Embed Size (px)
Citation preview

of October 31, 2018.This information is current as
Induced Enteropathy−Epithelial Regeneration following T Cell
Regulates Small Intestinal Crypt Damage and The Cytosolic Microbial Receptor Nod2
Catherine Streutker, Dana J. Philpott and Kenneth CroitoruDavid Prescott, Susan J. Robertson, Charles Maisonneuve, Galliano Zanello, Ashleigh Goethel, Sandrine Rouquier,
http://www.jimmunol.org/content/197/1/345doi: 10.4049/jimmunol.16001852016;
2016; 197:345-355; Prepublished online 20 MayJ Immunol
MaterialSupplementary
5.DCSupplementalhttp://www.jimmunol.org/content/suppl/2016/05/20/jimmunol.160018
Referenceshttp://www.jimmunol.org/content/197/1/345.full#ref-list-1
, 8 of which you can access for free at: cites 47 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2016 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on October 31, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on October 31, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
The Cytosolic Microbial Receptor Nod2 Regulates SmallIntestinal Crypt Damage and Epithelial Regenerationfollowing T Cell–Induced Enteropathy
Galliano Zanello,* Ashleigh Goethel,† Sandrine Rouquier,* David Prescott,‡
Susan J. Robertson,† Charles Maisonneuve,† Catherine Streutker,x Dana J. Philpott,† and
Kenneth Croitoru*,†,{,‖
Loss of function in the NOD2 gene is associated with a higher risk of developing Crohn’s disease (CD). CD is characterized by
activation of T cells and activated T cells are involved in mucosal inflammation and mucosal damage. We found that acute T cell
activation with anti-CD3 mAb induced stronger small intestinal mucosal damage in NOD22/2 mice compared with wild-type mice.
This enhanced mucosal damage was characterized by loss of crypt architecture, increased epithelial cell apoptosis, delayed
epithelial regeneration and an accumulation of inflammatory cytokines and Th17 cells in the small intestine. Partial microbiota
depletion with antibiotics did not decrease mucosal damage 1 d after anti-CD3 mAb injection, but it significantly reduced crypt
damage and inflammatory cytokine secretion in NOD22/2 mice 3 d after anti-CD3 mAb injection, indicating that microbial
sensing by Nod2 was important to control mucosal damage and epithelial regeneration after anti-CD3 mAb injection. To deter-
mine which cells play a key role in microbial sensing and regulation of mucosal damage, we engineered mice carrying a cell-
specific deletion of Nod2 in villin and Lyz2-expressing cells. T cell activation did not worsen crypt damage in mice carrying either
cell-specific deletion of Nod2 compared with wild-type mice. However, increased numbers of apoptotic epithelial cells and higher
expression of TNF-a and IL-22 were observed in mice carrying a deletion of Nod2 in Lyz2-expressing cells. Taken together, our
results demonstrate that microbial sensing by Nod2 is an important mechanism to regulate small intestinal mucosal damage
following acute T cell activation. The Journal of Immunology, 2016, 197: 345–355.
In the gastrointestinal tract, regulation of host–microbiomeinteractions is crucial to maintain intestinal homeostasis (1).Nod2, a cytosolic sensor of peptidoglycan, is one of the im-
portant regulators of this interaction. Nod2 is expressed in a varietyof cell types from hematopoietic and nonhematopoietic origins,including myeloid and lymphoid cells, stromal cells, intestinal ep-ithelial cells (IEC), and Paneth cells (2–6), yet it is unclear whichNod2-expressing cell is key for a balanced host–microbial rela-tionship. After stimulation with Nod2-specific ligand muramyl di-peptide (MDP), Nod2 signaling regulates gene expression of a
number of mediators thought to promote maintenance of intestinalbarrier integrity, the activation of antimicrobial function and auto-phagy, and the regulation of immune homeostasis (7). More recently,a crucial role of Nod2 in intestinal epithelial stem cell survival wasreported, highlighting the importance of Nod2 in intestinal epi-thelial regeneration (8).Loss-of-function mutations in the NOD2 gene were the first de-
fined genetic risk factor identified for Crohn’s disease (CD) (9, 10),and carriage of NOD2 risk alleles is associated with ileal diseaselocation (11). Although the etiology of CD remains unclear, the highnumbers of T cells in the inflamed intestinal mucosa, the secretion oflarge amounts of T cell–derived proinflammatory cytokines, and therequirement for T cells in various animal models of chronic intestinalinflammation strongly suggest a role of T cells in the pathogenesisof CD (12). Our group previously showed that, although Nod2 isexpressed and functionally active in murine CD4+ T cell subsets, theexpression of Nod2 is not required for the development or theprevention of the T cell transfer model of colitis in Rag-deficientmice (6). It remains possible that Nod2 modulates immune ho-meostasis in the small intestine in response to acute T cell activation.In this study, we used a model of T cell–induced enteropathy
triggered by i.p. injection with T cell activating monoclonal anti-CD3 Ab (anti-CD3 mAb) (13) and investigated the role of Nod2on T cell activation, T cell–induced mucosal damage and recov-ery, specifically within the small intestine. Our results showed thatacute T cell activation led to more severe crypt damage, increasedcrypt IEC apoptosis, and delayed epithelial regeneration in Nod2-deficient mice. Microbial sensing by Nod2 was important for theregulation of mucosal damage and epithelial regeneration. Dele-tion of Nod2 in intestinal epithelial cells and in Lyz2-expressingphagocytes was not sufficient to recapitulate the crypt damage
*Department of Medicine, University of Toronto, Toronto, Ontario M5S 1A8,Canada; †Department of Immunology, University of Toronto, Toronto, Ontario M5S1A8, Canada; ‡Department of Laboratory Medicine and Pathobiology, University ofToronto, Toronto, Ontario M5S 1A8, Canada; xDepartment of Pathology and LaboratoryMedicine, St. Michael’s Hospital, Toronto, Ontario M5B 1W8, Canada; {Zane CohenCentre for Digestive Diseases, The Lunenfeld-Tanenbaum Research Institute, SinaiHealth System, Toronto, Ontario M5G 1X5, Canada; and ‖Division of Gastroenterology,Mount Sinai Hospital, Toronto, Ontario M5G 1X5, Canada
ORCIDs: 0000-0002-0856-2759 (A.G.); 0000-0002-1508-0934 (S.R.); 0000-0002-9798-9526 (D.P.); 0000-0003-1231-0180 (K.C.).
Received for publication January 29, 2016. Accepted for publication April 21, 2016.
This work was supported by the Canadian Association of Gastroenterology, Crohn’sand Colitis Canada, the Canadian Foundation of Innovation, the Canadian Institutesof Health Research, and the Mount Sinai Hospital Department of Medicine.
Address correspondence and reprint requests to Dr. Kenneth Croitoru, University ofToronto, 600 University Avenue, Room 437, Toronto, ON M5G 1X5, Canada. E-mailaddress: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: CD, Crohn’s disease; IBD, inflammatory boweldisease; IEC, intestinal epithelial cell; ISC, intestinal stem cell; MDP, muramyldipeptide; Treg, regulatory T; WT, wild-type.
Copyright� 2016 by The American Association of Immunologists, Inc. 0022-1767/16/$30.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1600185
by guest on October 31, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

observed in NOD22/2 mice. However, increased numbers of apo-ptotic epithelial cells and higher expression of TNF-a and IL-22 wereobserved in mice carrying a deletion of Nod2 in Lyz2-expressing cells,indicating that Nod2 expression in phagocytes can alter the immuneresponse and the mucosal damage induced by anti-CD3 mAb injection.
Materials and MethodsMice
C57BL/6, NOD22/2, NOD2flox/flox, Villin-Cre, and LyzM-Cre mice weremaintained under standard pathogen-free conditions at the University ofToronto animal facility. Wild-type (WT) and NOD22/2 mice were used asseparate strains and were crossed to generate littermate control mice.NOD22/2 mice were obtained from Dr. J.-P. Hugot (Hopital Robert Debre,Universite Paris Diderot, Paris, France) (14), Villin-Cre (B6.Cg-Tg(Vil-cre)997Gum/J) and LyzM-Cre (B6.129P2-Lyz2tm1(cre)Ifo/J) mice were pur-chased from The Jackson Laboratory (Bar Harbor, ME), and NOD2flox/flox
mice were obtained from Dr. P. Rosenstiel (Institute for Clinical MolecularBiology, Christian-Albrechts-University, Kiel, Germany) (15).
NOD2flox/flox mice were generated by genOway (Lyon, France). TheloxP sites were inserted after exons 1 and 3 of the NOD2 gene in 129SVmouse embryonic stem cells. The distal loxP site was inserted togetherwith a FRT-neomycin selection cassette. The engineered mouse line wasbred with FLP deleter-mice allowing the deletion of the FRT-flankedneomycin selection cassette. The resultant NOD2flox/+ mice expressedtwo loxP sites in a genomic region including exons 2 and 3. Mice werebackcrossed onto C57BL/6 background for 10 generations.
Mice expressing a conditional deletion of NOD2 in villin-expressingcells (NOD2DIEC) and in lysozyme 2–expressing cells (NOD2DLyz2) wereobtained by crossing NOD2flox/flox mice and Villin-Cre or LyzM-Cre mice. Ex-pression of the Cre recombinase resulted in the deletion of both exons 2 and 3.
All mouse experiments were conducted as approved by the University ofToronto animal care committee in accordance with the regulations of theCanadian Council on animal care (University of Toronto approved protocolnumber 20010966).
Material and reagents
The following Abs were used for the experiments: anti-CD3ε (145-2C11;BioLegend, San Diego, CA), anti–CD3ε-PE.Cy5 (145-2C11; eBioscience,San Diego, CA), anti–CD4-A780 (GK1.5; eBioscience), anti–Foxp3-PE(FJK-16s; eBioscience), anti–IL-17A-Brilliant Violet (TC11-18H10.1;eBioscience), anti–IFN-g-FITC (XMG1.2; eBioscience), anti-CD11b (M1/70;eBioscience), anti–Gr-1 (RB6-8C5; eBioscience), brefeldin A, and Foxp3Fixation/Permeabilization Kit (eBioscience). PMA ( P8139) and ionomycin(I0634) were purchased from Sigma-Aldrich (St. Louis, MO). Collagenase Dwas purchased from Roche Applied Science (Penzberg, Germany).
T cell–induced enteropathy
Mice received an i.p. injection of 50 mg monoclonal anti-CD3ε Ab (145-2C11; BioLegend). Mice were assessed for clinical signs of enteropathy,including activity level, appearance of the coat, perianal state and behavior.Mice were sacrificed at days 1, 3, and 5 postinjection.
Antibiotic treatment consisted of administration of ampicillin, neomycin,and metronidazole each at 1 g/l, in drinking water provided ad libitum from10 d prior to anti-CD3 mAb injection until sacrifice, 1 and 3 d postinjection.
Histological examination
Small-bowel tissues were fixed in 10% buffered formalin and embedded inparaffin sections. H&E, TUNEL, and Ki67 staining were performed by thePathology Core, Center for Phenogenomics (Toronto, ON, Canada). Smallbowels were graded for severity of crypt damage by an experienced pa-thologist (C.S.) blinded to the experimental set-up. Each segment was gradedon a scale from 1 (no change) to 4 (severe damage and loss of crypts).
Analysis of mRNA relative expression using quantitativereal-time PCR
Total RNA was isolated from T cells using RNeasy Mini Kit (Qiagen,Germantown, MD). RNA samples were treated with DNAse Amp I Grade(Invitrogen) and cDNA were synthesized using Mu-MLV reverse tran-scriptase (Eurogentec, Liege, Belgium). For RT-PCR, cDNA samples werecombined with primer/probe sets and Power SYBR Green Master Mix(Applied Biosystems, Warrington, U.K.), according to the manufacturer’srecommendations. Samples were normalized internally using the averagecycle quantification of GUSB, HMBS, and TBP simultaneously. The primersequences were as follows: GUSB sense 59-AGC-CCT-TCG-GGA-CTT-
TAT-TG-39, GUSB antisense 59-AAT-GGG-CAC-TGT-TGA-TCC-TC-39,HMBS sense 59-GTA-CCC-TGG-CAT-ACA-GTT-TG-39, HMBS antisense59-CCT-TGG-TAA-ACA-GGC-TCT-TC-39, TBP sense 59-GCA-ACA-GCA-GCA-GCA-ACA-AC-39, TBP antisense 59-CAA-CGG-TGC-AGT-GGT-CAG-AG-39, CCL20 sense 59-TTG-CTT-TGG-CAT-GGG-TAC-TG-39, CCL20 antisense 59-TTC-ATC-GGC-CAT-CTG-TCT-TG-39, CXCL10sense 59-CAT-CCT-GCT-GGG-TCT-GAG-TG-39, CXCL10 antisense 59-AGG-ATA-GGC-TCG-CAG-GGA-TG-39, IL-1b sense 59-TGT-CTT-TCC-CGT-GGA-CCT-TC-39, IL-1b antisense 59-TCA-TCT-CGG-AGC-CTG-TAG-TG-39, IL-6 sense 59-ACA-AAG-CCA-GAG-TCC-TTC-AG-39, IL-6antisense 59-TGG-ATG-GTC-TTG-GTC-CTT-AG-39, IL-22 sense 59-CAC-AGA-TGT-CCG-GCT-CAT-CG-39, IL-22 antisense 59-CCT-GCA-TGT-AGG-GCT-GGA-AC-39, TNF-a sense 59-GTC-AGC-CGA-TTT-GCT-ATC-TC-39, TNF-a antisense 59-AGA-CTC-CTC-CCA-GGT-ATA-TG-39, LGR5 sense 59-AGA-CTA-CGC-CTT-TGG-AAA-CC-39, LGR5antisense 59-TGG-AGA-GTG-TCT-TGA-TTG-CAG-39, OLFM4 sense 59-CAA-AAG-TGA-CCT-TGT-GCC-TG-39, and OLFM4 antisense 59-ACC-ATG-ACT-ACA-GCT-TCC-AAG-39. Real-time assays were run on a Bio-Rad C1000 Touch Thermal Cycler (Bio-Rad, Hercules, CA). Expressiondata are expressed as relative values after Genex macro analysis (Bio-Rad).
Cytokine and myeloperoxidase quantification
For quantification of cytokine level, one centimeter of medial small-boweltissue was crushed in tubes containing ceramic beads and 500 ml HBSS-1%protease inhibitors (Sigma-Aldrich). The level of IFN-g, IL-10, IL-17A, andmyeloperoxidase (MPO) were quantified by ELISA following the manu-facturer recommendations (R&D Systems, Minneapolis, MN).
Cell sorting and flow cytometry
The small bowel was extracted, washed in PBS, and incubated (37˚C, 20min) in stripping buffer (PBS, 5 mM EDTA). After stripping, the smallbowel was cut in 0.5-cm pieces and digested in buffer containing RPMI1640 medium, 20% FBS, and 2 mg/ml collagenase D for 1 h at 37˚C.Digested material was passed through 100- and 70-mm cell strainers, andthe cells were collected by centrifugation.
For detection of intracellular cytokines, cells were cultured in 1 ml RPMI1640 medium-20% FBS containing 50 ng/ml PMA, 1000 ng/ml ionomycin,and 1 ml brefeldin A for 4 h at 37˚C. For detection of Foxp3, cells were notsimulated and were directly stained with anti-CD3 and anti-CD4 fluo-rescently labeled mAbs for 30 min. Cells were permeabilized with Foxp3Fixation/Permeabilization Kit (eBioscience) and then incubated with anti–IL-17A, anti–IFN-g, or anti-Foxp3 fluorescently labeled mAb for 30 min.Data were acquired on a FACSFortessa (BD Biosciences, Franklin Lakes,NJ) and analyzed using FlowJo Software (Tree Star, Ashland, OR).
Assessment of NOD2 deletion in NOD2DIEC and NOD2DLyz2 mice
To quantify NOD2 deletion in IEC, villi of the small intestine were re-moved by scraping and washed with cold PBS. Small intestinal tissue wascut into 2- to 3-mm pieces, carefully rocked in PBS containing 2 mMEDTA for 30 min at 4˚C, and then vigorously washed in PBS. The crypt-enriched supernatant fractions were filtered through a 70-mm cell strainer,centrifuged at 300 3 g for 5 min at 4˚C and resuspended in 50 ml Matrigel(Corning, New York, NY). The crypt-containing organoid cultures wereplated onto a 24-well plate and maintained in 500 ml crypt culture mediumcontaining growth factors (EGF at 0.05 mg/ml, R-spondin 1, and Noggin at10% of the conditioned medium). Organoids were maintained for 7 d inculture, and the medium was changed once a week.
Deletion of NOD2 in Lyz2-expressing cells was assessed after i.p. in-jection of 50 mg MDP or vehicle (PBS) in NOD2DLyz2 and WT littermatemice. The peritoneal cavity was lavaged 24 h later with 5 ml cold PBS. Theresulting cellular exudate was collected and incubated with anti-CD11b andanti–Gr-1 fluorescently labeled mAb to assess neutrophil recruitment.
Statistical analysis
The results are depicted as mean 6 SEM. Statistical analysis was per-formed using nonparametric Mann–Whitney U test. The differences be-tween two groups were considered significant when p# 0.05. All calculationswere performed using GraphPad Prism software (GraphPad, La Jolla, CA).Results are representative of $3 experiments.
ResultsNod2 regulates mucosal damage and small intestinal cryptarchitecture after acute T cell activation
To investigate whether Nod2 regulates acute T cell activation andT cell–induced mucosal damage in the small intestine, we treated
346 Nod2 REGULATES MUCOSAL DAMAGE IN THE SMALL INTESTINE
by guest on October 31, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

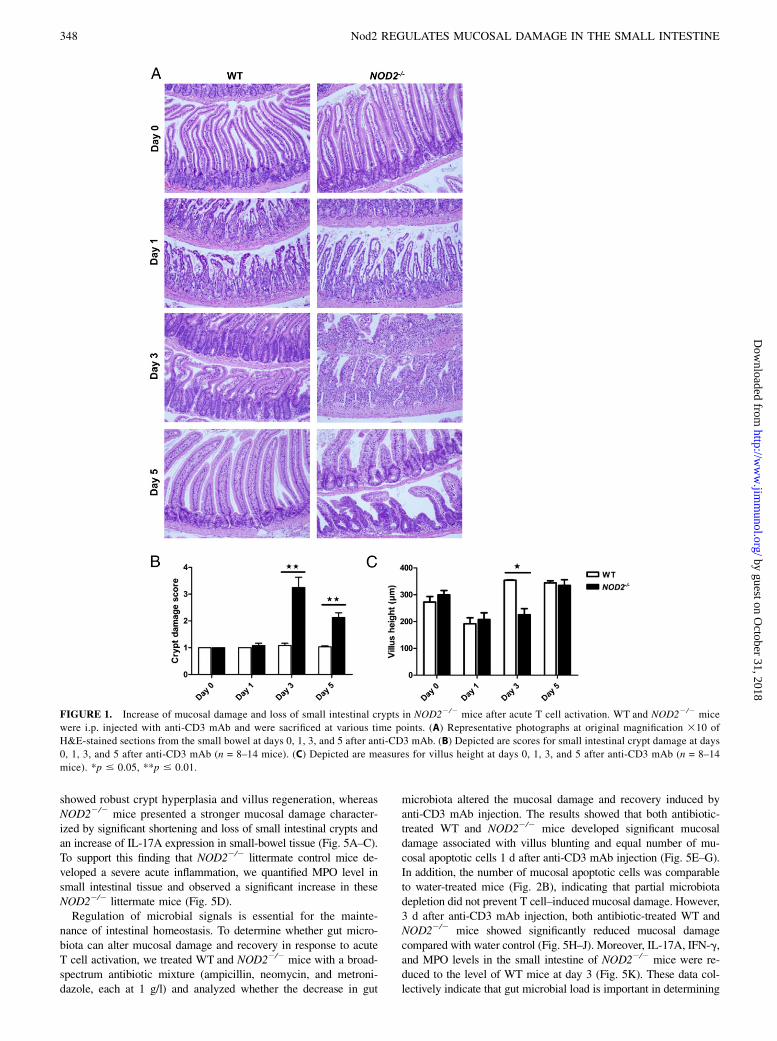
WT and NOD22/2 mice with a single i.p. injection of 50 mg anti-CD3 mAb. One day after T cell activation, mucosal damage in bothWT and NOD22/2 mice was characterized by increased IEC apo-ptosis and villus blunting (Fig. 1A). Three days after anti-CD3 mAbinjection, WT mice began to recover as shown by crypt hyperplasiaand villus regeneration, with full recovery achieved by day 5(Fig. 1A). In contrast, 3 d after anti-CD3 mAb injection, NOD22/2
mice continued to show severe mucosal damage. This was char-acterized not only by increased cell infiltration in the lamina propriaand villous blunting but also with significant shortening and loss ofsmall intestinal crypts compared with WT mice (Fig. 1). NOD22/2
mice started to recover 5 d after anti-CD3 mAb injection as ob-served by crypt hyperplasia and villus regeneration (Fig. 1). How-ever, NOD22/2 mice did not restore a normal crypt architecturecompared with WT mice at day 5 (Fig. 1).
Nod2 regulates intestinal epithelial cell regeneration inresponse to T cell–induced enteropathy
Because NOD22/2 mice developed more severe mucosal injurywith a delayed recovery phase after acute T cell activation, weexplored the factors that contributed to epithelial regeneration. Inorder to quantify the degree of apoptosis after mucosal injury andrepair, we used an immunohistochemical TUNEL assay to identifyapoptotic bodies and found an equal number in the mucosa of WTand NOD22/2 mice 1 d after anti-CD3 mAb injection (Fig. 2A,2B). The number of mucosal apoptotic bodies started to decreasein WT mice 3 d after T cell activation, whereas it remained as highas on day 1 in NOD22/2 mice (Fig. 2A, 2B). Five days after anti-CD3 mAb injection, the number of apoptotic bodies was sig-nificantly higher in NOD22/2 mice as compared with WT mice(Fig. 2A, 2B). In addition, these apoptotic bodies were specificallylocated in the epithelium of small intestinal crypts in NOD22/2
mice, as opposed to the lamina propria (Fig. 2C, 2D). Paneth cellsand intestinal stem cells (ISC) are located in the crypt epithelium;therefore, we counted the number of granulated Paneth cells percrypt and assessed the expression of LGR5 and OLFM4, ISCmarkers in the small intestine. One and three days after T cellactivation, the number of Paneth cells significantly decreased inboth WT and NOD22/2 mice (Fig. 2E). Five days after anti-CD3mAb injection, the number of Paneth cells recovered in WT micebut not in NOD22/2 mice (Fig. 2E). Our data also showed that themRNA expression levels of LGR5 and OLFM4 were significantlydecreased in both WT and NOD22/2 mice after T cell activation(Fig. 2F). LGR5 level was not different between WT and NOD22/2
mice at days 1 and 3 after anti-CD3 mAb injection. On day 5,LGR5 expression was upregulated in both WT and NOD22/2
mice, but this upregulation was significantly greater in WT mice(Fig. 2F). OLFM4 level was significantly decreased in both WTand NOD22/2 mice on day 1, and this decrease was more pro-nounced in NOD22/2 mice (Fig. 2F). On day 3, OLFM4 levelrecovered in WT mice but not in NOD22/2 mice, who showedrecovery only by day 5 after anti-CD3 mAb injection (Fig. 2F).To assess the degree of epithelial cell proliferation during the
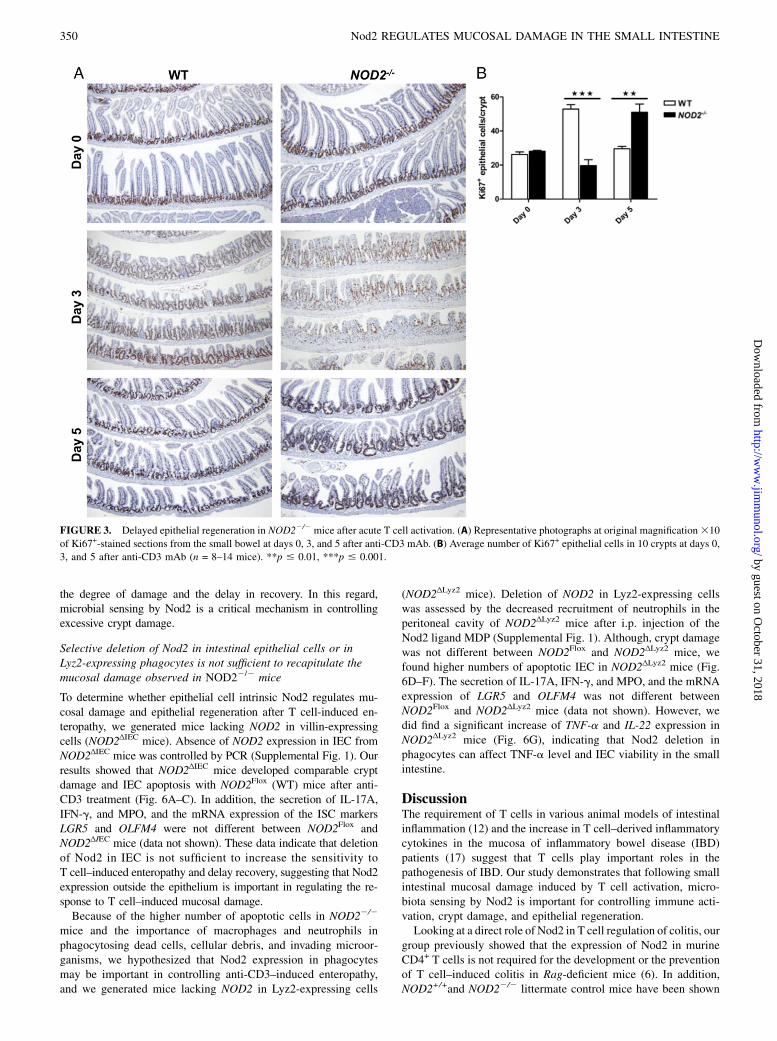
recovery phase, we used Ki67 staining. In WT mice, 3 d after anti-CD3 mAb injection, most of the Ki67+ cells were located in thehyperplastic small intestinal crypts, corresponding to the normalproliferative zone of the small intestine, whereas few Ki67+ cellswere found in the shortened crypts of NOD22/2 mice (Fig. 3).Moreover, we observed an atypical pattern of Ki67+ within epi-thelial cells extending along the villus in NOD22/2 mice (Fig. 3A).Five days after anti-CD3 mAb injection, Ki67+ cells were confinedto the crypt epithelium in WT mice, whereas a high number ofKi67+ cells were located in the hyperplastic crypts and along thevillus in NOD22/2mice (Fig. 3). Taken together, these data indicate
that acute T cell activation with anti-CD3 mAb induces strongerapoptosis of small intestinal mucosal cells, particularly crypt epi-thelial cells of NOD22/2mice compared with WT mice. The highernumber of apoptotic cells and the delayed epithelial regeneration inthe small intestine of NOD22/2 mice contribute to the severe mu-cosal injury, the loss of crypt architecture, and the delayed recoveryphase.
Small intestinal mucosal damage is associated with strongerimmune activation and accumulation of Th17 cells inNOD22/2 mice
Repeated low-dose injections (20 mg) of anti-CD3 mAb havebeen shown to induce IL-6–dependent generation and CCL20-dependent accumulation of Th17 cells in the small intestine ofmice (16). To explore the mechanisms by which acute T cell ac-tivation induced more severe small intestinal crypt damage inNOD22/2 mice, we examined the expression of T cell–associatedcytokines and the accumulation of T cell subsets within the smallintestinal lamina propria of WT and NOD22/2 mice. Our resultsshowed that anti-CD3 mAb injection significantly upregulated thelevel of IL-17A, IFN-g, and IL-10 cytokines in NOD22/2 micecompared with WT mice (Fig. 4A). IL-17A level was significantlyhigher in NOD22/2 mice at days 3 and 5 after anti-CD3 mAbinjection, whereas IFN-g and IL-10 levels were significantlyhigher only at day 3 (Fig. 3A). In parallel, we found that IL-1b,IL-6, and CCL20 transcripts were significantly upregulated inNOD22/2 mice (Fig. 4B), suggesting that these factors maycontribute to the generation and the accumulation of Th17 cells inthe lamina propria of NOD22/2 mice. The expression of IL-22,TNF-a, and CXCL10 (also known as IFN-g–induced protein 10)were also increased in NOD22/2 mice. Taken together, the in-creased secretion and expression in these cytokines indicated astronger inflammatory response in NOD22/2 mice (Fig. 4B).We then examined lamina propria lymphocytes from WT and
NOD22/2 mice and found that total CD4+ T cell number, IL-17A–expressing T cells, IFN-g–expressing T cells, and Foxp3+ regu-latory T (Treg) cell numbers were no different in untreated WTand NOD22/2 mice (Fig. 4D). We observed that anti-CD3 mAbinjection reduced total CD4+ T cell numbers within the laminapropria of both WT and NOD22/2 mice, but these numbers werenot different, suggesting that WT and Nod2-deficient CD4+ T cellspresented the same sensitivity to apoptosis (Fig. 4C, 4D). Threedays after anti-CD3 mAb, we found a significant increase in IL-17A–expressing T cells within the lamina propria of NOD22/2
mice compared with WT mice (Fig. 4C, 4D). In contrast, Foxp3+
Treg cell number was lower in NOD22/2 mice compared with WTmice, suggesting that the Th17/Treg ratio may in part explain thesevere mucosal damage in NOD22/2 mice (Fig. 4C, 4D). Last, wefound that the number of IFN-g–expressing T cells was not dif-ferent between WT and NOD22/2 mice 3 d after anti-CD3 mAbinjection (Fig. 4C, 4D). Taken together, these data indicate thatsmall intestinal mucosal damage is associated with accumulationof IL-17A–expressing T cells in NOD22/2 mice.
Sensing of gut microbiota by Nod2 is important for theregulation of T cell–induced enteropathy
To rule out the possibility that NOD22/2 mice are more sensitiveto T cell–induced enteropathy because of major differences inmicrobiota composition with WT mice, we crossed WT andNOD22/2 mice to generate littermate controls. NOD2+/+andNOD22/2 littermate control mice were sacrificed 3 d after anti-CD3 mAb injection because the strongest difference in mucosaldamage was shown at that time point. As previously described innonlittermate mice, we found that WT littermate control mice
The Journal of Immunology 347
by guest on October 31, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
showed robust crypt hyperplasia and villus regeneration, whereasNOD22/2 mice presented a stronger mucosal damage character-ized by significant shortening and loss of small intestinal crypts andan increase of IL-17A expression in small-bowel tissue (Fig. 5A–C).To support this finding that NOD22/2 littermate control mice de-veloped a severe acute inflammation, we quantified MPO level insmall intestinal tissue and observed a significant increase in theseNOD22/2 littermate mice (Fig. 5D).Regulation of microbial signals is essential for the mainte-
nance of intestinal homeostasis. To determine whether gut micro-biota can alter mucosal damage and recovery in response to acuteT cell activation, we treated WT and NOD22/2 mice with a broad-spectrum antibiotic mixture (ampicillin, neomycin, and metroni-dazole, each at 1 g/l) and analyzed whether the decrease in gut
microbiota altered the mucosal damage and recovery induced byanti-CD3 mAb injection. The results showed that both antibiotic-treated WT and NOD22/2 mice developed significant mucosaldamage associated with villus blunting and equal number of mu-cosal apoptotic cells 1 d after anti-CD3 mAb injection (Fig. 5E–G).In addition, the number of mucosal apoptotic cells was comparableto water-treated mice (Fig. 2B), indicating that partial microbiotadepletion did not prevent T cell–induced mucosal damage. However,3 d after anti-CD3 mAb injection, both antibiotic-treated WT andNOD22/2 mice showed significantly reduced mucosal damagecompared with water control (Fig. 5H–J). Moreover, IL-17A, IFN-g,and MPO levels in the small intestine of NOD22/2 mice were re-duced to the level of WT mice at day 3 (Fig. 5K). These data col-lectively indicate that gut microbial load is important in determining
FIGURE 1. Increase of mucosal damage and loss of small intestinal crypts in NOD22/2 mice after acute T cell activation. WT and NOD22/2 mice
were i.p. injected with anti-CD3 mAb and were sacrificed at various time points. (A) Representative photographs at original magnification 310 of
H&E-stained sections from the small bowel at days 0, 1, 3, and 5 after anti-CD3 mAb. (B) Depicted are scores for small intestinal crypt damage at days
0, 1, 3, and 5 after anti-CD3 mAb (n = 8–14 mice). (C) Depicted are measures for villus height at days 0, 1, 3, and 5 after anti-CD3 mAb (n = 8–14
mice). *p # 0.05, **p # 0.01.
348 Nod2 REGULATES MUCOSAL DAMAGE IN THE SMALL INTESTINE
by guest on October 31, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
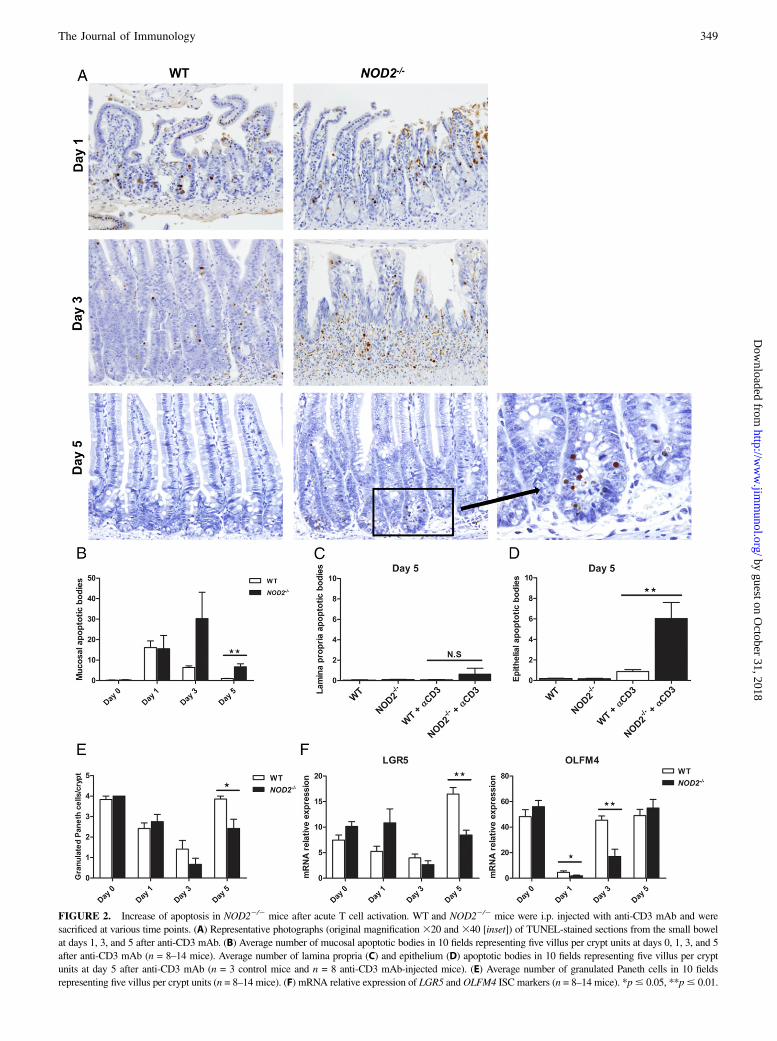
FIGURE 2. Increase of apoptosis in NOD22/2 mice after acute T cell activation. WT and NOD22/2 mice were i.p. injected with anti-CD3 mAb and were
sacrificed at various time points. (A) Representative photographs (original magnification 320 and 340 [inset]) of TUNEL-stained sections from the small bowel
at days 1, 3, and 5 after anti-CD3 mAb. (B) Average number of mucosal apoptotic bodies in 10 fields representing five villus per crypt units at days 0, 1, 3, and 5
after anti-CD3 mAb (n = 8–14 mice). Average number of lamina propria (C) and epithelium (D) apoptotic bodies in 10 fields representing five villus per crypt
units at day 5 after anti-CD3 mAb (n = 3 control mice and n = 8 anti-CD3 mAb-injected mice). (E) Average number of granulated Paneth cells in 10 fields
representing five villus per crypt units (n = 8–14 mice). (F) mRNA relative expression of LGR5 andOLFM4 ISC markers (n = 8–14 mice). *p# 0.05, **p# 0.01.
The Journal of Immunology 349
by guest on October 31, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
the degree of damage and the delay in recovery. In this regard,microbial sensing by Nod2 is a critical mechanism in controllingexcessive crypt damage.
Selective deletion of Nod2 in intestinal epithelial cells or inLyz2-expressing phagocytes is not sufficient to recapitulate themucosal damage observed in NOD22/2 mice
To determine whether epithelial cell intrinsic Nod2 regulates mu-cosal damage and epithelial regeneration after T cell-induced en-teropathy, we generated mice lacking NOD2 in villin-expressingcells (NOD2ΔIEC mice). Absence of NOD2 expression in IEC fromNOD2ΔIEC mice was controlled by PCR (Supplemental Fig. 1). Ourresults showed that NOD2ΔIEC mice developed comparable cryptdamage and IEC apoptosis with NOD2Flox (WT) mice after anti-CD3 treatment (Fig. 6A–C). In addition, the secretion of IL-17A,IFN-g, and MPO, and the mRNA expression of the ISC markersLGR5 and OLFM4 were not different between NOD2Flox andNOD2ΔIEC mice (data not shown). These data indicate that deletionof Nod2 in IEC is not sufficient to increase the sensitivity toT cell–induced enteropathy and delay recovery, suggesting that Nod2expression outside the epithelium is important in regulating the re-sponse to T cell–induced mucosal damage.Because of the higher number of apoptotic cells in NOD22/2
mice and the importance of macrophages and neutrophils inphagocytosing dead cells, cellular debris, and invading microor-ganisms, we hypothesized that Nod2 expression in phagocytesmay be important in controlling anti-CD3–induced enteropathy,and we generated mice lacking NOD2 in Lyz2-expressing cells
(NOD2ΔLyz2 mice). Deletion of NOD2 in Lyz2-expressing cellswas assessed by the decreased recruitment of neutrophils in theperitoneal cavity of NOD2ΔLyz2 mice after i.p. injection of theNod2 ligand MDP (Supplemental Fig. 1). Although, crypt damagewas not different between NOD2Flox and NOD2ΔLyz2 mice, wefound higher numbers of apoptotic IEC in NOD2ΔLyz2 mice (Fig.6D–F). The secretion of IL-17A, IFN-g, and MPO, and the mRNAexpression of LGR5 and OLFM4 was not different betweenNOD2Flox and NOD2ΔLyz2 mice (data not shown). However, wedid find a significant increase of TNF-a and IL-22 expression inNOD2ΔLyz2 mice (Fig. 6G), indicating that Nod2 deletion inphagocytes can affect TNF-a level and IEC viability in the smallintestine.
DiscussionThe requirement of T cells in various animal models of intestinalinflammation (12) and the increase in T cell–derived inflammatorycytokines in the mucosa of inflammatory bowel disease (IBD)patients (17) suggest that T cells play important roles in thepathogenesis of IBD. Our study demonstrates that following smallintestinal mucosal damage induced by T cell activation, micro-biota sensing by Nod2 is important for controlling immune acti-vation, crypt damage, and epithelial regeneration.Looking at a direct role of Nod2 in T cell regulation of colitis, our
group previously showed that the expression of Nod2 in murineCD4+ T cells is not required for the development or the preventionof T cell–induced colitis in Rag-deficient mice (6). In addition,NOD2+/+and NOD22/2 littermate control mice have been shown
FIGURE 3. Delayed epithelial regeneration in NOD22/2 mice after acute T cell activation. (A) Representative photographs at original magnification310
of Ki67+-stained sections from the small bowel at days 0, 3, and 5 after anti-CD3 mAb. (B) Average number of Ki67+ epithelial cells in 10 crypts at days 0,
3, and 5 after anti-CD3 mAb (n = 8–14 mice). **p # 0.01, ***p # 0.001.
350 Nod2 REGULATES MUCOSAL DAMAGE IN THE SMALL INTESTINE
by guest on October 31, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
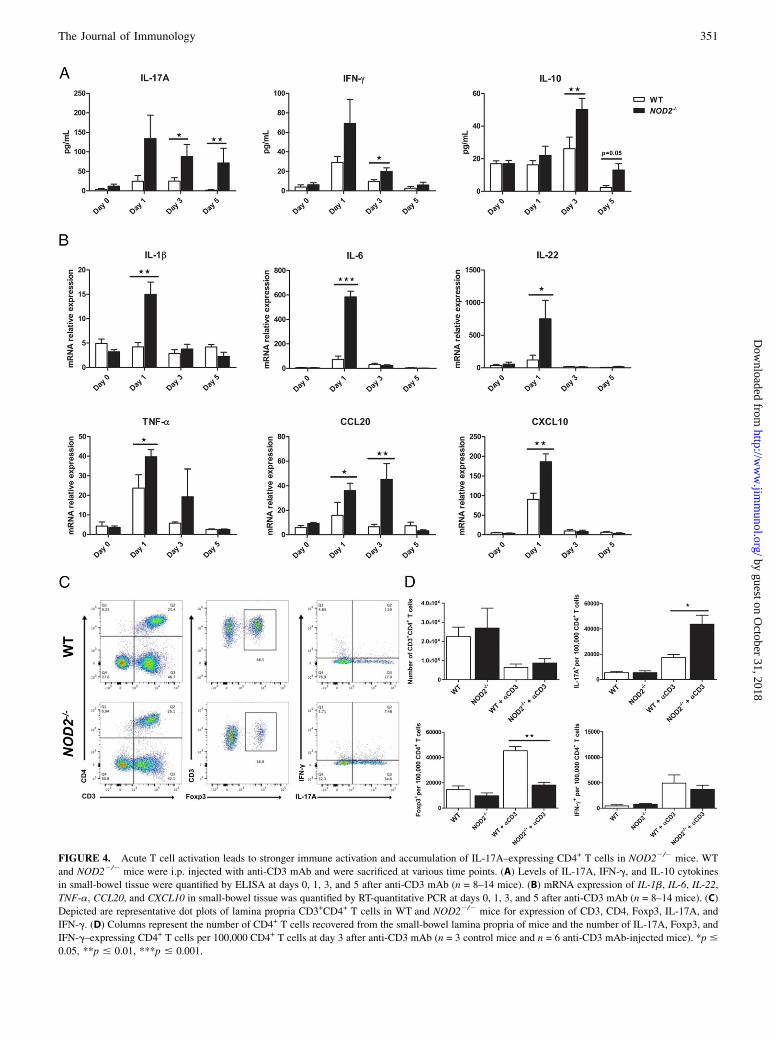
FIGURE 4. Acute T cell activation leads to stronger immune activation and accumulation of IL-17A–expressing CD4+ T cells in NOD22/2 mice. WT
and NOD22/2 mice were i.p. injected with anti-CD3 mAb and were sacrificed at various time points. (A) Levels of IL-17A, IFN-g, and IL-10 cytokines
in small-bowel tissue were quantified by ELISA at days 0, 1, 3, and 5 after anti-CD3 mAb (n = 8–14 mice). (B) mRNA expression of IL-1b, IL-6, IL-22,
TNF-a, CCL20, and CXCL10 in small-bowel tissue was quantified by RT-quantitative PCR at days 0, 1, 3, and 5 after anti-CD3 mAb (n = 8–14 mice). (C)
Depicted are representative dot plots of lamina propria CD3+CD4+ T cells in WT and NOD22/2 mice for expression of CD3, CD4, Foxp3, IL-17A, and
IFN-g. (D) Columns represent the number of CD4+ T cells recovered from the small-bowel lamina propria of mice and the number of IL-17A, Foxp3, and
IFN-g–expressing CD4+ T cells per 100,000 CD4+ T cells at day 3 after anti-CD3 mAb (n = 3 control mice and n = 6 anti-CD3 mAb-injected mice). *p #
0.05, **p # 0.01, ***p # 0.001.
The Journal of Immunology 351
by guest on October 31, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
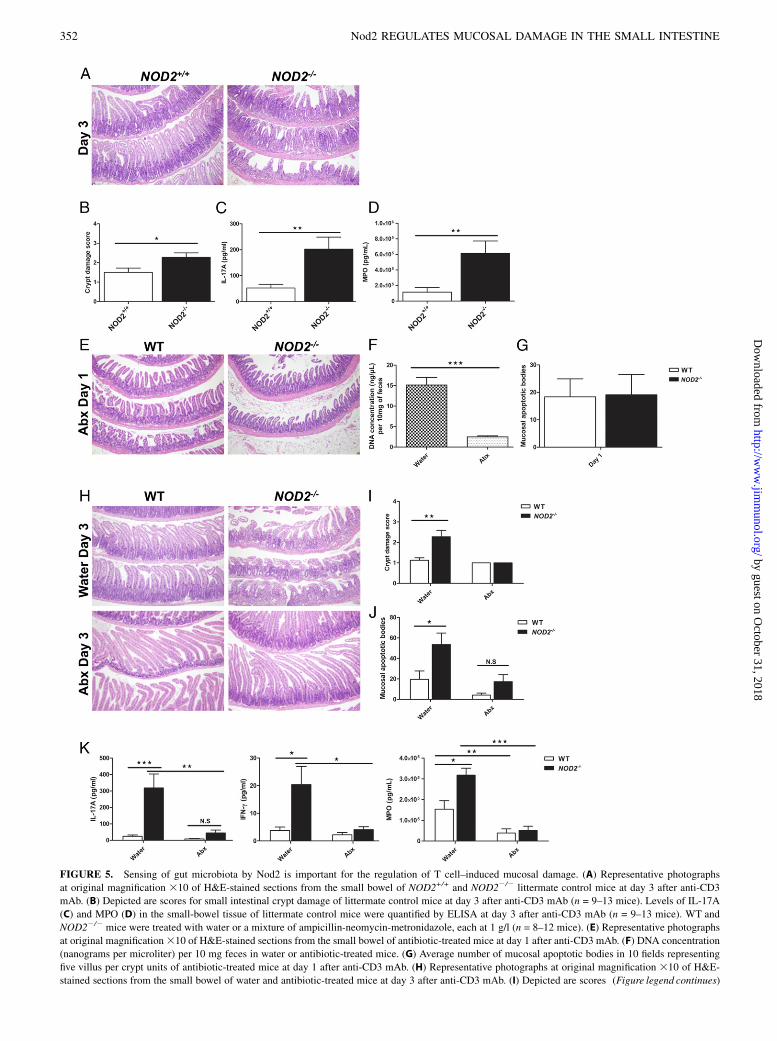
FIGURE 5. Sensing of gut microbiota by Nod2 is important for the regulation of T cell–induced mucosal damage. (A) Representative photographs
at original magnification 310 of H&E-stained sections from the small bowel of NOD2+/+ and NOD22/2 littermate control mice at day 3 after anti-CD3
mAb. (B) Depicted are scores for small intestinal crypt damage of littermate control mice at day 3 after anti-CD3 mAb (n = 9–13 mice). Levels of IL-17A
(C) and MPO (D) in the small-bowel tissue of littermate control mice were quantified by ELISA at day 3 after anti-CD3 mAb (n = 9–13 mice). WT and
NOD22/2 mice were treated with water or a mixture of ampicillin-neomycin-metronidazole, each at 1 g/l (n = 8–12 mice). (E) Representative photographs
at original magnification310 of H&E-stained sections from the small bowel of antibiotic-treated mice at day 1 after anti-CD3 mAb. (F) DNA concentration
(nanograms per microliter) per 10 mg feces in water or antibiotic-treated mice. (G) Average number of mucosal apoptotic bodies in 10 fields representing
five villus per crypt units of antibiotic-treated mice at day 1 after anti-CD3 mAb. (H) Representative photographs at original magnification 310 of H&E-
stained sections from the small bowel of water and antibiotic-treated mice at day 3 after anti-CD3 mAb. (I) Depicted are scores (Figure legend continues)
352 Nod2 REGULATES MUCOSAL DAMAGE IN THE SMALL INTESTINE
by guest on October 31, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

to present the same sensitivity to dextran sulfate sodium–inducedcolitis (18). These results and the association between Nod2mutations and ileal CD suggest a more specific role of Nod2 in theregulation of small intestinal homeostasis. The model of anti-CD3mAb i.p. injection was developed to study T cell driven inflam-matory diseases such as IBD, celiac disease, or graft-versus-hostdisease (13). In mice, acute T cell activation following i.p. in-jection of T cell–activating anti-CD3 mAb leads to severe smallintestinal mucosal damage characterized by villus blunting andapoptosis of IEC (13). Therefore, i.p. injection of anti-CD3 mAbis a helpful model to study the role of Nod2 in regulating T cell–induced small intestinal mucosal damage.
The inability to control inflammation and to regenerate the in-testinal epithelial barrier is a common characteristic of IBD. Re-generation of the epithelium is a highly regulated process that isorchestrated by multipotent stem cells located in the bottom of thecrypts (19, 20). Study of ISC biology has been facilitated by theidentification of ISC markers such as LGR5 and OLFM4 (21, 22).Nod2 is highly expressed in ISC and has an important function inISC survival (8). Our data showed that the delayed epithelial re-generation in NOD22/2 mice is associated with an increase ofcrypt IEC apoptosis and a decrease of LGR5 and OLFM4 ISCmarker expression, suggesting that Nod2-deficient ISC may bemore sensitive to apoptosis induced by anti-CD3 mAb injection.
for small intestinal crypt damage of water and antibiotic-treated mice at day 3 post anti-CD3 mAb. (J) Average number of mucosal apoptotic bodies in 10
fields representing five villus per crypt units of water and antibiotic-treated mice at day 3 after anti-CD3 mAb. (K) Levels of IL-17A, IFN-g, and MPO in the
small-bowel tissue of water and antibiotic-treated mice at day 3 after anti-CD3 mAb. *p # 0.05, **p # 0.01, ***p # 0.001.
FIGURE 6. Deletion of Nod2 in intestinal epithelial cells or Lyz2-expressing phagocytes does not recapitulate mucosal damage induced by acute T cell
activation in NOD22/2 mice. NOD2Flox, NOD2ΔIEC, and NOD2ΔLyz2 mice were i.p. injected with anti-CD3 mAb and were sacrificed at day 3. (A) Representative
photographs at original magnification 310 of H&E-stained sections from the small bowel of NOD2Flox and NOD2ΔIEC. (B) Depicted are scores for small in-
testinal crypt damage (n = 6 mice). (C) Average number of epithelial apoptotic bodies in 10 fields representing five villus per crypt units (n = 6 mice). (D)
Representative photographs at original magnification 310 of H&E-stained sections from the small bowel of NOD2Flox and NOD2ΔLyz2. (E) Depicted are scores
for small intestinal crypt damage (n = 10–11 mice). (F) Average number of epithelial apoptotic bodies in 10 fields representing five villus per crypt units (n = 10–
11 mice). (G) mRNA expression of TNF-a and IL-22 in small-bowel tissue was quantified by RT-quantitative PCR (n = 10–11 mice). *p # 0.05, **p # 0.01.
The Journal of Immunology 353
by guest on October 31, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

In addition, we found a more severe loss of granulated Paneth cellnumber in NOD22/2 mice. Nod2 is expressed in Paneth cells (4),and secretion of IFN-g by immune cells has been shown to inducePaneth cell degranulation and death (23), suggesting that the in-crease of IFN-g level in NOD22/2 mice can affect Paneth cellnumber and function.Intraperitoneal injection of mice with anti-CD3 mAb has been
used to study the role of Nod2 on T cell activation and function. In amurine model of low-dose (15 mg) i.p. injection of anti-CD3 mAbat days 0 and 2, Nod2 deletion has been shown to decreaseCXCL9/CXCL10-dependent recruitment of CD8+ T cells in thesmall intestinal lamina propria and the subsequent expression ofIL-10 by CD8+ T cells (24). In addition, the authors found a de-crease in IL-17A and IFN-g mRNA expression in the small in-testine of NOD22/2 mice (24). In our study, we found that a singlehigh-dose (50 mg) i.p. injection of anti-CD3 mAb induced higherlevels of IL-17A, IFN-g, IL-10, and CXCL10 in the small intestineof NOD22/2 mice compared with WT mice. These opposite resultsmay indicate that the intensity of T cell activation and the degree ofmucosal damage can strongly modulate the immune response.Another possibility is that the use of different NOD22/2 mice strainleads to contradictory results regarding the role of Nod2 (6, 25, 26).Yet in another model, repeated i.p. injection of 20 mg anti-CD3
mAb promoted generation and accumulation of Th17 cells withinthe small intestine of WT mice (16). In this model, Th17 cellgeneration was dependent on IL-6 secretion from macrophages,and their accumulation within the small intestine was triggered bythe CCR6–CCL20 axis. In our data, we observed a significantincrease of IL-6, CCL20 expression, IL-17A secretion, and IL-17A–expressing T cell accumulation within the small intestine ofNOD22/2 mice. After antibiotic treatment, the secretion of IL-17A, IFN-g, and MPO was decreased to the level of WT mice. Inaddition, antibiotic-treated NOD22/2 mice did not display themarked loss of small intestinal crypts seen in untreated NOD22/2
mice, highlighting the role of the microbiota in perpetuatingmucosal damage induced by acute T cell activation in NOD22/2
mice. Indeed, Nod2-deficient mice show increased sensitivity toseveral microbial pathogens (27–30). The altered response to injuryand infection that is seen in NOD22/2 mice and the development ofileal disease in CD patients carrying NOD2 risk alleles are con-sistent with our results supporting the importance of microbialsensing by Nod2 in the regulation of small-intestine homeostasis.CD is associated with structural damage and destruction of the
bowel wall (31). Epithelial regeneration is a crucial step in mu-cosal healing and this mechanism requires the coordinated activityof IEC, Paneth cells, and goblet cells to restore intestinal barrierfunction (32–34). Mice carrying both Nod1 and Nod2 deletionhave decreased paracellular permeability and a higher suscepti-bility to dextran sulfate sodium–induced colitis, highlighting apotential role of Nod receptors in the maintenance of intestinalbarrier function (35). Moreover, another study reported a newfunction for Nod2 in affording ISC survival and epithelial re-generation (8). The authors showed that stimulation with MDPenhanced in vitro intestinal organoid survival and in vivo stem cellsurvival following doxorubicin treatment in mice (8). With regardsto T cell–induced mucosal damage, our data has identified theimportance of Nod2 signaling in sustaining crypt architecture andepithelial regeneration and suggests that apoptotic epithelial cellsobserved in the small intestinal crypts of NOD22/2 mice mightbe ISC. Surprisingly, we found that Nod2 deletion in IEC did notexacerbate mucosal damage, suggesting that Nod2 function in othercellular compartments regulates intestinal homeostasis (7, 36–38).Resolution of acute inflammation requires the elimination of
invading microorganisms, apoptotic cells, and cellular debris from
the intestinal lamina propria. This mechanism is achieved via therecruitment of professional phagocytes (39, 40). Stimulation ofNod2 in human monocyte–derived macrophages and in mouseintestinal macrophages increases autophagy and bacterial clear-ance (41). We described that i.p. injection of anti-CD3 mAb in-duced higher numbers of mucosal apoptotic bodies in the smallintestinal lamina propria of NOD22/2 mice. In addition, disruptionof the intestinal epithelial barrier was associated with a microbiota-dependent perpetuation of mucosal damage in NOD22/2 mice.These results suggest that Nod2 deletion in macrophages can alterbacterial clearance and consequently the mucosal damage inducedby anti-CD3 mAb injection. We found that deletion of Nod2 inLyz2-expressing cells did not recapitulate the mucosal damageobserved in NOD22/2 mice. Indeed, the shortening and the lossof small intestinal crypts were comparable between WT andNOD2ΔLyz2 mice. However, NOD2ΔLyz2 mice showed highernumbers of epithelial apoptotic bodies and increased expressionof TNF-a and IL-22 compared with WT mice. TNF-a plays acrucial role in the pathogenesis of CD and high levels of TNF-acan alter intestinal epithelial barrier integrity (42–44). Chronicstimulation of Nod2 in human monocyte–derived macrophageshas been associated with a downregulation of TNF-a secretion(45), indicating that expression of Nod2 is important for both theinduction and the regulation of this response. These results suggestthat deletion of Nod2 in Lyz2-expressing cells can trigger higherlevels of TNF-a, thus inducing a stronger IEC apoptosis and theincreased expression of IL-22, a cytokine involved in gut barrierintegrity and epithelial regeneration (46, 47).In conclusion, we describe a role for Nod2 in regulating small
intestinal crypt damage and epithelial regeneration after acuteT cell activation. Understanding the function of Nod2 in mucosalhealing will help define the pathophysiology of inflammatorydiseases of the gut such as IBD.
AcknowledgmentsWe thank Dr. Philip Rosenstiel (Institute for Clinical Molecular Biology,
Christian-Albrechts-University, Kiel, Germany) for providing NOD2-
floxed mice.
DisclosuresThe authors have no financial conflicts of interest.
References1. Brown, E. M., M. Sadarangani, and B. B. Finlay. 2013. The role of the immune
system in governing host-microbe interactions in the intestine. Nat. Immunol. 14:660–667.
2. Barnich, N., J. E. Aguirre, H. C. Reinecker, R. Xavier, and D. K. Podolsky. 2005.Membrane recruitment of NOD2 in intestinal epithelial cells is essential fornuclear factor-kB activation in muramyl dipeptide recognition. J. Cell Biol. 170:21–26.
3. Gutierrez, O., C. Pipaon, N. Inohara, A. Fontalba, Y. Ogura, F. Prosper,G. Nunez, and J. L. Fernandez-Luna. 2002. Induction of Nod2 in myelomono-cytic and intestinal epithelial cells via nuclear factor-kB activation. J. Biol.Chem. 277: 41701–41705.
4. Ogura, Y., S. Lala, W. Xin, E. Smith, T. A. Dowds, F. F. Chen, E. Zimmermann,M. Tretiakova, J. H. Cho, J. Hart, et al. 2003. Expression of NOD2 in Panethcells: a possible link to Crohn’s ileitis. Gut 52: 1591–1597.
5. Takahashi, Y., K. Isuzugawa, Y. Murase, M. Imai, S. Yamamoto, M. Iizuka,S. Akira, G. M. Bahr, E. Momotani, M. Hori, et al. 2006. Up-regulation of NOD1and NOD2 through TLR4 and TNF-a in LPS-treated murine macrophages.J. Vet. Med. Sci. 68: 471–478.
6. Zanello, G., A. Goethel, K. Forster, K. Geddes, D. J. Philpott, and K. Croitoru.2013. Nod2 activates NF-kB in CD4+ T cells but its expression is dispensable forT cell‑induced colitis. PLoS One 8: e82623.
7. Philpott, D. J., M. T. Sorbara, S. J. Robertson, K. Croitoru, and S. E. Girardin.2014. NOD proteins: regulators of inflammation in health and disease. Nat. Rev.Immunol. 14: 9–23.
8. Nigro, G., R. Rossi, P. H. Commere, P. Jay, and P. J. Sansonetti. 2014. Thecytosolic bacterial peptidoglycan sensor Nod2 affords stem cell protection andlinks microbes to gut epithelial regeneration. Cell Host Microbe 15: 792–798.
354 Nod2 REGULATES MUCOSAL DAMAGE IN THE SMALL INTESTINE
by guest on October 31, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

9. Hugot, J. P., M. Chamaillard, H. Zouali, S. Lesage, J. P. Cezard, J. Belaiche, S. Almer,C. Tysk, C. A. O’Morain, M. Gassull, et al. 2001. Association of NOD2 leucine-richrepeat variants with susceptibility to Crohn’s disease. Nature 411: 599–603.
10. Ogura, Y., D. K. Bonen, N. Inohara, D. L. Nicolae, F. F. Chen, R. Ramos, H. Britton,T. Moran, R. Karaliuskas, R. H. Duerr, et al. 2001. A frameshift mutation in NOD2associated with susceptibility to Crohn’s disease. Nature 411: 603–606.
11. Bonen, D. K., and J. H. Cho. 2003. The genetics of inflammatory bowel disease.Gastroenterology 124: 521–536.
12. Shale, M., C. Schiering, and F. Powrie. 2013. CD4+ T-cell subsets in intestinalinflammation. Immunol. Rev. 252: 164–182.
13. Merger, M., J. L. Viney, R. Borojevic, D. Steele-Norwood, P. Zhou, D. A. Clark,R. Riddell, R. Maric, E. R. Podack, and K. Croitoru. 2002. Defining the roles ofperforin, Fas/FasL, and tumour necrosis factor a in T cell induced mucosaldamage in the mouse intestine. Gut 51: 155–163.
14. Barreau, F., U. Meinzer, F. Chareyre, D. Berrebi, M. Niwa-Kawakita, M. Dussaillant,B. Foligne, V. Ollendorff, M. Heyman, S. Bonacorsi, et al. 2007. CARD15/NOD2 isrequired for Peyer’s patches homeostasis in mice. PLoS One 2: e523.
15. Kim, D., Y. G. Kim, S. U. Seo, D. J. Kim, N. Kamada, D. Prescott, D. J. Philpott,P. Rosenstiel, N. Inohara, and G. Nunez. 2016. Nod2-mediated recognition of themicrobiota is critical for mucosal adjuvant activity of cholera toxin. Nat. Med.22: 524–530.
16. Esplugues, E., S. Huber, N. Gagliani, A. E. Hauser, T. Town, Y. Y. Wan,W. O’Connor, Jr., A. Rongvaux, N. Van Rooijen, A. M. Haberman, et al. 2011.Control of TH17 cells occurs in the small intestine. Nature 475: 514–518.
17. Neurath, M. F. 2014. Cytokines in inflammatory bowel disease. Nat. Rev.Immunol. 14: 329–342.
18. Smith, P., J. Siddharth, R. Pearson, N. Holway, M. Shaxted, M. Butler, N. Clark,J. Jamontt, R. P. Watson, D. Sanmugalingam, and S. J. Parkinson. 2012. Hostgenetics and environmental factors regulate ecological succession of the mousecolon tissue-associated microbiota. PLoS One 7: e30273.
19. Barker, N. 2014. Adult intestinal stem cells: critical drivers of epithelial ho-meostasis and regeneration. Nat. Rev. Mol. Cell Biol. 15: 19–33.
20. Tan, D. W., and N. Barker. 2014. Intestinal stem cells and their defining niche.Curr. Top. Dev. Biol. 107: 77–107.
21. Barker, N., J. H. van Es, J. Kuipers, P. Kujala, M. van den Born, M. Cozijnsen,A. Haegebarth, J. Korving, H. Begthel, P. J. Peters, and H. Clevers. 2007. Identificationof stem cells in small intestine and colon bymarker gene Lgr5.Nature 449: 1003–1007.
22. Schuijers, J., L. G. van der Flier, J. van Es, and H. Clevers. 2014. Robust cre-mediated recombination in small intestinal stem cells utilizing the olfm4 locus.Stem Cell Rep. 3: 234–241.
23. Farin, H. F., W. R. Karthaus, P. Kujala, M. Rakhshandehroo, G. Schwank,R. G. Vries, E. Kalkhoven, E. E. Nieuwenhuis, and H. Clevers. 2014. Paneth cellextrusion and release of antimicrobial products is directly controlled by immunecell-derived IFN-g. J. Exp. Med. 211: 1393–1405.
24. Wu, X., A. Lahiri, G. K. Haines, III, R. A. Flavell, and C. Abraham. 2014. NOD2regulates CXCR3-dependent CD8+ T cell accumulation in intestinal tissues withacute injury. J. Immunol. 192: 3409–3418.
25. Caetano, B. C., A. Biswas, D. S. Lima, Jr., L. Benevides, T. W. Mineo,C. V. Horta, K. H. Lee, J. S. Silva, R. T. Gazzinelli, D. S. Zamboni, andK. S. Kobayashi. 2011. Intrinsic expression of Nod2 in CD4+ T lymphocytes isnot necessary for the development of cell-mediated immunity and host resistanceto Toxoplasma gondii. Eur. J. Immunol. 41: 3627–3631.
26. Shaw, M. H., T. Reimer, C. Sanchez-Valdepenas, N. Warner, Y. G. Kim,M. Fresno, and G. Nunez. 2009. T cell‑intrinsic role of Nod2 in promoting type 1immunity to Toxoplasma gondii. Nat. Immunol. 10: 1267–1274.
27. Chassaing, B., N. Rolhion, A. de Vallee, S. Y. Salim, M. Prorok-Hamon, C. Neut,B. J. Campbell, J. D. Soderholm, J. P. Hugot, J. F. Colombel, and A. Darfeuille-Michaud. 2011. Crohn disease‑associated adherent-invasive E. coli bacteria target mouseand human Peyer’s patches via long polar fimbriae. J. Clin. Invest. 121: 966–975.
28. Frutuoso, M. S., J. I. Hori, M. S. Pereira, D. S. Junior, F. Sonego, K. S. Kobayashi,R. A. Flavell, F. Q. Cunha, and D. S. Zamboni. 2010. The pattern recognitionreceptors Nod1 and Nod2 account for neutrophil recruitment to the lungs of miceinfected with Legionella pneumophila. Microbes Infect. 12: 819–827.
29. Geddes, K., S. J. Rubino, J. G. Magalhaes, C. Streutker, L. Le Bourhis,J. H. Cho, S. J. Robertson, C. J. Kim, R. Kaul, D. J. Philpott, and S. E. Girardin.2011. Identification of an innate T helper type 17 response to intestinal bacterialpathogens. Nat. Med. 17: 837–844.
30. Hruz, P., A. S. Zinkernagel, G. Jenikova, G. J. Botwin, J. P. Hugot, M. Karin,V. Nizet, and L. Eckmann. 2009. NOD2 contributes to cutaneous defense againstStaphylococcus aureus through a-toxin-dependent innate immune activation.Proc. Natl. Acad. Sci. USA 106: 12873–12878.
31. Neurath, M. F. 2014. New targets for mucosal healing and therapy in inflam-matory bowel diseases. Mucosal Immunol. 7: 6–19.
32. Krishnan, K., B. Arnone, and A. Buchman. 2011. Intestinal growth factors:potential use in the treatment of inflammatory bowel disease and their role inmucosal healing. Inflamm. Bowel Dis. 17: 410–422.
33. Paclik, D., K. Lohse, B. Wiedenmann, A. U. Dignass, and A. Sturm. 2008.Galectin-2 and -4, but not galectin-1, promote intestinal epithelial wound healingin vitro through a TGF-b‑independent mechanism. Inflamm. Bowel Dis. 14:1366–1372.
34. Sturm, A., and A. U. Dignass. 2008. Epithelial restitution and wound healing ininflammatory bowel disease. World J. Gastroenterol. 14: 348–353.
35. Natividad, J. M., V. Petit, X. Huang, G. de Palma, J. Jury, Y. Sanz, D. Philpott,C. L. Garcia Rodenas, K. D. McCoy, and E. F. Verdu. 2012. Commensal andprobiotic bacteria influence intestinal barrier function and susceptibility to colitisin Nod1‑/‑;Nod2‑/‑ mice. Inflamm. Bowel Dis. 18: 1434–1446.
36. Cooney, R., J. Baker, O. Brain, B. Danis, T. Pichulik, P. Allan, D. J. Ferguson,B. J. Campbell, D. Jewell, and A. Simmons. 2010. NOD2 stimulation inducesautophagy in dendritic cells influencing bacterial handling and antigen presen-tation. Nat. Med. 16: 90–97.
37. Homer, C. R., A. L. Richmond, N. A. Rebert, J. P. Achkar, and C. McDonald.2010. ATG16L1 and NOD2 interact in an autophagy-dependent antibacterialpathway implicated in Crohn’s disease pathogenesis. Gastroenterology 139:1630‑1641, 1641 e1631-1632. doi:10.1053/j.gastro.2010.07.006
38. Travassos, L. H., L. A. Carneiro, M. Ramjeet, S. Hussey, Y. G. Kim,J. G. Magalhaes, L. Yuan, F. Soares, E. Chea, L. Le Bourhis, et al. 2010. Nod1and Nod2 direct autophagy by recruiting ATG16L1 to the plasma membrane atthe site of bacterial entry. Nat. Immunol. 11: 55–62.
39. Bain, C. C., and A. M. Mowat. 2014. Macrophages in intestinal homeostasis andinflammation. Immunol. Rev. 260: 102–117.
40. Scott, C. L., S. Henri, and M. Guilliams. 2014. Mononuclear phagocytes of theintestine, the skin, and the lung. Immunol. Rev. 262: 9–24.
41. Lahiri, A., and C. Abraham. 2014. Activation of pattern recognition receptorsup-regulates metallothioneins, thereby increasing intracellular accumulation ofzinc, autophagy, and bacterial clearance by macrophages. Gastroenterology 147:835–846.
42. Fries, W., C. Muja, C. Crisafulli, G. Costantino, G. Longo, S. Cuzzocrea,and E. Mazzon. 2008. Infliximab and etanercept are equally effective inreducing enterocyte APOPTOSIS in experimental colitis. Int. J. Med. Sci. 5:169–180.
43. Goretsky, T., R. Dirisina, P. Sinh, N. Mittal, E. Managlia, D. B. Williams,D. Posca, H. Ryu, R. B. Katzman, and T. A. Barrett. 2012. p53 mediates TNF-induced epithelial cell apoptosis in IBD. Am. J. Pathol. 181: 1306–1315.
44. Watson, A. J., and K. R. Hughes. 2012. TNF-a‑induced intestinal epithelial cellshedding: implications for intestinal barrier function. Ann. N. Y. Acad. Sci. 1258:1–8.
45. Hedl, M., J. Li, J. H. Cho, and C. Abraham. 2007. Chronic stimulation of Nod2mediates tolerance to bacterial products. Proc. Natl. Acad. Sci. USA 104: 19440–19445.
46. Lindemans, C. A., M. Calafiore, A. M. Mertelsmann, M. H. O’Connor,J. A. Dudakov, R. R. Jenq, E. Velardi, L. F. Young, O. M. Smith, G. Lawrence,et al. 2015. Interleukin-22 promotes intestinal-stem-cell-mediated epithelial re-generation. Nature 528: 560–564.
47. Zenewicz, L. A., G. D. Yancopoulos, D. M. Valenzuela, A. J. Murphy,S. Stevens, and R. A. Flavell. 2008. Innate and adaptive interleukin-22 protectsmice from inflammatory bowel disease. Immunity 29: 947–957.
The Journal of Immunology 355
by guest on October 31, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from





![php [Mode de compatibilité]croitoru/php.pdf · Practicals Th d f l tThe password form element](https://img.pdfslide.us/doc/110x75/5b893be77f8b9abe1e8d25bf/php-mode-de-compatibilite-croitoruphppdf-practicals-th-d-f-l-tthe-password.jpg)























