Embed Size (px)
Citation preview

The dynamics of bursting in neurons and networks
Professor Jonathan Rubin January 20, 2011

Outline:
• Introduction to neurons
• Introduction to bursting
• Mathematical analysis of bursting in single cells
• Frontiers in research on bursting dynamics (21-53)
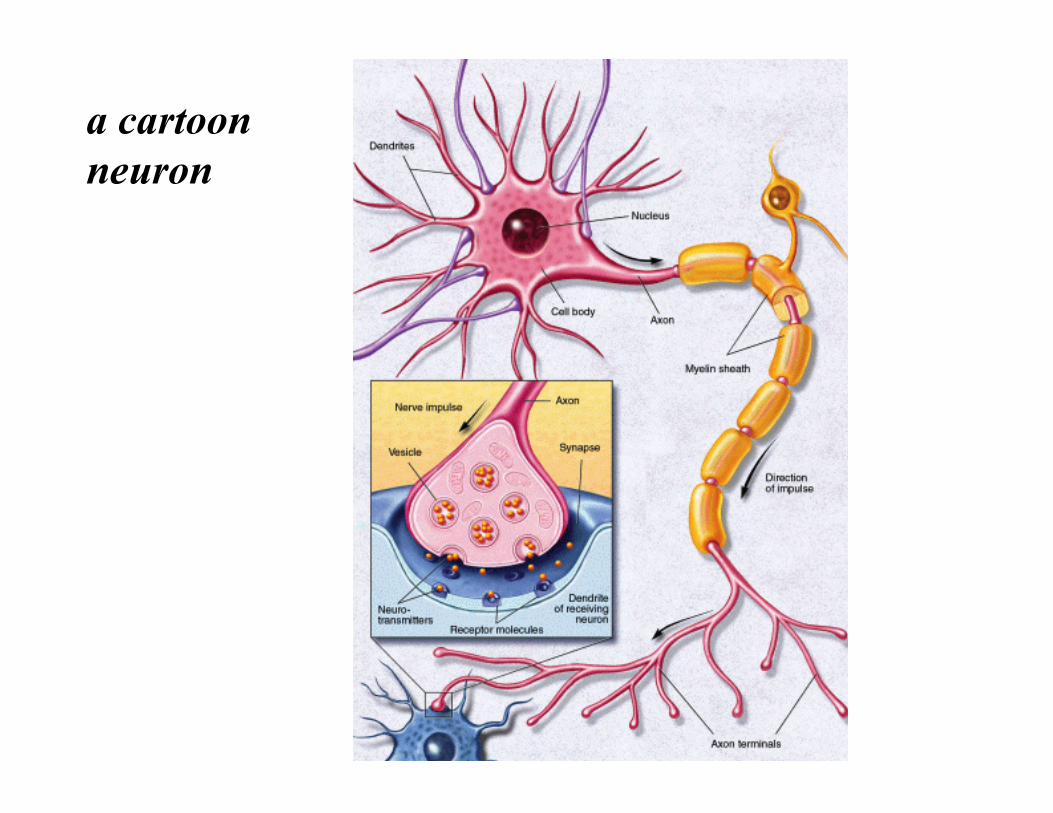
a cartoon neuron
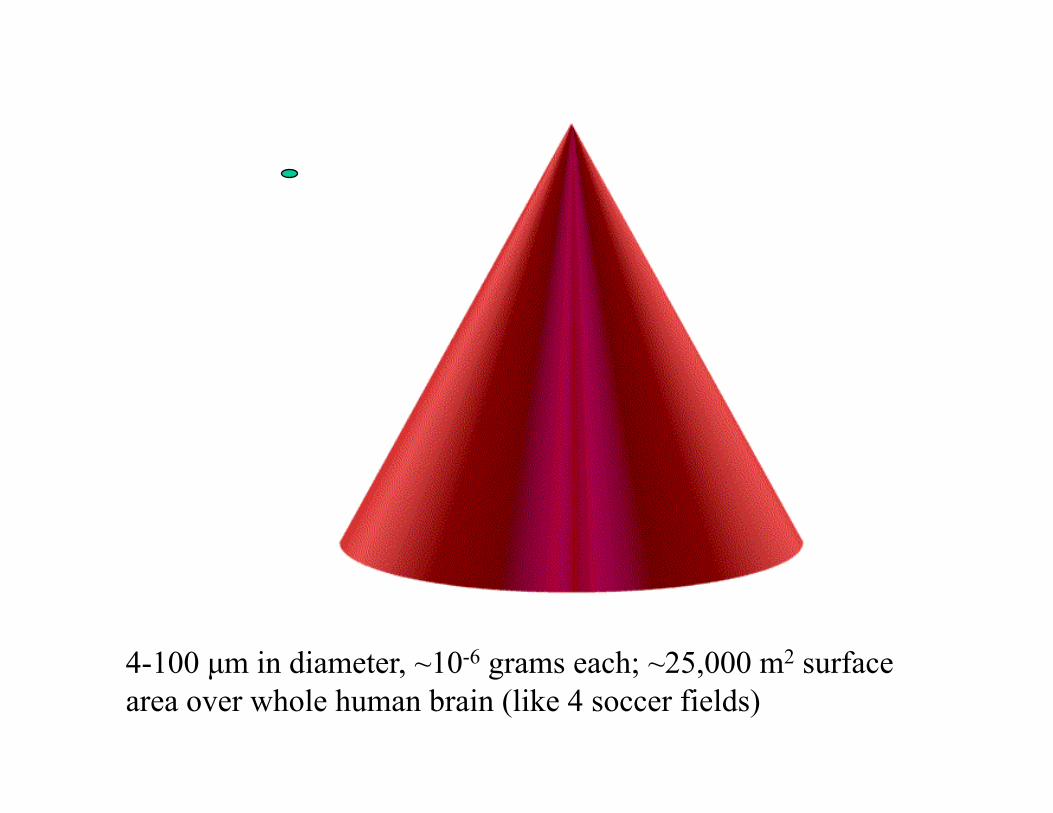
4-100 µm in diameter, ~10-6 grams each; ~25,000 m2 surface area over whole human brain (like 4 soccer fields)
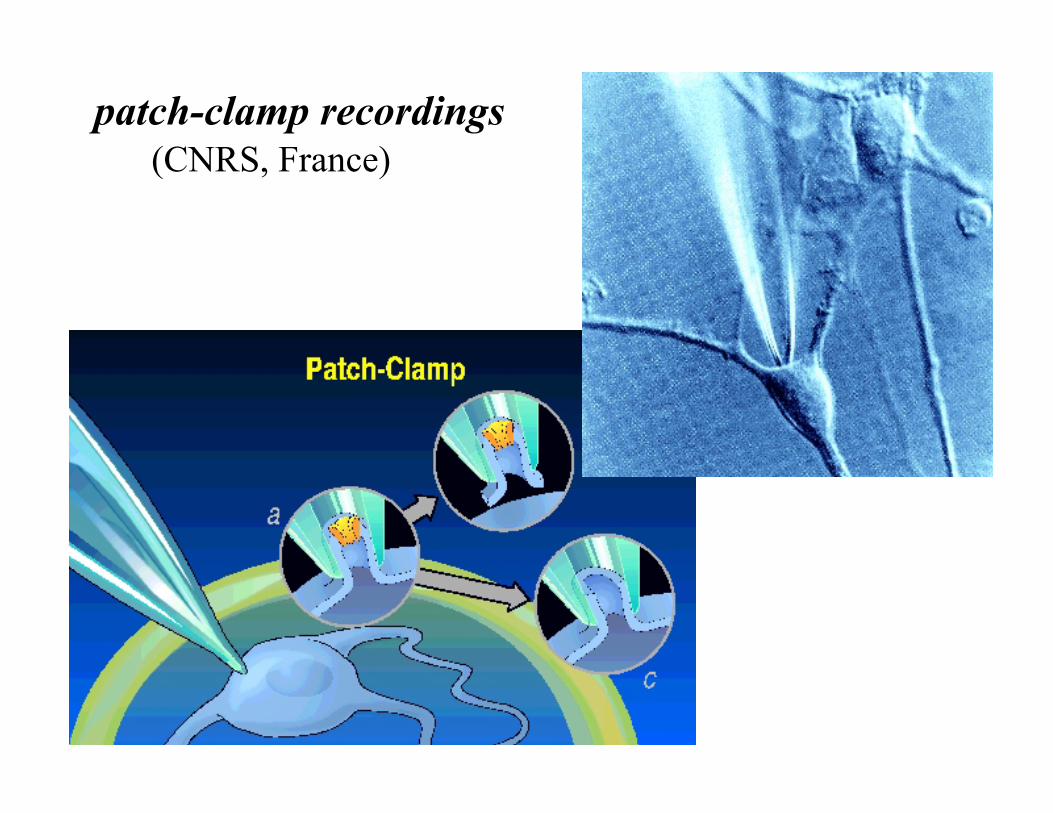
patch-clamp recordings (CNRS, France)
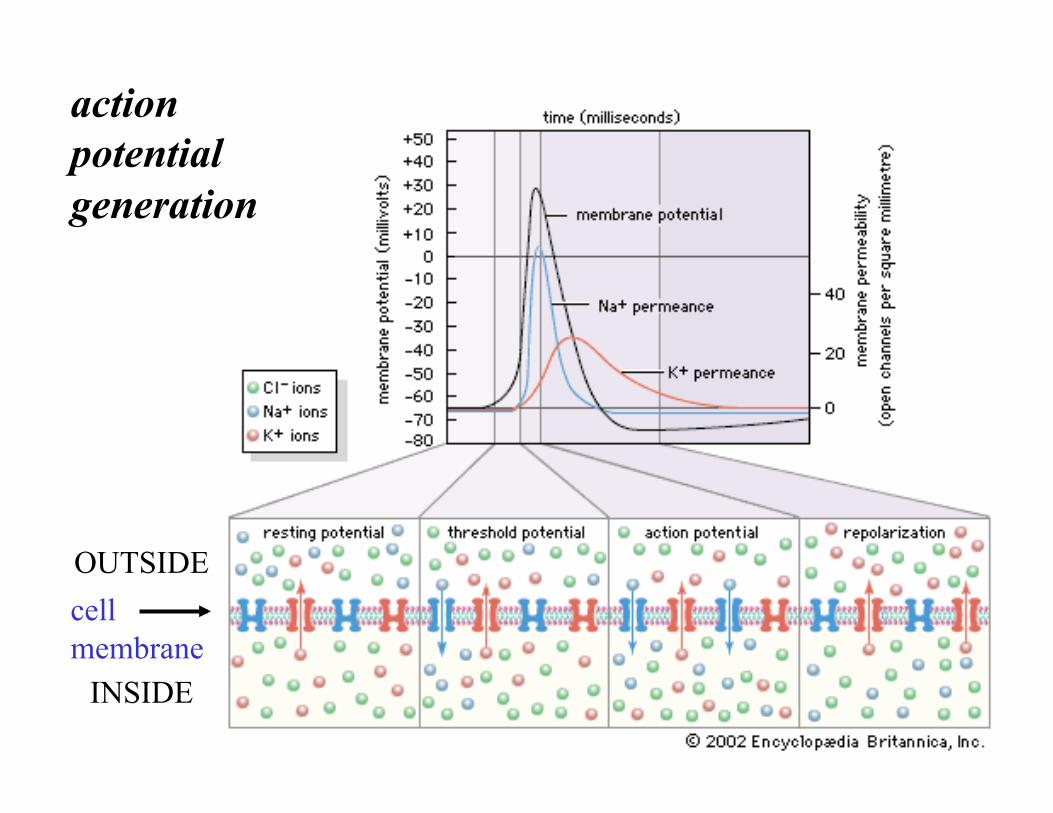
action potential generation
cell membrane
OUTSIDE
INSIDE
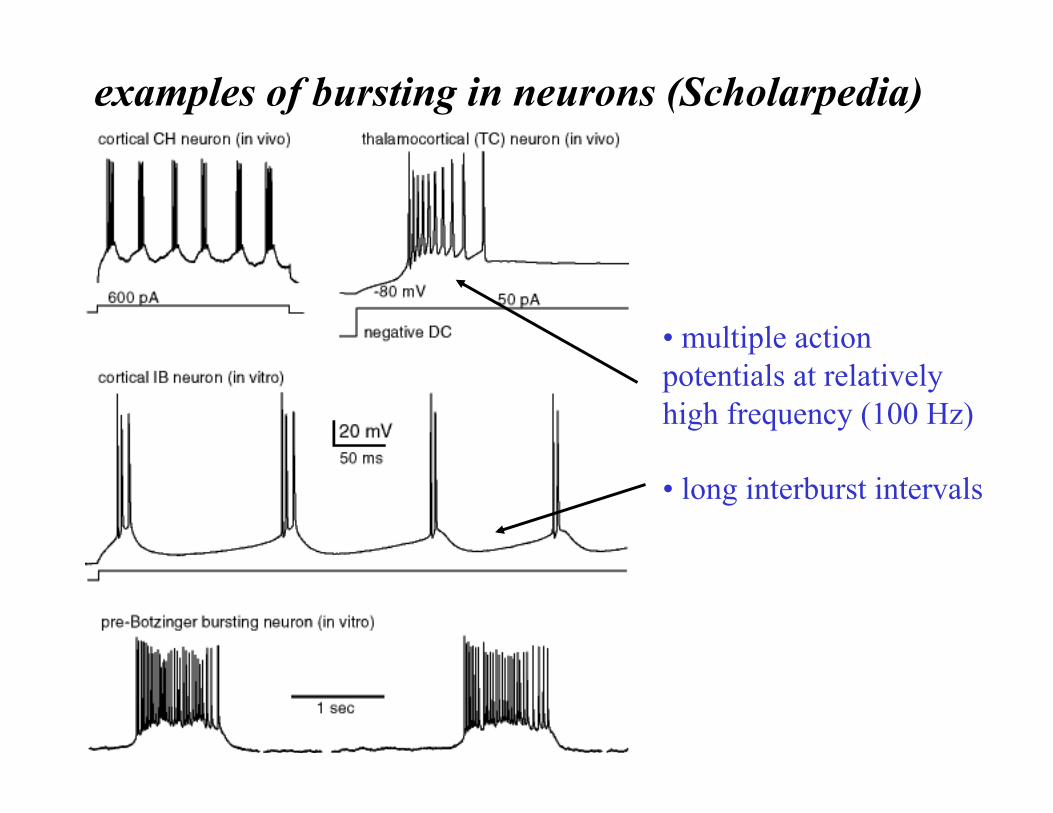
examples of bursting in neurons (Scholarpedia)
• multiple action potentials at relatively high frequency (100 Hz)
• long interburst intervals

why care about bursting?
1. mathematically interesting
2. plays a role in the brain
(a) sleep
(b) novelty detection
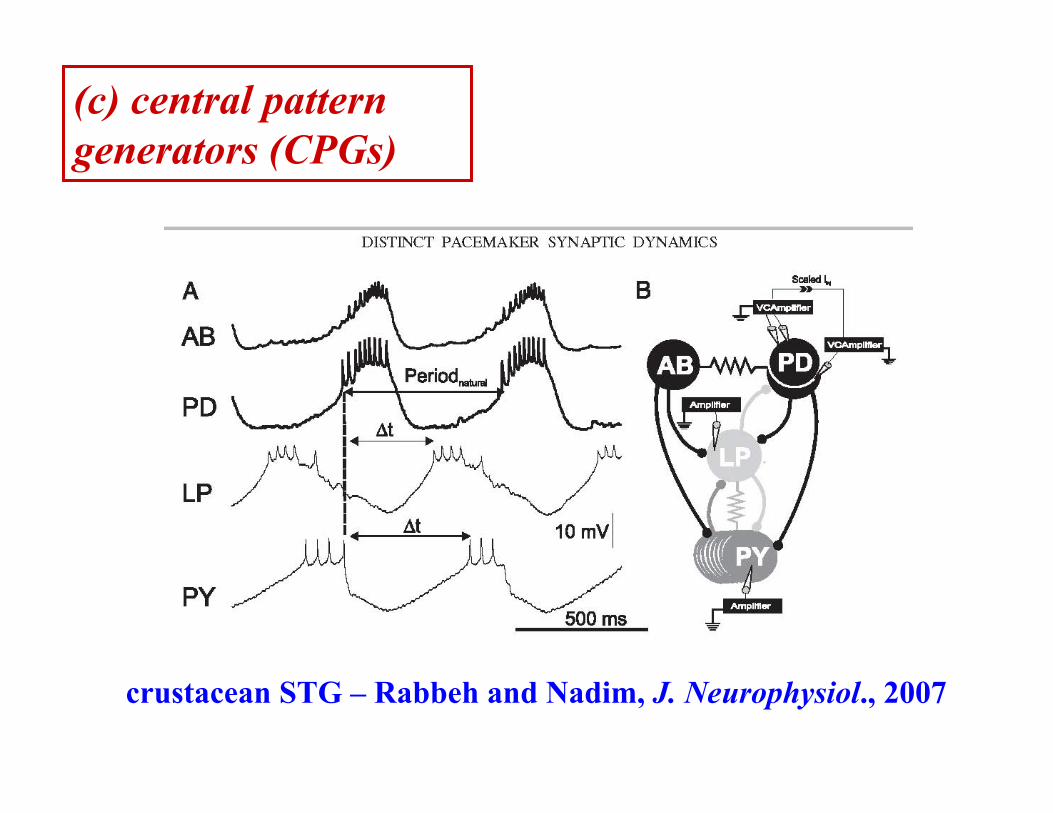
(c) central pattern generators (CPGs)
crustacean STG – Rabbeh and Nadim, J. Neurophysiol., 2007
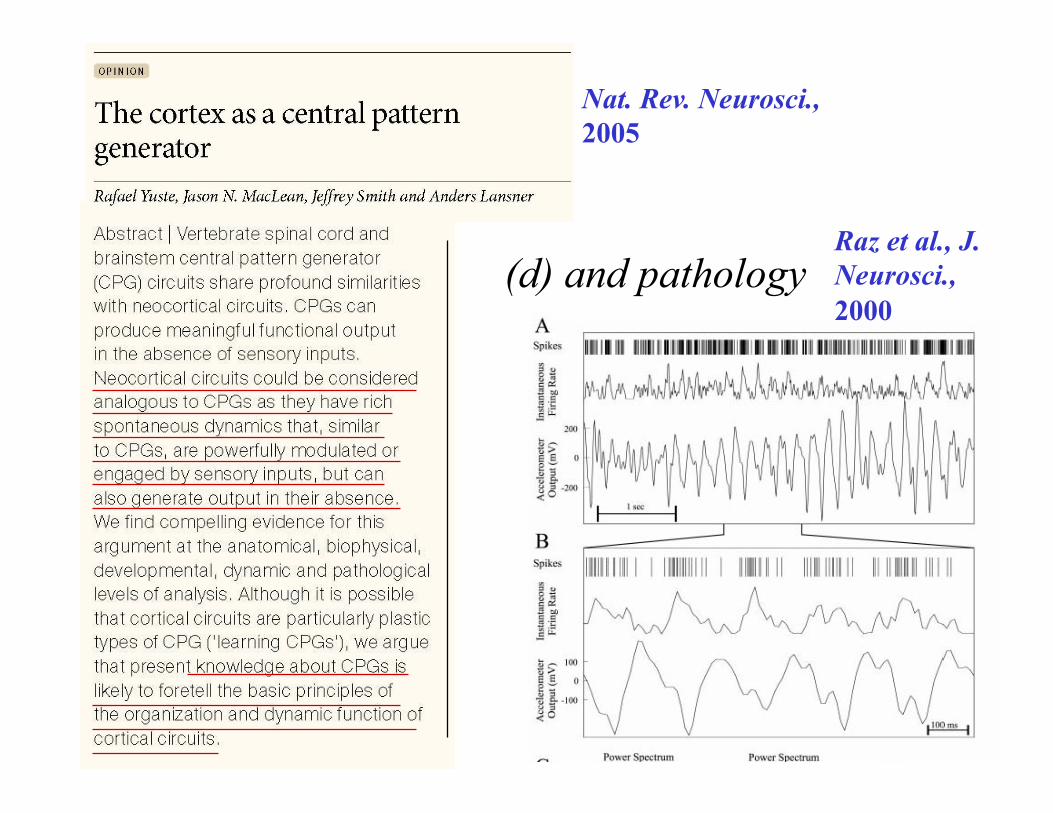
Nat. Rev. Neurosci., 2005
(d) and pathology Raz et al., J. Neurosci., 2000
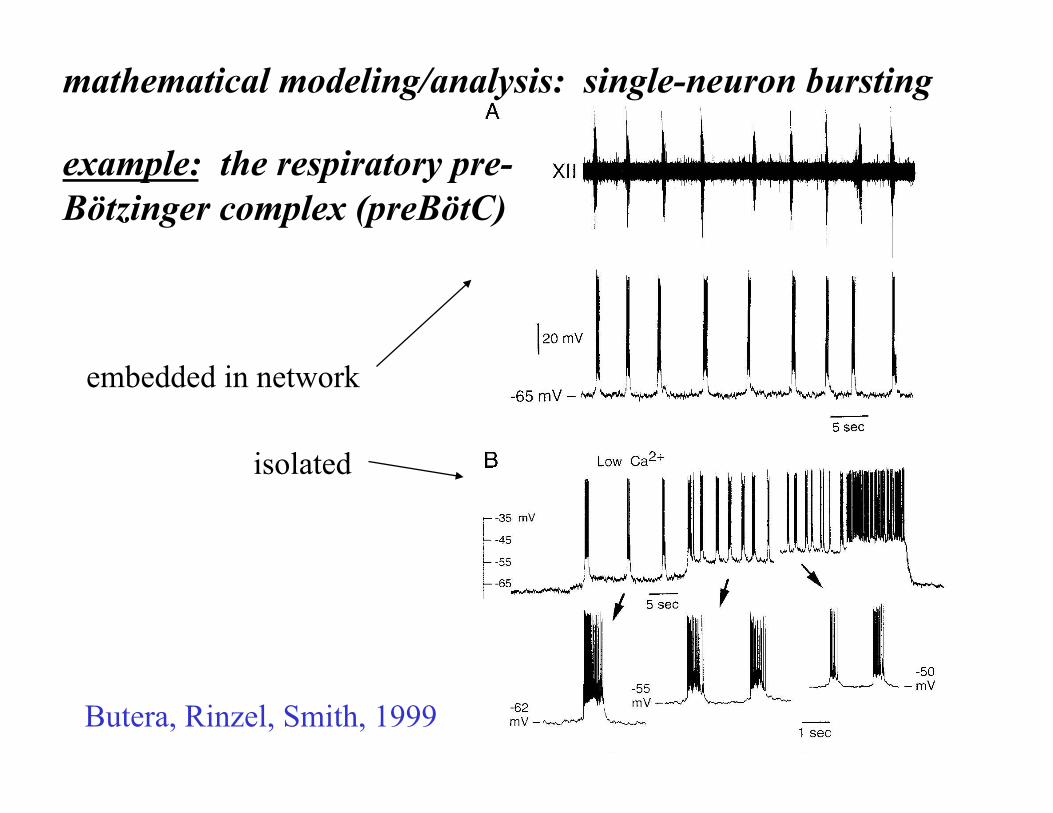
example: the respiratory pre-Bötzinger complex (preBötC)
embedded in network
isolated
Butera, Rinzel, Smith, 1999
mathematical modeling/analysis: single-neuron bursting
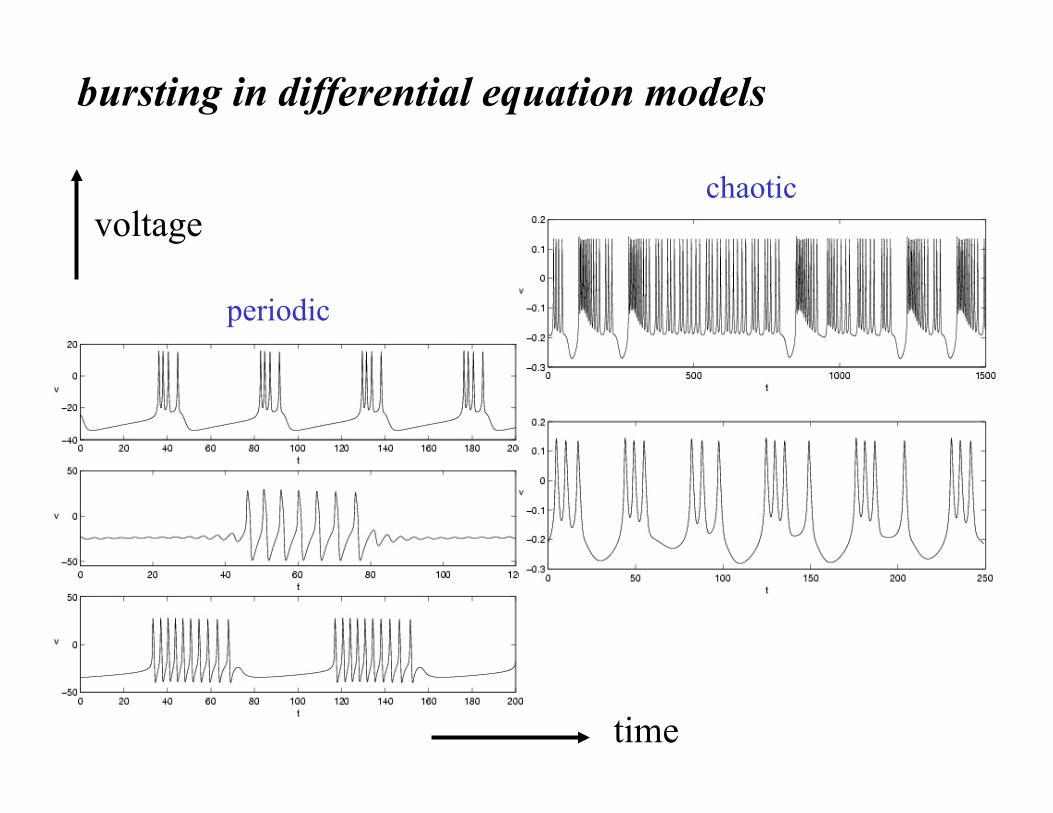
time
voltage
bursting in differential equation models
periodic
chaotic

Butera et al. model
Butera, Rinzel, Smith, 1999

fast-slow decomposition for bursting analysis (Rinzel, 1985+)
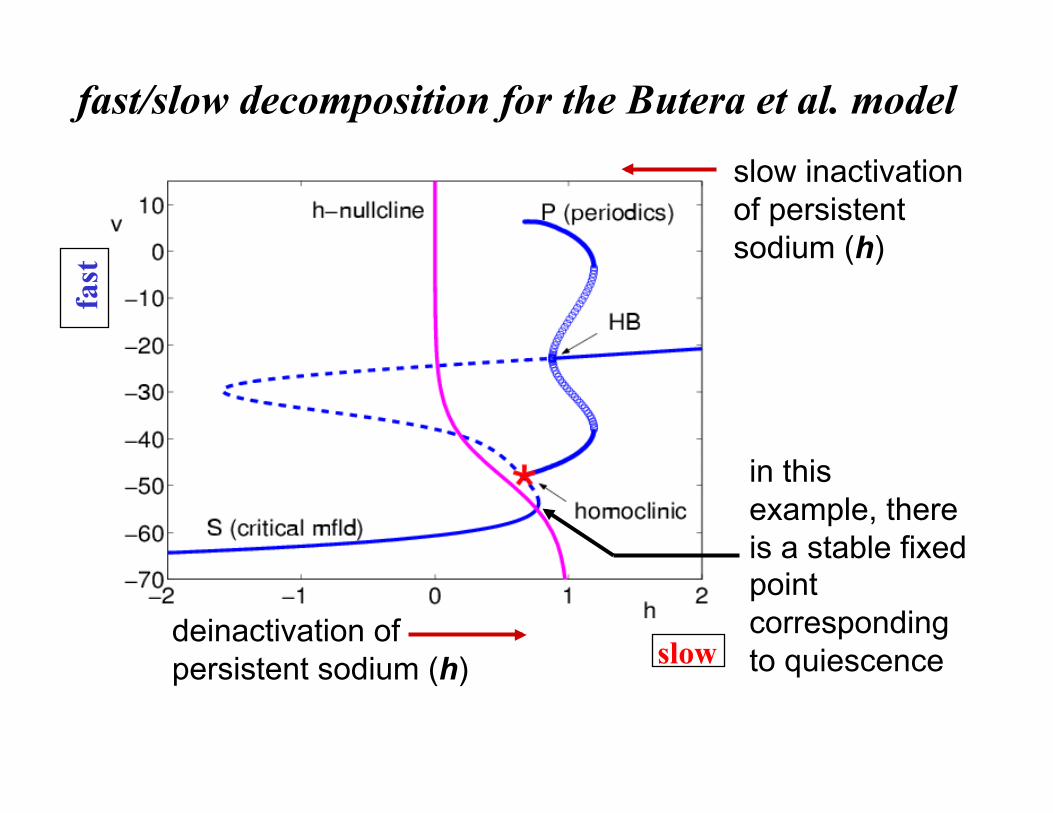
slow inactivation of persistent sodium (h)
fast/slow decomposition for the Butera et al. model
deinactivation of persistent sodium (h)
in this example, there is a stable fixed point corresponding to quiescence slow
fast
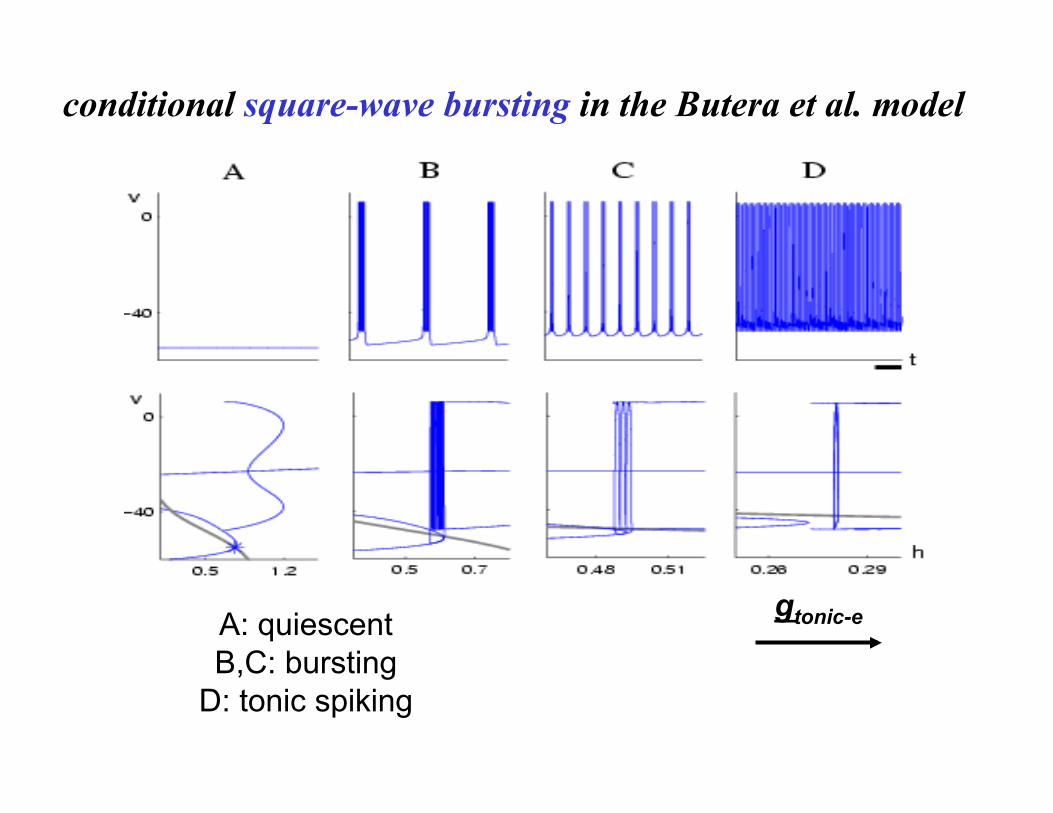
gtonic-e A: quiescent B,C: bursting
D: tonic spiking
conditional square-wave bursting in the Butera et al. model
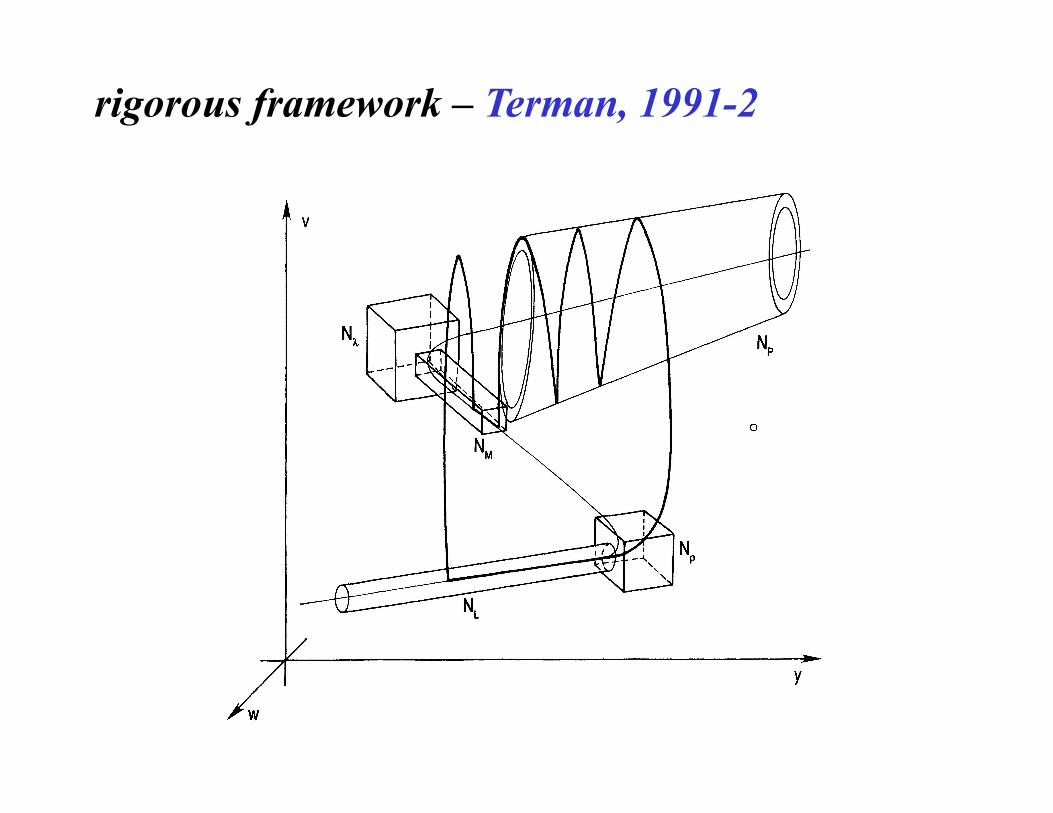
rigorous framework – Terman, 1991-2
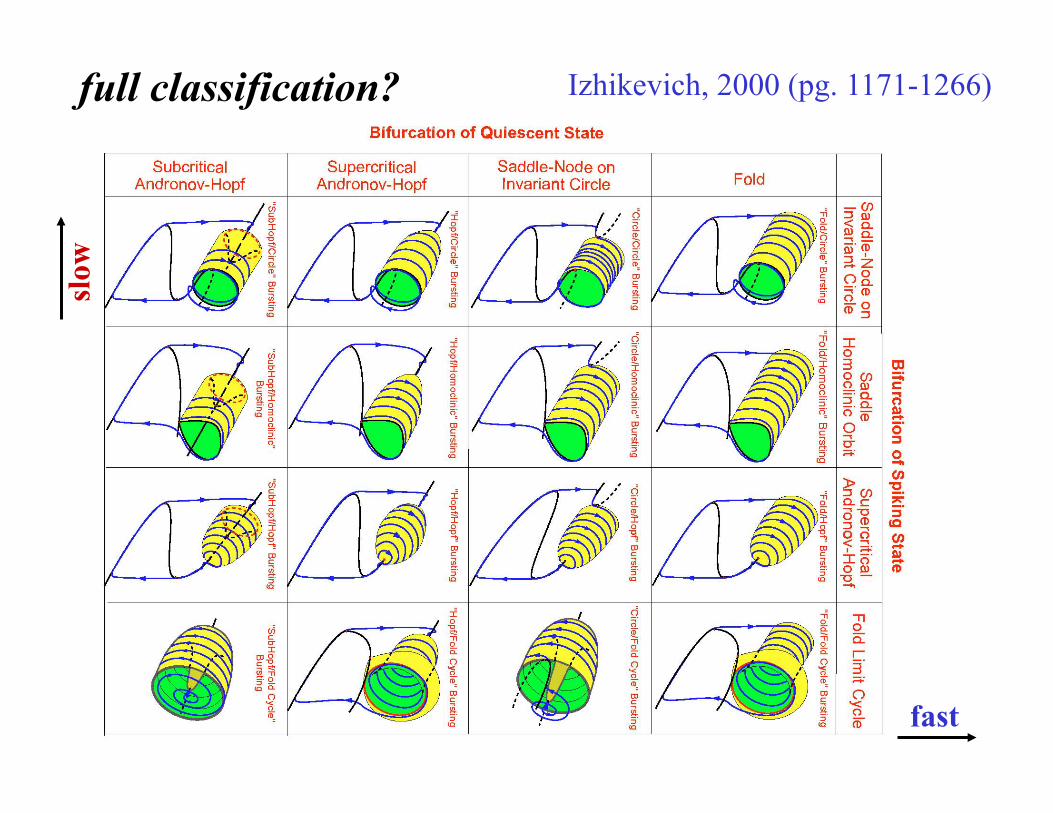
full classification? Izhikevich, 2000 (pg. 1171-1266) sl
ow
fast

case NOT closed – e.g., the Bursting book, 2005
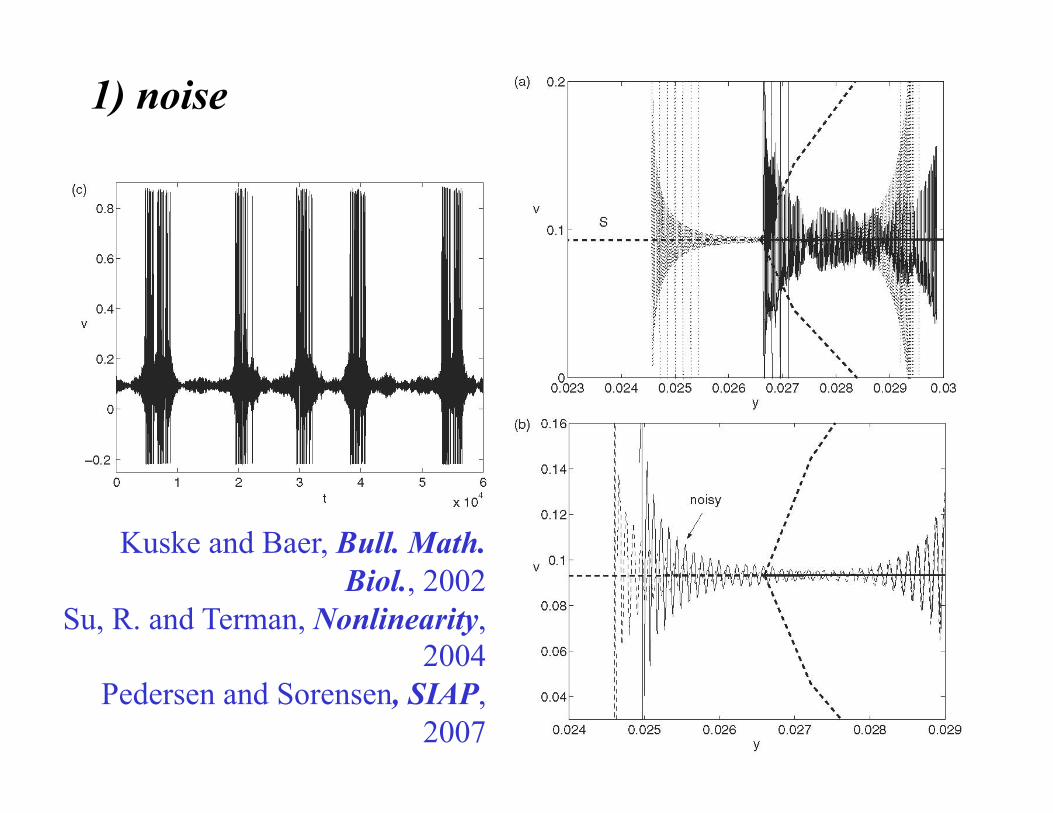
1) noise
Kuske and Baer, Bull. Math. Biol., 2002
Su, R. and Terman, Nonlinearity, 2004
Pedersen and Sorensen, SIAP, 2007
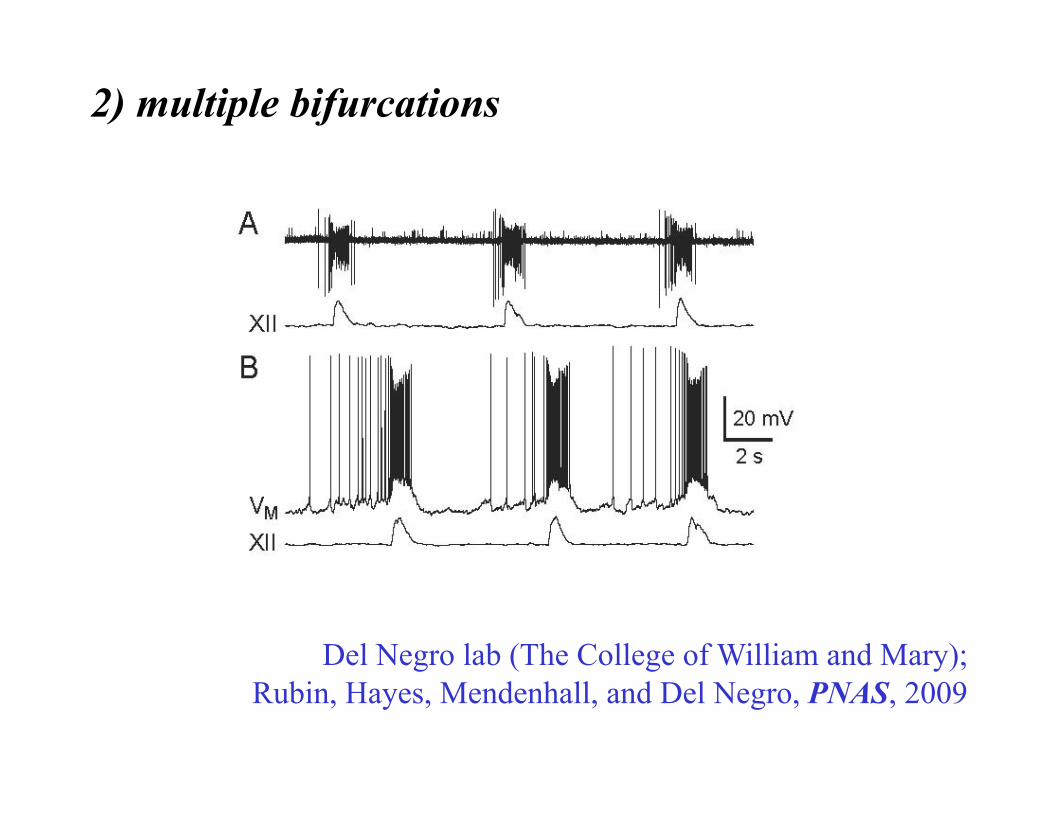
Del Negro lab (The College of William and Mary); Rubin, Hayes, Mendenhall, and Del Negro, PNAS, 2009
2) multiple bifurcations
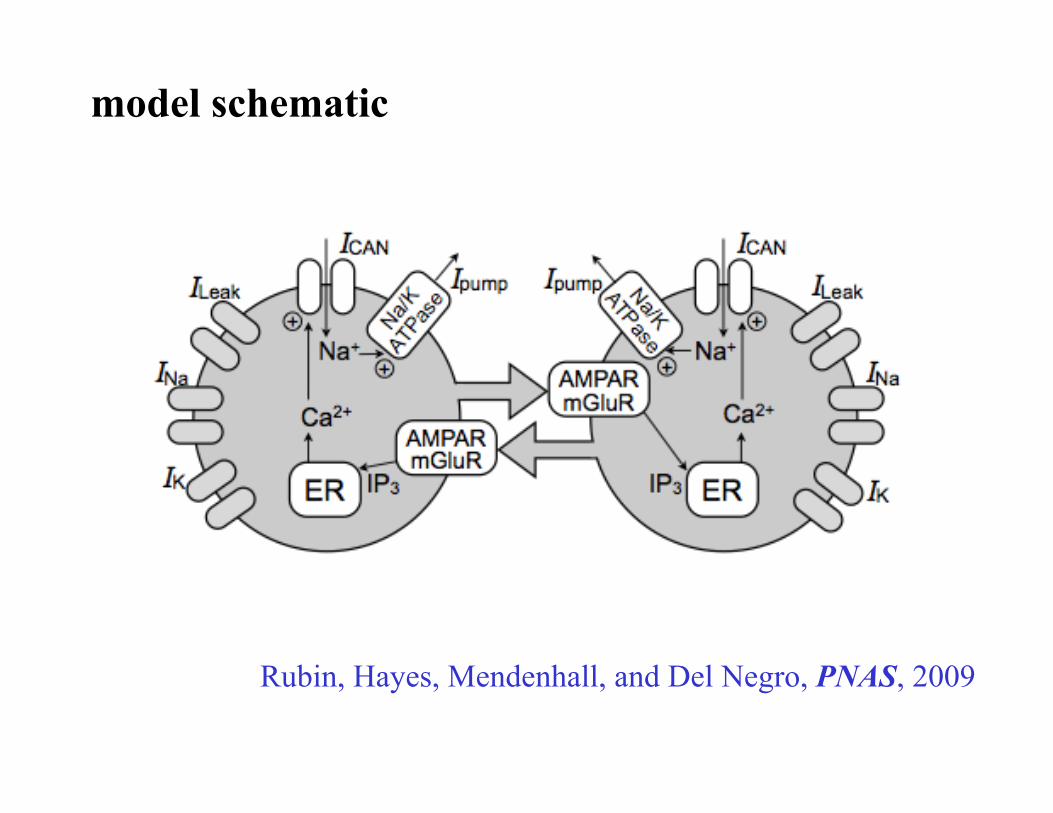
model schematic
Rubin, Hayes, Mendenhall, and Del Negro, PNAS, 2009
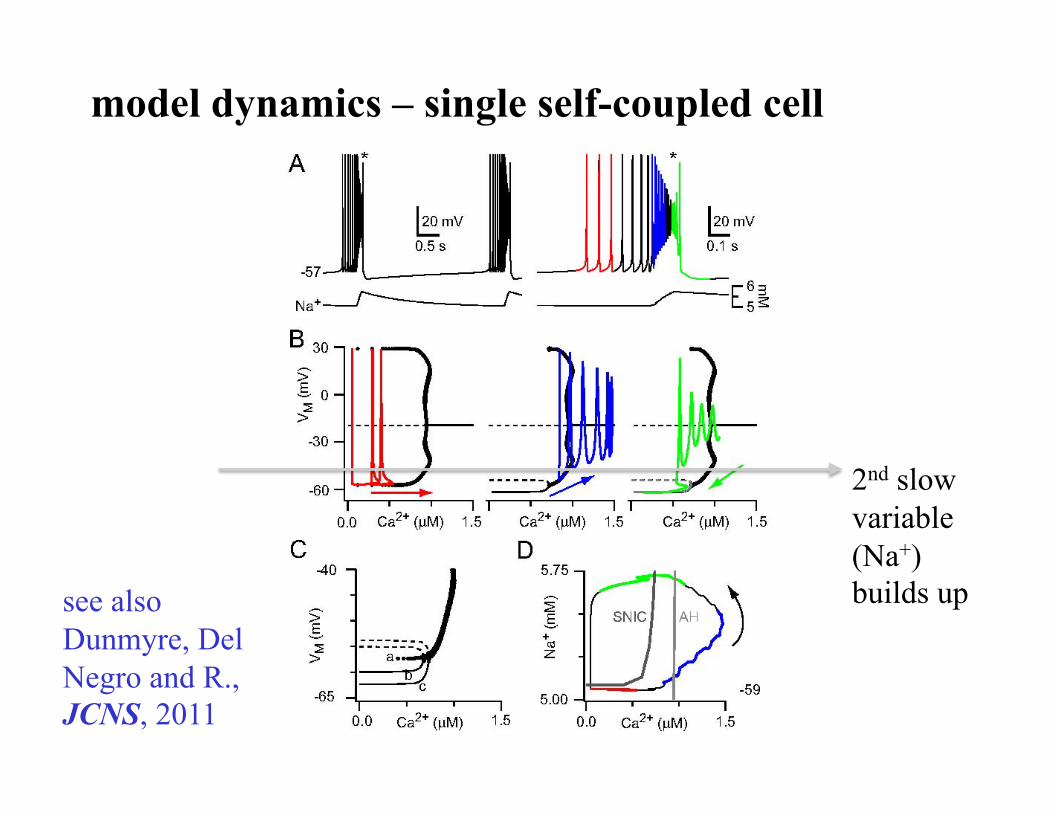
model dynamics – single self-coupled cell
2nd slow variable (Na+) builds up see also
Dunmyre, Del Negro and R., JCNS, 2011
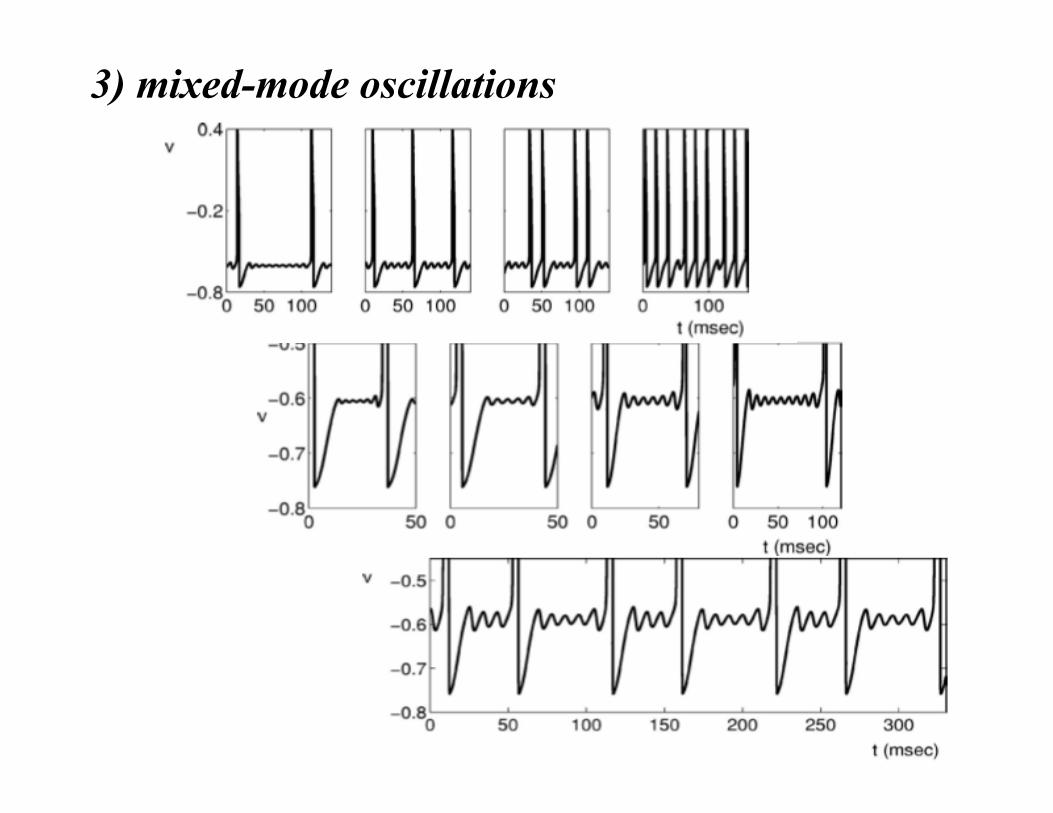
3) mixed-mode oscillations
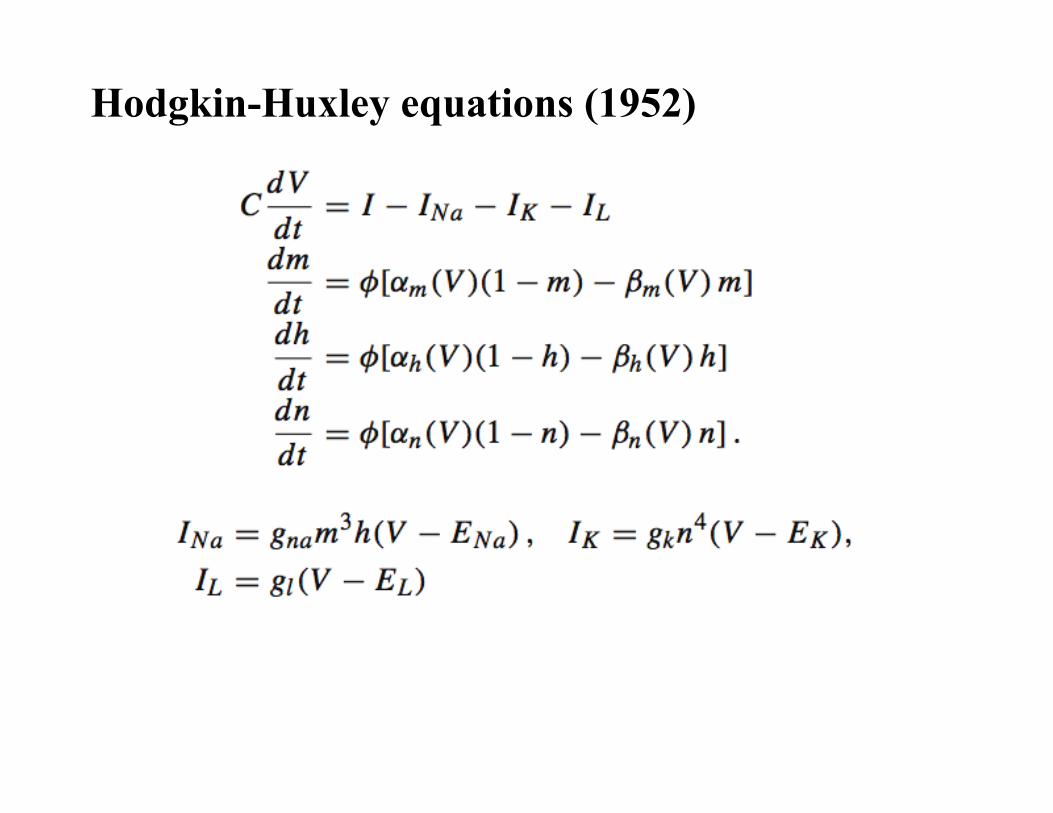
Hodgkin-Huxley equations (1952)
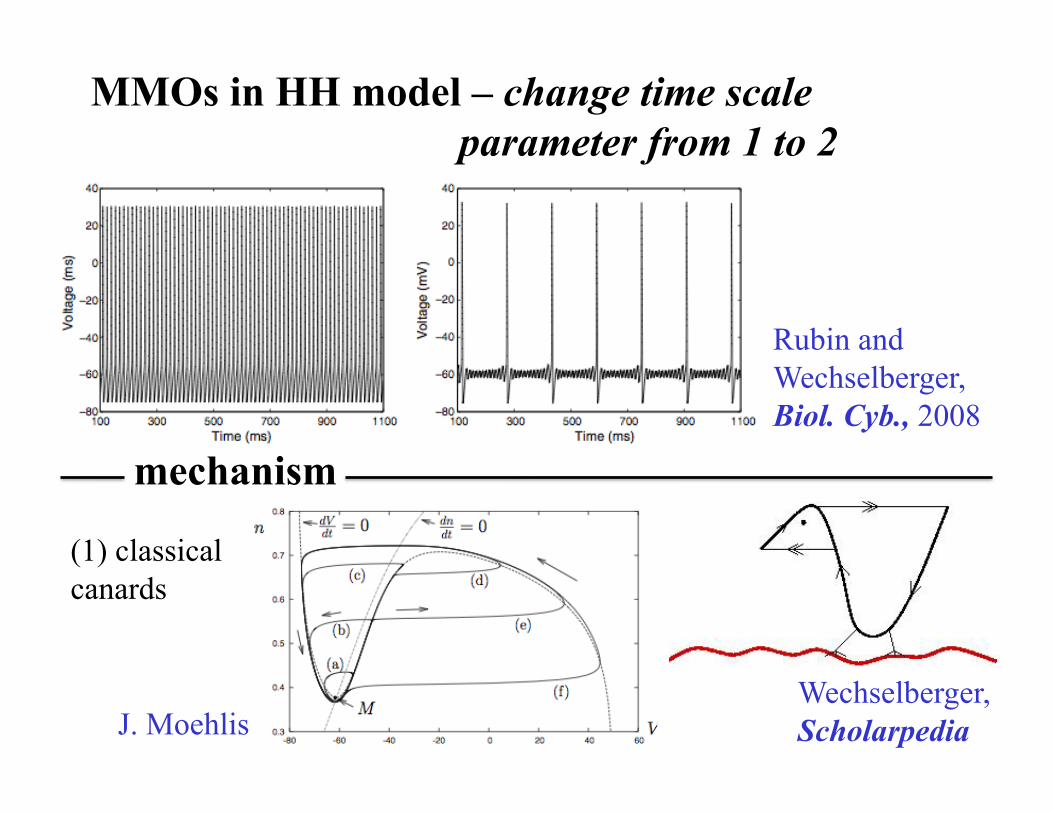
MMOs in HH model – change time scale parameter from 1 to 2
(1) classical canards
mechanism
Rubin and Wechselberger, Biol. Cyb., 2008
J. Moehlis Wechselberger, Scholarpedia
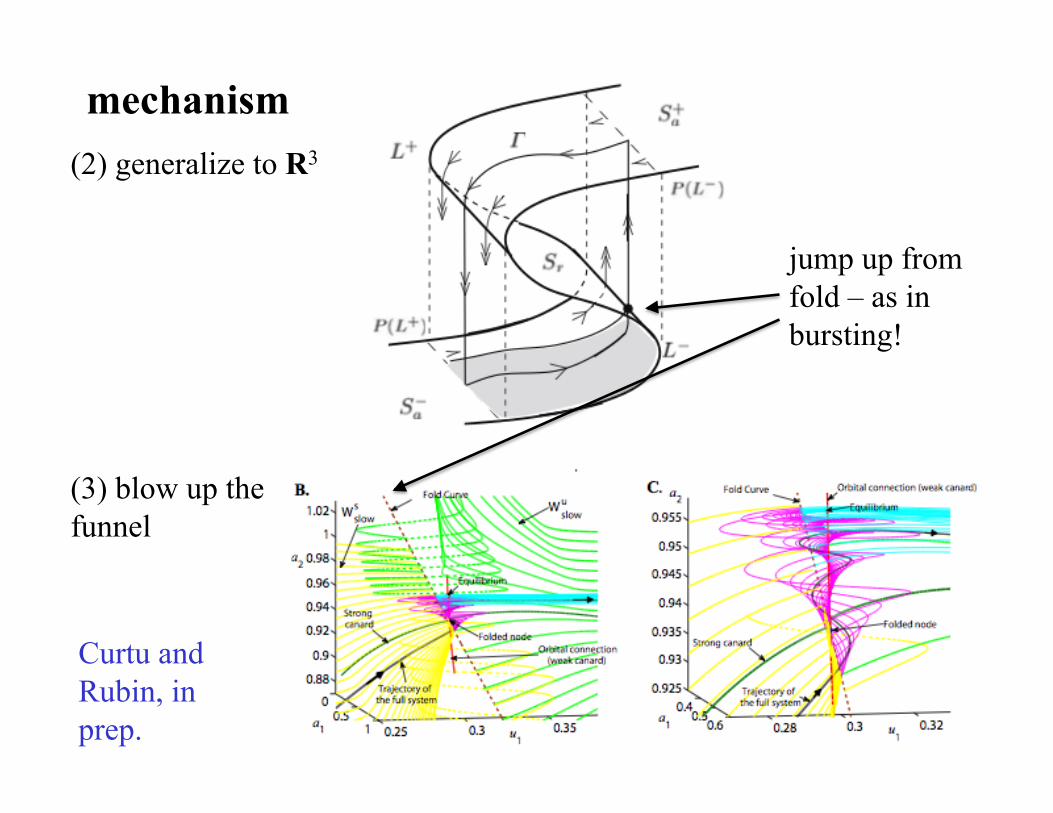
(2) generalize to R3
mechanism
jump up from fold – as in bursting!
(3) blow up the funnel
Curtu and Rubin, in prep.
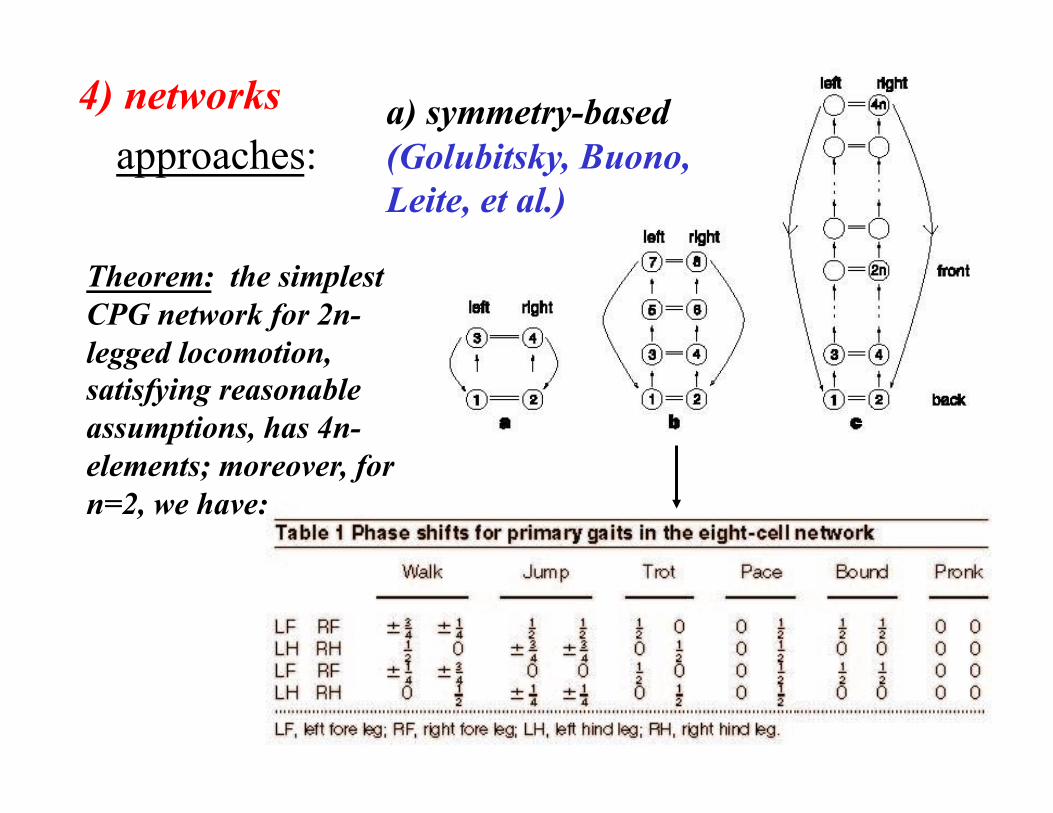
4) networks approaches:
a) symmetry-based (Golubitsky, Buono, Leite, et al.)
Theorem: the simplest CPG network for 2n-legged locomotion, satisfying reasonable assumptions, has 4n-elements; moreover, for n=2, we have:

b) connectivity-based (Pecora, Belykh, Hasler, et al.) – mostly for synchronization
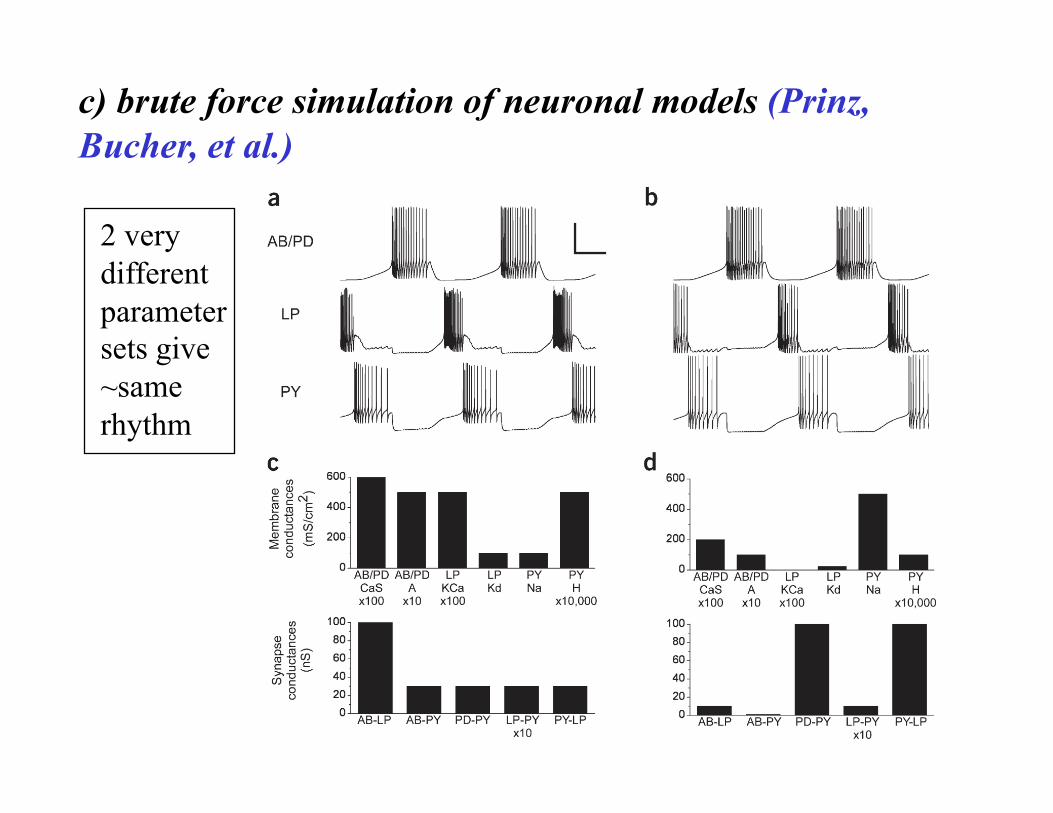
c) brute force simulation of neuronal models (Prinz, Bucher, et al.)
2 very different parameter sets give ~same rhythm
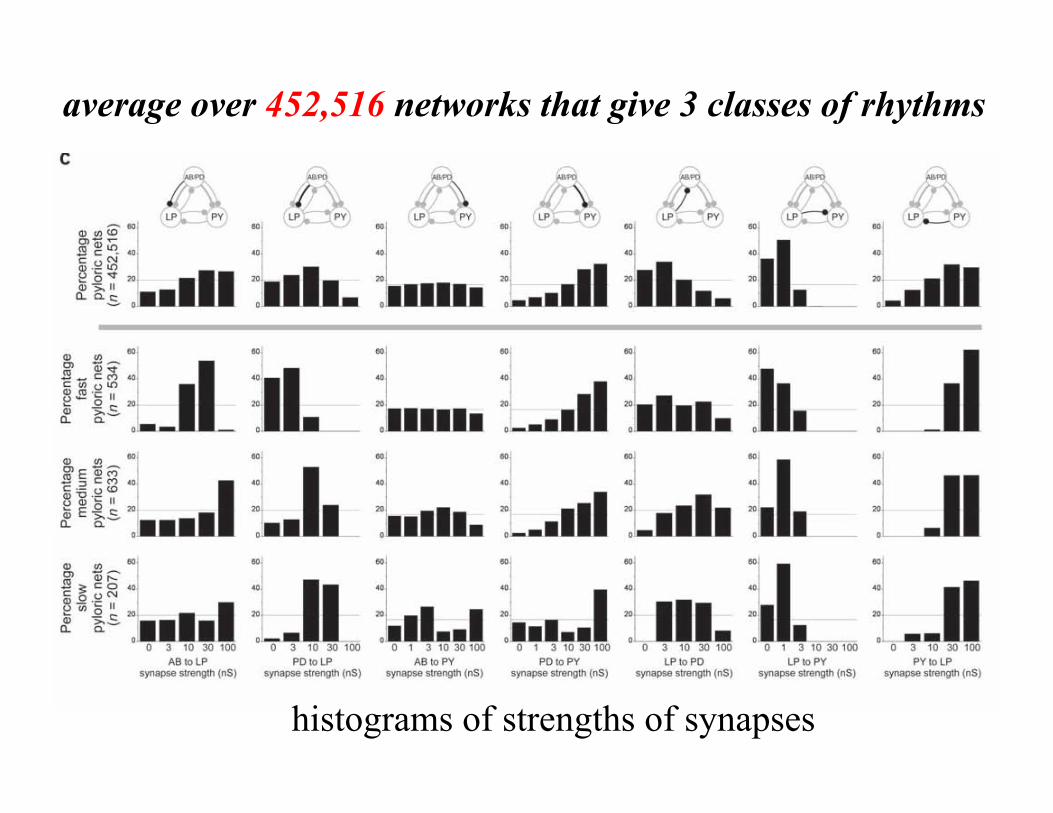
average over 452,516 networks that give 3 classes of rhythms
histograms of strengths of synapses
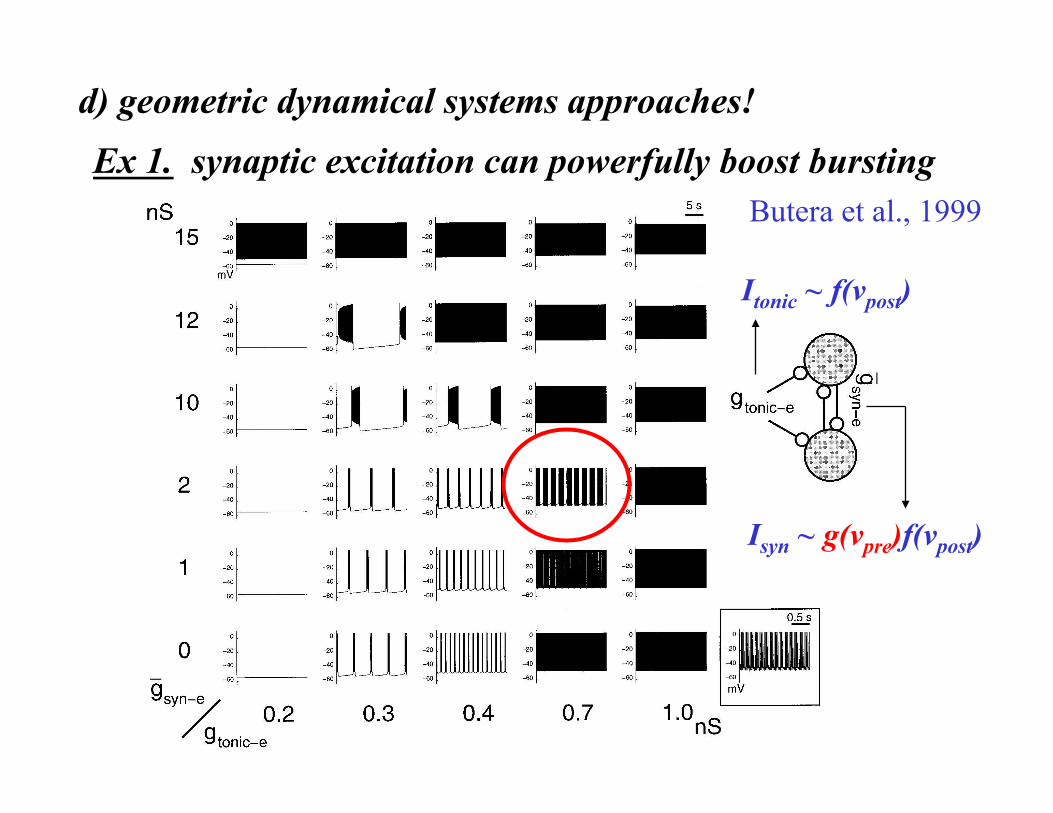
Ex 1. synaptic excitation can powerfully boost bursting
gtonic-e
Butera et al., 1999
Isyn ~ g(vpre)f(vpost)
Itonic ~ f(vpost)
d) geometric dynamical systems approaches!
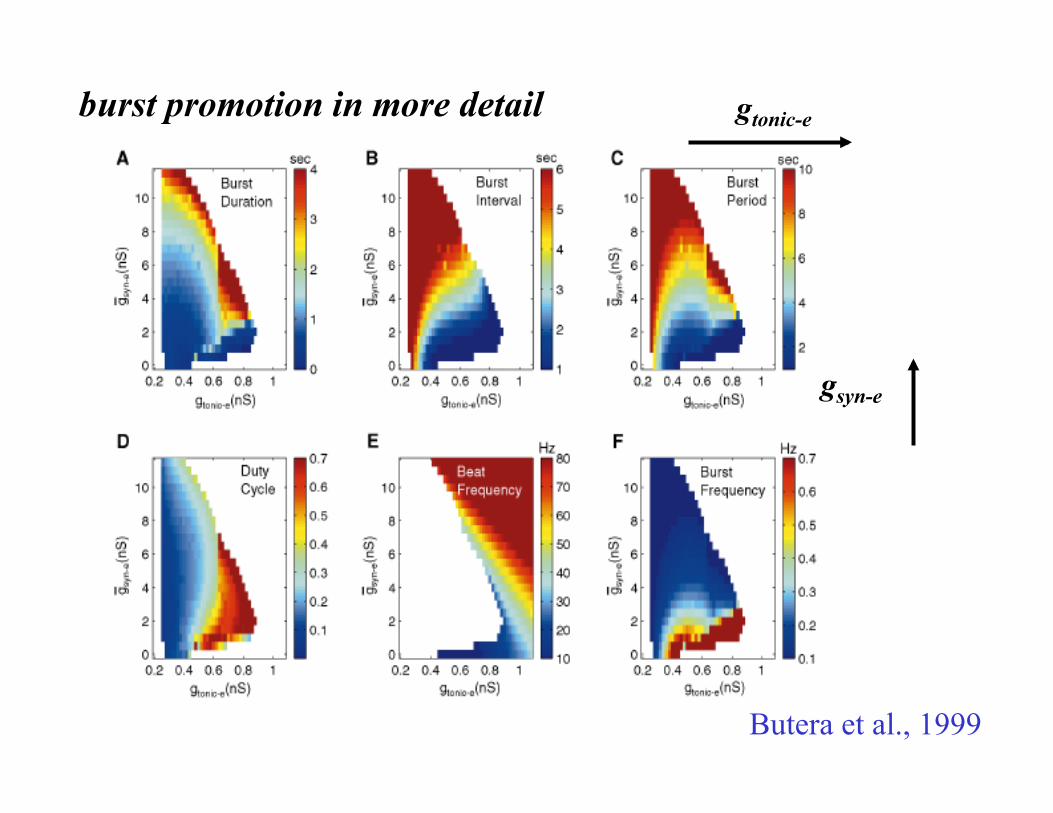
burst promotion in more detail
Butera et al., 1999
gtonic-e
gsyn-e

depend on vj
Butera model – but now we need 2 copies

• spike synchrony observed numerically to be unstable in all cases
Best, Borisyuk, Rubin, Terman, and Wechselberger, SIADS, 2005
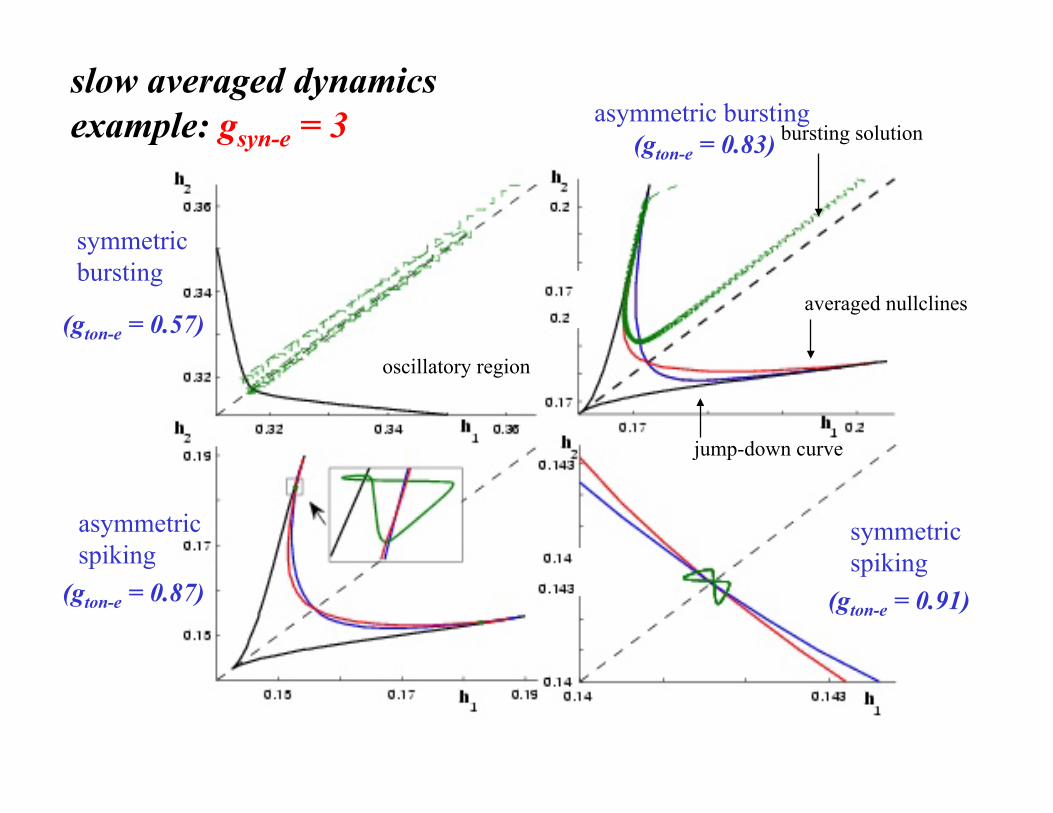
slow averaged dynamics example: gsyn-e = 3
(gton-e = 0.57)
(gton-e = 0.91) (gton-e = 0.87)
asymmetric bursting (gton-e = 0.83) bursting solution
averaged nullclines
jump-down curve
oscillatory region
symmetric bursting
asymmetric spiking
symmetric spiking
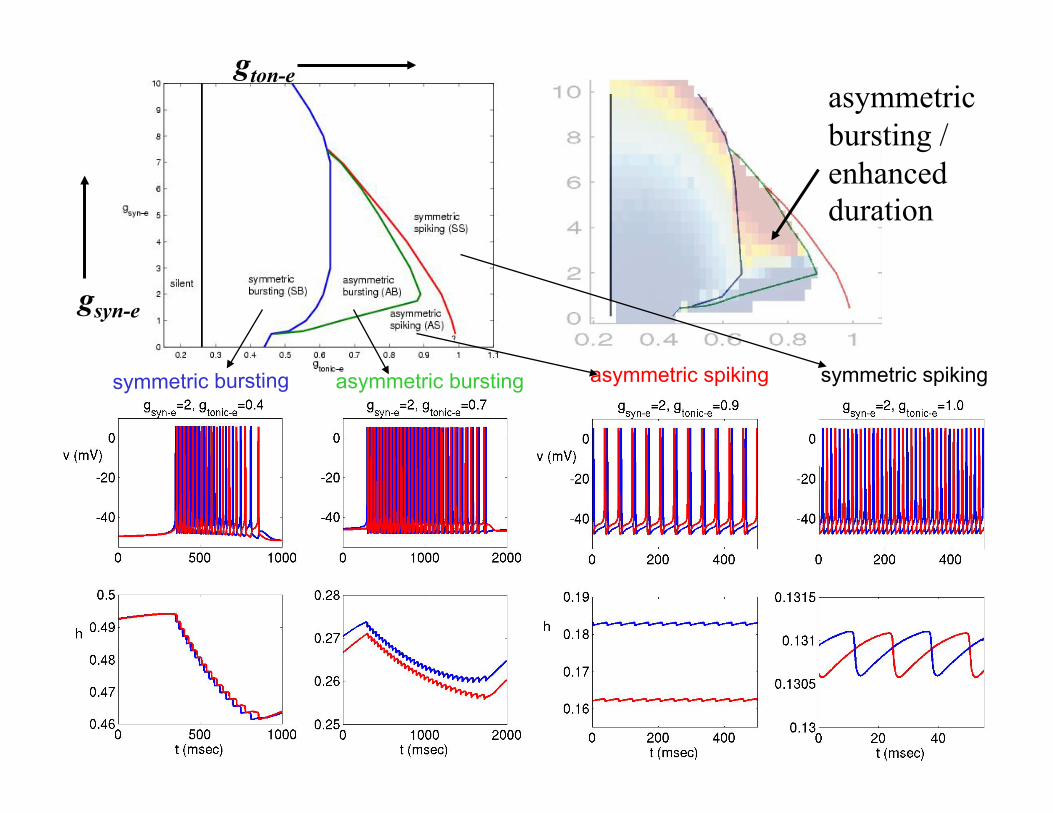
symmetric bursting asymmetric bursting asymmetric spiking symmetric spiking
gton-e
gsyn-e
asymmetric bursting / enhanced duration
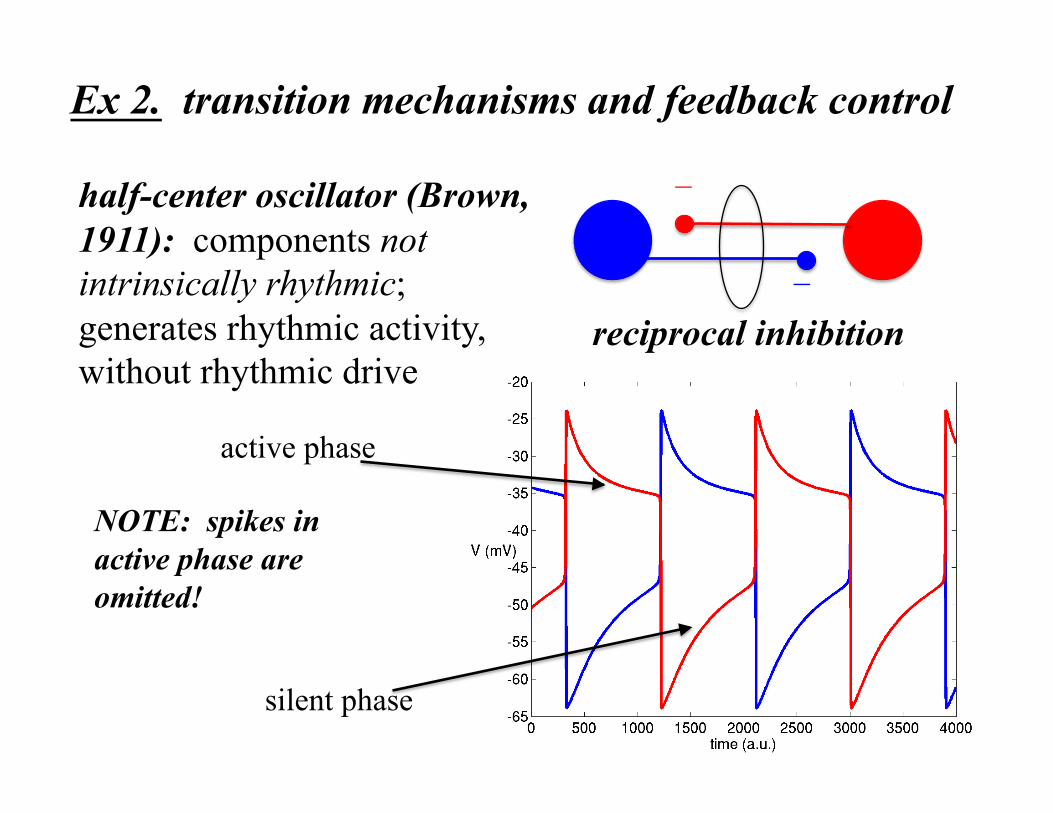
half-center oscillator (Brown, 1911): components not intrinsically rhythmic; generates rhythmic activity, without rhythmic drive
reciprocal inhibition
−
−
silent phase
active phase
Ex 2. transition mechanisms and feedback control
NOTE: spikes in active phase are omitted!
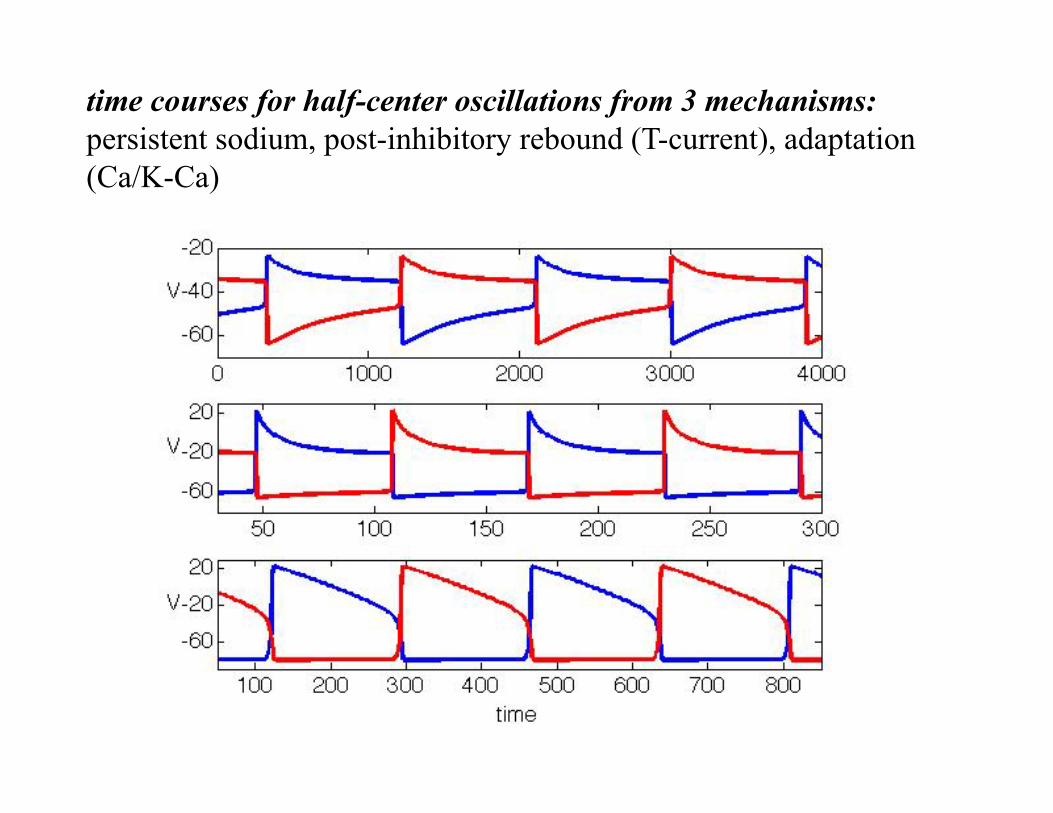
time courses for half-center oscillations from 3 mechanisms: persistent sodium, post-inhibitory rebound (T-current), adaptation (Ca/K-Ca)
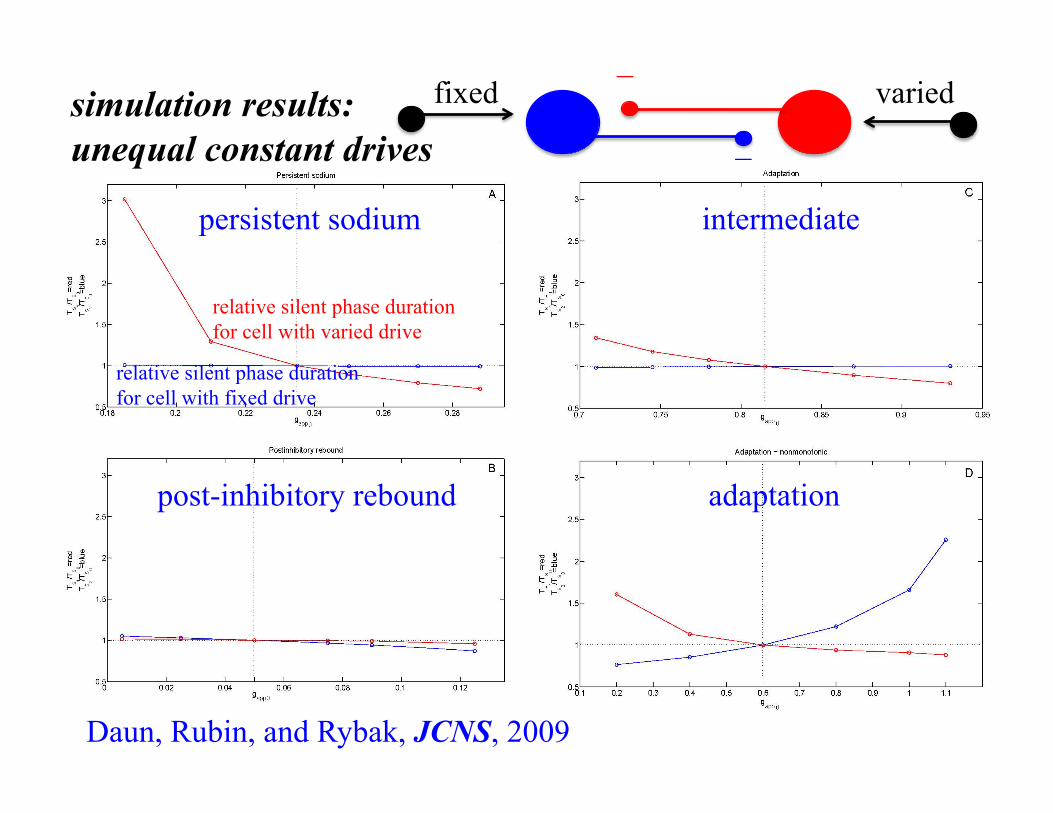
simulation results: unequal constant drives
intermediate
adaptation
persistent sodium
post-inhibitory rebound
relative silent phase duration for cell with varied drive
relative silent phase duration for cell with fixed drive
Daun, Rubin, and Rybak, JCNS, 2009
fixed varied −
−
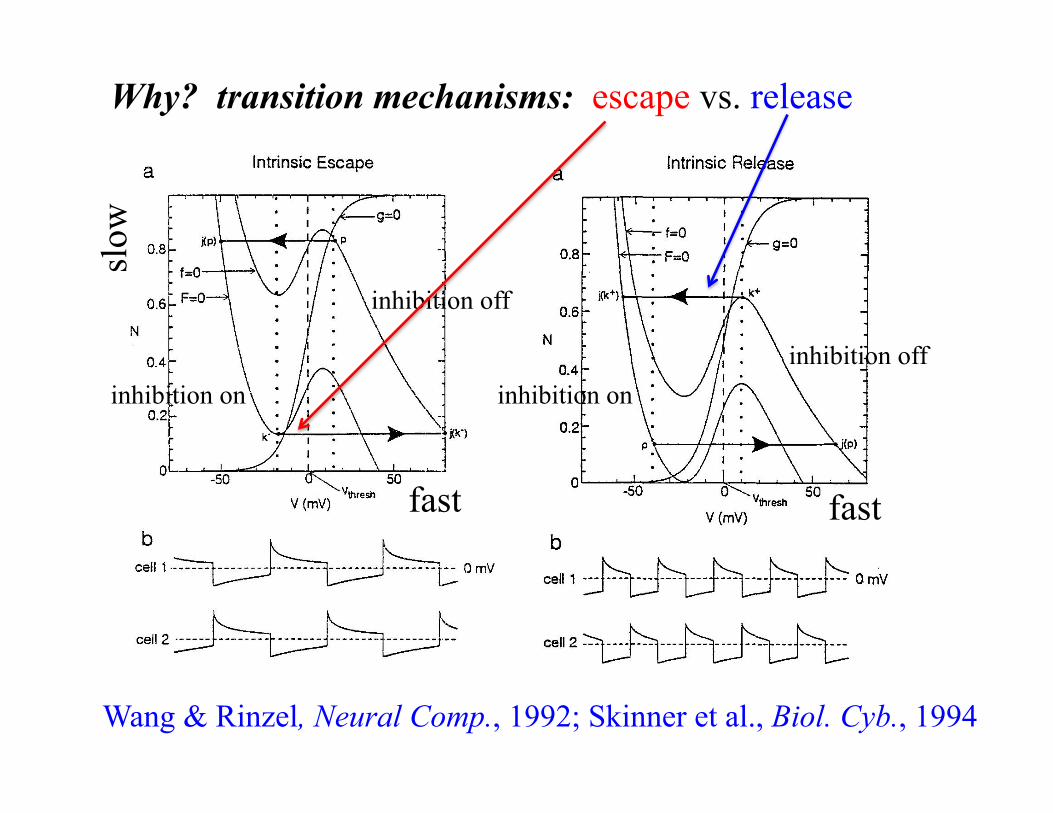
Why? transition mechanisms: escape vs. release
Wang & Rinzel, Neural Comp., 1992; Skinner et al., Biol. Cyb., 1994
inhibition on
inhibition off
inhibition on inhibition off
fast fast
slow

Summary
• escape: independent phase modulation (e.g., persistent sodium current)
• release: poor phase modulation (e.g., post-inhibitory rebound)
• adaptation = mix of release and escape: phase modulation NOT independent (e.g., Ca/K-Ca currents)
Daun, Rubin, and Rybak, JCNS, 2009
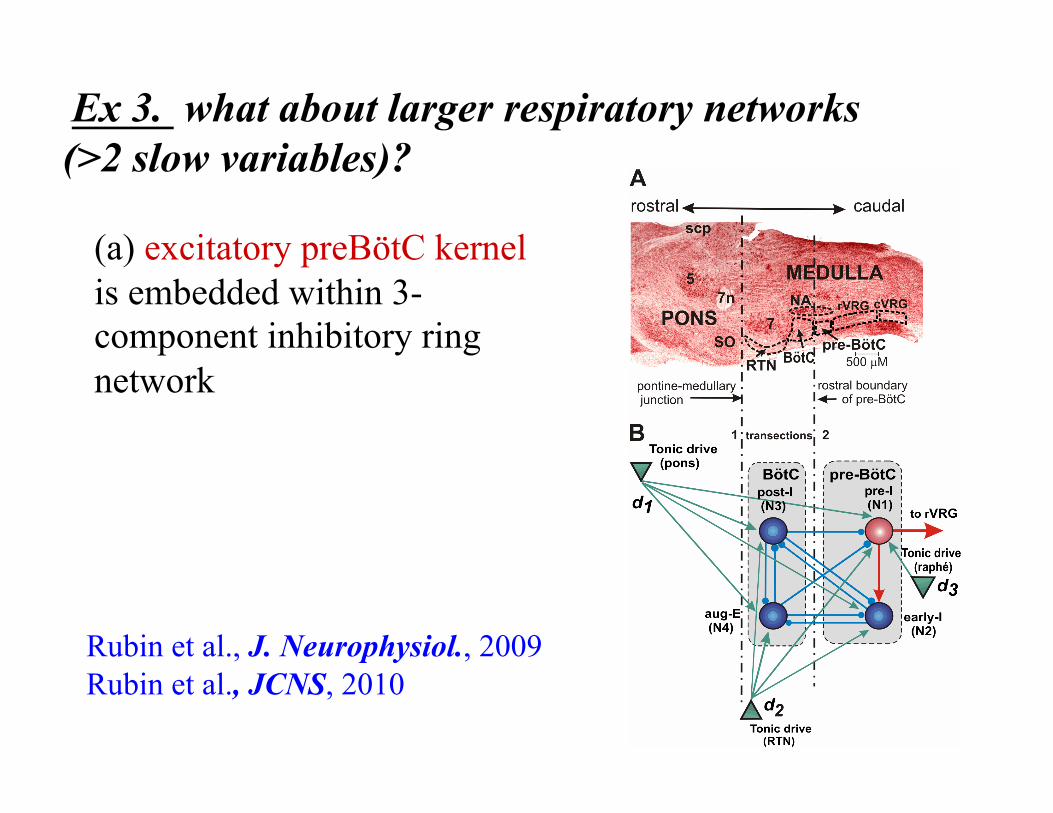
Ex 3. what about larger respiratory networks (>2 slow variables)?
(a) excitatory preBötC kernel is embedded within 3-component inhibitory ring network
Rubin et al., J. Neurophysiol., 2009 Rubin et al., JCNS, 2010
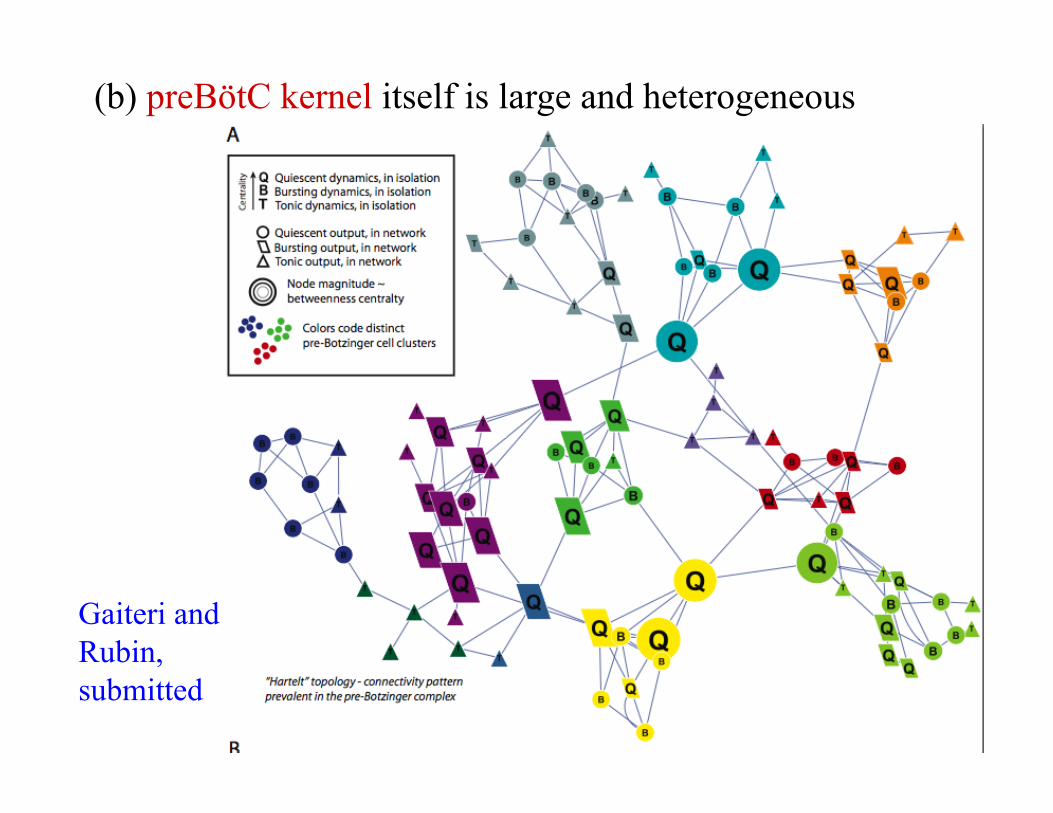
(b) preBötC kernel itself is large and heterogeneous
Gaiteri and Rubin, submitted
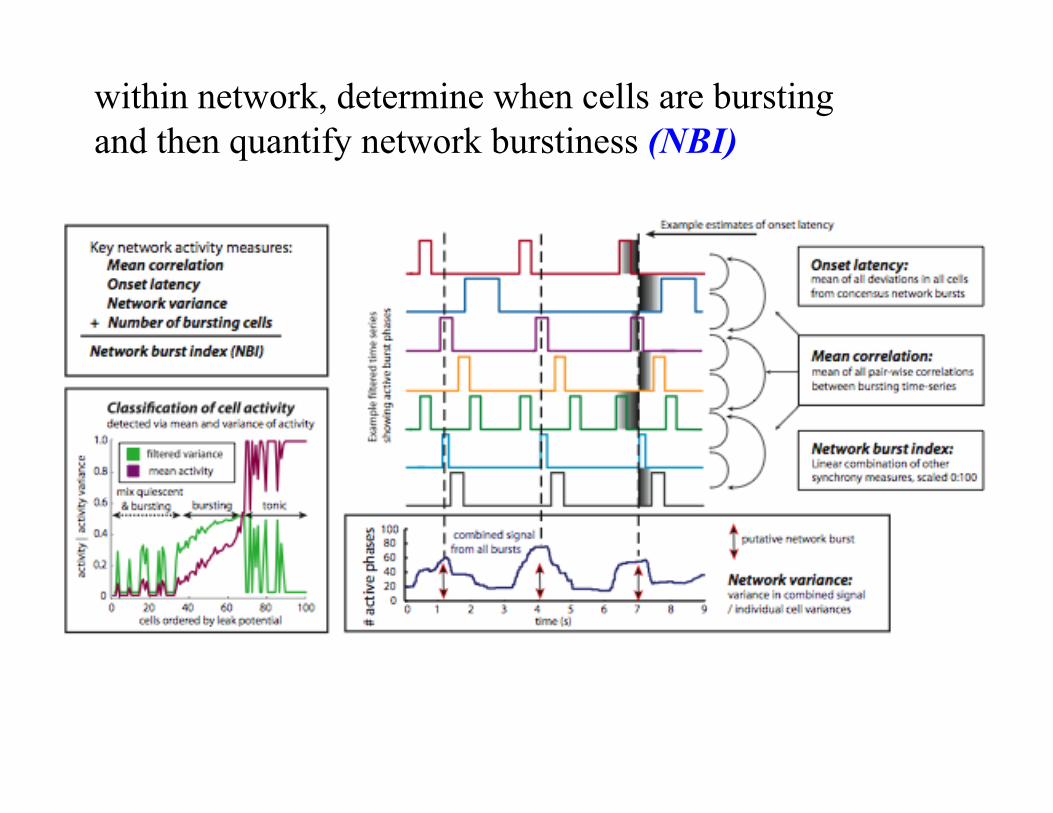
within network, determine when cells are bursting and then quantify network burstiness (NBI)
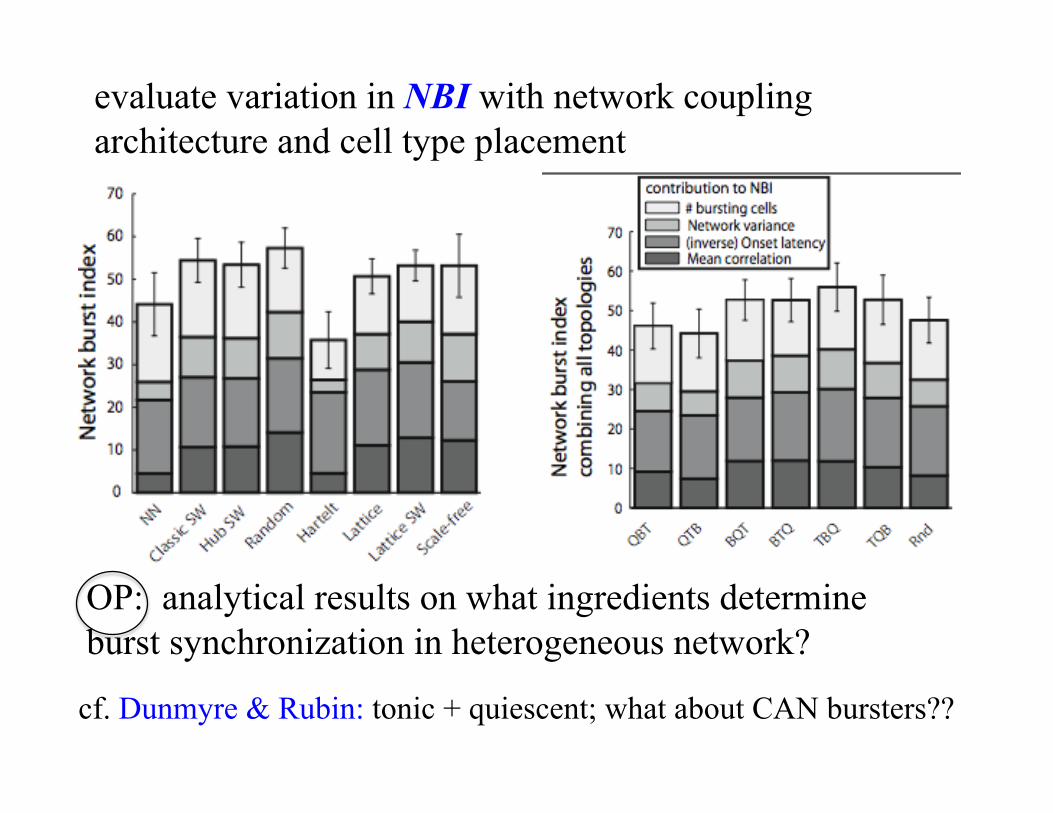
evaluate variation in NBI with network coupling architecture and cell type placement
OP: analytical results on what ingredients determine burst synchronization in heterogeneous network?
cf. Dunmyre & Rubin: tonic + quiescent; what about CAN bursters??
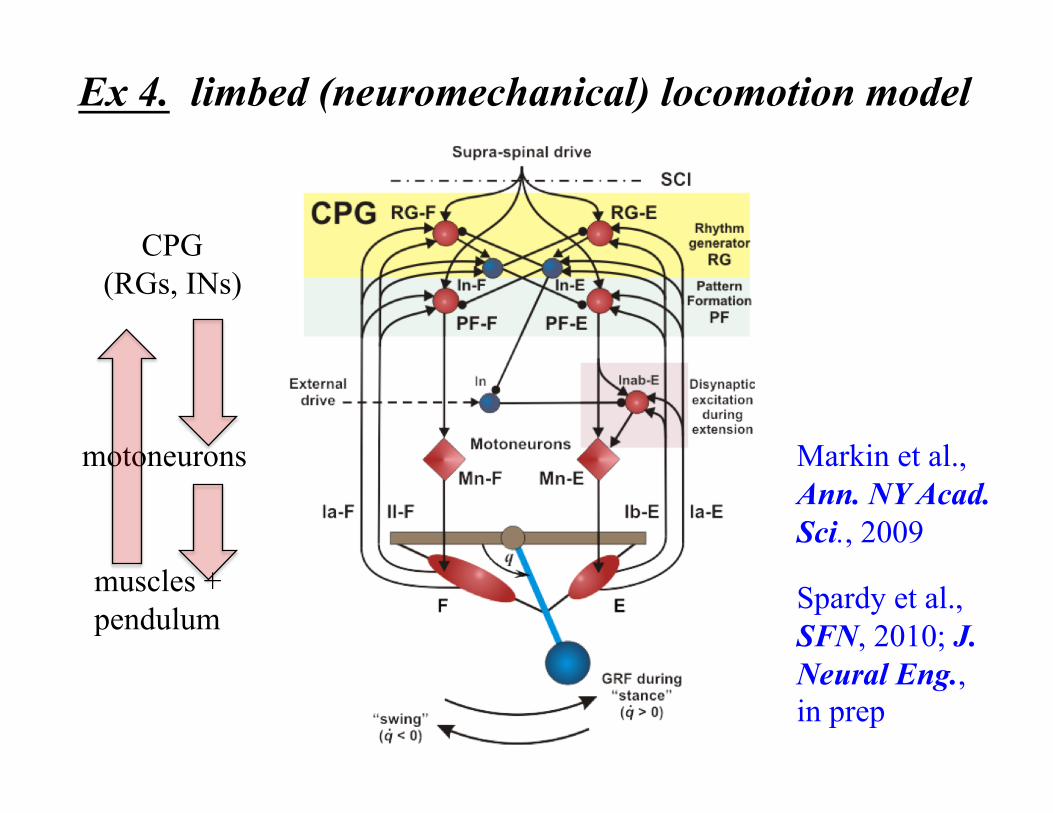
Ex 4. limbed (neuromechanical) locomotion model
Markin et al., Ann. NY Acad. Sci., 2009
Spardy et al., SFN, 2010; J. Neural Eng., in prep
CPG (RGs, INs)
motoneurons
muscles + pendulum
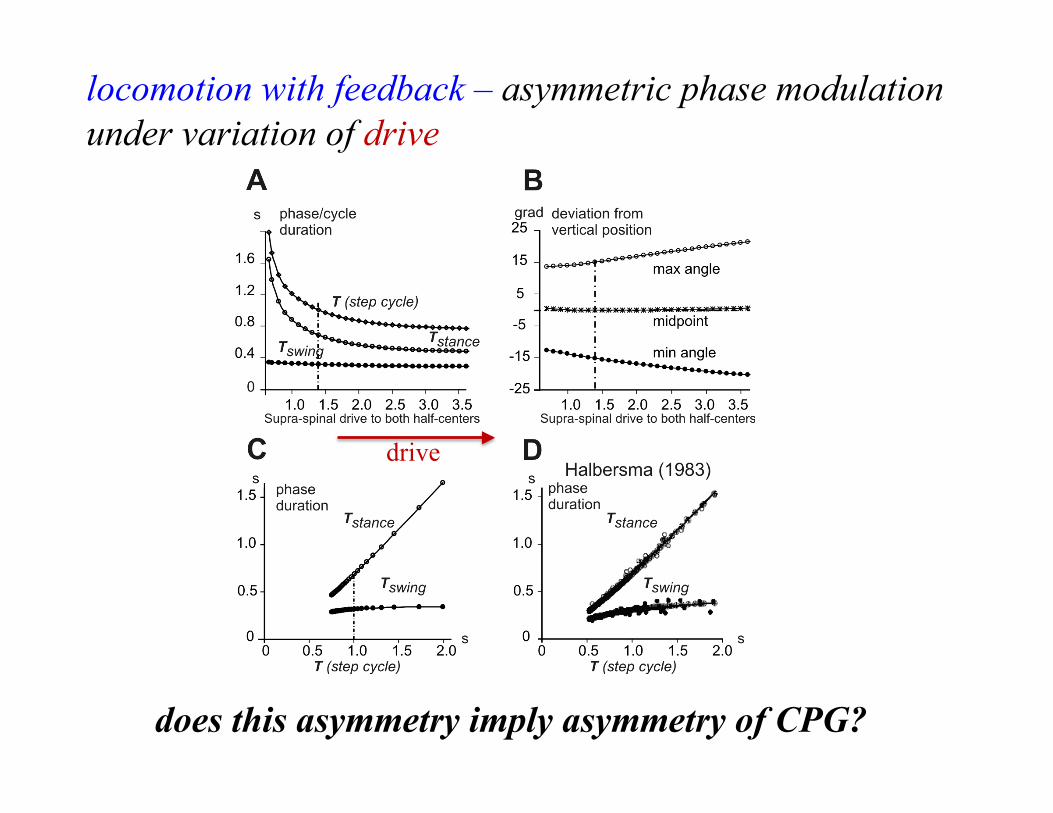
drive
locomotion with feedback – asymmetric phase modulation under variation of drive
does this asymmetry imply asymmetry of CPG?
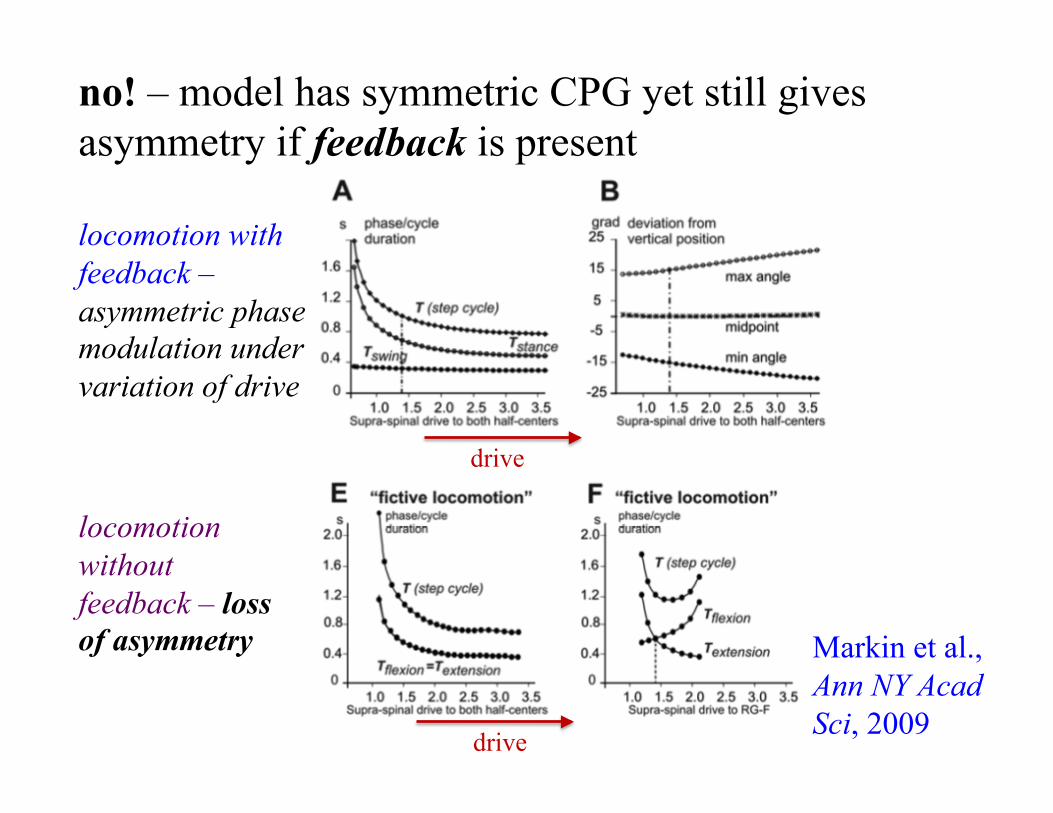
locomotion with feedback – asymmetric phase modulation under variation of drive
locomotion without feedback – loss of asymmetry
drive
drive
no! – model has symmetric CPG yet still gives asymmetry if feedback is present
Markin et al., Ann NY Acad Sci, 2009

rhythm with/without feedback: what is the difference?
with feedback
IN escape controls phase transitions
Lucy Spardy

without feedback
RG escape controls phase transitions
Lucy Spardy
rhythm with/without feedback: what is the difference?
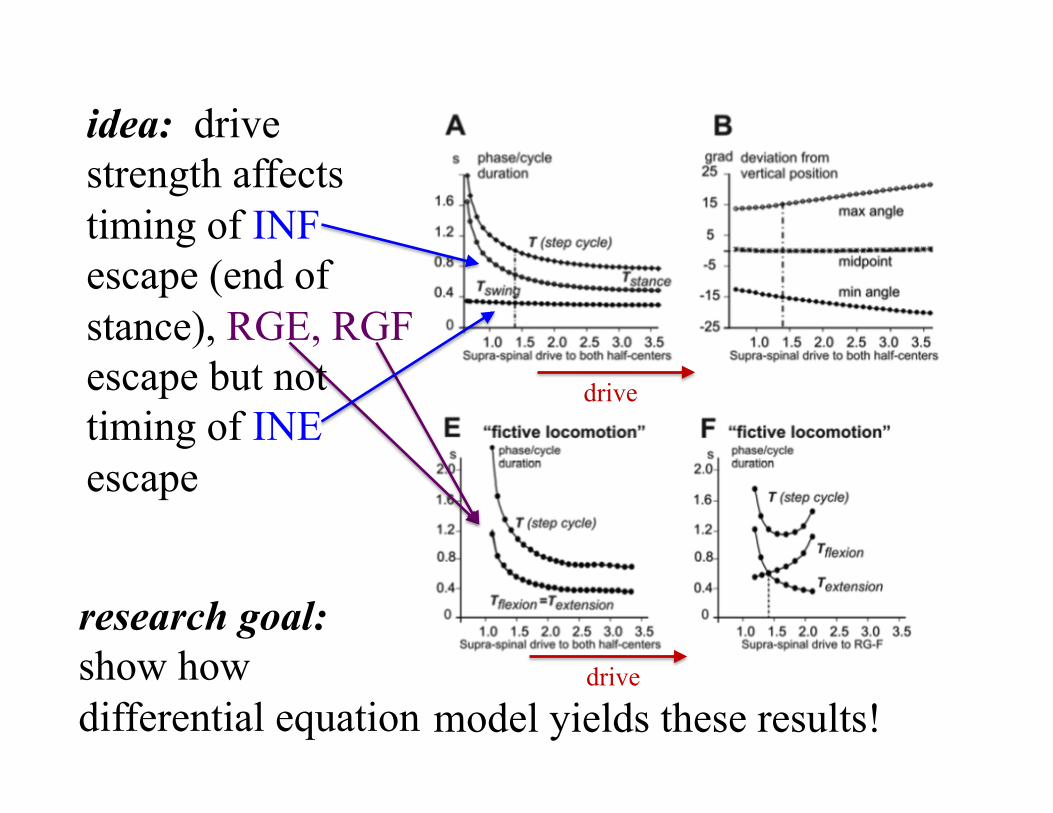
drive
drive
idea: drive strength affects timing of INF escape (end of stance), RGE, RGF escape but not timing of INE escape
research goal: show how differential equation model yields these results!

1. some neurons/networks exhibit dynamically rich behavior called bursting
2. single cell bursting can be studied via fast/slow decomposition and bifurcation analysis
3. a classification of bursting in single cells exists but is arguably not complete (noise, multibif, MMOs)
4. bursting in networks can be studied in various ways 5. synaptic excitation promotes bursting by
desynchronizing spikes: slow averaged dynamics 6. responses to feedback/drive depend on transition
mechanisms within bursting rhythms 7. remains to determine key ingredients for synchronized
bursting in heterogeneous networks
summary
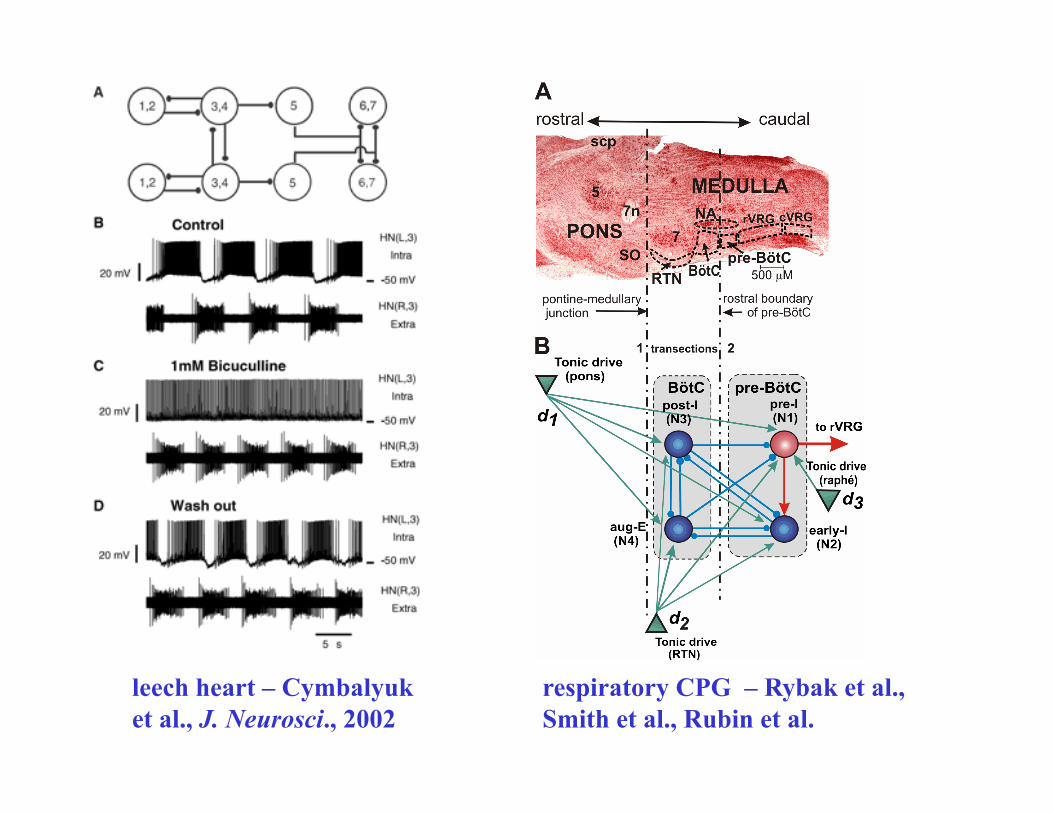
leech heart – Cymbalyuk et al., J. Neurosci., 2002
respiratory CPG – Rybak et al., Smith et al., Rubin et al.
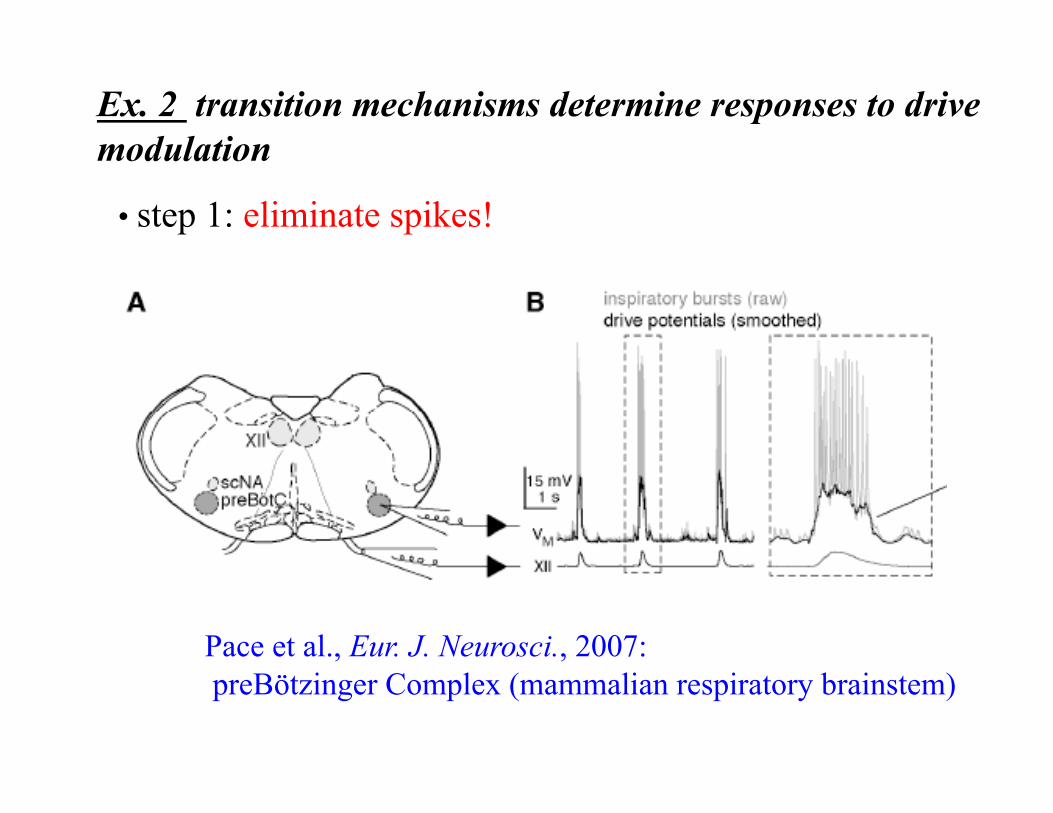
Ex. 2 transition mechanisms determine responses to drive modulation
• step 1: eliminate spikes!
Pace et al., Eur. J. Neurosci., 2007: preBötzinger Complex (mammalian respiratory brainstem)

asymmetric drive results (cont.)
highly tunable
high phase independence
robust

![The bubble bursting acoustic signature · 2 Introduction Vidal et al.[1] [1] V. Vidal et al., Dynamics of soap bubble bursting and its implications to volcano acoustics, Geophys.Res](https://img.pdfslide.us/doc/110x75/5e4503e1988bf560fd13a4f2/the-bubble-bursting-acoustic-2-introduction-vidal-et-al1-1-v-vidal-et-al.jpg)



























