Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY,0021-9193/98/$04.0010
Aug. 1998, p. 4116–4122 Vol. 180, No. 16
Copyright © 1998, American Society for Microbiology. All Rights Reserved.
The Cyanobacterium Synechocystis sp. Strain PCC 6803 Expressesa DNA Methyltransferase Specific for the Recognition
Sequence of the Restriction Endonuclease PvuIMATTHIAS SCHARNAGL,1 STEFAN RICHTER,2† AND MARTIN HAGEMANN1*
FB Biologie, Universitat Rostock, D-18051 Rostock,1 and Max-Planck-Institutfur Molekulare Genetik, D-14195 Berlin,2 Germany
Received 2 March 1998/Accepted 30 May 1998
By use of restriction endonucleases, the DNA of the cyanobacterium Synechocystis sp. strain PCC 6803 wasanalyzed for DNA-specific methylation. Three different recognition sites of methyltransferases, a dam-like siteincluding N6-methyladenosine and two other sites with methylcytosine, were identified, whereas no activities ofrestriction endonucleases could be detected in this strain. slr0214, a Synechocystis gene encoding a putative meth-yltransferase that shows significant similarities to C5-methylcytosine-synthesizing enzymes, was amplified byPCR and cloned for further characterization. Mutations in slr0214 were generated by the insertion of an aphIIgene cassette. Analyses of chromosomal DNAs of such mutants demonstrated that the methylation pattern waschanged. The recognition sequence of the methyltransferase was identified as 5*-CGATCG-3*, correspondingto the recognition sequence of PvuI. The specific methyltransferase activity was significantly reduced in proteinextracts obtained from mutant cells. Mutation of slr0214 also led to changed growth characteristics of the cellscompared to wild-type cells. These alterations led to the conclusion that the methyltransferase Slr0214 mightplay a regulatory role in Synechocystis. The Slr0214 protein was also overexpressed in Escherichia coli, and thepurified protein demonstrated methyltransferase activity and specificity for PvuI recognition sequences invitro. We propose the designation SynMI (Synechocystis methyltransferase I) for the slr0214-encoded enzyme.
DNA of prokaryotic and eukaryotic cells is usually modifiedby methylation. This modification is carried out by DNA-spe-cific methyltransferases, which transfer methyl groups from theuniversal substrate S-adenosylmethionine (AdoMet) to spe-cific target sequences in the host DNA. On the basis of differ-ent chemical reactions, methyltransferases can be divided intothree different groups leading to the appearance of N6-methyl-adenosine, C5-methylcytosine, or C4-methylcytosine. Amino acidsequence alignments of enzymes from these groups revealedconserved motifs characteristic of each methyltransferase fam-ily (25). In the enteric bacterium Escherichia coli, three methyl-transferases are present: (i) Dam (DNA adenine methyltrans-ferase), creating N6-methyladenosine in the specific targetsequence 59-GmATC-39; (ii) Dcm (DNA cytosine methyltrans-ferase), leading to an internal C5-methylcytosine in the specifictarget sequence 59-CmC(A/T)GG-39; and (iii) methyltransfer-ase M z EcoK, modifying the second adenine in the sequenceAmAC(N6)GTCG, which is specifically recognized as an un-methylated sequence by restriction endonuclease EcoK, whichis part of the host-specific restriction-modification (R-M) sys-tem (for a review, see reference 22). Methylation of cytosine inCG dinucleotides dominates in cells of higher eukaryotes. Inthese cells, the degree of methylation is tightly controlled andplays an important role in the regulation of gene expression,organization of chromatin, and developmental and DNA re-pair processes (4).
Methyltransferases involved in R-M systems have beenfound in all taxonomic groups of eubacteria examined so far.
R-M systems typically comprise two enzymatic activities, arestriction endonuclease and a methyltransferase. The genes ofboth enzymes are often organized in one operon. By extensivescreening, more than 2,750 type II restriction endonucleaseswith a total of 211 different specificities have been found (27).Cyanobacteria represent a rich source of restriction enzymes.In several strains, two or three different enzymes are present;among them, isoschizomers of the enzyme AvaII have beenoften found (10). Six restriction endonucleases with differentspecificities were detected in the cyanobacterium Dactylococ-copsis salina (19). The cognate methyltransferases have beeninvestigated less. The genes encoding AvaI and AquI methyl-transferases were cloned and sequenced from Anabaena varia-bilis and Agmenellum quadruplicatum, respectively. The aminoacid sequences of both proteins show significant similarities tothose of functionally related enzymes from heterotrophic bac-teria (13, 30).
Genetic tools are well developed only for a few cyanobacte-rial model strains. A general problem in establishing suchsystems for other strains is the barriers made by endogenousrestriction systems that prevent stable access of foreign DNAinto the cells. As has been found before in many instances, invitro methylation of DNA also improves the efficiency of DNAtransfer into cyanobacterial strains (35). The cyanobacteriumSynechocystis sp. strain PCC 6803 (hereafter referred to as Syn-echocystis) belongs to the group of naturally competent strainswhich are transformable by free exogenous DNA (6). A Ca21-dependent nuclease located in the cytoplasmic membrane isinvolved in DNA uptake and is assumed to convert double-stranded DNA into the single-stranded form (3). Recently, theentire genome of Synechocystis was sequenced, making thisstrain the favored cyanobacterium for genetic studies (11).
In this work, we examined the chromosomal DNA of Syn-echocystis for modifications by methylation. Sequence-specificmethylations were detectable, whereas no restriction endonu-
* Corresponding author. Mailing address: Universitat Rostock, FBBiologie, Doberaner Str. 143, D-18051 Rostock, Germany. Phone: 49-381-4942076. Fax: 49-381-4942079. E-mail: [email protected].
† Present address: Department of Molecular Genetics and Cell Bi-ology, University of Chicago, Chicago, IL 60637.
4116
on April 10, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
on April 10, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
on April 10, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

clease activities were found. The open reading frame (ORF)slr0214 (11), which shows high similarities to genes encodingDNA-specific methyltransferases, was inactivated on the chro-mosome. In the mutant, alterations of the DNA methylationpattern were detected, demonstrating that slr0214 encodes amethyltransferase which recognizes a target sequence of 6 bp.Significantly, compared to the wild-type (WT) strain, the mu-tant could not grow in CO2-enriched cultures under standardconditions, indicating an important function of this methyl-transferase in Synechocystis. Furthermore, the slr0214 genewas subcloned and expressed in E. coli. The resulting proteinshowed methyltransferase activity in vitro.
MATERIALS AND METHODS
Strains and culture conditions. A derivative of Synechocystis with enhancedtransforming capacity was used in all experiments and was obtained from S.Shestakov (Moscow State University, Moscow, Russia). Axenic cells were cul-tured on agar plates at 30°C under constant illumination with mineral medium C(16). Transformants were initially selected on medium containing 10 mg ofkanamycin (Sigma) ml21, while the segregation of clones and the cultivation ofmutants were performed with 50 mg of kanamycin ml21. For physiological char-acterization, axenic cultures of cyanobacteria (about 108 cells ml21) were grownphotoautotrophically under continuous illumination with 180 mmol of photonss21 m22 (warm light; Osram L58 W32/3) at 29°C in batch cultures bubbling withair enriched with CO2 (2.5%) in potassium nitrate-containing growth medium(1). Contamination by heterotrophic bacteria was checked by microscopy orspreading of 0.2 ml of culture on Luria-Bertani (LB) plates. E. coli JM101 (31)was used for routine DNA manipulations. For the overexpression of protein,protease-deficient E. coli BL21 (31) was used. E. coli was cultured in LB mediumat 37°C. Growth was monitored by measuring the optical density at 750 nm(OD750) for Synechocystis and at 500 nm (OD500) for E. coli.
DNA manipulations. The isolation of total DNA from Synechocystis was doneas described previously (2). All other DNA techniques, such as plasmid isolation,transformation of E. coli, ligation, and restriction analysis (restriction enzymeswere obtained from Life Technologies and New England BioLabs), were stan-dard methods (31). For Southern hybridization experiments, the DNA probeswere labelled with digoxigenin by use of a PCR DIG probe synthesis kit (Boehr-inger Mannheim Biochemicals). For restriction analyses with chromosomalDNA from Synechocystis, restriction endonucleases were used in a 10-fold excessand were incubated for at least 16 h at 37°C in order to ensure completedigestion. The following synthetic primers were specifically used for amplifica-tion of the sequence encoding the DNA-specific methyltransferase (slr0214) (11):59-GGCGCCGGATCCATGGCCAGACCCATTGCCAT-39 (Met-fw; BamHIrecognition sequence underlined; start codon of slr0214 in boldface) and 59-CGGCGCGAATTCTTAGGAATGGGATTTGGAC-39 (Met-re; EcoRI recogni-tion sequence underlined; stop codon of slr0214 in boldface). In order togenerate an slr0214 deletion mutant, a fragment containing slr0214 and twoneighboring ORFs was amplified with the following primers: 59-CGCGGATCCCCGAGTTTTATTAGCCCTTT-39 (MetD-fw; BamHI recognition sequence un-derlined) and 59-AAAACTGCAGCTTTAGTCTCAGTGTGGCGA-39 (MetD-re; PstI recognition sequence underlined). For the in vitro methylation assay, aninternal fragment of the clpC gene of Synechocystis was amplified by PCR withthe following primers: 59-CTCAAATCCATGGGGGTTAA-39 (Clpfw) and 59-GAGGTCATGATCAACAGGGT-39 (Clpcre) (custom oligonucleotide synthe-sis; Pharmacia). For PCR, PCR-supermix (Life Technologies) and the followingtemperature cycles (30 times) were applied: 15 s at 94°C, 30 s at 52°C, and 1 minat 72°C. The plasmid vectors pUC19 (38) and pGEM7 (Promega) were used.
Generation of insertion and deletion mutants. For the generation of slr0214mutants, the aphII gene cartridge (encoding aminoglycoside phosphotransferase
II) from E. coli plasmid pUC4K (37) was integrated at selected restriction sitesinto the sequence of slr0214 cloned into E. coli vectors (Table 1). Plasmid DNAof these constructs was isolated from E. coli with a QIAprep spin plasmid minikit (Qiagen). About 1 mg of DNA was used for the transformation of Synecho-cystis, and kanamycin-resistant clones were selected (9).
Protein overexpression. For overexpression and purification of the Slr0214protein, the glutathione S-transferase (GST) gene fusion system (Pharmacia)was used. The slr0214 ORF was amplified from chromosomal DNA of Synecho-cystis by PCR with primers Met-fw and Met-re. The translational start codon(boldface letters in the Met-fw primer) was located immediately behind a BamHIsite used to clone the fragment in frame with the GST ORF into pGEX-4T-3(Pharmacia). Constructs showing the correct insertion were transformed into E.coli BL21, and a transformant was selected. The cells were cultured at 30°C in LBmedium. The expression of the protein was induced at an OD500 of 1.0 by theaddition of isopropyl-b-D-thiogalactopyranoside (IPTG) (100 mM), and incuba-tion was continued for 150 min. The proteins were extracted from E. coli bysonication, and the fusion protein was bound to a glutathione-Sepharose slurry(Pharmacia). The Slr0214 protein was released from the matrix by cleavage ofthe protease thrombin (Boehringer), for which a cleavage site was insertedbetween GST and Slr0214. The extracted proteins were separated in sodiumdodecyl sulfate gels containing acrylamide gradients from 7.5 to 15% in thebuffer system of Laemmli (17).
Protein extraction and methyltransferase assay. The extraction of proteinsfrom frozen cell material of Synechocystis was done by ultrasonic treatment (30W, 2 min) in 10 mM HEPES-NaOH buffer (pH 7.3) under ice cooling. Crudeprotein extracts were purified by centrifugation (15 min, 5,000 3 g, 4°C). Theprotein content was estimated by the method of Lowry et al. (20). DNA-specificmethyltransferase activity was assayed by incubation of unmethylated chromo-somal DNA of Micrococcus lysodeikticus (Sigma) with [3H]AdoMet (Amersham)as described by Gunthert et al. (7). The radioactivity incorporated in the DNAwas determined by liquid scintillation counting. In vitro methylation of theinternal fragment of the clpC gene was performed after mixing about 1 mg ofDNA, AdoMet (10 mM final concentration), and low-salt restriction buffer (LifeTechnologies) in a total volume of 200 ml. The reaction was started by addingabout 0.5 mg of the Slr0214 protein, and the mixture was incubated for 12 h at37°C. After concentration by precipitation, the DNA was directly used for re-striction analysis.
RESULTS
The occurrence of sequence-specific DNA methylation wasexamined by incubation of chromosomal DNA isolated fromSynechocystis with different restriction enzymes to test whetheror not their activities are specifically influenced by methylationof their recognition sequences. In these experiments, it wasfound that the DNA was resistant to the action of MboI, ApaI,EaeI, HaeIII, PvuI, and SgfI, while other restriction endonucle-ases were able to cut (Table 2). Different sensitivities of theDNA to different restriction endonucleases indicated that atleast three methylation activities seemed to be present. Onerepresented a Dam-like methyltransferase, which was demon-strated by the inhibition of the restriction enzyme MboI andthe activities of DpnI and Sau3A (Table 2). The activities ofthese enzymes were differentially influenced by methylation ofthe adenine in their recognition sequence, 59-GATC-39: MboIcould not cleave Dam-modified DNA; DpnI could cleave onlyDam-modified DNA; and Sau3A was not influenced by Dammodification, since its cognate methyltransferase modifies the
TABLE 1. Plasmids and mutants of Synechocystis used and constructed in this study (see also Fig. 2)
Designation Size (kb) Description
pGGM02 4.32 pGEM7 containing a 1.3-kb BamHI/EcoRI fragment, the coding sequence of slr0214 amplified by PCRpUC19GM 4.0 pUC19 containing a 1.3-kb BamHI/EcoRI fragment, the coding sequence of slr0214 amplified by PCRpGGMK292 5.6 pGGM02 containing inactivated slr0214 (aphII gene inserted at the HincII site opposite to the direction
of transcription of slr0214)pGGML05 6.3 pGEM7 containing a 3.3-kb BamHI/PstI fragment, the coding sequence of slr0214 with an additional 1 kb
upstream and 1 kb downstream amplified by PCRpGGMDE122 6.3 pGGM02 containing partially deleted slr0214 (aphII gene inserted, after deletion of the internal BalI frag-
ment, opposite to the direction of transcription of slr0214)pGEXGM 6.2 pGEX4T3 containing a 1.3-kb BamHI/EcoRI fragment, the coding sequence of slr0214 fused to GSTslr0214 mutant Synechocystis mutant obtained after transformation of the WT with pGGMK292slr0214D mutant Synechocystis mutant obtained after transformation of the WT with pGGMDE122
VOL. 180, 1998 DNA METHYLATION IN SYNECHOCYSTIS 4117
on April 10, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

cytosine (24). Comparison of the recognition sequences ofthe other inhibited restriction endonucleases showed that atleast two different cytosine-specific methyltransferases wereinvolved. The activities of ApaI, EaeI, and HaeIII were mostprobably affected by one enzyme recognizing the sequence59-GGCC-39. PvuI and SgfI should have been inhibited by acytosine-specific methyltransferase modifying one of the twocytosines within the sequence 59-CGATCG-39, which includesthe Dam recognition site. The complete digestion of chromo-somal DNA by Sau3A excluded methylation of the secondcytosine. Thus, this cytosine-specific methyltransferase mostprobably modified the first cytosine of a PvuI site (Table 2).Statistical analyses showed that the frequency of occurrence ofPvuI and SgfI recognition sequences was increased severalfoldin the chromosomal DNA of Synechocystis compared to theirtheoretical frequency in a random sequence. The frequenciesof the recognition sequences of EaeI and HaeIII were alsoincreased but to a lesser extent, whereas the occurrence of theMboI site corresponded to its frequency in a random sequence(Table 2).
In order to test if these methyltransferase activities were partof a strain-specific R-M system, the restriction endonucleaseactivity of Synechocystis was examined. This was done by incu-bating unmethylated DNA of phage l (MBI Fermentas) withprotein extracts (150 mg) in the presence of the basic reactionbuffers (React 1 to 4; Life Technologies) recommended formost restriction endonucleases at 37°C overnight (19). Theseexperiments clearly showed that there was no restriction en-donuclease activity, since the l DNA was not cut into definedfragments after incubation with the large amounts of proteinsused over a long time under different reaction conditions (datanot shown).
On the genome of Synechocystis, there is one ORF (slr0214)encoding a putative cytosine-specific methyltransferase (11).The amino acid sequence of Slr0214 was aligned with those ofother methyltransferases. The proteins showing the best align-ments all belonged to the group of C5-cytosine methyltrans-ferases (Fig. 1). The highest amino acid identity (48.2%) overthe whole sequence was found with the XorII methyltrans-
ferase from Xanthomonas oryzae (5). The homology patterncharacteristic for the group of C5-cytosine methyltransferaseswas found to be well conserved in the predicted amino acidsequence for Synechocystis (Fig. 1). All of the 10 conservedregions (26) could be identified. The variable region betweenmotifs XIII and IX has been shown to determine the specificityof DNA sequence recognition (14). Especially within the vari-able region the amino acid sequences of Slr0214 from Synecho-cystis and the XorII methyltransferase showed the highest ho-mology (41.5% identical amino acid residues; less than 30%identity to those of other enzymes), indicating that these twoenzymes also might have similarities in their recognition se-quences. XorII is an isoschizomer of PvuI, whose recognitionsequence was found to be modified (Table 2). Therefore, theprotein encoded by slr0214 could be a candidate for a methyl-transferase that modifies the PvuI recognition sequence (59-CGATCG-39) in Synechocystis. Since XorII is also inhibited byDam modification (5), the following experiments were per-formed with PvuI.
In order to analyze the function of the putative C5-cytosinemethyltransferase, mutants impaired in slr0214 were gener-
TABLE 2. Restriction enzymes used for identification ofmethylation sites on chromosomal DNA of Synechocystisa
Restrictionenzyme
Recognitionsequenceb
Cleavageof chromo-somal DNA
Frequency (in bp)of occurrence in:
Chromoso-mal DNAc
Randomsequence
PvuId C G A T C G Noe 1/698 1/4,096SgfI G C G A T C G C Noe 1/1,131 1/65,536ApaI G G G C C C No 1/3,738 1/4,096EaeI (C/T)G G C C(G/A) No 1/544 1/2,048HaeIII G G C C Noe 1/186 1/256DpnI G A T C Yese 1/239 1/256MboI G A T C Noe 1/239 1/256Sau3AI G A T C Yese 1/239 1/256
a The following enzymes were additionally tested and found able to cut Syn-echocystis DNA: AflII, AflIII, AhaII, AluI, Asp718I, AspEI, AvaI, AvaII, BamHI,BglII, BstNI, CelII, ClaI, DdeI, DpnI, Eco47I, Eco47III, EcoRI, EcoRII, EcoRV,EspI, Fnu4HI, HindIII, HpaI, HpaII, MvaI, NcoI, PstI, PvuII, SacI, SmaI, XbaI,and XhoI. For enzymes in boldface, see footnote e.
b Underlining indicates that methylation inhibited restriction (24). Boldfacingindicates that N6-methyladenine is a prerequisite for restriction (24). Italicizingindicates that restriction is not influenced by methylation (24).
c Based on the sequence of the entire genome of Synechocystis (11).d Methylation of the first cytosine probably inhibited restriction (see Results).e Restriction was controlled additionally to gel electrophoresis by Southern
hybridization experiments.
FIG. 1. Amino acid sequence comparison between the Slr0214 protein(SynMI) (11) and the C5-methylcytosine-synthesizing methyltransferases XorII(5), DdeIM (34), NgoII (33), and HphIM (21). Uppercase letters and a shadedbackground indicate amino acids identical to those in SynMI. The conservedmotifs characteristic for C5-cytosine methyltransferases (26) are indicated byRoman numerals.
4118 SCHARNAGL ET AL. J. BACTERIOL.
on April 10, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

ated. After cloning of a PCR fragment containing Slr0214(resulting in plasmid pGGM02) (Table 1), an aphII gene car-tridge (conferring kanamycin resistance) whose transcriptionoccurred opposite to the direction of transcription of slr0214was inserted into the central HincII site (resulting in plasmidpGGMK292) (Fig. 2 and Table 1). Furthermore, a PCR frag-ment containing about 1.6 kb in addition to the sequence ofslr0214 was cloned (resulting in plasmid pGGML05) (Table 1).In plasmid pGGML05, about 60% of the slr0214 sequence wasdeleted by cutting with BalI, and the deleted fragment wasreplaced by an aphII gene cartridge (resulting in plasmidpGGMDE122) (Fig. 2 and Table 1). Synechocystis was trans-formed with these constructs. Since these plasmids do not rep-licate in Synechocystis, continued selection on kanamycin de-mands the integration of the aphII gene into the chromosome.After cultivation for several generations at an increased con-centration of the antibiotic, we isolated kanamycin-resistantclones. With this strategy, mutants impaired in slr0214 wereobtained by insertion of the kanamycin resistance cassette(slr0214 or insertion mutant) or by replacement of the cassette(slr0214D or deletion mutant).
The lesion in the DNA of these mutants was characterizedby Southern hybridization experiments (Fig. 3; compare to Fig.2). In NcoI-digested DNA of the slr0214 mutant, one fragmentof 4.5 kb was found with the slr0214 gene probe. This fragmentwas 1.2 kb larger than the signal detected in the WT DNA,consistent with the integration of the 1.2-kb aphII gene car-tridge (Fig. 3A). One fragment of only 2.4 kb became visible byhybridization with the slr0214D mutant DNA. This size exactlymatched the size of the fragment expected for the deletion mu-tant (Fig. 3A). After cutting of chromosomal DNA with HindIII,in the WT only one fragment of 2.5 kb was detectable with theslr0214 gene probe, while in the mutants two fragments (theaphII gene contains one HindIII site) of the expected sizes (2.2and 1.7 kb for the slr0214 mutant and 2.0 and 1.0 kb for theslr0214D mutant [with the slr0214 gene probe, the 1.0-kb frag-ment of the deletion mutant was detectable only as a very faintband, since it shared only 30 bp with the DNA probe]) became
visible (Fig. 3A). The same pattern was obtained for the DNAof the mutants when the aphII gene was used as a probe (Fig.3B). Again, after digestion with NcoI, fragments of 4.5 and 2.4kb were detected. After cutting with HindIII, the aphII geneprobe hybridized with a 2.2-kb fragment and a 1.7-kb fragmentof the slr0214 mutant DNA, whereas a 2.0-kb band and a 1.0-kb band were clearly visible with the slr0214D mutant DNA.No specific signal could be detected by hybridization with theWT DNA. These hybridization patterns indicated that the plas-mid constructs used for the transformation of Synechocystis toobtain the mutants were correctly integrated by double cross-ing over and replaced completely the WT copy of the slr0214gene.
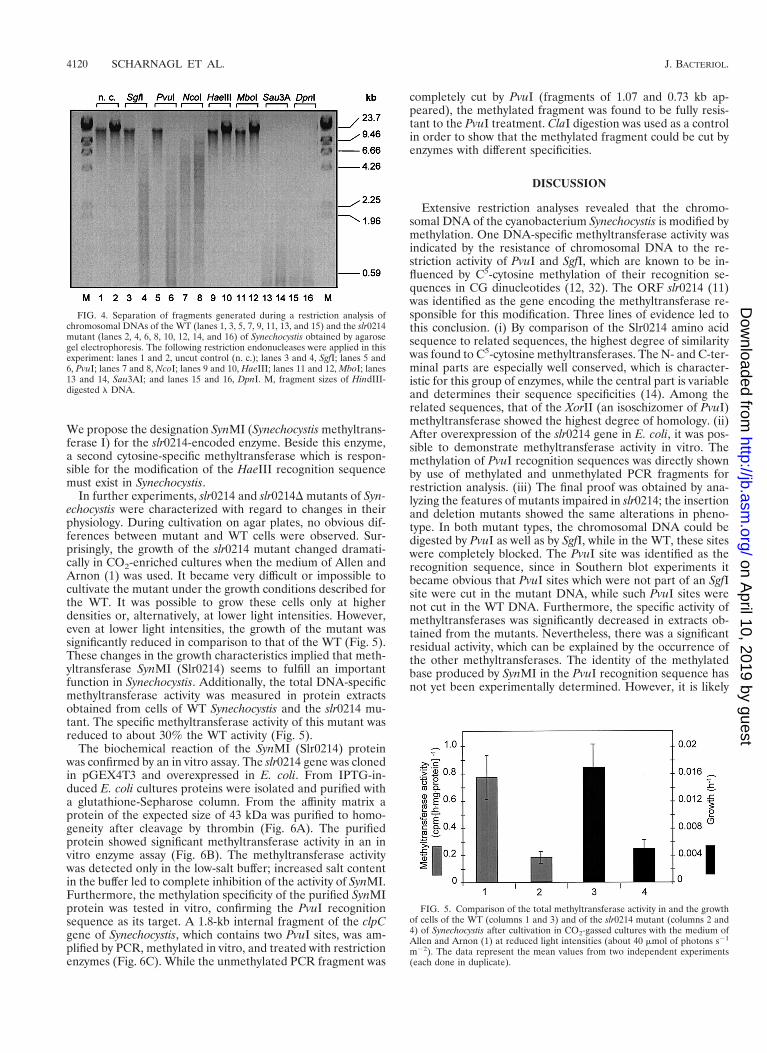
In order to analyze changes in the methylation pattern, chro-mosomal DNA isolated from the mutants was incubated withdifferent restriction enzymes which were found to be inhibitedby the WT DNA (Table 2). Digestion with NcoI was used as acontrol reaction in order to show that the DNA preparationscould be cleaved under the selected reaction conditions. Incontrast to the WT DNA, the DNA of the slr0214 mutant (Fig.4) as well as the DNA of the slr0214D mutant (data not shown)became accessible to restriction by PvuI and SgfI. The othermethylation activities, the Dam-like methylation and the sec-ond cytosine-specific methylation, remained intact, because theDNA of both mutants was still resistant to MboI as well asHaeIII and was digested by DpnI (Fig. 4; compare to Table 2).The ability of PvuI and SgfI to cut mutant DNA was confirmedby Southern hybridization experiments. With PvuI- and SgfI-cut DNAs from the WT and the mutants, three and two frag-ments (PvuI and SgfI, respectively) (Fig. 2) of the expectedsizes appeared after application of the slr0214 gene probe,while with the WT DNA only hybridization to the uncut high-molecular-weight DNA was visible (data not shown). There-fore, the methyltransferase encoded by slr0214 is specificallydirected to the recognition sequence of PvuI or its isoschizo-mer XorII. Furthermore, the methylation is directed to a cy-tosine within the recognition sequence, since the internal ad-enine remained modified, as shown by the inhibition of MboI.
FIG. 2. Schematic drawing showing the genetic organization, restriction map,and protein-encoding regions of the chromosomal region encoding Slr0214(SynMI) in Synechocystis (11). (A and B) The insertion of the aphII gene inselected sites to generate mutants is shown. (A) BalI was used to obtain theslr0214D mutant. (B) HincII was used to obtain the slr0214 mutant. (C) Protein-encoding region. (D) The binding sites of the primers are represented by trian-gles under the arrows indicating the protein-encoding regions in panel C.
FIG. 3. Southern blot experiments for characterization of complete segrega-tion of the slr0214 mutant and the slr0214D mutant. The digoxigenin-labelledPCR fragment containing slr0214 (A) or the digoxigenin-labelled aphII gene (B)was used as a probe for hybridization to HindIII digested (lanes 1 to 3) andNcoI-digested (lanes 4 to 6) chromosomal DNAs from cells of the WT (lanes 1and 4), the slr0214D mutant (lanes 2 and 5), and the slr0214 mutant (lanes 3 and6) of Synechocystis. M, fragment sizes of digoxigenin-labelled HindIII-digested lDNA.
VOL. 180, 1998 DNA METHYLATION IN SYNECHOCYSTIS 4119
on April 10, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
We propose the designation SynMI (Synechocystis methyltrans-ferase I) for the slr0214-encoded enzyme. Beside this enzyme,a second cytosine-specific methyltransferase which is respon-sible for the modification of the HaeIII recognition sequencemust exist in Synechocystis.
In further experiments, slr0214 and slr0214D mutants of Syn-echocystis were characterized with regard to changes in theirphysiology. During cultivation on agar plates, no obvious dif-ferences between mutant and WT cells were observed. Sur-prisingly, the growth of the slr0214 mutant changed dramati-cally in CO2-enriched cultures when the medium of Allen andArnon (1) was used. It became very difficult or impossible tocultivate the mutant under the growth conditions described forthe WT. It was possible to grow these cells only at higherdensities or, alternatively, at lower light intensities. However,even at lower light intensities, the growth of the mutant wassignificantly reduced in comparison to that of the WT (Fig. 5).These changes in the growth characteristics implied that meth-yltransferase SynMI (Slr0214) seems to fulfill an importantfunction in Synechocystis. Additionally, the total DNA-specificmethyltransferase activity was measured in protein extractsobtained from cells of WT Synechocystis and the slr0214 mu-tant. The specific methyltransferase activity of this mutant wasreduced to about 30% the WT activity (Fig. 5).
The biochemical reaction of the SynMI (Slr0214) proteinwas confirmed by an in vitro assay. The slr0214 gene was clonedin pGEX4T3 and overexpressed in E. coli. From IPTG-in-duced E. coli cultures proteins were isolated and purified witha glutathione-Sepharose column. From the affinity matrix aprotein of the expected size of 43 kDa was purified to homo-geneity after cleavage by thrombin (Fig. 6A). The purifiedprotein showed significant methyltransferase activity in an invitro enzyme assay (Fig. 6B). The methyltransferase activitywas detected only in the low-salt buffer; increased salt contentin the buffer led to complete inhibition of the activity of SynMI.Furthermore, the methylation specificity of the purified SynMIprotein was tested in vitro, confirming the PvuI recognitionsequence as its target. A 1.8-kb internal fragment of the clpCgene of Synechocystis, which contains two PvuI sites, was am-plified by PCR, methylated in vitro, and treated with restrictionenzymes (Fig. 6C). While the unmethylated PCR fragment was
completely cut by PvuI (fragments of 1.07 and 0.73 kb ap-peared), the methylated fragment was found to be fully resis-tant to the PvuI treatment. ClaI digestion was used as a controlin order to show that the methylated fragment could be cut byenzymes with different specificities.
DISCUSSION
Extensive restriction analyses revealed that the chromo-somal DNA of the cyanobacterium Synechocystis is modified bymethylation. One DNA-specific methyltransferase activity wasindicated by the resistance of chromosomal DNA to the re-striction activity of PvuI and SgfI, which are known to be in-fluenced by C5-cytosine methylation of their recognition se-quences in CG dinucleotides (12, 32). The ORF slr0214 (11)was identified as the gene encoding the methyltransferase re-sponsible for this modification. Three lines of evidence led tothis conclusion. (i) By comparison of the Slr0214 amino acidsequence to related sequences, the highest degree of similaritywas found to C5-cytosine methyltransferases. The N- and C-ter-minal parts are especially well conserved, which is character-istic for this group of enzymes, while the central part is variableand determines their sequence specificities (14). Among therelated sequences, that of the XorII (an isoschizomer of PvuI)methyltransferase showed the highest degree of homology. (ii)After overexpression of the slr0214 gene in E. coli, it was pos-sible to demonstrate methyltransferase activity in vitro. Themethylation of PvuI recognition sequences was directly shownby use of methylated and unmethylated PCR fragments forrestriction analysis. (iii) The final proof was obtained by ana-lyzing the features of mutants impaired in slr0214; the insertionand deletion mutants showed the same alterations in pheno-type. In both mutant types, the chromosomal DNA could bedigested by PvuI as well as by SgfI, while in the WT, these siteswere completely blocked. The PvuI site was identified as therecognition sequence, since in Southern blot experiments itbecame obvious that PvuI sites which were not part of an SgfIsite were cut in the mutant DNA, while such PvuI sites werenot cut in the WT DNA. Furthermore, the specific activity ofmethyltransferases was significantly decreased in extracts ob-tained from the mutants. Nevertheless, there was a significantresidual activity, which can be explained by the occurrence ofthe other methyltransferases. The identity of the methylatedbase produced by SynMI in the PvuI recognition sequence hasnot yet been experimentally determined. However, it is likely
FIG. 5. Comparison of the total methyltransferase activity in and the growthof cells of the WT (columns 1 and 3) and of the slr0214 mutant (columns 2 and4) of Synechocystis after cultivation in CO2-gassed cultures with the medium ofAllen and Arnon (1) at reduced light intensities (about 40 mmol of photons s21
m22). The data represent the mean values from two independent experiments(each done in duplicate).
FIG. 4. Separation of fragments generated during a restriction analysis ofchromosomal DNAs of the WT (lanes 1, 3, 5, 7, 9, 11, 13, and 15) and the slr0214mutant (lanes 2, 4, 6, 8, 10, 12, 14, and 16) of Synechocystis obtained by agarosegel electrophoresis. The following restriction endonucleases were applied in thisexperiment: lanes 1 and 2, uncut control (n. c.); lanes 3 and 4, SgfI; lanes 5 and6, PvuI; lanes 7 and 8, NcoI; lanes 9 and 10, HaeIII; lanes 11 and 12, MboI; lanes13 and 14, Sau3AI; and lanes 15 and 16, DpnI. M, fragment sizes of HindIII-digested l DNA.
4120 SCHARNAGL ET AL. J. BACTERIOL.
on April 10, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

that the first cytosine represents the target for methylation bySynMI, a conclusion which could be drawn from the data ob-tained for the activity and inhibition of different restrictionenzymes affecting the PvuI site and its internal sequence, 59-GATC-39, by use of DNAs from the WT and the Slr0214mutant (Table 2 and Fig. 4).
At least a second type of methyltransferase activity is presentin Synechocystis; this activity resembles the Dam modificationcharacterized in E. coli, in which the adenine in the sequence59-GATC-39 is modified. This finding was clearly shown by theinhibition of MboI, which is affected by the adenine methyl-ation, and by the activity of DpnI, which cuts only if the ade-nine is methylated (24). In E. coli, Dam is involved in multiplefunctions: (i) mismatch repair, (ii) regulation of initiation ofchromosome replication, and (iii) modification of expressionof several genes (for a review, see reference 22). Dam-likemodification of chromosomal DNA had been found in severalcyanobacteria (e.g., 18). In Synechocystis, mutation of slr0214did not affect the Dam-like methylation type. The correspond-ing methyltransferase probably is encoded by ORF slr1803(11), which encodes a putative adenine-specific methyltrans-ferase in Synechocystis and shows the highest similarity (41.2%identical amino acid residues) to MboA, the cognate methyl-transferase of the MboI restriction endonuclease in Moraxellabovis (36).
In addition, another cytosine-specific methyltransferase ac-tivity modifying the HaeIII recognition sequence was not af-fected by inactivation of slr0214. Except for slr0214 and slr1803,no other ORF encoding a putative DNA-specific methyltrans-ferase has been identified on the sequenced chromosome ofSynechocystis (11). Thus, the HaeIII site-specific methyltrans-ferase might be encoded by one of the three extrachromosomalelements (with a total size of 280 kb) (15) which were identifiedin Synechocystis or, alternatively, this enzyme might representa new type of methyltransferase. The latter is not very likelybecause of the high level of conservation of cytosine-specificmethyltransferases (26).
In many bacteria, DNA methylation is related to a strain-specific R-M system. However, in Synechocystis, no restrictionendonuclease activity was demonstrated by incubation of lamb-da DNA with large amounts of cellular protein. The capacity ofthis strain for natural transformation (6, 35) is a further indi-
cation for the absence of such enzymes; in addition, on theentire chromosome no ORF showed homologies to genes forknown restriction endonucleases (11). Therefore, Synechocystisexpresses methyltransferases which are not part of a host-spe-cific R-M system. Nevertheless, at least the enzyme SynMI(Slr0214) seems to be have an important function, since themutants showed altered growth characteristics.
The recognition sequence of the methyltransferase SynMI ispart of the HIP1 sequence (highly iterated palindrome) (8),which is identical to the SgfI site. HIP1 is very abundant in theDNA of several but not all cyanobacteria, where it occurs incoding and intergenic regions (28). The complete genome se-quence of Synechocystis (11) allowed us to estimate their fre-quencies. SynMI or PvuI sites were found every 698 bp and SgfIor HIP1 sequences were found every 1,131 bp in the DNA(Table 2). In Synechocystis sp. strain PCC 6301, HIP1 wasfound to be extremely frequent (every 320 bp). However, inthis strain, HIP1 is not methylated at all or by the same meth-ylase as in Synechocystis, since its chromosomal DNA was com-pletely digested by PvuI (28). The HIP1 sequence seems to beinvolved in excision and gene rearrangement (29).
In Synechocystis, the target sequences of the SynMI methyl-transferase, PvuI, including the HIP1 sequence, are highly over-represented and are completely methylated in the WT. Hypo-thetically, these methylated sequences could serve as bindingsites for chromatin-like proteins. The same function was sug-gested for another repetitive sequence found in filamentouscyanobacterial strains (23). The proteins associated with theDNA might have an influence on the rate of gene expression,as was found for histones in eukaryotic cells. Interestingly, thesequences of some highly expressed genes in the genome ofSynechocystis (e.g., genes encoding for rRNAs and for subunitsof photosystem I [psaF, -D, and -L] and of photosystem II[psbA1 to psbA3, psbCD]) (11) do not contain any PvuI site andare therefore also not methylated in WT cells. The completeabsence of methylation in the slr0214 mutants of Synechocystiscould disturb the balanced gene expression program, leadingto problems in growth at higher light intensities in CO2-en-riched cultures. In future experiments, we will analyze the phys-iological function of the SynMI methyltransferase in moredetail by growing the WT and the mutants under different
FIG. 6. Overexpression of the Slr0214 protein in E. coli BL21 by use of the GST gene fusion system (Pharmacia). (A) Coomassie blue staining of proteins afterseparation by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Lanes: 1, total extract from E. coli cells after induction by IPTG; 2, crude extract aftersonication and centrifugation at 25,000 3 g for 20 min; 3, isolated Slr0214 protein after thrombin cleavage; M, prestained broad-range markers (Bio-Rad). (B)Methyltransferase activity of the purified Slr0214 protein in in vitro assays with buffers containing different NaCl concentrations. Lanes: 1, assays done with 100 mMNaCl; 2, assays done with 50 mM NaCl; 3, assays done under NaCl-free conditions. (C) Restriction analysis of a DNA fragment with (lanes 4 to 6) and without (lanes1 to 3) methylation by SynMI in vitro. Lanes: 1 and 4, uncut; 2 and 5, PvuI; 3 and 6, ClaI; M, fragment sizes of EcoRI/HindIII-digested l DNA.
VOL. 180, 1998 DNA METHYLATION IN SYNECHOCYSTIS 4121
on April 10, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

conditions and by comparing the expression of genes with orwithout PvuI sites in both the WT and the mutants.
ACKNOWLEDGMENTS
The excellent technical assistance of B. Brzezinka, I. Dorr, and K.Sommerey is acknowledged. W. Messer and Jurgen Villert from theMax-Planck-Institut fur Molekulare Genetik, Berlin, Germany, are ac-knowledged for their help in the establishment of the methyltrans-ferase assay and helpful discussions.
REFERENCES
1. Allen, M. B., and D. I. Arnon. 1955. Studies on nitrogen fixing blue-greenalgae. II. The sodium requirement of Anabaena cylindrica. Physiol. Plant. 8:653–660.
2. Apte, S. K., and R. Haselkorn. 1990. Cloning of salinity stress-induced genesfrom the salt-tolerant nitrogen-fixing cyanobacterium Anabaena torulosa.Plant Mol. Biol. 15:723–733.
3. Barten, R., and H. Lill. 1995. DNA uptake in the naturally competentcyanobacterium Synechocystis sp. PCC 6803. FEMS Microbiol. Lett. 129:83–88.
4. Cedar, H. 1990. DNA methylation and gene activity. Cell 53:3–4.5. Choi, S. H., and J. E. Leach. 1994. Identification of the Xor II methyltrans-
ferase gene and a vsr homolog from Xanthomonas oryzae pv. oryzae. Mol.Gen. Genet. 244:383–390.
6. Grigorieva, G., and S. V. Shestakov. 1982. Transformation in the cyanobac-terium Synechocystis sp. 6803. FEMS Microbiol. Lett. 13:367–370.
7. Gunthert, U., M. Freund, and T. A. Trautner. 1981. Restriction and modi-fication in Bacillus subtilis: two DNA methyltransferases with BsuRI speci-ficity. J. Biol. Chem. 256:9340–9345.
8. Gupta, A., A. P. Morby, J. S. Turner, B. A. Whitton, and N. J. Robinson.1993. Deletion within the metallothionein locus of cadmium-tolerant Syn-echococcus PCC 6301 involving a highly iterated palindrome (HIP1). Mol.Microbiol. 7:189–195.
9. Hagemann, M., and E. Zuther. 1992. Selection and characterization of mu-tants of the cyanobacterium Synechocystis sp. PCC 6803 unable to toleratehigh salt concentrations. Arch. Microbiol. 158:429–434.
10. Houmard, J., and N. Tandeau de Marsac. 1988. Cyanobacterial genetictools: current status. Methods Enzymol. 167:808–850.
11. Kaneko, T., S. Sato, H. Kotani, A. Tanaka, E. Asamizu, Y. Nakamura, N.Miyajima, M. Hirosawa, M. Sugiura, S. Sasamoto, T. Kimura, T. Hosouchi,A. Matsuno, A. Muraki, N. Nakazaki, K. Nruo, S. Okumura, S. Shimpo, C.Takeuchi, T. Wada, A. Watanabe, M. Yamada, M. Yasuda, and S. Tabata.1996. Sequence analysis of the genome of the unicellular cyanobacteriumSynechocystis sp. strain PCC 6803. II. Sequence determination of the entiregenome and assignment of potential protein-coding regions. DNA Res. 3:109–136.
12. Kappelman, J. R., M. Brady, K. Knoche, E. Murray, T. Schoenfeld, R.Williams, and N. Vesselinova. 1995. Sgf I, a new type-II restriction endonu-clease that recognizes the octanucleotide sequence 59-GCGAT/CGC-39.Gene 160:55–58.
13. Karreman, C., and A. DeWaard. 1990. Agmenellum quadruplicatum M AquI,a novel modification methylase. J. Bacteriol. 172:266–272.
14. Klimasauskas, S., J. L. Nelson, and R. J. Roberts. 1991. The sequencespecificity domain of cytosine-C5 methylases. Nucleic Acids Res. 19:6183–6190.
15. Kotani, H., A. Tanaka, T. Kaneko, S. Sato, M. Sugiura, and S. Tabata. 1995.Assignment of 82 known genes and gene clusters on the genome of theunicellular cyanobacterium Synechocystis sp. strain PCC6803. DNA Res. 2:133–142.
16. Kratz, W. A., and J. Myers. 1955. Nutrition and growth of several blue-greenalgae. Am. J. Bot. 42:282–287.
17. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4. Nature 227:680–685.
18. Lambert, G. R., and N. G. Carr. 1984. Resistance of DNA from filamentousand unicellular cyanobacteria to restriction endonuclease cleavage. Biochim.Biophys. Acta 781:45–55.
19. Laue, F., L. R. Evans, M. Jarsch, N. L. Brown, and C. Kessler. 1991. Acomplex family of class-II restriction endonucleases, Dsa I-VI, in Dactylo-coccopsis salina. Gene 97:87–95.
20. Lowry, O. H., N. J. Rosebrough, A. L. Farr, and R. J. Randall. 1951. Proteinmeasurement with the Folin phenol reagent. J. Biol. Chem. 193:265–275.
21. Lubys, A., J. Lubiene, K. Stankevicius, and A. Janulaitis. 1996. Cloning andanalysis of the gene encoding type II restriction-modification system HphI.Nucleic Acids Res. 97:87–95.
22. Marinus, M. G. 1996. Methylation of DNA, p. 782–791. In F. C. Neidhardt,R. Curtiss III, J. L. Ingraham, E. C. C. Lin, K. B. Low, B. Magasanik, W. S.Reznikoff, M. Riley, M. Schaechter, and H. E. Umbarger (ed.), Escherichiacoli and Salmonella: cellular and molecular biology, 2nd ed. ASM Press,Washington, D.C.
23. Mazel, D., J. Houmard, A. M. Castets, and N. Tandeau de Marsac. 1990.Highly repetitive sequences in cyanobacterial genomes. J. Bacteriol. 172:2755–2761.
24. Nelson, M., E. Raschke, and M. McClelland. 1993. Effect of site-specificmethylation on restriction endonucleases and DNA modification methylases.Nucleic Acids Res. 21:3139–3154.
25. Noyer-Weidner, M., and T. A. Trautner. 1993. Methylation of DNA inprokaryotes. Birkhauser, Basel, Switzerland.
26. Posfai, J., A. S. Bhagwat, G. Posfai, and R. J. Roberts. 1989. Predictivemotifs derived from cytosine methyltransferases. Nucleic Acids Res. 17:2421–2435.
27. Roberts, R. J., and D. Macelis. 1996. Restriction enzymes and methylases.Nucleic Acids Res. 24:223–235.
28. Robinson, N. J., P. J. Robinson, A. Gupta, A. J. Bleasby, B. A. Whitton, andA. P. Morby. 1995. Singular over-representation of an octameric palindrome,HIP1, in DNA from many cyanobacteria. Nucleic Acids Res. 23:729–735.
29. Robinson, P. J., R. M. Cranenburgh, I. M. Head, and N. J. Robinson. 1997.HIP1 propagates in cyanobacterial DNA via nucleotide substitutions butpromotes excision at similar frequencies in Escherichia coli and Synechococ-cus PCC 7942. Mol. Microbiol. 24:181–189.
30. Ruan, H., K. D. Lunnen, M. E. Scott, L. S. Moran, B. E. Slatko, J. J.Pelletier, E. J. Hess, I. J. Benner, G. G. Wilson, and S.-Y. Xu. 1996. Cloningand sequence comparison of AvaI and BsoBI restriction-modification sys-tems. Mol. Gen. Genet. 252:695–699.
31. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual. Cold Spring Harbor Laboratory Press, Cold Spring Har-bor, N.Y.
32. Smith, M. D., M. Longo, G. F. Gerard, and D. K. Chatterjee. 1992. Cloningand characterization of genes for the PvuI restriction and modification sys-tem. Nucleic Acids Res. 20:5743–5747.
33. Sullivan, K. M., and J. R. Saunders. 1988. Sequence analysis of the NgoPIImethyltransferase gene from Neisseria gonorrhoeae P9: homologies withother enzymes recognizing the sequence 59-GGCC-39. Nucleic Acids Res. 16:4369–4387.
34. Szynter, L. A., B. Slatko, L. Moran, K. H. O’Donnell, and J. E. Brooks. 1987.Nucleotide sequence of the DdeI restriction-modification system and char-acterization of the methylase protein. Nucleic Acids Res. 15:8249–8266.
35. Thiel, T. 1995. Molecular genetic techniques for analysis of cyanobacteria,p. 581–611. In D. A. Bryant (ed.), Molecular biology of cyanobacteria.Kluwer Academic Publishers, Dordrecht, The Netherlands.
36. Ueno, T., H. Itoh, F. Kimizuka, H. Kotani, and K. Nakajima. 1993. Genestructure and expression of MboI restriction-modification system. NucleicAcids Res. 21:2309–2313.
37. Vieira, J., and J. Messing. 1982. The pUC plasmids, an M13mp7-derivedsystem for insertion mutagenesis and sequencing with synthetic universalprimers. Gene 19:259.
38. Yanisch-Perron, C., J. Vieira, and J. Messing. 1985. Improved M13 phagecloning vectors and host strains: nucleotide sequence of the M13mp18 andpUC19 vectors. Gene 33:103–119.
4122 SCHARNAGL ET AL. J. BACTERIOL.
on April 10, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

AUTHORS’ CORRECTIONS
The Highly Conserved, Coregulated SNO and SNZ GeneFamilies in Saccharomyces cerevisiae Respond to
Nutrient LimitationPAMELA A. PADILLA, EDWINA K. FUGE, MATTHEW E. CRAWFORD, ALLISON ERRETT,
AND MARGARET WERNER-WASHBURNE
Volume 180, no. 21, p. 5718–5726, 1998. Page 5725, Acknowledgments, second paragraph, lines 3–5: “and by MBRS grant5SO6GM52576-03 from the National Science Foundation and a Patricia Roberts Harris Fellowship to P.A.P.” should read “andby MBRS and Patricia Roberts Harris grants from NIH to P.A.P.”
The Cyanobacterium Synechocystis sp. Strain PCC 6803 Expressesa DNA Methyltransferase Specific for the Recognition
Sequence of the Restriction Endonuclease PvuIMATTHIAS SCHARNAGL, STEFAN RICHTER, AND MARTIN HAGEMANN
FB Biologie, Universitat Rostock, D-18051 Rostock, and Max-Planck-Institutfur Molekulare Genetik, D-14195 Berlin, Germany
Volume 180, no. 16, p. 4116–4122, 1998. Pages 4116–4122: The DNA methyltransferase from Synechocystis sp. strain PCC 6803named SynMI should have been named M.Ssp6803I in accordance with the usual nomenclature for DNA methyltransferases.
6794

![Synechocystis sp. PCC 6803 in a bio-photoelectrolysis cell ... · 4-resu e efficienc ncentration ated. )6] 4-. Applic ells) did no lted in an i ies of H2 s and tem ation of a t produce](https://img.pdfslide.us/doc/110x75/5f80e05ab8e29d00bc4e36a8/synechocystis-sp-pcc-6803-in-a-bio-photoelectrolysis-cell-4-resu-e-efficienc.jpg)



























