Embed Size (px)
Citation preview

Cell, Vol. 59, 751-765, November 17, 1989, Copyright 0 1969 by Cell Press
The Drosophila patched Gene Encodes a Putative Membrane Protein Required for Segmental Patterning
Joan E. Hooper and Matthew I? Scott Howard Hughes Medical Institute and Department of Molecular, Cellular
and Developmental Biology University of Colorado Boulder, Colorado 80309-0347
Summary
The patched (ptc) gene is one of several segment polarity genes required for correct patterning within every segment of Drosophila. The absence of pfc gene function causes a transformation of the fate of cells in the middle part of each segment so that they form pattern elements characteristic of cells posi- tioned around the segment border. Analysis of the mu- tant phenotype demonstrates that both segment and parasegment borders are included in the duplicated pattern of pfc mutants. We have cloned the pfc gene and deduced that the product is a 1288 amino acid pro- tein with at least seven putative transmembrane a helices. pfc RNA is expressed in embryos in broad stripes of segmental periodicity that later split into two stripes per segment primordium. The pattern of expression does not directly predict the transforma- tion seen in pfc mutant embryos, suggesting that pfc participates in cell interactions that establish pattern within the segment.
Introduction
Early in development, the Drosophila embryo is divided along the anterior-posterior axis into a series of homolo- gous segments. Many genes required for this segmentation process have been identified by systematic mutagenesis (Niisslein-Volhard and Wieschaus, 1980; Niisslein-Volhard et al., 1984; Jiirgens et al., 1984; Weischaus et al., 1984; Schiipbach and Wieschaus, 1986; F’errimon et al., 1989). These genes fall into four broad groups: those that affect the anterior-posterior polarity of the whole embryo (re- viewed in Niisslein-Volhard et al., 1987), those that affect several contiguous segments (gap genes), those that af- fect alternate segments (pair-rule genes), and those that affect every segment (segment polarity genes) (reviewed in Akam, 1987; Scott and Carroll, 1987; Ingham, 1988). These four classes of genes act during oogenesis and embryogenesis to subdivide the embryo first into broad domains, then into smaller domains, and finally into seg- mental metameres.
Genes of the segment polarity class are required for proper development of the iterated segmental pattern. Mutations in segment polarity genes result in the deletion of a homologous part of the segmental pattern in every segment, often accompanied by mirror-image duplica- tion of the remaining pattern (NOsslein-Volhard and Wies- chaus, 1980). On the ventral surface of a mature embryo,
the segmental pattern consists of rows of fine bristles (denticles) in the anterior part of each segment and naked cuticle in the posterior part of each segment (Figure 1). Among the segment polarity mutations are those that de- lete ventral naked cuticle, that is, posterior pattern ele- ments, and replace it with denticles. This group of genes includes armadillo (arm), Cell, cubitis interruptus, dishev- elled, fused, gooseberry (gsb), hedgehog (hh), porcupine, smooth, and wingless (wg) (Niisslein-Volhard and Wies- chaus, 1980; Niisslein-Volhard et al., 1984; Jiirgens et al., 1984; Perrimon and Mahowald, 1987; Orenic et al., 1988; Perrimon et al., 1989). Mutations in a second type of seg- ment polarity gene delete ventral denticles, that is, an- terior pattern elements, and replace them with naked cuti- cle. In this class are naked(nkd) and zestewhite3 (Jiirgens et al., 1984; Perrimon et al., 1989). Mutations in genes of a third class result in the deletion of central pattern ele- ments and their replacement by segment border pattern elements. patched @tc) and costal- are known to be in this class (Niisslein-Volhard and Wieschaus, 1980; Grau and Simpson, 1967). engrailed (en; Niisslein-Volhard and Wieschaus, 1980; Kornberg, 1981) and lines (Ntisslein- Volhard et al., 1984) may be members of a fourth class, in which mutations cause the absence of the anterior part of each denticle band, including the segment border.
Four segment polarity genes have been cloned and their products identified. The en gene is required for the proper formation of segment borders (Kornberg, 1981) and is expressed in the posterior part of each segment primor- dium (Fjose et al., 1985; Kornberg et al., 1985; DiNardo et al., 1985; lngham et al., 1985b). en contains a homeobox and is therefore likely to encode a transcription factor (reviewed in Scott et al., 1989). gsb, wg, and arm are all required for correct patterning of the posterior half of the segment. wg, which encodes a putative cell surface or secreted protein (Rijsewijk et al., 1987), is expressed in the cells immediately anterior to the cells that contain en prod- ucts (Baker, 1987). gsb-distal, another homeobox-con- taining gene, is expressed in the posterior half of the seg- ment primordia, overlapping the expression of en and of wg (Baumgartner et al., 1987). arm, which encodes a putative cytoskeleton-associated protein, is expressed uniformly in the embryo (Riggleman et al., 1989).
Parasegmental grooves form at the junction between en-and wg-expressing cells while segment borders form at the edge of en-expressing cells, where wg is not ex- pressed (Martinez-Arias and Lawrence, 1985; see Figure 10). The same relationship between en and wgexpression and the morphology of the segment is also seen in ptc and nkd mutants (Martinez-Arias et al., 1988). ptc mutant em- bryos have ectopic segment borders and parasegment borders in every segment that are presaged by an ectopic stripe of en-expressing cells just anterior to the wg- expressing cells. This produces an ectopic junction be- tween en- and wg-expressing cells and also an ectopic edge of en-expressing cells where wg is not expressed. The nkd phenotype is similarly predicted by ectopic en

Cdl 752
ventral dorsal wt Ptc
Figure 1. The Patched Embryonic Cuticle Phenotype
wt = wild-type embryos. ptc = embryos lacking all ptc function (Df(2R)44CE/Df(2R)P14TE). Anterior is up. Ventral views are of the fifth abdominal segment; dorsal views are of the third thoracic segment. Schematic diagrams of these cuticle patterns are shown below the photomicrographs; solid lines represent segment borders (s) and dashed lines represent parasegment borders (ps). Dorsal sensory hairs are indicated by heavy curved lines. The territory affected by ptc is indicated on the wild-type schematic as a shaded box. Large, shaded arrows (labeled A-D) beside the schematics are a further abstraction of the segmental pattern. (A) Pattern elements near the segment border; (D) pattern elements near the parasegment border; (B and C) remaining territories and patterns. Directions of the arrows indicate polarities of the patterns.
In wild-type embryos the ventral surface of abdominal segments consists of seven rows of denticles (arrows A and 8). Each row is made up of denticles with stereotyped size and polarity. The posterior half of the segment has naked cuticle (arrows C and D). The ventral muscles, which attach between the first and second rows of denticles, define the segment border.
In wild-type embryos the dorsal surface of thoracic segments consists of naked cuticle at the segment border (arrow A), followed posteriorly by a row of small denticle-like hairs that point posteriorly (arrow B), then a region of fine hairs (arrow C), and finally naked cuticle (arrow D). In abdominal segments the posterior naked cuticle (arrow D) is replaced by larger denticle-like hairs pointing anteriorly. The dorsal muscles, which attach in the naked cuticle just anterior to the row of small denticle-like hairs (arrow A), identify segment borders. Small arrows on the photographs of the dorsal views point to the sensory hairs; white dots indicate the dorsal midline.
Dorsally, the middle half of the segment (arrows B and C) of ptc- embryos is transformed to express pattern elements that include duplicated segment and parasegment borders in reversed polarity (inverted arrows A and D). The small denticle-like hairs (arrow B) are replaced by naked cuticle in thoracic segments (arrow A) and by large denticle-like hairs in abdominal segments (data not shown). Posteriorly oriented fine hairs (arrow C) are replaced by disordered fine hairs (arrow D). In the seventh abdominal segment of wild-type embryos, virtually all the fine dorsal hairs point anteriorly (Campos-Ortega and Hartenstein, 1995). In pfc- embryos there is a plane of polarity reversal in the middle of the fine dorsal hairs; the posterior half of the fine dorsal hairs (arrow D) point anteriorly as in wild type, while the anterior half of the fine dorsal hairs point posteriorly. Therefore the plane of mirror-image symmetry that defines the posterior edge of the territory affected by ptc mutants is approximately in the middle of the fine hairs (between arrows C and D).
and wg expression. The intimate coupling of the expres- sion patterns of en and wg (and probably other, yet un- characterized segment polarity genes) to the final mor- phology of the segment in both wild-type and mutant development suggests that the expression patterns of these and other segment polarity genes indicate the cell identities that ultimately determine the segmental pattern.
Substantial cell communication is required to establish, maintain, or evolve the final pattern of segment polarity gene expression. For example, the en and wg genes are not expressed in the same cells, yet en function is required to maintain the expression of wg in anterior neighboring cells and wg function is required to maintain the expres- sion of en in posterior neighboring cells (Martinez-Arias et al., 1988; DiNardo et al., 1988). Cell communication is also revealed by clones of wg (Morata and Lawrence, 1977; Baker, 1988a), hh (Mohler, 1988), gsb (Wieschaus and Riggleman, 1987), and ptc (R. Phillips, I. Roberts, P Ing- ham, and R. Whittle, personal communication) cells when they are surrounded by wild-type cells. hh or ptc clones can confer a mutant phenotype on wild-type neighboring cells, while wild-type neighboring cells can rescue the mu- tant phenotype of wg or gsb clones.
Several features of the phenotype of null mutations in the ptc gene suggest that ptc plays an early and critical role in patterning the segment. The duplicated segment borders in ptc- embryos (Niisslein-Volhard and Wies- chaus, 1980; see Figure 1) are made at the expense of pat- tern elements from the middle of the segment. The pheno- type is caused by a transformation of cell fate early in embryogenesis rather than cell death and ensuing repair processes (Martinez-Arias et al., 1988). Early manifesta- tions of the transformation include ectopic parasegmental grooves and the ectopic expression of en and wg men- tioned above (Martinez-Arias et al., 1988). Early transfor- mations in the central nervous system mirror the dele- tion/duplication phenotype of the cuticle in ptc mutants (Pate1 et al., 1989). The transformations of cell fate in ptc mutant embryos suggest that either ptc mutations prevent correct signals from being sent early in development or they cause early misreading of positional cues.
In this paper we describe the molecular cloning of the ptc gene. Two surprising results follow from the initial characterization of the ptc gene. First, sequences of ge- nomic and cDNA clones suggest that, unlike most other segmentation genes, ptc encodes an integral membrane
patched Transmembrane Protein Controls Segment Pattern 753
protein with multiple membrane-spanning domains. Sec- ond, the pattern of expression of the prc RNA does not directly predict the defects seen in the pfc mutant em- bryos. ptc is therefore one of the few currently identified segmentation genes that does not encode a transcription factor but instead encodes a protein that is likely to medi- ate cell interactions directly.
Results
The patched Embryonic Phenotype The wild-type ventral cuticle pattern consists of denticles in the anterior third of each segment; the posterior two- thirds of each segment is naked cuticle. Dorsally the posterior three-fourths of the segment is covered with a pattern of setae, fine hairs, and large sensory hairs. Thus the ventral pattern is best for monitoring the fate of the an- terior part of the segment while the dorsal cuticle pattern allows analysis of the posterior part of the segment. Both dorsally and ventrally, the major muscle attachment sites define the segment border, so it is possible to align the dorsal and ventral patterns according to their homologous positions within the segment. The wild-type ventral and dorsal cuticle patterns are shown in Figure 1, where let- tered arrows illustrate the relationship between the dorsal and ventral cuticle patterns.
In ptc- embryos, cells in the middle half of each seg- ment express pattern elements appropriate to the margins of the segment (e.g., segment and parasegment borders; Figure 1). The polarity of these duplicated pattern ele- ments is reversed relative to the overall polarity of the em- bryo. The planes of mirror-image symmetry delineate that part of the segment that is affected by prc mutations. The ventral cuticle phenotype shows that the anterior limit of the territory defective in pfc- embryos is approximately at the level of the third row of the abdominal denticle belt (be- tween arrows A and B, Figure 1). The dorsal cuticle pheno- type is consistent with this anterior limit for pfc require- ment and, in addition, defines the posterior limit of the territory defective in pfc mutants (between arrows C and D in Figure 1). Since this is well to the anterior of the parasegment border (arrow D), the dorsal cuticle pattern confirms that parasegment borders are duplicated in ptc mutants,
Polarity is a product of local cell interactions since the polarity of the pattern elements is linked to the identities of neighboring cells and not to the overall head-to-tail polarity of the embryo. This is clear ventrally where denti- cles in the most anterior row of an abdominal denticle belt point away from the nearest segment border and toward the nearest parasegment border, whether the denticle row is in its normal position in wild-type embryos or is ectopic, with reversed orientation relative to the head and tail, in ptc mutants.
The cuticular sensory structures of wild-type embryos include dorsal, lateral, and ventral hairs in the abdomen, and Keilin’s organs, ventral and lateral black dots, lateral hairs, and two dorsal hairs in the thorax (Campos-Ortega and Hartenstein, 1985). A subset of these structures are affected in ptc- embryos. Thoracic segments are missing
- centromere
QLCPl Q536 k&mere -
Q2304 -
Q Df(2R)SA14 R+l 1 : I 1 1 T(2;3)eve2.26 D
I Df(2R)193Au : i
Df(2R)GR15 L2 0: ; w Df(2R)44CE
w Df(SR)R+l TE
1 Df(2R)GRlti TE
- Df(2R)P14 TE
’ ptc’ region
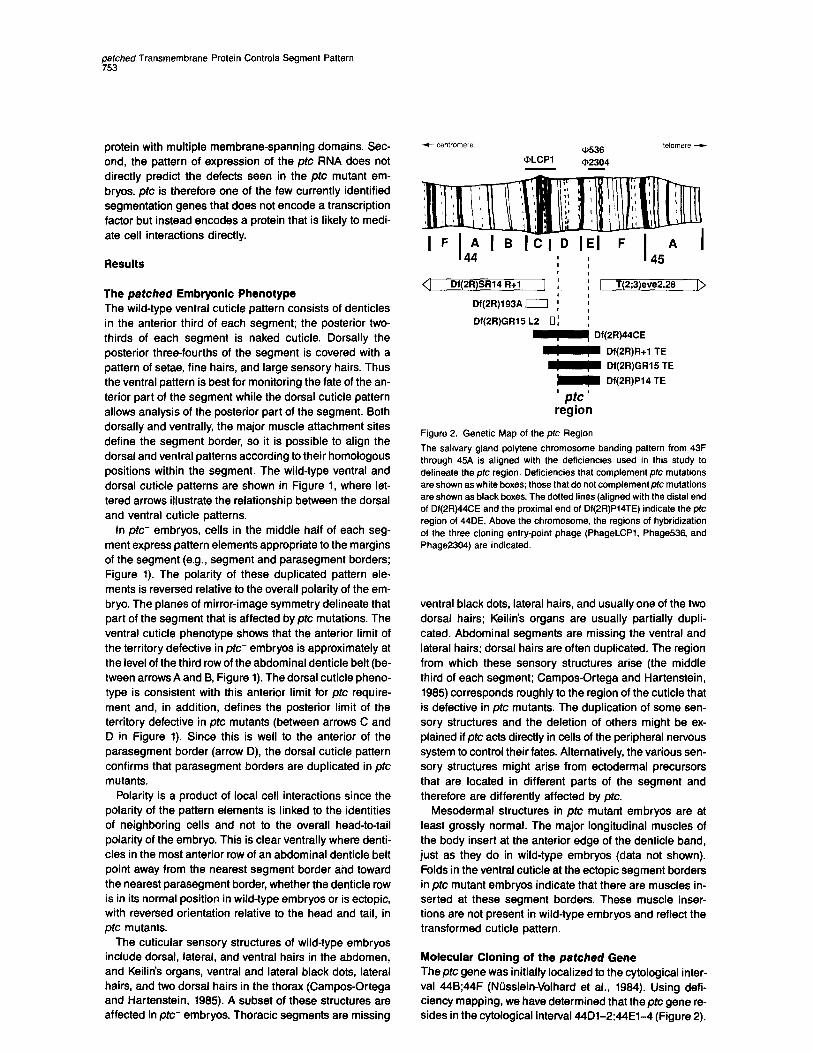
Figure 2. Genetic Map of the plc Region
The salivary gland polytene chromosome banding pattern from 43F through 45A is aligned with the deficiencies used in this study to delineate the ptc region. Deficiencies that complement ptc mutations are shown as white boxes; those that do not complementptc mutations are shown as black boxes. The dotted lines (aligned with the distal end of Df(2R)44CE and the proximal end of Df(2R)P14TE) indicate the ptc region of 44DE. Above the chromosome, the regions of hybridization of the three cloning entry-point phage (PhageLCPl. Phage536, and Phage2304) are indicated.
ventral black dots, lateral hairs, and usually one of the two dorsal hairs; Keilin’s organs are usually partially dupli- cated. Abdominal segments are missing the ventral and lateral hairs; dorsal hairs are often duplicated. The region from which these sensory structures arise (the middle third of each segment; Campos-Ortega and Hartenstein, 1985) corresponds roughly to the region of the cuticle that is defective in ptc mutants. The duplication of some sen- sory structures and the deletion of others might be ex- plained if ptc acts directly in cells of the peripheral nervous system to control their fates. Alternatively, the various sen- sory structures might arise from ectodermal precursors that are located in different parts of the segment and therefore are differently affected by pm.
Mesodermal structures in prc mutant embryos are at least grossly normal. The major longitudinal muscles of the body insert at the anterior edge of the denticle band, just as they do in wild-type embryos (data not shown). Folds in the ventral cuticle at the ectopic segment borders in ptc mutant embryos indicate that there are muscles in- serted at these segment borders. These muscle inser- tions are not present in wild-type embryos and reflect the transformed cuticle pattern.
Molecular Cloning of the patched Gene Thepfc gene was initially localized to the cytological inter- val 448;44F (Niisslein-Volhard et al., 1984). Using defi- ciency mapping, we have determined that the@ gene re- sides in the cytological interval 44Dl-2;44El-4 (Figure 2).

Cell 754
+195 185 175 185 155 145 135 125 115 105
transcription _1.7 6.3 2.4 units -1.2
> I<
cDNAs
exons
Figure 3. Molecular Map of the ptc Region
The most proximal 95 kb of the chromosome walk around ptc (+105- +195) is shown. Thirteen cosmid and two phage clones covering the
ptc region are aligned with the BamHl (B) and Bglll (Bg) restriction sites. The deficiencies that delineate the prc region, Df(2R)44CE and Df(2R)P14TE, are described in Figure 2 and their mapped endpoints are indicated. Df(2R)44CE begins in a 4.8 kb Hindlll-Notl restriction fragment between +108 and +lll (dashed line) and extends prox- imally beyond this chromosome walk (heavy arrow). Df- (2R)P14TE be- gins in a 4.7 kb BamHI-EcoRI restriction fragment between +188 and +173 (dashed line) and extends distally for about 100 kb (heavy arrow).
The approximate molecular positions of ptc1N’Q and pew, the le- sions used to identify the ptc transcription unit, are indicated. ptcRXm is localized in the 1.8 kb BamHl fragment around +148. prclNIOB maps within a region as small as +144-+173 (stippled line) or as large as +118-+178 (dashed lines; see Figure 4 for details). The transcription units in this region that are active at 4-8 hr are aligned beneath the pfc lesions. The size in kilobases of the detected transcripts based on RNA size markers (Boehringer Mannheim) is indicated. Arrows indi- cate the direction of transcription for those transcription units where it was determined. The 1.2 kb transcript is likely to be the Jonah tran- script (Carlson and Hogness, 1985). based on its genomic localization. Transcripts 4.0 and 4.O’may be the same or different transcription units, both with mature 4.0 kb products.
The three largest cDNA clones isolated, which together cover 5.3 kb of the ptc transcript, are shown. J20 is polyadenylated. The six exons of the ptc gene (A-F) are indicated, as well as the polyadenylation site at the 3’ end of exon F. The 5’ end of the gene has not been mapped, and therefore it is possible that another exon lies 5’of exon A. The large ORF is indicated as the solid boxes, beginning at an AUG codon in exon A and ending at an UAG codon in exon F. The 3’ untranslated re- gion is indicated by the dashed box in exon F; the putative 5’ untrans- lated region is indicated by the dashed box in exon A. Restriction en- donuclease sites within the ptc gene: Hindlll (H), Bglll (Bg), EcoRl (R), BamHl (B), Xbal (X), Sall (S), Apal (A), Kpnl (K), Notl (N). At least three additional EcoRl sites between +151.5 and +157.5 have not been pre- cisely mapped and are not shown.
Figure 4. Mapping of Polymorphisms Genetically Linked to the eNIOe Lesion
Polymorphisms are shown between the ptdN’OB (solid line) and ptc+ (CyO; stippled line) chromosomes in theptc region. Polymorphisms in- clude a 0.2 kb insertion at +185; H (Hindlll); E (EcoRI); A (Alul); R (Rsal); TTAAA (a pentanucleotide repeated three or five times within the third intron at +144; the sequence was found by sequencing the respective DNAs). Three independent revertants of ptcINIOB @t@+‘, pt@+‘, and pt~?‘+~) are indicated below with their associated polymor- phisms. The proximal crossover of pk@ must lie between the two Alul sites near +175, because pt@+’ has both the proximal Alul site characteristic of the ptclNqos chromosome and the slightly more distal Alul site characteristic of the ptc+ chromosome. The uncertainty in the location of the crossover is indicated by the overlap of the solid line and the stippled line. Similarly, the proximal crossovers for p&R+2 and pk? lie between the Alul site al +175 and the EcoRl site at +184. The distal crossovers for all three revertants lie between the TTAAA repeat at +144 and the EcoRl site at +118. The actual exchange event could be as small as 29 kb or as large as 58 kb for ptcR+‘, or 29-85 kb for pt@+’ and pf~~+~.
To locate the pfc gene relative to the deficiency break- points, complementation tests between ptc mutations and deficiencies that terminate within 448;44F were scored for the ptc phenotype. ptc function was found to require sequences between the centromere-proximal break of Df(2R)P14TE (44Dl-2; Gubb et al., 1988) and the centro- mere-distal break of Df(2R)44CE (44El-4). We shall refer to this genetically defined interval as the ptc region.
A chromosome walk that includes the ptc region is shown in Figure 3. The deficiency breakpoints that define the ptc region were approximately mapped onto the chro- mosome walk by hybridizing probes in situ to chromo- somes from larvae heterozygous for the appropriate defi- ciency. This located the Df(2R)P14TE proximal break between +159 and +199, and the Df(2R)44CE distal break between +112 and +127. Genomic DNA blots were used to pinpoint the breakpoints to specific genomic re- striction fragments: a 3.7 kb BamHI-Sall fragment (+188- +172) for Df(2R)P14TE proximal and a 3.0 kb Hindlll frag- ment (+lll-+114) for Df(2R)44CE distal (data not shown). The genetically defined ptc region, and therefore an es- sential part of the ptc gene, lies between coordinates +lll and +172.
One X-ray-induced ptc allele and 14 ethyl methanesul- fonate (EMS)-induced ptc alleles were screened for rear- rangements within the ptc region by genomic DNA blot- ting. No changes were detected in the DNA from the EMS-induced mutants. Novel restriction fragments were found between +145 and +148.4 in ptcRm, the X-ray-

perched Transmembrane Protein Controls Segment Pattern 755
induced allele. No other lesions were detected in the prc region of the pfcRm chromosome by Southern blotting or by in situ hybridization. Therefore the lesion in the pfc re- gion of the pfcR~ chromosome is limited to the 1.6 kb BamHl fragment. This lesion is probably responsible for the loss of pfc function associated with the pfcRXGI chro- mosome, and therefore is probably in or near thepfc gene.
The pfclNIOa lesion was mapped between +116 and +175 using linkage to the restriction site polymorphisms that distinguish the pfciNIOB chromosome from a pfc’ (CyO) chromosome. Three independent revertants of ,,~lNlOtJ were isolated after premeiotic gamma-ray muta- genesis of females heterozygous for the ptc’N1o* and the pfc+ (CyO) chromosomes. The revertants retained the flanking cinnibar (cn) and brown (bw) markers of the ptc’u’~ chromosome while recovering ptc+ function from the CyO chromosome. By meiotic recombination, the pfc+ function of all three revertants mapped to the pfc lo- cus f 2 CM, suggesting that mutant sequences at theptc locus of the pfrYN1m chromosome had been replaced by functional pfc+ sequences from the CyO chromosome. In each revertant the reversion to of& function was as- sociated with a conversion of sequences on the pfc’N1oB chromosome to those of the CyO chromosome some- where between +118 and +183 (Figure 4). Polymor- phisms characteristic of the pfc’NIOB chromosome at +116, +183, and +195 were retained in all three rever-
tams while polymorphisms characteristic of the CyO chro- mosome between +145 and +175 were picked up in all three revertants. The Alul site at +175 of the pfc’N1m chro- mosome was retained in pfcR+l, defining the maximal proximal extent of the ptc’N1oB lesion. The EcoRl site at +118 of the CyO chromosome, which is found in none of
the revertants, defines the maximal distal extent of the prctNIOB lesion. Thus the ptc’NIOB lesion, and therefore an essential part of the pfc gene, lies between +116 and +l75.
Identification of the pfc Transcription Unit Deficiency mapping, the location of pfcRXm, and the map- ping of ptc’NIOB using genetically linked polymorphisms indicate that at least part of theptc gene must be between +lll-+118 and +172-+175. To locate prc transcripts within this region, transcription units between +104 and +199 were roughly mapped. Polyadenylated embryonic RNA from 4-8 hr embryos (a stage when ptc function is required; Martinez-Arias et al., 1988) was screened by hybridizing RNA blots with genomic probes from across the region. Seven regions of genomic DNA between coor- dinates +104 and +183 were found to hybridize to tran- scripts that are detectable between 4 and 8 hr of em- bryonic development (Figure 3). Of these, five transcripts (1.2, 2.4, 4.0, 4.0’, and 6.3 kb) lay between the deficiency breakpoints defining the pfc region. The pfcRXU lesion maps within the transcription unit encoding the 6.3 kb transcript and at least 20 kb from the next closest tran- scription unit. We conclude that the 6.3 kb transcript be- tween +138 and +162 (Figure 3) is the best candidate for the product of the pfc gene, and we will refer to it as the ptc transcript. This assignment is in agreement with that
of l? lngham et al. (personal communication), who have used different alleles to map the prc gene.
The 6.3 kb mature pfc transcript is virtually undetectable in polyadenylated RNA isolated from preblastoderm (O-2 hr) embryos (data not shown) but is detectable in RNA from O-4 hr embryos (Figure 5). It is present in significant amounts in RNA isolated from extended germband em- bryos (4-8 hr) and slightly older embryos (8-12 hr) but falls to lower levels in late embryos, larvae, and pupae. This temporal pattern of expression is consistent with the requirements for pfc function during embryogenesis (the temperature-sensitive period for the prc cuticle phenotype is 4-12 hr; data not shown) and imaginal development (the temperature-sensitive period for pfc adult lethality ex- tends through the larval stages; data not shown). The other transcripts within theptc region (1.2,2.4,4.0, and 4.0 kb) do not peak at the extended germband stage and therefore do not have developmental profiles reflecting the prc temporal requirements.
Spatial Distribution of the ptc Transcript In situ hybridization to embryos was used to determine the spatial distribution of the ptc RNA during development (Figures 6 and 7). The prc RNA is not detectable in em- bryos before the thirteenth nuclear cleavage (for descrip- tions of developmental stages see Foe and Alberts, 1983; Campos-Ortega and Hartenstein, 1985). During cellulari- zation of the blastoderm (stage 5) prc RNA is detected throughout the cortical region of the embryo except for a dorsal anterior patch and a small region around the posterior pole including the pole cells (Figures 6A, 6D, 7A). prc RNA is also associated with the yolk nuclei at this stage. The anterior region that does not express prc corre- sponds to the primordium for the unsegmented acron or procephalic lobe, while the posterior region contains the germ cells and the anlagen of the posterior midgut and malphigian tubules (Jiirgens et al., 1986; Jiirgens, 1987).
During gastrulation and at least through mid-germband extension (early stage 8) ptc RNA remains homogene- ously distributed through most of the embryo (not shown). By the end of stage 8, all epidermal cells have divided and the homogeneous pattern of ptc transcript has been modulated into 15 stripes with segmental periodicity and into two other broad regions (Figures 6B, 6E, 78). The stripes correspond to part of each of parasegments O-14 within the ectoderm and mesoderm. The additional re- gions of ptc expression are the ectodermally derived hind- gut/analia posteriorly and the labrum in the dorsal head. The anteriormost of the parasegmental stripes lies an- terior to the cephalic furrow and posterior to the anterior midgut invagination. Segmentation of the mesoderm be- comes visible at the end of stage 6. The ptc stripes in the mesoderm are narrower than those in the ectoderm, oc- cupying the anterior one-third to one-half of each meta- mere (Figures 6B, 6E, 6J). Even at the earliest stage of segmental modulation of ptc transcription, the ectodermal and mesodermal stripes are not aligned.
During the extended germband stage there are two rounds of cell division in the ectoderm while neurogenesis begins (Campos-Crtega and Hartenstein, 1985). The pat-

Cell 756
tubulin -
Figure 5. Time Course of pfc RNA Accumulation
A blot of polyadenylated RNA (IO wg per lane) from embryos (O-4 hr, 4-6 hr, etc.), larvae (Ll; first larval instar, etc.), or pupae (EP [l-2 days after pupal formation] or LP 13-4 days after pupal formation]) of the in- dicated stages was probed with the 1.6 kb BamHl genomic DNA frag- ment derived from the region near +145. Below, the same blot was re- hybridized with a tubulin probe to control for even loading of mRNA.
tern of pfc transcripts in the embryo changes as each wide ectodermal stripe splits into two stripes (Figures 6C, 6F, 7C, 7D). The cells in the middle of the original stripe no longer express ptc RNA; the cells at the edges retain or increase their accumulation of ptc RNA. In the head, the involution of the foregut brings the labral region of ptc ex- pression to lie at the anterior end of the embryo (the clypeolabrum) where it splits into two regions. At least three additional regions of ptc transcript are found in the head at this stage in the vicinity of the stomodeum: one at the dorsal lip of the invagination (possibly in the interca- lary segment), one at the ventral lip of the invagination, and at least one in the roof of the stomodeum, which may correspond to the primordium of the stomatogastric ner- vous system. Posteriorly, ptc transcripts become restricted to a region around the incipient malphigian tubules.
The ptc expression pattern evolves differently in the different germ layers. Within the developing nervous sys- tem pfc transcripts are detected in some but not all of the
neuroblasts and their progeny that underlie the initial broad ectodermal stripe, but not in those that underlie the “interstripe” where en-expressing cells are found (Figures 6J and 6K). The ptc expression in the nervous system is consistent with the demonstrated requirement for ptc in the nervous system (Pate1 et al., 1969). In the mesoderm, some planes of section show rather diffuse labeling while other planes show labeling restricted to the anterior por- tion of the mesoderm of each metamere.
Following germband shortening the expression of ptc in the mesoderm and nervous system becomes more diffi- cult to detect. The epidermal stripes narrow to single-cell width (Figure 7C). Little ptc transcript is detectable in ma- ture embryos by this technique.
The position of ptc expression within each segment can be established relative to morphological landmarks and relative to the expression of other segment polarity genes. en is expressed in the anterior quarter of each paraseg- ment (Fjose et al., 1965; Kornberg et al., 1965; lngham et al., 1965b), and wg is expressed in the posterior quarter of each parasegment (Baker, 1967), when theptc RNA pat- tern is first modulated into stripes covering threequarters of each parasegment (Figures 6J-6L). The posterior edge of each ptc stripe corresponds to the wg-expressing cells, and the interstripes correspond to the cells expressing en. Parasegmental grooves appear slightly later at the pos- terior edge of the wide ptc stripes (Figures 6G and 6l-l). Mixed ptc and wg probes (Figures 7D-7F) illustrate that the wg stripe is coincident with the more anterior of the paired ptc stripes, that is, the stripe derived from the posterior edge of the early wide ptc stripe. Thus the pos- teriormost cells that express ptc are coincident with the wg-expressing cells and abut en-expressing cells.
The anterior limit of pfc expression is most easily estab- lished after germband shortening, when a scalloping of the ectoderm becomes apparent at the segment border (Figures 61 and 7C). The stripe that defines the posterior limit of ptc expression falls about two-thirds of the way back in each segment. The second stripe, which defines the anterior limit of ptc expression, lies at or just behind the segment border. Thus the cells expressing ptc in late extended germband embryos are the anteriormost CSIIS of every segment and the posteriormost cells of every para- segment (Figure 10).
cDNA and Transcript Structures and Sequence Three cDNAs that collectively account for 5.3 kb of the 6.3 kb matureptc transcript are shown in Figure 3. A compari- son of genomic and cDNA sequences indicated that the
Figure 6. The Pattern of ptc RNA Expression in Wild-Type Embryos
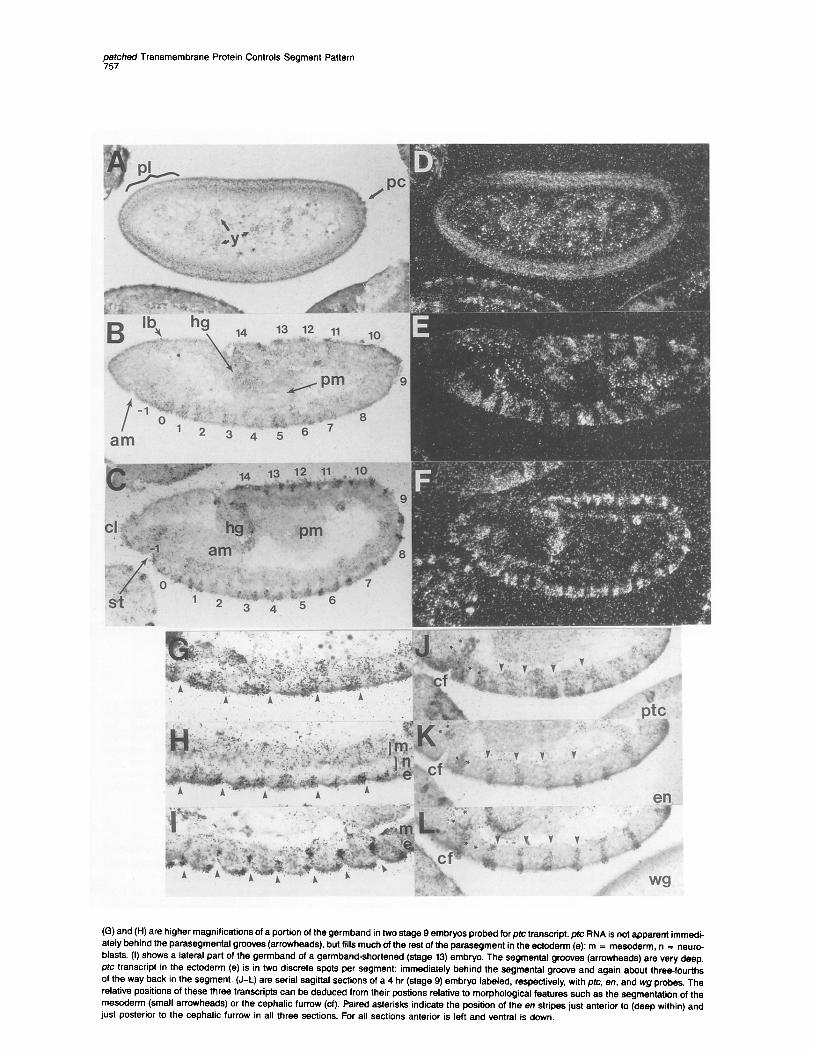
Midsagittal sections of embryos hybridized in situ with ptc antisense 3H-riboprobe. (A) and (D) are bright-field and dark-field photomicrographs, respectively, of a blastoderm stage embryo (about 2.5 hr; stage 5). Silver grains indicating ptc transcripts are localized throughout the cortical region except around the pole cells (PC) and the primordium for the procephalic lobe (pl). Silver grains are also localized over the yolk nuclei (y). (B) and (E) are bright-field and dark-field photomicrographs of an early extended germband embryo (about 3.5 hr; stage 6/9). The invaginations of the anterior midgut (am) and the posterior midgut (pm) are clearly seen. ptc transcripts are located in segment-periodicity stripes in parasegments -1 through 14 as well as in the primordium for the labrum (lb) and the hindgut (hg). (C)and(F) are bright-field and dark-field photomicrographs of a mid-extended germband embryo (about 5.5 hr; stage 11). ptc transcript is found in two stripes per parasegment (-1 through 14) as well as in two small regions of the clypeolabrum (cl), a region in the roof of the stomodeum (St), and a region at the junction of the hindgut (hg) and the posterior midgut (pm).
ghed Transmembrane Protein Controls Segment Pattern
(G) and (H) are higher magnifications of a portion of the germband in two stage 9 embryos probed for pfc transcript. ptc RNA is not apparent immedi- ately behind the parasegmental grooves (arrowheads), but fills much of the rest of the parasegment in the ectoderm (e); m = mesoderm, n = neuro- blasts. (I) shows a lateral part of the germband of a germband-shortened (stage 13) embryo. The segmental grooves (arrowheads) are very deep. ptc transcript in the ectoderm (e) is in two discrete spots per segment: immediately behind the segmental groove and again about three-fourths of the way back in the segment. (J-L) are serial sagittal sections of a 4 hr (stage 9) embryo labeled, respectively, with ptc, en, and wg probes, The relative positions of these three transcripts can be deduced from their postions relative to morphological features such as the segmentation of the mesoderm (small arrowheads) or the cephalic furrow (cf). Paired asterisks indicate the position of the en stripes just anterior to (deep within) and just posterior to the cephalic furrow in all three sections. For all sections anterior is left and ventral is down.
Cell 759
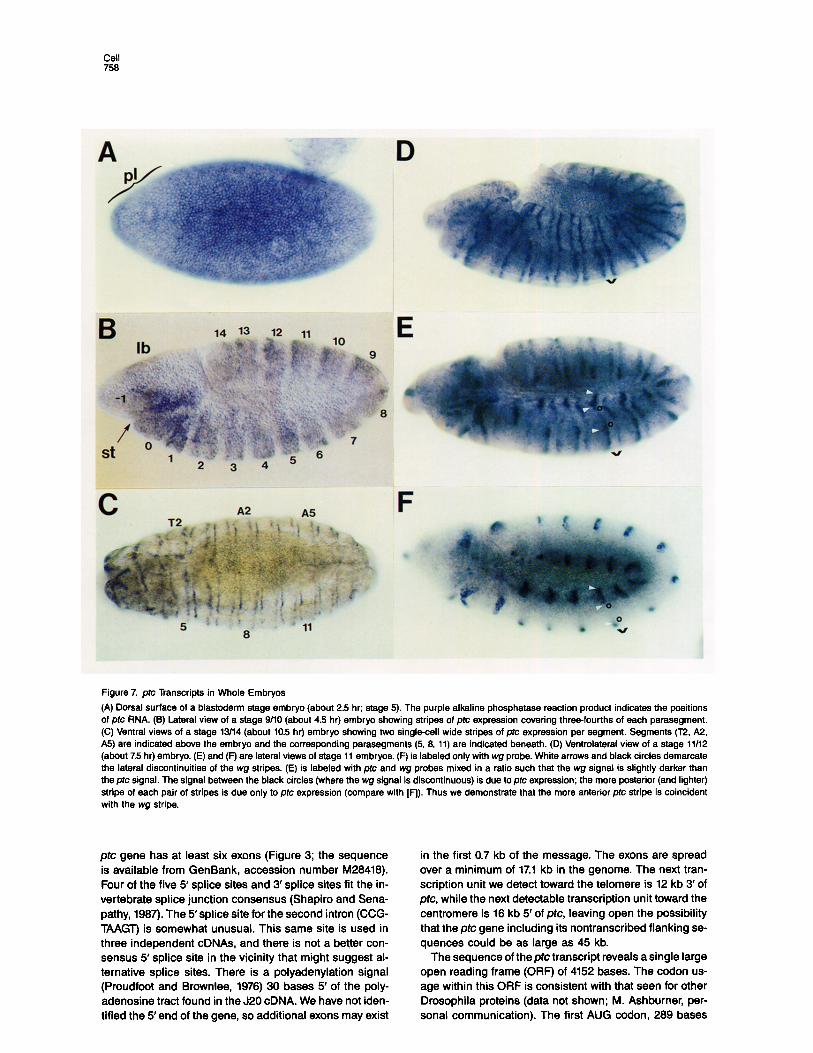
Figure 7. pfc Transcripts in Whole Embryos
(A) Dorsal surface of a blastoderm stage embryo (about 2.5 hr; stage 5). The purple alkaline phosphatase reaction product indicates the positions of ptc RNA. (B) Lateral view of a stage g/IO (about 4.5 hr) embryo showing stripes of ptc expression covering three-fourths of each parasegment. (C) Ventral views of a stage 13/14 (about 10.5 hr) embryo showing two single-cell wide stripes of ptc expression per segment. Begments (T2, A2, A5) are indicated above the embryo and the corresponding parasegments (5, 8, 11) are indicated beneath. (D) Ventrolateral view of a stage 11112 (about 75 hr) embryo. (E) and (F) are lateral views of stage 11 embryos. (F) is labeled only with wg probe. White arrows and black circles demarcate the lateral discontinuities of the wg stripes. (E) is labeled with ptc and wg probes mixed in a ratio such that the wg signal is slightly darker than the ptc signal. The signal between the black circles (where the wg signal is discontinuous) is due to ptc expression; the more posterior (and lighter) stripe of each pair of stripes is due only to ptc expression (compare with [F]). Thus we demonstrate that the more anterior ptc stripe is coincident with the wg stripe.
ptc gene has at least six exons (Figure 3; the sequence is available from GenBank, accession number M28418). Four of the five 5’ splice sites and 3’ splice sites fit the in- vertebrate splice junction consensus (Shapiro and Sena- pathy, 1987). The 5’splice site for the second intron (CCG- TAAGT) is somewhat unusual. This same site is used in three independent cDNAs, and there is not a better con- sensus 5’ splice site in the vicinity that might suggest al- ternative splice sites. There is a polyadenylation signal (Proudfoot and Brownlee, 1978) 30 bases 5’ of the poly- adenosine tract found in the J20 cDNA. We have not iden- tified the 5’end of the gene, so additional exons may exist
in the first 0.7 kb of the message. The exons are spread over a minimum of 17.1 kb in the genome. The next tran- scription unit we detect toward the telomere is 12 kb 3’ of ptc, while the next detectable transcription unit toward the centromere is 18 kb 5’ of ptc, leaving open the possibility that the ptc gene including its nontranscribed flanking se- quences could be as large as 45 kb.
The sequence of the ptc transcript reveals a single large open reading frame (ORF) of 4152 bases. The codon us- age within this ORF is consistent with that seen for other Drosophila proteins (data not shown; M. Ashburner, per- sonal communication). The first AUG codon, 289 bases
patched Transmembrane Protein Controls Segment Pattern 759
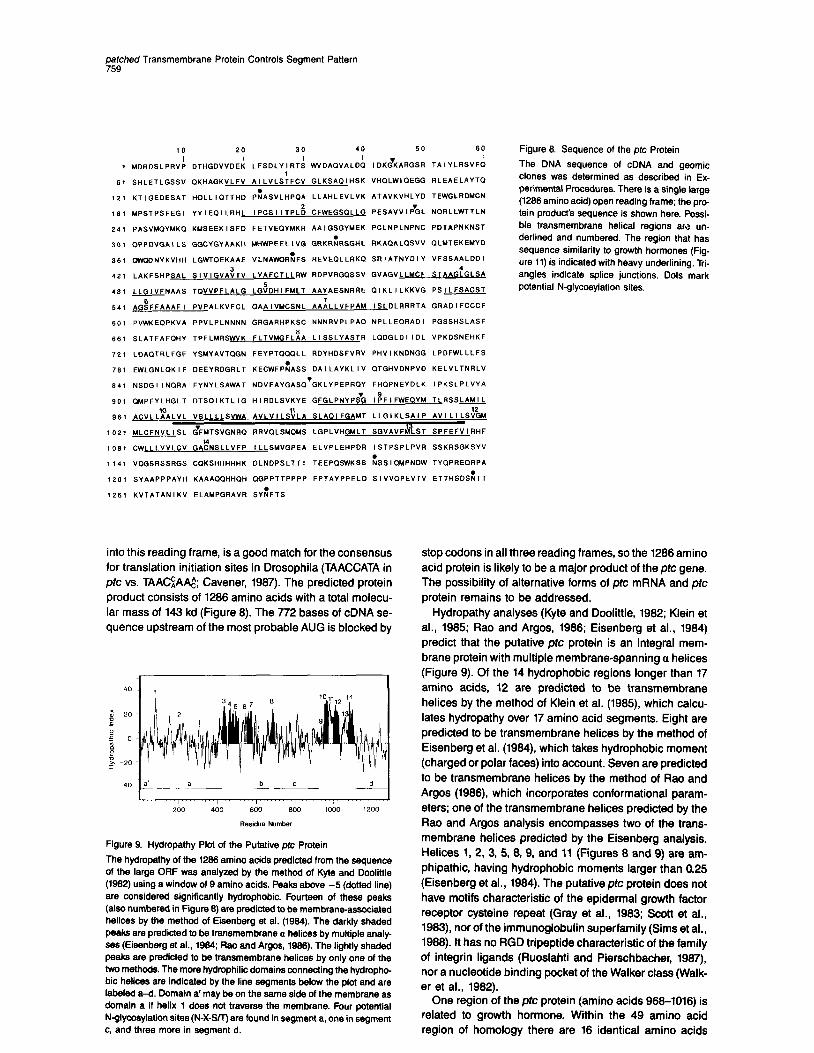
10 20 30 40 50 60
I I I I I I 1 MDRDSLPRVP DTHGDVVDEK LFSOLYIRTS WVOAOVALDO IDK&ARGSR TAIYLRSVFO
1 61 SHLETLGSSV OKHAGKVLFV AILVLSTFCV GLKSAOIHSK VHGLWIOEGG RLEAELAYTO
12, KTlGEDESAT HOLLIOTTHO PiASVLHPOA LLAHLEVLVK ATAVKVHLYD TEWGLRDMCN
,S, MPSTPSFEG, YYIEDILRHL ,PCS,,TPL; CFWEGSOLLG PESAVVIF%L NORLLWTTLN
241 PASVHOYMKO KMSEEKISFD FETVEOYMKR AAIGSGYMEK PCLNPLNPNC PDTAPNKNST
30, OPPDVGAILS GGCYGYAAKH MHWPEELLVG GRKRZRSGHL RKAGALGSVV QLMTEKEMYD
36, DWODNYKVHH LGWTOEKAAE “LNAWORAFS REVEOLLRKO SRIATNYDIY VFSSAALDDI
42, LAKFSHPSAL S,VlGVA:TV LYAFCTLLRW RDPVRGOSSV GVAGVLLMCF 4
S TAAGLGLSA
48, LLGlVFNAAS TOVVPFLALG LGtDHIFMLT AAYAESNRRE DTKLILKKVG PSILFSACST
54, AG:FFAAAF, PVPALKVFCL OAAIVMCSNL AA:LLVFPAM ISLDLRRRTA GRADIFCCCF
601 PVWXEOPKVA PPVLPLNNNN GRGARHPKSC NNNRVPLPAO NPLLEORADI PGSSHSLASF
661 SLATFAFCIHY TPFLMRSWVK FLTVMGFLBA LISSLYASTR LODGLDIIDL VPKDSNEHKF
72, LDAOTRLFGF YSMYAVTQGN FEYPTOQQLL RDYHDSFVRV PHVIKNONGG LPDFWLLLFS
78, EWLGNLDKIF DEEYRDGRLT KECWFPEASS DAILAYKLIV OTGHVDNPVD KELVLTNRLV
84, NSDGIINQRA FYNYLSAWAT NDVFAYGASO.GKLYPEPRQY FHOPNEYDLK IPKSLPLVYA
90, OMPFYLHGLT DTSOIKTLIG HIRDLSVKYE GFGLPNYP& l;FlFWEQYM TLRSSM
36, )&“LL:OALVL “SLLLLSVWA AVLV,L&LA SLAOIFGAMT LLGIKLSAIP A”lLILS&A
102, MLCFNVLISL ZFMTSVGNRQ RRVOLSMOUS LGPLVHGMLT SGVAVF i&T SPFEFVIRHF
,081 CWLLLVVLCV GAhLLVFP ,LLSMVGPEA ELVPLEHPDR ISTPSPLPVR SSKRSGKSYV
,141 “OGSRSSRGS COKSHHHHHK DLNDPSLTTI TEEPQSWKSS &.SIOMPNDW TYQPREORPA
,201 SYAAPPPAYH KAAAOOHHOH OGPPTTPPPP FPTAYPPELQ SIVVOPEVTV ETTHSDSiTT
,261 KVTATANIKV ELAUPGRAVR SYi?FTS
into this reading frame, is a good match for the consensus for translation initiation sites in Drosophila (TAACCATA in pfc vs. TAACgAA,A; Cavener, 1987). The predicted protein product consists of 1286 amino acids with a total molecu- lar mass of 143 kd (Figure 8). The 772 bases of cDNA se- quence upstream of the most probable AUG is blocked by
-c b A... ,,, ,,,,,,,, ,,,,,,,,, ,,,,,,,,, m.,,,,, ,,,,,,,,, . ..J
200 400 600 800 ,000 1200
Reshs Number
Figure 9. Hydropathy Plot of the Putative pfc Protein
The hydropathy of the 1266 amino acids predicted from the sequence of the large ORF was analyzed by the method of Kyte and Doolittle (1962) using a window of 9 amino acids. Peaks above -5 (dotted line) are considered significantly hydrophobic. Fourteen of these peaks (also numbered in Figure 6) are predicted to be membraneassociated helices by the method of Eisenberg et al. (1964). The darkly shaded peaks are predicted to be transmembrane a helixes by multiple analy- ses (Eisenberg et al., 1964; Rao and Argos, 1966). The lightly shaded peaks are predicted to be transmembrane helices by only one of the two methods. The more hydrophilic domains connecting the hydropho- bic helices are indicated by the line segments below the plot and are labeled a-d. Domain a’ may be on the same side of the membrane as domain a if helix 1 does not traverse the membrane. Four potential Nglycosytation sites (NXsrr) are found in segment a, one in segment c, and three more in segment d.
Figure 6. Sequence of the ptc Protein
The DNA sequence of cDNA and geomic clones was determined as described in Ex- perimental Procedures. There is a single large (1266 amino acid) open reading frame; the pro- tein product’s sequence is shown here. Possi- Me transmembrane helical regions am un- derlined and numbered. The region that has sequence similarity to growth hormones (Fig- ure 11) is indicated with heavy underlining. Tri- angles indicate splice junctions. Dots mark potential N-glycosylation sites.
stop codons in all three reading frames, so the 1286 amino acid protein is likely to be a major product of the ptc gene. The possibility of alternative forms of ptc mRNA and ptc protein remains to be addressed.
Hydropathy analyses (Kyte and Doolittle, 1982; Klein et al., 1985; Rao and Argos. 1986; Eisenberg et al., 1984) predict that the putative ptc protein is an integral mem- brane protein with multiple membrane-spanning a helices (Figure 9). Of the 14 hydrophobic regions longer than 17 amino acids, 12 are predicted to be transmembrane helices by the method of Klein et al. (1985) which calcu- lates hydropathy over 17 amino acid segments. Eight are predicted to be transmembrane helices by the method of Eisenberg et al. (1984), which takes hydrophobic moment (charged or polar faces) into account. Seven are predicted to be transmembrane helices by the method of Rao and Argos (1986), which incorporates conformational param- eters; one of the transmembrane helices predicted by the Rao and Argos analysis encompasses two of the trans- membrane helices predicted by the Eisenberg analysis. Helices 1, 2, 3, 5, 8, 9, and 11 (Figures 8 and 9) are am- phipathic, having hydrophobic moments larger than 0.25 (Eisenberg et al., 1984). The putative ptc protein does not have motifs characteristic of the epidermal growth factor receptor cysteine repeat (Gray et al., 1983; Scott et al., 1983) nor of the immunoglobulin superfamily (Sims et al., 1988). It has no RGD tripeptide characteristic of the family of integrin ligands (Ruoslahti and Pierschbacher, 1987) nor a nucleotide binding pocket of the Walker class (Walk- er et al., 1982).
One region of the ptc protein (amino acids 988-1016) is related to growth hormone. Within the 49 amino acid region of homology there are 16 identical amino acids

Cell 760
cuticle pattern
2.5h 3.5h 5.5h
and 9 conservatively substituted amino acids (Figure 11). Growth hormone is composed of four a-helical rods linked by short connecting regions (AbdeCMeguid et al., 1987). Helices 2 and 3 are amphipathic helices that form a struc- tural backbone for somatotropins and apparently do not carry specific ligand information (Cunningham et al., 1989). The region of growth hormone that is related to ptc includes all of helix 2, a sharp turn, and most of helix 3. Virtually all of the homologies withptc lie on the hydropho- bic faces of the helices, the faces of the helices that pack against each other. In ptc the homology includes the last 9 amino acids of putative helix 10, helix 11, and the first 9 amino acids of helix 12 (Figure 8). This region of ptc is highly hydrophobic (39149 amino acids), so the homology may reflect the hydrophobicity of the two regions. The ho- mology does not necessarily suggest that the ptc protein is a ligand or is released in a soluble form. It seems more likely to reflect conserved structural features such as helix packing.
iu'LU~iLiiSU~~flUiUlijULRSLflQlFGR~TL~Fl~~SfliPR~iLl:-l016
,“:y;” I* *llll* I I II * * *II ** II * II I LLRISLLLI~SULGPL~FLSRUFTNSLUFETSDRUVEKLKDLEEGlLflL-I23
Bouine GH LLRlSLLLlQSULGPLQFLUAUFTNSLUFGTSDRUVEKLKDLEEGlLflL Porcine GH LLRFSLLLIPSULGPUOFLSRUFTNSLUFGTSDRUVEKLKDLEEGlQflL Ruian GH LLRFSLULlQSULTPU~ULSKUFTNNLUFGTSDRUFEKLKDLEEFlQflL Horse GN LLRFSLLLIQSULGPUOLLSRUFTNSLUFGTSORUYEKLRDLEEGIORL Rat GH LLRFSLLLI~SULGPUQFLSRIFTNSLMFGTSDRUVEKLKDLEEGIORL Humm GN LLRlSLLLl~SULEPUOFLRSUFRNSLUVGflSDS~VDLLKDLEEGl~TL
N Monkey GH LLRISLLLIQSULEPUOFLRSUFRNSLUVGTSVS~VDLLKDLEEGIQRL
N It-H.llx 2 --WI Turn Id-Hallx 3 -
Figure 11. Similarity between the ptc Protein and a Region of Soma- totropins
A 49 amino acid region of homology between the ptc protein and growth hormone (GH) sequences is shown. Hormone sequences are from Abdel-Meguid et al. (1967) and references therein. The numbers refer to the amino acid positions in the ptc protein and ovine growth hormones. The human and monkey sequences have an additional asparagine, as shown. Vertical bars indicate amino acid identities be- tween prc and the ovine growth hormones (W49). and asterisks indi- cate conservative substiWons (9/49). The positions of the growth hor- mone sequences that comprise two of the four a-helical regions in the crystallized protein are indicated. The two helices have a sharp turn between them. Dots indicate the positions of amino acids that form all of one largely hydrophobic face of each helix in growth hormones; many of the conserved residues lie on these faces. Conserved in ptc are lO/ll residues that form the hydrophobic face on helix 2, and 617 of those in helix 3.
Figure 10. Summary of the Ectodermal Pat- tern of ptc Transcript within the Segment and Relative to the Cuticle Pattern
A two-segment repeat is shown for blastoderm stage (2.5 hr), early extended germband (3.5 hr), mid-extended germband (5.5 hr), and ma- ture embryo cuticle pattern (ventral abdominal segments). The transcript patterns for ptc (solid shading), en (hatched shading), and wg (light shading) at each stage are indicated rela- tive lo the segment borders (solid lines labeled ‘V), parasegment borders (dashed lines la- beled “ps”), and the cells (circles and ovals). The cuticle pattern is explained in Figure 1.
Discussion
ptc Expression in the Ectoderm pfc is initially expressed homogeneously across the em- bryo except for primordia of the unsegmented acron and telson regions. This homogeneous expression is main- tained while en, wg, and gsb transcripts appear in seg- ment-periodicity stripes on the blastoderm. The homo- geneous pattern of ptc expression is rather abruptly modulated into segment-periodicity stripes over a period of about 15 min at 3:15-3:30 hr, as the germband extends, tit about the time of the first postblastoderm mitosis in the ventral epidermis. The stripes appear as transcript levels drop in the interstripes, judging by the density of silver grains (compare Figures 8A and 8B). A half-life of less than 15 min for ptc RNA, as suggested by the rapid evolu- tion of the segmental ptc stripes, is consistent with rapid turnover for the mRNAs of some other segmentation genes (e.g., 8 min for fushi rafazu mRNA; Edgar et al., 1988). In the ectoderm, ptc RNA disappears from the posterior of each segment primordium, at precisely the time and place where en protein first accumulates. The en protein is capable of blocking transcriptional activation in cultured cells (Jaynes and O’Farrell, 1988), so the first change in the pfc RNA pattern could be a direct negative effect of the en protein on pfc transcription. The continuing presence of ptc RNA in the posterior region of segments in embryos that lack en function (unpublished data) sup- ports this hypothesis.
ptc Expression in the Mesoderm ptc expression in the mesoderm evolves differently from the pattern in the ectoderm. ptc is expressed in an anterior group of mesodermal cells in every segment from the ini- tial segmentation of the mesoderm (late stage 8) through germband shortening (stage 13). ptc expression in the mesoderm does not have the same relationship with en that it does in the ectoderm (Figures 8J and 8K). The mesodermal stripes of ptc are shifted one-quarter to one- third of a segment anterior relative to the ectodermal stripes. This cannot be due to a mesodermal “shift” (Mar- tinez-Arias and Lawrence, 1985) because wg and en in the mesoderm are still aligned with their ectodermal stripes. Therefore prc and en may be expressed in some of the same cells in the mesoderm, in contrast to their mutually

petched Transmembrane Protein Controls Segment Pattern 761
exclusive relationship in the ectoderm. Furthermore, the mesodermal stripes of ptc are unaffected in en mutants (unpublished data), suggesting that@ is differently regu- lated in the two germ layers.
ptc is the only segmentation gene that has yet been found to be expressed in the mesoderm in more than a transient way. For example, the RNAs encoded by en, wg, and gsb disappear from the mesoderm soon after gastru- lation (Fjose et al., 1985; Kornberg et al., 1985; Baker, 1987; Bopp et al., 1988; C&6 et al., 1987), suggesting a limited role for these genes in mesodermal development. In fact, en function is not required for mesodermal devel- opment (Lawrence and Johnston, 1984). In ptc mutants, mesodermal segmentation apparently proceeds normally (unpublished data), but it remains to be determined whether more subtle mesodermal processes, such as the proper formation of the visceral mesoderm and precise muscle patterns, require ptc function.
The Role of pfc in Patterning the Ectoderm At no stage does the evolving pattern of expression of ptc RNA predict the cuticle defects of ptc mutants (Figure 10). The anteriormost cells of each segment, which express ptc at both 3.5 and 5.5 hr of development, are not affected by loss of ptc function, yet cells in the middle of the seg- ment, which do not express ptc at 5.5 hr, are severely af- fected by loss of ptc function. These cells are apparently sensitive to the lack of ptc expression in neighboring cells. This suggests that the ptc protein is involved in sending or receiving signals within the segment primordium.
The role of ptc in normal embryogenesis, broadly de- fined, is to organize the pattern within the anterior three- fourths of the segment (Figures 1 and 10). This region cor- responds to the broad initial stripes of ptc expression. In ptc- embryos the cuticle defects, including the ectopic segment and parasegment borders, are limited to this an- terior part of the segment, as are the ectopic stripes of en and wg expression (Martinez-Arias et al., 1988; DiNardo et al., 1988). The ectopic expression patterns of en and wg indicate the changing cell identities that ultimately result in the ptc cuticle phenotype. These early changes of cell identity that result from lack of ptc function can be inter- preted as the result of inappropriate positional informa- tion.
Mosaic analysis also suggests that the ptc protein is in- volved in signaling between neighboring cells (R. Phillips, I. Roberts, t? Ingham, and R. Whittle, personal communi- cation). Small clones of ptc- cells are not phenotypically rescued by surrounding wild-type cells, but small clones of homozygous mutant cells induced in the midst of ptc+ cells can bestow mutant phenotype on their wild-type neighbors. The phenotypic autonomy of clones of ptc- cells suggests that the ptc gene product does not travel between cells; the ability of ptc- cells to affect their neighbors suggests that a lack of ptc function generates anomalous signals or a lack of normal signals, leading to the mutant morphology of neighboring cells. One possibil- ity is that ptc is a receptor whose activation changes the character of cells in which it is expressed. A manifestation of this changed character is new signals to neighboring
cells. The absence of ptc function then has two conse- quences: inappropriate states for the mutant cells and in- appropriate signals to neighboring cells, ultimately result- ing in inappropriate states for those neighboring cells. The ptc protein is almost certainly an integral membrane pro- tein with multiple membrane-spanning domains. Such a structure is consistent with the ptc gene product function- ing at the cell surface to respond to a ligand and transmit a signal across the cell membrane.
In ptc- embryos, the presence of ectopic segment and parasegment borders reduces the number of cells that do not express en, so it is difficult to discern other roles for
ptc in patterning these anterior cells. In embryos homozy- gous for the weak allele ptcullz, ectopic segment and parasegment borders do not form (unpublished data). Nevertheless, the denticles (the pattern elements in the anterior part of the segment) are disordered, demonstrat- ing a second, more subtle requirement for ptc in pattern- ing the anterior part of each segment. The late stripes of ptc expression define the margins of this territory (of non- en-expressing cells) and are thus positioned to transmit or relay a signal(s) that organizes the anterior part of the segment.
How Segment Polarity Genes Establish the Segmental Pattern Of the other four segment polarity genes that have been cloned and characterized, in only one case does the do- main of gene expression predict the subsegmental region that is defective in mutants. gsb transcripts are normally expressed in the posterior half of every segment (Bopp et al., 1988; Cat6 et al., 1987), and this is the region that is defective in gsb mutants. In contrast, wg affects more cells than those in which it is expressed (Morata and Lawrence, 1977; Baker, 1987), as does en (Kornberg, 1981; Kornberg et al., 1985; Fjose et al., 1985; DiNardo et al., 1985). arm is not required for patterning the anterior cells of each seg- ment, although it is expressed there (Wieschaus and Rig- gleman, 1987; Riggleman et al., 1989). Clearly, many seg- ment polarity genes do not act as genetic switches in the sense of homeotic genes or of most of the other segmen- tation genes.
Segment polarity gene products participate in a network of local cell interactions that lead to the patterning of seg- ments. Pattern formation within the segment or imaginal disc evolves as cells monitor and respond to the identities of their neighbors. If cells are apposed to inappropriate neighbors through damage or mutation, adjustments take place that reestablish appropriate neighbor relationships. This has been extensively documented for abdominal cu- ticle (Locke, 1959; Lawrence, 1988; Wright and Lawrence, 1981), imaginal discs (Bryant, 1975), and thoracic append- ages (French, 1978; Bryant et al., 1981) and is likely to be true for embryonic pattern formation as well. The instabil- ity of en expression in the absence of wg (DiNardo et al., 1988; Martinez-Arias et al., 1988), the instability of wg ex- pression in the absence of en, and the spreading of en in the absence of ptc or n/d (Martinez-Arias et al., 1988) are manifestations of these adjustments. The polarity rever- sals of the duplicated pattern elements in segment polar-

ity mutants are another manifestation of these neighborly relationships. An understanding of segment polarity gene function at the molecular level will reveal how cell interac- tions establish position and pattern within the segment.
Experimental Pmcedures
Drosophila Stocks, Mutagenesis, and Cmases b-7, an isogenized y; cn bw sp line, was obtained from Dr. J. Kennison (National Institutes of Health). Thirteen EMS-induced pc alleles and T(2;3)e~e~.~ (Niissiein-blhard and Wieschaus, 1980; Niissiein- Voihard et al., 1984) were obtained from the T8bigen stock center. PtcR”, recovered after X-ray mutagenesis, was a gift from Dr. P Simpson (Strasbourg). Df(2R)44CE was a gift from Dr. G. Yannopoulos (Patras, Greece). Df(2R)193A is a cytologically invisible lesion that we recovered as a white- revertant after gamma-ray mutagenesis (4000 R) of males carrying the TE78 chromosome, which has a white+ insert in 44C (ising and Block, 1981). it was classified as a deficiency based on its lack of complementation with the two nonoverlapping deficien- cies Df(2R)SR14LR+lR and Df(2R)GR15LTER. Autosynaptic stocks LS(2)GRl5//DS(2)GRl5. LS(2)ScoR+‘//DS(2)ScoR+‘, LS(2)ScoR+‘//DS- (2)TE1L146R, and LS(2)P14//DS(2)ScoR+r were generously provided by Dr. D. Gubb (Cambridge). We recovered DS(2)l2 over LS(2)GR15 after X-ray mutagenesis (Gubb et al., 1988). Df(2R)SR14LR+1R, Df- (2R)GR15LL2R, Df(2R)R+lLTER, and Df(2R)P14LTER are synthetic deficiencies that were made by recombination between autosynaptic chromosomes with slightly offset breakpoints. This generated pericen- tric inversions with one break in 2L (Gubb et al., 1988) and the other break in the ptc region. inversions deficient in 2R were identified by lethality over Df(2R)44CE and Df(2R)193A. Df(2R)44CE, Df(PR)SR- 14LR+1R, and T(2;3)e~e*.~ are cytologically visible. The other defi- ciencies were ordered by complementation of lethality and by com- plementation of the ptc phenotype of ptc’NIOB and ptc2F8”. Other marker mutants and chromosomes are described in Lindsley and Grell (1966).
Revertants of ptclNIOB were isolated as follows. cn ~tc’~‘~ bw spl CyC virgin females received 4500 rad from @%o and were mated to sfwpwn cn pk.?= bw/CyC males, and eggs were collected for 6 days at 25OC. Three CyO+ progeny homozygous for cn and bw were recov- ered, each from a different bottle; a total of about 1.5-3.5 x IO5 prog- eny were screened. The ptc+ function in each case segregated with the mutagenized cn bw sp chromosome and mapped by meiotic re- combination between cn and en, at or near the ptc locus. Mechanisti- cally, the exchange could have been either double recombination or gene conversion. The frequency of the reversion events (3/75,000- 160,000 = 2.5-6 x 10m4) is consistent with either mechanism (Na- goshi and Geibart, 1987; Symmington and Pete& 1966).
Larval cuticles were prepared as described by Van der Meer (1977) and photographed using phase-contrast optics. We have ordered 14 ptc alleles in the following allelic series based on the severity of the cuticle phenotype, particularly with respect to the sensory hairs: U112 < F85 (IS’%) < F65 (25%) < R87 < 9828 < X103, B98, C22, E51, W109, C84,6P43,7M59, IN108 RX67, Df(2R)44CE/Df(2R)P14TE. ptc- in this paper refers to ptcrNIOB or to Df(2R)44CE/Df(2R)P14TE.
isolation of Genomic DNA and cDNA Clones A chromosome walk was initiated from three previously isolated lambda bacteriophage clones mapped in 44D-E (PhageLCPl [Snyder et al., 19611, Phage536 [Levy et al., 19621, and Phage2304 [Carlson and Hogness, 19651). A group of clones covering 114 kb around PhageLCPl was found by in situ hybridization to salivary gland chro- mosomes to lie entirely within the proximal part of Df(2R)44CE, outside Df(2R)P14TE and therefore outside the ptc region. A second chromo- some walk, including both Phage536 and Phage2304 and crossing both ends of Df(2R)P14TE. covered 204 kb. Chromosome walks uti- lized cosmid libraries constructed from a partial Sau3A digest of DNA from iso- flies in CosPneo or CoSpeR vectors (Haenlin et al., 1965) modified by Dr. R. Deuring to introduce Notl sites in the polylinker flanking the BamHl cloning site. The libraries were constructed and provided by Dr. J. Tamkun. The cosmids were isolated by standard chromosome walking techniques. To cross a region of clustered cos- mid ends (+165, Figure 3). the 11.5 kb BamHl fragment around +I65
was cloned by isolation of that BamHl fragment from a complete BamHi digest of iso DNA inserted into lambda EMBU (Kaiser and Murray, 1985). A cosmid (b3, Figure 3) containing the entire ptc tran- scription unit was isolated from a cosmid library constructed from partial BamHl digests of iso- DNA inserted into the Not-Bam-Not- CoSpeR vector. The orientation of the cloned DNA relative to the chro- mosome and to the deficiencies was determined by in situ hybridixa- tion to salivary gland polytene chromosomes (Bonner and Pardue, 1976). DNA probes were labeled by Klenow extension of random hexanucleotide-primed DNA (Feinberg and Vogelstein, 1963) incor- porating bio-11-UTP or bio-16UTP (ENZO) and detected with strep tavidin-alkaline phosphatase using the DNA detection kit (Bethesda Research Laboratories).
The 3-12 hr embryonic cDNA library in lambda gtl0 (Poole et al., 1965; a gift from Drs. L. Kauvar and T Kornberg, University of Califor- nia, San Francsico) wasscreened with the 1.6 kb and the 1.8 kb BamHl restriction fragments from the middle of theptc region (Figure 3 around +145), which were 32P-labeled by random primer extension (Feinberg
and Vogelstein, 1963). A O-24 hr iso- embryonic cDNA library in lambda gtli (made by Dr. J. Tamkun) was screened with the 4.6 kb BamHl fragment (Figure 3, +140) and a 5.5 kb BamHI-Sali fragment (Figure 3, +160). A total of 11 cDNAs were isolated using standard screening methods (Maniatis et al., 1982). ptc cDNAs were recovered at a frequency of 1 per 20,000 plaques screened. cDNA covering a 316 base gap (genomic) between J20 and K20 cDNAs (Figure 3) was directly sequenced after polymerase chain reaction amplification of cDNA (Saiki et al., 1988) synthesized from oligo(dT)-primed 4-8 hr em- bryonic polyadenylated RNA.
Preparation and Analysis of RNA and DNA Embryos were collected at defined times after oviposition. Larvae and pupae were staged from the time of egg hatching and harvested after examination of their morphology to confirm their developmental ages; all procedures were performed at 25OC. Staged embryos, larvae, and pupae were frozen in small batches in liquid nitrogen and stored at -80% until needed. Adult flies were taken from laboratory stock bot-
tles grown at room temperature. Total RNA was extracted by grinding frozen samples over dry ice, guanidine-HCI extraction, LiCl precipita- tion, and phenol extraction (Laughon et al., 1986). Poly(A)+ RNA was purified by one cycle of binding to oligo(dT)-cellulose (Maniatis et al., 1982). RNA was fractionated in formaldehyde-agarose gels, trans- ferred to Zetabind (AMF Cuno), and probed with 32P-labeled probe prepared by random primer extension according to standard methods (Feinberg and Volestein, 1983; Maniatis et al., 1982). DNA was isolated from adult flies as described by Scott et al. (1983). DNA blots were pre- pared and probed as described by Maniatis et al. (1982).
In situ hybridizations to 6-10 pm paraffin sections of embryos or larva were performed using 3H-labeied RNA probes as described by lngham et al. (1965a). The direction of transcription was determined using plus- and minus-strand probes for ptc and for its flanking tran- scripts (Figure 5). pc probes utilized antisense transcripts from the 1.8 kb BamHl genomic fragment around +I45 (Figure 3). Sense-strand transcripts were used as controls (not shown). The patterns of expres- sion of the 1.7 kb transcript (around +175) and the 2.4 kb transcript (around +127) were also determined using antisense probes; both transcripts were ubiquitiously distributed in O-12 hr embryos.
in situ hybridization to whole embryos utilized digoxigenin-deriva- tized DNA probes (Genius kit; Boehringer Mannheim) synthesized from the J20 cDNA as described by Tautz and Pfeiffle (1989).
DNA Sequencing and Computer Analysis Genomic BamHi restriction fragments and cDNAs were subcloned into pGEM7 or pGEM1 (Promega). Nested deletions were obtained using exonuclease ill (Erase-a-Base system, Promega) and sequenced using the dideoxy chain termination method (Sanger et al., 1977) as modified for use with Sequenase (United States Biochemical Corp.). The sense strands of cDNAs were sequenced, and the corresponding antisense strands of genomic DNAs were sequenced. GC-rich regions where sequence was ambiguous were sequenced again using dlTP When nested deletion endpoints were more than 250 bases apart, syn- thetic oligonucleotides were used as primers to fill the gaps. Five in- trons were detected in the cDNAs by comparison of the cDNA and genomic sequences. Double-stranded sequence around the splice

patched Transmembrane Protein Controls Segment Pattern 763
junctions, 3’ of the polyadenylation site and 5’ of the K12 cDNA, was obtained by sequencing the sense strand of genomic subclones using nested deletions or synthetic oligonucleotide primers. The comparison of sense and antisense sequences (isogenized y; cn bw sp vs. Oregon R strain) revealed 16 base substitution polymorphisms, 15 of which were located within the large open reading frame, 14 as silent changes and 1 histidine-to-arginine substitution (amino acid 139). The identifi- cation of only 16 differences over 5.4 kb, all but one of which were silent substitutions, validated the approach of sequencing cDNAs on one strand and genomic DNA on the other strand. DNA sequences were compiled using the Bionet GEL program (NIH grant P41RR01685) and analyzed using the lntelligenetics Inc. PC/Gene programs. The de- duced amino acid sequence was compared with the SWISSPROT and PIR databases (versions 10 and 19, respectively) using the program FASTA (Pearson and Lipman, 1968) and with other membrane protein sequences using the ALTSCHUL program of EuGene (Molecular Biol- ogy Information Resource, Department of Cell Biology, Baylor College of Medicine).
Acknowledgments
This work was initiated in the laboratory of Peter Lawrence. We are par- ticularly grateful to him and to Robert Holmgren, Nick Baker, and Iva Greenwald for their input early in this work. We thank Phil lngham for continuing open communication about ptc. A. Laughon, J. Tamkun, J. Bermingham, D. Gubb, S. McGill, and J. Kennison provided invaluable advice. We are indebted to C. Niisslein-Volhard, J. Kennison, G. Ising, D. Gubb, I? Simpson, and G. Yannopoulos for stocks; to L. Kauvar and J. Tamkun for libraries; to N. Baker, I? O’Farrell, M. Nell, D. Hogness, J. Manning, J. Fristrom, and R. Deuring for probes; to M. Ashburner for his Drosophila codon usage table; to D. Botstein and J. Mulholland for help with data base searching; and to J. Tamkun, C. Doe, M. Fuller, and W. Wood for careful readings of the manuscript. The research was supported by NIH grant HD24584, by an NIH postdoctoral fellowship (F32HDO6784) to J. E. H., and by a Searle Scholar Award to M. P S. M. P S. is an investigator of the Howard Hughes Medical Institute.
The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received July 6. 1989; revised September 21, 1989.
References
AbdeCMeguid, S. S., Shieh, H.-S., Smith, W. W., Dayringer, H. E., Violand, B. N., and Bentle, L. A. (1987). Three-dimensional structure of a genetically engineered variant of porcine growth hormone. Proc. Natl. Acad. Sci. USA 84, 8434-6437. Akam, M. (1987). The molecular basis for metameric pattern in the Dro- sophile embryo. Development 701, i-22.
Baker, N. E. (1987). Molecular cloning of sequences from wing/ess, a segment polarity gene in Dmsophifa: the spatial distribution of a tran- script in embryos. EMBO J. 6, 1765-1773.
Baker, N. E. (1988a). Embryonic and imaginal requirements for wing- less, a segment-polarity gene in Drosophila. Dev. Biol. 725, 96-108.
Baker, N. E. (1988b). Localization of transcripts from the wing/ess gene in whole Dmsophifa embryos. Development 703, 289-298.
Baumgartner, S., Bopp, D., Burri. M., and Nell, M. (1987) Structure of two genes at the goosebeny locus related to the paired gene and their spatial expression during Drosophila embryogenesis. Genes Dev. 1, 1247-1267
Banner, J. J., and Pardue. M. L. (1976). Ecdysone-stimulated RNA syn- thesis in imaginal discs of Dmsophile melanogaster Chromosoma 58, 87-98.
Bopp, D., Burri, M., Baumgartner, S., Frigerio, G., and Nell, M. (1986). Conservation of a large protein domain in the segmentation gene paired and in functionally related genes of Drosophila. Cell 47 1033- 1040.
Bryant, P J. (1975). Pattern formation in the imaginal wing disc of Dm-
sophi/a melanogastar: fate map, regeneration and duplication. J. Exp. Zool. 193, 49-78.
Bryant, S. V., French, V, and Bryant, P J. (1981). Distal regeneration and symmetry. Science 212, 993-1002.
Campos-Ortega, J. A., and Hartenstein. V. (1985). The Embryonic De velopment of Dmsophile melanogaster (Berlin: Springer-Verlag).
Carlson, J. R., and Hogness, D. S. (1985). The Jonah genes: a new multigene family in Drosophila melanogaste~ Dev. Biol. 103, 341-354.
Cavener, D. R. (1987). Comparison of the consensus sequence flank- ing translational start sites in Dmsophile and vertebrates. Nucl. Acids Res. 15, 1353-1381.
C&B, S., Preiss, A., Hailer, J., Schuh, R., Kienlin, A., Seifert, E., and Jtickle, H. (1987). The gooseberry-zipper region of Dmsophila: five genes encode spatially restricted transcripts in the embryo. EMBO J. 6, 2793-2801.
Cunningham, B. C., Jhurani, P., Ng, P., and Wells, J. A. (1989). Recep tor and antibody epitopes in human growth hormone identified by homolog-scanning mutagenesis. Science 243, 1330-1336.
DiNardo, S., Kuner, J. M., Theis, J., and O’Farrell, I? H. (1985). Develop ment of embryonic pattern in D. melanogaster as revealed by accumu- lation of the nuclear engrailed protein. Cell 43, 59-89.
DiNardo, S., Sher, E., Heemskerk-Jongens, J., Kassis, J. A., and O’Far- rell, P H. (1988). Two-tiered regulation of spatially patterned engmi/ed gene expression during Dmsophile embryogenesis. Nature 332,8&l- 609.
Edgar, B. A., Weir, M. P, Schiibiger, G., and Kornberg, T (1986). Repression and turnover pattern fushi tarazu RNA in the early Dro- sophila embryo. Cell 47, 747-754.
Eisenberg, D., Schwarz. E., Komaromy, M., and Wall, R. (1984). Analy- sis of membrane and surface protein sequences with the hydrophobic moment plot. J. Mol. Biol. 179, 125-142.
Feinberg, A., and Vogelstein, B. (1983). A technique for radiolabeling DNA restriction endonuclease fragments to high specific activity. Anal. B&hem. 132, 6-13.
Fjose, A., McGinnis, W. J., and Gehring, W. J. (1985). Isolation of a homoeobox-containing gene from the engrailed region of Drosophila and the spatial distribution of its transcript. Nature 313, 284-289.
Foe, V. E., and Alberts, B. (1983). Studies of nuclear and cytoplasmic behavior during the five mitotic cycles that precede gastrulation in Dro- sophila embryogenesis. J. Cell Sci. 61, 31-70.
French, V. (1976). Leg regeneration in the cockroach, B/ate//a german- ica. Il. Regeneration from a non-congruent tibia1 graft host junction. J. Embryol. Exp. Morphol. 35, 267-301.
Grau, Y., and Simpson, P. (1987). The segment polarity gene costal- in Drosophile. I, The organization of both primary and secondary em- bryonic fields may be affected. Dev. Biol. 122. 186-200.
Gray, A., Dull, T J., and Ullrich, A. (1983). Nucleotide sequence of epidermal growth factor cDNA predicts a 128,000-molecular weight protein precursor. Nature 303, 236-240.
Gubb, D., McGill, S., and Ashburner, M. (1988). A selective screen to recover chromosomal deletions and duplications in Dmsophils me. lanogaster. Genetics 119, 377-390.
Haenlin, M., Steller, H., Pirrotta, V., and Mohier, E. (1985). A 43 kilobase cosmid P transposon rescues the f.s(l)KIO morphogenetic locus and three adjacent Drosophila developmental mutants. Cell 40, 827-837.
Hafen, E., Kuroiwa, A., and Gehring, W. J. (1984). Spatial distribution Of transcripts from the segmentation gene fushi tamzu during Drosoph- ila embryonic development. Cell 37; 833-841.
lngham, P W. (1988). The molecular genetics of embryonic pattern for- mation in Drosophila. Nature 335, 25-34.
Ingham, P W., Howard, K. R., and Ish-Horowitz, D. (1985a). Transcrip tion pattern of the Dmsophile segmentation gene hairy. Nature 318, 439-445.
Ingham, I? W., Martinez-Arias, A., Lawrence, P. A., and Howard, K. (1985b). Expression of engrai/ed in the parasegment of Drosophila. Na- ture 377, 634-636.
Ising, G., and Block, K. (1961). Derivation-dependent distribution of in-

Cell 784
SertiOn Sites for a Dmsophila transposon. Cold Spring Harbor Symp. Quant. Biol. 45, 527-544.
Jaynes, J. B., and O’Farrell. f? H. (1988). Activation and repression of transcription by homeodomain-containing proteins that bind a com- mon site. Nature 336, 744-749.
Jiirgens, G. (1987). Segmental organisation of the tail region in the em- bryo of Dmsophi/a melanogaster. Roux’s Arch. Dev. Biol. 196, 141-157.
Jiirgens, G., Lehmann, Ft., Schardin, hf., and Niisslein-Volhard, C. (1986). Segmental organisation of the head in the embryo of Dmsoph- i/a melanogaster Roux’s Arch. Dev. Biot. 195, 359-377
Jiirgens, G., Wieschaus, E., Niisslein-Volhard, C., and Kluding, H. (1984). Mutations affecting the pattern of the larval cuticle in DmsophL melanogastef. II. Zygotic loci on the third chromosome. Roux’s Arch, Dev. Biol. 193, 283-295.
Kaiser, K., and Murray, N. E. (1985). The use of phage lambda replace- ment vectors in the construction of representative genomic DNA libraries. In DNA Cloning: A Practical Approach, Volume 1, D. M. Glover, ed. (Oxford: IRL Press), pp. l-47.
Klein, P., Kanehisa, M., and DeLisi, C. (1985). The detection and clas- sification of membrane-spanning proteins. Biochim. Biophys. Acta 615, 468-476.
Kornberg, T (1981). engra%d: a gene controlling compartment and segment formation in Dmsophile. Proc. Natl. Acad. Sci. USA 78, 1095-1099.
Kornberg, T., Siden, I., C’Farrell, P H., and Simon, M. (1985). The en- grailed locus of Drosophila: in situ localization of transcripts reveals compartment-specific expression. Cell 40, 45-53.
Kyte, J., and Doolittle, R. F (1982). A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 157, 133-148.
Laughon, A., Boulet, A. M., Bermingham, J. R., Laymon, R. A., and Scott, M. P (1986). The structure of transcripts from the homeotic An- tennapedia gene of Drosophila: two promoters control the major protein-coding region. Mol. Cell. Biol. 6, 4678-4889.
Lawrence, l? A. (1988). Gradients in the insect segment: the orientation of hairs in the milkweed bug Dncopeltus fasciatus. J. Exp. Biol. 44, 807-620.
Lawrence, P A., and Johnston, t? (1984). On the role of the engraileti gene in the internal organs of Dmsophile. EMBO J. 3, 2839-2844.
Levine, M., Hafen, E., Garber, R. L., and Gehring, W. J. (1983). Spatial distribution of Antennapedia transcripts during Dmsophile develop ment. EMBO J. 2, 2037-2048.
Levy, L. S.. Ganguly, R., Ganguly, N., and Manning, J. E. (1982). The selection, expression and organization of a set of head-specific genes in Dmsophila. Dev. Biol. 94, 451-464.
Lindsley, D. L., and Grell, E. H. (1988). Genetic Variations of Dmsoph- i/a melanogaster. Carnegie Inst. Wash. Publ. 827.
Locke, M. (1959). The cuticular pattern in an insect, Rhodnius pmlixus Sta. J. Exp. Biol. 36, 459-477.
Maniatis. 1, Fritsch, E. F., and Sambrook, J. (1982). Molecular Clon- ing: A Laboratory Manual (Cold Spring Harbor, New York: Cold Spring Harbor Laboratory).
Martinez-Arias, A., and Lawrence, l? A. (1985). Parasegments and compartments in the Drosophila embryo. Nature 313, 839-842.
Martinez-Arias, A., Baker, N. E., and Ingham, t? W. (1988). Role of seg- ment polarity genes in the definition and maintenance of cell states in the Dmsophi/a embryo. Development 103, 157-170.
Mohler, J. (1988). Requirements for hedgehog. a segmental polarity gene, in patterning larval and adult cuticle of Dmsophile. Genetics 120, 1081-1072.
Morata, G.. and Lawrence, P A. (1977). The development of wingless, a homeotic mutation of Drosophila. Dev. Biol. 56, 227-240.
Nagoshi, R. N., and Gelbart, W. M. (1987). Molecular and recombina- tonal mapping of mutations in the Ace locus of Dmsophila melanogas- fer Genetics 117, 487-502.
Nirsslein-Volhard, C., and Wieschaus, E. (1980). Mutations affecting segment number and polarity in Dmsophila. Nature 267: 795-801.
Niisslein-Volhard, C., Frohnhiifer, H. G., and Lehman% R. (1987). De-
termination of anteroposterior polarity in Drosophila. Science 238, 1875-1881.
Ntisslein-Volhard, C., Wieschaus, E., and Kluding, H. (1984). Mutations affecting the pattern of the larval cuticle in Drosophila melanogaster. I. Zygotic loci on the second chromosome. Roux’s Arch. Dev. Biol. 193, 287-282.
Drenic, T., Chidsey, J., and Holmgren, R. (1987). Cell and cubitus interruptus-Dominant: two segment polarity genes on the fourth chro- mosome in Drosophila. Dev. Biol. 124, 50-58.
Patel, N. H., Schafer, B., Goodman, C. S., and Holmgren, R. (1989). The role of segment polarity genes during Dmsophila neurogenesis. Genes Dev. 3, 890-904.
Pearson, W. R., and Lipman, D. J. (1988). Improved tools for biological sequence comparison. Proc. Natl. Acad. Sci. USA 85,2444-2448.
Perrimon. N., and Mahowald, A. t? (1987). Multiple functions of seg- ment polarity genes in Dmsophils. Dev. Biol. 119, 587-800.
Perrimon, N., Engstrom, L., and Mahowald, A. R (1989). Zygotic lethals with specific maternal effect phenotypes in Drosophila melanogastec I. Loci on the X chromosome. Genetics 121, 333352.
Poole, S. J., Kauvar, L. M., Drees, B., and Kornberg, T (1985). The en- grailed locus of Drosophila: structural analysis of an embryonic tran- script. Cell 40, 37-43.
Proudfoot, N. J., and Brownlee, G. G. (1978). 3’ non-coding region se- quences in eucaryotic mRNA. Nature 263, 211-214.
Rao, J. K. M., and Argo& P (1988). A conformational preference pa- rameter to predict helices in integral membrane proteins. Biochim. Bio- phys. Acta 869, 197-214.
Riggleman, B., Wieschaus, E., and Schedl, F! (1989). Molecular analy- sis of the armadillo locus: uniformly distributed transcripts and a pro- tein with novel internal repeats are associated with a Drosophila sep ment polarity gene. Genes Dev. 3, 98113.
Rijsewijk, R., Schuermann, M., Wagenaar, E., Parren, P.. Weigel, D., and Nusse, R. (1987). The Drosophila homolog of the mouse mam- mary oncogene in&l is identical to the segment polarity gene wingless. Cell 50, 849857.
Ruoslahti, E., and Pierschbacher, M. D. (1987). New perspectives in ceil adhesion: RGD and integrins. Science 238, 491-497.
Saiki, R. K., Gelfand, D. H., Stoffel, S., Scharf, S. J., Higuchi, R., Horn, G. T, Mullis, K. B., and Erlich, H. A. (1988). Primer-directed enzymatic amplification of DNA with a thermostable DNA polymerase. Science 239, 487-491.
Sanger, F., Nicklen, S., and Coulson, A. R. (1977). DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74, 5483- 5487.
Schiipbach, T., and Wieschaus, E. (1988). Germline autonomy of maternal-effect mutations altering the embryonic body pattern of Dm- sophila. Dev. Biol. 113, 443-448.
Scott, J., Urdea, M., Quiroga, M. R., Sanchez-Pescador, R., Fong, N., Selby, M., Rutter, W. J., and Bell, G. I. (1983). Structure of a mouse submaxillary messenger RNA encoding epidermal growth factor and seven related proteins. Nature 221, 238-240.
Scott, M. P, and Carroll, S. B. (1987). The segmentation and homeotic gene network in early Drosophila development. Cell 51, 689-898.
Scott, M. P.. Tamkun, J. W.. and Hartzell, G. W.. Ill (1989). The struc- ture and function of the homeodomain. Biochim. Biophys. Acta 969, 25-48.
Scott, M. P. Weiner, A. J., Hazelrigg, T. I., Polisky, B. A., Pirrotta, V., Scalenghe, F., and Kaufman, T. C. (1983). The molecular organization of the Antennapedia locus of Drosophila. Cell 35, 783-778.
Shapiro, M. B., and Senapathy, P (1987). RNAsplice junctionsof differ- ent classes of eukaryotes: sequence statistics and functional implica- tions in gene expression. Nucl. Acids Res. 15, 7l55-7l74.
Sims, J. E.. March, C. J., Cosman, D., Widmer, M. B., MacDonald, H. R., McMahan, C. J., Grubin, C. E., Wignall, J. M., Jackson, J. L., Call, S. M., Friend, D., Alpert, A. R., Gillis, S., Urdal, D. L., and Dower, S. K. (1988). cDNA expression cloning of the IL-1 receptor, a member of the immunoglobulin superfamily. Science 241, 585-589.

gF/red Transmembrane Protein Controls Segment Pattern
Snyder, M.. Hirsch, J., and Davidson, N. (1961). The cuticle genes of Drosophila: a developmentally regulated gene cluster. Cell 25, 165- 177.
Symington, L. S., and Pete& T D. (1968). Expansions and contractions of the genetic map relative to the physical map of yeast chromosome Ill. Mol. Cell. Biol. 8, 595-604.
Tautz, D., and Pfeiffle, C. (1969). A non-radioactive in situ hybridization method for the localization of specific RNAs in Drosophda embryos re- veals translational control of the segmentation gene hunchback. Chro- mosoma 98, 61-65.
Van der Meer, J. M. (1977). Optical clean and permanent whole mount preparation for phase-contrast microscopy of cuticular structures of in- sect larvae. Dros. Inf. Serv. 52, 160.
Walker, J. E., Saraste, M., Runswick, M. J.. and Gay, N. J. (1962). Dis- tantly related sequences in the alpha- and beta-subunits of ATP syn- thase, myosin, kinases and other ATP-requiring enzymes and a com- mon nucleotidabinding fold. EMBO J. 8, 945-951.
Wieschaus. E., and Riggleman, R. (1967). Autonomous requirements for the segment polarity gene armadillo during Drosophila embryogen- esis. Cell 49, 177-164.
Wieschaus, E.. Niisslein-Volhard, C., and Jiirgens, G. (1984). Muta- tions affecting the pattern of the larval cuticle in Drosophila melanogas- fer. Ill. Zygotic loci on the X-chromosome and fourth chromosome. Roux’s Arch. Dev. Biol. 793, 296-367
Wright, D. A., and Lawrence, P A. (1981). Regeneration of the segment boundary in Oncopeltus. Dev. Biol. 85, 317-327.
GenBank Acceselon Number
The accession number for the sequence reported in this paper is M26416.





























