Embed Size (px)
Citation preview

Ti.wie Ailr;,qens 1993. 41' 227-237 Prinrcd in Deimork . Al l righrs resrrvrd
C u p y r i x h r Munksffaard 1993
T I S S U E A N T I G E N S ISSN 0001-281j
The HLA polymorphsm of two distinctive South-American Indian tribes: The Kaingang and the Guarani
M. L. Petzl-Erler, R. Luz, V. S . Sotomaior. The HLA polymorphism of two distinctive South American Indian tribes: The Kaingang and the Guarani. Tissue Antigens 1993: 41: 227-237. 0 Munksgaard, 1993
Abstract: The HLA-A. B, C, DR and DQ antigens of 240 Kaingang and 98 Guarani individuals have been characterized. The most frequent antigens found among the Kaingang are A3 I , 2 , 24; B35, 5 I , 39.48; Cw4, 7. 3. 1; DR8, 4, 2; DQ blank, 3. In the Guarani, they are A2, 28, 31; B40. 62, "53G"; Cw3, 4; DR2, 4, 8, 6; DQ3, blank. B"53G" is an unusual antigen of the B5 cross-reactive group. DQ blank possibly corresponds to DQ4. not tested in this study, The reaction patterns of B35, B40 and DR4 indicate intra-tribal (of B35 and B40), and inter-tribal (DR4, B40 and 835) heterogeneity of these antigens. 408 Kaingang and 141 Guarani haplotypes were defined by segregation analysis. Of the commonest 10 Guarani and 9 Kaingang haplotypes, only one is shared by both tribes. Significant, positive linkage disequilibrium values for HLA-A,B; HLA- A.C; HLA-B,DR and most HLA-B,C antigen pairs were also different for the two populations. Genetic distance estimates between these two and another seven South-American Indian populations. and relative to the major human races (negroids, Caucasoids, and mongoloids) reveal a com- paratively high degree of divergence between the Kaingang and the Gua- rani, which is uncommon for Amerindian populations living close one
1 to another.
Introduction The Kaingang and the Guarani are the two major Amerindian tribes of southern Brazil. Culturally they are quite distinct from each other. The Gua- rani belong to the Tupi linguistic group. In ancient times, the Guarani occupied a wide territory in what is nowadays southern Brazil and neighboring countries. They lived close to the basins of the major rivers and were organized into several sub- groups. The Guarani were, as many still are, no- mads. Their subsistence activities included slash- and-bum agriculture, hunting and gathering. The first contacts with the Spanish and Portuguese colonizers occurred during the 16th century. Pres- ent-day Guarani live in southeastern to southern Brazil, in Paraguay, northern Argentina and Boliv- ia. The Kaingang are GO-speaking Amerindians. Before the second half of the 18th century, when
Marla Luiza Petzl-Erlsr, Roberto Loz and Vanessa Santot Sotomaior Department of Genetics, Federal University of Parana. Curitiba, Brazil
Key words: HLA - MHC - genetic pdymorphism - Amerindians - population genetics
Received 19 Janualy, accepted for publication 18 February 1993
contacts with the European colonizers became more permanent, these people occupied very wide- spread areas in southern Brazil and northern Ar- gentina. They lived mainly in the highlands and adopted a semi-nomadic way of life, having agri- culture, hunting and gathering as subsistence ac-, tivities. They practiced patrilinear exogamy, which restricted incest. According to their fathers, indi- viduals belonged to one of two subtribes, named Kame and Kadnieru. On marriage, forbidden be- tween individuals belonging to the same subtribe, women changed from their fathers' to their hus- bands' sub-tribe. Formerly, polygamy was fre- quent. In spite of the reduction in population size, today the Kaingang are still distributed over almost all of their ancient territory, living in reservation areas. In several of these areas, shared with the Guarani, Kaingang and Guarani live in different villages. Both tribes are in an advanced stage of
227

Petzl-Erler et al.
acculturation. However, they di ffere n t ia te them- selves clearly from each other, from other Amerin- dians and from neo-Brazilians, remaining predom- inan tly endogamic.
The Guarani and Kaingang were among the first Amerindians examined for genetic polymorphisms ( 1 , 2). Especially the Kaingang were studied exten- sively for plasmatic and erythrocytic polymorph- isms. during the 1960s and 1970s, by F. M. Salzano and co-workers (reviewed in (3)). Recently we started the investigation of the HLA and other genetic polymorphisms of these tribes. Results on the BF and C3 genes of the Kaingang and cDNA sequencing of several HLA class I alleles of both tribes have been reported elsewhere (4, 5).
The major histocompatibility complex (MHC) is polymorphic in all human populations and in most of the other higher vertebrates studied thus tar (6). In fact, the high degree of polymorphism of the HLA system (the human MHC) is un- equalled by any other genetic system in the human genome. The sets of HLA alleles and haplotypes and their frequencies may vary markedly between different ethnic groups (7-9). Apart from this, the degree of polymorphism is. not surprisingly, lower in genetic isolates, including the different Amerind- ian and Oceanian populations (7, 10-12). The mechanisms of generation and maintenance of the unusual polymorphism of the M H C are poorly understood. The explanations remain conjectural and controversial (13-18, and others). The careful analysis of the HLA system of different ethnic groups is an essential step to better understand its evolution and functions. In this context. the study of “primitive”. genetically isolated populations. like the Amerindians, is especially informative. At the same time, the HLA polymorphism is highly informative in the study of interrelationships be- tween human populations.
We hereby report data on the HLA polymorph- ism of the Kaingang and Guarani tribes, including the identification of their characteristic HLA anti- genic specificities, antigen, allele and haplotype fre- quencies, and linkage disequilibria. Further, the HLA-A and B allele, and HLA-A,C.B haplotype frequencies are used to evaluate intra- and inter- tribal genetic distances.
Material and methods Study populations and sample collection The Amerindian population of the State of Parana amounts altogether approximately 8000 individ- uals, distributed over 17 reservations occupying a total area of roughly 80000 hectares. Two of these reservations, Rio das Cobras (18 682 hectares) and Ivai (7306 hectares) were selected for this study
(Fig. I ) . There are approximately 1800 Kaingang and 400 Guarani in Rio das Cobras, living in differ- ent villages of the reservation. In Ivai, there are only Kaingang. numbering nearly 880 individuals. All three populations, designated here as KIV (Kaingang from Ivai), KRC (Kaingang from Rio das Cobras), and GRC (Guarani from Rio das Cobras) were investigated.
The sizes of the three population samples are: KIV, n = 127; KRC, n = 113; GRC, n=98. Data referring to each individual and his/ her relatives were collected. Since most individuals in each population are related to one another, the samples include consanguineous as well as supposedly unre- lated individuals. It was not feasible to select repre- sentative subgroups of unrelated individuals.
Every night, the blood samples obtained during the preceding day were transported by road to the laboratory in Curitiba, in boxes, at a temperature of 15°C to 20°C. Lymphocyte separation was com- pleted within 18 to 30 hours of sample collection. by ficoll-hypaque density barrier centrifugation. Lymphocytes were frozen immediately after iso- lation and were kept frozen in liquid nitrogen until serological HLA typings were completed.
HLA typing L y m p hocytoto xici t y tests with cy to fl uo roch romas- ia (19. 20) were carried out on T- and B-cell en- riched suspensions (31) with a panel of 204 antisera (146 anti-class I and 58 anti-class I l ) , plus controls. Antisera were obtained from other laboratories of several countries and from our own screening pro- gram. The antisera were selected to emphasize spe- cificities most often detected in South American Indians. The serum panel allowed the detection of the following specificities: HLA-A 1, 2, 3. 9. 23. 24.
51. 52, 8, 12. 44, 45, 13, 14, IS, 62, (63). 16. 38, 39, 17, 57, 18, 21. 22, 27, 35, 37, 40. 60, (41). (46), 47, 48, 53. w4, w6; HLA-Cwl, w2, w3. w4. w5, w6.
HLA-DQI, 2, 3. The antisera used in this study were screened with a non-Amerindian cell panel, to check their reactivity and to compare reaction patterns of Kaingang and Guarani specificities with those of non-Amerindians. For antigens in brackets, we did not know the reactivity of the used antisera, since we had only one or no reference cell exhibiting these specificities.
LO. 25, 26, 1 I , 28, 29. 30, 31, 32. 33, 36; HLA-B5,
~ 7 , ~ 8 ; HLA-DRI, 2, 3, 4, 5, 1 1 , 6, 7. 8. (9). ~ 5 2 ;
Methods of analysis Anrigen frequencies were obtained by direct count- ing. Allele frequencies were estimated by the maximum
228

HLA polymorphism of Kaingang and Guarani Amerindians
likelihood method (22, 23). Each HLA locus was considered to present k-1 co-dominant and one recessive allele. The recessive allele corresponds to the sum of unidentified alleles (blank). The fre- quencies of the co-dominant alleles are denoted by p, (i = 1, 2, ..., k-1), and the frequency of the unidentified alleles, by pblank. The formulae used for the calculations were:
k- I
C nij + (n, x hl) J = l
2N PI =
and k- I
Pbhnk = - c Pi I = I
where ni, is the number of individuals heterozygous for alleles i and j, n, is the number of individuals exhibiting only antigen i, and h, is the expected proportion of individuals homozygous for allele i
PI h, = PI + 2Pblmk
and
N = Zn,+L-++ nbldnL
where nbldnk is the number of individuals presenting none of the typable antigens at the considered lo- cus. The initial values of allele frequencies (plJ were obtained by Bernstein’s formula:
PI0 = 1 -q- and
k-l
1- I
The values of p and h were iterated for convergence until the difference between two consecutive esti- mates of each allele frequency was smaller than
Linkage disequilibrium coefficients were calcu- lated as detailed by Mattiuz et al. (24):
AIJ = E-fFT- (b+d) ( c f d )
where n is the total number of individuals pheno- typed for both loci, and a, b, c and d are the numbers of individuals having particular pheno- typic combinations of two specificities Si and S,: + +, + -, - + and - -, respectively.
Point estimates of the two-locus haplotype fre- quencies were calculated by
where p,p, is the product of the gene frequencies of the ith allele at one locus and the jlh allele at the other locus.
The hypothesis of linkage equilibrium (A =0) was tested by Fisher’s two-tailed exact probability test for 2 x 2 contingency tables.
HLA-A, C, B. DR, DQ haplotypes were identified by segregation analysis in families. Considering the low frequency of “blank” at most loci and esti- mates of linkage disequilibrium in the populations, haplotypes of several individuals in truncated nu- clear families were deduced. Haplotype frequencies were estimated by direct counting.
Genetic differences between populations were es- timated by genetic distance measurements accord- ing to Edwards (25):
- 3 8( 1 - k = l i V L G ) E’ -
U - ( 1 + ; f i ) ( l + ifi) k = I k = l
where A is the number of alleles at a given locus. pIL and P , ~ are the frequencies of the kIh allele in populations i and j. The values of Et obtained for loci HLA-A and HLA-B were added and the result used to estimate E, which expresses the genetic distance in units of genetic substitutions, using the following formula:
Amerindian populations with a sample size 5 5 0 or with 2 10% “non-Amerindian” alleles for at least one HLA locus were not used for estimates of genetic distances. Considering the technical dif- ferences of serological typings between studies, due especially to the different times at which they were conducted, the following standardization was ap- plied to all data used to estimate genetic distances: counts for splits were combined in broad speci- ficities. Thus, counts for B51 and B52; B62, B63. B75, B76 and B77; B38 and B39; and B60, B61 and B48 were combined in the broad specificities B5, B15, B16 and B40, respectively. The frequen- cies of the splits of A10, A28, B12, BI4, B17, B21, B22 were also added. B53 and the Guarani specificity B“53G” were considered equivalent. Furthermore, for Amerindian populations, counts for A30, A3 1 , A32 and A33 were combined as A19 (excludin_g A29).
In addition, HLA-A,C,B haplotype frequencies were used to estimate genetic distances.
229
Petzl-Erler et al.
i i
SOUTH
AMERICA ;lii a Figure I . M a p of S o u t h A m e r i c a showing the location of the G u a r a n i and Kaingang and of the seven o t h e r A m e r i n d i a n populations included in the genetic distance analysis.
Results Antigen and allele frequencies
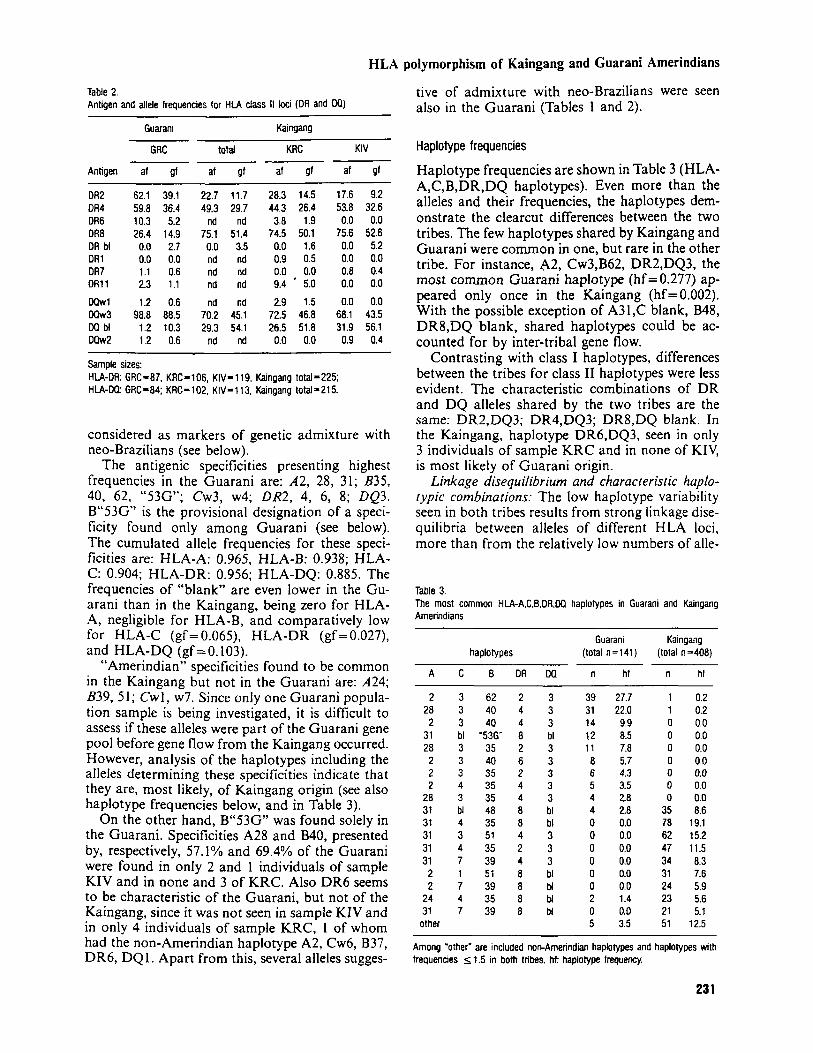
Antigen and allele frequencies for the three popula- tions are presented in Table 1 (HLA-A, B and C) and in Table 2 (HLA-DR and DQ). In both tribes, low variability was observed at all HLA loci.
The overall similarity of the frequencies of speci- ficities shared by both Kaingang population samples (KRC and KIV) indicates that no gross bias resulted from the unavoidable inclusion of consanguineous individuals in each sample. Be- sides this, the availability of two Kaingang popula- tion samples facilitates the discrimination of the specificities and haplotypes which characterize the tribe, and of those probably introduced in the population by inter-ethnic gene flow.
Several of the specificities detected among the Kaingang are shared by both population samples (KIV and KRC): A2, 24, 31; B35, 39, 40, 48, 51, 62; Cwl, w2, w3, w4, w7; DR2, 4, 8; DQ3. The alleles determining these specificities account for cumulated frequencies of 0.926 at the HLA-A lo- cus, 0.935 at HLA-B, 0.830 at HLA-C, 0.928 at HLA-DR, and 0.451 at HLA-DQ. The frequencies
230
Table 1. Antigen and allele frequencies for HIA class I lcci (A, 8 and C)
Guarani Kamgang
GRC (n-98) total (11-240) KRC (1-11113) KIV (11-127)
Antigen at gl af gf at gf at gf
A2 A24 A28 A31 A bl A3 A1 1 A26 A29 A30
835 639 840 048 851 062 8'536' 8 bl 88 813 B1 4 818 822 837 653
6w4 Bw6
Cwl CW2 cw3 cw4 cw7 C bl cw5 Cw6 CW8 'Cwll'
81.6 52.6 2.0 1.0
57.1 32.7 21.4 11.2 0.0 0.0 0.0 0.0 1.0 0.5 0.0 0.0 1.0 0.5 3.1 1.5
36.7 20.9 3.1 1.5
69.4 38.2 5.1 2.6 1.0 0.5 50.0 28.1 13.3 6.6 0.0 0.1 3.1 1.5 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
16.3 7.7 100.0 92.3
0.0 0.0 0.0 0.0
96.9 83.8 12.2 6.6 5.1 2.6 0.0 6.5 1.0 0.5 0.0 0.0 0.0 0.0 0.0 0.0
27.0 14.7 12.1 6.3
nd nd 90.4 71.6 0.0 1.7 nd nd nd nd nd nd nd nd nd nd
57.0 36.7 35.4 19.2
1.7 0.9 15.0 8.1
7.5 3.8 0.0 0.0 0.0 1.3 0.0 0.0 nd nd nd nd nd nd nd nd nd nd nd nd
47.9 28.5 90.8 71.5
18.7 9.9 3.7 1.9
26.2 14.3 60.8 37.7 35.0 19.2
1.7 13.5 0.0 0.0 nd nd nd nd nd nd
42.9 24.8
34.5 19.4 20.5 10.5 12.4 6.2 11.8 6.3 0.0 0.0 1.6 0.8
88.5 64.8 92.1 77.7 0.0 0.5 0.0 2.7 0.9 0.4 0.0 0.0 4.4 2.2 0.0 0.0 2.7 1.3 0.0 0.0 9.7 5.2 0.0 0.0 0.0 0.0 3.9 2.0
54.9 34.4 59.8 38.8 38.1 20.5 33.1 18.0
2.7 1.4 0.8 0.4 13.3 7.1 16.5 9.0 42.5 25.3 43.3 24.3
6.2 3.1 8.7 4.3 0.0 0.0 0.0 0.0 0.0 2.5 0.0 0.4 0.0 0.0 0.0 0.0 8.0 4.0 0.0 0.0 2.7 1.3 0.0 0.0 0.0 0.0 0.8 0.4 0 0 0.0 3.1 1.6 0.9 0.4 0.0 0.0 0.0 0.0 5.5 2.8
48.7 30.1 47.2 27.2 88.5 69.9 92.9 72.8
21.2 11.3 16.5 8.7 1.8 0.9 5 5 2.8
20.4 10.9 31.5 17.3 59.3 37.3 62.2 38.1 37.2 20.7 33.1 18.5
1.8 13.8 1.6 13.0 0.0 0.0 0.0 0.0 8.8 4.4 0.0 0.0 2.7 1.3 0.0 0.0 0.0 0.0 3.1 1.6
ag: antigen frequency; gf: allele (gene) frequency; nd (not determined): Ire- quencies corresponding to specificities found in only one of Me two Kaingang population samples were not determined for the whole sample. Speaficities most likely introduced in the populations by gene flow from nM1- Amerindians are listed for each locus below Me blanks (bl).
of "blank" is very low for HLA-A and HLA-B (gf 5 0.01 7), low for HLA-DR (gf = 0.035), but high for HLA-C (gf=0.135) and HLA-DQ (gf= 0.541). However, it should be noted that DQ4, common in some Amerindian Tribes (26, 27), was not tested for in this study.
In general, specificities found in only one of the Kaingang populations presented frequencies lower than shared specificities. With the exception of A28, DR6 and perhaps A30 and DR 1 1, these speci- ficities are not found in non-admixed South-Amer- ican Indian populations. Therefore, they can be
HLA polymorphism of Kaingang and Guarani Amerindians
tive of admixture with neo-Brazilians were seen also in the Guarani (Tables 1 and 2).
Table 2. Antigen and allele frequencies for HLA dass II loci (OR and w)
Guarani Kaingang
GRC total KRC KIV
Antigen af gf at gt af gf af gf ~~~ _____~
OR2 62.1 39.1 22.7 11.7 28.3 14.5 17.6 9.2 DR4 59.8 36.4 49.3 29.7 44.3 26.4 53.8 32.6 OR6 10.3 5.2 nd nd 3.8 1.9 0.0 0.0 OR8 26.4 14.9 75.1 51.4 74.5 50.1 75.6 526 OR bl 0.0 2.7 0.0 3.5 0.0 1.6 0.0 5.2 OR1 0.0 0.0 nd nd 0.9 0.5 0.0 0.0 OR7 1.1 0.6 nd nd 0.0 0.0 0.8 0.4 DR11 2.3 1.1 nd nd 9.4 ’ 5.0 0.0 0.0
Wwl 1.2 0.6 nd nd 2.9 1.5 0.0 0.0 DQw3 98.8 88.5 70.2 45.1 72.5 46.8 68.1 43.5 W M 1.2 10.3 29.3 54.1 26.5 51.8 31.9 56.1 o h 2 1 2 0.6 nd nd 0.0 0.0 0.9 0.4
Sample sizes: HLA-DR: GRC-87. KRC-106, KIV-119. Kaingang total-225: HLA-W: GRC-84: KRC-102. KIV=113. Kaingang total=215.
considered as markers of genetic admixture with neo-Brazilians (see below).
The antigenic specificities presenting highest frequencies in the Guarani are: A2, 28, 31; B35, 40, 62, “53G”; Cw3, w4; DR2, 4, 6, 8; DQ3. B“53G“ is the provisional designation of a speci- ficity found only among Guarani (see below). The cumulated allele frequencies for these speci- ficities are: HLA-A: 0.965, HLA-B: 0.938; HLA- C: 0.904; HLA-DR: 0.956; HLA-DQ: 0.885. The frequencies of “blank” are even lower in the Gu- arani than in the Kaingang, being zero for HLA- A, negligible for HLA-B, and comparatively low for HLA-C (gf = 0.069, HLA-DR (gf= 0.027), and HLA-DQ (gf = 0.103).
“Amerindian” specificities found to be common in the Kaingang but not in the Guarani are: A24; B39, 5 1 ; Cw 1, w7. Since only one Guarani popula- tion sample is being investigated, it is difficult to assess if these alleles were part of the Guarani gene pool before gene flow from the Kaingang occurred. However, analysis of the haplotypes including the alleles determining these specificities indicate that they are, most likely, of Kaingang origin (see also haplotype frequencies below, and in Table 3).
On the other hand, B“53G” was found solely in the Guarani. Specificities A28 and B40, presented by, respectively, 57.1% and 69.4% of the Guarani were found in only 2 and 1 individuals of sample KIV and in none and 3 of KRC. Also DR6 seems to be characteristic of the Guarani, but not of the Kaingang, since it was not seen in sample KIV and in only 4 individuals of sample KRC, 1 of whom had the non-Amerindian haplotype A2, Cw6, B37, DR6, DQI. Apart From this, several alleles sugges-
Haplotype frequencies
Haplotype frequencies are shown in Table 3 (HLA- A,C,B,DR,DQ haplotypes). Even more than the alleles and their frequencies, the haplotypes dem- onstrate the clearcut differences between the two tribes. The few haplotypes shared by Kaingang and Guarani were common in one, but rare in the other tribe. For instance, A2, Cw3,B62, DR2,DQ3, the most common Guarani haplotype (hf= 0.277) ap- peared only once in the Kaingang (hf= 0.002). With the possible exception of A31,C blank, B48, DR8,DQ blank, shared haplotypes could be ac- counted for by inter-tribal gene flow.
Contrasting with class I haplotypes, differences between the tribes for class I1 haplotypes were less evident. The characteristic combinations of DR and DQ alleles shared by the two tribes are the same: DR2,DQ3; DR4,DQ3; DR8,DQ blank. In the Kaingang, haplotype DR6,DQ3, seen in only 3 individuals of sample KRC and in none of KIV, is most likely of Guarani origin.
Linkage disequilibrium and characteristic huplo- rypic combinations: The low haplotype variability seen in both tribes results from strong linkage dise- quilibria between alleles of different HLA loci, more than from the relatively low numbers of alle-
Table 3. The most common HLA-A.C.B.0R.W haplotypes in Guarani and Kaingang Amerindians
Guarani Kaingang
n hf
haplotypes (total n=141) (total n-408)
A C B D R W n hf
2 3 6 2 2 3 39 27.7 1 0.2 28 3 40 4 3 31 22.0 1 0.2
0 0.0 2 3 4 0 4 3 14 9.9 0 0.0 31 bl ’536’ 8 bl 12 8.5
11 7.8 0 0.0 28 3 35 2 3 0 0.0 2 3 4 0 6 3 8 5.7 0 0.0 2 3 3 5 2 3 6 4.3
2 4 3 5 4 3 5 3.5 0 0.0 28 3 35 4 3 4 28 0 0.0 31 bl 48 8 bl 4 2.8 35 8.6 31 4 35 8 bl 0 0.0 78 19.1 31 3 51 4 3 0 0.0 62 15.2 31 4 35 2 3 0 0.0 47 11.5 31 7 39 4 3 0 0.0 34 8.3
2 1 51 8 bl 0 0.0 31 7.6 2 7 3 9 8 M 0 0.0 24 5.9
24 4 35 8 M 2 1.4 23 5.6 31 7 39 a M 0 0.0 21 5.1
other 5 3.5 51 12.5
Among ‘other‘ are included nowAmerindian haplotypes and haplotypes with trequenaes 5 1.5 in both tribes. M haplotype frequency.
23 1

Petzl-Erler et al.
Table 4. Most important' pusibve linkage disequilibria for two-point haplotypes in both tribes
Guarani Kaingang
Hap I o ty p e s hf A p hf A p
A2.851 A2.862 A28,B40 A28.835 A31,8'53G'
A2,Cwl
B35.Cw4 839.Cw7 848,C bl 051 ,Cwl B51.Cw3 8'53C.C bl
B35.DR2 B51.DR4 8'536'. DR8 B62.DR2
DR2.003 DR4.W3 DR8.W bl
0.249 0.236 0.145 0.068
0.058 0.01 0
0.063
0.072 0.240
0.107
0.101 21 .lo-' 0.112 5.9.10m5 0.077 3.0. lo-' 0.061 5.0.1 0-6
0.044 4.7.10-5 0.010 6.0.10-3
0.059 3.3.
0.061 4.9.10-7 0.134 9.6.10-7
0.092 1.1 .lo-'
0.077 0.041 1.1. lo-'
0.075 0.060 5.9.1 0-' 0.351 , 0.213 2.1 . l o - ' 0.1 90 0.1 53 4.8. lo-' 0.079 0.068 3.3. lo-' 0.091 0.066 5.0.1 0-' 0.1 31 0.096 2.3.1 0-'
0.105 0.062 3.1 0.148 0.074 3.2.10-6
0.118 0.065 1.3.10-6 0.296 0.163 3.1 . 0.506 0.228 3.5.1 0-6
' p s 0.001 in at least one of ~ I E tribes. Low frequency alleles and alleles present in only one of the two Kangang popuhtion samples were exduded from the analyses.
les. Two-loci haplotypes (A-B,A-C,B-C,B-DR and DR-DQ) showing positive linkage disequilibrium are shown in Table 4. Several of the allele pairs showing positive linkage disequilibrium seen in the Guarani or in the Kaingang are found also in other Amerindians from South America (26, 28, 29), Central America (30, 31) and North America (32-34). Furthermore, several allele pairs are found to be in linkage disequilibrium not only in Amerin- dians, but also in Caucasoids, Negroids and Mong- oloids (8). The most notable exceptions are DR2.DQ3 and DR6,DQ3. positive disequilibria shared exclusively with some Amerindians (26, 33) and B48,C blank and B51,Cw3, presenting signifi- cant positive disequilibrium only in the Kaingang.
Evidently absence of significant linkage disequi- librium does not mean that all combinations be- tween alleles occur. Linkage disequilibrium is not detected for combinations including high-fre- quency alleles, e.g. A31 in the Kaingang and Cw3 and DQ3 in the Guarani. This is best exemplified by B,C haplotypes in the Guarani. Cw3 has a very high frequency in this population (gf = 0.838) and thus most of the combinations involve this allele. B40,Cw3 and B62,Cw3 are shared by many ethnic groups; B35,Cw3 is less common in other popula- tions. On the other hand, non-random associations with the lower-frequency allele Cw4 are readily
revealed (Table 4): All Guarani Cw4 alleles occur in B35 haplotypes; the combination B35,Cw4 is found in all major racial groups (8).
Identification of the antigens and possible new antigenic specificities: The comparison of the reac- tion patterns of anti-sera, given by cells of Kain- gang, Guarani, and non-Amerindians, provided evidence for new variants of some antigens, and/or for differences (heterogeneity) between seemingly equivalent Kaingang and Guarani antigens.
Antigens of the B5 cross-reactive group (CREG): 1) B51, common among the Kaingang, gave an inconspicuous reaction pattern. 2) There were no clear differences of the patterns given by antigen B35 of Kaingang, Guarani, and non-Amerindians. A possible variant of B35 occurring in part of the Kaingang could not be confirmed, due to inconsist- ency of the alternative reaction patterns and lack of informative families for segregation analysis. Sequencing of B35 cDNA of 2 Kaingang and 1 Guarani individuals revealed two new B35 alleles among the Kaingang (B*3505 and B*3506), one of which was shared with the Guarani (B*3505) (5) . 3) An uncommon pattern was found with several Guarani cells, indicating the existence of a new antigen of the B5 CREG in this tribe. I t could be clearly differentiated from B5, from B51 and B52 (splits of BS), and from B35. It differed also from antigen B53, found in one of the Kaingang popula- tion samples and from B53 of a non-Amerindian control cell. The specificity received the provisional designation of B"53G". because its pattern re- sembled that of B53 and the molecule carried the Bw4 epitope. 4) Antigen B53 was found in 7 Kain- gang individuals from Ivai (KIV), of whom 4 were known to be related. It occurred on a Cw2-positive haplotype. Cw2 was not seen in other haplotypes of this population, and was not found in the non- admixed South-American Indian populations studied thus far. Therefore, it seems likely that this haplotype is of non-Amerindian origin. However, the possibility that it was not seen in sample KRC due to sampling bias cannot be ruled out.
Antigens of the B40 CREG: 1) The Guarani B40 can be differentiated from B60, of split of B40. Some of the anti-B40 sera failed to react with part of the B40-positive Guarani cells, which indicates heterogeneity of B40 in this tribe. In fact, two different B40 alleles (B*4003 and B*4004) were found in the Guarani (5). 2) The reaction pattern of B48 can be clearly differentiated from B60 and B61, the two splits of B40. Its pattern was also compared with a B48-positive control. Antiserum KRC-109, an anti-B48 of Kaingang origin, did not react with the control B48.
HLA-C: The presence of serological triplets for HLA-C is noteworthy. The specificity Cwl + Cw3
232

HLA polymorphism of Kaingang and Guarani Amerindians
segregates with haplotype A24,B62,DR4,DQ3, found in 4 individuals of sample KIV, of whom 2 typed as Cwl,Cw3,Cw4 (having a Cw4-positive second haplotype) and 2 as Cwl,Cw3 (having a Cw3-positive second haplotype). The combination A24,B62 occurs only in this haplotype, among KIV. The putative Cwl l (=Cwl +Cw3) specificity was recognized during the 10th International Histocompatibility Workshop and later shown to be the combination of a Cwl product and an epi- tope shared by Cw3 and B46 antigens (35).
Class I / anfigens: 1) The reaction pattern of antigen DR4 of the Guarani (DR4G) but not of the Kain- gang (DR4K) was unusual. While DR4K behaved like the DR4 found in non-Amerindian controls, DR4G exhibited unusually strong reactions with anti-DR5 (but riot anti-DR11) sera. This indi- cates that the DR4 molecules of the Kaingang and Guarani differ. 2) Anti-DQ3 and most anti-DQ1 sera reacted with all DR2-positive cells. DQ3 was assigned to the DR2 haplotype; reactivity with DQl antisera is likely due to concomitant anti- DR2 activity of many anti-DQl sera (26).
Genetic admixture
The cumulated frequencies of the non-Amerindian alleles permits a rough estimate of genetic admix- ture with neo-Brazilians. This is 0.053 for the Kain- gang and 0.025 for the Guarani. These values cor- respond to the sum of the frequencies of "non- Amerindian" alleles of the most informative locus (HLA-A, in this study). They mean that approxi- mately 5.3% of the present Kaingang and 2.50/0 of the present Guarani gene pool are of non-Amerind- ian origin.
Based on A,C,B,DR,DQ haplotypes and their frequencies, approximately 1.4% of the present Gu- arani gene pool is of Kaingang origin, and 0.5% of the Kaingang gene pool is derived from the Guarani.
Genetic distances
Genetic distances between the Kaingang, Guarani and other South-American Indian populations can be seen in Table 5 (distances based on HLA-A and HLA-B allele frequencies) and Table 6 (based on HLA-A,C,B haplotype frequencies). Table 7 lists the distances relative to the major racial groups, derived from HLA-A and -B allele frequencies. Fig. 1 shows the location of the reservation areas of Ivai and Rio das Cobras, and those of seven other Amerindian populations being considered for estimates of genetic distances. The shortest dis- tances are seen between tribes belonging to the
Table 5. Genetic distances ( x 1000) based on HLA-A and HLA-6 allele frequencies. between the Guarani, the two Kangang, other seven SouthAmerican Indian populations. and the South-American Indian mean
GRC' KIV' KRC' MAT3 PAR' ASU' WAI' KAY' UKA' WAR' SAM
GRC - 808 KIV - KRC MAT PAR ASU WAI KAY . UKA WAR
841 570 443 608 562 497 605 352 717 573 620 620 467 581 - 736 633 648 645 499 603
- 547 586 602 577 668 - 403 362 289 462
- 378 364 298 - 408 318
- 355 -
568 572 503 532 502 307 307 347 31 0
575 507 505 455 41 0 350 344 343 340 328
Populations or tribes, and references: h n g a n p l v a i (KIV). Kaingang-Rio das Cobras (KRC). Guarani-Rio das Cabras (GRC): this study. Mataco (MAT): (36); Parakard. (PAR), Asurini (ASU), Waiipi (WAI). Kayapo (KAY). Unrbu-Kaapor (UKA): (37-39); Warn (WAR): (26). SAM: average SouthAmerican Indian gene frequencies. obtained from the tribes listed here. Linguistic groups: 'Tupi. 'GC. 3Mataco, 'Warao.
Table 6. Genehc distances ( x 1000) between the Guarani. the two Kangang. and other five South-American Indian populations, based on HIA-A.C.6 hapbfype frequencies
GRC' KIV' KRC' PAR' ASU' KAY' UKA' WAR'
GRC' - 966 954 772 802 782 728 781 KIV' - 375 745 780 820 852 918 KRC' - 750 737 797 828 894 PAR' - 468 490 612 798 ASU' - 489 556 723 KAY' - 569 670 UKA' - 692
Tribes and linguistic groups: see Table 5. References: UKA. ASU, PAR: (39); KAI: (28).
same linguistic group (e.g. Urubu-Kaapor and As- urini) and/or between tribes separated by the shor- test geographic distances (e.g. Parakanl and Kaya- PO, Urubu-Kaapor and Asurini). The Guarani dif- fer least from the Parakanii, also a tribe of the Tupi linguistic group, and the Kaingang from the Kayapo, a Gt-speaking tribe. However, there is no overall correlation between genetic and linguistic affinities, a result in accordance with previous reports for the Tupi linguistic group (44,45). Inter- estingly, the greatest distance is that between the Guarani and the Kaingang, followed by that be- tween Kaingang and Mataco. These are the only three southern tribes included in the analysis. They also differ most from the South-American mean. The distance between the two Kaingang popula- tions results mainly from their different sets of non-Amerindian alleles.
233

Petzl-Erler et al.
Table 7. Genetic distances ( x 1000) based on HLA-A and HLA-B allele frequencies. between the Guarani and the Kaiigang and the major h u m races
SAM NAM OR1 CAU NEG
GRC 575 71 7 857 909 950 KIV 507 659 890 983 1070 KRC 505 61 4 809 903 1027 SAM - 393 701 a44 966 NAM - 636 746 902 OR1 - 573 727 CW - 495
GRC Guarani. Rio das Cobras; KRC: Kaingang. Rio das Cobras, KIV: Kaingang. Ivai. SAM: average SouthAmerkan lndian Qene frequencies, see Table 5 for refwences: NAM: average NorthAmerican lndian gene frequencies, obtained from the following tnbes Zuni (40), Fqaga (41), Pima (42). Cherokee (a), Hopi (321, Navap (32. 33); ORI: Orientals, CAU: Caucasoids,-NEG: Negroids (9).
Genetic distances based on haplotype frequen- cies (Table 6) show better the very close relation- ship between the two Kaingang populations. This approach measures divergence due to recombi- nation, in addition to that resulting from mutation and other evolutionary factors, reflecting better the degree of divergence over a shorter period of time, being less informative for more divergent groups. Thus , it is not surprising to see that the distance between the two Kaingang populations, which are not isolated genetically one from another, is lower than that between any other pair of populations, when haplotype data are used (Table 6). On the other hand, the largest distances are observed again between the Kaingang and the Guarani, reflecting the high degree of dissimilarity between these tribes.
Discussion This investigation of the HLA polymorphism of the Kaingang and Guarani is one of a series of studies to analyze the MHC and other genes, gen- etic systems, and DNA markers of these two Amer- indian Tribes of Southern Brazil. Results on the C3 and BF genes, the MHC class I11 region, and sequencing of several HLA class I alleles have been reported elsewhere [4, 46 (submitted), 51.
The antigenic specificities detected belong to the groups already well-known to occur in South American Indians. However, the analyses of the serological reaction patterns and their comparison between Kaingang, Guarani and non- Amerindian controls revealed differences between seemingly equivalent antigens and the presence of specificities unusual in the other major ethnic groups. Thus, the DR4 antigens of both tribes differ one from
another; the Guarani DR4 is characterized by reac- tions with anti-DR4 and most anti-DR5 sera, while the Kaingang DR4 does not differ from the non- Amerindian controls. The reaction patterns of anti- gens DR2, DR6 and DR8 do not differ between Guarani, Kaingang and non-Amerindian controls. Antigen DR3/4/5 or DR3 x 6, described in Pim- ans, Hopi and Navajo from North-America (27, 47) was not seen in these tribes. All HLA-A and HLA-C specificities, as well as B51, B62 and B39 were inconspicuous. Specificities Cw 1 + Cw3 segre- gated together in haplotype A24,B62,DR4,DQ3 in sample KIV, a fact reminiscent of the putative Cwll antigen, recently shown to be the combi- nation of a Cwl product and an epitope shared by Cw3 and B46 antigens (35). The occurrence of B48 was suggested by a short reaction pattern with anti-B40 sera and confirmed by DNA sequencing of one Kaingang individual (5). B48 is common among North-American Indians and Eskimos (33, 48) but until recently (5 , 49) was not identified in South-American Indians. However, B48 may be more usual among Amerindians than previously reported, since B40 and B48 molecules present strong serological cross-reactivity, which may re- sult in misclassification of B48 as B40. B"53G" is an unusual specificity found exclusively in the Guarani and can be differentiated from B5 and its splits and from B35 and B53, being more closely related to 853, and carrying epitope Bw4. I t differs from antigen B5 of the Pimans (50), also related to B53, since B"53G" did not react with anti-B5 sera. B35 and B40 gave variable reaction patterns. Heterogeneity of B35 has been suggested also in other Amerindians (5 1.52). The serological hetero- geneity of B35 and B40 might result from antigenic differences of the molecules encoded by different alleles sequenced from individuals of these two tribes: B*4003 and B*4004 in the Guarani; B*3505 (Kaingang and Guarani) and B*3506 (Kaingang) (5) . The genetic determination of these different serological patterns could not be tested by segre- gation analysis due to lack of informative families. The evaluation of the serological patterns in com- parison to the DNA sequences and deduced pro- tein structure is of interest, because the products of recently sequenced South-Amerindian HLA-B alleles belonging to the same serologic group (e.g. B35) differ in their peptide-binding site ( 5 , 49), in residues of the beta-strands thought to be inaccess- ible to antibodies and of prime functional import- ance.
The detailed serological analyses show that at least HLA-B and HLA-DR are more heterogen- eous in Amerindians than previously reported. In fact, sequencing of HLA class I alleles of only a few selected Guarani, Kaingang and Waorani
234

HLA polymorphism of Kaingang and Guarani Amerindians
individuals already revealed differences between HLA-B alleles whose products could not be clearly distinguished by serology (5,49). A similar hetero- geneity will probably be found also for HLA-DR. We expect that population analyses at the DNA level will detect additional HLA-B and HLA-DR alleles in these and other Amerindian tribes.
The Guarani and Kaingang differ clearly in their HLA polymorphism, a fact evidenced at several levels: 1) Existence of specificities characteristic of one but not the other tribe: A28, B40, B“53G”, and DR6 in the Guarani, and A24, B39, B5L, Cwl, and Cw7 in the Kaingang: 2) Frequencies corresponding to shared specificities are clearly dif- ferent. 3) All but one of the HLA-A,C,B,DR,DQ haplotypes differ between Guarani and Kaingang. 4) The majority of allele pairs in linkage disequilib- rium differ too.’On the other hand, the combi- nations of shared class I1 (DR and DQ) alleles are the same and correspond to those described in other South-American Indians: DR2,DQ3 and DR6,DQ3 were for the first time described in the Venezuelan Warao (26). Conversely, in the North- American Pimans, DR2 is associated to DQI (27), the combination most usual in other examined eth- nic groups. DR4,DQ3 is the commonest DR4-DQ combination in all major human races (8). The DQ specificity of the Guarani and Kaingang DR8 haplotypes was not determined, but DQ I , DQ2 and DQ3 were excluded; in other Amerindian and non-Amerindian populations DQ4, not tested in this study, is associated with DR8.
The above discussed differences between the Gu- arani and the Kaingang reflect the great genetic distance between these tribes, which becomes ap- parent by the comparison of genetic distances be- tween these two ethnic groups and other South- American and North-American Indians, and rela- tive to the major racial groups. Based on the HLA polymorphism, the greatest divergence between nine South-American Indian populations is seen between the Kaingang and the Guarani, followed by that between the Kaingang and the Mataco. It is noteworthy that the genetic distances between the Kaingang and the Guarani, and the Kaingang and Mataco are greater than those between the Kaingang and the tribes from northern South- America. Thus the genetic distances between these three southern tribes do not correlate with geo- - graphic distance, but rather with cultural and linguistic distinctiveness. Such a high level of dis- similarity between populations living within the same geographic area is uncommon (3). The dis- tinctiveness of these tribes provides a model to test hypotheses regarding the evolution of the HLA polymorphism.
An estimate of the degree of genetic admixture
was obtained by direct gene counting. This simple approach, applicable only to data from poly- morphisms presenting private alleles of given eth- nic groups, and to populations exhibiting a charac- teristic polymorphism, results, in our view, in less distorted estimates than those obtained by methods which require knowledge of the gene frequencies of the original populations. Two sources of bias may influence this estimate: first, it may be dis- torted because of the inclusion of consanguineous individuals in the samples. Second, it may be an underestimate, because some of the specificities found in the Amerindians, e.g. A2 and B35, are common also in other populations, including neo- Brazilians. Nevertheless, the inspection of haplo- types indicates that at least this second source of bias is negligible, because the vast majority of the specificities shared with neo-Brazilians are, in the Kaingang and Guarani, inserted in haplotypes characteristic of Amerindians and rare in neo-Bra- zilians (Petzl-Erler, unpublished data). The degree of non-Amerindian admixture was estimated as 5.3% for the Kaingang and 2.5% for the Guarani. The analysis of 58 South-American Indian popula- tions, based on 3 to 13 genetic markers, showed that 42 (72.40/) of them had less than 10% of admixture with non-Amerindians (3). For the Kaingang, this was estimated as 8.8%. No previous data are avaiiable for the Guarani. The clearcut differences of the HLA haplotypes of the Kaingang and the Guarani allowed an estimate of the intertri- bal gene flow: the Guarani have 1.4‘K of Kaingang admixture, while the Kaingang present 0.5”/0 of Guarani admixture. Taken together, these esti- mates indicate that both tribes still retain more than 90% of their ancient genetic constitution.
Acknowledgments We thank the Guarani and Kaingang populations for their cooperation; Misses Maria Cecilia Trip- pia, Elizabeth Setim,, Iara M. Pereira, Teresa H. Nakamura and Mr. Fernando Sedor for help dur- ing the field work; Miss Marisol Ferraz and Mrs Sbnia C. M. Costa for technical assistance; and the FundaCGo Nacional do indio for providing facilities at the posts. This work was supported by grants from the Conselho Nacional de Desenvolvi- mento Cientifico e Tecnologico (CNPq).
Rafsrancst
I . Ribeiro L. Berardinelli W, Roiter M. Grupo sanguine0 dos indios Guaranys. Arch Med Leg Idmt 1935: 5: 59-65.
2. Junqueira PC, Wishart PJ, Ottensooser E Pasqualin R. Fernandez PL, Kalmus H. The Diego blood factor in Brazi- lian Indians. Nurure 1956: 177: 41.
235

Petzl-Erler et al.
3.
4.
5.
6.
7.
8.
9.
10.
11.
Salzano FM, Callegari-Jacques SM. South American Indi- ans. A Case Study in Evolution. Oxford: Clarendon Press, 1988. Guerra LK, Rehfeldt IR. Lima MT. Petzl-Erler ML, Messi- as IJT. BF and C3 genetic polymorphism in Kaingang Indians from Southern Brazil. Hum Hered 1992: 42:
Belich MP. Madrigal JA. Hildebrand WH, et al. Unusual HLA-B alleles in two tribes of Brazilian Indians. Nature 1992: 357: 326-329. Klein J. Natural History of the Major Histocompatibility Complex. New York: Wiley, 1986. Dausset J, Colombani J (eds). Histocompatibility Testing 1972. Copcnhagen: Munksgaard, 1973. Baur MP, Neugebauer M, Albert ED. Reference tables of two-locus haplotype frequencies for all MHC marker loci. In: Albert ED, Baur MP, Mayr WR. eds. Histocompatibility Testing 1984. Berlin: Springer-Verlag. 1984: 677. Baur MP, Neugebauer M. Deppe H, el al. Population analysis on the basis of deduced haplotypes from random families. In Albert ED, Baur MP, Mayr WR. eds. Histocom- patibility Tisting 1984. Berlin: Springer-Verlag. 1984:
Kostyu DD. Amos B. Mysteries of the Amerindians. Tissue Anfigens 1981: 16: I I 1-123. Kostyu D. Dawson D, Ciftan E. et. al. HLA in two islands of French Polynesia. Tissw Anrizens 1984: 23: 217-228.
153-1 56.
333-341.
26. Laynsse 2. Heinen HD, Balbas 0. Garcia E. Stoikow Z. Unique HLA-DR/DQ associations revealed by family studies in Warao Amerindians. Haplotype and homozygos- ity frequencies. Hum lmmunol 1988: 23: 45-57.
27. Williams RC, McAuley JE. HLA class I 1 variation in the Gila River Indian community of Arizona: Alleles, haplo- types. and a high frequency epitope at the HLA-DR locus. Hum Immwol 1992 33: 29-38.
28. Black FL. Interrelationships between Amerindian Tribes of lower Amazonia as manifest by HLA haplotype disequilib- ria. Am J Hum Genet 1984: 36: 1318-1331.
29. Haas EJC, Salzano FM, Araujo HA, et al. HLA antigens and other genetic markers in the Mapuche Indians of Ar- gentina. Hum Hered 1985: 35: 306313.
30. Corky RB, Specs EK, Cabrcra GM. Swanson JL, Amos DB. HL-A antigens of the Guatemalan Ixils. In: Terasaki PI. ed. Histocompatibility Testing 1970. Copenhagen: Munksgaard, 1970: pp. 351-357.
31. Gorodezky C, Castro-Escobar LE, Escobar-Guitiirrez A. The HLA system in the prevalent Mexican Indian group: the Nahuas. Tissue Antigens 1985: 25: 38-46.
32. Williams RC, Morse HG, Bonnell MD. Rate RG, Kuberski TT. The HLA loci of the Hopi and Navajo. Am J Phys Antliropol 1981: 56: 291-296.
33. Troup GM, khanfield MS, Singaraju CH, et al. Study of HLA alloantigens of the Navajo Indians of North America. 11. HLA-A, B. C, DR and other genetic markers. Tissue
12. Crane G. Bnthia K, HoneymanM, et al. HLA studies of highland and coastal New Guineans. Him lmniunul 1985: 12: 247-260.
Anrigens 1982: 20: 339-35 I . 34. Amos DB. Kostyu DD, Stewart AD, Ward FE. North
American Indians: Pima lndians. In: Aizawa M. Natori T. 13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
Black FL,-Salzano FM. Evidence for heterosis in the HLA system. Arrt J Him Gener 1981: 33: 894499. Gustafsson K. Wiman K. Emmoth E. et al. Mutations and selection in the generation of' class I I histocompatibility antigen polymorphism. EMBO J 1984: 3: 1655-1661. Klein J. Origin of major histocompatibility complex poly- morphism: The trans-species hypothesis. Hurlitin b m u - no/o.q- 1957: 1 9 155-162. Hedrick PW. Evolulion of HLA: Possible explanations for the deliciency of homozygotes in two populations. Him Hcretl 1990: 40: 21 3-220. Takahata N. Nei M. Allelic genealogy under overdominant and frequency-dependent selection and polymorphism of major histocompatibility complex loci. Generics 1990: 124:
Klein J. Klein D (eds). Molecular Evolution of the Major Histocompatibility Complex. Berlin: Springer-Verlag. 1991. Terasaki PI. McClelland JD. Microdroplet assay of human serum cytotoxins. Nurure 1964: 204: 998-1000. Bodmer WF. Bodmer J. Cytofluorochromasia. In: Ray JG. Hare DB, Pedersen PD, Mullally DI, eds. NlAlD Manual of Tissue Typing Techniques. DREW Publ. No. (NIH) 78-
Werner C. Klouda PT, C o m a MC, Vassali P. Jeannet M. Isolation of B- and T-lymphocytes by nylon fiber columns. ?'isme Anrigens 1977: 9: 227-229. Ceppclini R, Siniscalco M, Smith CAB. The estimation of gene frequencies in a random-mating population. Ann Hum Genet 1955: 20: 97. Yasuda N, Kimura M. A gene-counting method of maxi- mum likelihood for estimating gene frequencies in ABO and ABO-like systems. Ann Hum Gener 1968: 31: 409. Mattiuz PL, Ihde D, Piazza R, Cepellini R. Bodmer WE New approaches to the population genetic and segregation analysis of the HLA system. In: Terasaki PI, ed. Histocom- patibility Testing 1970. Copenhagen: Munksgaard. 1970: pp. 193-105. Edwards AWF. Distances between populations on the basis of gene frequencies. Biometrics I97 1 : 27: 573-88 I .
967-978.
545. 1976: pp. 31-34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
Wakisaka A. Konoeda Y, eds. Proceedings of the Third Asia-Oceania Histocompatibility Workshop and Confer- ence. Sapporo: Hokkaiclo University Press, 1986: pp.
Zemmour J. Gumperz J. Marsh SGE, Ward F. Williams R. Parham P. HLA-CwI I is a combination of Cwl and a public epitope of HLA-Cw3 and HLA-846. In: Tsuji K. Aizawa M. Sasazuki T, eds. HLA 1991. Oxford: Oxford University Press, 1992. Vullo CM. Celis EM. Serra HM, Rierd CM. Study of HLA system in a Mataco population: A geographically isolated American Indian tribe. Tissuc Anrigcns 1984: 23: 33-40. Black FL. Salzano FM. Layrisse 2, Franco MHLP. Harris NS, Weimer TA. Restriction and persistence of polymorph- isms of HLA and other genetic traits in the Parakani Indians of Brazil. Ant J Phys An!/irupo/ 1980: 52: 119-132. Black FL, Salzano FM, Berman LL. et al. Failure of linguistic relationships to predict genetic distances between the WaiIpi and other tribes of lower Amazonia. Am J Phys Anlhropol 1983: 60: 327-335. Black FL. Santos SEB, Salzano FM, et al. Genetic variation within the Tupi linguistic group: New data on three Ama- zonian tribes. Ann Hum Biol 1988: 15: 337-351. Troup GM. Harvey RL. Walford RL, Smith GS, Sturgeon P. Analysis of the HL-A, erythrocyte and gamma globulin antigen systems in the Zuni Indians of New Mexico. In: Dausset J, Colombani J. eds. Histocompatibility Testing 1972. Copenhagen: Munksgaard, 1973. Perkins HA, Payne RO. Kidd KK. Huestis DW. HLA-A and Gm typing of Papago Indians. In: Dausset J. Colom- bani J. eds. Histocompatibility Testing 1972. Copenhagen: Munksgaard, 1973: 333-338. Spees EK, Kostyu DD. Elston RC. Amos DB. HL-A pro- tiles of the Pima Indians of Arizona. In: Dausset J, Colom- bani J, eds. Histocompatibility Testing 1971. Copenhagen: Munksgaard. 1973: 345-349. Spees EK, Woodbury MA. Gottsch LG, Nicely PR. Amos DB. Heterogeneity of HL-A specificities in the Cherokee Indians. In: Kissmeyer-Nielsen F. ed. Histocompatibility Testing 1975. Copenhagen: Munksgaard. 1975: 219-222.
287-290.

HLA polymorphism of Kaingang and Guarani Amerindians
44. Callegari-Jacques S, Salzano FM. Genetic variation within two linguistic Amerindian groups: relationships to geogra- phy and population size. Am J Phys Anthropol 1989: 79:
45. Aguiar GFS. Ethnohistory, intertribal relationships, and genetic diversity among Amazonian Indians. Hum Biol 1991: 63: 743-162.
46. Weg-Remers S. Brenden M, Schwarz E, et al. MHC class 111 genetics in three South American Indian populations from Brazil (submitted).
47. Ward FE, Kostyu DD, Stewart AD, Amos DB. HLA anti- gens in the Piina Indians. In: Aizawa M, Natori T, Waki- saka A, Konoeda Y, eds. P r d i n g s of the Third Asia- Oceania Histocompatibility Workshop and Conference. Sapporo: Hokkaido University Press, 1,9861 pp. 53 1-534.
48. Petersen GM. Ward JI, Terasaki PI, et al. Genetic poly- morphisms in Southwest Alaskan Eskimos. Hum Hered 1991: 41: 236-247.
49. Watkins DI, McAdam SN, Liu X, et al. New recombinant HLA-B alleles in a tribe of South American Amerindians
3 13-320.
indicate rapid evolution of MHC class I loci. Nurure 1992: 357: 329-332.
SO. Williams RC. McAuley JE. HLA class I variation con- trolled for genetic admixture in the Gila River Indian com- munity of Arizona: A model for the Paleo-Indians. Hum Immunol 1992: 33: 39-46.
51. Kostyu DD, Amos DB, Hinostroza S. An analysis of the 4c complex of HL-A based on Indian populations. Tissue Antigens 1975: 5: 420430.
52. Johnson AH, Noreen H, Spees EK, et al. The distribution of HLA antigens in the Motilones Indians of Venezuela. Tirsue Anrigens 1978: 12: 163-169.
Address: Dr. Maria Luka Petzl-Erler Department of Genetics Federal University of Parana Caixa Postal 19071 81531-970 Curitiba, Brazil Fax: (41) 266-2042 Tel: (41) 366-3144, ext 255 or 259
237





























