand these express low orundetectable levels of Id3. J and L,Id3 is highly upregulated in humanPDA lesions, yellow arrowheads inJ and L are morphologicstructures similar to those in I andK, respectively. Blue nuclearcounterstain is DAPI. Scale bars,A–I¼ 50 mmexcept insets in A andC ¼ 5 mm. G–L ¼ 100 mm.
Id3
Muc
inH
&E
Normal
Normal Pancreatic ductal adenocarcinoma
A
G I K
LJH
B D F
C EPanlN
H&
EId
3 C
K19
Id3/E47 Pancreatic Adenocarcinoma
www.aacrjournals.org Mol Cancer Res; 9(6) June 2011 785
Research. on February 20, 2020. © 2011 American Association for Cancermcr.aacrjournals.org Downloaded from
Published OnlineFirst April 15, 2011; DOI: 10.1158/1541-7786.MCR-10-0535
unknown (30–33). To address this question and to extendthe relevance of our observations to human disease, weanalyzed Id3 expression in human PDA tissues. Id3protein was weakly expressed or undetectable in normal,nonneoplastic areas of exocrine tissue (Fig. 2G and H;Supplementary Fig. S3). In contrast, prominent Id3staining was observed in the majority of cytokeratin 19(CK19)-positive ductal cells within neoplastic tissue areas(Fig. 2I–L; Supplementary Fig. S3), confirming that Id3expression in the human exocrine pancreas is associatedwith PDA.
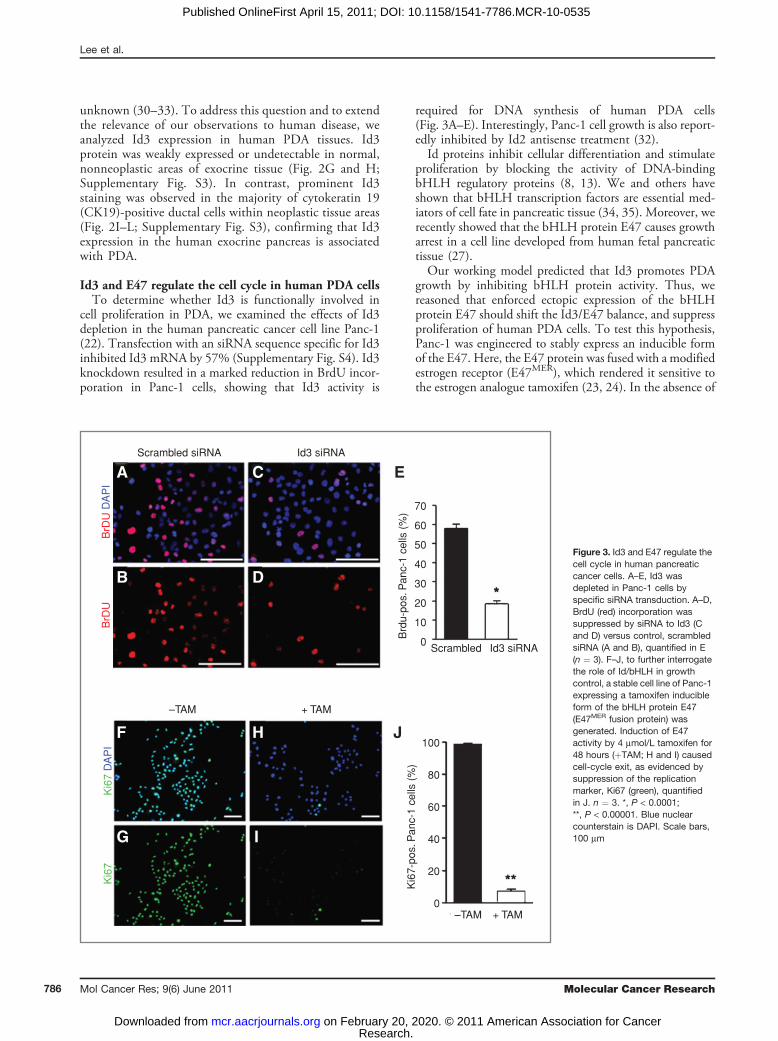
Id3 and E47 regulate the cell cycle in human PDA cellsTo determine whether Id3 is functionally involved in
cell proliferation in PDA, we examined the effects of Id3depletion in the human pancreatic cancer cell line Panc-1(22). Transfection with an siRNA sequence specific for Id3inhibited Id3 mRNA by 57% (Supplementary Fig. S4). Id3knockdown resulted in a marked reduction in BrdU incor-poration in Panc-1 cells, showing that Id3 activity is
required for DNA synthesis of human PDA cells(Fig. 3A–E). Interestingly, Panc-1 cell growth is also report-edly inhibited by Id2 antisense treatment (32).Id proteins inhibit cellular differentiation and stimulate
proliferation by blocking the activity of DNA-bindingbHLH regulatory proteins (8, 13). We and others haveshown that bHLH transcription factors are essential med-iators of cell fate in pancreatic tissue (34, 35). Moreover, werecently showed that the bHLH protein E47 causes growtharrest in a cell line developed from human fetal pancreatictissue (27).Our working model predicted that Id3 promotes PDA
growth by inhibiting bHLH protein activity. Thus, wereasoned that enforced ectopic expression of the bHLHprotein E47 should shift the Id3/E47 balance, and suppressproliferation of human PDA cells. To test this hypothesis,Panc-1 was engineered to stably express an inducible formof the E47. Here, the E47 protein was fused with a modifiedestrogen receptor (E47MER), which rendered it sensitive tothe estrogen analogue tamoxifen (23, 24). In the absence of
Scrambled siRNA Id3 siRNA
A C E
B
F
D
H J
G I
70
60
50
40
30
20
10
0
100
80
60
40
20
0
Brd
u-po
s. P
anc-
1 ce
lls (
%)
Ki6
7-po
s. P
anc-
1 ce
lls (
%)
Scrambled Id3 siRNA
+ TAM–TAM
+ TAM–TAM
BrD
UB
rDU
DA
PI
Ki6
7 D
AP
IK
i67
Figure 3. Id3 and E47 regulate thecell cycle in human pancreaticcancer cells. A–E, Id3 wasdepleted in Panc-1 cells byspecific siRNA transduction. A–D,BrdU (red) incorporation wassuppressed by siRNA to Id3 (Cand D) versus control, scrambledsiRNA (A and B), quantified in E(n ¼ 3). F–J, to further interrogatethe role of Id/bHLH in growthcontrol, a stable cell line of Panc-1expressing a tamoxifen inducibleform of the bHLH protein E47(E47MER fusion protein) wasgenerated. Induction of E47activity by 4 mmol/L tamoxifen for48 hours (þTAM; H and I) causedcell-cycle exit, as evidenced bysuppression of the replicationmarker, Ki67 (green), quantifiedin J. n ¼ 3. *, P < 0.0001;**, P < 0.00001. Blue nuclearcounterstain is DAPI. Scale bars,100 mm
Lee et al.
Mol Cancer Res; 9(6) June 2011 Molecular Cancer Research786
Research. on February 20, 2020. © 2011 American Association for Cancermcr.aacrjournals.org Downloaded from
Published OnlineFirst April 15, 2011; DOI: 10.1158/1541-7786.MCR-10-0535
tamoxifen, E47MER is sequestered in the cytoplasm of Panc-1/E47MER cells and is thus functionally inactive. Additionof tamoxifen induced translocation of E47MER to the nucleus(Supplementary Fig S5) allowing it to become transcription-ally active. Consistent with our hypothesis, tamoxifen-induced E47 activity resulted in cell-cycle exit as illustratedby downregulation of Ki67 expression (Fig. 2F–J). Takentogether, the Id3 depletion and E47 overexpression studiesestablish that the Id3/bHLH axis directly controls theproliferative status of PDA cells. These data suggest that itis the balance of activating (e.g., E47) and repressing (e.g.,Id3) helix-loop-helix factors, rather than the absolute level ofa particular factor, that regulates proliferation.
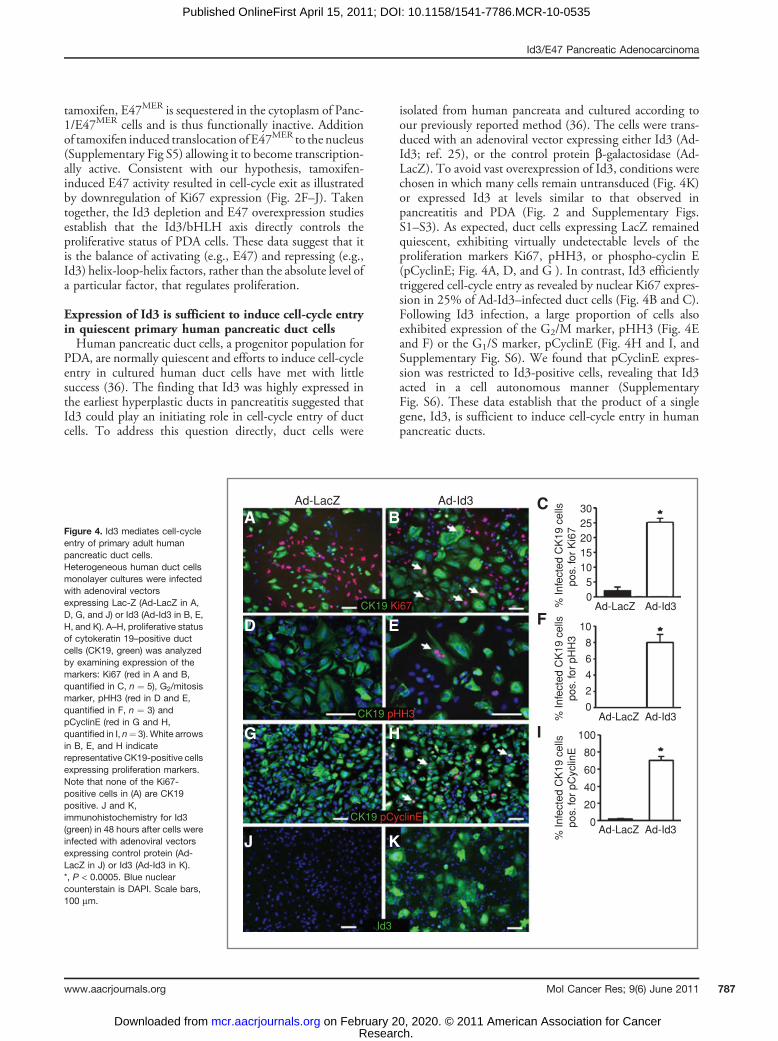
Expression of Id3 is sufficient to induce cell-cycle entryin quiescent primary human pancreatic duct cellsHuman pancreatic duct cells, a progenitor population for
PDA, are normally quiescent and efforts to induce cell-cycleentry in cultured human duct cells have met with littlesuccess (36). The finding that Id3 was highly expressed inthe earliest hyperplastic ducts in pancreatitis suggested thatId3 could play an initiating role in cell-cycle entry of ductcells. To address this question directly, duct cells were
isolated from human pancreata and cultured according toour previously reported method (36). The cells were trans-duced with an adenoviral vector expressing either Id3 (Ad-Id3; ref. 25), or the control protein b-galactosidase (Ad-LacZ). To avoid vast overexpression of Id3, conditions werechosen in which many cells remain untransduced (Fig. 4K)or expressed Id3 at levels similar to that observed inpancreatitis and PDA (Fig. 2 and Supplementary Figs.S1–S3). As expected, duct cells expressing LacZ remainedquiescent, exhibiting virtually undetectable levels of theproliferation markers Ki67, pHH3, or phospho-cyclin E(pCyclinE; Fig. 4A, D, and G ). In contrast, Id3 efficientlytriggered cell-cycle entry as revealed by nuclear Ki67 expres-sion in 25% of Ad-Id3–infected duct cells (Fig. 4B and C).Following Id3 infection, a large proportion of cells alsoexhibited expression of the G2/M marker, pHH3 (Fig. 4Eand F) or the G1/S marker, pCyclinE (Fig. 4H and I, andSupplementary Fig. S6). We found that pCyclinE expres-sion was restricted to Id3-positive cells, revealing that Id3acted in a cell autonomous manner (SupplementaryFig. S6). These data establish that the product of a singlegene, Id3, is sufficient to induce cell-cycle entry in humanpancreatic ducts.
Figure 4. Id3 mediates cell-cycleentry of primary adult humanpancreatic duct cells.Heterogeneous human duct cellsmonolayer cultures were infectedwith adenoviral vectorsexpressing Lac-Z (Ad-LacZ in A,D, G, and J) or Id3 (Ad-Id3 in B, E,H, and K). A–H, proliferative statusof cytokeratin 19–positive ductcells (CK19, green) was analyzedby examining expression of themarkers: Ki67 (red in A and B,quantified in C, n ¼ 5), G2/mitosismarker, pHH3 (red in D and E,quantified in F, n ¼ 3) andpCyclinE (red in G and H,quantified in I, n¼ 3). White arrowsin B, E, and H indicaterepresentative CK19-positive cellsexpressing proliferation markers.Note that none of the Ki67-positive cells in (A) are CK19positive. J and K,immunohistochemistry for Id3(green) in 48 hours after cells wereinfected with adenoviral vectorsexpressing control protein (Ad-LacZ in J) or Id3 (Ad-Id3 in K).*, P < 0.0005. Blue nuclearcounterstain is DAPI. Scale bars,100 mm.
Ad-LacZ
A
D
G
J
BC
F
I
E
H
K
Ad-Id3302520151050
10
8
6
4
2
0
100
80
60
40
20
0
Ad-LacZ Ad-Id3
Ad-LacZ Ad-Id3
Ad-LacZ Ad-Id3
% In
fect
ed C
K19
cel
lspo
s. fo
r K
i67
% In
fect
ed C
K19
cel
lspo
s. fo
r pH
H3
% In
fect
ed C
K19
cel
lspo
s. fo
r pC
yclin
E
CK19 Ki67
CK19 pHH3
CK19 pCyclinE
Id3
Id3/E47 Pancreatic Adenocarcinoma
www.aacrjournals.org Mol Cancer Res; 9(6) June 2011 787
Research. on February 20, 2020. © 2011 American Association for Cancermcr.aacrjournals.org Downloaded from
Published OnlineFirst April 15, 2011; DOI: 10.1158/1541-7786.MCR-10-0535

Discussion
Dysregulated expression of Id proteins has been notedin a variety of cancers, and this has prompted interest inId proteins as both diagnostic markers and as potentialtherapeutic targets (28). In the present study, we hypothe-sized that Id3 plays both early and sustained roles inpathogenic pancreatic duct cell growth. Our results areconsistent with this hypothesis and strongly suggest thatId3 activity contributes to the pronounced growth that is ahallmark of PDA.To test the hypothesis that Id3 expression and duct
hyperplasia are temporally and spatially linked, we exam-ined the PDL model of pancreatitis in which duct hyper-plasia reproducibly occurs within 7 days. Our rapid modelof pancreatitis allowed us to precisely define the temporalrelationship between Id3 expression and the onset of ductalhyperplasia. These studies show that Id3 expression isupregulated in all duct cells at the inception of hyperplasia.The fact that not all cells in hyperplastic ducts were Ki67positive, although they are BrdU positive (ref. 21), suggeststhat not all duct cells are cycling at one time.In contrast to Id3, Id1 was present in pancreatic tissue at
low tomoderate levels, but the expressionwas not affected byPDL. Id2 and Id4 expression increased after PDL, but thestaining patterns were diffuse in contrast to the nuclearstaining of Id3. One study examining Id mRNA and proteinexpression in pancreatic tissue from chronic pancreatitis(CP) patients (37) noted that Id mRNA levels were similarin CP and normal pancreas but that Id1, Id2, and Id3proteins were more highly expressed in dysplastic pancreaticducts than in normal tissue. This latter finding is similar butnot identical to our observations in murine pancreatitis.Activation of the mutant Kras oncogene is one of the
earliest transformative events in human PDA, and mutatedKras is present in up to 90% of human PDA cases (27). ThePdx-1-cre;LSL-KrasG12D transgenic mouse is an excellentexperimental model, as it faithfully recapitulates the pro-gression of PanIN lesion severity observed in the develop-ment of human PDA (22). Elevated Id3 expression wasobserved in PanIN lesions of animals at 8 weeks of age,when most lesions are at the earliest stage (1A/1B; ref. 22).Collectively, the results in the pancreatitis and PanINs placeincreased Id3 activity at a very early point in the dysregula-tion of ductal cell growth.Interestingly, only about 10% of Pdx-1-cre;LSL-
KrasG12D mice progress to invasive adenocarcinoma, sug-gesting that additional genetic or epigenetic events areneeded to reconstitute the advanced human disease. Con-sistent with this possibility, mice engineered to express botha pancreas-specific KrasG12D mutation and deletion of aconditional p16INK4A allele display accelerated developmentof PanIN lesions which progress to invasive adenocarci-noma. In contrast, mice carrying the single deletion ofp16INK4A display no pancreatic pathology (38). Transform-ing mutations in several other oncogenes and tumor sup-pressor genes are common in human PDA and accumulateas early, intermediate, and late events. Allelic deletions of
p16INK4A occur in human PanIN1A/1B lesions as often asdo Kras mutations (39), whereas mutated p53,SMAD/DPC4, or BRCA2 genes appear in more advancedPanIN lesions (40). Thus, although Kras is the initiatinggenetic defect in Pdx-1-cre;KrasG12D mice, the observationswith p16INK4A/Kras mice support the idea that additionalinsults or changes are necessary for the invasive phenotypeto develop.Understanding the relationship between Kras activation
and additional targets in development of pancreatic cancer isan area of intense study. It seems unlikely that the Idproteins are bona fide oncogenes, as there are no knowntumor-associated Id3 mutations (28, 40). However, theincreased Id3 expression found in the earliest PanIN lesionsstrongly suggests that Id3 is a downstream effector of Kras orcooperates with Kras to initiate/increase ductal proliferationPdx-1-cre;KrasG12D mice. In support of this possibility, adirect pathway between Ras activation and Id3 transcriptionhas been shown in thymocytes (41).In human PDA tissue, we detected intense nuclear
expression of Id3 in neoplastic ductal cells, confirmingthe relevance of the murine PanIN studies to humandisease. These results extend and clarify previous studies,which have identified varying degrees of Id1, Id2, and Id3expression in human PDA tissue (30–34, 37, 42). In onestudy, slightly increased Id3 in PDA tissues was observedcompared with normal pancreas. However, they observedmoderate and occasionally strong staining of Id3 expressionin normal pancreas, which we did not see (37). Severalfactors may explain the different observations includingvariations in analytic techniques (mRNA, protein, immu-nostaining methods). Our Id3 immunohistologic results arein agreement with a recent study of Id3 and p48 in PDA(43).Id1 and Id3 undoubtedly have overlapping roles in many
tissues and their expression patterns are similar (44). How-ever, many functional studies have simultaneously targetedId1 and Id3 expression, making it difficult to obtain a clearunderstanding of their individual roles. Id1/Id3 knockdownin pancreatic cancer cells inhibits their metastatic potentialin an ectopic tumor model (45). Similarly, Id1/Id3 knock-down has an inhibitory effect on gastric cancer (29). In arecent study, Id3 silencing reduced the size and weight ofectopic small cell lung cancer tumors (46). However,aberrant Id3 expression is not always associated with aproproliferative effect. For example, ovarian adenocarcino-mas display decreased Id3 expression (47), and overexpres-sion of Id3 suppresses the invasiveness of human squamouscell carcinoma (48). Thus, it seems likely that the impact ofaberrant Id protein expression on cancer growth is cell-typespecific, as is the case in development and differentiation.In functional studies, using siRNA-mediated knockdown
of Id3 in Panc-1 cells, we established that Id3 activity isrequired for DNA synthesis. Similarly, overexpression of thebHLH protein E47 also induces cell-cycle exit in Panc-1.Therefore, altering the balance between Id3 and E47 is bothnecessary and sufficient to regulate the cell cycle in PDAcells. Interestingly, Panc-1 cells carry transforming gene
Lee et al.
Mol Cancer Res; 9(6) June 2011 Molecular Cancer Research788
Research. on February 20, 2020. © 2011 American Association for Cancermcr.aacrjournals.org Downloaded from
Published OnlineFirst April 15, 2011; DOI: 10.1158/1541-7786.MCR-10-0535

mutations in p53, p16, and DPC4 in addition to Kras. Ourfinding that suppression of Id3 activity alone can inhibitcell-cycle progression in Panc-1 cells therefore implicates theId3/E47 axis as a critical convergence point for oncogenicsignals in PDA.In a previous study, we showed that normally quiescent
human pancreatic duct cells were not efficiently stimulatedto proliferate in response to high serum or growth factors(36). The observation that Id3 expression was elevated ineven the earliest pancreatitis and PanIN lesions suggestedthat Id3 might be an initiating event in duct pathogenesis.Our hypothesis predicted that Id3 overexpression would besufficient to induce cell-cycle entry in primary humanpancreatic ductal cells. Remarkably, we found this to bethe case. Ectopic expression of Id3 using adenovirus-induced primary human duct cells to enter and progressthrough the cell cycle as shown by the proliferation markersKi67, pCyclinE, and pHH3; although, we cannot rule outthe possibility that lower Id3 expression would be insuffi-cient to induce cell-cycle entry. To our knowledge, this isthe first demonstration that overexpression of a single Idprotein is sufficient to induce cell-cycle entry in quiescentprimary human ductal cells. The fact that our previousattempts to stimulate cell-cycle entry in primary humanduct cells by exposure to growth factors did not induceproliferation speaks to the potency and specificity of the Id/bHLH axis in their growth control (36).Together the gain and loss of function studies presented
here implicate dysregulation of the Id/bHLH axis as an earlyeffector of duct cell pathogenesis with continued involve-ment throughout the spectrum of PDA development.
Further investigation is necessary to determine whetherrestoring balance to the Id/bHLH axis will prove aneffective therapeutic approach to pancreatitis and PDA.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Authors' Contributions
S.-H. Lee carried out experiments and analysis. E. Hao conducted animalsurgeries. A. Kiselyuk carried out experiments and analysis. J. Shapiro providedvaluable reagents. A. Lowy and D.J. Shields provided valuable reagents, data analysis,and discussion. S.-H. Lee, F. Levine, and P. Itkin-Ansari wrote the manuscript. P.Itkin-Ansari and F. Levine designed experiments and P. Itkin-Ansari provided overalldirection for project planning and execution.
Acknowledgments
The authors thank Dr. Colleen McNamara for Adeno-Id3 and Adeno-lacZviruses, Tatsuya Kin at the University of Alberta, Betsy Holbrook at Emory, ICRCenters, particularly City of Hope, for their invaluable gift of human pancreatic cells,Dr. Rati Fotedar for helpful discussion, and Li Huang and Kaitlyn Kirk for technicalassistance.
Grant Support
This study was funded by an Academic Senate Award from UCSD (P. Itkin-Ansari), DERC award from UCSD/UCLA (P. Itkin-Ansari), CIRM (S.-H. Lee), andthe JDRF Regeneration Program (P. Itkin-Ansari and F. Levine).
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
Received November 23, 2010; revised February 23, 2011; accepted April 5, 2011;published OnlineFirst April 15, 2011.
References1. Jemal A, Siegel R, Ward E, Hao Y, Xu J, Thun MJ. Cancer statistics,
2009. CA Cancer J Clin 2009;59:225–49.2. Li J, Wientjes MG, Au JL. Pancreatic cancer: pathobiology, treatment
options, and drug delivery. AAPS J 2010;12:223–32.3. Jemal A, Siegel R, Xu J, Ward E. Cancer statistics, 2010. CA Cancer J
Clin 2010;60:277–300.4. Strosberg J, Gardner N, Kvols L. Survival and prognostic factor
analysis in patients with metastatic pancreatic endocrine carcinomas.Pancreas 2009;38:255–8.
5. Stathis A, Moore MJ. Advanced pancreatic carcinoma: current treat-ment and future challenges. Nat Rev Clin Oncol 2010;7:163–72.
6. Koliopanos A, Avgerinos C, Paraskeva C, Touloumis Z, Kelgiorgi D,Dervenis C. Molecular aspects of carcinogenesis in pancreatic can-cer. Hepatobiliary Pancreat Dis Int 2008;7:345–56.
7. Strimpakos A, Saif MW, Syrigos KN. Pancreatic cancer: from mole-cular pathogenesis to targeted therapy. Cancer Metastasis Rev2008;27:495–522.
8. Norton JD. ID helix-loop-helix proteins in cell growth, differentiationand tumorigenesis. J Cell Sci 2000;113:3897–905.
9. Zebedee Z, Hara E. Id proteins in cell cycle control and cellularsenescence. Oncogene 2001;20:8317–25.
10. Benezra R, Davis RL, Lockshon D, Turner DL, Weintraub H. Theprotein Id: a negative regulator of helix-loop-helix DNA binding pro-teins. Cell 1990;61:49–59.
11. Norton JD, Atherton GT. Coupling of cell growth control and apop-tosis functions of Id proteins. Mol Cell Biol 1998;18:2371–81.
12. Sikder HA, Devlin MK, Dunlap S, Ryu B, Alani RM. Id proteins in cellgrowth and tumorigenesis. Cancer Cell 2003;3:525–30.
13. Perk J, Iavarone A, Benezra R. Id family of helix-loop-helix proteins incancer. Nat Rev Cancer 2005;5:603–14.
14. Wilson JW, Deed RW, Inoue T, Balzi M, Becciolini A, Faraoni P, et al.Expression of Id helix-loop-helix proteins in colorectal adenocarci-noma correlates with p53 expression and mitotic index. Cancer Res2001;61:8803–10.
15. Damdinsuren B, Nagano H, Kondo M, Yamamoto H, Hiraoka N,Yamamoto T, et al. Expression of Id proteins in human hepatocellularcarcinoma: relevance to tumor dedifferentiation. Int J Oncol2005;26:319–27.
16. Kim D, Peng XC, Sun XH. Massive apoptosis of thymocytes in T-cell-deficient Id1 transgenic mice. Mol Cell Biol 1999;19:8240–53.
17. MorrowMA, Mayer EW, Perez CA, AdlamM, Siu G. Overexpression ofthe Helix-Loop-Helix protein Id2 blocks T cell development at multiplestages. Mol Immunol 1999;36:491–503.
18. Wice BM, Gordon JI. Forced expression of Id-1 in the adult mousesmall intestinal epithelium is associated with development of adeno-mas. J Biol Chem 1998;273:25310–9.
19. Lasorella A, Boldrini R, Dominici C, Donfrancesco A, Yokota Y, InserraA, et al. Id2 is critical for cellular proliferation and is the oncogeniceffector of N-myc in human neuroblastoma. Cancer Res 2002;62:301–6.
20. Sakaguchi Y, Inaba M, Kusafuka K, Okazaki K, Ikehara S. Establish-ment of animal models for three types of pancreatitis and analyses ofregeneration mechanisms. Pancreas 2006;33:371–81.
21. Demeterco C, Hao E, Lee SH, Itkin-Ansari P, Levine F. Adult humanbeta-cell neogenesis? Diabetes Obes Metab 2009;11 Suppl 4:46–53.
Id3/E47 Pancreatic Adenocarcinoma
www.aacrjournals.org Mol Cancer Res; 9(6) June 2011 789
Research. on February 20, 2020. © 2011 American Association for Cancermcr.aacrjournals.org Downloaded from
Published OnlineFirst April 15, 2011; DOI: 10.1158/1541-7786.MCR-10-0535

22. Hingorani SR, Petricoin EF, Maitra A, Rajapakse V, King C, JacobetzMA, et al. Preinvasive and invasive ductal pancreatic cancer and itsearly detection in the mouse. Cancer Cell 2003;4:437–50.
23. Schwartz R, Engel I, Fallahi-Sichani M, Petrie HT, Murre C. Geneexpression patterns define novel roles for E47 in cell cycle progres-sion, cytokine-mediated signaling, and T lineage development. ProcNatl Acad Sci U S A 2006;103:9976–81.
24. Kiselyuk A, Farber-Katz S, Cohen T, Lee SH, Geron I, Azimi B, et al.Phenothiazine neuroleptics signal to the human insulin promoter asrevealed by a novel high-throughput screen. J Biomol Screen2010;15:663–70.
25. Matsumura ME, Li F, Berthoux L, Wei B, Lobe DR, Jeon C, et al.Vascular injury induces posttranscriptional regulation of the Id3 gene:cloning of a novel Id3 isoform expressed during vascular lesionformation in rat and human atherosclerosis. Arterioscler Thromb VascBiol 2001;21:752–8.
26. Lowenfels AB, Maisonneuve P. Epidemiology and risk factors forpancreatic cancer. Best Pract Res Clin Gastroenterol 2006;20:197–209.
27. Lemoine NR. Molecular advances in pancreatic cancer. Digestion1997;58:550–6.
28. Fong S, Debs RJ, Desprez PY. Id genes and proteins as promisingtargets in cancer therapy. Trends Mol Med 2004;10:387–92.
29. Tsuchiya T, Okaji Y, Tsuno NH, Sakurai D, Tsuchiya N, Kawai K, et al.Targeting Id1 and Id3 inhibits peritoneal metastasis of gastric cancer.Cancer Sci 2005;96:784–90.
30. Lee KT, Lee YW, Lee JK, Choi SH, Rhee JC, Paik SS, et al. Over-expression of Id-1 is significantly associated with tumour angi-ogenesis in human pancreas cancers. Br J Cancer 2004;90:1198–203.
31. Iacobuzio-Donahue CA, Ashfaq R, Maitra A, Adsay NV, Shen-Ong GL,Berg K, et al. Highly expressed genes in pancreatic ductal adeno-carcinomas: a comprehensive characterization and comparison of thetranscription profiles obtained from three major technologies. CancerRes 2003;63:8614–22.
32. Kleeff J, Ishiwata T, Friess H, Buchler MW, Israel MA, Korc M. Thehelix-loop-helix protein Id2 is overexpressed in human pancreaticcancer. Cancer Res 1998;58:3769–72.
33. Grutzmann R, Boriss H, Ammerpohl O, Luttges J, Kalthoff H, Schack-ert HK, et al. Meta-analysis of microarray data on pancreatic cancerdefines a set of commonly dysregulated genes. Oncogene2005;24:5079–88.
34. Gradwohl G, Dierich A, LeMeur M, Guillemot F. neurogenin3 isrequired for the development of the four endocrine cell lineages ofthe pancreas. Proc Natl Acad Sci U S A 2000;97:1607–11.
35. Itkin-Ansari P, Marcora E, Geron I, Tyrberg B, Demeterco C, HaoE, et al. NeuroD1 in the endocrine pancreas: localization and
dual function as an activator and repressor. Dev Dyn 2005;233:946–53.
36. Hao E, Tyrberg B, Itkin-Ansari P, Lakey JR, Geron I, Monosov EZ, et al.Beta-cell differentiation from nonendocrine epithelial cells of the adulthuman pancreas. Nat Med 2006;12:310–6.
37. Maruyama H, Kleeff J, Wildi S, Friess H, Buchler MW, Israel MA,et al. Id-1 and Id-2 are overexpressed in pancreatic cancer and indysplastic lesions in chronic pancreatitis. Am J Pathol 1999;155:815–22.
38. Aguirre AJ, Bardeesy N, Sinha M, Lopez L, Tuveson DA, Horner J,et al. Activated Kras and Ink4a/Arf deficiency cooperate to producemetastatic pancreatic ductal adenocarcinoma. Genes Dev 2003;17:3112–26.
39. Schutte M, Hruban RH, Geradts J, Maynard R, Hilgers W, RabindranSK, et al. Abrogation of the Rb/p16 tumor-suppressive pathwayin virtually all pancreatic carcinomas. Cancer Res 1997;57:3126–30.
40. Hustinx SR, Cao D, Maitra A, Sato N, Martin ST, Sudhir D, et al.Differentially expressed genes in pancreatic ductal adenocarcinomasidentified through serial analysis of gene expression. Cancer Biol Ther2004;3:1254–61.
41. Bain G, Cravatt CB, Loomans C, Alberola-Ila J, Hedrick SM, Murre C.Regulation of the helix-loop- helix proteins, E2A and Id3, by the Ras-ERK MAPK cascade. Nat Immunol 2001;2:165–71.
42. Cao D, Ashfaq R, Goggins MG, Hruban RH, Kern SE, Iacobuzio-Donahue CA. Differential expression of multiple genes in associationwith MADH4/DPC4/SMAD4 inactivation in pancreatic cancer. Int JClin Exp Pathol 2008;1:510–7.
43. Dufresne M, Clerc P, Dieng M, Edir A, Couvelard A, Delisle MB, et al.Id3 modulates cellular localization of bHLH Ptf1-p48 protein. Int JCancer 2010 Sep 9. [Epub ahead of print].
44. Lim RW, Wu JM. Molecular mechanisms regulating expression andfunction of transcription regulator inhibitor of differentiation 3. ActaPharmacol Sin 2005;26:1409–20.
45. Shuno Y, Tsuno NH, Okaji Y, Tsuchiya T, Sakurai D, Nishikawa T, et al.Id1/Id3 knockdown inhibits metastatic potential of pancreatic cancer.J Surg Res 2010;161:76–82.
46. Kamalian L, Forootan SS, Bao ZZ, Zhang Y, Gosney JR, Foster CS,et al. Inhibition of tumourigenicity of small cell lung cancer cells bysuppressing Id3 expression. Int J Oncol 2010;37:595–603.
47. Arnold JM,Mok SC, Purdie D, Chenevix-Trench G. Decreased expres-sion of the Id3 gene at 1p36.1 in ovarian adenocarcinomas. Br JCancer 2001;84:352–9.
48. Higashikawa K, Yoneda S, Tobiume K, Saitoh M, Taki M, Mitani Y,et al. DeltaNp63alpha-dependent expression of Id-3 distinctivelysuppresses the invasiveness of human squamous cell carcinoma.Int J Cancer 2009;124:2837–44.
Lee et al.
Mol Cancer Res; 9(6) June 2011 Molecular Cancer Research790
Research. on February 20, 2020. © 2011 American Association for Cancermcr.aacrjournals.org Downloaded from
Published OnlineFirst April 15, 2011; DOI: 10.1158/1541-7786.MCR-10-0535

2011;9:782-790. Published OnlineFirst April 15, 2011.Mol Cancer Res Seung-Hee Lee, Ergeng Hao, Alice Kiselyuk, et al. Pancreatic Ducts and AdenocarcinomaThe Id3/E47 Axis Mediates Cell-Cycle Control in Human
Updated version
10.1158/1541-7786.MCR-10-0535doi:
Access the most recent version of this article at:
Material
Supplementary
http://mcr.aacrjournals.org/content/suppl/2011/06/17/1541-7786.MCR-10-0535.DC1
Access the most recent supplemental material at:
Cited articles
http://mcr.aacrjournals.org/content/9/6/782.full#ref-list-1
This article cites 47 articles, 13 of which you can access for free at:
Citing articles
http://mcr.aacrjournals.org/content/9/6/782.full#related-urls
This article has been cited by 1 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
SubscriptionsReprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. (CCC)Click on "Request Permissions" which will take you to the Copyright Clearance Center's
.http://mcr.aacrjournals.org/content/9/6/782To request permission to re-use all or part of this article, use this link
Research. on February 20, 2020. © 2011 American Association for Cancermcr.aacrjournals.org Downloaded from
Published OnlineFirst April 15, 2011; DOI: 10.1158/1541-7786.MCR-10-0535





![Lexer input and Output i d 3 = 0 LF w i d 3 = 0 LF w id3 = 0 while ( id3 < 10 ) id3 = 0 while ( id3 < 10 ) lexer Stream of Char-s ( lazy List[Char] ) class](https://img.pdfslide.us/doc/110x75/56649f275503460f94c3f36f/lexer-input-and-output-i-d-3-0-lf-w-i-d-3-0-lf-w-id3-0-while-id3-.jpg)

![A Comparison of Efficiency and Robustness of ID3 and C4.5 ... · of the popular ones are ID3 [1] and C4.5 [2] by J.R Quinlan. II. ID3 VS. C4.5 ID3 algorithm selects the best attribute](https://img.pdfslide.us/doc/110x75/5f0f2afd7e708231d442d273/a-comparison-of-efficiency-and-robustness-of-id3-and-c45-of-the-popular-ones.jpg)




















