Embed Size (px)
Citation preview

The fish: What potential for awareness?
(C-fibres, cognition and consciousness)
ANZCCART conference – Auckland 2008

The information and/or views in this presentation does not necessarily represent the views and/or policy position of the Ministry of Agriculture & Forestry.
Disclaimer:

The question of consciousnessWhat do we mean by consciousness?
Do fish consciously process sensory stimuli?
The biggest debate in the last few years…
…pain sensation
nociception or nociperception?
Using the pain debate to examine some aspects of current cognitive theory and implications for fish.

C-fibre
Aδ-fibre

Reticular formation
Periaqueductal grey
Thalamus
Somatosensory cortex
Prefrontal cortex
Insula
Cingulate gyrus

PAG
Rostral ventrolateral medulla
Locuscoeruleus
NAdr
5-HT
CVS - sympathetic
Vocalisation, motor responses
Nucleus retroambiguus
Raphe
Reticulospinal tract

Human Teleost
nociceptors
Aδ-fibres
C-fibres
spinothalamic tracts
*
PAG, locus coeruleus
*
thalamus *
The nociceptive functional unit
* equivalent structures

• Nociceptors• 22 – 58 receptors on the head of trout• polymodal, mechanothermal, mechanochemical
From Sneddon et al. 2003
Free nerve endings in skin adapted to respond to different stimuli

• Neurochemistry of teleosts• Substance P• Glutamate• Enkephalin• β-endorphin• μ-opioid receptors• α-adrenergic receptors• response to morphine(FSBI 2002, Håstein 2004)
• Functional homology in nociceptive system
• What’s necessary for conscious processing of the nociceptive stimuli i.e. nociperception?
• We have to look at the telencephalon (forebrain)

The cortex and consciousness… Necessity for a large, complex cortex
From Rose (2002)

The compulsory cortex (continued)
• Avoidance learning / “Fear” conditioning = implicit learning• Pavlovian conditioning of HR and RR• migration by olfactory imprinting• implicit learning happens in humans e.g. driving (cars & golf balls)• Macphail (1989) – “learning abilities are a universal capability of vertebrates, even decerebrate vertebrates”
• Overmier and Papini (1986) carried out ablations of fish telencephalons• find and eat food• social behaviours – schooling, spawning, intraspecies aggression• courtship, nest building, parental care• avoidance learning, although possible, was much more difficult (reflex motor response present, modulation of intensity lost)
Consciousness requires a large, complex cortex…Fish have a small cortex…Fish cannot be conscious… … … A valid hypothesis?

The alternative view
The alternative must be that there is a possibility that the fish brain can carry out more than just innate behavioural reflex responses…
…this will take a little more explaining…there’s quite a lot to think about.
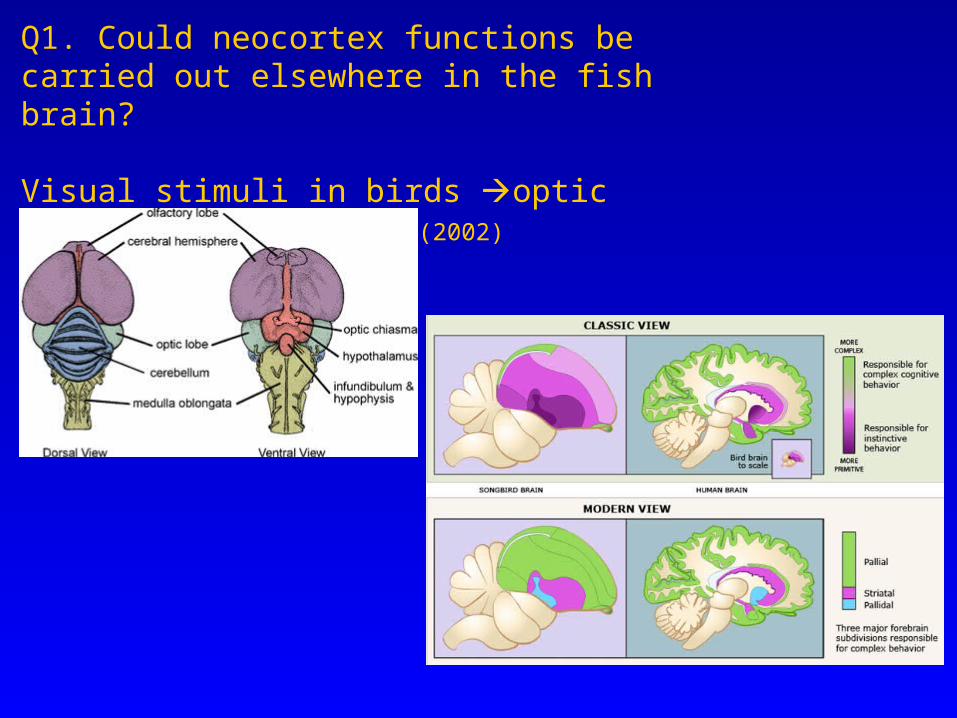
Q1. Could neocortex functions be carried out elsewhere in the fish brain?
Visual stimuli in birds optic tectum (midbrain) FSBI(2002)

Q2. What evidence exists for apparently complex cognitive functions in the fish brain?
Mental representation of environment:goldfish in mazes (Rodriguez et al. 1994)butterfly fish on coral reefs (Reese 1989)rockpool jumping gobies (Aronson 1971)paradise fish avoid areas where they encountered predators (Czanyi & Doka 1993)carp have been shown to avoid hooking (Beukema 1970)
Conspecific recognition in guppies and seahorses (Swaney et al. 2001, Vincent and Sadler 1995)

Defining consciousness
Primary consciousness – “ability to generate mental scene in which diverse information is integrated for the purpose of directing oneself”.
Secondary consciousness – higher order cognition incorporating self knowledge and self awareness.

The generation of consciousness
Not related solely to the cortex…
1. Normal individuals, normal IQ…but…lack normal degree of neocortical expansion (Edelman and Tononi 2000)
2. PVS can occur following thalamic damage, even with almost normal cortical metabolic rates (Schiff et al. 2002)
3. PET showed decreased connectivity (thalamus-cortex) in PVS. This activity was restored in patients who recovered from PVS (Laureys et al. 2000)
Evidence indicates that the generation of consciousness is dependent on thalamocortical neural networks rather than just the existence of the cortex in isolation.

The generation of consciousness
It is not purely an anatomical requirement……there is a functional component too
“Dynamic Core Hypothesis”(Edelman and Tononi 2000, Baars 2002)
Human brain – awake/active vs deep dreamless sleep• Distinct differences in consciousness and self-awareness• Distinct differences in brain wave activity• No physical difference

Resonance networks and γ-frequencies
Consciousness supported by neural resonance in the thalamocortical networks.
Feedback loops / neural signal re-entry
Resonance in gamma frequencies (20-50 Hz)Greatest activity and signal amplification at 40 Hz
Thalamocortical neurones – intrinsic, oscillatory activity in dendritic processes. Occur at 40 Hz when strongly depolarised
awake = depolarised
Slow wave sleep = hyperpolarised
(Llinas et al 1998)

The significance of 40 Hz
10 Hz
40 Hz

Thalamocortical networks in teleost fish
Reports of telencephalon (forebrain) of fish receiving neural projections from thalamic structures
• somatosensory (Ito et al. 1986)• olfactory (Laberge & Hara 2001)• acoustic (Goodson and Bass 2002)
Anatomical requirements are present.Does it prove conscious awareness for e.g. noxious stimuli?

• Dunlop & Laming (2005)
• Can thalamocortical or cortical radiation in response to nociception be demonstrated in fish?
• goldfish & trout
• very fine electrodes – spinal cord, cerebellum, tectum, telencephalon
• noxious stimulus polymodal nociceptor
• Categorised activity by Aδ, C1 or C2 fibres

goldfish
trout
from Dunlop & Laming 2005

Sneddon (2006) reports: -
• fMRI – detected activity in midbrain (expected) and forebrain of fish during noxious stimuli
• gene expression in the forebrain in response to noxious stimuli
• why is this significant?
• substance P upregulate expression of Immediate Early Genes (IEG’s)

Cognitive theory and the fish
Consider 2 theorised mental states: -
1.Conscious cognition
2.Motivational affective states

Conscious cognitionDepends on primary consciousness (Heyes 1993, Dickinson & Balleine 1994)
Ability to retain learned information as mental representations: -
a) “Procedural representations” : stimulus-response/implicit learning
b) “Declarative representations” : selective attention (internal/external stimuli) + expectations Flexible behaviour
e.g. Siamese fighting fish• will chose to observe two other males interacting• are more willing to fight a fish they have observed lose to
another fish• females display a preference for males they have observed
“win” an interaction(Oliveira et al. 1998, Doutrelant & McGregor 2000, McGregor et al. 2001)

e.g. hunger, thirst, fear
Biological adaptation motivate behaviour in situations where response needs to be more flexible than a rigid reflex response
Involved in memory and learning
2 systems theorised: -
Limbic system
Dopaminergic system
Considered relatively primitive systems, conserved through evolution (Le Doux 2000)
Motivational Affective States

Limbic system• humans – amygdala, hippocampus, (hypothalamus, habenulae, etc.)
• amygdala – emotional behaviours & emotional memory (+fear/avoidance)
• hippocampus – spatial learning, relational memory, temporal attributes of stimuli & declarative representation (Eichenbaum 1997)
• ablation of telencephalon of goldfish resulted in similar effects to lesions in the amygdala and hippocampus of humans
• medial pallium amygdala (Portavella et al. 2002,2004)
• lateral pallium hippocampus (Salas et al. 1996, Portavella et al. 2004)
If lesions in the fish telencephalon (forebrain) mirror effects in humans fish have functional homologues for the amygdala and hippocampus (and thus a functional limbic system?)
emotional memory? relational memory? declarative representation?

Dopaminergic systemmesolimbic neurones – associated with limbic system
mediate learning, reward, cost/benefit decisions
• dopamine agonists (apomorphine and amphetamine) can be used to alter state of affect of fish similarly to humans(Lett and Grant 1989, Mok and Munro 1998)
mesostriatal neurones
can induce Parkinson’s Disease like neurochemical and behavioural changes in fish (Pollard et al. 1996)
evidence of a dopaminergic nervous system in fish?
experimental evidence of behaviour that supports a functional limbic and dopaminergic nervous system?

Dunlop et al. (2006) – avoidance learning in goldfish & trout• implanted electrodes – discrete stimulation of local nociceptors• gradient of electrical stimuli; repeated d1 & d2• presence/absence of a conspecific• blood [cortisol]
Is the response flexible? (c.f. declarative representation – limbic system)Do the fish display changing behaviour according to cost/benefit? (i.e. mesolimbic dopaminergic system)i.e. is there evidence to suggest responses change due to cognitive elements
from Dunlop et al. 2006

• Gradient of electrical stimulus / learning = flexible response?
goldfish
trout
from Dunlop et al. 2006

• Presence/absence of conspecific = cost/benefit decision?
goldfish
troutfrom Dunlop et al. 2006
Expt 1 – conspecifc absent
Expt 2 – conspecific present
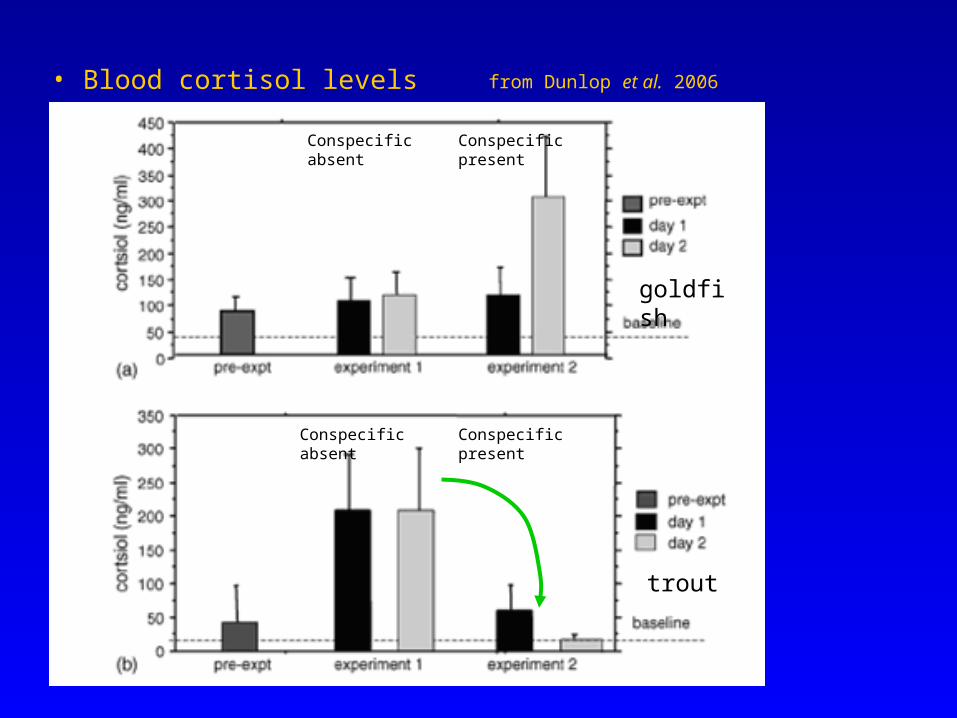
• Blood cortisol levels
goldfish
trout
from Dunlop et al. 2006
Conspecific absent
Conspecific presentConspecific absent
Conspecific present

In summary, teleost fish…• posses the basic unit of nociception (nociceptors, Aδ & C fibres, spinal tracts, neurotransmitters, midbrain involvement)
• respond to μ-opioid agonists (e.g. morphine) which could function either as an analgesic or as an anti-nociceptive
• display complex behaviours associated with learning, memory and spatial relationships
• display thalamocortical connectivity, although we don’t know significance (if any) of resonant frequency
• radiate noxious stimuli beyond the midbrain to the cortex, inducing activity in cortical Aδ- and C- fibres, as well as inducing gene expression
• demonstrate evidence of flexibility of response associated with declarative representation and functional limbic and dopaminergic systems

Thank you
For your attention

The stress response• Primary stress response
• neural mediated release of Adr & NAdr from chromaffin tissue• activation of hypothalmus-pituitary-interrenal axis cortisol
• Secondary stress response• changes to concentrations of pituitary hormones• changes in dopamine and serotonin secretion• elevated HR, elevated Qgill ionic loss/gain• breakdown of CHO and lipid reserves
• Tertiary stress response• changes in immune status & disease resistance• changes in reproductive status

Longer term effects• Upregulation of gene expression
• C-fibres substance P
• stimulation of cell growth in culture
• increased IL-6 and IL-8 production chronic inflammatory effects
• increased TNFα • cellular apoptosis• insulin resistance• hepatic production of acute phase proteins• hyperlipidaemia• chronic long term wt loss, protein catabolism, subendocardial inflammation and hepatosplenomegaly
• increased cFos and cJun expression (IEGs – proto-oncogenes)• fos-jun dimers• increased inducible transcription factor P1• increased DNA transcription, cell division etc.





























