Embed Size (px)
Citation preview

THE HEAD PATTERN IN AMBLYSTOMA STUDIED BY VITAL STAINING AND TRANSPLANTATION
METHODS
ESTHER CARPENTER Oaborn zoo+togiod Laboratories, Yale University '.
TEN TEXT FIQUEES AND O N E PLATE ("WELAVE FIQUBES)
INTRODUCTION
The experiments presented here were undertaken for the purpose of 1) locating in the neurula stage of Amblystoma punctatum the ectoderm involved in the formation of the prospective head pattern and 2) determining the relative potencies at this stage of prospective balancer, lens and nasal regions.
Part I deals with the results obtained by methods of local vital staining of prospective head region of embryos of Amblystoma punctatum. Four cases are given in detail and the composite diagrams of the slit blastopore and open medullary plate stages compared with other similar work on urodeles.
Part I1 deals with transplantation experiments carried out between stained and unstained embryos of the same age for the purpose of testing the relative potencies of prospective balancer, lens and nasal ectoderm and the extent to which the ectoderm of these regions can be interchanged without inter- fering with the development of normal structures. The de- gree of determination was assumed to be indicated by the fate
'The experiments cited were performed in the spring of 1929, 1930 and 1931 and presented in partial fulfillment of the requirements for the degree of Doctor of Philosophy at Yale University.
103
* Contributions from the Department of Zoology, Smith College, no. 186.
THE J O ~ N A L OF EXPEB,IYENTAL zoboay, VOL 76, NO. 1

104 ESTHER CARPENTER
of the graft when placed in an abnormal position on the head. The normality of the organ developing in situ was judged by comparison with that of the side of the animal on which no operation had been performed.
ACKNOWLEDGMENTS
I am deeply indebted to Dr. Ross. G. Harrison for sug- gestions received during the course of this investigation. I wish also to thank Mr. E. B. Bosworth for giving me the benefit of his experience in vital staining technique.
PART I MATERIALS AND METHODS
The results presented are based on local vital staining ex- periments on embryos of Amblystoma punctatum.
The methods used were essentially those described by Vogt ('25). Bits of agar containing either Nile blue sulfate or neutral red were applied during the gastrula stage with the vitelline membrane intact. After staining the embryos were changed to fresh tap water and camera lucida drawings made to show the position and extent of the stained areas. If the embryo was still round, lateral and anterior views were ob- tained by means of a Zeiss prism rotator. In most cases the stained areas were apparent until the gill swellings and balancers became evident and in some instances could be followed up to stage 39. After this, fading was very rapid, probably due both to thinning out of the stained material as the animal increased in size and to metabolic activity.
Except for a few cases in which stained areas were large and numerous there was no evidence of abnormality due to toxic effect of the dye. It seems doubtful that even these deaths can be attributed to the staining, since embryos may be stained in toto with either dye and develop normally unless overstained (Detwiler, '17).
THE HEAD PATTERN IN AMBYLSTOMA 105
DESCRIPTION OF FIGURES
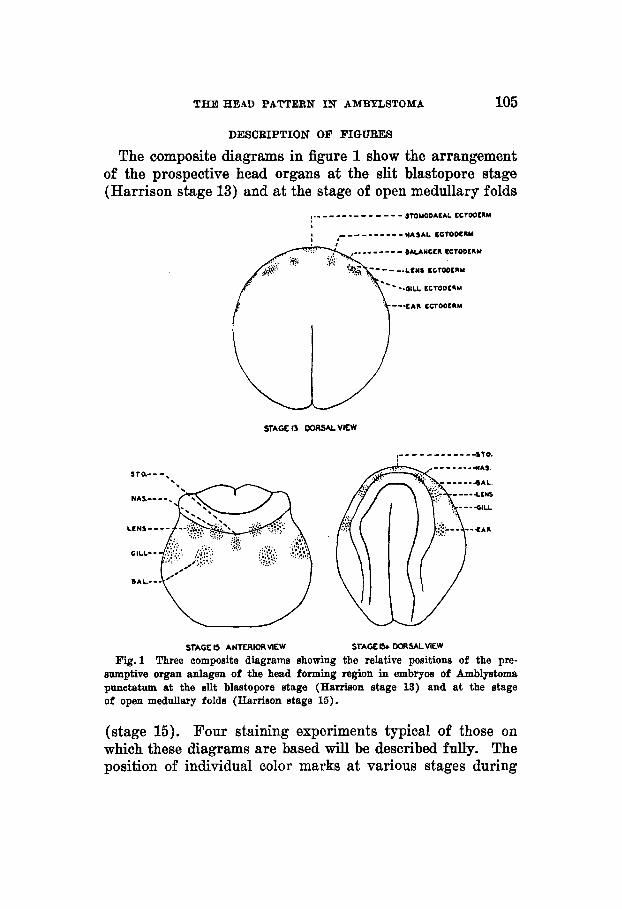
The composite diagrams in figure 1 show the arrangement of the prospective head organs at the slit blastopore stage (Harrison stage 13) and at the stage of open medullary folds
.
STAGE 6 ANTERIORVIEW STAGE Is, DORSAL VIEW
Fig. 1 Three aompoaite diagrams showing the relative positions of the pre- sumptive organ anlagen of the head forming region in embryos of Amblyatoma pruretatum at the slit blastopore stage (Harrison atage 13) and at the stage of open medullary folda (Harrison stage 15).
(stage 15). Four staining experiments typical of those on which these diagrams are based will be described fully. The position of individual color marks at various stages during

106 ESTHER CARPENTER
development may be followed by reference to figures 2 to 5. The numbers at the lower left in each case indicate the stage of development.
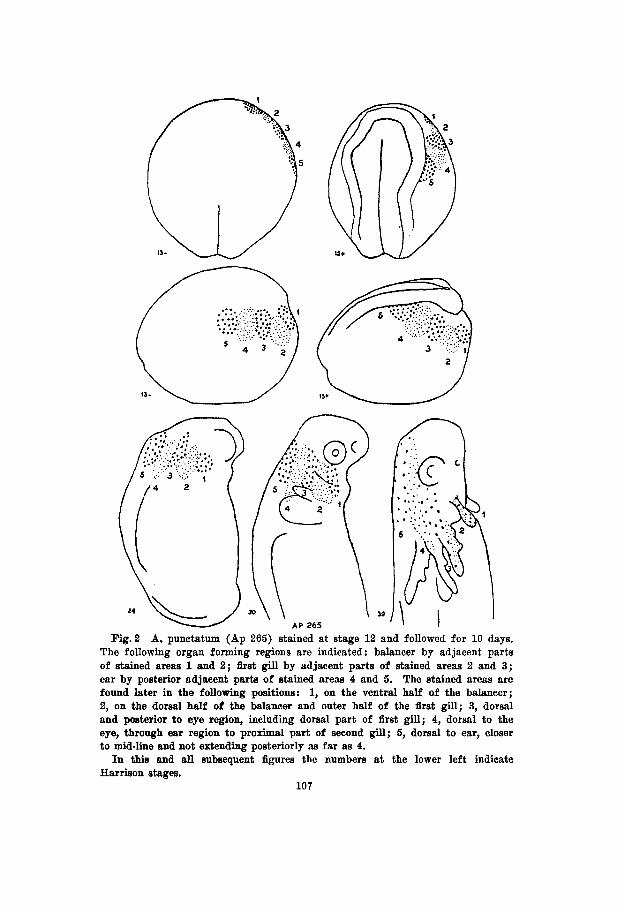
The embryo shown in figure 2 (Ap 265) was stained during the yolk plug stage and the marks followed for 10 days there- after. A few hours later at stage 13-, as seen from the dorsal surface, the five stained areas formed a border at the outer edge of the right anterior quadrant of the embryo. Viewed from the side areas 2 and 4 could be seen to extend somewhat more ventrally than areas 1, 3 and 5. At stage 15+ the medullary fold had appeared adjacent to mark 5 and the transverse fold lay posterior to the other stains as one would expect if in urodeles generally the prospective anterior part of the forebrain lies directly opposite the slit blastopore (Vogt, '29). During the movements concomitant with the formation of the medullary plate, adjacent material is also displaced. The greatest change in position is shown by stained area 5 which is in contact with the fold, while the least is shown by stained area 1. As a result of the different degrees of displacement incident to the formation of the head of the embryo the more posterior stained areas come to lie dorsal to the anterior ones as shown in stages 24 and 30 of figure 2. Thus it comes about that in the fully formed embryo the prospective epidermis which at stage 13- lies at the outer edge of the right anterior quadrant as seen from the dorsal surface, extends from the dorsal mid-line and anterior ear region to the ventral part of the balancer including the area between the eye and the base of the gills. Areas 4 and 5 move anteriorly as the animal straightens out, while the more ventrally lying areas 1 , 2 and 3 become involved in the forma- tion of the balancer and the first two gills.
Reference to figure 2 shows that the material for the balancer lies in the region indicated by area 1 and the adjacent part of 2. The first gill is formed partially from the ventral part of area 3 and the ventral-posterior part of area 2. The prospective ear ectoderm lies in the adjacent parts of areas 4 and 5 at a short distance from the outer edge of the medul- lary fold.
39 AP 265
Fig .2 A. punctatum (Ap 265) stained at stage 12 and followed for 10 days. The following organ forming regions are indicated : balancer by adjacent parts of stained areas 1 and 2; &st gill by adjacent parts of stained areas 2 and 3; ear by posterior adjacent parts of stained areas 4 and 5. The stained areas are found later in the following positions: 1, on the ventral half of the balancer; 2, on the dorsal half of the balancer and outer half of the first gill; 3, dorsal and posterior to eye region, including dorsal part of first gill; 4, dorsal to the eye, through ear region to proximal part of second gill; 5, dorsal to ear, closer to mid-line and not extending posteriorly as far as 4. In this and all subsequent figures the numbers at the lower left indicate
Harrison stages. 107
108 ESTHER CARPENTER
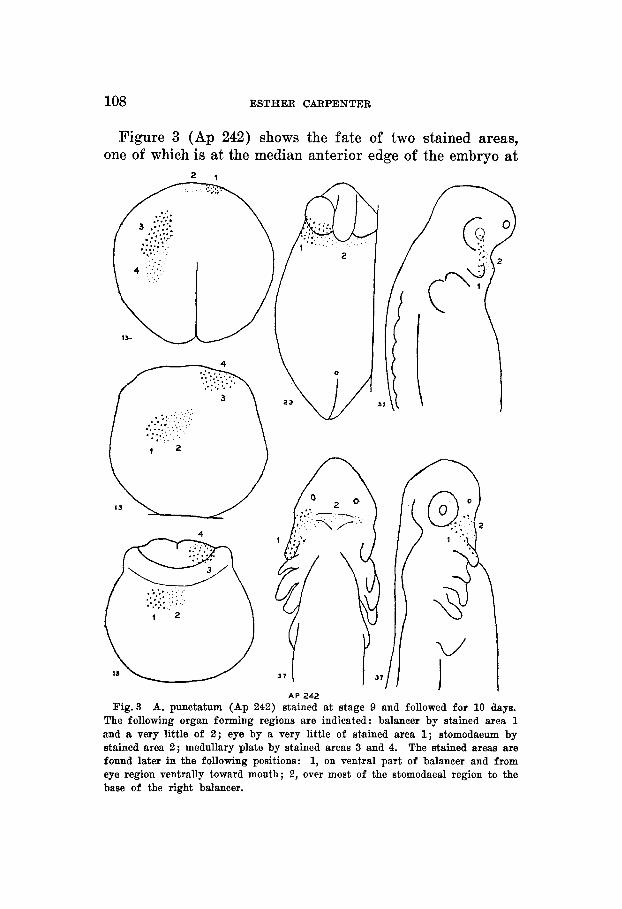
A P 242 Fig.3 A. punctatum (Ap 242) stained at stage 9 and followed for 10 days.
The following organ forming regions are indicated: balancer by stained area 1 and a very little of 2 ; eye by a very little of stained area 1; stomodaeum by stained area 2 ; medullary plate by stained areas 3 and 4. The stained areas are found later in the following positions: 1, on ventral part of balancer and from eye region ventrally toward mouth; 2, over most of the stomodaeal region to the base of the right balancer.
THE HEAD PATTERN I N AMBYLSTOMA 109
stage 13- and the other adjacent as seen in a dorsal view. These fll in the gap in the previous figure. Area 2 covers the stomodaeal region and the base of the right balancer. Area 1 covers the ventral surface of the balancer and extends to the postero-ventral part of the eye.
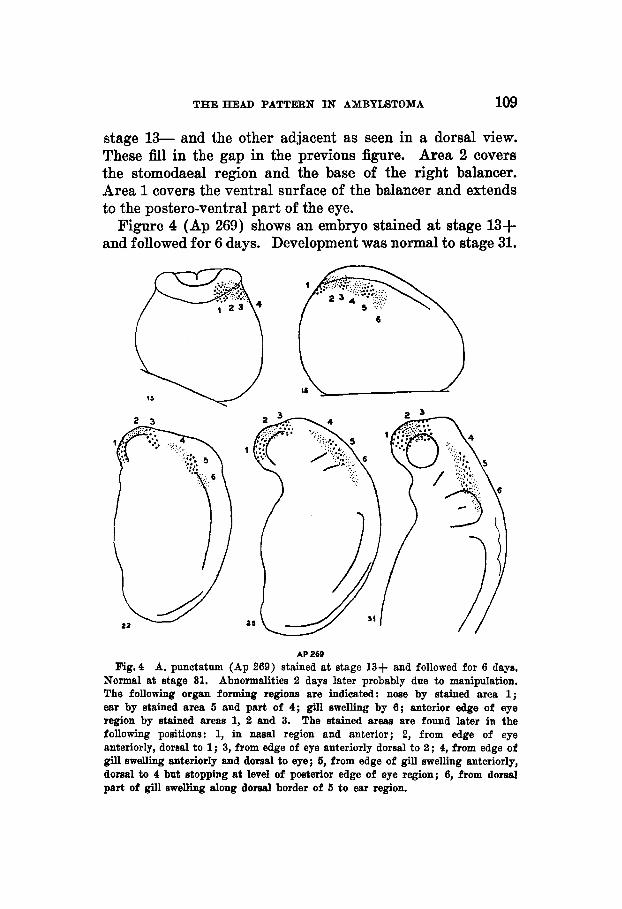
Figure 4 (Ap 269) shows an embryo stained at stage 13+ and followed for 6 days. Development was normal to stage 31.
AP 269 Fig.4 A. punotatum (Ap 269) stained at stage 13+ and followed for 6 days,
Normal at stage 31. Abnormalities 2 days later probably due to manipulation. The following organ forming regions are indicated: nose by stained area 1; ear by stained area 5 and part of 4; gill swelling by 6 ; anterior edge of eye region by stained area8 1, 2 and 3. The stained areas are found later in the following positions: 1, in nasal region and anterior; 2, from edge of eye anteriorly, domal t o 1; 3, from edge of eye anteriorly dorsal to 2; 4, from edge of gill swelling anteriorly and dorsal to eye; 5, from edge of gill swelling anteriorly, dorsal to 4 but stopping at level of posterior edge of eye region; 6, from dorsal part of gill swelling along dorsal border of 5 to ear region.
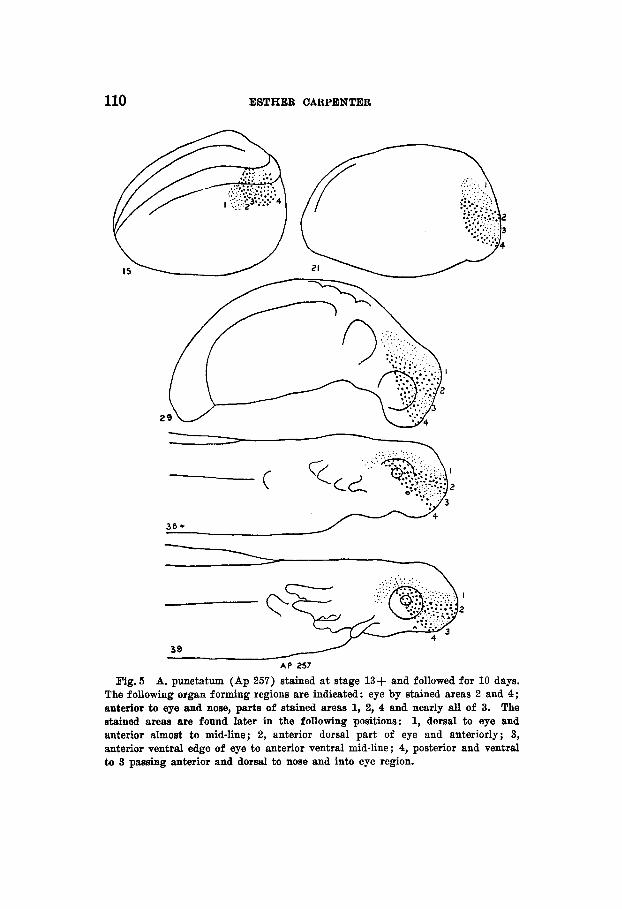
110 ESTHER CARPENTER
39 A P 257
Fig. 5 A. punctatum (Ap 257) stained a t stage 13+ and followed for 10 days. The following organ forming regions are indicated: eye by stained areas 2 and 4; anterior to eye and nose, parts of stained areas 1, 2, 4 and nearly all of 3. The stained areas are found later in the following positions: 1, dorsal to eye and anterior almost to mid-line; 2, anterior dorsal part of eye and anteriorly; 3, anterior ventral edge of eye to anterior ventral mid-line; 4, posterior and ventral to 3 passing anterior and dorsal to nose and into eye region.

THEHEAD PATTERN I N AMBPLSTOMA 111
Two days later the embryo appeared somewhat abnormal and was discarded. Bismarck brown was used instead of neutral red and may have been slightly toxic. The most interesting result here is the evidence that the nasal material (mark 1) lies at least partially on the outer side of the medullary fold while the lens material, ventral to marks 1, 2 and 3, seems not to encroach on the fold at all. Prospective ear ectoderm likewise lies off the medullary fold and a short distance from it. The change in shape of marks 4, 5 and 6 to produce a shingle effect shows that considerable extension of the ecto- derm in an anterior posterior direction takes place in the region dorsal to the balancer.
N
STAGE 13 S T A G E 2 2
Fig. 6 Diagrams to show the relative change in position of balancer, lens and nasal ectoderm of A. punctatum between stages 13 and 22. The points of the triangle are approximately in the oenter of the areas involved in the formation of them three organa.
Figure 5 (Ap 257) shows an embryo stained at stage 13+ and followed for 10 days. From the fate of the originally ventral parts of marks 2 , 3 and 4 it is apparent that the lens material lies off the medullary fold and that the prospective head ectoderm close to the fold and anterior to the ear region rotates through an angle of more than 90" between the stage when the folds first become visible and the stages in which the head is apparent as such. Some indication of this relative shift in position of balancer, lens and nasal ectoderm during neurulation is given by the diagrams in figure 6.

112 ESTHER CARPENTER
DISCUSSION OF FIGURES
The composite diagrams of figure 1 represent the arrange- ment in the slit blastopore and neural fold stages (stages 13 and 15) of the ectoderm involved in the formation of nose, lens, ear, balancer, gills and stomodaeum as found by local vital staining experiments on embryos of Amblystoma punc- tatum such as those cited (figs. 2 to 5).
In stage 13, an approximate sphere, the stomodaeal ecto- derm is found at the anterior median border of the embryo as seen from above (mark 2, fig. 3). The transverse neural fold forms a short distance back of this region. As the embryo develops the stomodaeal ectoderm becomes greatly stretched in a lateral direction. If one considers the stomodaeal ecto- derm as located at 90" at stage 13, the position of the other regions may be indicated as follows: Balancer 55 to 65" at the margin (areas 1 and 2, fig. 2) ; gills 30 to 45" at the margin (parts of 2, 3 and 4, fig. 2 ) ; ear 15 to 20" at the margin (areas 4 and 5, fig. 2) ; nasal placode somewhat back from the anterior margin at 70 to 75" (area 1, fig. 4).
In the neural plate stage the epidermal material extends to about the middle of the part outlined as neural fold. Thus the nasal ectoderm lies closest to the boundary between epi- dermal and medullary material, the lens lies at the outer border of the fold and the ear ectoderm just off the latter.
With the elevation of the neural folds and their approach toward the mid-line, adjacent material is carried along as shown by the relative positions of the ear ectoderm in stages 13 and 15+ (fig. 1). Due to more rapid growth in the dorsal region, as the folds fuse and head formation begins, the anterior part of the neural folds and the immediately adjacent material move forward and downward (ventrally) thus giving the originally ventral material a relative movement in a posterior dorsal direction. In this way the balancer and gill ectoderm finally assume their typical relation to the eye and ear (figs. 2 and 6).

THE HEAD PATTERN IN AMBYLSTOMA 113
COMPARISON WITH RESULTS ON OTHER URODELES
Comparison of the neural fold stage of Amblystoma shown in figure 1 with that of Riihlich ('31) for Triton and of Schechtman ('32) for Triturus shows a similar general ar- rangement of anlagen but with variations in detail. Rohlich's diagram indicates that in Triton nose, lens and ear ectoderm lie in contact with the neural fold and extend equal distances from it at this stage. In Amblystoma the nasal material reaches up on the outer side of the fold a short distance and the ear ectoderm does not quite come in contact with the fold. Although the method is such that it would be difficult to de- limit exactly the amount of ectoderm involved in each case, the lens material in Amblystoma undoubtedly extends further ventrally than the nasal at stage 15 and covers a larger area (fig. 4). It does not seem to extend up on the fold at all. The position of lens material thus ahecks with that given by both Manchot and Rohlich for Triton. Schechtman shows prospective lens material extending on to the outer side of the medullary fold in Triturus torosus while the position of nasal ectoderm seems to approximate more closely that found in Amblystoma than that given by Rohlich for Triton. Ea; ectoderm seems to be farther anterior in Amblystoma than in Triton. Figures for Triturus (Schechtman, '32, plate 36) indicate that here as in Amblystoma the ear ectoderm does not extend up to the edge of the neural fold.
The movements of material of particular interest in the stages studied are those brought about by the dorsal con- vergence of the neural folds together with the adjacent pre- sumptive epidermis and the subsequent forward and down- ward growth of the head region anteriorly in such a way that material at the center of the transverse fold comes to lie just in front of the stomodaeum and ventral to the eye, i.e., a shifting of the material nearest the dorsal mid-line in an anterior and ventral direction while that originally ventral moves caudally and dorsally (see fig. 6). These movements refer to the ectoderm of the prospective head region only. Balancer and gill ectoderm are only partially visible from the

114 ESTHER CARPENTER
dorsal surface of stages 13 and 15. The gill material covers a larger area and extends further ventrally. Both regions move in a dorsal and posterior direction as the neural folds close. If one considers the stomodaeal region as a relatively fixed point, the balancer and gill regions may be pictured as rotating about this point from their marginal position as shown in figure 1 toward the mid-line. The gill material thus shows a greater change in position than the balancer.
The diagram of the slit-blastopore stage may be considered to partially fill the gap between the stages worked on by Vogt and those worked on by Rohlich since the results in general are in harmony with those obtained by both these in- vestigators.
PART I1 INTRODUCTION
In the experiments to be described the potencies of balancer, lens and nasal ectoderm were tested in the neurula stage by two methods: 1) transplantation to other parts of the em- bryo, and 2) extirpation with replacement either partially or completely by material from some other region of the embryo. Donor and host embryos were always of the same age. It would be desirable for comparison to carry out a similar series of experiments in which the age of donor and host varied. I n the case of the balancer and nose, rotation of the grafts might be expected to yield still further informa- tion concerning the order of events.
MATERIALS AND METHODS
All the operations were carried out on embryos of Amblystoma punctatum between stages 13 and 26. All transplantations were reciprocal, each embryo serving both as donor and as host. I n all cases the embryos of a pair were from the same bunch of eggs. Ectoderm only was trans- planted. Previous to the operations half of the embryos were stained in toto in Nile blue sulfate, in order that the actual extent of the grafted region might be more easily ascertained.

THE HEAD PATTERN I N AMBYLSTOMA 115
Unless otherwise stated the graft was placed on the host in normal orientation with respect to the medullary plate. Camera lucida drawings were made at intervals during de- velopment.
Operations were carried out in dilute Ringer’s solution and incisions made by means of glass needles and hair loops. Care was taken to keep the embryos cool until the grafts were well healed.
Although a limited number were allowed to develop for 6 weeks, most of the embryos were fixed in Bouin’s fluid be- tween stages 35 and 42. For histological purposes good re- sults were obtained by clearing in aniline oil and wintergreen and staining with Mallory’s triple connective tissue stain.
The fate of the graft when placed in an atypical position on the head was used as an indication of the degree of de- termination at the time of transplantation of the graft, and the normality of the organ developing in situ was determined by comparison with that of the side of the animal on which no operation had been performed.
The results presented are based on fifty-seven animals, in which either in the normal or the transplanted region the balancer was involved twenty-two times, the lens twenty-one times and the nasal placode forty-four times.
OPERATIONS INVOLVING BALANCER ECTODERM
I n the earliest case (NL2 blue) shown in figure 7, the balancer was inhibited by nasal ectoderm placed in this region at stage 13+. No balancer developed from the prospective balancer ectoderm transplanted, nor did it inhibit the growth of a normal nasal placode in the host (fig. 11). In eight additional cases operated on from stages 16 to 26 the balancer was suppressed by either lens or nasal ectoderm. In six of these there was a slight thickening of the basement membrane in the balancer region. In the other two animals nasal placodes had developed from the transplants. One is shown in figure 10.
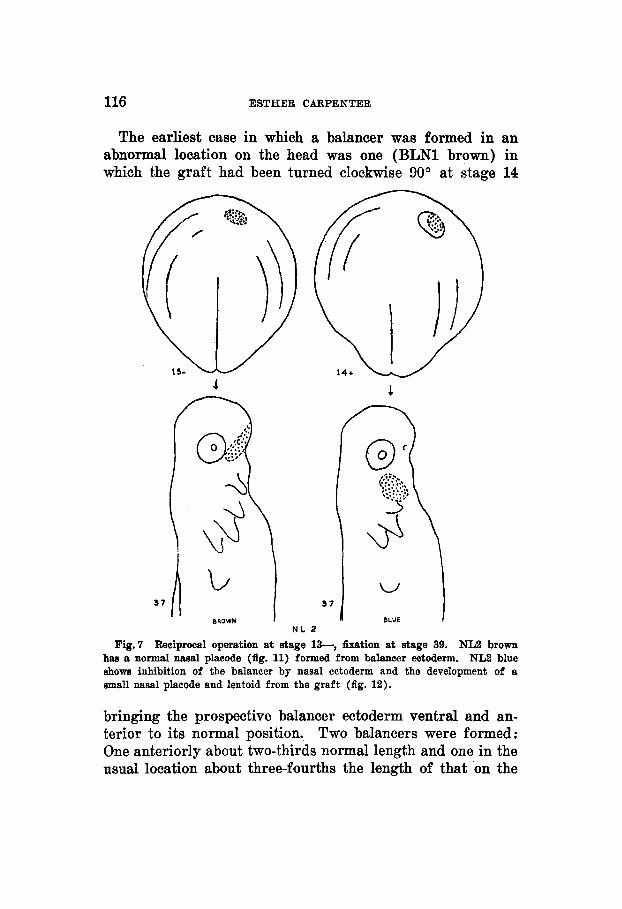
116 ESTHER UARPENTER
The earliest case in which a balancer was formed in an abnormal location on the head was one (BLN1 brown) in which the graft had been turned clockwise 90° at stage 14
37 A 37
N L 2
Fig.7 Reciprocal operation at stage 13-, fixcttion at stage 39. NL2 brown has a normal nasal placode (fig. 11) formed from balancer ectoderm. NL2 blue shows inhibition of the balancer by nasal ectoderm and the development of a small nasal placode and lentoid from the graft (fig. 12).
bringing the prospective balancer ectoderm ventral and an- terior to its normal position. Two balancers were formed: One anteriorly about two-thirds normal length and one in the usual location about three-fourths the length of that on the
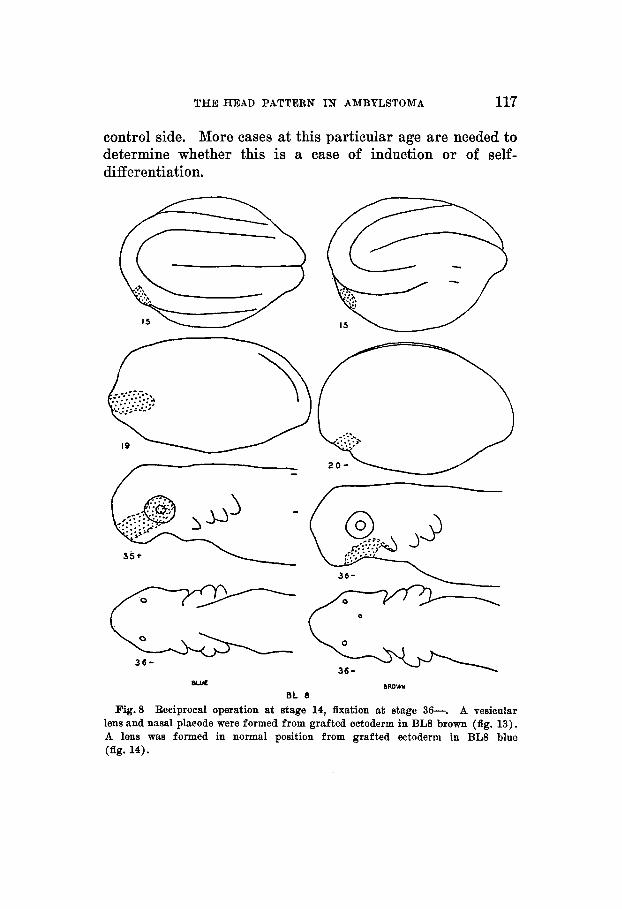
THE HEAD PATTERN IN AMBYLSTOMA 117
control side. More cases at this particular age are needed to determine whether this is a case of induction or of self- differentiation.
36-
mow BWI 0L 8
Fig.8 Reciprocal operation at stage 14, fixation at stage 36-. A vesicular lens and nasal placode were formed from grafted ectoderm in BL8 brown (fig. 13). A lens was formed in normal position from grafted ectoderm in BL8 blue (fig. 14).

118 ESTHER CARPENTER
When prospective balancer ectoderm was transplanted to the nasal region at stages 19- and 20 (BN38 and BN31) self- differentiation did occur as indicated in figures 9, 19 and 20.
B N 38 Fig.9 Reciprocal operation at stage 19-. BN38 brown, fixed at stage 40,
shows a double balancer formed by ectoderm grafted anterior to the nasal region (fig. 19). BN38 blue, fixed at stage 4 6 , shows retarded development of the first gill.
In the first (figs. 9 and 19) histological examination revealed two balancers fused together. The membranes were distinctly blue and the more posterior one contained a good number

THE HEAD PATTERN IN AMBYLSTOMA 119
of mesenchyme cells. The scarcity of mesenchyme cells in the other seemed to be due to loss of connection with the source of supply. I n the second case (fig. 20) the balancer after growing part way out was either partially resorbed or else a piece broke off and the end healed. The differentiation of balancers in this region indicates a lack of localization of specific potencies in the mesectodermal substrate at the time of operation.
These experiments differ from Mangold's ('31) in the method employed and from Harrison's ('25) in that the operations were always between animals of the same age. The latter obtained independent differentiation of balancer ectoderm when transplanted from stages with open neural folds to the gill region of hosts varying in age from stages 20 to 24. More experiments are needed to determine whether a balancer can be produced when grafts are made between animals both of which are in the early medullary plate stage.
OPERSTIONS INVOLVING LENS ECTODERM
The earliest case (BL8) of self-differentiation of lens ectoderm in an abnormal location was operated upon at stage 14 (fig. 8). From the graft placed ventral to the normal region a vesicular lens developed (fig. 13). This was in contact with the optic stalk anteriorly. Of eight other cases ranging in age from 13+ to 25+ at the time of operation in only one instance (NL31 blue) was a well-developed lens obtained. Instead there were found lentoids, masses of cells of a doubtful nature and difficult of interpretation. A true segregation of lens material (Lillie, '29) has not occurred up to the time of fusion of the neural folds.
I n four cases the lens region was completely covered by other head ectoderm. From ectoderm ventral to the eye transplanted at stage 14- (NL11 blue) an apparently normal lens was obtained. At stage 14 a similar operation (BL8 blue) resulted in a lens normal in structure but slightly smaller than the control (fig. 14). Ectoderm of the dorsal balancer region grafted over the lens area at stage 14f (BL9
T H E JOURNAL O F EXPERIMENTAL Z&LOGY, VOL. 75, 10. 1

120 ESTHER CARPENTER
blue) produced a fairly typical lens of nearly normal size with B cavity in the center (fig. 16). Ectoclerm of the balancer ~ e g i o n transplanted a t stage 20 (RL21 blue) resulted in a lens normal in stage of development but markedly smaller than the control (figs. 21 and 2 2 ) . The optic cup was also smaller.
EL 21
Fig. 10 Reciprocal operation a t stage 20, fixation a t stage 40. BL21 brown shows inhibition of the balancer by lens ectoderm and the formation of a small nasal placodr by the graft . BL21 blue shows a small balancer developed from ecto- derm grafted anterior to the normal one. A lens developed in normal position fiom the grafted ectoderm (fig. 2 2 ) .

THE HEAD PATTERN IN AMBYLSTOMA 121
Seven cases in which lens ectoderm was partially replaced by other head ectoderm at stages varying from 13+ to 23 indicate that this has little or no effect before the closure of the neural folds (NL2, BN4 and BL30). After this point there is a marked reduction in the size of the developing lens (NL28, NL41, BL19 and BL32).
OPERATIONS INVOLVING NASAL ECTODERM
This series contains the largest number of cases and shows the most constant results. Good nasal placodes were obtained in eighteen cases in which the prospective nasal ectoderm was transplanted to other parts of the head region at stage 14 or later up to stage 26+. I n one additional case (NL2 blue) in which the operation was performed at stage 13+ a somewhat doubtful nasal placode was produced anterior to the balancer region (fig. 12). The three earliest cases (BL8 brown, BL9 brown and BN4 blue) in which clearly recog- nizable nasal placodes were formed were operated on at stages 14, 14f and 15 respectively and are shown in section in figures 13,15 and 17. In the first of these a vesicular lens also developed from the graft which was placed ventral to the posterior part of the optic cup. In the second a mass of nerve tissue is in contact with the placode developing dorsal to the balancer region. The third reveals only a nasal placode developing from the graft which is between the eye and the balancer. No nerve tissue was found near it. Nerve tissue apparently developing from the graft was found in four (NL37 brown, BN38 blue, BN35 brown and BN40 brown) of the remaining fifteen cases. These were operated on at stages 18, 19-, 19- and 22 respectively. A section through the first (NL37 brown) is shown in figure 18. That the de- velopment of the nasal placodes was not dependent on the presence of this nerve tissue is indicated by its absence in other cases, operated on at the same ages, in which placodes alone developed from the transplanted ectoderm. I n other words this series indicates that the capacity for self-differenti- ation is fixed in the nasal ectoderm even before the appearance
TREI JOURNAL OF EXPERIMENTAL ZOOLOGY, VOL. 75, NO. 1

122 ESTHER CARPENTER
of the neural folds. That no specific substrate is necessary for the development of the nasal placode is indicated by the fact that placodes differentiated in the balancer region, be- tween the balancer and the eye, in the mouth region and on the surface of the first gill.
I n no instance in these experiments did a nasal placode fail to develop in the normal position. The earliest case (NL2 brown) and in fact the only one in which it was apparent that all of the nasal ectoderm had been replaced (fig. 7) was operated on at stage 13+ and shows in section a normal nasal placode (fig. 11). This is the same in which a doubtful nasal placode was formed in the balancer region from the trans- planted prospective nasal ectoderm (fig. 12). Replacement of about half the nasal ectoderm in four cases operated on at stage 14 produced little or no effect on the size of the placode developing in situ. Twenty-two other cases operated on from stages 14+ to 26+ in which from one-fourth to one- half of the prospective nasal ectoderm was replaced showed varying amounts of reduction in size of the nasal placode. In general the amount of reduction was less than the amount of material removed. In nine of these cases the telencephalon also showed some reduction in size as compared with that of the side on which no operation had been performed.
DISCUSSION
By the oriterion of self-diff erentiation when transplanted to other parts of the embryo the nasal ectoderm is a segregate, i.e., possesses the specific potency for forming a particular organ or part of an organ (Lillie, '29), earlier than either the balancer or the lens. The equipotentiality of the parts of the segregate is indicated by the fact that a fraction of the material is sufficient to produce a distinct nasal placode on other parts of the head of the embryo if the transplantation is made at stage 14 or later up to stage 26+. I n the earliest case of self-diff erentiation of nasal ectoderm obtained (figs. 8 and 13) about half the prospective nasal ectoderm was transplanted at stage 14 and the embryo examined histo-

THE HEAD PATTERN IN AMBYLSTOMA 123
logically at stage 36-. None of the medullary plate was in- cluded and no nerve tissue developed from the graft. If nasal placodes are induced by the prospective forebrain as sug- gested by Holtfreter there would seem to be a very early transference of the induction potency to the induced material (Raven, '33).
In one case (BN35 brown) less than one-fourth of the prospective nasal material transplanted at stage 19- dif- ferentiated into a nasal placode in the balancer region. The placode of the donor was only slightly smaller than that of the control side. I n this and in other cases in which nerve tissue developed in association with transplanted nasal placodes it seems probable that a small amount of medullary material was included in the graft. It would be desirable to know whether prospective forebrain transplanted alone to some other part of the head region would induce the formation of a nasal placode.
The fact that nasal ectoderm differentiated whether placed in the balancer region, between the balancer and the eye, in the mouth region or on the surface of the first gill indicates that no specific substrate is necessary.
Suppression of the balancer by covering the region with other head ectoderm was obtained as early as stage 13+ (fig. 7) . The only indication of any inductive capacity in the substrate of this region was a slight thickening of the base- ment membrane and in some cases of the lower layer of the covering ectoderm.
The earliest case of self-differentiation of the balancer at a distance from the normal region was obtained by transplanta- tion at stage 19- to the nasal region (figs. 9 and 19). No indication of self-diff erentiation of balancer ectoderm was found at stage 13+ (NL2 brown) or at stage 16 (BN33 blue). There is some evidence (BLN1 brown) that at stage 14 balancer ectoderm will differentiate if transplanted a short distance from the normal region. This case is not strictly comparable with the other experiments, however, since it in- volved turning a graft containing part of the balancer

124 ESTHER CARPENTER
material 90" to the right while in all other operations the normal orientation was maintained.
From the fact that differentiation of balancer ectoderm oc- curred when the graft was taken from the open neural fold stage and placed on other parts of the head region of slightly older embryos (Harrison, '25), it seems probable that further experiments between embryos of the same age will reveal a capacity for self-differentiation in prospective balancer ecto- derm earlier than stage 19-.
Prospective lens ectoderm appears to be in a labile condi- tion during the neural fold stages. In one animal (BLS brown) the whole prospective lens transplanted at stage 14 formed a typical vesicular lens in the stomodaeal region (figs. 8 and 13). This was the only case of that degree of differentiation in operations performed up to stage 25. Lentoids were more often found formed from transplanted lens ectoderm and were variable in size and degree of dif- ferentiation. From the results obtained it is tentatively 'as- sumed that the development of a lens from prospective lens ectoderm transplanted to some other part of the head region before the closure of the neural folds is dependent on the de- gree of determination of the surrounding tissue. That prospective lens tissue may be subordinated by adjacent nasal ectoderm was shown in two cases (BL9 brown and BL21 brown) in which all the lens ectoderm was transplanted to the balancer region at stages 14+ and 20 respectively. I n both grafts a small amount of nasal ectoderm was also in- cluded and from each a nasal placode developed but no lens (figs. 10 and 15).
In no case was the formation of either a lens or a nasal placode in the normal position completely suppressed in these experiments.
CONCLUSIONS
1. As shown by the test of self-differentiation when trans- planted to other parts of the head region of the embryo, nasal ectoderm is determined earlier than either bazancer or lens in Amblystoma punctatum.

THE HEAD PATTERN IN AMBYLSTOMA 125
2. The lens ectoderm is in a labile condition longer than either the balancer or nose.
3. The balancer can be inhibited at an earlier stage than either the lens or nose by covering the region with other head ectoderm.
LITERATURE CITED
CABPENTER, E. 1932 The head pattern in Amblystoma studied by vital staining and transplantation methods. Proc. SOC. Exp. Biol. and Med.,
DETWILEFC, 8. R. On the use of Nile blue sulphate in embryonic tissue transplantation. Anat. Rec., pol. 13, pp. 493-497.
HARRISON, R. G. 1920 Experiments on the lens in Amblystoma. Proc. Soe. Exp. Biol. and Med., vol. 17, pp. 199-200.
- 1925 The development of the balancer in Amblystoma studied by the method of transplantation and in relation to the connective tissue problem. J. Exp. Zool., vol. 41, pp. 249-420. 1933 Some difEcu1ties of the determination problem. Am. Nat.,
vol. 67, pp. 306-321. HOLTFEETER, J. 1933 Organisierungsstufen nach regionaler Kombination von
Entomesoderm mit Ektoderm. Biol. Zentr., Bd. 53, 9. 404-431. - 1933 Der Einfluss von Wirtsalter und verschiedenen Organ- bezirken auf die Diff ereneierung von angelagertem Gastrulaektoderm. Arch. f . Entwmech. d. Organ., Bd. 127, 8. 619-775.
LILLIE, F. R. 1929 Embryonic segregation and its role in the life history. Arch. f. Entwmech. d. Organ., Bd. 118, 8. 499-533.
MANCHOT, E. 1929 Abgrenzung des Augenmaterials nnd anderer Teilbezirke in der Medullarplatte ; die Teilbewegungen wahrend der Auff altung (Farbmarkierungsversuche an Keimen von Urodelen). Arch. f. Entwmech. d. Organ., Bd. 116, S. 689-708.
MANQOLD, 0. 1931 Versuche e m Analyse der Entwicklnng dea Haftfadens bei Urodelen; ein Beispiel fur die Induktion artfremder Organe. Die Naturwiss., 19 Jahrg., 8. 905-911. - 1933 Isolationaversuche zur Analyse der Entwicklung bestimmter Kopforgane. Die Naturwise., 21 Jahrg., 8. 394-397.
RAVEN, C. P. 1933 Zur Entwicklung der Ganglienleiste. 111. Die Induktions- fahigkeit des Kopfganglienleistenmaterials von Rana fneca. Arch. f. Entwmech. d. Organ., Bd. 130, 8. 517-561.
R~HLICH, K. 1931 Gestaltungsbewegungen der prasumptiven Epidermis wahrend der Neurulation und Kogfbildung bei Triton taeniatus. Arch. f. Entwmech. d. Organ., Bd. 124, 8. 66-81.
SCIIZCHTMAN, A. M. 1932 Movement and localization of the presumptive epi- dermis in Triturus torosus (Rathke). Univ. of Cal. Pub. in Zool.,
V O ~ . 30, pp. 370-372. 1917
V O ~ . 36, pp. 325-346.

126 ESTHER CARPENTER
VWT, W. 1925 Oestaltungsanalyse am Amphibienkeim mit Grtlieher Vital- farbung. Vorwort iiber Wege und Ziele. I. Teil. Methodik und Wirkungsweise der ortlichen Vitalfiirbung mit Agar als Farbtrager. Arch. f . Entwmech. d. Organ., Bd. 106, 8. 542-610.
-- 1929 11. Teil. Gastrulation und Yesodermbildung bei Urodelen und Anuren. Arch. f. Entwmech. d. Organ., Bd. 120, 8. 384-706.
WEISSMBEEQ, R. 1929 Vitalmarkierungen mit Bismarekbraun und ihre Kon- servierung zweeke Untersuehung auf Paraffinschnitten. Arch. f. Entwmech. d. Organ., Bd. 118, 8. 485-496.

PLATE
127

PLATE 1
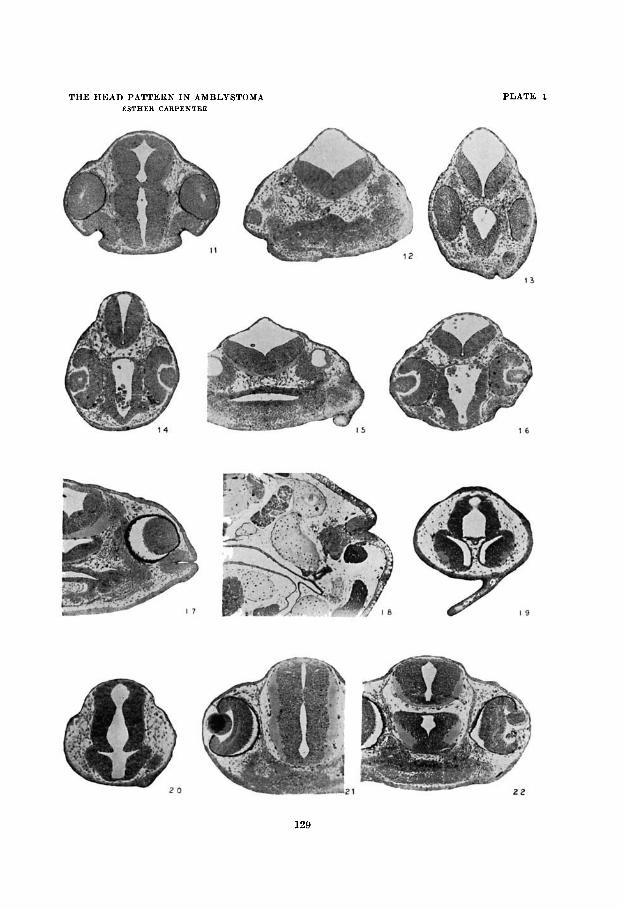
EXPLANATION OF FIGURES
Figures 11 to 22 represent sections of experimental animals fised in Bouin’e fluid and stained with Mallory’s triple stain. The microphotographs were taken with a Bausch and Lomb outfit and a Zeiss microscope with a K7 ocular and 10 apochromatic objective. The microphotographs are reduced one-half. Final magnification is approximately x 35.
11 Section through NL2 brown of figure 7 showing normal nasal plaeode formed from presumptive balancer ectoderm grafted a t stage 13+.
12 Section through NL2 blue showing lentoid attached to ectoderm anil a small nasal placode (fig. 7).
13 Section through BLS brown showing extra nasal placode and vesicular lens formed from ectoderm grafted ventrally at stage 14 (fig. 8).
14 Section through BL8 blue showing lens formed from grafted eetoderm
15 Section through BL9 brown a t stage 39+ showing the nasal plaeode and lierve tissue which developed dorsal t o the balancer from ectoderm transplanted at stage 14+.
16 Section through BL9 blue a t stage 35+ showing the lens formed from presumptive balancer ectoderm transplanted at stage 14+.
17 Section through BN4 blue at stage 42- showing extra nasal placode formed from ectoderm grafted near the eye region at stage 15.
18 Section through NL37 brown, larva 18 mm. long, showing nasal placode and nerve tissue formed from ectoderm grafted at stage 18.
19 Section through BN38 brown showing section through balancer formed anterior to the basal region from ectoderm grafted a t stage 19- (fig, 9).
20 Section through BN31 blue showing partially resorbed balancer formed iu the nasal region from ectoderm transplanted a t stage 20.
21 Section through normal lens of BL21 blue. 22 Section through BL21 blue at stage 40 showing lens formed from pre-
(fig. 8 ) .
sumptive balancer eetoderm transplanted at stage 20 (fig. 10).
128
THE HEAD PATTERN IN AMBLYSTOMA ESTHEB CARPENTER
PLATE: 1


























![Kidney Transplantation (Renal Transplantation) Auto Saved]](https://img.pdfslide.us/doc/110x75/577d22b31a28ab4e1e9807d7/kidney-transplantation-renal-transplantation-auto-saved.jpg)


