Embed Size (px)
Citation preview

The Creation of Functional Pseudoislets Using Modular Tissue Engineering
by
Yarden Gratch
A thesis submitted in conformity with the requirements for the degree of Master of Applied Science
Department of Chemical Engineering and Applied Chemistry University of Toronto
© Copyright by Yarden Gratch 2016

ii
The Creation of Functional Pseudoislets Using Modular Tissue
Engineering
Yarden Gratch
Master of Applied Science
Department of Chemical Engineering and Applied Chemistry
University of Toronto
2016
Abstract
Pancreatic islet transplantation exists as an option for treating type I diabetes but is not widely
used as there is a shortage of donors and it requires intensive immunosuppression. We
investigated aggregating islet cells in modules (injectable microtissue structures) to create
pseudoislets that have better vasculature and ability to reverse hyperglycemia. Blood outgrowth
endothelial cells (BOEC) are autologous and easily obtained, making them an ideal endothelial
cell source. Results showed there were no significant differences in vascularization when using
BOEC rather than human umbilical vein endothelial cells (HUVEC) or when embedding them in
modules compared to coating them. Diabetic SCID-Bg mice returned to normoglycemia faster
when subcutaneously implanted with BOEC coated modules or modules containing αTC1-6 cells
along with MIN6 cells (in a 33/67 mix). Future work with stem cell sources could eliminate the
strain on donor supply and lead to a long-term solution for those with type I diabetes.

iii
Acknowledgments
I would like to thank Professor Michael V. Sefton for supervising my thesis work and
encouraging my continued research work over the last few years; I have learned a lot from you
and really appreciate all the guidance you have provided. I would also like to thank Chuen Lo for
his expertise in animal surgery, and the entire Sefton Lab for their help along the way. I also
thank Professor Alison McGuigan and Damien Noone for being part of my thesis committee and
providing me with much appreciated advice.
I would like to express my appreciation to my friends from "Imagine Green", my band
VolpeMantra, and those from high school for providing me with fun, laughs and exciting
experiences to keep me going. Additionally, I would like to thank my family for always
providing their support, particularly in keeping me well fed with a roof over my head. Lastly, I
would like to express my gratitude to Pendar Aryafar for his continued encouragement and
ability to always help me relax no matter how challenging things got.
I look forward to moving on to the next chapter of my life and will always remember none of
this would have been possible without all of you.

iv
Table of Contents
Acknowledgments.......................................................................................................................... iii
Table of Contents ........................................................................................................................... iv
List of Tables ................................................................................................................................ vii
List of Figures .............................................................................................................................. viii
List of Appendices ...........................................................................................................................x
List of Abbreviations ..................................................................................................................... xi
Chapter 1 Introduction .....................................................................................................................1
1 Introduction .................................................................................................................................1
1.1 Hypothesis and Research Objectives ...................................................................................1
1.2 Clinical Impetus ...................................................................................................................2
1.3 Islet Transplantation and Limitations ..................................................................................3
1.3.1 Islet Transplantation Background ............................................................................3
1.3.2 Improving the Function and Viability of Islet Transplants ......................................5
1.4 Modular Tissue Engineering ................................................................................................7
1.4.1 Endothelial Progenitor Cells as an Alternative to HUVEC in Modular Tissue
Engineering ..............................................................................................................7
1.5 Islet Cell Aggregation ........................................................................................................10
1.5.1 Motivation for Re-aggregation of Islets .................................................................10
1.5.2 The Formation of Pseudoislets from Pancreatic Cell Lines...................................11
1.5.3 Pseudoislets from Mixed Islet Cell Types .............................................................12
1.5.4 Pseudoislets with Endothelial Cells .......................................................................12
Chapter 2 Methodology .................................................................................................................13
2 Methodology .............................................................................................................................13
2.1 Tissue Culture ....................................................................................................................13
2.2 Module Fabrication ............................................................................................................14

v
2.3 BOEC Characterization - Sprouting Assay........................................................................16
2.4 Immunofluorescence Staining and Imaging ......................................................................17
2.4.1 BOEC Morphology on Modules ............................................................................17
2.4.2 Morphology of αTC1-6 and MIN6 Cells in Modules ............................................17
2.5 In Vivo Vascularization ......................................................................................................18
2.5.1 Modules Coated in BOEC or HUVEC ..................................................................18
2.5.2 Modules Embedded with BOEC or HUVEC .........................................................18
2.6 Pseudoislet Modules In Vivo ..............................................................................................19
2.6.1 Diabetic Mice .........................................................................................................19
2.6.2 Pseudoislet Module Transplants ............................................................................19
2.6.3 Metabolic Follow Up .............................................................................................20
2.6.4 Histology ................................................................................................................20
Chapter 3 Results ...........................................................................................................................21
3 Results .......................................................................................................................................21
3.1 Modules Coated in BOEC or HUVEC ..............................................................................21
3.1.1 Sprouting Assay .....................................................................................................21
3.1.2 Morphology of EC Coated Modules ......................................................................22
3.1.3 In Vivo Vascularization of EC Coated Modules ....................................................23
3.2 Modules Embedded with BOEC or HUVEC .....................................................................29
3.2.1 Morphology of Modules Embedded with ECs ......................................................29
3.2.2 In Vivo Vascularization of Modules Embedded with ECs.....................................30
3.3 Pseudoislet Module Morphology .......................................................................................32
3.4 Pseudoislet Module Activity In Vivo ................................................................................34
3.4.1 Overview of Test Parameters and Treatment Groups ............................................34
3.4.2 Blood Glucose Measurements ...............................................................................35
3.4.3 Glucose Tolerance Test (GTT) ..............................................................................38

vi
3.4.4 Histology ................................................................................................................39
Chapter 4 Discussion .....................................................................................................................47
4 Discussion .................................................................................................................................47
4.1 BOEC vs. HUVEC in Modular Tissue Engineering ..........................................................47
4.2 The Effects of Embedding BOEC or HUVEC in Modules ...............................................48
4.3 Pseudoislet Modules ..........................................................................................................49
Chapter 5 Conclusions and Future Work .......................................................................................51
5 Conclusions and Future Work ...................................................................................................51
References ......................................................................................................................................53
Appendices .....................................................................................................................................65
Appendix A: Embedded BOEC Explants at Day 3 ...................................................................65
Appendix B: Pseudoislet Modules with Fewer MIN6 ..............................................................66
Appendix C: Glucagon Staining of Pseudoislet Modules .........................................................68

vii
List of Tables
Table 1: Culture media used for various cell types. ..................................................................... 13
Table 2: Cell make-up of module groups..................................................................................... 14
Table 3: Cell make-up of pseudoislet modules for diabetic animal studies. ................................ 19
Table 4: Size distribution of CD31+ vessels in implants with pseudoislet modules (n = 5) ....... 45
Table 5: Size distribution of UEA-1+ vessels in implants with pseudoislet modules (n = 5) ..... 45

viii
List of Figures
Figure 1: Summary of the islet isolation and transplantation procedure ....................................... 3
Figure 2: Representative confocal images of human and mouse islets. ........................................ 4
Figure 3: Diagram of a collagen module coated in cells ............................................................... 7
Figure 4: Summary of characteristics of early EPCs and OECs .................................................... 9
Figure 5: Module cutting set-up ................................................................................................... 15
Figure 6: Average sprout length and number for BOEC or HUVEC in fibrin assay. ................. 22
Figure 7: Confocal images of collagen modules coated in BOEC or HUVEC at day 7 ............. 23
Figure 8: Representative photographs of tissues with BOEC and HUVEC coated modules ...... 24
Figure 9: CD31 and UEA-1 staining for BOEC and HUVEC coated modules .......................... 25
Figure 10: H&E and Trichrome taining of BOEC and HUVEC coated modules ....................... 26
Figure 11: F4/80 staining for BOEC and HUVEC coated modules ............................................ 27
Figure 12: Total number of vessels on day 14 for BOEC and HUVEC modules ....................... 28
Figure 13: Number of UEA-1 vessels on day 14 for BOEC and HUVEC modules ................... 28
Figure 14: Confocal images of collagen modules embedded with BOEC or HUVEC at day 7 . 29
Figure 15: Representative photographs of tissues with EC embedded modules ......................... 30
Figure 16: Histology results for modules embedded with adMSC and BOEC or HUVEC ........ 31
Figure 17: Pseudoislet modules contract over one week ............................................................. 32
Figure 18: Pseudoislet module morphology using confocal microscopy .................................... 33
Figure 19: Summary of treatment groups used for diabetic mouse study. .................................. 34

ix
Figure 20: Average daily blood glucose measurements .............................................................. 36
Figure 21: Dot plots of blood glucose measurements on days 7, 14, and 21. ............................. 37
Figure 22: Line plot of glucose tolerance test results .................................................................. 38
Figure 23: Representative photographs of pseudoislet module tissues explanted at day 21 ....... 39
Figure 24: CD31 and UEA-1 staining for pseudoislet modules .................................................. 41
Figure 25: H&E and trichrome staining of pseudoislet modules................................................. 42
Figure 26: Insulin staining of pseudoislet modules ..................................................................... 43
Figure 27: CD31 vessel density and size for implants with pseudoislet modules ....................... 44
Figure 28: UEA-1 vessel density and size for implants with pseudoislet modules ..................... 44
Figure 29: Insulin pixel density for implants with pseudoislet modules at day 21 ..................... 46
Figure 30: Histology samples for modules embedded with adMSC and BOEC at day 3 ........... 65
Figure 31: Daily blood glucose for 50/50 MIN6/αTC1-6 mixed pseudoislet modules ............... 66
Figure 32: Representative histology images for 50/50 MIN6/αTC1-6 pseudoislet modules ...... 67
Figure 33: Glucagon staining for implants with pseudoislet modules after 21 days ................... 68

x
List of Appendices
Appendices .....................................................................................................................................65
Appendix A: Embedded BOEC Explants at Day 3 ...................................................................65
Appendix B: Pseudoislet Modules with Fewer MIN6 ..............................................................66
Appendix C: Glucagon Staining of Pseudoislet Modules .........................................................68
xi
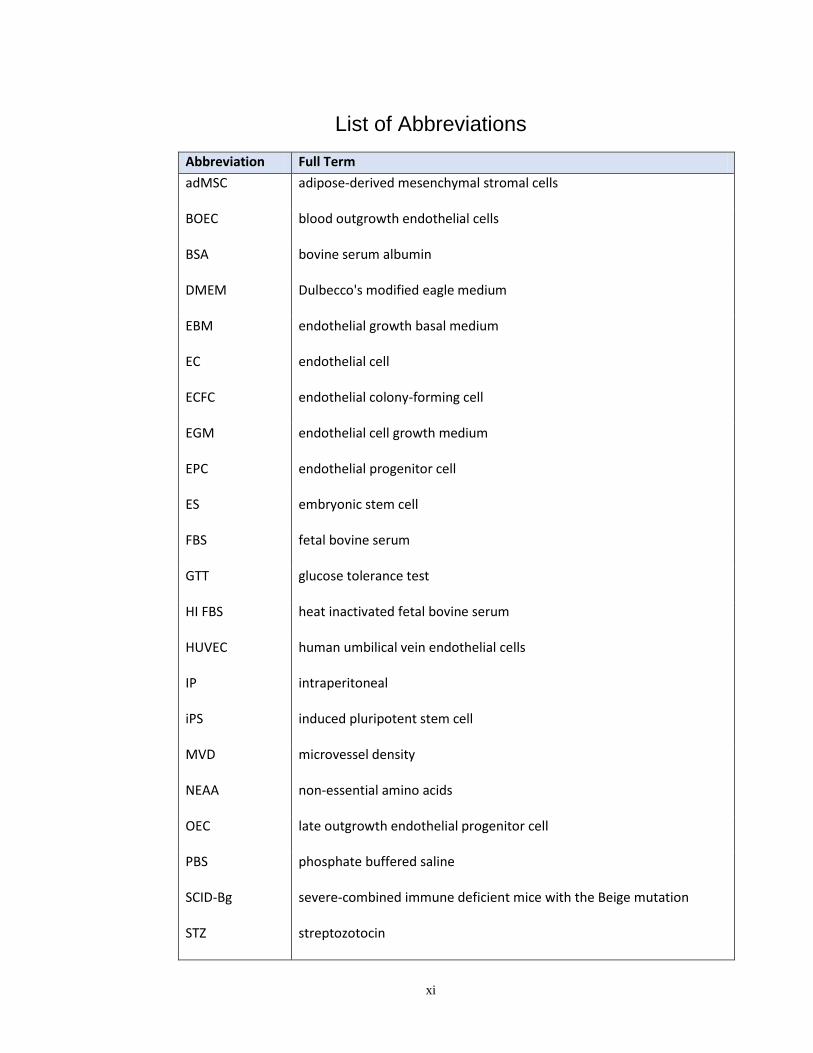
List of Abbreviations
Abbreviation Full Term
adMSC adipose-derived mesenchymal stromal cells
BOEC blood outgrowth endothelial cells
BSA bovine serum albumin
DMEM Dulbecco's modified eagle medium
EBM
EC
endothelial growth basal medium
endothelial cell
ECFC endothelial colony-forming cell
EGM
EPC
endothelial cell growth medium
endothelial progenitor cell
ES embryonic stem cell
FBS fetal bovine serum
GTT glucose tolerance test
HI FBS heat inactivated fetal bovine serum
HUVEC
IP
iPS
human umbilical vein endothelial cells
intraperitoneal
induced pluripotent stem cell
MVD microvessel density
NEAA non-essential amino acids
OEC late outgrowth endothelial progenitor cell
PBS
SCID-Bg
STZ
phosphate buffered saline
severe-combined immune deficient mice with the Beige mutation
streptozotocin

1
Chapter 1 Introduction
1 Introduction
1.1 Hypothesis and Research Objectives
In this thesis we investigate aggregating islet cells in modules to create pseudoislets that have
better vasculature and ability to reverse hyperglycemia. While HUVEC are currently used in
modules, an alternative endothelial cell option would be BOEC. Since the long-term goal is to
find a clinically viable means of treating diabetes, BOEC are preferable because they are
relatively easy to obtain and are autologous.
The hypothesis of this research project is that modules containing a mixture of α and β cells and
BOEC (pseudoislets) injected subcutaneously will result in vascularized insulin producing tissue
with the potential to restore normoglycemia in diabetic mice. In assessing this hypothesis, the
following aims will be met:
1. Show that BOEC can be used instead of HUVEC in standard modules without
significant impacts on function or morphology. Confocal microscopy and a sprouting
assay will be used to examine selected in vitro features of BOEC on modules. The in vivo
vasculature will be assessed histologically.
2. Determine whether BOEC can be embedded in modules or if they must be coated on
the outside. Histology and vessel counts will be used to compare vessel formation in
severe-combined immune deficient mice with the Beige mutation (SCID-Bg) mice
implanted with modules embedded with BOEC or coated with BOEC.
3. Evaluate the ability of modular pseudoislets to restore normoglycemia in diabetic
mice. Modules (containing the functional insulin generating capacity equivalent of 750
islets) will be subcutaneously injected into diabetic mice and glucose levels examined
over time to show the return to normoglycemia. This rat islet dose has been found to be
sufficient in some but not all animals.

2
1.2 Clinical Impetus
In 2014, it was reported that over 650 million people aged 18 or older worldwide were living
with diabetes [1]. In Canada alone, there are 10 million people living with diabetes and an
estimated 20 Canadians diagnosed with the disease every hour [2]. Diabetes continues to become
more prevalent and while treatments do exist, no cure has yet been developed. Of these, 10% are
cases of type 1 diabetes [2].
Diabetes is a chronic disease caused by either lack of insulin production or the inability of the
body to adequately use insulin produced in the body. These are referred to as type 1 and type 2
diabetes, respectively. More specifically, type 1 diabetes results when insulin producing cells
(i.e. β cells) are destroyed in the pancreas by the immune system [2]. Insulin is responsible for
metabolizing glucose in the body and without it, glucose accumulates to high levels in the blood.
Type 1 diabetes requires stringent monitoring of blood glucose levels and daily insulin injections
(sometimes more than once per day) or insulin pumps. This treatment allows people with type 1
diabetes to live, but it does not prevent the degenerative effects of diabetes such as blindness,
nerve damage, or vascular problems [3, 4]. Additionally, there can be stress from periods of
hypoglycemia, when blood glucose levels are too low, which can also be fatal [5]. Thus, there
exists strong motivation to develop better treatment options for type 1 diabetes.

3
1.3 Islet Transplantation and Limitations
1.3.1 Islet Transplantation Background
Developing a better means of treating this disease involves restoration of the body's natural
ability to produce insulin. One approach to this is pancreas transplantation but the patient must
take immunosuppressant drugs, weakening the immune system [6]. The actual transplantation
procedure itself also has a high level of risk associated with it [6].
Another option is pancreatic islet transplantation, albeit, still requiring immunosuppression (see
Figure 1 for a summary of the procedure). Islets of Langerhans, or islets, make up only 1 - 2% of
the pancreas and are responsible for regulating blood sugar in the body [7]. Islets are cell clusters
largely made up of insulin-producing β cells (about 60%) and glucagon-producing α cells (about
30%) [8]. The other cell types present are somatostatin-producing δ (<10%), pancreatic
polypeptide-producing γ cells (<5%), and ghrelin-producing ε cells [8]. Islets vary both in size
(between 0.5 – 1.3 cm3) and the total number of cells (between 3.6 x 10
6 – 14.8 x 10
6 cells) [8].
The organization and function of these cells are unique to species. In mice, for example, there is
a core of β cells surrounded by α cells, whereas other varying patterns have been reported in
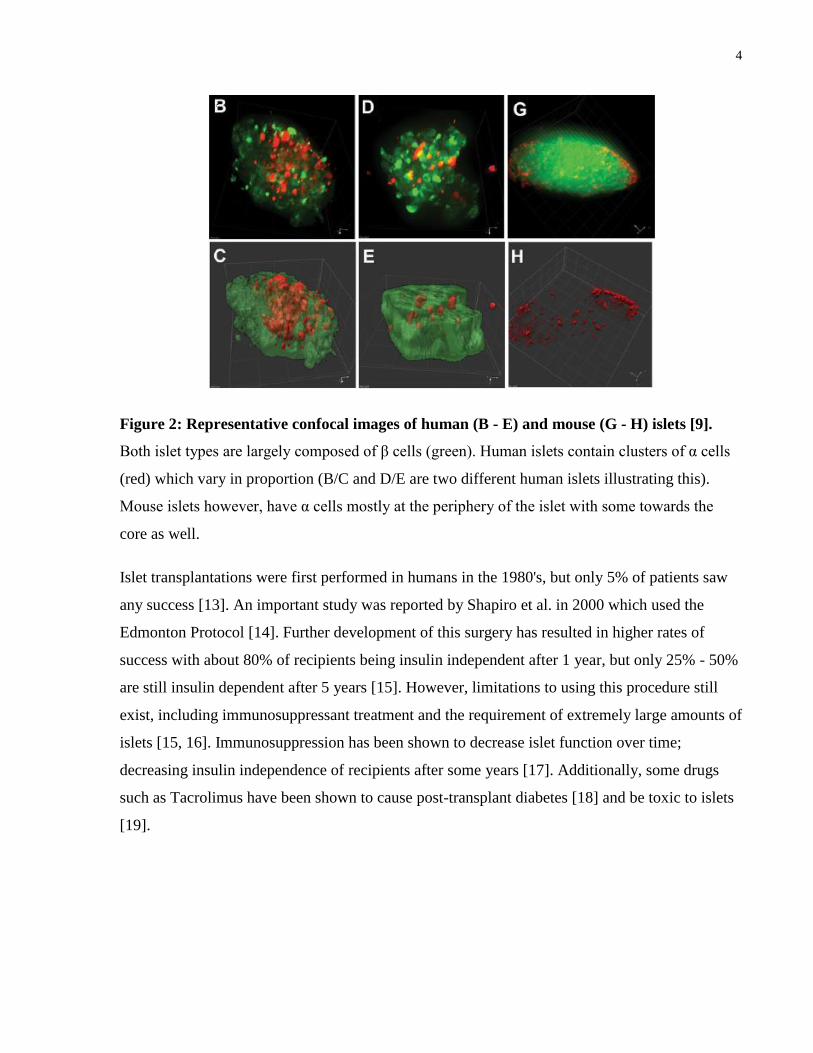
human islets [9, 10, 11]. Figure 2 shows some representative images of islets in these species.
Figure 1: Summary of the islet isolation and transplantation procedure [12].
4
Figure 2: Representative confocal images of human (B - E) and mouse (G - H) islets [9].
Both islet types are largely composed of β cells (green). Human islets contain clusters of α cells
(red) which vary in proportion (B/C and D/E are two different human islets illustrating this).
Mouse islets however, have α cells mostly at the periphery of the islet with some towards the
core as well.
Islet transplantations were first performed in humans in the 1980's, but only 5% of patients saw
any success [13]. An important study was reported by Shapiro et al. in 2000 which used the
Edmonton Protocol [14]. Further development of this surgery has resulted in higher rates of
success with about 80% of recipients being insulin independent after 1 year, but only 25% - 50%
are still insulin dependent after 5 years [15]. However, limitations to using this procedure still
exist, including immunosuppressant treatment and the requirement of extremely large amounts of
islets [15, 16]. Immunosuppression has been shown to decrease islet function over time;
decreasing insulin independence of recipients after some years [17]. Additionally, some drugs
such as Tacrolimus have been shown to cause post-transplant diabetes [18] and be toxic to islets
[19].

5
1.3.2 Improving the Function and Viability of Islet Transplants
1.3.2.1 Effects of Size on Islet Transplants
The literature agrees that smaller islets (generally of a size less than 100 μM) function better than
those of a larger size [20, 21]. More specifically, these smaller islets have been found to have a
greater viability (99.7% ± 0.1% vs. 72.4% ± 2.8%), increased glucose penetration (about 95% vs.
15 - 20%), increased insulin expression after exposure to high glucose levels, and better
maintenance of normoglycemia in diabetic rats [20]. Size affects the function of the islets
because upon isolation and transplantation, islets are no longer highly vascularized [16]. It is
more difficult for oxygen and nutrients to diffuse through larger islets which results in lower
viability and function [20]. Finding a means to revascularize islets would improve their viability
and function over time after transplantation.
1.3.2.2 The Importance of Vasculature on Islet Function and Viability
In their native environment, islets are close to major blood vessels and contain a highly
vascularized intraislet network as well to facilitate large amounts of blood flow. Islets have 5
times the amount of blood flow as the exocrine tissue relative to tissue mass and islet capillary
networks are 5 times denser than that found in the exocrine tissue [22]. Islet isolations result in
the loss of arterial and venous connections and thus, transplanted islets must rely on
revascularization from the recipient which may end up taking too long or be inadequate [23]. In
addition to connecting with host vasculature, intraislet endothelial cells have been found to
contribute to revascularization [23]. It is possible to preserve intraislet endothelial cells during
isolation, but a better method may be to transplant islets with supporting cells that drive
vascularization.

6
1.3.2.3 The Use of Biomaterials to Improve Islet Transplantation
Islet transplantation can also be improved by use of biomaterials to protect them from the body's
inflammatory response [21, 24] and improve their functionality [25, 26]. Encapsulating islets in
semi-permeable biomaterials can provide immunoisolation by preventing direct contact with the
host's immune cells and still allowing essential nutrients to pass through, eliminating the need for
immunosuppressive therapy [21, 27, 28]. A number of different materials, often alginate-based,
have shown promising results [26, 29]. For example, Duvivier-Kali et al. showed prolonged
survival of allogeneic and syngeneic islets over 350 days when encapsulated in barium-alginate
and without added immunosuppression [21].
However, these materials are generally not biocompatible, which may then result in poor islet
function from the formation of a pericapsular overgrowth from the host immune response [30].
The formation of fibrotic cell layers on the surface of the microcapsules by immune-mediated
foreign-body responses leads to donor tissue necrosis and nutrient isolation [30, 31, 32]. A
strategy to address this concern is the administration of anti-inflammatory drugs, but these drugs
have also been shown to negatively affect the function of the encapsulated islets [33]. Therefore,
a major concern for islet encapsulation moving forward to clinical trials is the use of a
biocompatible material that would protect the transplanted cells without eliciting an immune
response from the material itself [29, 34]. Recently, Vegas et al. evaluated a large number of
alginate variants for their ability to significantly decrease foreign-body responses in rodents and
non-human primates and found 3 triazole-containing alginates that were highly successful in
doing so [35].
In order to further improve islet transplantation, it is desirable to use small islets in the presence
of a vasculature while minimizing the immune and inflammatory response. Modular tissue
engineering presents a means of doing at least some of this by allowing for a high degree of
customization of cell type and biomaterials used.

7
1.4 Modular Tissue Engineering
Modular tissue engineering uses injectable microtissue structures, modules, which allow for the
creation of vascularized, scalable constructs (see Figure 3). Previously, the Sefton Lab developed
modules coated in human umbilical vein endothelial cells (HUVEC) using collagen embedded
with adipose-derived mesenchymal stem cells (adMSC) [36]. This model allows for the
combination of many additional cells types, while the HUVEC are able to form inter-connected
channels which enable blood perfusion. Subcutaneous transplants of these modular constructs
form vascular beds which allow for the delivery of therapeutic cells [36].
Figure 3: Diagram of a collagen module coated in cells. Each module is about 2 mm long
before shrinkage.
1.4.1 Endothelial Progenitor Cells as an Alternative to HUVEC in Modular Tissue Engineering
Modules are being studied as a means of transplanting islets as the endothelial cell (EC) coating
drives vessel formation around the implant, improving islet viability [16]. Additionally, coating
of islets with ECs have been shown to add protection from inflammatory reactions [37]. HUVEC
are currently used to coat modules, however, it is desirable to use an EC source that would be
more clinically viable. Some disadvantages of HUVEC are their limited life span and their
ability to rarely be isolated from patients with vascular abnormalities [38]. A particularly
interesting alternative is endothelial progenitor cells (EPCs).

8
Some fundamental properties of EPCs are the ability to form lumenized tubes in vitro, being a
circulating cell that gives rise to progeny with proliferative potential and restricted differentiation
for only those of an endothelial lineage, and the ability to form human blood vessels that become
integrated with host vasculature when implanted [39]. Since EPCs were first reported by Asahara
et al. in 1997 [40], much discussion and controversy has occurred over what an EPC is exactly;
unique cell surface markers have not been defined. Due to this, a number of different names exist
and often the methods used to isolate the cells are used to better describe them.
EPCs can be divided into 2 categories: early EPCs and blood outgrowth endothelial cells
(BOEC) (refer to Figure 4 for a summary of their characteristics). BOEC are also known as late
outgrowth EPCs (OEC) and endothelial colony-forming cells (ECFC). Early EPCs appear early
in culture, but they lack long-term proliferative potential, express hematopoietic markers, and
have been shown to differentiate into macrophages in vivo rather than forming vessels [41, 42].
In fact, studies have shown that early EPCs are actually hematopoietic cells - not true endothelial
cells - by phenotype [42, 43]. BOEC, on the other hand, do show a long-term ability to
differentiate in culture, express endothelial markers without hematopoietic markers, and can
form integrated vasculature in vivo [41, 44, 45]. Since both of these cell populations are present
in low numbers in the blood, ex vivo expansion would be required for any applications with
them. BOEC have been shown to have enhanced proliferation potential and can be kept longer in
culture [46]. Plus, BOEC can be obtained relatively easily from circulating EPCs in adult
peripheral blood, and can generate up to 1019
cells from just 50 - 100 mL of blood [44]. Given
these findings, we would want to use BOEC, not early EPCs in modular tissue engineering.
BOEC can be obtained from either peripheral blood or cord blood. While studies have shown
that cord blood BOEC can last longer in vivo and are more proliferative, they are much more
difficult to obtain. Given that BOEC, from either source, should function at least as well as
HUVEC [47, 48, 49, 50, 51, 52], it would be preferable to use BOEC from peripheral blood as it
presents an easily obtained autologous cell source for modules.

9
Figure 4: Summary of characteristics of early EPCs and OECs [53].
Evidence for the benefits of using BOEC in islet transplantation have also been reported. Co-
transplanting islets with BOEC has been shown to improve blood glucose levels in diabetic mice,
increase plasma C-peptide, reduce β cell death, and increase β cell proliferation independent of
the delivery technique [54, 55]. Tracing BOEC at these implant sites showed that BOEC did not
expand, but instead remained at the site, increasing their clinical potential [54]. Finally, BOEC
isolated from patients with type 1 diabetes were able to demonstrate the same benefits; this
confirms its ability to be used as an autologous cell source [54]. Since BOEC present a number
of possible advantages over HUVEC moving forward, they will be compared to HUVEC in
modules to confirm no adverse effects exist in this system.

10
1.5 Islet Cell Aggregation
1.5.1 Motivation for Re-aggregation of Islets
While improvements continue to be made in islet transplantation, a number of challenges still
persist, including: a shortage of donor islets, the loss of islet viability during isolation or upon
transplantation, the common need for multiple islet transplants per recipient, and the risk of
immunosuppressive therapies for the recipient and transplanted islets. Currently, 2 - 3 donor
pancreata are required for a single islet transplant [20]. Of these isolated islets, some may not be
able to be used for transplantation if they do not meet the required purity, viability, or function
standards [56]. Further strain is placed on these limited donor numbers when one considers that
most diabetic patients actually require multiple transplants to maintain insulin dependence over
time [57]. This is largely because at least 50% of the transplanted islet cells die within 10 weeks
after transplantation [57].
The main reason for this cell death is the loss of vasculature and blood supply upon isolation
[20]. Diffusion then becomes the main means for islets to receive oxygen, glucose, and other
nutrients. However, diffusion can be limited by the size of islets, which varies greatly. Studies
have shown larger islets (with diameters above 150 μm) had poorer viability and function than
islets with diameters under 125 μm [57, 58, 59]. Simply dispersing islet cells would decrease
these diffusion barriers, but also vastly decrease the function of these cells; islet cells rely on
paracrine interactions and cell-to-cell communication which are enhanced when cells are
clustered [20]
All the cells, or just the β cells, can be isolated from native islets and cultured in conditions
which allow them to re-associate into aggregates. These aggregates, termed pseudoislets, can
form structures with a similar morphology to native islets when in collagen [60]. Initial interest
in this activity was to further examine the makeup of islets, however, this also presents further
possibilities for addressing the concerns of islet transplantations mentioned above. Although islet
cells can spontaneously re-aggregate, they often form large clusters which face the same
diffusion limitations as native islets.

11
Ramachandran et al. was able to consistently form small pseudoislets (< 125 μm in diameter) by
allowing the dispersed cells to re-aggregate on a glass mold with microrecesses [20]. By
controlling the aggregation of islet cells, pseudoislets with greater viability and function can be
created to more efficiently take advantage of the donor islets available. Islet cell aggregates have
better oxygen-consumption rate, nuclei counts, and insulin-to-DNA ratio [25] and show
comparable ability to reduce blood glucose in diabetic mice when compared to native islets [20,
25]. They also have significantly higher viability and increased glucose diffusion [20]. Islet
aggregation can be controlled to produce pseudoislets more consistent in size than isolated islets.
Since this can be controlled, smaller pseudoislets can be created as well, which has been shown
to increase function and viability [20].
1.5.2 The Formation of Pseudoislets from Pancreatic Cell Lines
In order to address the concern of donor availability, pseudoislets can also be created by
aggregating insulin producing cell lines [61]. A number of options exist, including, MIN, INS-1,
βTC, HIT, and RIN [62]. For the purpose of the proof-of-principle experiments in this study,
mice were chosen since they are a low order animal model. MIN6 are a mouse insulinoma line
that have been shown to be responsive to glucose levels [62, 63]. Other groups have already
shown the ability of MIN6 to bring diabetic mice into a normoglycemic state [63]. Thus, since
MIN6 are commonly used and easily available to us, they were chosen as the insulin producing
cell line for this study.
Another possibility to form pseudoislets that would be better in future studies involves inducing
fully differentiated β cells from stem cells such as induced pluripotent stem cells (iPS) or
embryonic stem cells (ES). This would further reduce the need for large donor numbers. Studies
have already shown the ability to form pseudoislets from a number of stem cell sources including
human adult pancreatic endocrine progenitors [64] combined with MSCs from pancreatic islets
[65], adipose tissue [66], or bone marrow [67]. Recently, Vegas et al. used human stem cell-
derived β cells to form pseudoislets in biocompatible capsules made from triazole-
thiomorpholine dioxide (TMTD) alginate [68]. They successfully achieved long-term correction
of diabetic mice using these pseudoislets [68]. In conclusion, it appears that islet aggregates
provide improvements on their native counterparts, in part due to the ability to decrease their size
and the potential to reduce donor numbers, if starting with a stem cell source.

12
1.5.3 Pseudoislets from Mixed Islet Cell Types
While many groups have made pseudoislets from β cells alone, the literature suggests that islet
aggregates of mixed cells (i.e. α and other endocrine cells) perform at least as well, if not better
than, pure β cell pseudoislets [25, 69, 70]. Such mixed cell aggregates normalized glucose levels
faster, for example [70]. Additionally, metabolic control was lost faster in patients with pure β
islets compared to the mixed aggregates [70]. Since α cells are the other major cell type in islets,
it would be ideal to aggregate them with β cells to form better pseudoislets. In this study we have
used αTC1-6 cells which are a mouse cell line that produce glucagon and were readily available
to us.
1.5.4 Pseudoislets with Endothelial Cells
Islet aggregates from β cell and islet derived ECs have also been prepared and showed improved
insulin production and response to glucose in vitro compared to aggregates without the ECs [61].
Similarly, pseudoislets containing late outgrowth EPCs produced significantly more insulin in
response to a high glucose stimulus compared to the low condition in vitro, whereas pseudoislets
without the late outgrowth EPCs showed no response at all [71].
Given this data and the benefits reported of using BOEC in islet transplantation, it would be
expected that pseudoislets made with BOEC would be superior to those without. Although
modules are coated in ECs, simply embedding them may be sufficient for vascularization; the
ECs should migrate to the surface of the collagen [72]. This would allow pseudoislet modules to
more closely mimic the intravascular network found in native islets [23]. Data must still be
obtained comparing modules embedded with ECs rather than coated to confirm this theory. The
long-term goal for this research project is to make pseudoislets in modules since these would be
consistent in size, should improve islet cell viability, promote vasculature, and reduce the need of
a large number of islet donors. The initial work is to use the various cell lines noted in proof-of-
concept studies.
13
Chapter 2 Methodology
2 Methodology
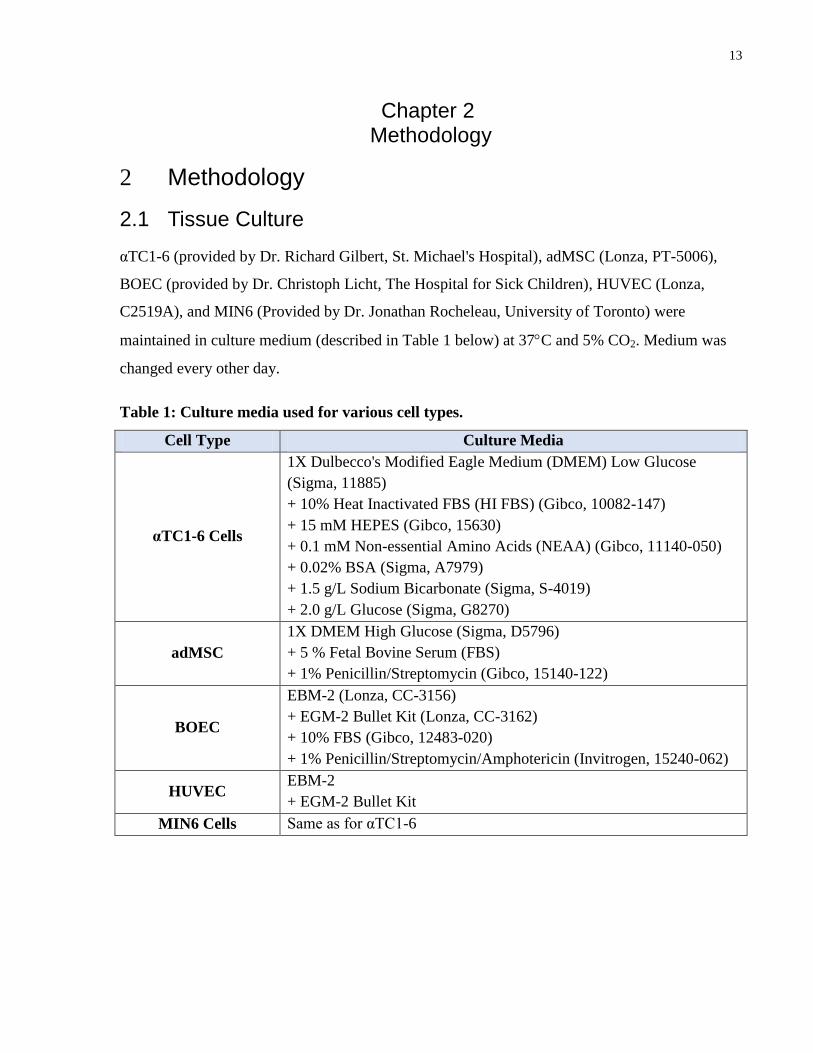
2.1 Tissue Culture
αTC1-6 (provided by Dr. Richard Gilbert, St. Michael's Hospital), adMSC (Lonza, PT-5006),
BOEC (provided by Dr. Christoph Licht, The Hospital for Sick Children), HUVEC (Lonza,
C2519A), and MIN6 (Provided by Dr. Jonathan Rocheleau, University of Toronto) were
maintained in culture medium (described in Table 1 below) at 37C and 5% CO2. Medium was
changed every other day.
Table 1: Culture media used for various cell types.
Cell Type Culture Media
αTC1-6 Cells
1X Dulbecco's Modified Eagle Medium (DMEM) Low Glucose
(Sigma, 11885)
+ 10% Heat Inactivated FBS (HI FBS) (Gibco, 10082-147)
+ 15 mM HEPES (Gibco, 15630)
+ 0.1 mM Non-essential Amino Acids (NEAA) (Gibco, 11140-050)
+ 0.02% BSA (Sigma, A7979)
+ 1.5 g/L Sodium Bicarbonate (Sigma, S-4019)
+ 2.0 g/L Glucose (Sigma, G8270)
adMSC
1X DMEM High Glucose (Sigma, D5796)
+ 5 % Fetal Bovine Serum (FBS)
+ 1% Penicillin/Streptomycin (Gibco, 15140-122)
BOEC
EBM-2 (Lonza, CC-3156)
+ EGM-2 Bullet Kit (Lonza, CC-3162)
+ 10% FBS (Gibco, 12483-020)
+ 1% Penicillin/Streptomycin/Amphotericin (Invitrogen, 15240-062)
HUVEC EBM-2
+ EGM-2 Bullet Kit
MIN6 Cells Same as for αTC1-6
14
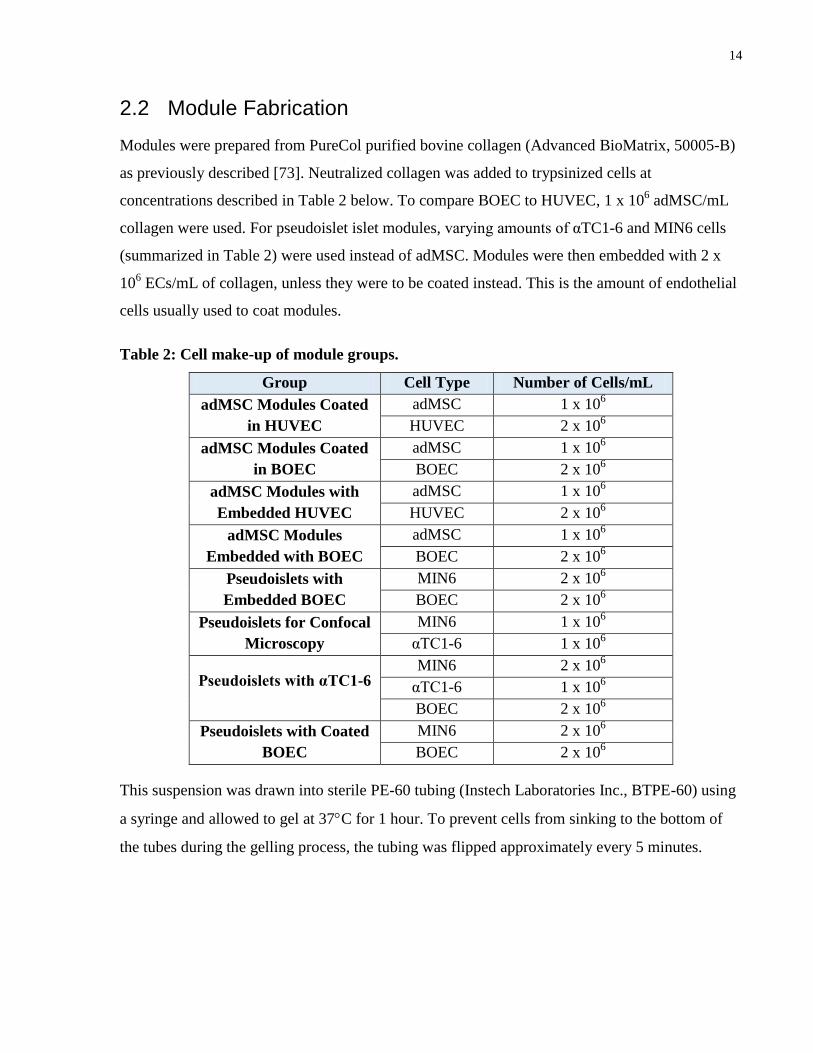
2.2 Module Fabrication
Modules were prepared from PureCol purified bovine collagen (Advanced BioMatrix, 50005-B)
as previously described [73]. Neutralized collagen was added to trypsinized cells at
concentrations described in Table 2 below. To compare BOEC to HUVEC, 1 x 106 adMSC/mL
collagen were used. For pseudoislet islet modules, varying amounts of αTC1-6 and MIN6 cells
(summarized in Table 2) were used instead of adMSC. Modules were then embedded with 2 x
106 ECs/mL of collagen, unless they were to be coated instead. This is the amount of endothelial
cells usually used to coat modules.
Table 2: Cell make-up of module groups.
Group Cell Type Number of Cells/mL
adMSC Modules Coated
in HUVEC
adMSC 1 x 106
HUVEC 2 x 106
adMSC Modules Coated
in BOEC
adMSC 1 x 106
BOEC 2 x 106
adMSC Modules with
Embedded HUVEC
adMSC 1 x 106
HUVEC 2 x 106
adMSC Modules
Embedded with BOEC
adMSC 1 x 106
BOEC 2 x 106
Pseudoislets with
Embedded BOEC
MIN6 2 x 106
BOEC 2 x 106
Pseudoislets for Confocal
Microscopy
MIN6 1 x 106
αTC1-6 1 x 106
Pseudoislets with αTC1-6 MIN6 2 x 10
6
αTC1-6 1 x 106
BOEC 2 x 106
Pseudoislets with Coated
BOEC
MIN6 2 x 106
BOEC 2 x 106
This suspension was drawn into sterile PE-60 tubing (Instech Laboratories Inc., BTPE-60) using
a syringe and allowed to gel at 37C for 1 hour. To prevent cells from sinking to the bottom of
the tubes during the gelling process, the tubing was flipped approximately every 5 minutes.
15
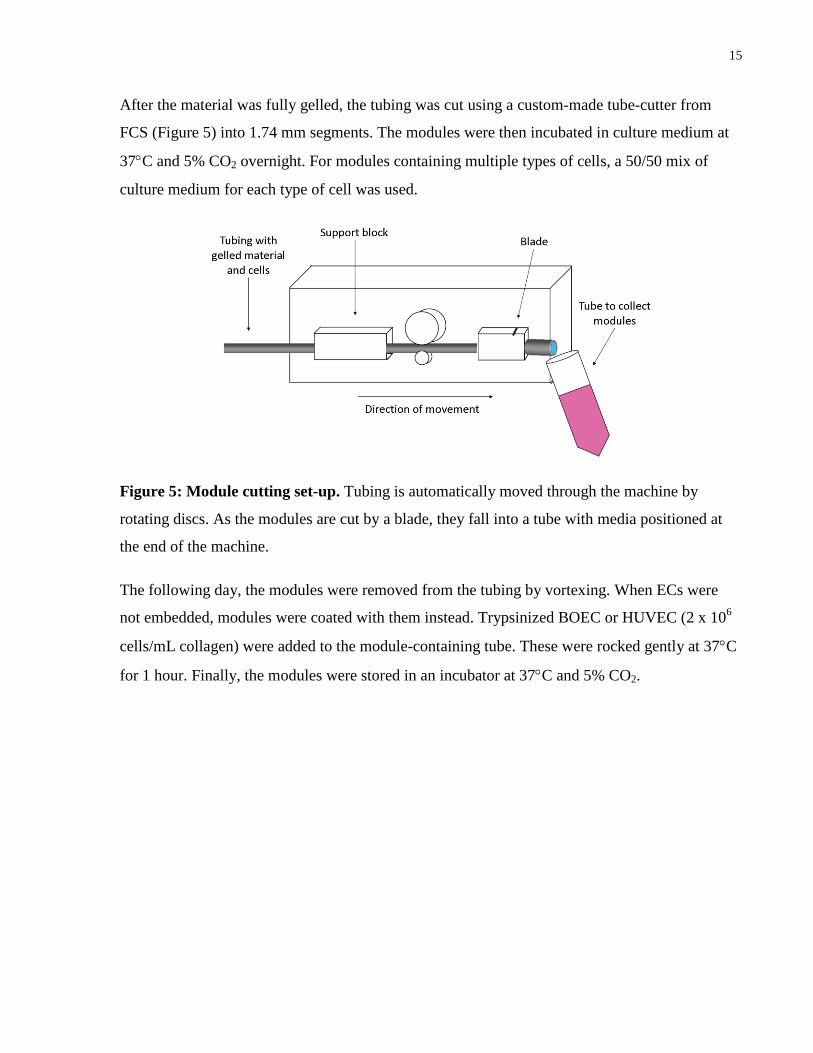
After the material was fully gelled, the tubing was cut using a custom-made tube-cutter from
FCS (Figure 5) into 1.74 mm segments. The modules were then incubated in culture medium at
37C and 5% CO2 overnight. For modules containing multiple types of cells, a 50/50 mix of
culture medium for each type of cell was used.
Figure 5: Module cutting set-up. Tubing is automatically moved through the machine by
rotating discs. As the modules are cut by a blade, they fall into a tube with media positioned at
the end of the machine.
The following day, the modules were removed from the tubing by vortexing. When ECs were
not embedded, modules were coated with them instead. Trypsinized BOEC or HUVEC (2 x 106
cells/mL collagen) were added to the module-containing tube. These were rocked gently at 37C
for 1 hour. Finally, the modules were stored in an incubator at 37C and 5% CO2.

16
2.3 BOEC Characterization - Sprouting Assay
Sprouting of BOEC from Cytodex-3 beads (Amersham Pharmacia, 17-0485-01) in fibrin gel was
compared to the same for HUVEC. Beads coated with BOEC or HUVEC were prepared one day
prior to the assay (i.e. day -1). About 2500 beads were mixed with 1 x 106 BOEC or HUVEC and
incubated at 37C for 4 hours. After this incubation, the coated beads were transferred to a non-
tissue culture Petri dish containing 5 mL of medium and incubated overnight.
The fibrin gel was prepared by adding 0.15 units/mL aprotinin (Sigma, A-1153) to a 2.0 mg/mL
fibinogen type 1 (Sigma, F-8630) solution on day 0. The coated beads were re-suspended in the
fibrinogen solution at a concentration of approximately 400 beads/mL. In a 24-well plate, 0.625
units/mL of thrombin (Sigma, T-3399) was added to each well. To this, 0.5 mL of the
fibrinogen-bead mixture was added and mixed. The plate was incubated at 37C for 10 - 15
minutes to generate a clot.
After this incubation time, medium was changed to a 50/50 mixture of BOEC or HUVEC
medium and adMSC medium. After incubation at 37C for 30 minutes, half of the wells had 1 x
105 adMSC added to them. The plate was stored at 37C and medium was changed every other
day. Images were obtained on days 1, 3, and 7 using a Zeiss Axiovert light microscope with a
5X objective lens and CCD camera. Sprouts were quantified by manually measuring and
counting in ImageJ.

17
2.4 Immunofluorescence Staining and Imaging
2.4.1 BOEC Morphology on Modules
Modules were transferred to a 15 mL tube and let settle. Culture medium was removed and
replaced with 2% paraformaldehyde solution in 1X phosphate buffered saline (PBS) (Gibco,
10010) to be fixed for 30 minutes.
Excess paraformaldehyde solution was removed and the modules were washed 3 times with
wash buffer (0.1% Tween-20 (Sigma, 9005-64-5) in PBS). Modules were resuspended in
blocking buffer (0.05% Tween-20 and 2% BSA in PBS) and blocked for 30 minutes.
Blocking buffer was removed by rinsing with PBS and goat anti-VE-cadherin primary antibody
(Santa Cruz Biotechnology, sc-6458) was added in a 1:50 dilution with incubation buffer (0.05%
Tween-20 in 1X PBS). Samples were rocked gently in the dark at 4C overnight.
The following day, primary antibody was removed, samples washed, and donkey anti-goat IgG
H&L secondary antibody (Alexa Fluor 405) (abcam, ab175664) was added at a 1:400 dilution.
Modules were incubated in the dark at 4C while stirring overnight.
After this time, secondary antibody was removed and modules were resuspended in PBS to be
imaged using a Nikon A1si confocal microscope (Centre for Microfluidic Systems in Chemistry
and Biology, University of Toronto). DAPI (Molecular Probes, R37606) was added immediately
before imaging.
2.4.2 Morphology of αTC1-6 and MIN6 Cells in Modules
Staining of pseudoislet modules was performed as described in the previous section. Primary
antibodies were monoclonal mouse anti-insulin (Sigma, I2018) and rabbit anti-glucagon (Sigma,
SAB4501137) at a 1:200 dilution. Secondary antibodies were goat anti-mouse IgG H&L Alexa
Fluor 488 (Life Technologies, A-11001) and goat anti-rabbit IgG H&L Texas Red (Life
Technologies, T-2767) at a 1:1500 dilution.

18
2.5 In Vivo Vascularization
2.5.1 Modules Coated in BOEC or HUVEC
Modules containing adMSC were prepared and coated with either BOEC or HUVEC and then
implanted into male 6 - 8 week old SCID-Bg mice (Charles River) to confirm the ability of
BOEC to form vasculature in tissue comparable to HUVEC. Each implant consisted of 1.5 mL of
collagen modules embedded with 2 x 106 adMSC/mL collagen embedded. Two million BOEC or
HUVEC per mL of collagen were used to coat the modules 1 hour after being cut. Modules were
incubated at 37C and 5% CO2 in culture media for 72 hours prior to subcutaneous implantation;
the number attached was not determined, but past experience using the same conditions has
indicated that EC are confluent by this time. The modules were subcutaneously delivered using
an 18-gauge needle in about 0.3 mL of PBS.
Tissues were explanted on days 3 and 14 and stained for CD31, UEA-1, H&E, Trichrome, and
f4/80 at Toronto General Hospital by the Pathology Research Laboratory. Slides were imaged
using a Zeiss Axiovert light microscope with a CCD camera. Microvessel density (MVD)
method counts were performed at 400X magnification in 5 representative areas and the average
of the counts obtained.
2.5.2 Modules Embedded with BOEC or HUVEC
Each implant consisted of 1.5 mL collagen modules embedded with 2 x 106 adMSC/mL and 2 x
106 BOEC/mL collagen. Modules were implanted, again after a 72 hour incubation period, and
explants analyzed as described in the above section.
19
2.6 Pseudoislet Modules In Vivo
2.6.1 Diabetic Mice
Adult male 6 - 8 week old SCID-Bg mice were individually housed and fed ad libitum. The mice
were rendered diabetic via intraperitoneal (IP) injection of 200 mg/kg streptozotocin (STZ)
(Sigma-Aldrich, S0130) in pH 4.5 citrate buffer (Sigma-Aldrich, 82585). After injection, the
animal’s water was replaced with filtered sucrose solution (10% w/v) to avoid the hypoglycemia
that may be caused by insulin being released from the STZ damaging the pancreas. The sucrose
solution was replaced with normal sterile water the following day.
Daily blood glucose measurements were obtained using blood from the tail vein and measured
with a glucometer (Onetouch Ultrasmart, LifeScan, Milpitas, CA). Animals that show
hyperglycemic blood glucose levels (those above 20 mM) for at least 3 consecutive days were
considered diabetic and subsequently used for transplantation studies. The study was approved
by the University of Toronto animal care committee.
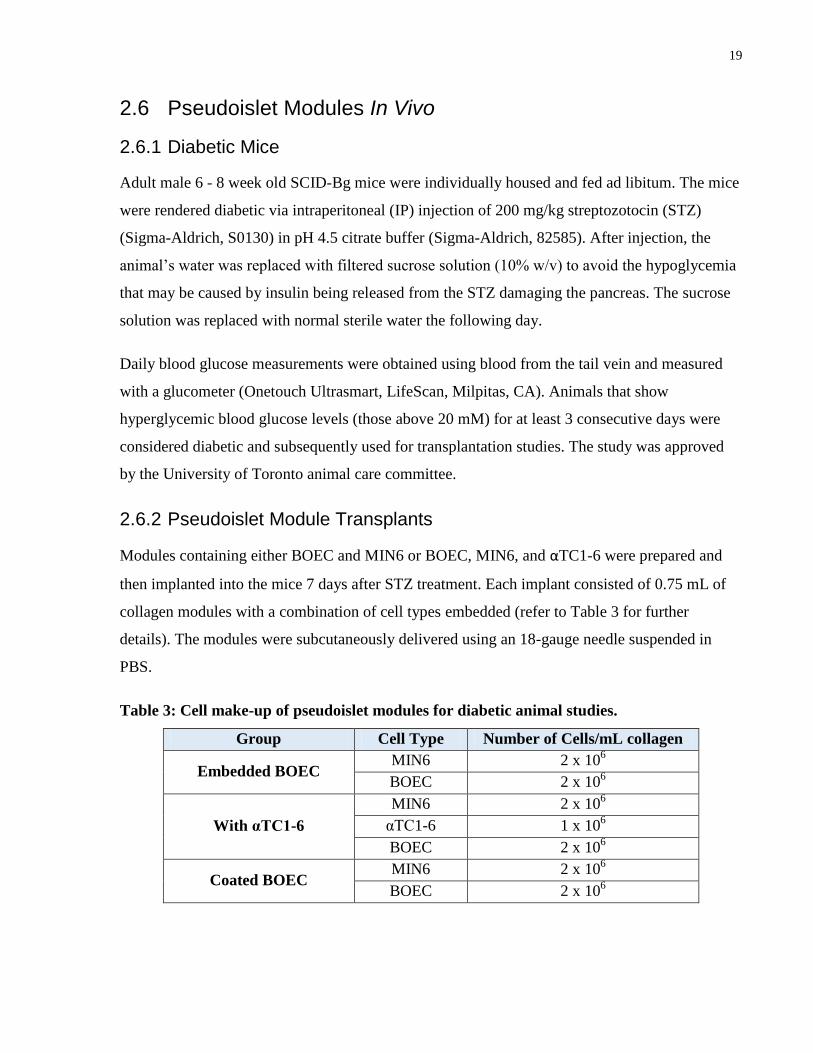
2.6.2 Pseudoislet Module Transplants
Modules containing either BOEC and MIN6 or BOEC, MIN6, and αTC1-6 were prepared and
then implanted into the mice 7 days after STZ treatment. Each implant consisted of 0.75 mL of
collagen modules with a combination of cell types embedded (refer to Table 3 for further
details). The modules were subcutaneously delivered using an 18-gauge needle suspended in
PBS.
Table 3: Cell make-up of pseudoislet modules for diabetic animal studies.
Group Cell Type Number of Cells/mL collagen
Embedded BOEC MIN6 2 x 10
6
BOEC 2 x 106
With αTC1-6
MIN6 2 x 106
αTC1-6 1 x 106
BOEC 2 x 106
Coated BOEC MIN6 2 x 10
6
BOEC 2 x 106

20
2.6.3 Metabolic Follow Up
Blood was sampled daily using the tail vein and blood glucose measured with a glucometer.
Fourteen days after module implantation, animals were subjected to a glucose tolerance test
(GTT). After 4 hours of fasting, a 2 g/kg glucose solution was administered via IP injection and
then blood glucose was measured from 0 - 120 minutes via the tail vein. Measurements were
taken at 0 minutes, 15 minutes, 30 minutes, 60 minutes, and 120 minutes.
2.6.4 Histology
Tissues were explanted 21 days after module implantation, fixed in 4% buffered formalin
(Sigma-Aldrich) for 48 hours and stained for CD31, UEA-1, H&E, Trichrome, Insulin, and
Glucagon at Toronto General Hospital by the Pathology Research Laboratory. Slides were
imaged using a Zeiss Axiovert light microscope with a CCD camera.
The total number of blood vessels (CD31+ staining) and the BOEC-derived blood vessels (UEA-
1+ staining) were manually counted at the implant site using digitized histology slides
(ScanScope XT brightfield scanner, Aperio Technologies) and the Aperio ImageScope software
(Aperio Technologies). The number of vessels in the implant area was normalized to the area
occupied by the implant on the whole histological section to obtain vessel density at the implant
site. The diameter of vessels was manually measured using the ImageScope software and
categorized based on their size as follows: < 9 µm capillaries, 9 – 15 µm small arterioles or
venules, 15 – 75 µm large arterioles or venules, and ≥ 75 µm for other (abnormal) as defined by
Chamberlain et al. previously [74]. The Positive Pixel Count Algorithm available with the
ImageScope software was used to determine the insulin density at the implant site.

21
Chapter 3 Results
3 Results
3.1 Modules Coated in BOEC or HUVEC
Currently, HUVEC are used in modules, and while they are sufficient to form a high degree of
vascularization, they are not ideal when thinking of a clinical setting. They are not autologous,
difficult to obtain, and limited in how much they can be expanded in culture. In contrast, BOEC
are highly proliferative, easily obtained, and can be expanded more in culture. Therefore, it is
desirable to switch to BOEC for future studies as they are more clinically relevant. Before doing
so, we first had to confirm that no adverse effects existed in the vasculature formed when using
BOEC to make modules.
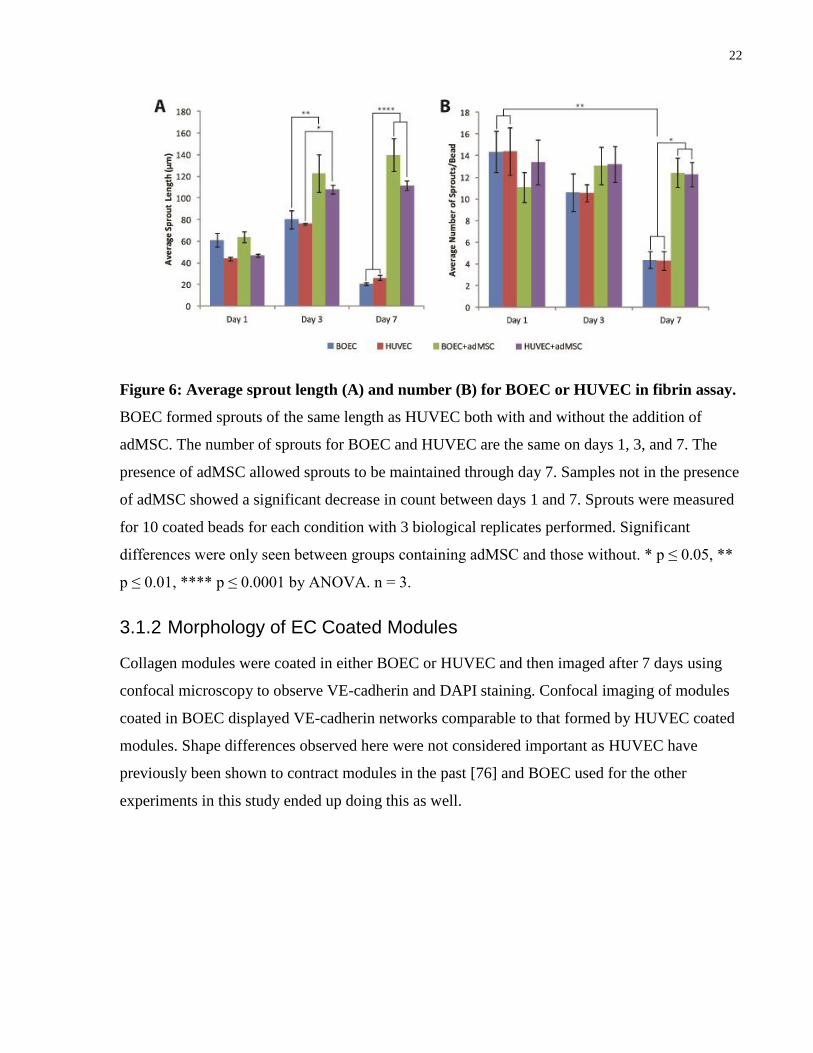
3.1.1 Sprouting Assay
Cytodex-3 beads were coated in either BOEC or HUVEC (1 x 106 cells/2500 beads) and placed
in a fibrin assay to compare resulting sprouts on days 1, 3, and 7. Half of the samples were
incubated in the presence of adMSC (1 x 105 adMSC/well). No significant differences in sprout
number or length (Figure 6) were observed in using BOEC rather than HUVEC (p ≤ 0.0001 by
ANOVA). As expected, samples in the presence of adMSC were able to maintain sprouts over a
longer period of time [75, 72].
22
Figure 6: Average sprout length (A) and number (B) for BOEC or HUVEC in fibrin assay.
BOEC formed sprouts of the same length as HUVEC both with and without the addition of
adMSC. The number of sprouts for BOEC and HUVEC are the same on days 1, 3, and 7. The
presence of adMSC allowed sprouts to be maintained through day 7. Samples not in the presence
of adMSC showed a significant decrease in count between days 1 and 7. Sprouts were measured
for 10 coated beads for each condition with 3 biological replicates performed. Significant
differences were only seen between groups containing adMSC and those without. * p ≤ 0.05, **
p ≤ 0.01, **** p ≤ 0.0001 by ANOVA. n = 3.
3.1.2 Morphology of EC Coated Modules
Collagen modules were coated in either BOEC or HUVEC and then imaged after 7 days using
confocal microscopy to observe VE-cadherin and DAPI staining. Confocal imaging of modules
coated in BOEC displayed VE-cadherin networks comparable to that formed by HUVEC coated
modules. Shape differences observed here were not considered important as HUVEC have
previously been shown to contract modules in the past [76] and BOEC used for the other
experiments in this study ended up doing this as well.
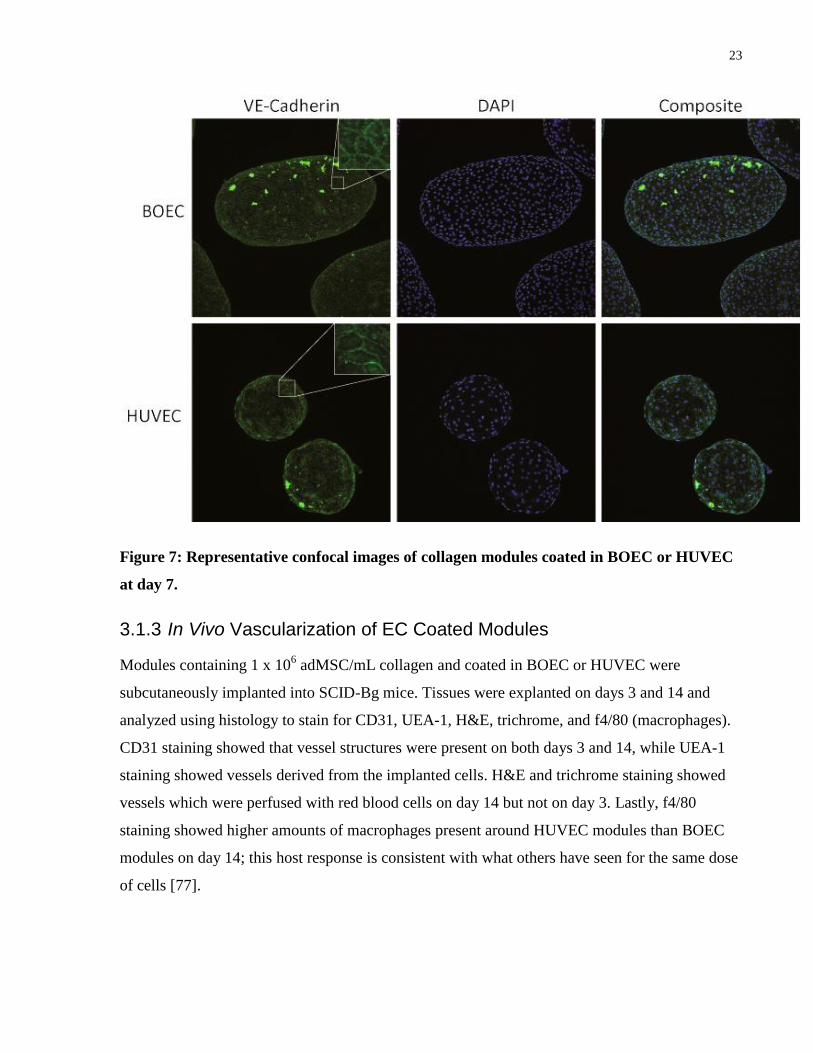
23
Figure 7: Representative confocal images of collagen modules coated in BOEC or HUVEC
at day 7.
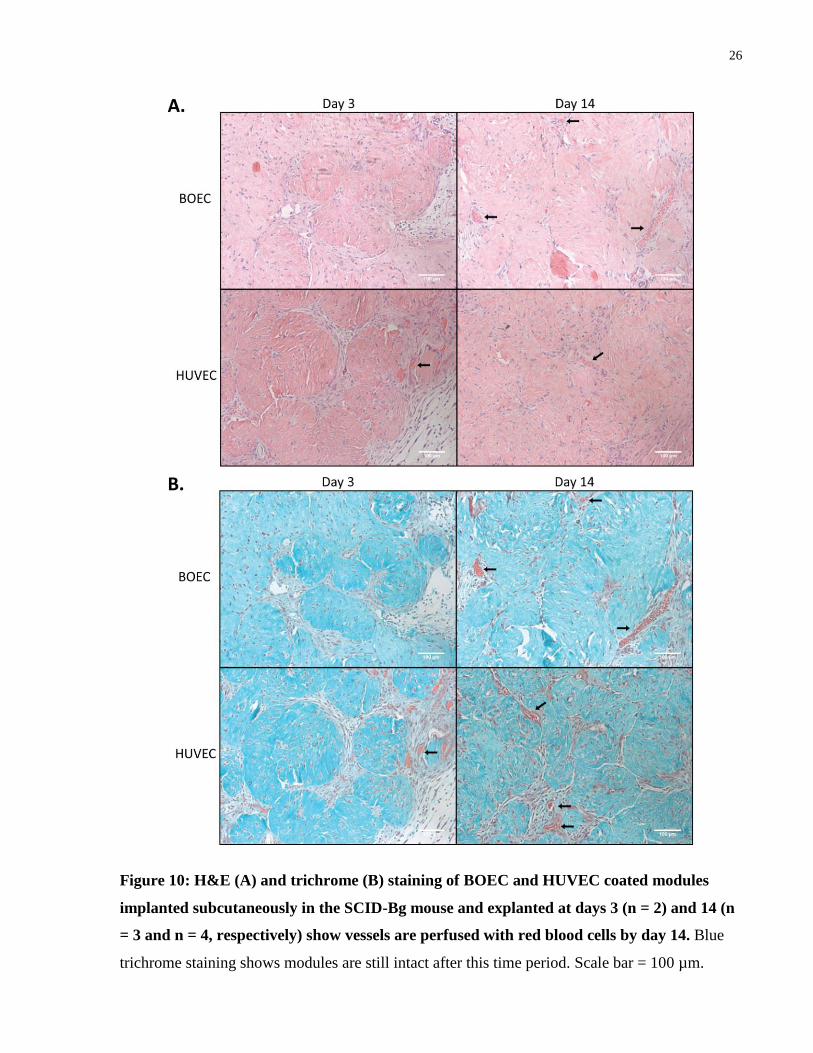
3.1.3 In Vivo Vascularization of EC Coated Modules
Modules containing 1 x 106 adMSC/mL collagen and coated in BOEC or HUVEC were
subcutaneously implanted into SCID-Bg mice. Tissues were explanted on days 3 and 14 and
analyzed using histology to stain for CD31, UEA-1, H&E, trichrome, and f4/80 (macrophages).
CD31 staining showed that vessel structures were present on both days 3 and 14, while UEA-1
staining showed vessels derived from the implanted cells. H&E and trichrome staining showed
vessels which were perfused with red blood cells on day 14 but not on day 3. Lastly, f4/80
staining showed higher amounts of macrophages present around HUVEC modules than BOEC
modules on day 14; this host response is consistent with what others have seen for the same dose
of cells [77].
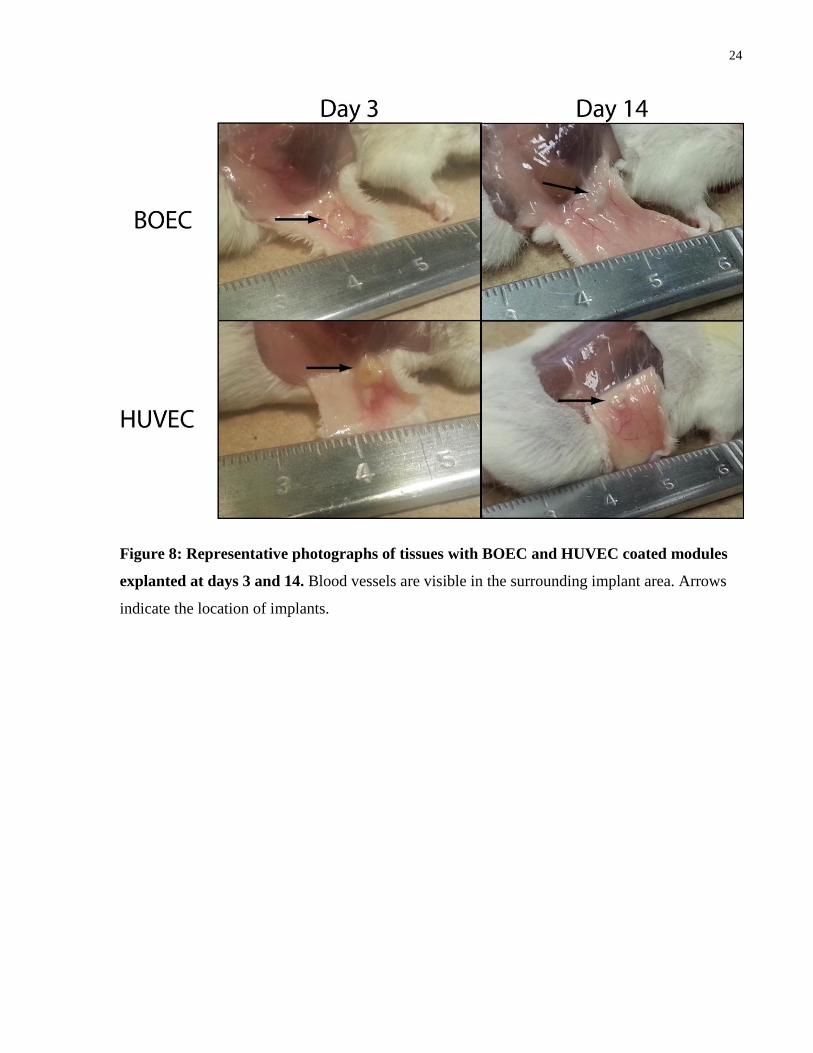
24
Figure 8: Representative photographs of tissues with BOEC and HUVEC coated modules
explanted at days 3 and 14. Blood vessels are visible in the surrounding implant area. Arrows
indicate the location of implants.

25
Figure 9: CD31 (A) and UEA-1 (B) staining for BOEC and HUVEC coated modules
implanted subcutaneously in the SCID-Bg mouse and explanted at days 3 (n = 2) and 14 (n
= 3 and n = 4, respectively). CD31 staining shows the formation of vessel-like structures around
the modules. UEA-1 staining shows vessel structures formed by the implanted human ECs. Scale
bar = 100 µm.
26
Figure 10: H&E (A) and trichrome (B) staining of BOEC and HUVEC coated modules
implanted subcutaneously in the SCID-Bg mouse and explanted at days 3 (n = 2) and 14 (n
= 3 and n = 4, respectively) show vessels are perfused with red blood cells by day 14. Blue
trichrome staining shows modules are still intact after this time period. Scale bar = 100 µm.

27
Figure 11: F4/80 staining for BOEC and HUVEC coated modules implanted
subcutaneously in the SCID-Bg mouse and explanted at days 3 (n = 2) and 14 (n = 3 and n =
4, respectively) shows macrophages were not present around the implant site at day 3. A
higher presence of macrophages were observed around the HUVEC implant site compared to the
BOEC implant site. Scale bar = 100 µm.
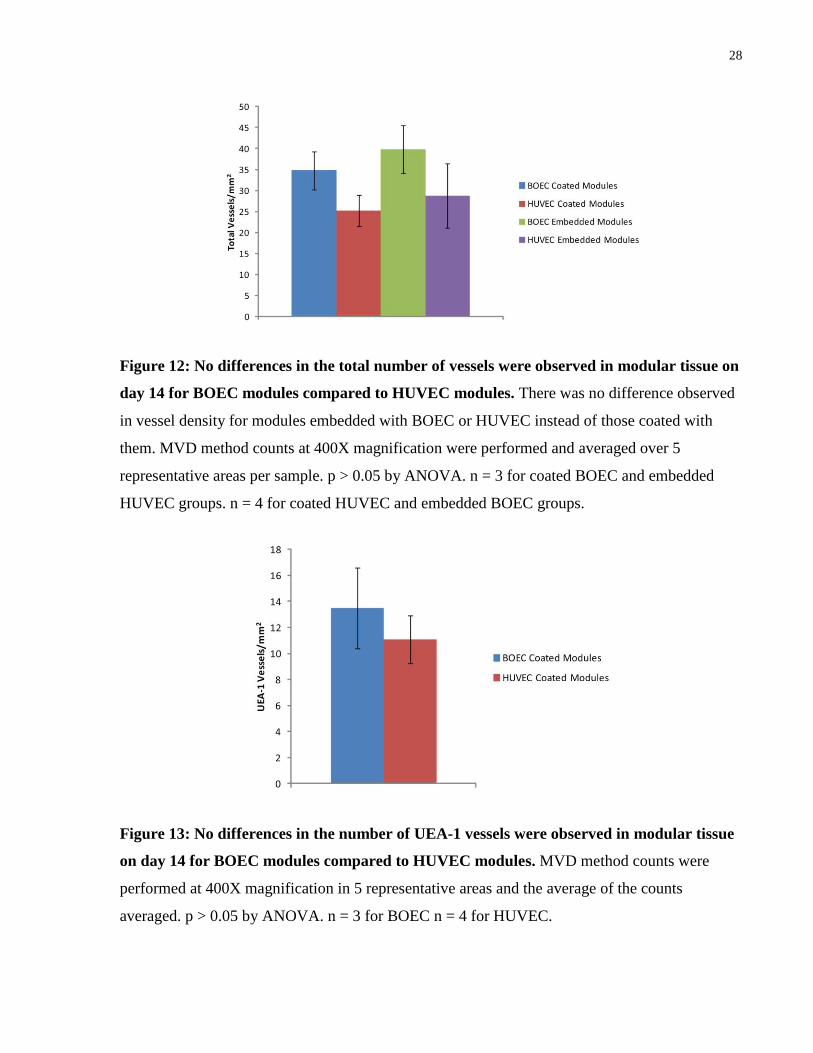
Vessel counts were performed for day 14 samples to better compare the degree of vascularization
of BOEC modules against HUVEC modules. Tissues implanted with BOEC modules were
shown to have no significant differences in both the number of total vessels and UEA-1 vessels
(p > 0.05 by ANOVA). The embedded EC groups are discussed below.
28
Figure 12: No differences in the total number of vessels were observed in modular tissue on
day 14 for BOEC modules compared to HUVEC modules. There was no difference observed
in vessel density for modules embedded with BOEC or HUVEC instead of those coated with
them. MVD method counts at 400X magnification were performed and averaged over 5
representative areas per sample. p > 0.05 by ANOVA. n = 3 for coated BOEC and embedded
HUVEC groups. n = 4 for coated HUVEC and embedded BOEC groups.
Figure 13: No differences in the number of UEA-1 vessels were observed in modular tissue
on day 14 for BOEC modules compared to HUVEC modules. MVD method counts were
performed at 400X magnification in 5 representative areas and the average of the counts
averaged. p > 0.05 by ANOVA. n = 3 for BOEC n = 4 for HUVEC.
29
3.2 Modules Embedded with BOEC or HUVEC
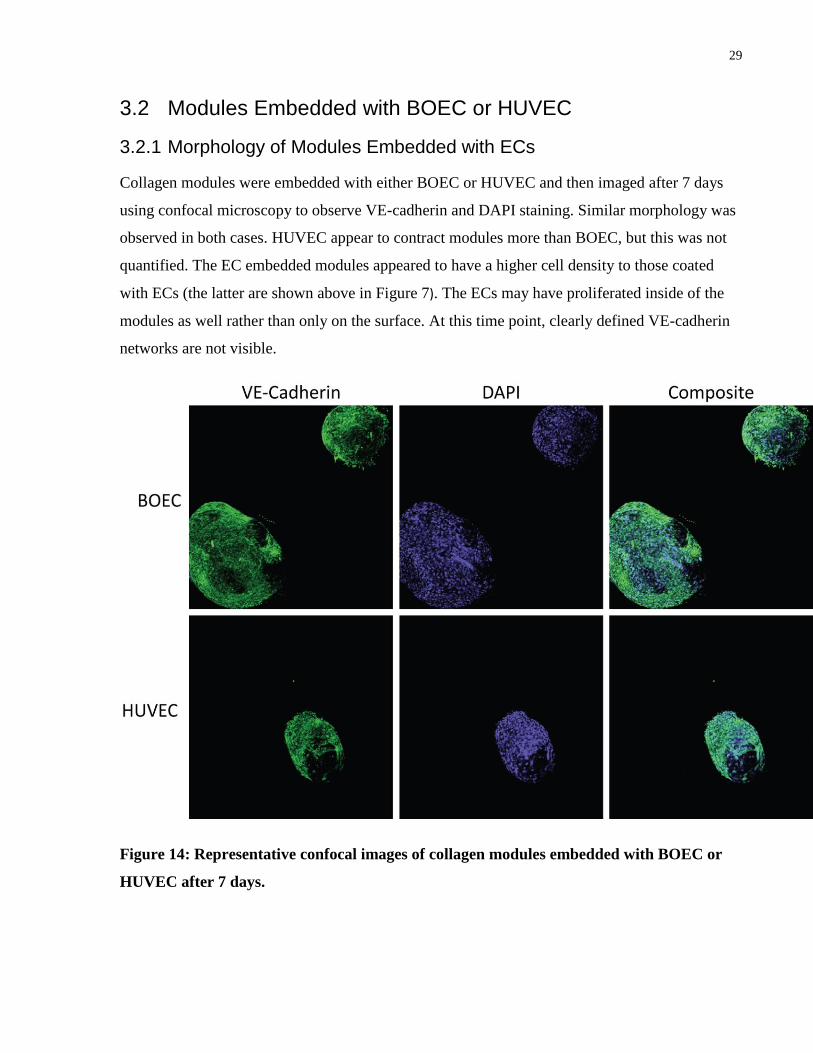
3.2.1 Morphology of Modules Embedded with ECs
Collagen modules were embedded with either BOEC or HUVEC and then imaged after 7 days
using confocal microscopy to observe VE-cadherin and DAPI staining. Similar morphology was
observed in both cases. HUVEC appear to contract modules more than BOEC, but this was not
quantified. The EC embedded modules appeared to have a higher cell density to those coated
with ECs (the latter are shown above in Figure 7). The ECs may have proliferated inside of the
modules as well rather than only on the surface. At this time point, clearly defined VE-cadherin
networks are not visible.
Figure 14: Representative confocal images of collagen modules embedded with BOEC or
HUVEC after 7 days.

30
3.2.2 In Vivo Vascularization of Modules Embedded with ECs
Modules containing 1 x 106 adMSC/mL collagen and about 2 x 10
6 BOEC or HUVEC per mL
collagen were subcutaneously implanted into SCID-Bg mice. Tissues were explanted on day 14
and analyzed using histology to stain for CD31, H&E, Trichrome, and f4/80. Tissues implanted
with embedded BOEC modules were also explanted at day 3 (see Appendix A: Embedded
BOEC Explants at Day 3). As seen for modules coated with ECs, the vessels formed were not
yet integrated with the host network at day 3.
Figure 15: Representative photographs of tissues explanted at day 14. Blood vessels are
visible in the surrounding implant area. Arrows indicate the location of implants.
It is evident from the histology images at day 14 (Figure 16) that modules embedded with ECs
rather than being coated with them are still able to vascularize mouse tissue in vivo. Blood
vessels formed around the modules in the implant area as shown by CD31 staining. By day 14,
H&E and trichrome stained red blood cells were visible inside some of these vessels which
suggests they were perfused and integrated with the host vasculature. These same observations
were made with modules coated in ECs, as seen above. Vessel counts shown in Figure 12
confirmed that there were no significant differences in vessel density for modules embedded
with ECs compared to those coated in ECs (p > 0.05 by ANOVA).
F4/80 staining was inconsistent between animals. For 2 BOEC animals, there was much f4/80
staining visible but almost none for the other 2 animals. One HUVEC animal had essentially no
f4/80 staining while the other 2 animals had some. There was no observable difference in the
number of vessels. As a result, it is hard to draw any conclusions on macrophage presence.
Overall, these results suggest that embedding ECs within collagen modules is sufficient for
vascularization, simplifying the process of module fabrication.

31
Figure 16: Histology results for modules embedded with adMSC and BOEC or HUVEC
implanted subcutaneously in the SCID-Bg mouse and explanted at day 14. CD31 staining
shows the formation of vessel-like structures around the modules. H&E and trichrome stains
show vessels perfused with erythrocytes visible in red. F4/80 staining shows higher amounts of
macrophages for BOEC than for HUVEC in half the animals. Scale bar = 100 µm. n = 3 for
HUVEC and n = 4 for BOEC.
32
3.3 Pseudoislet Module Morphology
Pseudoislet modules were prepared by embedding a 50/50 mix of αTC1-6 and MIN6 cell lines
and coating the modules in BOEC. It was observed that MIN6 proliferate faster on collagen in
vitro, and thus the initial 50/50 mix resulted in about a 30/70 ratio of αTC1-6 to MIN6 by day 7.
From fabrication to day 7, the pseudoislet modules contracted as expected (Figure 17).
Figure 17: Pseudoislet modules contract over one week. Shown in the figure are modules at
5X magnification on days 0 (A), 2 (B), 5 (C), and 7 (D). Scale bars represent 200 μm.
Native murine islets contain a core primarily consisting of β cells with α cells around the
periphery of the islet [9, 11, 78]. However, the α cells do not form a complete ring; some β cells
are exposed to the outside of the islet and some α cells can be found closer to the core [9, 78].
Confocal imaging was used to visualize the cells in the pseudoislet modules. The arrangement
was similar to that seen in another study which used MIN6 and αTC1 cells to form pseudoislets
(albeit without collagen) [78]. Here, the modules had a larger number of α cells in the core.
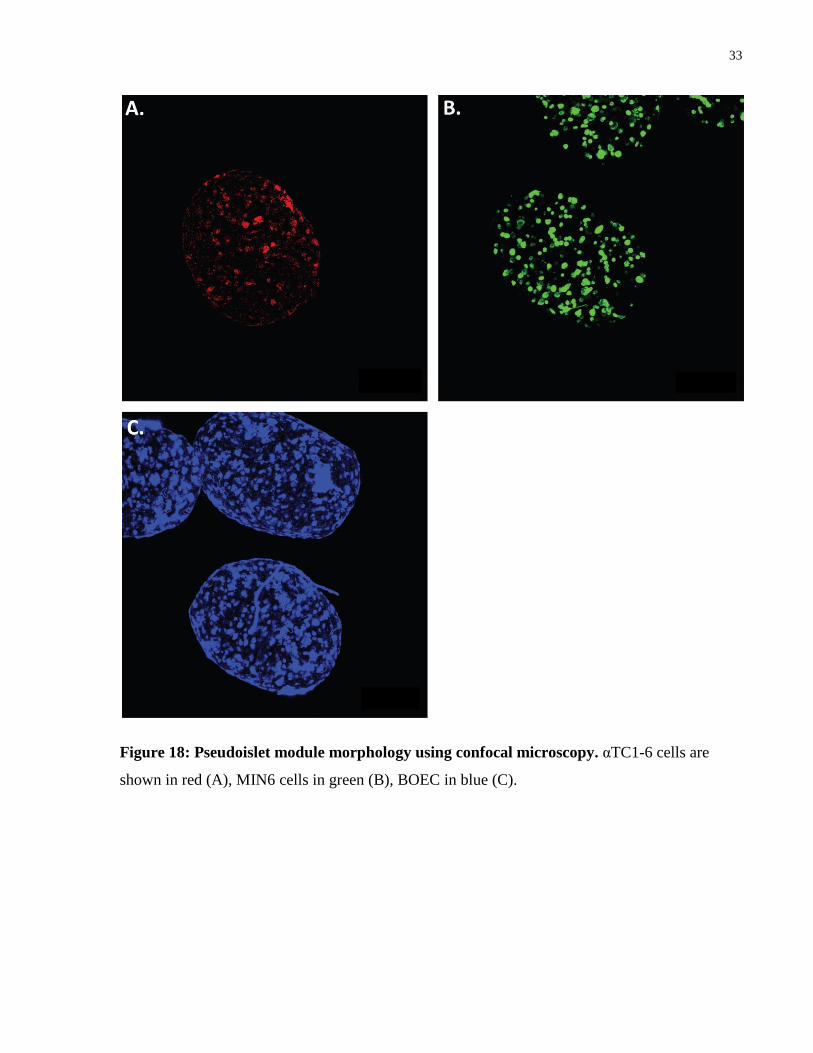
33
Figure 18: Pseudoislet module morphology using confocal microscopy. αTC1-6 cells are
shown in red (A), MIN6 cells in green (B), BOEC in blue (C).

34
3.4 Pseudoislet Module Activity In Vivo
3.4.1 Overview of Test Parameters and Treatment Groups
SCID-Bg mice were rendered diabetic via IP injection of STZ. Seven days later, animals with
blood glucose measurements above 20 mM for 3 consecutive days were considered diabetic and
subcutaneously implanted with 0.75 mL of collagen modules containing cells as described in
Table 3 and Figure 19. Blood glucose was measured daily, and tissues were explanted 21 days
after implantation for histology studies. In addition, a glucose tolerance test was performed on
day 14 to determine whether MIN6 regulated insulin production in response to a glucose
stimulus.
In evaluating the activity of pseudoislet modules in diabetic SCID-Bg mice, the following 2
parameters were investigated:
1. The presence of αTC1-6 cells
2. Embedding vs. coating modules with BOEC
Thus, 3 treatment groups were used as summarized in Figure 19 below:
Figure 19: Summary of treatment groups used for diabetic mouse study.
One final group containing half as many MIN6 in a 50/50 split with αTC1-6 and embedded
BOEC was also tested. A summary of the results for this group can be found in Appendix B:
Pseudoislet Modules with Fewer MIN6.

35
3.4.2 Blood Glucose Measurements
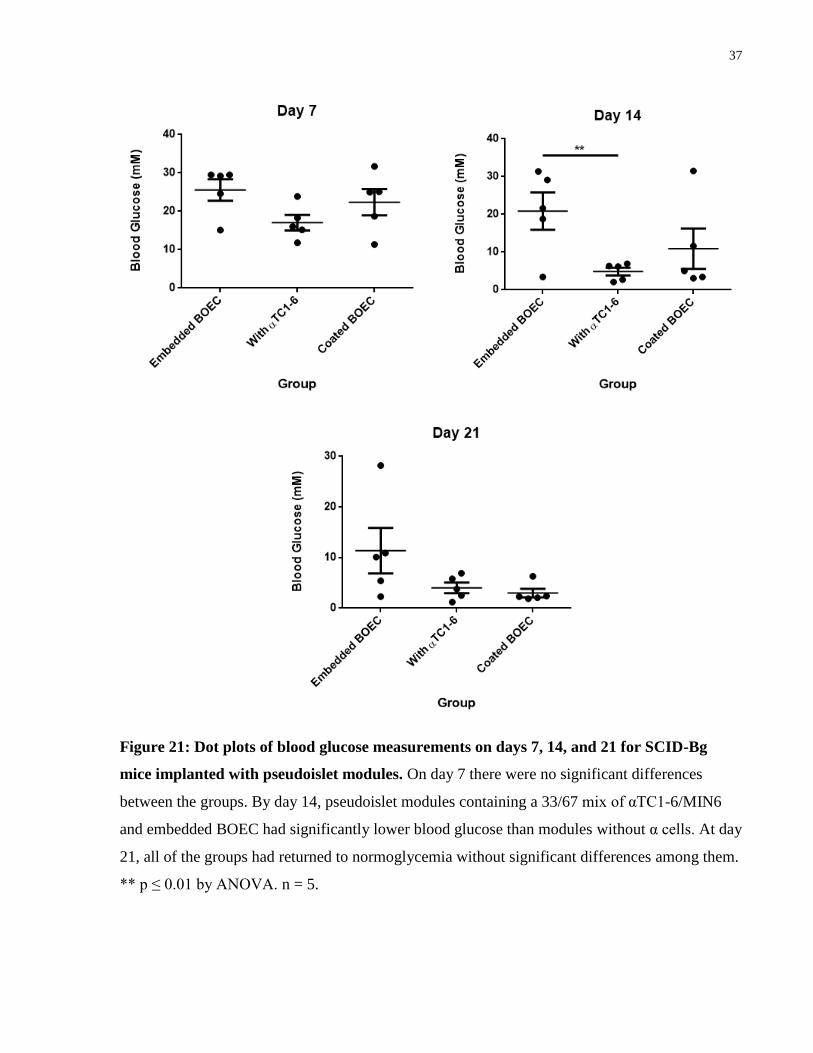
Blood glucose was measured daily for diabetic SCID-Bg mice implanted with pseudoislet
modules containing a variety of cell types. Daily blood glucose measurements showed that the
coated module group and the group with αTC1-6 were able to return to normoglycemia after
about 8 days (Figure 20). Embedded BOEC showed a delay in decreasing the blood glucose
levels and likely would have reached normoglycemia just after 21 days (Figure 20). After 2
weeks, the group with αTC1-6 had significantly lower blood glucose levels compared to those
without the αTC1-6 cells, but no statistical differences were seen with the BOEC coated group (p
> 0.05 by ANOVA) (Figure 21). By day 21, all of the groups were statistically the same,
although the average value of blood glucose for the embedded BOEC modules was not
considered normoglycemic (Figure 21).
Modules coated with BOEC had a faster and more drastic effect on blood glucose levels
compared to those embedded with BOEC, returning animals to normoglycemia approximately 2
weeks sooner. The collagen modules contracted due to traction forces exerted by the BOEC,
mostly in the axial direction as seen in previous studies done with HUVEC [79]. However, it
appeared that MIN6 modules coated in BOEC, contracted more than those with BOEC
embedded. This contraction increased cell density within the transplant and may have some
benefits on the vasculature [47] or function of MIN6.
The addition of αTC1-6 cells in a 33/67 ratio with MIN6 cells appeared to be beneficial, as
animals implanted with these modules showed significantly lower blood glucose on day 14 when
compared to modules without the αTC1-6 (p ≤ 0.01 on day 14 by ANOVA) (Figure 21). The
return to normoglycemia for modules with αTC1-6 was approximately the same, or even faster,
than that for modules coated in BOEC.
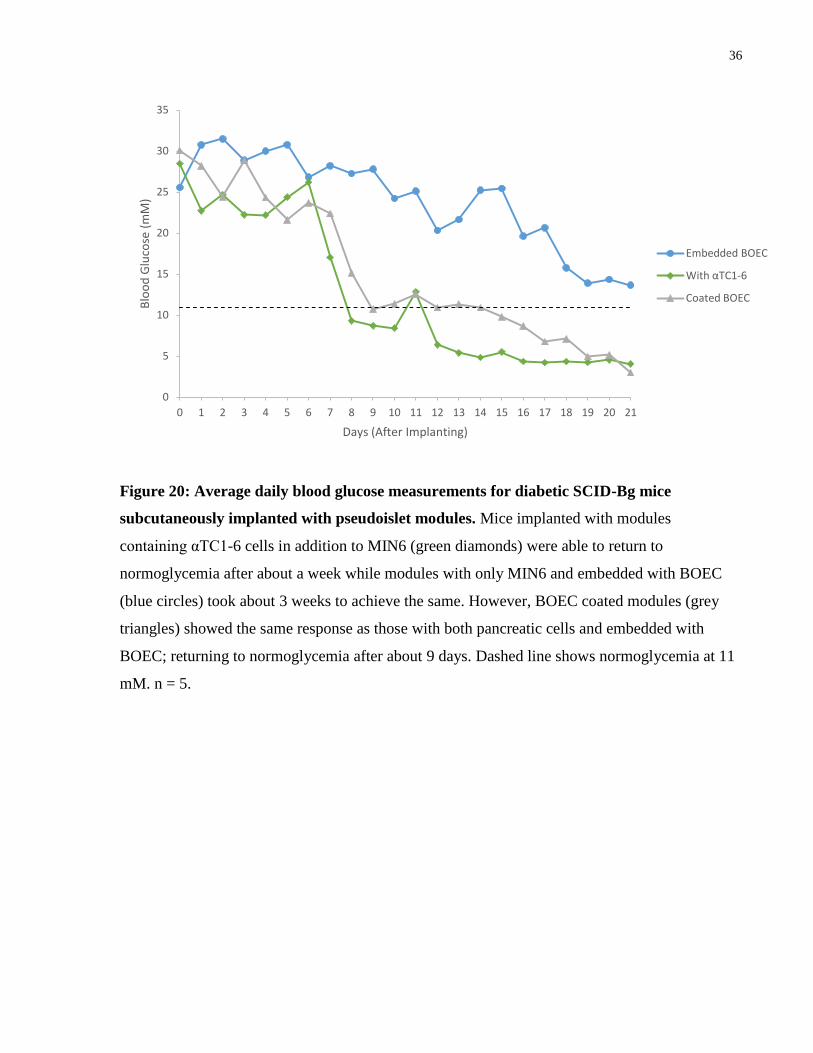
36
Figure 20: Average daily blood glucose measurements for diabetic SCID-Bg mice
subcutaneously implanted with pseudoislet modules. Mice implanted with modules
containing αTC1-6 cells in addition to MIN6 (green diamonds) were able to return to
normoglycemia after about a week while modules with only MIN6 and embedded with BOEC
(blue circles) took about 3 weeks to achieve the same. However, BOEC coated modules (grey
triangles) showed the same response as those with both pancreatic cells and embedded with
BOEC; returning to normoglycemia after about 9 days. Dashed line shows normoglycemia at 11
mM. n = 5.
0
5
10
15
20
25
30
35
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21
Blo
od
Glu
cose
(m
M)
Days (After Implanting)
Embedded BOEC
With αTC1-6
Coated BOEC
37
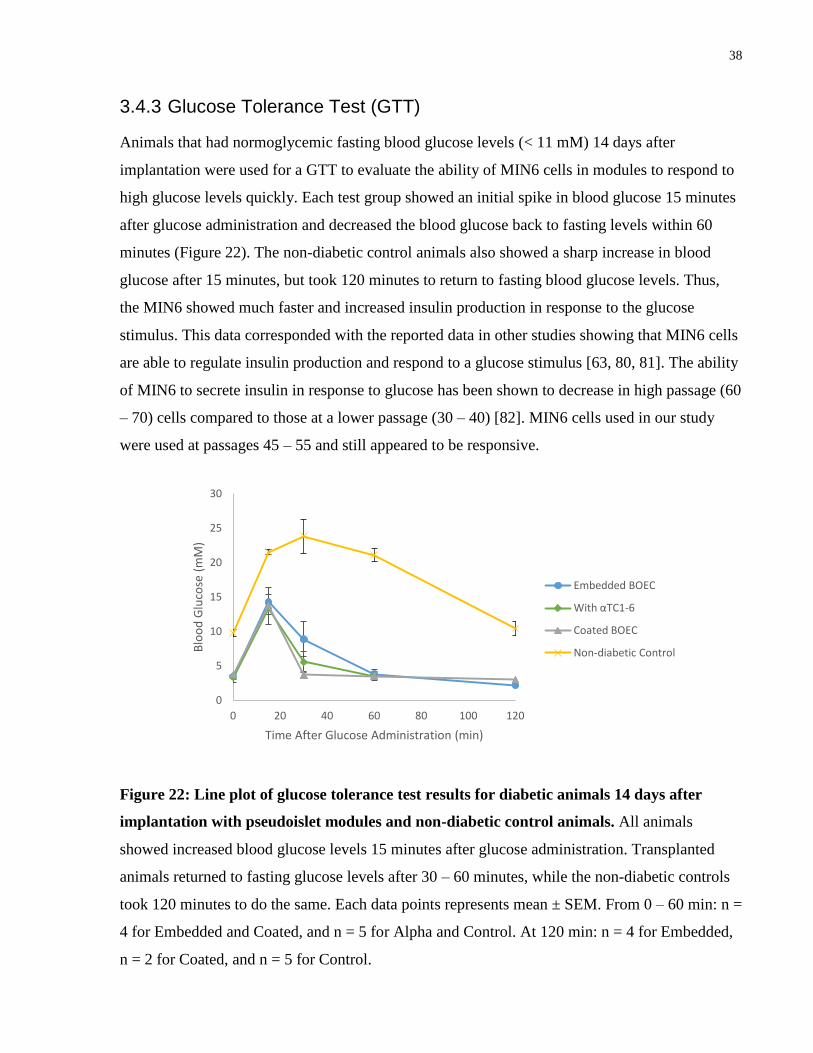
Figure 21: Dot plots of blood glucose measurements on days 7, 14, and 21 for SCID-Bg
mice implanted with pseudoislet modules. On day 7 there were no significant differences
between the groups. By day 14, pseudoislet modules containing a 33/67 mix of αTC1-6/MIN6
and embedded BOEC had significantly lower blood glucose than modules without α cells. At day
21, all of the groups had returned to normoglycemia without significant differences among them.
** p ≤ 0.01 by ANOVA. n = 5.
38
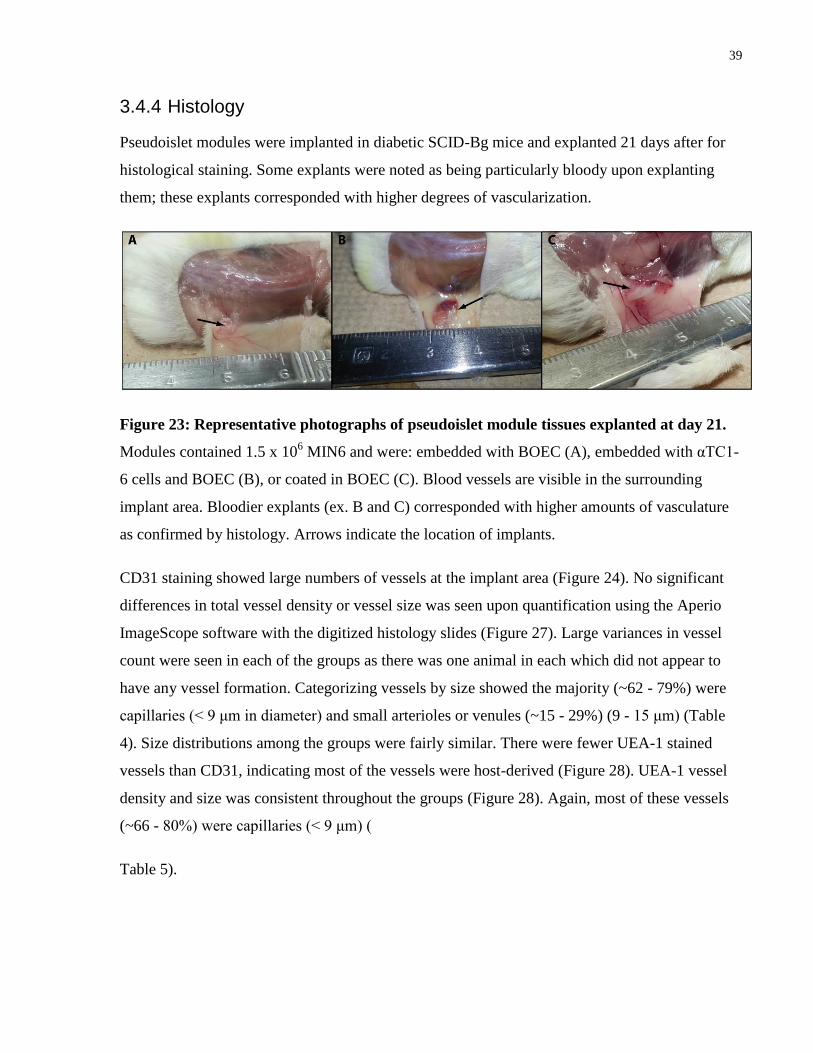
3.4.3 Glucose Tolerance Test (GTT)
Animals that had normoglycemic fasting blood glucose levels (< 11 mM) 14 days after
implantation were used for a GTT to evaluate the ability of MIN6 cells in modules to respond to
high glucose levels quickly. Each test group showed an initial spike in blood glucose 15 minutes
after glucose administration and decreased the blood glucose back to fasting levels within 60
minutes (Figure 22). The non-diabetic control animals also showed a sharp increase in blood
glucose after 15 minutes, but took 120 minutes to return to fasting blood glucose levels. Thus,
the MIN6 showed much faster and increased insulin production in response to the glucose
stimulus. This data corresponded with the reported data in other studies showing that MIN6 cells
are able to regulate insulin production and respond to a glucose stimulus [63, 80, 81]. The ability
of MIN6 to secrete insulin in response to glucose has been shown to decrease in high passage (60
– 70) cells compared to those at a lower passage (30 – 40) [82]. MIN6 cells used in our study
were used at passages 45 – 55 and still appeared to be responsive.
Figure 22: Line plot of glucose tolerance test results for diabetic animals 14 days after
implantation with pseudoislet modules and non-diabetic control animals. All animals
showed increased blood glucose levels 15 minutes after glucose administration. Transplanted
animals returned to fasting glucose levels after 30 – 60 minutes, while the non-diabetic controls
took 120 minutes to do the same. Each data points represents mean ± SEM. From 0 – 60 min: n =
4 for Embedded and Coated, and n = 5 for Alpha and Control. At 120 min: n = 4 for Embedded,
n = 2 for Coated, and n = 5 for Control.
0
5
10
15
20
25
30
0 20 40 60 80 100 120
Blo
od
Glu
cose
(m
M)
Time After Glucose Administration (min)
Embedded BOEC
With αTC1-6
Coated BOEC
Non-diabetic Control
39
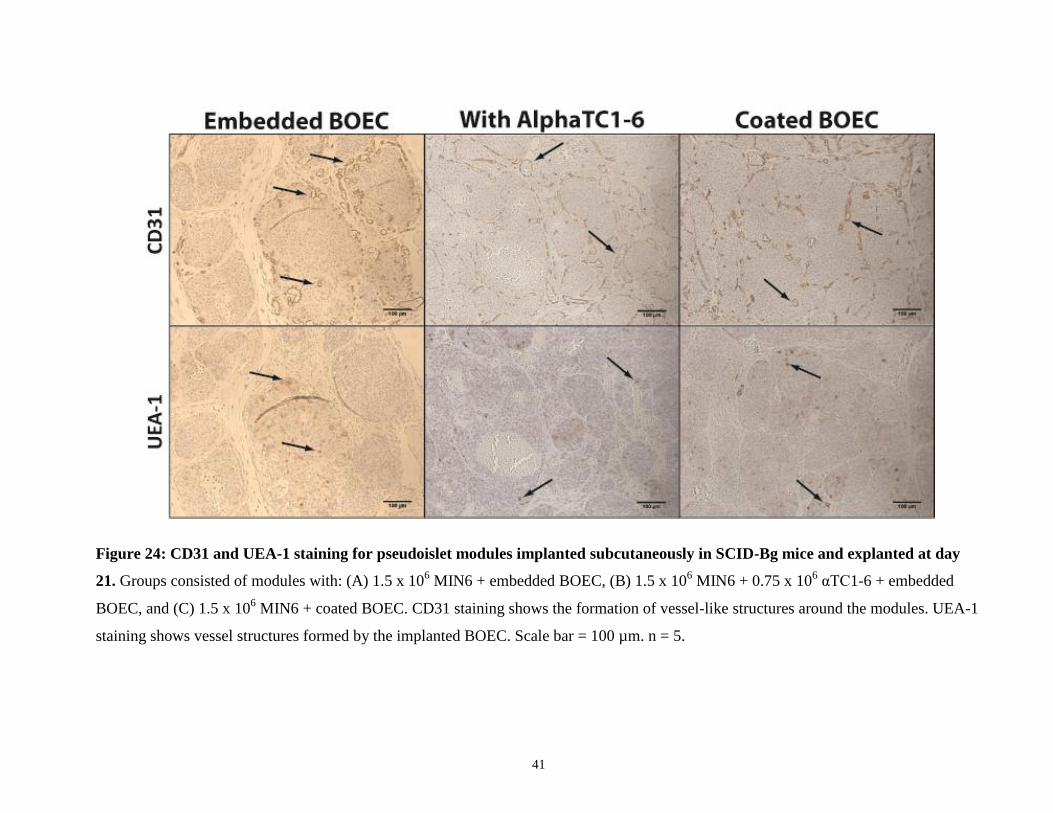
3.4.4 Histology
Pseudoislet modules were implanted in diabetic SCID-Bg mice and explanted 21 days after for
histological staining. Some explants were noted as being particularly bloody upon explanting
them; these explants corresponded with higher degrees of vascularization.
Figure 23: Representative photographs of pseudoislet module tissues explanted at day 21.
Modules contained 1.5 x 106 MIN6 and were: embedded with BOEC (A), embedded with αTC1-
6 cells and BOEC (B), or coated in BOEC (C). Blood vessels are visible in the surrounding
implant area. Bloodier explants (ex. B and C) corresponded with higher amounts of vasculature
as confirmed by histology. Arrows indicate the location of implants.
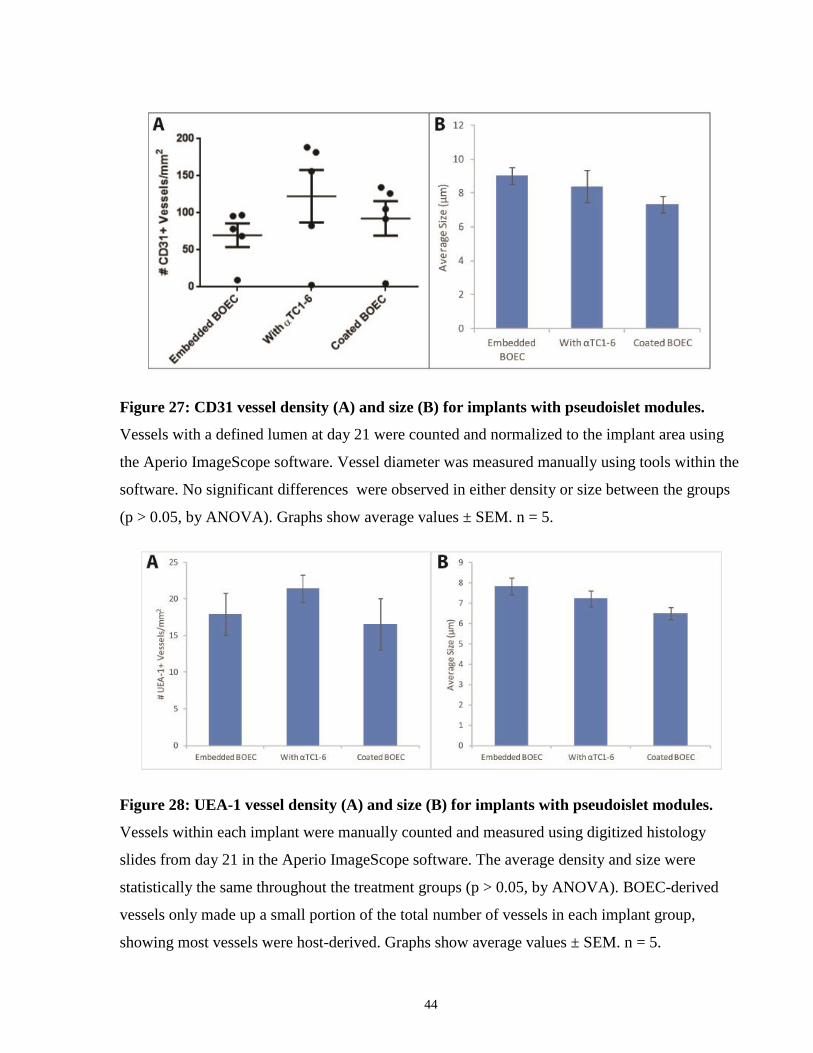
CD31 staining showed large numbers of vessels at the implant area (Figure 24). No significant
differences in total vessel density or vessel size was seen upon quantification using the Aperio
ImageScope software with the digitized histology slides (Figure 27). Large variances in vessel
count were seen in each of the groups as there was one animal in each which did not appear to
have any vessel formation. Categorizing vessels by size showed the majority (~62 - 79%) were
capillaries (< 9 μm in diameter) and small arterioles or venules (~15 - 29%) (9 - 15 μm) (Table
4). Size distributions among the groups were fairly similar. There were fewer UEA-1 stained
vessels than CD31, indicating most of the vessels were host-derived (Figure 28). UEA-1 vessel
density and size was consistent throughout the groups (Figure 28). Again, most of these vessels
(~66 - 80%) were capillaries (< 9 μm) (
Table 5).

40
H&E staining showed the presence of red blood cells stained in red within many of the vessels
indicating incorporation with the host vasculature (Figure 25). Trichrome showed little blue
staining for collagen, illustrating degradation of the modules, presumably by proliferation of the
implanted pancreatic cells. Some larger red pools are also present without vessels directly around
them in each of the groups. These indicate the vasculature is leaky. This, combined with the
highly proliferative nature of the cells, suggests a tumorigenic response upon implantation of
these cells [83, 84]. Since most of the vasculature formed is host-derived and given that the
pancreatic cells used are from cancer lines, this tumorigenic response is likely just from the
MIN6 and αTC1-6 rather than the BOEC. No differences were seen between the groups from
H&E and trichrome staining.
Brown insulin staining was visible throughout many of the cells in the modules, indicating that a
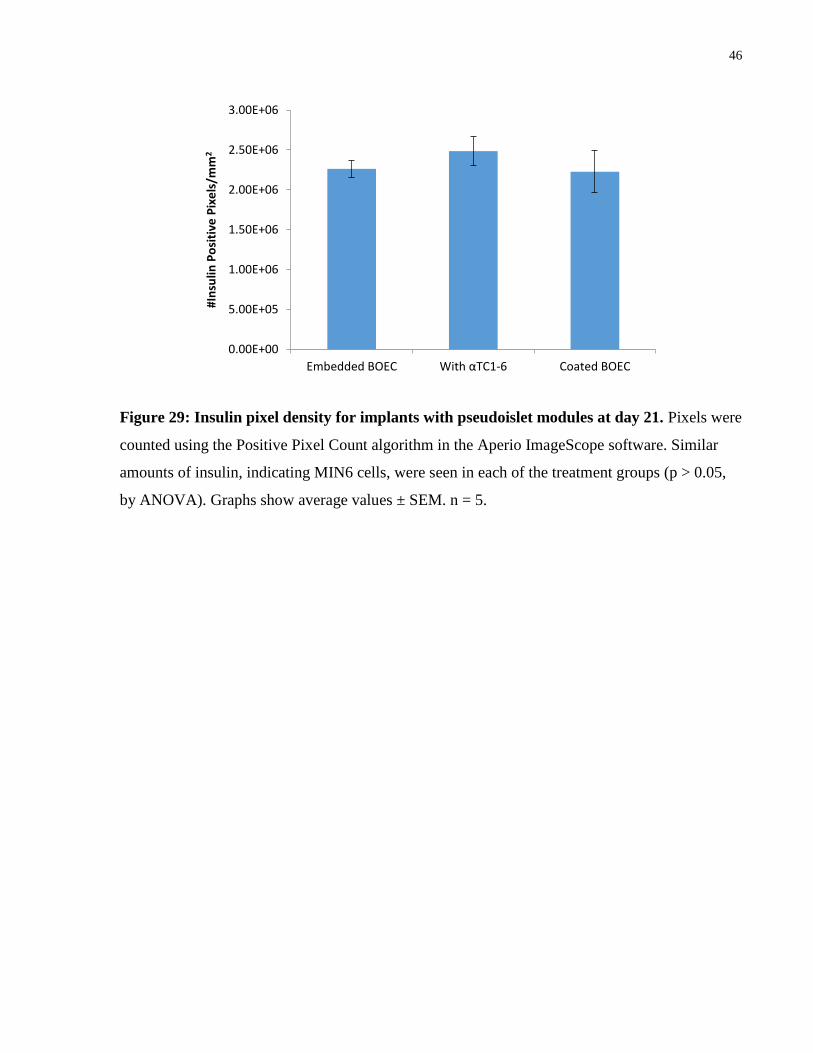
large number of MIN6 were present (Figure 26). There were no significant differences in the
amount of insulin staining among the treatment groups (Figure 29). Since all groups started with
the same number of MIN6 initially, this indicated that the addition of αTC1-6 cells does not
appear to change the proliferation of MIN6 in the modules. Glucagon stained histology slides
were also obtained (Figure 33). These showed similar staining to insulin; MIN6 cells also
produce small amounts of glucagon [85].
41
Figure 24: CD31 and UEA-1 staining for pseudoislet modules implanted subcutaneously in SCID-Bg mice and explanted at day
21. Groups consisted of modules with: (A) 1.5 x 106 MIN6 + embedded BOEC, (B) 1.5 x 10
6 MIN6 + 0.75 x 10
6 αTC1-6 + embedded
BOEC, and (C) 1.5 x 106 MIN6 + coated BOEC. CD31 staining shows the formation of vessel-like structures around the modules. UEA-1
staining shows vessel structures formed by the implanted BOEC. Scale bar = 100 µm. n = 5.
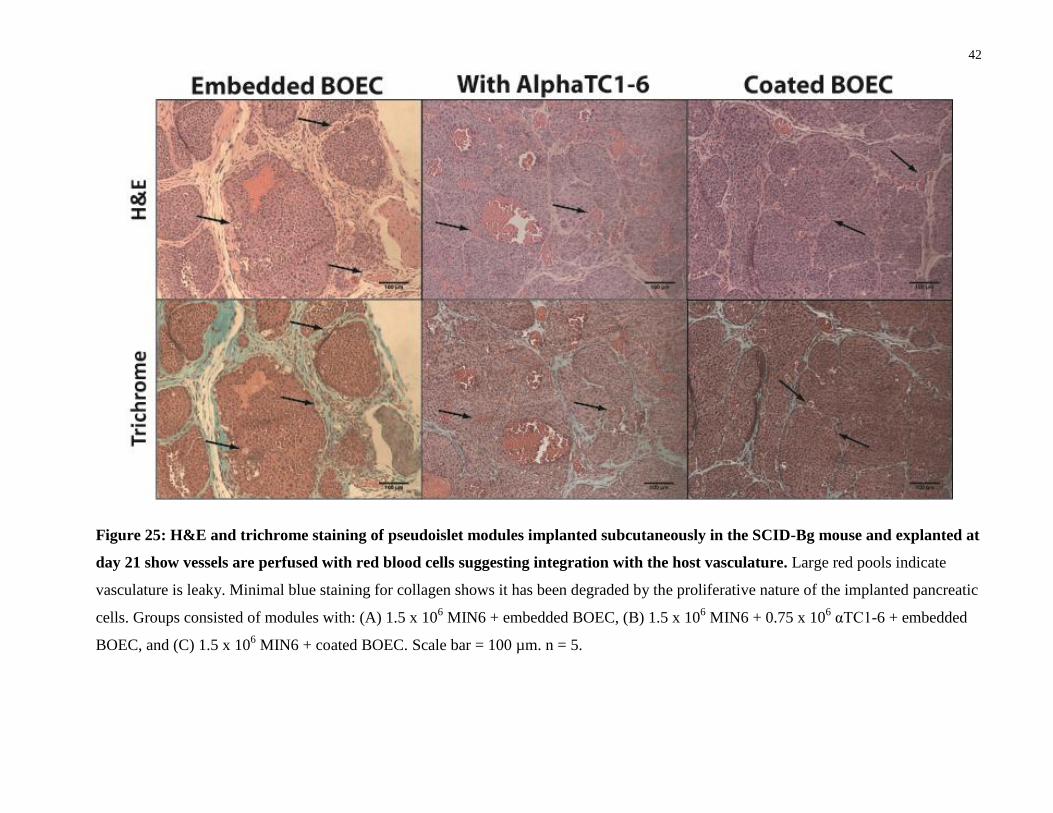
42
Figure 25: H&E and trichrome staining of pseudoislet modules implanted subcutaneously in the SCID-Bg mouse and explanted at
day 21 show vessels are perfused with red blood cells suggesting integration with the host vasculature. Large red pools indicate
vasculature is leaky. Minimal blue staining for collagen shows it has been degraded by the proliferative nature of the implanted pancreatic
cells. Groups consisted of modules with: (A) 1.5 x 106 MIN6 + embedded BOEC, (B) 1.5 x 10
6 MIN6 + 0.75 x 10
6 αTC1-6 + embedded
BOEC, and (C) 1.5 x 106 MIN6 + coated BOEC. Scale bar = 100 µm. n = 5.
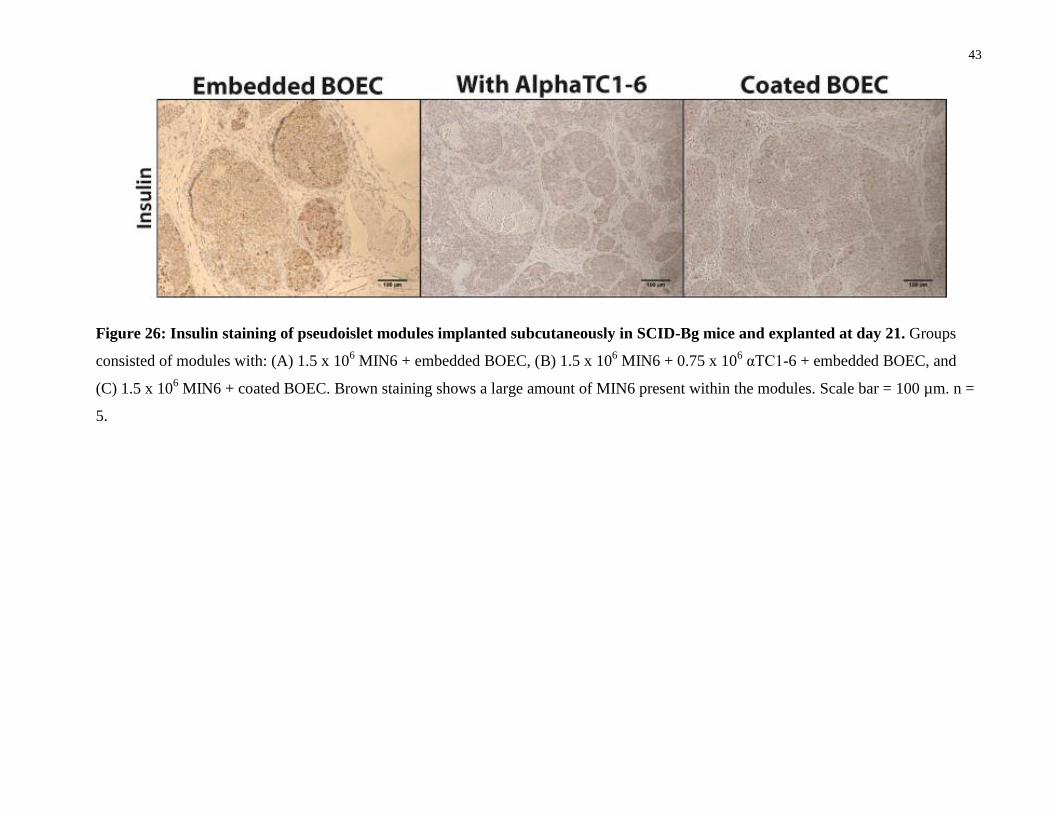
43
Figure 26: Insulin staining of pseudoislet modules implanted subcutaneously in SCID-Bg mice and explanted at day 21. Groups
consisted of modules with: (A) 1.5 x 106 MIN6 + embedded BOEC, (B) 1.5 x 10
6 MIN6 + 0.75 x 10
6 αTC1-6 + embedded BOEC, and
(C) 1.5 x 106 MIN6 + coated BOEC. Brown staining shows a large amount of MIN6 present within the modules. Scale bar = 100 µm. n =
5.
44
Figure 27: CD31 vessel density (A) and size (B) for implants with pseudoislet modules.
Vessels with a defined lumen at day 21 were counted and normalized to the implant area using
the Aperio ImageScope software. Vessel diameter was measured manually using tools within the
software. No significant differences were observed in either density or size between the groups
(p > 0.05, by ANOVA). Graphs show average values ± SEM. n = 5.
Figure 28: UEA-1 vessel density (A) and size (B) for implants with pseudoislet modules.
Vessels within each implant were manually counted and measured using digitized histology
slides from day 21 in the Aperio ImageScope software. The average density and size were
statistically the same throughout the treatment groups (p > 0.05, by ANOVA). BOEC-derived
vessels only made up a small portion of the total number of vessels in each implant group,
showing most vessels were host-derived. Graphs show average values ± SEM. n = 5.

45
Table 4: Size distribution of CD31+ vessels in implants with pseudoislet modules (n = 5).
Size Range Embedded BOEC With αTC1-6 Coated BOEC
Capillaries (< 9 μm) 62% 70% 79%
Small Arterioles or
Venules (9 - 15 μm) 29% 18% 15%
Large Arterioles or
Venules (15 - 75 μm) 10% 11% 6%
Other (abnormal, ≥
75 μm) 0% 0% 0%
Table 5: Size distribution of UEA-1+ vessels in implants with pseudoislet modules (n = 5).
Size Range Embedded BOEC With αTC1-6 Coated BOEC
Capillaries (< 9 μm) 69% 81% 87%
Small Arterioles or
Venules (9 - 15 μm) 28% 19% 13%
Large Arterioles or
Venules (15 - 75 μm) 0% 0% 0%
Other (abnormal, ≥
75 μm) 0% 0% 0%
46
Figure 29: Insulin pixel density for implants with pseudoislet modules at day 21. Pixels were
counted using the Positive Pixel Count algorithm in the Aperio ImageScope software. Similar
amounts of insulin, indicating MIN6 cells, were seen in each of the treatment groups (p > 0.05,
by ANOVA). Graphs show average values ± SEM. n = 5.
0.00E+00
5.00E+05
1.00E+06
1.50E+06
2.00E+06
2.50E+06
3.00E+06
Embedded BOEC With αTC1-6 Coated BOEC
#In
sulin
Po
siti
ve P
ixe
ls/m
m2

47
Chapter 4 Discussion
4 Discussion
4.1 BOEC vs. HUVEC in Modular Tissue Engineering
Vascular tissue engineering relies on the formation of a microvasculature network to supply
oxygen and nutrients throughout the implanted tissue. The choice of EC source is critical both in
the formation of this network as well as in making these engineered tissues clinically relevant. It
is desirable to have an EC source that is autologous, easily obtained from adults, and able to be
expanded into a large number of cells. Mature ECs are limited in their clinical use because of
their relatively low proliferation potential, invasive isolation procedures, and difficulty in
obtaining a sufficient number of cells from isolation [51]. BOEC are easy to obtain and can be
expanded much more in culture compared to other sources like HUVEC, making them a suitable
choice for tissue engineering applications.
HUVEC are only used up to passage 6 for module fabrication, whereas BOEC up to passage 15
were able to be used (higher passages may be possible but were not tested) [36, 77]. In this study
BOEC between passage 4 - 8 were typically used. The ability for BOEC to be expanded for
prolonged periods of time in culture is supported in studies done by others as well [47, 51, 52].
As BOEC are expanded in vitro, they may develop a lower degree of vascularization as a result
of maturation [51]. However, this can be compensated for by using more cells which are made
available through expanding the BOEC anyways [51]. The highly proliferative nature of BOEC
is particularly useful when considering the challenges of scaling up this technology. Moving
from murine to porcine and eventually to human trials will require much larger amounts of cells
and modules per implant; BOEC can be expanded to meet this demand without losing expression
of their endothelial markers or function [51].
In terms of their function, HUVEC coated modules with adMSC have been shown to form
vasculature networks when subcutaneously implanted [36]. This study showed that BOEC are
able to form vasculature with a similar number of both total and EC-derived vessels at day 14
(Figure 12 and Figure 13). Regardless of EC type, less than half of the total vessels were EC-
derived at this time point (Figure 13). Erythrocytes were visible in the vessels using H&E and

48
trichrome staining (Figure 10), indicating integration with the host vasculature. Vessels were also
present at day 3 for both BOEC and HUVEC coated modules, but were not yet integrated with
the host vasculature then (Figure 9 and Figure 10). It may be beneficial to examine other times
points before day 14 to see whether BOEC-derived vessels become integrated faster than
HUVEC as suggested in other studies [47]. For in vitro functional analysis, results from the
fibrin sprouting assay showed BOEC formed sprouts of similar lengths and numbers as HUVEC
at days 1, 3, and 7 both with and without adMSC (Figure 6).
While our study showed a similar degree of vascularization by BOEC and HUVEC, other studies
have demonstrated that BOEC may form better networks. Sieminski et al. showed that BOEC in
3D collagen gels in vitro had better cell elongation and generated more force (contracted the gels
more) [47]. The benefits seen from this contraction likely occur as a result of increased cell
density [47].
4.2 The Effects of Embedding BOEC or HUVEC in Modules
Modules are currently coated by gently mixing them with HUVEC for 1 hour after being cut and
separated from their tubing. This results in a confluent monolayer on the surface of the modules
after about 2 - 3 days [86]. This confluent layer is responsible for EC-derived vessel formation
and integration with host vasculature upon implantation. Being able to embed ECs in the
modules rather than coating them would simplify the fabrication process and also potentially
provide signaling to other embedded cells to improve their function. Results in this study showed
that both BOEC and HUVEC embedded modules were able to form the same number of vessels
as coated modules 14 days after subcutaneous implantation in SCID-Bg mice (Figure 12).
Similarities were also seen using other histological stains; red blood cells were present in vessels
at day 14 (Figure 16). These in vivo results demonstrated that embedding and coating modules
with ECs form equivalent vasculature networks with adMSC modules.
There may be other factors to consider, such as how many of the EC are able to migrate to the
surface and whether a confluent monolayer still forms after the same time. Confocal imaging of
the EC embedded modules did not show organized VE-cadherin networks (Figure 14) as was
seen for coated modules (Figure 7). This suggested that a confluent layer had not yet formed, in
vitro, at the surface of the modules at day 7; ECs form organized junctions when confluent [87,
88]. Despite this, the EC embedded modules demonstrated a similar degree of vascularization by

49
day 14, but further studies may be needed to ensure that embedding the ECs is not detrimental at
earlier time points.
4.3 Pseudoislet Modules
One of the major challenges with islet transplantation is the loss of function and viability upon
isolation from the donor pancreas and implantation into non-vascularized tissue. The loss of their
highly vascularized native environment causes islets to rely on diffusion alone to receive oxygen
and nutrients; many islets are too large to adequately receive these resources through diffusion
[20]. Vascular tissue engineering can enable the production and integration of a vasculature
network upon implantation of islets; improving their function and viability for longer periods of
time.
A further improvement to the islet transplantation procedure is the ability to form aggregates
(pseudoislets) from pancreatic cell lines. This has the potential to allow for the creation of
smaller islets and to eliminate the concern of donor shortages. In our study, subcutaneous
injection of pseudoislet modules into diabetic SCID-Bg mice was shown to return blood glucose
levels to normoglycemia after about 1 - 3 weeks depending on the treatment group (Figure 20).
Animals implanted with pseudoislet modules were able to return to fasting blood glucose levels
after a glucose stimulus in half the time it took non-diabetic controls to do the same (Figure 22).
A large amount of CD31 staining was visible after 21 days, indicating the presence of a large
number of vessels (Figure 24 and Figure 27). These vessels were integrated with the host
vasculature as evidenced through the presence of erythrocytes within the vessels (Figure 25).
Few BOEC appear to still be present at this time point (Figure 24 and Figure 28). The
transplanted MIN6 cells were highly proliferative and degraded most of the collagen by day 21
(Figure 25). Given this proliferative nature and the presence of leaky vasculature (large blood
pools were present in all of the explants), it can be concluded that the MIN6 (and αTC1-6 when
used) had a tumorigenic response. As these are cancer cell lines, this response was not
surprising; it further underlined the need to use a different cell source instead in future studies.
In our study we focused on looking at embedding versus coating pseudoislet modules with
BOEC and what effects adding glucagon-producing αTC1-6 cells would have. Pseudoislet
modules coated in BOEC returned to normoglycemia about 2 weeks sooner than those embedded
with BOEC (Figure 20), although no differences were seen in the number of vessels present after

50
21 days (Figure 27). This contradicted the findings in other studies that showed improved insulin
production of β cells when transplanted with ECs or late outgrowth EPCs [61, 71]. The presence
of ECs should help pseudoislets to mimic the intravascular network of native islets [23], but this
was not seen in our data. One reason for this could be the MIN6, as they would produce large
amounts of insulin even without the BOEC; it is possible any effects of the BOEC were
overpowered by the MIN6. Another possibility is that the coated modules were contracted more
than the embedded ones (this was noted visually before implantation). The BOEC generate a
force to contract the modules which would result in a higher cell density and possibly a better
vasculature network [47]. Studies with HepG2 and HUVEC showed a 7% increase in cell density
[79]. The BOEC should not be interfering directly with the function of the MIN6, as numerous
studies show that ECs actually promote the function of β cells in islets and aggregates [61, 71,
89, 90, 91]. Thus it is possible that the increased contraction seen in the coated pseudoislet
modules may benefit MIN6 function in some way, increasing insulin secretion.
Pseudoislets embedded with BOEC and containing αTC1-6 cells in a 33/67 ratio with MIN6
brought diabetic mice to normoglycemia after 8 days - about 2 weeks sooner than those with
MIN6 only (Figure 20). Blood glucose levels for modules with this mix of cells was significantly
lower at day 14 compared to those without the αTC1-6 (Figure 21). There appeared to be more
vessels present on day 21 in animals with αTC1-6 cells, but large variances in the animals
prevented a statistically different result from being seen (Figure 27). The presence of α cells was
expected to allow for signaling and communication with the MIN6 that would prolong and
improve their function, and improve their response to a glucose stimulus as seen in other reports
[70, 92, 93, 94, 95]. Notably, Keymeulen et al., used between 1.2 x 106 – 1.7 x 10
6 β cells with
0.6 x 106 – 0.7 x 10
6 α cells; both approximately the same ratio and number of cells used in our
study [70]. They found that animals transplanted with mixed aggregates were glucose tolerant
until 20 weeks after transplantation, and only 1 of the 12 animals in that group were diabetic
after 64 weeks [70]. In comparison, those transplanted only with β cells lost glucose tolerance
after 4 weeks, and 5 of 11 animals were diabetic at the end of the experiment [70]. These results
show promise for pseudoislet modules to maintain normoglycemia in longer studies in the future.

51
Chapter 5 Conclusions and Future Work
5 Conclusions and Future Work
The results in this study support the concept that transplantation of aggregated pancreatic cell
lines in modules with BOEC will result in effectively vascularized pseudoislets with the ability
to restore diabetic mice to a normoglycemic state. Initial experiments showed there were no
significant differences in the vascularization ability of BOEC compared to HUVEC in vitro or in
vivo. Since BOEC have clinically relevant advantages over HUVEC, stemming from their nature
as a progenitor cell line, it would be beneficial to use BOEC in subsequent studies in the future.
Evaluation of their use as an autologous cell source could be performed by obtaining the cells
from the blood of rats or larger animal models, and studying the immune response following
transplantation of the cells back into the host animals.
Embedding ECs in modules was shown to be sufficient to form a vasculature which was not
significantly different from that formed by modules coated in ECs. However, it appears that
modules coated with BOEC and embedded with MIN6 were able to return diabetic SCID-Bg
mice to normoglycemia faster than modules embedded with BOEC instead. Both of these groups
returned diabetic mice to fasting blood glucose levels within 60 minutes of glucose
administration in a similar way. Future work should examine possible differences in the degree
of contraction of EC coated vs. embedded modules, and what effect this may play in the function
of insulin-producing cells.
Pseudoislets containing a 33/67 mix of αTC1-6/MIN6 were able to return diabetic mice to
fasting blood glucose levels 60 minutes after receiving a glucose stimulus and successfully
reached normoglycemia about 1 week after transplantation in diabetic mice; faster than was seen
for modules without the αTC1-6. Histological analysis showed there were no significant
differences between the groups in terms of vessel density and size. Thus, differences seen in
terms of restoration of normoglycemia were likely due to other factors than more or less
vasculature. For example, αTC1-6 cells may enhance the function of MIN6 through signaling
and cell-to-cell interactions and coated modules may contract more which could affect MIN6

52
function. An additional test group in the future should contain the same mix of pancreatic cells,
but be coated with BOEC rather than embedded to see if results are improved further.
Overall, the results from these experiments warrant future studies with pancreatic cell aggregates
in modules with different cells since both αTC1-6 and MIN6 are insulinoma-derived cell lines. In
particular, two options are re-aggregating isolated islets or using insulin and glucagon secreting
cells induced from pluripotent stem cells or progenitor cells. Many donor islets cannot be
transplanted after isolation because they are too damaged. Re-aggregating islets post-isolation
may allow them to have better function and viability upon transplantation. This would partially
reduce the high demand of donors, but probably not enough. Instead, using stem or progenitor
cells would eliminate this concern. Also, this would allow for a higher degree of customization
of the aggregates and further optimization of the model. Therefore, it is recommended to use
pancreatic cells derived from stem or progenitor cells in pseudoislet modules in the future.
Possible parameters to explore include the ratio and amount of each cell used (including the
addition of BOEC either embedded in or coating the modules), whether the organization of the
cells within the aggregates may affect their function, and the ability of the cells to maintain
normoglycemia over long periods of time.

53
References
[1] World Health Organization, "Diabetes," January 2015. [Online]. Available:
http://www.who.int/mediacentre/factsheets/fs312/en/. [Accessed 23 June 2015].
[2] Canadian Diabetes Association, "Types of Diabetes," 2015. [Online]. Available:
http://www.diabetes.ca/about-diabetes/types-of-diabetes. [Accessed 23 June 2015].
[3] JDRF, "Type 1 Diabetes," [Online]. Available: http://www.jdrf.ca/news-and-media/fact-
sheets/type-1-diabetes/. [Accessed 23 June 2015].
[4] B. M. Frier, G. Schernthaner and S. Heller, "Hypoglycemia and Cardiovascular Risks,"
Diabetes Care, vol. 34, no. 2, pp. 132-137, 2011.
[5] S. Heller, "Sudden Death and Hypoglycemia," Diabetic Hypoglycemia, vol. 1, no. 2, pp. 2-
7, 2008.
[6] American Diabetes Association, "Pancreas Transplantation," 3 November 2013. [Online].
Available: http://www.diabetes.org/living-with-diabetes/treatment-and-
care/transplantation/pancreas-transplantation.html. [Accessed 23 June 2015].
[7] Diabetes Research Institute Foundation, "What are Islet Cells?," Diabetes Research Institute
Foundation, 2014. [Online]. Available: http://www.diabetesresearch.org/islet-cells.
[Accessed 3 February 2016].
[8] C. Ionescu-Tirgoviste, P. A. Gagniuc, E. Gubceac, L. Mardare, I. Popescu, S. Dima and M.
Militaru, "A 3D map of the islet routes throughout the healthy human pancreas," Scientific
Reports, vol. 14634, no. 5, pp. 1-14, 2015.
[9] M. Kharouta, K. Miller, A. Kim, P. Wojcik, G. Kilimnik, A. Dey, D. F. Steiner and M.
Hara, "No mantle formation in rodent islets - the prototye of islet revisited," Diabetes
Research and Clinical Practice, vol. 85, no. 3, pp. 252-257, 2009.

54
[10] J. Dolensek, M. S. Rupnik and A. Stozer, "Structural similarities and differences between
the human and the mouse pancreas," Islets, vol. 7, no. 1, p. e1024405, 2015.
[11] A. Kim, K. Miller, J. Jo, G. Kilimnik, P. Wojcik and M. Hara, "Islet architecture: A
comparative study," Islets, vol. 1, no. 2, pp. 129-136, 2009.
[12] S. Merani and A. M. J. Shapiro, "Current status of pancreatic islet transplantation," Clinical
Science, vol. 110, pp. 611-625, 2006.
[13] D. E. Sutherland, P. F. Gores, A. C. Farney, D. C. Wahoff, A. J. Matas, D. L. Dunn, R. W.
Gruessner and J. S. Najarian, "Evolution of Kidney, Pancreas, and Islet Transplantation for
Patients With Diabetes at the University of Minnesota," The American Journal of Surgery,
vol. 166, no. 5, pp. 456-491, 1993.
[14] A. J. Shapiro, J. R. Lakey, E. A. Ryan, G. S. Korbutt, E. Toth, G. L. Warnock, N. M.
Kneteman and R. V. Rajotte, "Islet Transplantation in Seven Patients with Type 1 Diabetes
Mellitus Using a Glucocorticoid-Free Immunosuppressive Regimen," The New England
Journal of Medicine, vol. 343, no. 4, pp. 230-238, 2000.
[15] N. Niclauss, R. Meier, B. Bedat, E. Berishvili and T. Berney, "Beta-cell replacement:
Pancreas and islet cell transplantation," Endocrine Development, vol. 31, pp. 146-162, 2016.
[16] R. Gupta and M. V. Sefton, "Application of an Endothelialized Modular Construct for Islet
Transplantation in Syngeneic and Allogeneic Immunosuppressed Rat Models," Tissue
Engineering: Part A, vol. 17, no. 15 and 16, pp. 2005-2015, 2011.
[17] T. Berney, L. Buhler, P. Majno, G. Mentha and P. Morel, "Immunosuppression for
pancreatic islet transplantation," Transplantation Proceedings, vol. 36, pp. 362S-366S,
2004.
[18] M. Milgrom, R. M. Jindal, R. A. Sidner and M. L. Milgrom, "Post-transplant diabetes
mellitus: The role of immunosuppression," Drug Safety, vol. 16, no. 4, pp. 242-257, 1997.

55
[19] K. K.-T. Cheung and P. A. Senior, "Tacrolimus toxicity in islet transplantation due to
interaction with macrolides," Clinical Diabetes and Endocrinology, vol. 2, 2016.
[20] K. Ramachandran, S. J. Williams, H.-H. Huang, L. Novikova and L. Stehno-Bittel,
"Engineering Islets for Improved Performance by Optimized Reaggregation in a
Micromold," Tissue Engineering: Part A, vol. 15, no. 5 and 6, pp. 604-612, 2013.
[21] V. F. Duvivier-Kali, A. Omer, R. J. Parent, J. J. O'Neil and G. C. Weir, "Complete
Protection of Islets Against Allorejection and Autoimmunity by a Simple Barium-alginate
Membrane," Diabetes, vol. 50, no. 8, pp. 1698-1705, 2001.
[22] I. Konstantinova and E. Lammert, "Microvascular development: Learning from pancreatic
islets," BioEssays, vol. 26, pp. 1069-1075, 2004.
[23] M. Brissova, M. Fowler, P. Wiebe, A. Shostak, M. Shiota, A. Radhika, P. C. Lin, M.
Gannon and A. C. Powers, "Intraislet Endothelial Cells Contribute to Revascularization of
Transplanted Pancreatic Islets," Diabetes, vol. 53, pp. 1318-1325, 2004.
[24] S. Kizilel, A. Scavone, X. Liu, J. M. Nothias, D. Ostrega, P. Witkowski and M. Millis,
"Encapsulation of Pancreatic Islets Within Nano-thin Functional Polyethylene Glycol
Coatings for Enhanced Insulin Secretion," Tissue Engineering Part A, vol. 16, no. 7, pp.
2217-2228, 2010.
[25] E. S. O'Sullivan, A. S. Johnson, A. Omer, J. Hollister-Lock, S. Bonner-Weir, C. K. Colton
and G. C. Weir, "Rat Islet Cell Aggregates are Superior to Islets for Transplantation in
Microcapsules," Diabetologia, vol. 53, pp. 937-945, 2010.
[26] D. Jacobs-Tulleneers-THevissen, M. Chintinne, Z. Ling, P. Gillard, L. Schoonjans, G.
Delvaux, B. L. Strand, F. Gorus, B. Keymeulen and D. Pipeleers, "Sustained function of
alginate-encapsulated human islet cell implants in the peritoneal cavity of mice leading to a
pilot study in a type 1 diabetic patient," Diabetologia, vol. 56, pp. 1605-1614, 2013.
[27] F. Lim and A. M. Sun, "Microencapsulated islets as bioartificial endocrine pancreas,"

56
Science, vol. 210, no. 4472, pp. 908-910, 1980.
[28] P. Soon-Shiong, E. Feldman, R. Nelson, R. Heintz, Q. Yao, Z. Yao, T. Zheng, N. Merideth,
G. Skjak-Braek, T. Espevik, O. Smidsrod and P. Sandford, "Long-term reversal of diabetes
by the injection of immunoprotected islets," Proceedings of the National Academy of
Sciences, vol. 90, pp. 5843-5847, 1993.
[29] D. W. Scharp and P. Marchetti, "Encapsulated islets for diabetes therapy: history, current
progress and critical issues requiring solution," Advanced Drug Delivery Reviews, Vols. 67-
68, pp. 35-73, 2014.
[30] M. de Groot, T. A. Schuurs and R. van Schilfgaarde, "Causes of limited survival of
microencapsulated pancreatic islet grafts," Journal of Surgical Research, vol. 121, pp. 141-
150, 2004.
[31] R. van Schilfgaarde and P. de Vos, "Factors influencing th eproperties and performance of
microcapsules for immunoprotection of pancreatic islets," Journal of Molecular Medicine,
vol. 77, pp. 199-205, 1999.
[32] W. M. Fritschy, P. de Vos, H. Groen, F. A. Klatter, A. Pasma, G. H. Wolters and R. va
Schilfgaarde, "The capsular overgrowth on microencapsulated pancreatic islet grafts in
streptozotocin and autoimmune diabetic rats," Transplant International, vol. 7, pp. 264-271,
1994.
[33] L. Tappy, D. Randin, P. Vollenweider, L. Vollenweider, N. Paquot and U. Scherrer,
"Mechanisms of dexamethasone-induced insulin resistance in healthy humans," Journal of
Clinical Endocrinology Metabolism, vol. 79, no. 4, pp. 1063-1069, 1994.
[34] E. Dolgin, "Encapsulate this," Nature, vol. 20, no. 1, pp. 9-11, 2014.
[35] A. J. Vegas, O. Veiseh, J. C. Doloff, M. Ma, H. H. Tam, K. Bratlie, J. Li, A. R. Bader, E.
Langan, K. Olejnik, P. Fenton, J. W. Kang, J. Hollister-Locke, M. A. Bochenek, A. Chiu, S.
Siebert, K. Tang, S. Jhunjhunwala, S. Aresta-Dasilva, N. Dholakia, R. Thakrar, T. Vietti, M.
Chen, J. Cohen, K. Siniakowicz, M. Qi, J. McGarrigle, S. Lyle, D. M. Harlan, D. L. Greiner,

57
J. Oberholzer, G. C. Weir, R. Langer and D. G. Anderson, "Combinatorial hydrogel library
enables identification of materials that mitigate the foreign body response in primates,"
Nature Biotechnology, vol. 34, no. 3, pp. 345-352, 2016.
[36] A. P. McGuigan and M. V. Sefton, "Vascularized Organoid Engineered by Modular
Assembly Enables Blood Perfusion," Proceedings of the National academy of Sciences of
the United States of America, vol. 103, no. 31, pp. 11461-11466, 2006.
[37] U. Johansson, G. Elgue, B. Nilsson and O. Korsgren, "Composite Islet-Endothelial Cell
Grafts: A Novel Approach to Counteract Innate Immunity in Islet Transplantation,"
American Journal of Transplantation, vol. 5, pp. 2632-2639, 2005.
[38] J. Matin-ramirez, M. Hofman, M. Van Den Biggelaar, R. P. Hebbel and J. Voorberg,
"Establishment of outgrowth endothelial cells from peripheral blood," Nature Protocols,
vol. 7, no. 9, pp. 1709-1715, 2012.
[39] M. C. Yoder, "Human endothelial progenitor cells," Cold Spring Harbor Perspectives in
Medicine, vol. 2, no. 7, p. a006692, 2012.
[40] T. Asahara, T. Murohara, A. Sullivan, M. Silver, R. van der Zee, T. Li, B. Witzenbichler, G.
Schatteman and J. M. Isner, "Isolation of putative progenitor endothelial cells for
angiogenesis," Science, vol. 275, no. 5302, pp. 964-967, 1997.
[41] M. C. Yoder, L. E. Mead, D. Prater, T. R. Krier, K. N. Mroueh, F. Li, R. Krasich, C. J.
Temm, J. T. Prchal and D. A. Ingram, "Redefining endothelial progenitor cells via clonal
analysis and hematopoietic stem/progenitor cell principals," Blood, vol. 109, no. 5, pp.
1801-1809, 2007.
[42] R. J. Medina, C. L. O'Neill, M. Sweeney, J. Guduric-Fuchs, T. A. Gardiner, D. A. Simpson
and A. W. Stitt, "Molecular analysis of endothelial progenitor cell (EPC) subtypes reveals
two distinct cell populations with different identities," BMC Medical Genomics, vol. 3, no.
18, 2010.
[43] K. K. Hirschi, D. A. Ingram and M. C. Yoder, "Assessing identity, phenotype, and fate of

58
endothelial progenitor cells," Arteriosclerosis, Thrombosis, and Vascular Biology, vol. 28,
pp. 1584-1595, 2008.
[44] Y. Lin, D. J. Weisdorf, A. Solovey and R. P. Hebbel, "Origins of circulating endothelial
cells and endothelial outgrowth from blood," Journal of Clinical Investigations, vol. 105,
no. 1, pp. 71-77, 2000.
[45] D. A. Ingram, L. E. Mead, H. Tanaka, V. Meade, A. Fenoglio, K. Mortell, K. Pollok, M. J.
Ferkowicz, D. Giller and M. C. Yoder, "Identification of a novel hierarchy of endothelial
progenitor cells using human peripheral and umbilical cord blood," Blood, vol. 104, no. 9,
pp. 2752-2760, 2004.
[46] R. T. van Beem, R. E. Verloop, M. Kleijer, W. A. Noort, N. Loof, P. Koolwijk, C. E. van
der Schoot, V. W. van Hinsbergh and J. J. Zwaginga, "Blood outgrowth endothelial cells
from cord blood and peripheral blood: angiogenesis-related characteristics in vitro," Journal
of Thrombosis and Haemostasis, vol. 7, no. 1, pp. 217-226, 2008.
[47] A. L. Sieminski, R. P. Hebbel and K. J. Gooch, "Improved microvascular network in vitro
by human blood outgrowth endothelial cells relative to vessel-derived endothelial cells,"
Tissue Engineering, vol. 11, no. 9-10, pp. 1332-1345, 2005.
[48] K. A. Ahmann, S. L. Johnson, R. P. Hebbel and R. T. Tranquillo, "Shear stress responses of
adult blood outgrowth endothelial cells seeded on bioartificial tissue," Tissue Engineering
Part A, vol. 17, no. 19-20, pp. 2511-2521, 2011.
[49] N. B. Thebaud, R. Bareille, M. Remy, C. Bourget, R. Daculsi and L. Bordenave, "Human
progenitor-derived endothelial cells vs. venous endothelial cells for vascular tissue
engineering: an in vitro study," Journal of Tissue Engineering and Regenerative Medicine,
vol. 4, no. 6, pp. 473-484, 2010.
[50] B. Hendrickx, K. Verdonck, S. Van den Berge, S. Dickens, E. Eriksson, J. J. Vranckx and
A. Luttun, "Integration of blood outgrowth endothelial cells in dermal fibroblast sheets
promotes full thickness wound healing," Stem Cells, vol. 28, no. 7, pp. 1165-1177, 2010.

59
[51] J. M. Melero-Martin, Z. A. Khan, A. Picard, X. Wu, S. Paruchuri and J. Bischoff, "In vivo
vasculogenic potential of human blood-derived endothelial progenitor cells," Blood, vol.
109, no. 11, pp. 4761-4768, 2007.
[52] X. Wu, E. Rabkin-Aikawa, K. J. Guleserian, T. E. Perry, Y. Masuda, F. W. Sutherland, F. J.
Schoen, J. J. Mayer and J. Bischoff, "Tissue-engineered microvessels on three-dimensional
biodegradable scaffolds using human endothelial progenitor cells," American Journal of
Physiology: Heart and Circularoty Physiology, vol. 287, no. 2, pp. H480-487, 2004.
[53] C.-H. Yoon, J. Hur, K.-W. Park, J.-H. Kim, C.-S. Lee, I.-Y. Oh, T.-Y. Kim, H.-J. Cho, H.-J.
Kang, I.-H. Chae, H.-K. Yang, B.-H. Oh, Y.-B. Paek and H.-S. Kim, "Synergistic
neovascularization by mixed transplantation of early endothelial progenitor cells and late
outgrowth endothelial cells," Circulation, vol. 112, pp. 1618-1627, 2005.
[54] V. Coppens, Y. Heremans, G. Leuckx, K. Suenens, D. Jacobs-Tulleneers-Thevissen, K.
Verdonck, T. Lahoutte, A. Luttun, H. Heimberg and N. De Leu, "Human Blood Outgrowth
Endothelial Cells Improve Islet Survival and Function When Co-transplanted in a Mouse
Model of Diabetes," Diabetologia, vol. 56, pp. 382-390, 2013.
[55] V. Coppens, Y. Heremans, G. Leuckx, K. Suenens, D. Jacobs-Tulleneers-Thevissen, K.
Verdonck, A. Luttun, H. Heimberg and N. De Leu, "Reversal of hyperglycemia in diabetic
mice by a marginal islet mass together with human blood outgrowth endothelial cells is
independent of the delivery technique and blood clot-induced processes," Islets, vol. 5, no.
5, pp. 196-200, 2013.
[56] J. Rheinheimer, A. C. Bauer, S. P. Silveiro, A. A. Estivalet, A. P. Boucas, A. R. Rosa, B. M.
de Souza, F. S. de Oliveira, L. A. Cruz, L. A. Brondani, M. J. Azevedo, N. E. Lemos, R.
Carlessi, T. S. Assmann, J. L. Gross, C. B. Leitao and D. Crispim, "Human pancreatic islet
transplantation: an update and description of the establishment of a pancreatic islet isolation
laboratory," Archives of Endocrinology and Metabolism, vol. 59, no. 2, pp. 161-170, 2015.
[57] R. R. MacGregor, S. J. Williams, P. Y. Tong, K. Kover, W. V. Moore and L. Stehno-Bittel,

60
"Small rat islets are superior to large islets in in vitro function and in transplantation
outcomes," American Journal of Physiology Endocrinology and Metabolism, vol. 290, pp.
E771-779, 2005.
[58] R. Lehmann, R. A. Zuellig, P. Kugelmeier, P. B. Baenninger, W. Moritz, A. Perren, P.-A.
Clavien, M. Weber and G. A. Spinas, "Superiority of small islets in human islet
transplantation," Diabetes, vol. 56, no. 3, pp. 594-603, 2007.
[59] S. J. Williams, H.-H. Huang, k. Kover, W. Moore, C. Berkland, M. Singh, I. V. Smirnova,
R. MacGregor and L. Stehno-Bittel, "Reduction of diffusion barriers in isolated rat islets
improves survival, but not insulin secretion or transplantation outcome," Oraganogenesis,
vol. 6, no. 2, pp. 115-124, 2010.
[60] R. Montesano, P. Mouron, M. Amherdt and L. Orci, "Collagen matrix promotes
reorganization of pancreatic endocrine cell monolayers into islet-like organoids," The
Journal of Cell Biology, vol. 97, pp. 935-939, 1983.
[61] M. G. Spelios, L. A. Kenna, B. Wall and E. M. Akirav, "In vitro formation of B cell
pseudoislets using islet derived endothelial cells," PLoS ONE, vol. 8, no. 8, p. e72260, 2013.
[62] M. Skelin, M. Rupnik and A. Cencic, "Pancreatic beta cell lines and their applications in
diabetes mellitus research," ALTEX, vol. 2, no. 10, pp. 105-113, 2010.
[63] S. Inada, S. Kaneko, K. Suzuki, J.-i. Miyazaki, H. Asakura and M. Fujiwara, "Rectification
of diabetic state in C57BL/KsJ-db/db mice by implantation of pancreatic beta cell line
MIN6," Diabetes Research and Clinical Practice, vol. 32, pp. 125-133, 1996.
[64] T. Koblas, L. Pektorova, K. Zacharovova, Z. Kerkova, P. Girman, E. Dovolilova, L.
Karasova and F. Saudek, "Differentiation of CD133-positive pancreatic cells into insulin-
producing islet-like cell clusters," Transplantation Proceedings, vol. 40, pp. 415-418, 2008.
[65] C. Zanini, S. Bruno, G. Mandili, D. Baci, F. Cerutti, G. Cenacchi, L. Izzi, G. Camussi and
M. Forni, "Differentiation of mesenchymal stem cells derived from pancreatic islets and
bone marrow into islet-like cell phenotype," PLoS ONE, vol. 6, no. 12, p. e28175, 2011.

61
[66] Q.-P. Xie, H. Huang, B. Xu, X. Dong, S.-L. Gao, B. Zhang and Y.-L. Wu, "Human bone
barrow mesenchymal stem cells differentiate into insulin-producing cells upon
microenvironmental manipulation in vitro," Differentiation, vol. 77, pp. 483-491, 2009.
[67] D.-Q. Tang, L.-Z. Cao, B. R. Burkhardt, C.-Q. Xia, S. A. Litherland, M. M. Atkinson and
L.-J. Yang, "In vivo and in vitro characterization of insulin-producing cells obtained from
murine bone marrow," Diabetes, vol. 53, no. 7, pp. 1721-1732, 2004.
[68] A. J. Vegas, O. Veiseh, M. Gurtler, J. R. Millman, F. W. Pagliuca, A. R. Bader, J. C. Doloff,
J. Li, M. Chen, K. Olejnik, H. H. Tam, S. Jhunjhunwala, E. Langan, S. Aresta-dasilva, S.
Gandham, J. J. McGarrigle, M. A. Bochenek, J. Hollister-Lock, J. Oberholzer, D. L.
Greiner, G. C. Weir, D. A. Melton, R. Langer and D. G. Anderson, "Lon-term glycemic
control using polymer-encapsulated human stem cell-derived beta cells in immune
competent mice," Nature, 2016.
[69] A. J. King, J. R. Fernandes, J. Hollister-Lock, C. E. Nienaber, S. Bonner-Weir and G. C.
Weir, "Normal Relationship of Beta- and Non-beta-cells Not Needed for Successful Islet
Transplantation," Diabetes, vol. 56, no. 9, pp. 2312-2318, 2007.
[70] B. Keymeulen, J. Anselmo and D. Pipeleers, "Length of Metabolic Normalization After Rat
Islet Cell Transplantation Depends on Endocrine Cell Composition of Graft and on Donor
Age," Diabetologia, vol. 40, pp. 1152-1158, 1997.
[71] D. Penko, D. Mohanasundaram, S. Sen, C. Drogemuller, C. Mee, C. S. Bonder, P. H. Coates
and C. F. Jessup, "Incorporation of endothelial progenitor cells into mosaic pseudoislets,"
Islets, vol. 3, no. 3, pp. 1-7, 2011.
[72] F. Dietrich and P. I. Lelkes, "Fine-tuning of a three-dimensional microcarrer-based
angiogenesis assay for the analysis of endothelial-mesenchymal cell co-cultures in fibrin
and collagen gels," Angiogenesis, vol. 9, pp. 111-125, 2006.
[73] A. P. McGuigan, B. Leung and M. V. Sefton, "Fabrication of Cells Containing Gel Modules
to Assemble Modular Tissue-engineered Constructs.," Nature Protocols, vol. 1, no. 6, pp.

62
2963-2969, 2006.
[74] M. D. Chamberlain, R. Gupta and M. V. Sefton, "Bone marrow-derived mesenchymal
stromal cells enhance vessel development driven by endothelial cell-coated microtissues,"
Tissue Engineering Part A., vol. 18, no. 3-4, pp. 285-294, 2012.
[75] S. Kachgal and A. J. Putnam, "Mesenchymal stem cells from adipose and bone marrow
promote angiogenesis viz distinct cytokine and protease expression mechanisms,"
Angiogenesis, vol. 14, pp. 47-59, 2011.
[76] L. Corstorphine and M. V. Sefton, "Effectiveness factor and diffusion limitations in
collagen gel modules containing HepG2 cells," Journal of Tissue Engineering and
Regenerative Medicine, vol. 5, no. 2, pp. 119-129, 2011.
[77] M. J. Butler and M. V. Sefton, "Cotransplantation of Adipose-Derived Mesenchymal
Stromal Cells and Endothelial Cells in a Modular Construct Drives Vascularization in
SCID/bg Mice," Tissue Engineering Part A, vol. 18, no. 15-16, pp. 1628-1641, 2012.
[78] H. Brereton, M. J. Carvell, S. J. Persaud and P. M. Jones, "Islet a-cells do not influence
insulin secretion from B-cells through cell-cell contact," Endocrinology, vol. 31, pp. 61-65,
2007.
[79] A. P. McGuigan and M. V. Sefton, "Design and fabrication of sub-mm-sized modules
containing encapsulated cells for modular tissue engineering," Tissue Engineering, vol. 13,
no. 5, pp. 1069-1078, 2007.
[80] H. Ishihara, T. Asano, K. Tsukuda, H. Katagiri, K. Inukai, M. Anai, M. Kikuchi, Y. Yazaki,
J. Miyazaki and Y. Oka, "Pancreatic beta cell line MIN6 exhibits characteristics of glucose
metabolism and glucose-stimulated insulin secretion similar to those of normal islets,"
Diabetologia, vol. 36, no. 11, pp. 1139-1145, 1993.
[81] J. Miyazaki, K. Araki, E. Yamato, H. Ikegami, T. Asano, Y. Shibasaki, Y. Oka and K.
Yamamura, "Establishment of a pancreatic beta cell line that retains glucose-inducible
insulin secretion: special reference to expression of glucose transporter isoforms,"

63
Endocrinology, vol. 127, no. 1, pp. 126-132, 1990.
[82] K. Cheng, V. Delghingaro-Augusto, C. J. Nolan, N. Turner, N. Hallahan, S. Andrikopoulos
and J. E. Gunton, "High passage MIN6 cells have impaired insulin secretion with impaired
glucose and lipid oxidation," PLoS ONE, vol. 7, no. 7, p. e40868, 2012.
[83] K. Jain, "Normalization of tumor vasculature: an emerging concept in antiangiogenic
therapy," Science, vol. 307, no. 5707, pp. 58-62, 2005.
[84] A. C. Dudley, "Tumor endothelial cells," Cold Spring Harbor Perspectives in Medicine,
vol. 2, no. 3, p. a006536, 2012.
[85] K. Nakashima, Y. Kanda, Y. Hirokawa, F. Kawasaki and M. Matsuki, "MIN6 is not a pure
beta cell line but a mixed cell line with other pancreatic endocrine hormones," Endocrine
Journal, vol. 56, no. 1, pp. 45-53, 2009.
[86] M. She, A. P. McGuigan and M. V. Sefton, "Tissue factor and thrombomodulin expression
on endothelial cell-seeded collagen modules for tissue engineering," Journal of Biomedical
Materials Research Part A, pp. 497-504, 2006.
[87] M. G. Lampugnani, M. Corada, L. Caveda, F. Breviario, O. Ayalon, B. Geiger and E.
Dejana, "The molecular organization of endothelial cell to cell junctions: differential
association of plakoglobin, B-catenin, and a-catenin with vascular endothelial cadherin (VE-
cadherin)," The Journal of Cell Biology, vol. 129, no. 1, pp. 203-217, 1995.
[88] E. Dejana, "Endothelial cell-cell junctions: happy together," Nature Reviews Moledular Cell
Biology, vol. 5, no. 4, pp. 261-270, 2004.
[89] H. Peris, C. S. Bonder, T. H. Coates, D. J. Keating and C. F. Jessup, "The B-cell/EC axis:
how do islet cells talk to each other?," Diabetes, vol. 63, pp. 3-11, 2014.
[90] A. Johansson, J. Lau, M. Sandberg, L. Borg, P. U. Magnusson and P. O. Carlsson,
"Endothelial cell signalling supports pancreatic beta function in the rat," Diabetologia, vol.
52, pp. 2385-2394, 2009.

64
[91] M. M. Zanone, E. Favaro and G. Camussi, "From endothelial to B cells: insights into
pancreatic islet microendothelium," Current Diabetes Review, vol. 4, pp. 1-9, 2008.
[92] B. Keymeulen, G. Korbutt, M. De Paepe, F. Gorus, G. Kloppel and D. G. Pipeleers, "Long-
term metabolic control by rat islet grafts depends on the composition of the implant,"
Diabetes, vol. 42, no. 12, p. 1814, 1996.
[93] D. Pipeleers, P. In't Veld, E. Maes and M. Van De Winkel, "Glucose-induced insulin release
depends on functional cooperation between islet cells," Proceedings of the National
Academy of Sciences, vol. 79, pp. 7322-7325, 1982.
[94] D. W. Hopcroft, D. R. Mason and R. S. Scott, "Structure-function relationships in
pancreatic islets: support for intraislet modulation of insulin secretion," Endocrinology, vol.
117, no. 5, pp. 2073-2080, 1985.
[95] P. A. Halban, C. B. Wollheim, B. Blondel, P. Meda, E. N. Niesor and D. H. Mintz, "The
possible importance of contact between pancreatic islet cells for the control of insulin
release," Endocrinology, vol. 111, no. 1, pp. 86-94, 1982.
[96] Z. Liu, W. Kim, Z. Chen, Y.-K. Shin, O. D. Carlson, J. L. Fiori, J. K. Napora, R. Short, J. O.
Odetunde, Q. Lao and J. M. Egan, "Insulin and glucagon regulate pancreatic a-cell
proliferation," PLoS ONE, vol. 6, no. 1, p. e16096, 2011.
[97] K. Ramachandran, X. Peng, K. Bokvist and L. Stehno-Bittel, "Assessment of Re-aggregated
Human Pancreatic Islets for Secondary Drug Screening," British Journal of Pharmacology,
vol. 171, pp. 3010-3022, 2014.
[98] C. Wittig, M. W. Laschke, C. Scheuer and M. D. Menger, "Incorporation of bone marrow
cells in pancreatic pseudoislets improves posttransplant vascularization and endocrine
function," PLoS ONE, vol. 8, no. 7, p. e69975, 2013.

65
Appendices
Appendix A: Embedded BOEC Explants at Day 3
Figure 30: Modules embedded with 1 x 106 adMSC/mL collagen and 2 x 10
6 BOEC were
implanted in SCID-Bg mice and explanted at day 3. Histological staining shows CD31 vessel
structures are present at this time point but are not yet perfused with red blood cells (H&E and
trichrome), and thus have not yet become integrated with the host vasculature. n = 2.
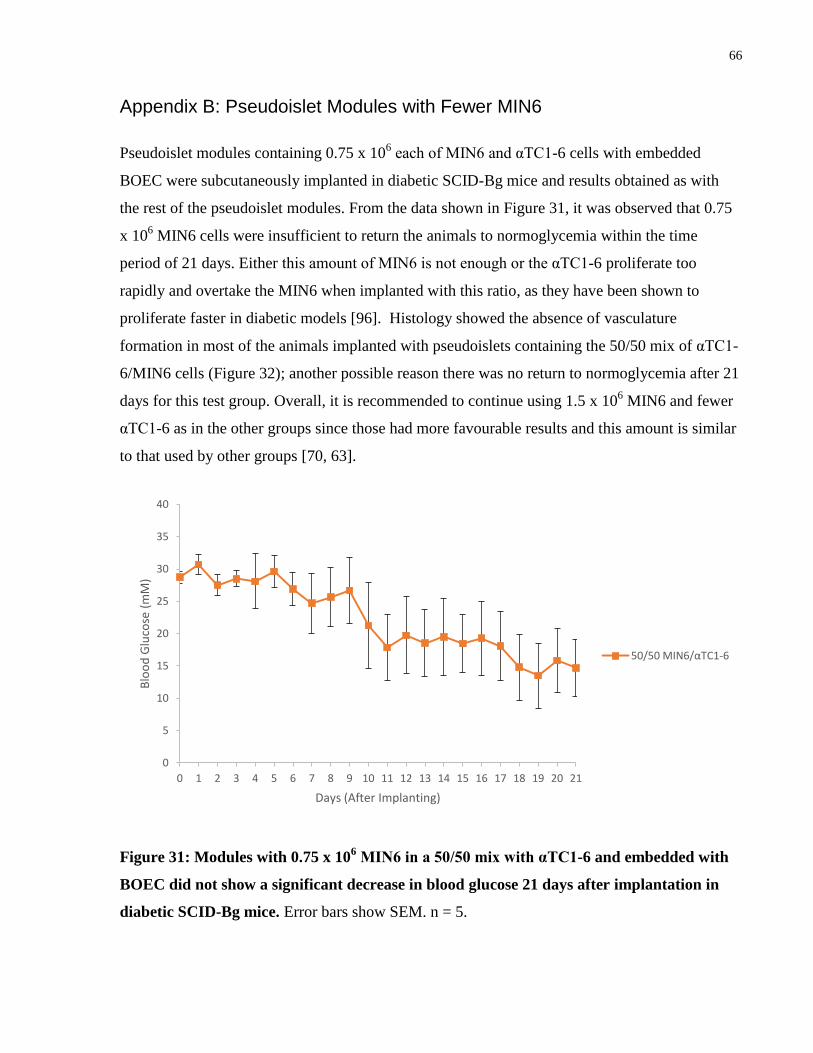
66
Appendix B: Pseudoislet Modules with Fewer MIN6
Pseudoislet modules containing 0.75 x 106 each of MIN6 and αTC1-6 cells with embedded
BOEC were subcutaneously implanted in diabetic SCID-Bg mice and results obtained as with
the rest of the pseudoislet modules. From the data shown in Figure 31, it was observed that 0.75
x 106 MIN6 cells were insufficient to return the animals to normoglycemia within the time
period of 21 days. Either this amount of MIN6 is not enough or the αTC1-6 proliferate too
rapidly and overtake the MIN6 when implanted with this ratio, as they have been shown to
proliferate faster in diabetic models [96]. Histology showed the absence of vasculature
formation in most of the animals implanted with pseudoislets containing the 50/50 mix of αTC1-
6/MIN6 cells (Figure 32); another possible reason there was no return to normoglycemia after 21
days for this test group. Overall, it is recommended to continue using 1.5 x 106 MIN6 and fewer
αTC1-6 as in the other groups since those had more favourable results and this amount is similar
to that used by other groups [70, 63].
Figure 31: Modules with 0.75 x 106 MIN6 in a 50/50 mix with αTC1-6 and embedded with
BOEC did not show a significant decrease in blood glucose 21 days after implantation in
diabetic SCID-Bg mice. Error bars show SEM. n = 5.
0
5
10
15
20
25
30
35
40
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21
Blo
od
Glu
cose
(m
M)
Days (After Implanting)
50/50 MIN6/αTC1-6
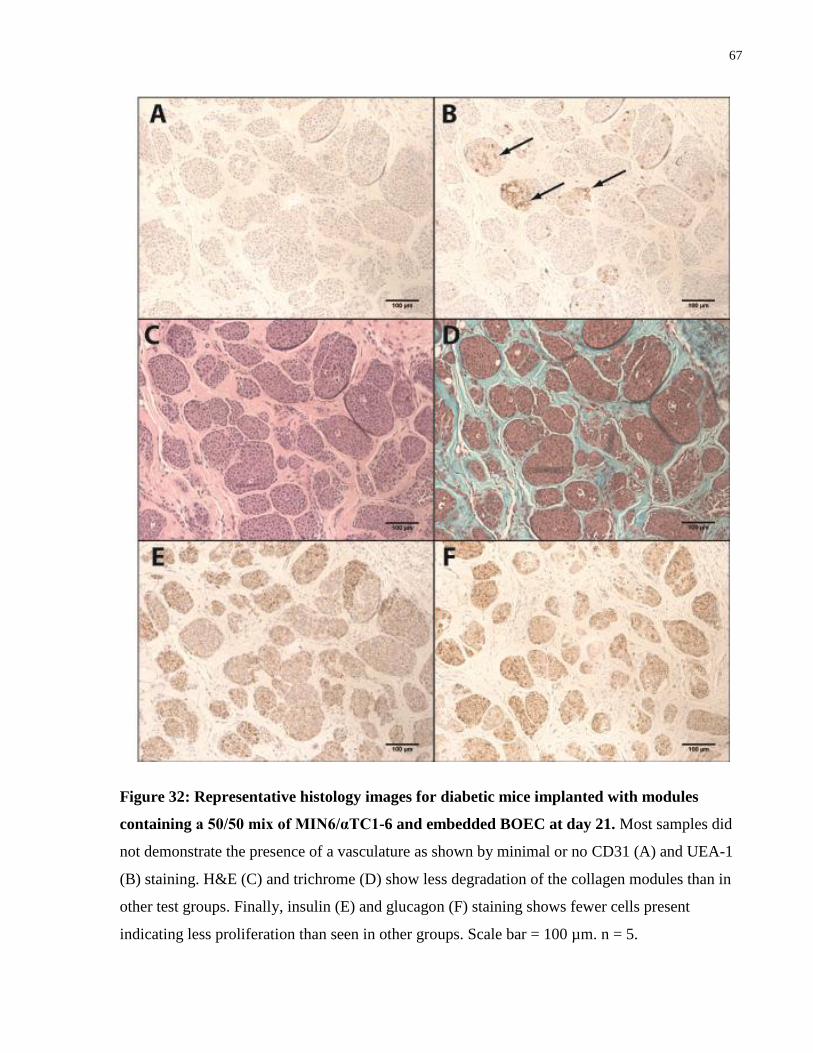
67
Figure 32: Representative histology images for diabetic mice implanted with modules
containing a 50/50 mix of MIN6/αTC1-6 and embedded BOEC at day 21. Most samples did
not demonstrate the presence of a vasculature as shown by minimal or no CD31 (A) and UEA-1
(B) staining. H&E (C) and trichrome (D) show less degradation of the collagen modules than in
other test groups. Finally, insulin (E) and glucagon (F) staining shows fewer cells present
indicating less proliferation than seen in other groups. Scale bar = 100 µm. n = 5.
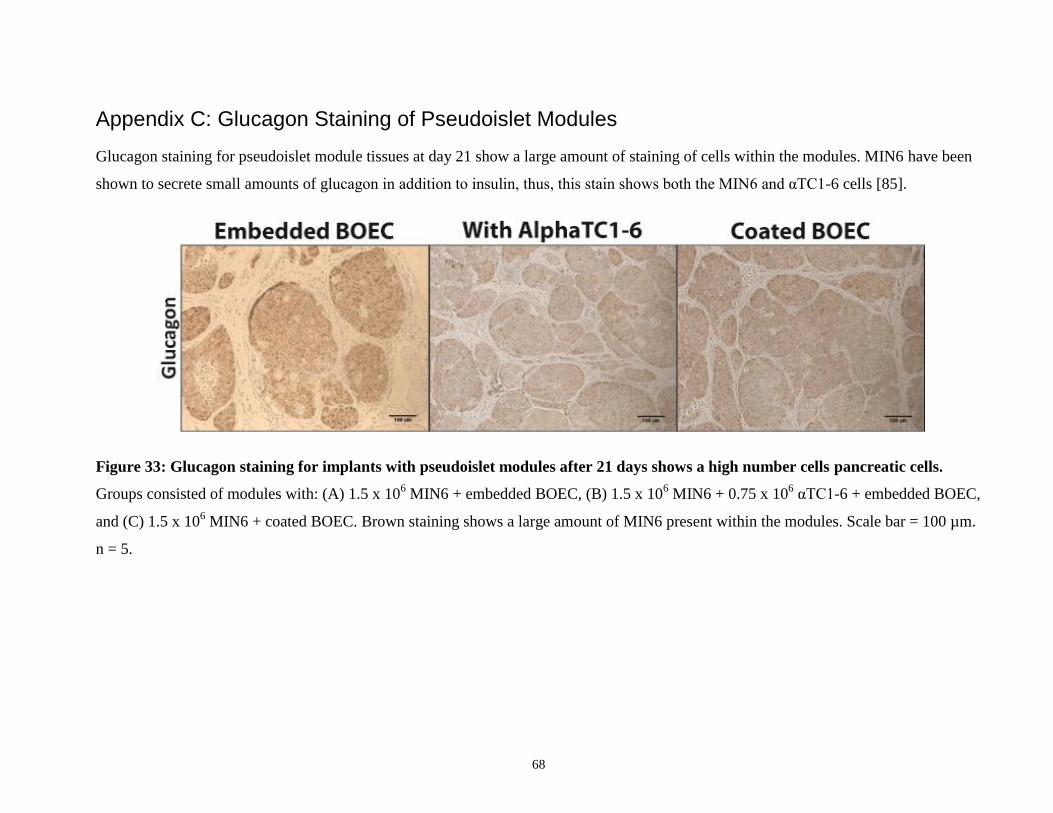
68
Appendix C: Glucagon Staining of Pseudoislet Modules
Glucagon staining for pseudoislet module tissues at day 21 show a large amount of staining of cells within the modules. MIN6 have been
shown to secrete small amounts of glucagon in addition to insulin, thus, this stain shows both the MIN6 and αTC1-6 cells [85].
Figure 33: Glucagon staining for implants with pseudoislet modules after 21 days shows a high number cells pancreatic cells.
Groups consisted of modules with: (A) 1.5 x 106 MIN6 + embedded BOEC, (B) 1.5 x 10
6 MIN6 + 0.75 x 10
6 αTC1-6 + embedded BOEC,
and (C) 1.5 x 106 MIN6 + coated BOEC. Brown staining shows a large amount of MIN6 present within the modules. Scale bar = 100 µm.
n = 5.













![Ani-Bot: A Modular Robotics System Supporting Creation ......the whole lifecycle of modular robotics, namely Creation, Tweaking, and Usage [29]. Therefore, we propose Ani-Bot, a modular](https://img.pdfslide.us/doc/110x75/5fece09f8387ec3643576562/ani-bot-a-modular-robotics-system-supporting-creation-the-whole-lifecycle.jpg)