Embed Size (px)
Citation preview

The Ecology and Digestive System of the Struthiolariidae(Gastropoda)
By J. E. MORTON{From the Department of Zoology, Auckland University College)
SUMMARY
The two neozelanic species Struthiolaria papulosa and Pelicaria vermis have beenstudied as regards ecology, feeding mechanism, and structure and function of thedigestive system. They are dwellers on sand or sand-mud-flat, with a feeding positionjust below the surface, where they construct paired siphonal tubes with the rostrum.A ciliary mode of feeding has been acquired by the modification of the gill filamentsand the pallial rejection system. The alimentary canal is adapted for deposit feedingand has developed a crystalline style. Food particles are conducted to the stomach bya functionally reduced mucus-secreting oesophagus, where they are subjected to theaction of the rotating style, and a complex system of ciliary currents. Digestible par-ticles are passed into paired diverticula, where absorption and intracellular digestion,takes place, while faecal material is surrounded with mucus and formed into firmpellets by the ciliary and muscular action of the intestine. The relationships of theStruthiolariidae are discussed, and their origin from the Aporrhaidae is postulated.
CONTENTSPAGE
I N T R O D U C T I O N . . . . . . . . . . . . . I
M A T E R I A L A N D M E T H O D S . . . . . . . . . . . 2
L I F E R E L A T I O N S . . . . . . . . . . . . Z
T h e H a b i t a t 2
E x t e r n a l C h a r a c t e r s a n d M o v e m e n t s . . . . . . . . 3
T h e F e e d i n g P o s i t i o n : F o r m a t i o n o f S i p h o n a l T u b e s . . . . . 4
T h e M e c h a n i s m o f F e e d i n g . . . . . . . . . . 7
T H E D I G E S T I V E S Y S T E M . . . . . . . . . . . 1 1
T h e B u c c a l a n d O e s o p h a g e a l R e g i o n s . . . . . . . . 1 2
T h e S t o m a c h a n d C r y s t a l l i n e S t y l e C a e c u m . . . . . . 1 3
T h e D i g e s t i v e D i v e r t i c u l a . . . . . . . . . . . 1 6
T h e M e c h a n i s m o f D i g e s t i o n . . . . . . . . . - 1 9
T h e I n t e s t i n a l R e g i o n . . . . . . . . . . . 2 0
C O M P A R A T I V E D I S C U S S I O N . . . . . . . . . . . 2 1
R E F E R E N C E S . . . . . . . . . . . . - 2 4
INTRODUCTION
' I "*HE family Struthiolariidae Fischer comprises a small group of proso-X branch molluscs, with an extensive time range and geographical dis-
tribution in Tertiary seas, but including only four recent genera, all in thesouthern hemisphere, of which three have but one surviving species. Perisso-donta Martens 1878 is represented by one species at Kerguelen Land and[Quarterly Journal Microscopical Science, Vol. 92, par t 1, March, 1951.]
2421.1 B

2 Morton—Ecology and Digestive System of.
another at South Georgia, and Tylospira Harris 1897 by a single species inNew South Wales. The present paper deals with the representatives of thetwo New Zealand genera—Siruthiolaria Lamarck 1818, as now restricted,and Pelicaria Finlay 1928. The two species Struthiolaria papulosa (Martyn1784) and Pelicaria vermis (Martyn 1784) have long been known to concho-logy, being accurately figured in Thomas Martyn's The New Conchologistfrom shells collected on Cook's first voyage, probably by Sir Joseph Banks inQueen Charlotte Sound. Our anatomical knowledge of the Struthiolariidaeis, however, extremely fragmentaiy. Quoy and Gaimard (1833) give fairlyaccurate figures of the external characters of the New Zealand species. Hutton(1882) contributed a note on the anatomy of Struthiolaria papulosa; hisdescription and drawings are rough and inaccurate, though the dentition andoperculum were for the first time correctly figured. As regards the biologyof the New Zealand forms, Powell (1937) has briefly concluded that Struthio-laria and Pelicaria are deposit feeders. A detailed examination of the structureand feeding mechanism of Struthiolaria papulosa has brought to light featuresof special biological interest. Pelicaria vermis has been compared withStruthiolaria throughout and found to agree in all essential characters.
MATERIAL AND METHODS
Specimens of both Struthiolaria papulosa and Pelicaria vermis used in thepresent work were collected in New Zealand at Cheltenham and Takapunabeaches on the shore of Rangitoto Channel near Auckland. The mode of lifewas studied in the field, while material was dissected, with the help of aZeiss binocular, both alive for the study of ciliary and digestive action, andafter fixation with Bouin's Fluid. This fixative was found excellent for generalhistological work, while Carnoy's without chloroform was used for the quickfixation of ciliated tissues. Paraffin sections were cut at 8 microns and double-stained with Delafield's haematoxylin and van Gieson's picrofuchsin.
LIFE RELATIONS
The habitat. Struthiolaria papulosa (Text-fig. 2) is found widely throughoutNew Zealand and extends to the Kermadec Islands. It occurs characteristi-cally on the lowermost littoral fringe of clean sand-mud-flats, of whichCheltenham Beach is a typical example, with a wide expanse of shore somethree-quarters of a mile long and 300 yards between tides. Wave action isrelatively subdued and an area of fine shell sand has become covered with athin mantle of organic sediment, plant detritus, and benthic diatoms. Watermovement is nevertheless sufficient to ensure a well-aerated substratum, themore stable conditions of mud-fiat being undeveloped and Zostera presentonly in isolated tufts. There is a comparatively rich fauna, including selectivedeposit feeders such as Struthiolaria, and at least six pelecypods; detrituseaters (Echiuris, Amphiura, the synaptid Trochodota, and a hitherto unrecordedenteropneust, Ptychodera) as well as carnivorous gastropods (Alcithoe andAncilla) and polychaetes (Glycera, Nephthys, and a maldanid).

Struthiolariidae {Gastropoda) 3
Pelicaria vermis (Text-fig. 1) is confined to the North Island—accompany-ing in general the rather more common Struthiolaria papulosa. It is also moretolerant of muddy conditions, being well represented at Waikowhai in theManukau Harbour, where Struthiolaria does not extend. In addition, bothspecies probably occur fairly widely in the soft benthic sub-littoral, being
TEXT-FIGS, I-Z
Fig. 1. Pelicaria vermis, shell x j . Takapuna, Auckland. Fig. 2. Struthiolaria papulosa,shell x | . Cheltenham, Auckland.
recorded by Powell (1937) in the Maoricolpus-\-Dosinula and Taioera-\-Venericardia formations.
External characters and movements. Struthiolaria has the general appearanceof a typical mesogastropod prosobranch; the most conspicuous external fea-ture is the highly extensible foot and head region, for the Struthiolariidae—though sedentary feeders—have, like most sand-flat inhabitants, retainedactive powers of locomotion. Struthiolaria papulosa measures two or threetimes the length of Pelicaria vermis and is further distinguished by its some-what handsome appearance—translucent milk-white in colour, with the ex-posed parts of the head, trunk, and foot marked with fine close-set lines oforange-red. Pelicaria vermis, on the other hand, is yellowish or clay-coloured,with rather less conspicuous lineations of rust-red. The cephalic tentacles inStruthiolaria are tapering and sharply pointed, in Pelicaria less slender andmore bluntly tipped.
The foot is very labile and possesses a broad oval creeping surface attachedto the trunk by a short, cylindrical 'waist' region capable of great elongation.The operculum is reduced to a small chitinous plate with the distal end pro-duced into a strong, sharp claw which is at times employed as an accessorylocomotor organ (Text-fig. 3). The Struthiolariidae exhibit two types of

B
TEXT-FIG. 3. S. papulosa, diagramsshowing the position of the foot, innormal creeping movement (A) and oper-cular movement (B).
4 Morton—Ecology and Digestive System of
movement, the first by using the plantar surface of the foot, the other bythe levering action of the operculum. The normal creeping movement on thesurface of the sand is performed by the widely expanded sole, the oper-
culum remaining out of use on the dor-sal surface of the foot. There is also asecond mode of progression by the useof the sole which is observed to bestadvantage upon a hard surface as atthe bottom of a glass dish. The sole isplaced firmly upon the substratum andthe body-whorl of the shell raised clearof the ground by the elongation of themuscular 'waist' region; the heavy shellis then allowed to fall some distanceforward in the direction of movement,and the sole is again moved forward inadvance of the shell for the repetitionof this rather clumsy lunging movement(cf. Yonge (1937), on Aporrhais).
When the operculum is employed in movement, the extent of the sole isgreatly reduced by withdrawal of blood from the pedal sinus, and the footand trunk elongate to form a narrow muscular column. The sole—now smalland oval—is displaced dorsally, and the muscular lobe carrying the operculumis thrust forward so that the sharp claw projects from the tip of the foot.The opercular claw is used mainly in the characteristic movement of 'righting'the shell when the animal is overturned; it is thrust round forcibly beneaththe left side of the shell and dug deeply into the substratum, when the con-siderable leverage of the foot serves to heave the shell over to its normalposition. The claw of the operculum may also be used as a defensive weaponwhen a specimen is taken up on the hand, the sharp-tipped foot beingextended for 2 to 3 in. and moved about vigorously seeking a point of pur-chase. The opercular movements of Struihiolaria are chiefly of interest inshowing how the normal mode of locomotion in the related highly specializedStrombidae may have arisen. In this group the sole is vestigial and theanimal progresses by a series of convulsive leaps by the flexion of the foot asthe opercular blade is thrust into the substratum.
The feeding position: formation of siphonal tubes. During feeding Struthio-laria lies buried immediately beneath the surface of the sand and constructsa pair of mucus-lined siphonal tubes. It is not in the strictest sense adaptedfor burrowing; despite Oliver's statement (1923) that it ploughs along belowthe surface, it is quite clear that progression does not take place while theanimal is buried, although the feeding position is abandoned at frequentintervals and the animal crawls about freely upon the surface. The noduloseturreted shell is characteristically that of a surface-dwelling gastropod, incontrast with burrowing forms at Cheltenham such as Andlla and Pervicada.

Struthiolariidae (Gastropoda) 5
The animal submerges itself by a continuation of the normal crawlingmovement, and anterior canal being depressed slightly and pushed below thesand, while the labile anterior margin of the foot is thrust downward, obtain-ing purchase in the sand and drawing the shell behind it. Gradually, the
exhalent siphonalapert • —
inhalant siphonalaperture
TEXT-FIGS. 4-7
Fig. 4. S. papulosa buried below the surface, showing the mode of formation of the siphonaltubes. Fig. 5. Surface view of the substratum, showing the inhalant siphonal aperture (right)and the exhalant aperture (left). Figs. 6 and 7. Diagrams showing the action of the pro-boscis in forming siphonal tubes.
pallial current and the entrenching movements of the foot clear a space intowhich the whole shell subsides. The buried animal may usually be locatedby a small raised mound of sand as well as by the two openings of the siphonaltubes (Text-fig. 5), approximately i£ in. apart and about one-sixth of aninch in diameter. These tubes, described originally in the related Aporrhais(Yonge, 1937), are unique among molluscs in being paired and widelyseparated—an inhalant tube directly above the left side of the animaland an exhalant tube on the right. The side-walls are very regular, beingcompacted with a thin secretion of mucus as in burrowing polychaetes andEnteropneusta.

6 Morton—Ecology and Digestive System of
The siphonal tubes of Struthiolaria are constructed by the action of theproboscis (Text-fig. 4), which exhibits a high degree of adaptation for its role.The retracted organ forms a short dorso-ventrally flattened tube, some three-quarters of an inch long, with its integument thrown into very regular, close-set annular rugae. By a copious inflow of blood from the body haemocoeleinto the rhynchocoele, the proboscis may be extended to form a cylindricalsiphon-like organ, almost 3 in. in length, terminated by a circular oral diskbearing the vertical slit-like mouth at its centre. The oral disk is radiallystreaked with yellowish and grey, and may be flat, depressed to form a shallowfunnel, or when fully expanded, somewhat convex. The integument of theproboscis is beset with numerous mucous cells, which are especially denseround the marginal rim of the oral disk. There are also large numbers offusiform sensory receptor cells, of the same type as occur in the integumentof the cephalic tentacles; it appears that the proboscis serves also as a sensoryorgan for maintaining contact with the surface while the animal is buried.
In the formation of the siphonal tubes the proboscis is at short intervalspushed up through the sand to establish a pair of holes. It is first narrowlycompressed by the contraction of the circular muscles passing round therugae, and at the same time greatly elongated and pointed at the tip by partialinflow of blood and the relaxation of its longitudinal muscles (Text-fig. 6).Except for the oral disk, the wall is not everted as in the pleurecbolic introverttype of proboscis. When the oral disk reaches the surface, the circular musclesare relaxed, and the rhynchocoele tensely engorged by the increased blood-supply, the dilation of the rostral artery being assisted by the contraction ofextrinsic muscle slips inserted on its wall. The oral disk is now widely ex-panded with its marginal rim extending slightly beyond the edge of thesiphonal tube (Text-fig. 4). The organ is then quickly withdrawn, and firmlymoulds the wall of the tube in its downward passage, expressing a coat ofmucus, especially from the periphery of the oral disk (Text-fig. 7). From timeto time during feeding the proboscis is extended through one or other tubelike a pelecypod siphon, removing obstructions, and maintaining sensorycontact with the surface. A specimen removed from sand in a laboratory dishwell exhibits the stereotyped tube-forming movements of this organ, withperiodical erection, engorgement, and rapid withdrawal.
The pallial cavity in Struthiolaria (Text-fig. 8) is extremely spacious, anda continuous current of water enters from the inhalant tube and leaves by theexhalant, serving respiratory, cleansing, and food-collecting functions. Ontaking up a specimen in the hand, a strong jet of water is expelled from theexhalant side by the sudden retraction of the head and foot, which servesto close the cavity in front. The aperture is otherwise unprotected and thefree margin of the mantle forms a continuous skirt covering the wide callusedperistome of the shell. It is liberally supplied with blood, and as the gill isprimarily specialized as a current-producing organ, apparently serves anaccessory respiratory function. Just behind the right mantle margin and ashort distance in front of the anus^ lies a single pallial tentacle (Text-fig. 8)
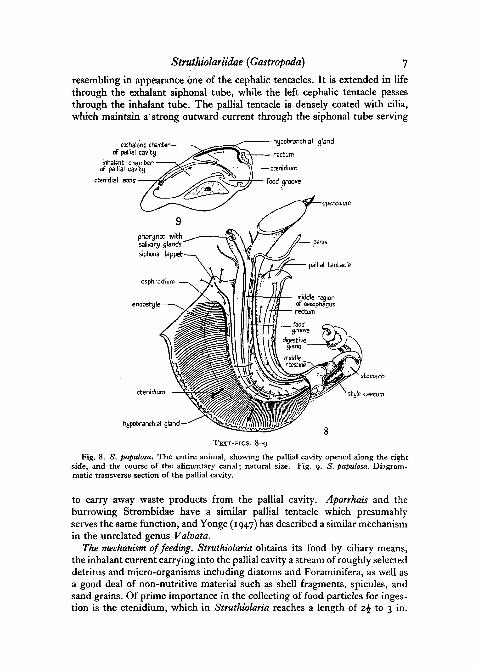
Struthiolariidae {Gastropoda) 7
resembling in appearance one of the cephalic tentacles. It is extended in lifethrough the exhalant siphonal tube, while the left cephalic tentacle passesthrough the inhalant tube. The pallial tentacle is densely coated with cilia,which maintain a' strong outward current through the siphonal tube serving
exhalant chamberof pallial cavity
inha!=nt chamberof pallial cavity
ctenidial axis
pharynx withsalivary glandssiphonal lappet
hypobranchial gland
rectum
ctenidium
food groove
operculum
ctenidium
hypobranchial gland
TEXT-FIGS. 8-9
Fig. 8. S. papulosa. The entire animal, showing the pallial cavity opened along the rightside, and the course of the alimentary canal; natural size. Fig. 9. 5. papulosa. Diagram-. matic transverse section of the pallial cavity.
to carry away waste products from the pallial cavity. Aporrhais and theburrowing Strombidae have a similar pallial tentacle which presumablyserves the same function, and Yonge (1947) has described a similar mechanismin the unrelated genus Valvata.
The mechanism of feeding. Struthiolaria obtains its food by ciliary means,the inhalant current carrying into the pallial cavity a stream of roughly selecteddetritus and micro-organisms including diatoms and Foraminifera, as well asa good deal of non-nutritive material such as shell fragments, spicules, andsand grains. Of prime importance in the collecting of food particles for inges-tion is the ctenidium, which in Struthiolaria reaches a length of z\ to 3 in.

8 Morton—Ecology and Digestive System of
It is immediately apparent on opening the pallial cavity (Text-fig. 8), extend-ing back from the margin of the mantle (across which the anterior filamentsmay protrude) to the narrow posterior end of the cavity, thus completelyencircling the body-whorl. The ctenidial axis (Text-fig. 13) lies along theleft side of the mantle, and the monopectinate lamina arches to the rightacross the whole-width of the mantle cavity. It is composed of some 300-400tubular filaments, each a narrow, laterally compressed rod, attached proxi-mally to the mantle wall on the left side, along the first half of its dorsal edge,while its distal half is entirely free. The great development of the ctenidiumdivides the pallial cavity (Text-fig. 9) obliquely into two longitudinal com-partments each roughly triangular in section. The left compartment or inha-lant chamber is the more venttfally placed, being floored by the dorsal surfaceof the trunk and bounded above by the curved ctenidial septum, extendingfrom the roof of the pallial cavity on the left to near the floor on the right.The right, or exhalant, chamber lies somewhat dorsally to the inhalant, itsfloor being the gill septum while it is roofed by the hypobranchial, or pallial,mucous gland (Text-fig. 9). The pallial epithelium is here extremely thickenedand thrown into wide, yellowish-brown transverse rugae, secreting muchcolourless viscid mucus.
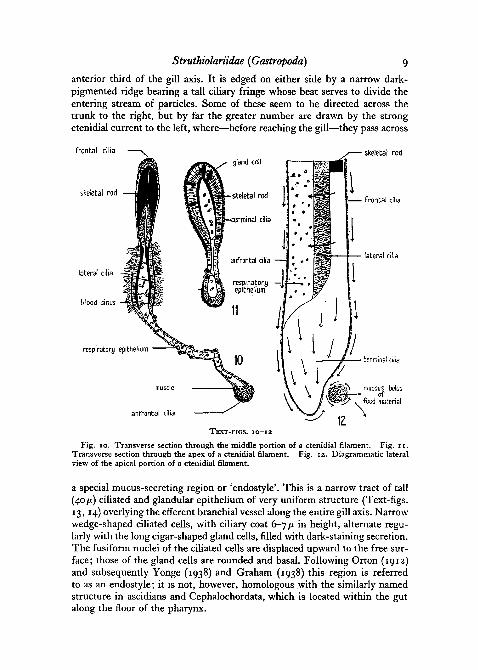
The individual gill filaments are highly adapted to the mode of life. Likethe triangular leaflet of the generalized prosobranch gill (Yonge, 1938), eachis formed of a membranous fold of integument enclosing a narrow respiratoryblood space, and basally attached to the mantle. In Struthiolaria the filamentapex is carried across to the right side of the pallial cavity, to form a long,free distal portion, strengthened by the well-developed, paired, chitinousskeletal rods. In histological structure (Text-figs. 10, 11) the filament is note-worthy for the relatively increased area occupied by the very long lateralcilia (Text-fig. 10). They maintain a strong flexual beat across the filamentfrom ventral to dorsal sides, creating a continuous water current from inha-. lant to exhalant chambers. In addition to the lateral field there are two princi-pal tracts of shorter cilia; the ventral edge of the filament is clad with short,fast-beating frontal cilia, which direct a current towards the apex, whilealong the dorsal edge runs a tract of abfrontal cilia, smaller and with a weakerbeat than the frontals. The original function of the frontal and abfrontal ciliawas no doubt to collect particles for transport to the free edge of the gilland rejection from the pallial cavity (Yonge, 1947). In Struthiolaria, as inother ciliary feeding Prosobranchia (Yonge, 1938), it was only a short stepto establish a feeding mechanism by which nutritive particles were collectedby the frontal and abfrontal currents.
The inhalant current bearing food and detritus is drawn into the pallialcavity on the left across the small triangular siphonal lappet (Text-fig. 8),passing inwards along the osphradium (Text-fig. 13) which may be regardedas an organ for the detection of entering sediment (Hulbert and Yonge,1937). In Struthiolaria, the osphradium forms a simple linear sensory tractwhich runs across the siphonal lappet, and passes backwards parallel to the
Strutkiolariidae {Gastropoda) 9
anterior third of the gill axis. It is edged on either side by a narrow dark-pigmented ridge bearing a tall ciliary fringe whose beat serves to divide theentering stream of particles. Some of these seem to be directed across thetrunk to the right, but by far the greater number are drawn by the strongctenidial current to the left, where—before reaching the gill—they pass across
frontal cilia
skeietal rod
skeletal rod
frontal cilia
ateral cilia
lateral cilia
blood sinus
respiratory epithelium
muscle
anfrontal cilia
TEXT-FIGS, IO-IZ
Fig. 10. Transverse section through the middle portion of a ctenidial filament. Fig. 11.Transverse section through the apex of a ctenidial filament. Fig. 12. Diagrammatic lateralview of the apical portion of a ctenidial filament.
a special mucus-secreting region or 'endostyle'. This is a narrow tract of tall(40 fi) ciliated and glandular epithelium of very uniform structure (Text-figs.13, 14) overlying the efferent branchial vessel along the entire gill axis. Narrowwedge-shaped ciliated cells, with ciliary coat 6-7 /x in height, alternate regu-larly with the long cigar-shaped gland cells, filled with dark-staining secretion.The fusiform nuclei of the ciliated cells are displaced upward to the free sur-face; those of the gland cells are rounded and basal. Following Orton (1912)and subsequently Yonge (1938) and Graham (1938) this region is referredto as an endostyle; it is not, however, homologous with the similarly namedstructure in ascidians and Cephalochordata, which is located within the gutalong the floor of the pharynx.

io Morton—Ecology and Digestive System of
The secretion of the endostyle entangles food and particles of detritus whileat the same time the ciliary field maintains a rapid transverse beat, carryinga continuous sheet of mucus with entrapped particles across the ctenidial axisto the frontal surface of the gill. The frontal cilia at once carry the particlesacross the ventral aspect to the right margin of the gill. The lateral cilia beat-ing inwards between the filaments serve as a sieve mechanism for straining offsolid particles from the respiratory current. Such smaller particles as maypass through the sieve between the filaments receive a small accretion ofmucus from the scattered gland cells in the respiratory epithelium of the fila-ment, and are finally carried to the apex by the abfrontal ciliary tract.
osphradiumciliated cell
gland eel!
TEXT-FIGS. 13-14
Fig. 13. Transverse section through the ctenidial axis, x 50. Fig. 14. Portion of the endostylarepithelium. X 500.
The termination of each filament (Text-fig. 12) is bluntly rounded andslightly expanded; there are no special cilia creating a forward current alongthe gill as in Crepidula and Vermetus (Yonge, 1938), but a uniform coat ofshort terminal cilia beating towards the tip, the columnar ciliated cells beinginterspersed with a few mucus-secreting cells. Just behind the tip of the fila-ment on the ventral (or frontal) side is a small depression where the frontaland abfrontal currents converge, the abfrontal current with its particles havingpassed around the apex. Within the depression the particles of the two
streams are intermingled by a rapid ciliary rotation, and—assisted by a smallamount of mucus—are rounded off into a tiny spherical bolus. The series ofdepressions on successive filaments together constitute a shallow longitudinalfood-collecting groove, and the mucous boluses become continuous to forma thin thread of food material clearly visible to the naked eye along the ventraledge of the gill.
In the living animal the down-curved gill projects into a well-markedexcavation of the dorsal surface of the trunk, occupying the right-hand portionof the pallial cavity floor, and referred to as the food groove (Text-fig. 8). Itcontinues forward in front of the pallial cavity along the right side of thetrunk as far as the base of the right tentacle, becoming considerably narrowerand bounded on either side by tall integumentary folds which may be tempo-rarily approximated to form a closed tube. The bounding fold along the

Struthiolariidae {Gastropoda) 11
right side encloses beneath its outer edge the ciliated genital furrow in bothsexes. The marginal folds of the groove are extremely labile, being liberallysupplied with blood and capable of considerable muscular movement. Theciliated epithelium is richly beset with unicellular mucous glands, v/hosecontents render the integument a characteristic greenish-grey in colour, andstain black in haematoxylin.
From time to time the narrow thread of mucus along the edge of the gill isrolled off by ciliary rotation into the posterior part of the food groove. Otherparticles enter the groove from the surface of the trunk, passing betweensmall muscular crenulations of the left wall. The wide floor of the groove,while less glandular than the margins, maintains a rapid ciliary current whichsweeps particles forward towards the region of the head. At the same time aliberal secretion of mucus is received, while the ciliated coat of the side-wallsbegins to rotate the contents passing forward through the temporary tube,so as to form a long spiral mucous cord, which finally issues from the spout-like opening of the groove near the base of the proboscis. The food string isusually greyish-brown in colour, with a large content of recognizable detritusfrom the substratum. In animals kept in clear water it becomes opaque whiteconsisting of almost pure mucus and resembling a strand of wool. The con-tents of the string are surrounded by a delicate pellicle of mucus which be-comes condensed on contact with the external medium.
At regular intervals the proboscis is turned backwards to the opening ofthe food groove and the paired jaws pluck at the tuft of issuing food material,which is pulled away in strands and either rapidly ingested, or allowed toaccumulate in a small heap at the side of the animal. The radula appears tobe used principally to rake food material through the pharynx, after a bolushas been picked up or detached by the jaws. It would seem likely that themethod of ciliary feeding provides the whole means of subsistence in Struthio-laria. The proboscis is evidently modified wholly for constructing the siphonaltubes; though frequently seen to explore the ground like -a sensory organ, itwas never observed to pick up particles and does not appear adapted forcollecting food.
THE DIGESTIVE SYSTEM
The alimentary canal in Struthiolaria is highly adapted for the slow regularintake of fine detrital particles. As in numerous other microphytophagousmesogastropods a crystalline style is present, and the most distinctive featuresof the digestive system are:
I. The loss of triturating function by the pharynx.
II. The reduction of the oesophagus to a mucus-secreting region convey-ing a food string to the stomach.
III. The reduction of muscular tissue in the gut in general and theincreased reliance on ciliary manipulation of food and faeces.
IV. The specialization of the stomach for the sorting of particles.

12 Morton—Ecology and Digestive System of
V. The absence of extracellular enzymes, apart from that of the style,and the ingestiori of particles by the digestive diverticula.
VI. The adaptation of the intestine for producing firm faecal pellets toavoid fouling of the pallial cavity.
VII. The frequent occurrence of wandering phagocytic cells performingan accessory digestive function.
The course of the alimentary canal (Text-fig. 8) is relatively simple. Themouth opens into a small pharyngeal bulb, leading into a long narrow oeso-phagus, which passes directly through the trunk cavity to open into thestomach on the left side. The stomach is a large rounded chamber, occupying
TEXT-FIG. 15. S. papulosa. A single row of radular teeth.
with the crystalline style caecum the whole left aspect of the first visceralwhorl. It gives exit to paired digestive diverticula which ramify to form thetwo massive, asymmetrical lobes of the digestive gland; the right or anteriorlobe is the smaller and embraces the deep aspect of the stomach, whilst theleft or posterior lobe is spirally coiled, comprising, with the gonad, thegreater part of the visceral hump. The style caecum and the proximal divi-sion of the intestine (Text-fig. 17) open forward together from the stomach,the caecum on the right side overlying and partly concealing the intestine.Just behind the pericardium the intestine turns sharply below the apex ofthe caecum, and emerges on the right side of the visceral spire as the narrowmiddle intestine which loops back around the renal organ and then passesforward along the right pallial wall into the wider rectum. The anus opensanteriorly upon a small spout-like papilla, immediately behind the tentacleon the right pallial margin.
The Buccal and Oesophageal RegionsThe pharynx presents no special features in Struthiolaria, being greatly
reduced in consequence of its loss of function. A pair of cuticular jaws isretained in the form of small triangular plates lining the pharynx wall justwithin the slit-like mouth aperture. This region is readily reversible, and thesharp chitinized margins of the jaws serve to strengthen the edges of themouth for grasping mucous boluses from the food groove. The radula,although equipped with sharp, curved marginal teeth as in other ciliaryfeeding gastropods, was not observed to come into play in seizing food. It isexceptionally small in relation to the animal, and its caecum is very short,scarcely emerging through the floor of the pharynx. There is the typicaltaenioglossan formula (Text-fig. 15) of seven teeth in each row: the laterals

Struthiolariidae (Gastropoda) 13
are rectangular in shape with a finely denticulate cusp at the mesial edge.The quadrangular or five-sided central tooth carries a broad, finely serratetriangular cusp. The salivary glands are vestigial, reduced to a pair of tinywhite lobules closely flattened against the roof of the pharynx at the base ofthe oesophagus. They are histologically simple, the epithelial cells each con-taining a large mucous spherule with no apparent enzymatic contents.
The ciliated, glandular dorsal food channel of the pharynx continues backalong the anterior division of the oesophagus, where it is bounded by a pairof rather prominent dorso-lateral folds. There are- other less permanent epi-thelial folds, and the whole region forms a thin-walled narrow tube, linedwith columnar cells bearing a tall ciliary coat (18/x) interspersed with verynumerous fusiform mucous cells of the same type as those in the margins ofthe food groove. The glandular tracts of the oesophagus are thus a con-spicuous grey-green in colour. The middle region of the oesophagus, com-mencing immediately behind the nerve ring, shows a condition somewhatless advanced than in the ciliary feeders Crepidula and Turritella (Graham,1939): its topographically dorsal portion forms a structural remnant of thespacious oesophageal crop of less specialized prosobranchs, the food channelpassing, as a result of torsion, around the left side to the floor of the oeso-phagus where it proceeds backward as a wide greenish epithelial tract. Themore extensive dorsal portion is transparent and non-ciliated with the liningthrown into small, papillose longitudinal folds. Its epithelium is small-celledand cubical, devoid of glandular elements.
The most posterior part of the oesophagus (Text-fig. 16) possesses—unlikethe rest of the alimentary canal—a fairly thick coat of circular muscle (some-times reaching 100/z). The lining epithelium is uniformly ciliated, throwninto 12-20 regular longitudinal folds. The mucous glands—though lessabundant than anteriorly—are still frequent, and it is in this region that thefood string receives its final shape, being carried back by the cilia along thesummits of the folds to the stomach.
The Stomach and Crystalline Style Caecum
The stomach, style caecum, and proximal intestine form a single functionalunit of the alimentary canal (Text-fig. 17). The stomach is roughly flask-shaped, consisting of a wide posterior chamber receiving the oesophagus, andopening forward into a smaller anterior chamber, which leads in frontby a common aperture to the style caecum and proximal intestine. The cae-cum remains in wide communication with the intestine for about three-quarters of its length, the two cavities being incompletely separated by a pairof typhlosoles—the dorsal one very large, and the ventral a low glandularridge.
The crystalline style projects in life into the anterior chamber, its headbearing like a pestle against the gastric shield, a triangular outgrowth of theventral stomach wall—transparent and brittle in texture and superficially
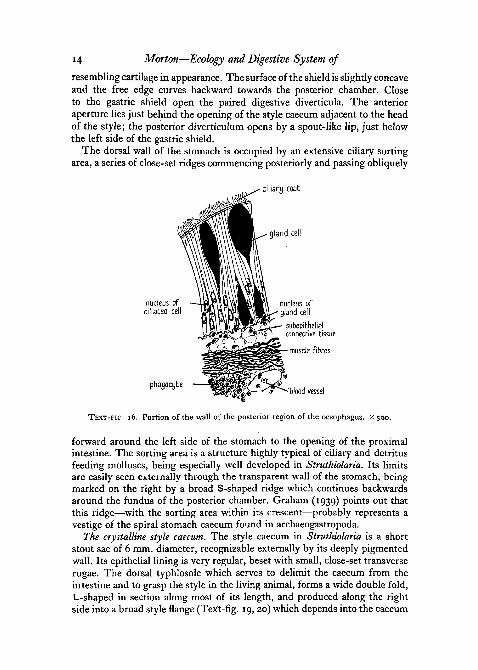
14 Morton—Ecology and Digestive System of
resembling cartilage in appearance. The surface of the shield is slightly concaveand the free edge curves backward towards the posterior chamber. Closeto the gastric shield open the paired digestive diverticula. The anterioraperture lies just behind the opening of the style caecum adjacent to the headof the style; the posterior diverticulum opens by a spout-like lip, just belowthe left side of the gastric shield.
The dorsal wall of the stomach is occupied by an extensive ciliary sortingarea, a series of close-set ridges commencing posteriorly and passing obliquely
ciliary coab
gland cell
phatjocyte•'blood vessel
TEXT-FIC 16. Portion of the wall of the posterior region of the oesophagus. X 500.
forward around the left side of the stomach to the opening of the proximalintestine. The sorting area is a structure highly typical of ciliary and detritusfeeding molluscs, being especially well developed in Struthiolaria. Its limitsare easily seen externally through the transparent wall of the stomach, beingmarked on the right by a broad S-shaped ridge which continues backwardsaround the fundus of the posterior chamber. Graham (1939) points out thatthis ridge—with the sorting area within its crescent—probably represents avestige of the spiral stomach caecum found in archaeogastropoda.
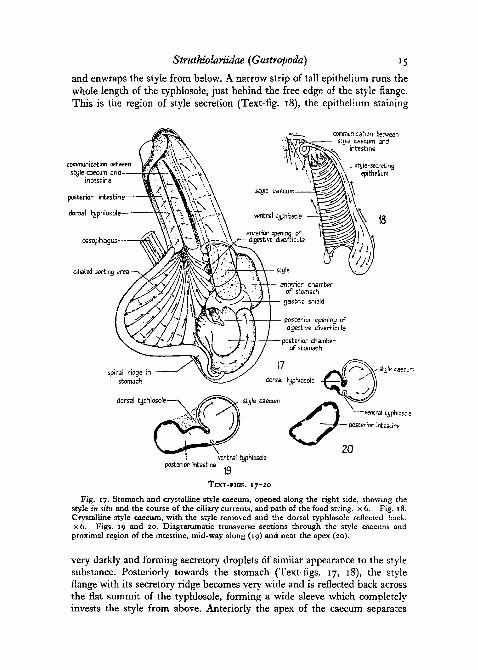
The crystalline style caecum. The style caecum in Struthiolaria is a shortstout sac of 6 mm. diameter, recognizable externally by its deeply pigmentedwall. Its epithelial lining is very regular, beset with small, close-set transverserugae. The dorsal typhlosole which serves to delimit the caecum from theintestine and to grasp the style in the living animal, forms a wide double fold,L-shaped in section along most of its length, and produced along the rightside into a broad style flange (Text-fig. .19, 20) which depends into the caecum
Stnithiolariidae {Gastropoda) 15
and enwraps the style from below. A narrow strip of tall epithelium runs thewhole length of the typhlosole, just behind the free edge of the style flange.This is the region of style secretion (Text-fig. 18), the epithelium staining
communication betweenstyle caecum snd-
incestine
posterior intestine
donal hjphbsole-
oesophagus
ciliated sorting area
communication betweensti/le caecum and
« intestine
styie-secrrtingepithelium
posterior intestine
TEXT-FIGS. 17-20
Fig. 17. Stomach and crystalline style caecum, opened along the right side, showing thestyle in situ and the course of the ciliary currents, and path of the food string, x 6. Fig. 18.Crystalline style caecum, with the style removed and the dorsal typhlosole reflected back.X 6. Figs. 19 and 20. Diagrammatic transverse sections through the style caecum andproximal region of the intestine, mid-way along (19) and near the apex (20).
very darkly and forming secretory droplets of similar appearance to the stylesubstance. Posteriorly towards the stomach (Text-figs. 17, 18), the styleflange with its secretory ridge becomes very wide and is reflected back acrossthe flat summit of the typhlosole, forming a wide sleeve which completelyinvests the style from above. Anteriorly the apex of the caecum separates

16 Morton—Ecology and Digestive System of
completely from the intestine (Text-fig. 20) by the coalescence of the lefttyphlosole margin with the ventral wall of the common caecum-intestinalchamber. The now narrow style flange remains in contact with the stylewithin the caecum. The line of fusion lies to the left of the small ventraltyphlosole which is thus also enclosed within the caecum, together with anarrow remnant closed off from the intestinal lining.
The crystalline style forms an extremely delicate taper.ed rod, 2 cm. inlength, hyaline golden brown in colour, and very flexible. It is rapidly dis-solved after cessation of feeding, and was best examined immediately uponremoving the animal from water, being completely resorbed within an houror two of collecting.
Histology. The lining epithelium of the stomach, in contrast to that of theoesophagus and of the intestine, is devoid of gland cells. There are two mainhistological regions, the ciliated sorting area, and the cuticulated area sur-rounding the base of the gastric shield. The sorting ridges are due entirelyto differences in the height of the cells, varying from 80 //. along the folds to30^. in the grooves. The nuclei are elongate-ovoid, binucleolate, forming asubcentral row, and the ciliary fringe is especially well developed (12-15 fi),supported by dense clusters of fibrillae. The epithelium is conspicuouslyinvaded by wandering phagocytes from the underlying zone of connectivetissue and a basal supporting reticulum traverses a series of intra-epithelialcanals. The cuticulate epithelial cells are extremely tall and narrow, reaching200 //, in height, while the cells secreting the gastric shield proper become astall as 750 JU.. The nuclei are compressed and rod-like, forming a crowdedsubcentral row, while the secreted cuticle (10 fi thick) is hyaline and structure-less, attached to the epithelium by fine perpendicular strands resembling cilia.
The epithelium of the style caecum (Text-fig. 21) possesses extremelyrobust cilia forming a dense coat 25 p in height. The ciliary basal granulesform a prominent refractile line, and the cell fibrillae form dense bundlespassing from the sides of the nuclei to the bases of the cells. The ovoid tospherical nuclei are izfj- in length, binucleolate and with sparse chromatin.The cytoplasm is uniformly granular and without secretory contents. At thebases of the cells is developed a system of intracellular supporting fibres,intra-epithelial canals appearing in section as small, non-staining vacuoles.
The Digestive DiverticulaThe digestive gland in Struthiolaria shows typical alveolar structure, each
diverticulum ramifying into a system of smaller ductules, which ultimatelygive rise to clusters of small, dark-coloured digestive lobules, each about200^ across. The ductules are lined throughout with columnar ciliated epi-thelium, the cells reaching 55/z in height, with a ciliary coat 5-6 [j. wide andfibrillar bundles well developed. The basal nuclei are ovoid, binucleolate, and7 jii in length.
The glandular cells of the terminal lobules (Text-fig. 22) are of two distincttypes, digestive and excretory. The lumen is generally triangular, bounded on

Struthiolariidae {Gastropoda) 17
three sides by tall columnar digestive cells which by increase in height mayreduce the space to a triradiate cleft. At the bottom of each crypt a smallerpyramidal group of darkly pigmented excretory cells is intercalated betweenthe digestive cells. The digestive cells possess small, darkly staining basalnuclei. The free surfaces—though always intact—are commonly rather con-vex, sometimes bulging and somewhat pseudopodial. There is some evidence
ciliary coat
phagocyteintra-epithelial
canalmuscle fibres
pigmented cell
TEXT-FIG, SI . Portion of the wall of the style caecum, showing ciliated cells. X 500.
that the border is normally ciliated in the resting condition, though cilia arenot apparent after fixation. A row of tiny refractile granules underlies thefree surface, with a short fan of fibrillae radiating through the superficialcytoplasm. The distal cytoplasm of the digestive cell is very coarsely granularand evidently contains ingested material, while the basal half of the cell ispacked with small greenish spherules, 2-3 p across, somewhat oily in appear-ance and containing irregular clumps of particulate matter. The digestiveepithelium is frequently invaded by phagocytes from the vascular interstitialconnective tissue.
The pigmented excretory cells in the crypts appear to be of several types.There is usually a centrally placed, broad-based, flask-shaped cell, with aconstricted neck, containing a single large dark pigment spherule, sometimes25-30/n. across, round and dark-brown or black with a greenish oily irides-cence. The nucleus is small and pressed flat against the cell base. Flankingthe central cell on each side are several columnar excretory cells, of thesecond type, with small round nuclei as in the digestive cells. The free sur-face is rounded and pseudopodial, and the distal third packed with rather
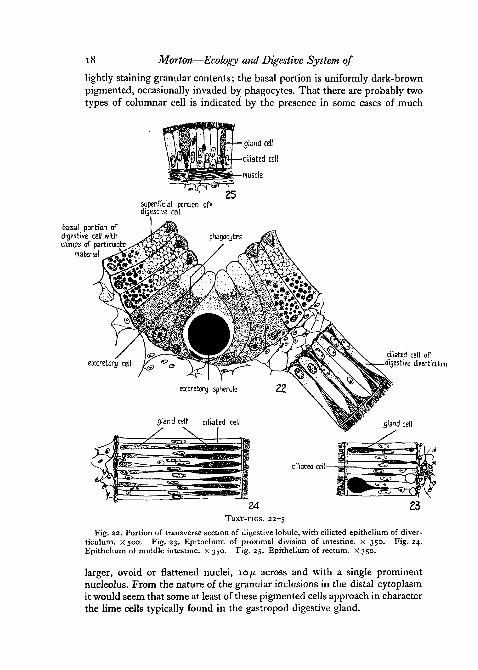
18 Morton—Ecology and Digestive System of
lightly staining granular contents; the basal portion is uniformly dark-brownpigmented, occasionally invaded by phagocytes. That there are probably twotypes of columnar cell is indicated by the presence in some cases of much
gland cell
ciliated cell
muscle
superficial portion of*digestive cell
basal portion ofdigestive cell withclumps of participate
material
excretory cell
excretory spherule
gland celt ciliated cell
ciliated cell ofdigestive diverticulum
gland cell
ciliated cell—-S;:
24TEXT-FIGS. 22-5
Fig. 22. Portion of transverse section of digestive lobule, with ciliated epithelium of diver-ticulum. X500. Fig. 23. Epithelium of proximal division of intestine. X 350. Fig. 24.Epithelium of middle intestine, x 350. Fig. 25. Epithelium of rectum. X 350.
larger, ovoid or flattened nuclei, 10 ft across and with a single prominentnucleolus. From the nature of the granular inclusions in the distal cytoplasmit would seem that some at least of these pigmented cells approach in characterthe lime cells typically found in the gastropod digestive gland.

Struthiolariidae {Gastropoda) 19
The Mechanism of DigestionIn Text-figs. 17 and 18 are illustrated the system of ciliary currents in the
stomach and style caecum which together comprise the most specializedregion of the gut. In this region the wall of the gut is firmly attached bymuscle strands to the external body-wall, and is incapable of peristalsis; theciliary fields are thus all-important in securing movement of contents. Thelong robust cilia of the style caecum serve by their transverse beat to rotatethe style on the bearings formed by the epithelial folds. The direction ofrotation is clockwise, and in the transparent veliger larva the speed wasobserved to be about 40 turns a minute. At the same- time the dense butshorter cilia covering the style flange of the typhlosole maintain a strong beattowards the stomach, by which the rotating style is gradually thrust down-ward so that its head bears against the gastric shield. Yonge (1932) hasemphasized the importance of the style as a mechanism for the continuousliberation of amylase in those molluscs, lamellibranchs, and micro-herbivorousprosobranchs, which have a slow continuous feeding process. Strong amylo-lytic action was detected by digestive experiments on the style of Struthiolaria,while no action was observed on either dissaccharides or cellulose. It wouldseem, however, that a possibly equally important function of the style is toafford mechanical assistance to the movement of the food contents in thestomach. The soft, rotating style head becomes securely attached to the endof the oesophagal food string, which is slowly but continuously drawn intothe stomach, around the fundus of the posterior chamber and forward onthe right side to the surface of the gastric shield. The cord is thrown into aclose spiral by the style rotation, while the attached end is broken down andthe contents freed, partly by attrition by the style head, but also, no doubt,by the lowered viscosity of the mucus in the stomach, brought about by thereduction of hydrogen ion concentration (Yonge, 1935).
The rotation of the style also effects a constant stirring of the dispersedstomach contents by which particles are repeatedly drawn across the ridgesof the ciliary sorting area. In Struthiolaria a large amount of non-nutritivematerial must enter the stomach. The sorting area consists of flat-toppedprimary ridges alternating with smaller triangular secondary ridges runningalong the intervening grooves. The primary ridges bear tall ciliary fringeskeeping up a steady transverse beat across the stomach wall at right-angles tothe direction of the ridges. Coarser, heavier particles such as sand grains,thrown against the sorting area by style rotation, sink into the grooves, andare carried directly forward by the cilia of the secondary ridges to the proxi-mal intestine. The lighter particles, on the other hand, are flicked across thesorting area by the ciliary tufts of the primary ridges, and are conveyed byspecial ciliary currents into the digestive diverticula.
The diverticula do not secrete; apart from the action of the style amylase,digestion is intracellular. The digestive gland forms the main absorptiveregion of the gut, and a series of simple spotting tests on extracts showed anormal complement of intracellular enzymes. Of proteins, fibrin and also

20 Morton—Ecology and Digestive System of
powdered peptone were readily digested, best in slightly acid media; 5 percent, methyl acetate was easily hydrolysed showing the presence of anesterase (probably a lipase), while a strong amylase was demonstrated bycomplete hydrolysis of starch solution. There was no action on the celluloseof cotton fibres and it is probable that, as in other detritus feeders, cellulose,if assimilated at all, demands previous bacterial or autolytic breakdown.
Fine particles appear to be ingested by the epithelium of the diges-tive lobules in its pseudopodial phase. The basal greenish spherules of thedigestive cells invariably contain clumps of solid particles, which apparentlyrepresent the non-assimilable residue after intracellular digestion, probablysurrounded by enzyme to form a fluid vacuole. At regular intervals these basalcell contents are returned to the stomach by the fragmentation of the digestivecells, and can be detected in sections in the form of tiny boluses of egesta,approximately 50 fi across, passing forward along the sorting area to theintestine. Each bolus contains closely compacted cell fragments containingthe greenish vacuoles, as well as nuclei, phagocytic cells, and brown excretoryspherules. Mansour (1946) claims that in the lamellibranchs enzymes areliberated into the stomach by the fragmentation of holocrine digestive cells.In Struthiolaria, however, it is quite clear that the fragmented particles arealways in the nature of egesta, and pass directly to the intestine. Small tracesof enzyme are doubtless liberated in this way into the stomach; in general,however, Struthiolaria conforms entirely to Yonge's rule as to the intra-cellular nature of digestion in style-bearers.
Following Macmunn (1900) it may be concluded that the large dark spherulesdischarged from the excretory cells consist of a chlorophyllous pigment de-rived from food substances and extracted from the blood by special cells in thedigestive lobules. Intracellular digestion of detrital particles must be accom-panied by large absorption of plant pigments, and in style-bearing gastropodsin particular the excretory mechanism of the digestive gland is well developed.
The abundance of wandering phagocytic cells in the subepithelial layer ofmost parts of the gut has already been mentioned. These cells readily invadethe epithelium and ingest solid particles, though it is questionable whetherthey are primarily nutritive or are concerned with the removal of waste par-ticles. It is certain that particles of no food value may be ingested, as wasdemonstrated by the accumulation of neutral red particles within the phago-cytic cells of the sorting area of the veliger larva. The most probable hypo-thesis, in light of the work of Yonge (1926), is that the phagocytes emergeinto the lumen where they ingest both free food particles, which are thenintracellularly digested, as well as particles of rejected material. The presenceof a very thick zone of phagocytes in the wall of the proximal intestine ofStruthiolaria would point also to rejectory function.
The Intestinal RegionThe intestine, adapted solely for compaction of faeces, consists of three
regions, the wide proximal intestine communicating with the style caecum,

Struthiolariidae {Gastropoda) 21
a harrow middle intestine, and a terminal rectum. The lining is ciliated andglandular throughout (Text-figs. 23, 24, 25), and in the proximal intestine theepithelium is disposed in tall longitudinal tracts, separated by narrowerintervening grooves lined by much shorter cells. The ventral wall has twobroad tracts, separated by a deeply incised groove along which are carriedindigestible particles from the grooves of the gastric sorting area. The dorsalwall consists also of two broad tracts with a narrower suture, while along theleft side runs a wide shallow channel of low-celled epithelium. The wall ofthe proximal intestine is not, as in the stomach, secured to the body-wall,and the edges of the broad tracts work freely upon the contents of the grooves,by means of a thin, continuous zone of muscle in the hind-gut wall. Thefaecal material is all the while liberally admixed with mucus, and particlesare carried by ciliary currents across the tracts into the grooyes, where theyare conveyed forward to the middle intestine.
The faeces are formed into pellets during passage through the middleintestine, which has a characteristic trihedral structure, the lumen beingbounded by three broad ciliated tracts sutured by three grooves of low epi-thelial cells. The ciliated cells of the broad tracts are extremely tall and nar-row (120/x in height as compared with 80ju. in the proximal intestine). Theypossess ovoid-elongate central nuclei, and are regularly interspersed with aseries of greatly attenuated mucous cells, with a distal rank of cigar-shapedsecretion droplets, and a basal row of rounded uninucleolate nuclei (Text-fig.24). In passing along the narrow grooves the faeces are firmly kneaded intoa coherent mucous string, from which individual pellets are from time to timenipped off by slight peristaltic movements of the bounding tracts. The pelletsare given their final compact form by ciliary rotation against the rectum wall,producing firm grey-green ovoid masses, approximately 0-25 mm. long.
The rectum is a long straight tube with its walls smooth or thrown whenempty into small impermanent folds. The columnar ciliated cells (Text-fig.25) are much shorter (40-50/x) than in preceding regions, with ovoid basalnuclei. The gland cells are simple and ovoid, with light-staining contentswhich form the transparent coat finally surrounding the large masses of faecalpellets. The rectal wall near the anus is somewhat muscular, and the faecesare expelled by slight contractions in coherent strings which are immediatelycarried away in the exhalant pallial current.
COMPARATIVE DISCUSSION
Ciliary feeding in prosobranchiate gastropods has now been reported as theresult of independent evolutionary change in five style-bearing families whichotherwise have little in common, namely, the sessile Calyptraeidae (Orton,1912), Vermetidae (Yonge, 1932), Capulidae (Yonge, 1938), and in the free-moving Turitellidae (Graham, 1938). To these the also active Struthiola-riidae must now be added. Similar pallial adaptations have been acquired ineach family, including strongly ciliated, linear gill filaments, and mucus-producing endostyle and food groove. The Calyptraeidae [Calyptraea and

22 Morton—Ecology and Digestive System of
Crepidula) are the most highly specialized group, being suspension feeders,largely on diatoms, and having the gill filaments free and rod-like along theirwhole length, with the blood space reduced, and the membranous respiratoryarea entirely lost. Yonge correlates the extreme specialization of the gill fila-ment with a reduction in respiratory needs of the sessile Crepidula; in theactively moving Struthiolaria the filament has, in the proximal region atleast, retained its respiratory function.
The Struthiolariidae most closely resemble in their biology the familyTurritellidae (Graham, 1938; Yonge, 1946), as typified in New Zealand byMaoricolpus rosea, examined during the present work. These two groups arethe only recorded ciliary feeding prosobranchs free-moving in a soft sub-stratum. In neither case has the gill filament become free along its wholelength, while in both groups there are convergent adaptations for dealingwith the large amounts of non-nutritive bottom material that must enter thepallial cavity during feeding. The unicellular mucous glands of the gill fila-ment, and the large hypobranchial gland are retained, while the food groove—a mere shallow depression in Crepidula—forms in Struthiolaria and Turri-tella a muscular, temporarily closed tube, capable of compacting largeamounts of imperfectly sorted surface detritus. In both forms the gastricciliary sorting area, and the faeces-compacting region of the proximal intes-tine, only slightly. represented in Crepidula, are prominently developed.Turritella, with its long tapering shell, is pre-eminently adapted for trailingthrough mud, and the pallial cavity is screened from sediment by a curtainof pinnate pallial tentacles. The absence of such a mechanism in Struthiolariamay probably be ascribed to the cleaner, sandy habitat, and to the efficientmucus-lined siphonal tubes.
The Struthiolariidae are classified by Thiele (1931) in his Stirps Strom-bacea, along with the families Aporrhaidae, Strombidae, and Xenophoridae.Unpublished observations of the present writer indicate that the Xenophori-dae are somewhat removed from the remaining three families, which togetherform a compact natural assemblage—with an especially close affinity betweenthe Aporrhaidae and Struthiolariidae. In general characters Aporrhais pes-pelicani, as described by Yonge (1937), is very similar to Struthiolaria. Thehabitat is alike in both groups, and in mode of locomotion Aporrhais agreeswith Struthiolaria in showing both creeping and lunging movements with thewidely expanded sole. Yonge states that the reduced operculum in Aporrhaisis never used in locomotion, and that the righting movement of the upturnedanimal is effected not with the operculum after the strombid fashion, but byleverage obtained by placing the sole flat on the substratum. However,examination of the operculum in preserved Aporrhais pes-pelicani revealsthat the distal edge is produced into a strong, spade-like plate, not figured byYonge, which has all the appearance of a locomotor organ like the claw inStruthiolaria.
Aporrhais, like Struthiolaria, constructs paired, mucus-lined siphonal tubesby the action of the proboscis. This adaptation, which is recorded nowhere

Struthiolariidae {Gastropoda) 23
else among gastropods, would appear to have arisen as a single evolutionarydevelopment in a common ancestral form of the Aporrhaidae and Struthio-lariidae. Aporrhais has not acquired a ciliary feeding mechanism; the pallialorgans, while forming an efficient cleansing and rejection system, are notmodified for food collecting, and the gill filaments remain triangular. Theinhalant current brings detrital particles to the vicinity of the rostrum andaccording to Yonge feeding takes place as readily upon the surface as whenburied. 'The animal presumably feeds normally by collecting by means ofthe extensible proboscis all particles of plant matter which occur in the mudin the region below and around the expanded lip.' Yonge stresses the creationof powerful water currents by the gill in Aporrhais and the elaboration ofciliary currents for the rejection of sediment. He emphasizes that thesedevelopments 'have an added interest in that they indicate the way in whichciliary feeding in gastropods may have been evolved'. The exhalant currentsmight further be modified to form the food channel, while modifications ofthe tips of the gill filaments would convert the gill into the food-collectingorgan found in Crepidula. Such a process is exactly what is now found tohave occurred in the Struthiolariidae, which apparently represent an almostdirect continuation of the aporrhaid trend of evolution. It is of considerableinterest, since Struthiolaria has not hitherto been investigated in life, to findYonge's hypothetical conclusion actually realized in the closest living relativesof the Aporrhaidae.
In the digestive system Struthiolaria shows in comparison with Aporrhaisa number of structural advances associated with ciliary feeding. The alimen-tary canal of Aporrhais (Yonge, 1937) examined by the present writer fromfixed material has an essentially similar plan to that of Struthiolaria] theforegut, however, is a good deal more generalized, and the condition of thedigestive and other systems indicates that the Aporrhaidae may have givenrise fairly directly both to the Struthiolariidae and to the somewhat lessclosely related Strombidae. The buccal bulb is much less reduced than inStruthiolaria, and the radula—while already small—remains large enough for thelong marginal teeth to be employed with the jaws in seizing particles. The anterioroesophagus is much wider than in Struthiolaria, and the mid-oesophagusretains as in the Strombidae a long cylindrical crop traversed by the ciliatedglandular food channel, ventral in position following torsion. The salivaryglands remain as large lobulated masses within the trunk cavity, sending for-ward through the nerve ring a pair of long ducts which run along the oeso-phagal roof to the pharynx. The stomach and style sac complex of Aporrhaisis closely similar to that of Struthiolaria, the figure of Yonge (1937) requiringcorrection by the transposition of the captions of the oesophagus and anteriordigestive diverticulum.
Struthiolaria possesses further specializations upon the aporrhaid plan,especially in the nervous system, and the reproductive organs and life-history.The Aporrhaidae are a somewhat ancient family with a world-wide Jurassicrange, and are represented by several genera in the New Zealand Cretaceous

24 Morton—Ecology and Digestive System of
(Struthioptera, Drepanochilus, Hemichenopus, Dicroloma, Perissoptera). Thefirst undoubted struthiolariids belong to the earliest South American Tertiary1
(Marwick, 1924).The present writer has examined the shell and animal of Perissodonta
georgiana, of South Georgia, one of the two species of the most archaic livinggenus. On both shell and dentition features this group appears to stand closeto the line of descent from an aporrhaid stock. In all essential features, how-ever, the animal is a true struthiolariid, and the pallial organs are already fullyadapted for ciliary feeding.
ACKNOWLEDGEMENTS
The writer wishes to thank Prof. W. R. McGregor, Head of the ZoologyDepartment, Auckland University College, for assistance during the prepara-tion of the thesis of which these results formed part, Mr. A. W. B. Powell,Assistant Director, Auckland Museum, for much helpful discussion andfrequent loan of specimens, and the Director, Marine Biological Associationof the United Kingdom, Plymouth, for generously supplying preservedmaterial of Aporrhais. Finally, the writer has had the special advantage ofProfessor C. M. Yonge's kindly criticism of the manuscript.
REFERENCES
GRAHAM, A., 1938. 'On a ciliary process of food-collecting in the gastropod Twritella com-munis Risso.' Proc. Zool. Soc. Lond. A, 108, 453.I939- 'On the structure of the alimentary canal in the style-bearing prosobranchs.'
Ibid. B, 109, 75.HULBERT, G. C. E. B., and YONGE, C. M., 1937. 'A possible function of the osphradium in
the Gastropoda." Nature, 139, 840.HUTTON, F. W., 1882. 'Notes on the structure of Struthiolaria papulosa'. Trans. N.Z. Inst.,
15, 117.MACMUNN, C. A., 1900. 'On the gastric gland of Mollusca and decapod Crustacea.' Phil.
Trans. Roy. Soc. B, 193.MANSOUR, K., 1946. 'Food and digestive organs of lamellibranchs.' Nature, 158, 378.MAECTN, T., 1784. The Universal Conchologist. London.MARWICK, J., 1924. 'The Struthiolariidae.' Trans. N.Z. Inst., 55, 161.OLIVER, W. R. B., 1923. 'Marine littoral plant and animal communities in New Zealand.'
Ibid., 54, 496.ORTON, J. H., 1912. 'The mode of feeding of Crepidula with an account of the current-
producing mechanism in the mantle cavity, and some remarks on the mode of feeding ingastropods and lamellibranchs.' J. mar. biol. Assoc. U.K., 9, 444.
POWELL, A. W. B., 1937. 'Animal communities of the sea bottom in the Auckland and Manu-kau Harbours.' Trans. Roy. Soc. N.Z., 66, 354.
QUOY and GAIMARD, 1833. Voyage autour du tnonde de I'Astrolabe, 1826-29. Zoologie. Illustra-tions, vol. i.
THIELE, J., 1931. Handbuch der systematischen Weichtierkunde, I. Jena, Fischer.YONGE, C. M., 1923. 'Studies on the comparative physiology of digestion. I. The mechanism
of feeding, digestion and assimilation in the lamellibranch Mya.' Brit. Journ. exp. Biol.,1,15.1926. 'The digestive diverticula in the lamellibranchs.' Trans. Roy. Soc. Edin., 54,
7O3-1932. 'Notes on feeding and digestion in Pterocera and Vermetus, with a discussion on
the occurrence of the crystalline style in the Gastropoda.' Sci. Repts. Gt. Barrier ReefExped. 1928-1929, Brit. Mus. (Nat. Hist.), 1, 259.

Struthiolariidae {Gastropoda) 25
YONGE, C. M., 1935. 'On some aspects of feeding and digestion in ciliary feeding animals.' J.mar. biol. Assoc. U.K., 20, 341.1937- 'The biology ot Aporrhais pes-pelicani, Linn, and A. serresiana (Mich.).' Ibid.,
21,687.1938. 'Evolution of ciliary feeding in the Prosobranchia with an account of feeding in
Capulus ungaricus.' Ibid., 22, 453.1946. 'On the habits of Turritella communis Risso.' Ibid., 26, 377.1947- 'The pallial organs in the aspidobranch Gastropoda and their evolution through-
out the Mollusca.' Phil. Trans. Roy. Soc. B, 232, 443.





























