Embed Size (px)
Citation preview

J. Cell Sci. 71, 17-35 (1984) 17Printed in Great Britain © The Company of Biologists Limited 1984
THE COMPETITIVE EFFECTS OF SERUM PROTEINS
ON CELL ADHESION
A. S. G. CURTISDepartment of Cell Biology, University of Glasgow, Glasgow GJ2 OTW, U.K.
AND J. V. FORRESTERUniversity Department of Ophthalmology, Aberdeen Royal Infirmary, Foresterhill,Aberdeen, U.K.
SUMMARY
Binding curves for the adsorption of plasma fibronectin, alpha-1-antitrypsin, alpha-2-macroglobulin, ceruloplasmin, transferrin and bovine serum albumin to plain and to hydroxylatedpolystyrene surfaces were measured. These curves were correlated with the adhesion of BHK cellsand leucocytes to these adsorbed protein surfaces in protein-free culture media. Hydroxylatedpolystyrene adsorbed less of alpha-1-antitrypsin, alpha-2-macroglobulin and albumin than the plainpolystyrene. On the other hand the hydroxylated surfaces bound more fibronectin than the plainpolystyrene surfaces. Hydroxylated polystyrene surfaces were also more adhesive for both BHKcells and leucocytes than plain polystyrene: a result confirming earlier work. The competition offibronectin for adsorption to plain polystyrene with alpha-1-antitrypsin, alpha-2-macroglobulin andceruloplasmin was measured and correlated with effects on cell adhesion. The results suggest thatthe low adhesiveness of BHK cells and leucocytes on plain polystyrene in sera-containing media isdue both to the low binding of fibronectin and to the binding of serum albumin, alpha-1-antitrypsinand alpha-2-macroglobulin. The relative unimportance of fibronectin in adhesion to these surfacesis shown by the finding that cell attachment will not occur to polystyrene surfaces that have boundhigh levels of the antiadhesive proteins in the presence of fibronectin, even though attachment willoccur in the absence of fibronectin provided that the antiadhesive proteins are lacking.
INTRODUCTION
A vast literature has been published on the effects of various sera and sera fractionson cell adhesion. Certain proteins have been identified as being of particular relevanceto the adhesion of cells to non-living surfaces and to other cells. Chief amongst theseproteins has been fibronectin (Weston & Hendricks, 1972; Hynes, 1976; Yamada &Olden, 1978; Grinnell, 1978), which is present in two forms, one of which circulatesin plasma and the other is bound to cell surfaces. Grinnell & Feld (1979) reported thatBHK fibroblasts have an absolute requirement for fibronectin for attachment andspreading on polystyrene surfaces, because they would not adhere under conditionsin which surface-bound and endogenous fibronectin were totally lacking. Such studiesrefer primarily to the adhesion of cells to 'tissue culture grade' polystyrene, which hasbeen corona discharge-treated, but adhesion and spreading of cells on the untreatedmaterial hardly occur in the presence of serum unless very high levels of fibronectinhave been bound (Grinnell & Feld, 1981).
Curtis, Forrester, Mclnnes & Lawrie (1983), however, found that BHK cells willadhere to and spread on highly hydroxylated polystyrene in the complete absence of

18 A. S. G. Curtis andjf. V. Forrester
fibronectin when the same conditions, as those imposed by Grinnell and Feld, ob-tained. They would not spread, however, on the less highly hydroxylated surfacesprovided by 'tissue culture' grade culture dishes. This result argues that fibronectinis not an essential requirement for the adhesion and spreading of fibroblasts or similarcell types.
Grinnell & Feld (1981, 1982) and Klebe, Bentley & Schoen (1981) found that theadsorption of fibronectin to polystyrene surfaces is influenced by the wettability of9uch surfaces; namely, the ratio of their hydrophobicity/hydrophilicity. The first twoof these three papers also showed that competition with various other serum proteinsoccurs when adsorption takes place from a serum medium. Forrester, Lackie & Brown(1983) found that the plasma proteins, alpha-1-antitrypsin and alpha-2-macroglobulin diminished leucocyte adhesion to glass coverslips. Nevertheless, wedescribe in this paper experiments that show that adsorption of these proteins rendersthe polystyrene surfaces wettable. Thus the question arises as to whether the dif-ferences in adhesion found in the presence of serum on 'bacteriological grade' and'tissue culture' grade dishes are due to the differing extents of fibronectin adsorption,influenced perhaps by competition with other plasma proteins, or whether they aredue to the differential adsorption of antiadhesive proteins such as alpha-1 -antitrypsin.Finally, the question must be raised again as to the nature of the feature shared byfibronectin and hydroxylated polystyrene, which aids cell spreading and is not foundin the plasma antiproteases.
This paper addresses these problems. The range of proteins that has been studiedhas been extended beyond those used earlier (Grinnell & Feld, 1981; Forrester,Lackie & Brown, 1983) to include the other two most common plasma proteins,namely transferrin and ceruloplasmin.
MATERIALS AND METHODS
CellsBHK21 C13 cells were obtained from stocks routinely grown in this laboratory, by trypsinization
of confluent or near-confluent cultures, using the method of Edwards & Campbell (1971). Poly-morphonuclear leucocytes were obtained from fresh human blood by the technique used by Forres-ter & Wilkinson (1981). The BHK cells were suspended in either Dulbecco's phosphate-bufferedsaline or Ham's F10 medium plus 3 % (w/v) foetal calf serum and the insulin-transferrin-selenitesupplement (ITS) (Collaborative Research, Waltham, Md, U.S.A.) with insulin and transferrineach at S/igml"1 at a cell density of 2 x 105ml~'. Leucocytes were suspended at a density of 1-5 X10* ml"1 in Hepes-buffered Hanks' balanced salts solution with or without 10% foetal calf serum(purchased from Gibco-Biocult, Paisley, U.K.).
Culture dishesPolystyrene culture dishes, bacteriological and tissue culture grade were purchased from the
following makers: Sterilin, Teddington, Middx, U.K., Nunc through Gibco-Biocult, Paisley,U.K., and Corning, Inc through MacFarlane Ribson, Glasgow, U.K.
The bacteriological dishes were given the following treatments: chloric acid treatment by adding3 ml of 70 % perchloric acid and 2 ml saturated aqueous potassium chlorate to the dishes andallowing these to react for 10 min. Control dishes (bacteriological) were stripped of residual mould-release agent by treatment for 30 min with ethanolic sodium hydroxide (5 M in 70 % (v/v) ethanol)

Adsorbed proteins and cell adhesion 19
followed by extensive washing with tap water. Subsidiary experiments showed that tap water wasno different from distilled water as a wash.
Measurement of cell adhesionBHK cells (0-6 X 106 in a final volume of 3 ml) or 1 X 106 leucocytes (in a final volume of 4 ml)
were placed in each culture dish, which was then incubated at 37 °C for 15 min (BHK cells) or 30 min(leucocytes). Cell spreading during these incubation periods is inappreciable on control surfaces.Those cells that had not adhered in these culture periods were removed from the dishes by washingwith Hepes-buffered Hanks' balanced salts solution three times. The number of adherent cells ineach of 10 standard counting areas was then counted using phase-contrast microscopy to detect thecells and a Quantimet 720 image analysing computer to count the cells. The counting areas were0-0026 cm2. The results are expressed in terms of the number of cells adhering per cm2.
Proteins used in binding studiesHuman transfemn, alpha-1-antitrypsin, ceruloplasmin and bovine serum albumin were pur-
chased from Sigma Chemical Co., Poole, U.K. Human serum albumin was purchased fromBehringwerke, Marburg, West Germany. Alpha-2-macroglobulin was prepared by the methoddescribed by Forrester, Wilkinson & Lackie (1983). Fibronectin (bovine) was prepared from bovineserum in this laboratory by affinity chromatography on gelatin-Sepharose columns, using themethod described by Engvall & Ruoslahti (1977). The transferrin was converted to the iron-saturated form by reacting it with ferric nitrilotriacetate in stochiometric proportions and the excessand free nitrilotriacetic acid was removed by dialysis. Bovine immunoglobulin G was prepared frombovine serum by column chromatogTaphy on DEAE-cellulose at pH 8-0 taking the fraction elutedby 0-005 M-phosphate buffer. The purity of these proteins waa checked by sodium dodecylsulphate/polyacrylamide gel electrophoresis (SDS-PAGE) systems, fibronectin degradationproducts were found at low levels in the fibronectin and very low levels of two other proteins werefound in the serum albumin. No impurities could be detected in the alpha-2-macroglobulin,ceruloplasmin or alpha-1-antitrypsin.
Radioactive labelling of proteinsTransferrin, alpha-2-macroglobulin, ceruloplasmin, albumin and alpha-1-antitrypsin were
labelled with I2SI using the iodogen labelling method (Pierce Chemical Co.). Carrier-free sodiumiodide was obtained from Amersham International, Amersham, U.K. The labelled proteins wereseparated using column chromatography on Biogel P10 columns (Biorad, Watford, Herts, U.K.)with 0-1 M-Hepes buffer (pH8-0) as eluant. The labelled proteins were used within 3 weeks oflabelling, being stored at — 25 °C until just before use.
Binding of proteins to polystyrene surfacesProtein solutions diluted to levels of between 5-500 fig ml"1 with Ham's F10 saline (serum-free)
were placed in 3-ml samples in the dishes and allowed to bind by absorption for 30 min at 25 °C. Theexact conditions of binding are described separately for each experiment. At the end of the bindingperiod unbound protein was washed off with four washes of Ham's F10 saline.
Measurement of protein bindingProtein binding was measured by counting the activity of I25I, on a gamma counter (Wilj Interna-
tional, Ashford, Kent, England), of known amounts of the protein solutions and then counting thedishes to which radioactive proteins had been bound. In the earlier part of the work the dishes werecut into strips on a hot wire cutter and the pieces of dish were loaded into the counting tubes. Laterin the experiments dishes were extracted twice with small volumes of sodium hydroxide (2 M) andthe counts extracted were measured. Both methods gave almost identical results. In all thoseexperiments in which the attachment of cells to adsorbed protein surfaces was being measured, theindividual dishes in which attachment assays for adhesion had been carried out were then countedto measure the bound protein remaining after cell attachment. No evidence was obtained fromsubsidiary experiments that the short incubation of the dishes in the presence of cells caused any lossof bound protein into the medium.
20 A. S. G. Curtis andjf. V. Forrester
RESULTS
Binding offibronectin to various polystyrene surfaces and cell adhesion thereto
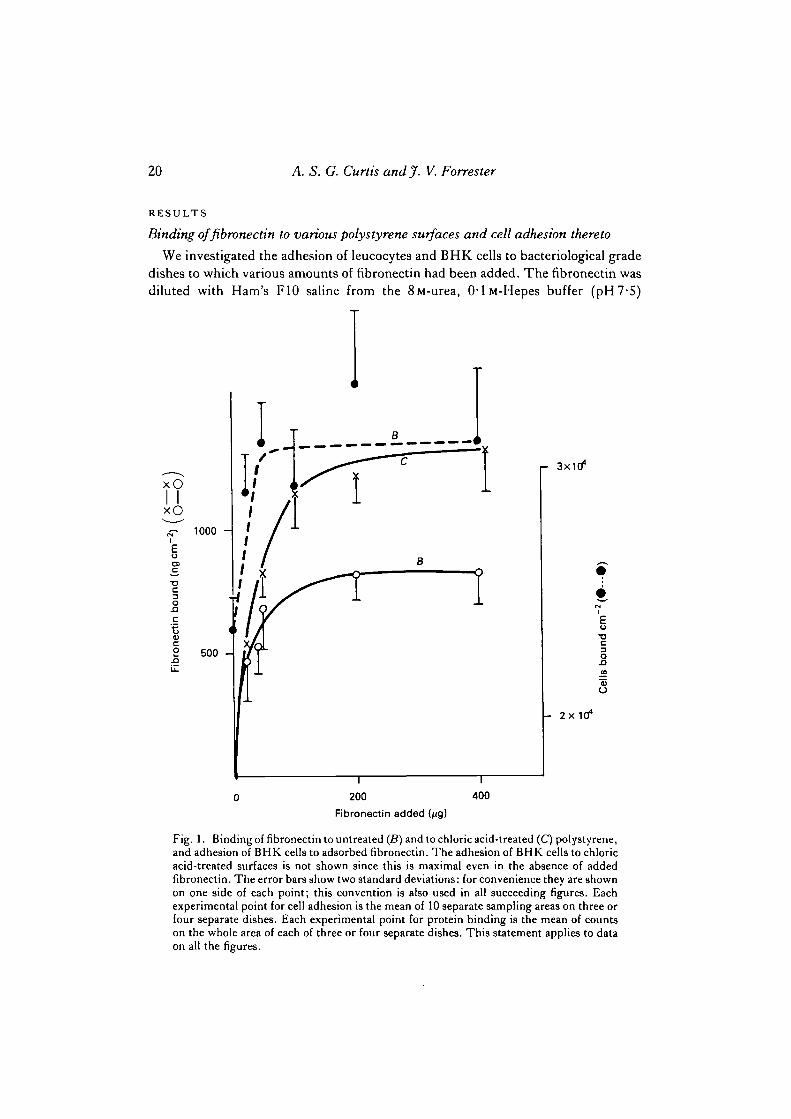
We investigated the adhesion of leucocytes and BHK cells to bacteriological gradedishes to which various amounts of fibronectin had been added. The fibronectin wasdiluted with Ham's F10 saline from the 8M-urea, 0-lM-Hepes buffer (pH7-5)
xO
,c- 1000 -
oCD
T3C•zo.a
t5IDCo 500 -
r 3X104
•oco.Q
- 2X104
0 200Fibronectin added (j/g)
Fig. 1. Binding of fibronectin to untreated (B) and to chloric acid-treated (C) polystyrene,and adhesion of BHK cells to adsorbed fibronectin. The adhesion of BHK cells to chloricacid-treated surfaces is not shown since this is maximal even in the absence of addedfibronectin. The error bars show two standard deviations: for convenience they are shownon one side of each point; this convention is also used in all succeeding figures. Eachexperimental point for cell adhesion is the mean of 10 separate sampling areas on three orfour separate dishes. Each experimental point for protein binding is the mean of countson the whole area of each of three or four separate dishes. This statement applies to dataon all the figures.

Adsorbed proteins and cell adhesion 21
Table
Dish type
1. The binding of fibronectin to polystyrene
f
dishes and cell adhesion
Cell adhesion
BHK cells Leucocytes
Controls: no fibronectin
Bacteriological grade 10677(30%) 49292(14%)Chloric acid treated 32485 (22%) 44285 (36%)Tissue culture grade 13435(24%) 44574(14%)
Experimental12JI bound
Fibronectin on bact. grade 292(18%) 17296(25%) 12489(37%)Fibronectin on chloric acid treated 394(24%) 36485(25%) 44423(17%)
Adhesion is expressed as cells bound cm"2. Binding is expressed as disints per min cm"2 per40/Jg of fibronectin added per dish.
Figures in parenthesis are the standard deviations expressed as percentages of the mean. Eachmeasurement is the mean of 10 individual measurements of three dishes for experimental measure-ments; for control results the measurements are the means of at least 10 counts per dish on at leastfour dishes. The maximum number of leucocytes available for adhesion was 52910 cm"2 and themaximum number of BHK cells 31750cm"2. The labelled fibronectin had a specific activity of0'3 [tCi mg"1. The measurements of disints refer to the mean of six measurements, one on each dish.The measurements were carried out in serum-free medium. See Fig. 1 for further data.
solution in which it had been stored, to 1 M in urea, immediately before addition tothe dishes. Adsorption was allowed to continue for 30 min and then excess unboundfibronectin was washed off with Ham's F10 saline, reducing the urea level to very lowvalues. The effect on the adhesion of BHK cells of binding varying amounts to thedishes on cell adhesion is shown in Fig. 1.
The data in Fig. 1 established the amount of fibronectin that should be bound inorder to increase the adhesion of BHK cells to the dishes appreciably and this levelwas used in the main experiments thereafter. Table 1 shows that fibronectin on plainpolystyrene has the effect of reducing the adhesion of the leucocytes. This resultconfirms an earlier report by Brown & Lackie (1981). The data presented in Fig. 1also demonstrate that chloric acid treatment enhances the saturation level and possiblythe affinity of polystyrene surfaces for fibronectin. This enhancement of binding bychloric acid treatment appears to be unique for fibronectin amongst the proteinsstudied in this work.
Table 1 and Fig. 1 show that appreciable fibronectin binding occurs on bacterio-logical grade dishes and that this raises the binding of BHK cells. Both results confirmthe results of many other groups, for instance Yamada & Olden (1978). Binding offibronectin to chloric acid-treated surfaces is greater than that seen on bacteriologicalgrade dishes. The binding of fibronectin to these treated dishes does not, however,increase the adhesion of BHK cells, which was of course already very high as a resultof the chloric acid treatment (Curtis et al. 1983). In contrast, leucocyte adhesion is

22 A. 5. G. Curtis andj. V. Forrester
reduced when fibronectin is bound to an untreated dish surface, but not when it isbound to a chloric acid-treated surface. In a subsequent section we describe the effectsthat result from competition between other proteins and fibronectin in binding totreated and untreated polystyrene surfaces, and the resulting effects on cell adhesion.
Binding of alpha-1-antitrypsin to various polystyrene surfaces and cell adhesionthereto
The data in Table 2 and Fig. 2 show that alpha-1-antitrypsin binds to untreatedsurfaces at a saturation level of 300 ngcrn"2 while chloric acid treatment of the dishes
500 -
I 450u
T3C
s 300 -
150 -
IItIt
r 3xio"
ITI\
- 2-5X104
- 2X104
I II I
- 1-5X104
I100AAT added
200
Fig. 2. The binding of alpha-1-antitrypsin (AAT) to untreated (B) and to chloric acid-treated polystyrene (C) and the adhesion of BHK cells to alpha-1-antitrypsin adsorbed tosuch surfaces. Note that the adhesion of BHK cells to the protein on untreated surfaces( • -—•) rises at very low levels of adsorbed protein but is reduced at higher levels.(X — X) Adhesion of cells to chloric acid-treated surface.

Adsorbed proteins and cell adhesion 23
Table 2. The binding of alpha-1-antitrypsin to polystyrene surfaces and cell adhesionthereto
Cell adhesion
Dish type BHK cells Leucocytes
Controls: no antitrypsin
Bact. grade 10677(30%) 49292(14%)Chloric acid-treated 32485 (9%) 34105 (15%)
Experimental12JI bound
Bact. grade +200 fig 3866(1%) 11340(8%) 26666(25%)Bact. grade +100 jig 2662 (3%) 14775 (12%) 32160 (20%)Chloric acid-treated +220 fig 2941(3%) 30811(7%) 34694(10%)
Adhesion is expressed as cells bound cm"2. Binding of protein is expressed disints per min cm"2.Additions of protein, 200 or 100/Jg as stated below.
Specific activity of antitrypsin was 4-85 ^Cimg"1. The maximal possible cell adhesion.Leucocytes, 52910cm"2; BHK cells, 31750cm"2. Cell counts, means of 10 areas on each of threedishes. Binding counts, means of three dishes for experimental dishes. Cell counts, means of fouror more dishes for controls. For fuller data on the binding of the protein to plain and to hydroxylatedpolystyrene and BHK cell adhesion see Fig. 2.
Table 3.
Dish type
The binding of alpha-2-macroglobulin to polystyrene and cell adhesion
Cell adhesion
BHK cells^
Leucocytes
Controls: no macroglobulin
Bact. grade 10677(41%) 49292(14%)Chloric acid-treated 31485(7%) 44285(5%)
Experimental12SI bound
Bact. grade 2176 (21%) 14664 (43%) 278 (88%)Chloric acid-treated 452(60%) 23758(17%) 45633(17%)
Adhesion is expressed as cells bound cm"2. Binding of protein is expressed as disints permin cm"1 per 100 fig of macroglobulin added per dish.
1251-labelled macroglobulin, sp. act. 19'5 ̂ Ci mg"1. The maximum number of leucocytes availablefor adhesion was 52910 cm"2, and the maximum number of BHK cells available was 31750 cm"2.The adhesion assay was in serum-free medium. The cell counts are the means and standard devia-tions (expressed as percentages of the mean) of 10 counts on each of three dishes for the experimentalseries, and of ten counts on each of ten dishes (control series). The binding counts are the meansand standard deviations of counts of three dishes in each group. See Fig. 3 for further data.
reduces the binding to about 70 % of this value. The adhesion of leucocytes is reducedsignificantly when they attempt to adhere to the alpha-1-antitrypsin surface onuntreated polystyrene, but there is no reduction of the adhesion of leucocytes attaching

24 A. S. G. Curtis andj. V. Forrester
50 -
c
ll25 -
EO
6
- 2x10*
1x10*
200
a-2-M added (/ig dish
oII
ou
Fig. 3. The binding of alpha-2-macroglobulin (a-2-M) to untreated polystyrene surfaces( # • ) and the adhesion of BHK cells to the adsorbed protein (O—-O). Note that,as with alpha-1-antitrypsin, very low levels of adsorbed alpha-2-macroglobulin enhanceBHK cell adhesion.
Table 4. A. The binding of55Fe-labelled iron-rich transferrin to polystyrene and celladhesion thereto
Cell adhesion
Dish type 5SFe bound BHK Leucocytes
Bact. gradeChloric acid-treatedChloric acid-treated
1033 (5%)469 (5%)
No transferrin
28483 (11%)27087 (12%)32485 (17%)
46062 (19%)43895 (30%)51562 (40%)
Bact. gradeChloric acid-treated
B. The binding of l2sI-labelled apotransferrin and cell adhesion
1152(5%) 27147(16%) 44050(26%)390 (4%) 26529 (14%) 49147 (27%)
Adhesion of cells is expressed as cells bound cm 2. Binding is expressed as disints per min cm 2
per 300 fig of transferrin added to each dish. This gives approx. 90% saturation.55 1fg55Fe-transferrin (sp. act.125
mg"1).(p f g )125I-labelled apotransferrin (sp. act. 3-0/iCimg"1). Maximal cell binding. Leucocytes,
52910cm"2; BHK cells, 31750cm"2. For other conditions see Table 1.
to alpha-1-antitrypsin bound to chloric acid-treated surfaces. The difference is fargreater than can be accounted for by the reduced protein binding. The protein whenbound to chloric acid-treated surfaces does not reduce the adhesion of BHK cells.Low levels of binding of alpha-1-antitrypsin on plain polystyrene raise the adhesionof BHK cells above the control level found in the absence of the protein but this effect

Adsorbed proteins and cell adhesion 25
disappears at higher levels of binding. The difference in the effects of the proteinwhen bound to untreated and to treated surfaces cannot be explained by the differencein binding since this is relatively slight.
Table 5. The binding of serum albumin to polystyrene surfaces and cell adhesionthereto
Cell adhesion
Dish type 125I bound BHK cells Leucocytes
Bact. gradeChloric acid-treated
1874960
287 (114%)7998 (35%)
19302 (38%)52766 (18%)
100jig albumin was added per dish. Adhesion is expressed as cells bound cm 2. Binding isexpressed as disints per mincm"2.
For controls without absorbed albumin see Tables 1-3. Specific activity of labelled albumin,3-15/<Cimg~'. Maximal cell attachment: leucocytes, 52910cm~2; BHK cells, 31750cm"2. Resultsare the means and standard deviations of 10 measurements on each of five dishes. The binding ofalbumin corresponds to 0-26 j*gcm~2 (bacteriological grade) and subsidiary experiments show thatthis is about 90% saturation.
100 - I
CD_cT3C3
o
COaa.o
50 -
100
Protein added (/jgdish)
200
Fig. 4. The adsorption of ceruloplasmin to untreated (X-treated ( • • ) polystyrene.
-X) and to chloric acid-
CEL71

26 A. S. G. Curtis andj. V. Forrester
Binding of alpha-2-macroglobulin to various polystyrene surfaces and cell adhesionthereto
Alpha-2-macroglobulin resembles antitrypsin in its effects on adhesion (see Table3 and Fig. 3). Its saturation binding to plain hydrophobic polystyrene is about fourtimes greater than to the hydroxylated polystyrene (chloric acid-treated). Alpha-2-macroglobulin reduces leucocyte adhesion when bound to plain untreatedpolystyrene, to a very marked degree (see Table 3). The level of binding achieved withthe addition of 100 fig of protein per dish (3 ml volume) is 52ngcm~2 and this issufficient to inhibit leucocyte adhesion, but leaves the unhydroxylated surface moreadhesive for BHK cells than the control surface (see Table 3). Examination of Fig.3 shows that, as with alpha-1-antitrypsin, alpha-2-macroglobulin adsorbed to plainpolystyrene surfaces enhances BHK adhesion most at very low levels of binding.Alpha-2-macroglobulin has little or no effect on the adhesion of leucocytes to chloricacid-treated surfaces at the rather low binding level of 8• 75 ng cm"2, but reduces BHKadhesion from about 31000 cells cm"2 to about 24000 cells cm"2. Alpha-2-macroglobulin and alpha-1-antitrypsin are alike in that they are most effective in
oCDc
•o
O
2X104 -="
Ceruloplasmin (;tg) 200
Fig. 5. The adhesion of leucocytes (O, A) and BHK cells (X, • ) to ceruloplasminadsorbed to untreated (O, X) or to chloric acid-treated (A, • ) polystyrene. The ordinateis calibrated in ng of ceruloplasmin added per dish. See Fig. 4 for details of adsorption ofthis protein.

Adsorbed proteins and cell adhesion 27
reducing adhesion when bound to the hydrophobic untreated surface. They affect
leucocyte adhesion to a greater extent than BHK cell adhesion.
Binding of transferrin and of albumin to various polystyrene surfaces and cell ad-hesion thereto
Both iron-saturated transferrin and apotransferrin bind more extensively to bac-teriological grade polystyrene than to the chloric acid-treated material (see Table 4).They have little effect on cell adhesion other than raising the adhesion of BHK cellsto untreated polystyrene. The results of experiments with bovine serum albumin areshown in Table 5: they indicate that hydroxylated polystyrene surfaces have a reducedbinding for albumin when compared with the bacteriological grade dishes. Albuminclearly reduces the adhesion of BHK cells to either type of polystyrene surface evenat relatively low bindings, and reduces leucocyte adhesion to bacteriological surfaces,but has no effect when bound to hydroxylated surfaces.
- 3X104
- 2-5x10*
u
CD
o
- 2x10*
1-5x104
50 100 150
Fibronectin added per dish (H
Fig. 6. The competitive adsorption of alpha-1-antitrypsin (AAT; 150/ig added) tountreated polystyrene ( • • ) in the presence of increasing concentrations of fibro-nectin. The effect on BHK cell adhesion (X x) is also shown. Note that fibronectinreduces the binding of alpha-1-antitrypsin.

28 A. S. G. Curtis andj. V. Forrester
The effects of bound ceruloplasmin on cell adhesion
Ceruloplasmin binds to a similar level on both chloric acid-treated polystyrene andthe untreated material (see Fig. 4). On both types of surface it acts to reduce theadhesion of both leucocytes and BHK cells (see Fig. 5), but the effect with plainpolystyrene plates is small presumably because of the low adhesion of these surfacesin the absence of adsorbed protein.
Does fibronectin compete with alpha-1-antitrypsin, alpha-2-macroglobulin orceruloplasmin for binding with the substrate?
The data presented above were obtained from experiments in which serum wasabsent and in which usually only a single species of protein had been bound to the dishsurface. It is pertinent to ask whether the proteins would compete with each other forbinding to the treated or untreated polystyrene surfaces. Alpha-1-antitrypsin bindsmore extensively to hydrophobic than to hydrophilic surfaces and depresses adhesion
400 -T i- 4xio4
300
i.Q
200 -
100 -
uCDC
aT3
- 1x10"
400
Alpha anti-trypsin added per dish I
Fig. 7. The converse of Fig. 6. The adsorption of fibronectin ( • • ) in the presenceof alpha-1-antitrypsin; 80 fig fibronectin was present in the dish during adsorption. Notea slight reduction in the adhesion of BHK cells (X X) to the adsorbed protein onuntreated polystyrene.

Adsorbed proteins and cell adhesion 29
- 3x10*
- 2X104
- 1X104 <3
T0 50 100 150
Fibronectin added (ftg)
Fig. 8. The competitive binding of alpha-2-macroglobulin (100 /ug added) in the presenceof fibronectin (O O) to untreated polystyrene. The effect on BHK cell adhesion isalso shown (X X).
of leucocytes and/or BHK cells-when so bound (Fig. 2, Table 2). Fibronectin in-creases the adhesion of BHK cells to the surfaces on which it is bound. In consequencewe ask whether fibronectin competes with alpha-1-antitrypsin for binding to variouspolystyrene surfaces and whether cell adhesion thereto is changed according to therelative binding. A constant amount of alpha-1-antitrypsin was used in competitionwith varying amounts of fibronectin, and the binding of the antitrypsin to ahydrophobic surface and the adhesion of the two cell types were determined: theresults are shown in Fig. 6. Clearly, fibronectin reduces alpha-1-antitrypsin bindingand the adhesion of BHK cells increases as the fibronectin level rises and alpha-1-antitrypsin decreases. The experiment was repeated in the converse mode, usinglabelled fibronectin and increasing amounts of antitrypsin (see Fig. 7) with parallelresults. The experiment was confined to hydrophobic surfaces since it is only on thesethat appreciable inhibition of adhesion occurs with bound alpha-1-antitrypsin.
A similar pair of experiments were carried out on the competition of alpha-2-macroglobulin with fibronectin (see Figs 8, 9). The results were similar to those withalpha-1-antitrypsin.

30 A. S. G. Curtis andjf. V. Forrester
"DcDO
XI
co
r 3x10"
-2x104
-1X10 4
u•oco
XI
40 60 80 100ct-2-Macroglobulin added (j/g dish)
Fig. 9. The converse of Fig. 8. The adsorption of fibronectin to untreated polystyrene( • • ) in the presence of alpha-2-macroglobulin; 80 ng was present in the dish duringadsorption. The effect on BHK cell adhesion is also shown (X X).
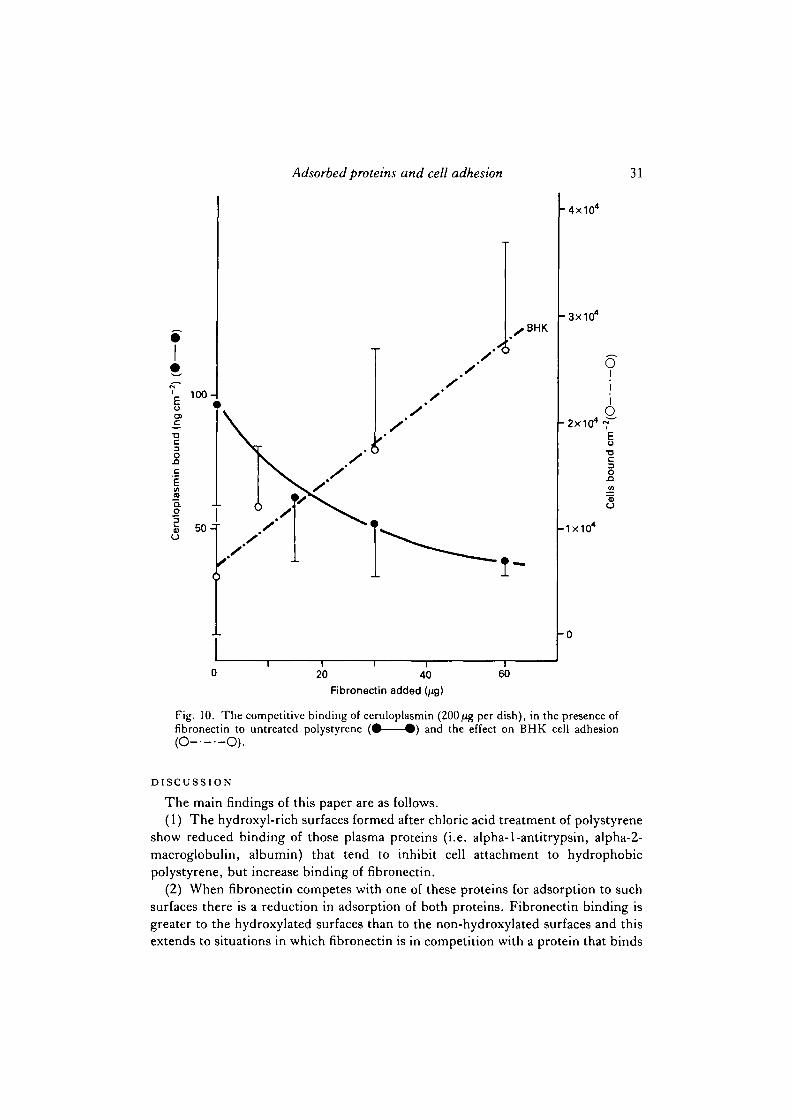
Fibronectin competes with saturating levels of ceruloplasmin to reduce binding ofthat protein to about 40% of its saturation level (Figs 10, 11). The adhesion level ofBHK cells to such surfaces falls to about a third of the level that would be found ona surface saturated with fibronectin, even though the bound fibronectin level is suf-ficient (see Fig. 1) by itself to maintain an adhesion level of 90 % of maximal values.In other words the depression in adhesion is not solely due to a lack of fibronectin butalso to the presence of ceruloplasmin.
Clearly, if fibronectin competes to reduce the binding of these antiadhesive proteinsto the dishes, cells should adhere well to hydrophobic dishes provided that theproteins could be removed from the serum used to prepare the culture medium.Unfortunately this does not appear to be practical, but it is possible to carry out theexperiment in a similar manner. We prepared an artificial serum from albumin,transferrin and immunoglobulin G (for composition see Table 6), and measured cellattachment in this medium with and without the addition' of antitrypsin or macro-globulin. The results (Table 6) show that our prediction is supported and is consistentwith the idea that fibronectin reduces the binding of alpha-1-antitrypsin, alpha-2-macroglobulin and ceruloplasmin to a hydrophobic surface and thus maintains itsadhesiveness.
Adsorbed proteins and cell adhesion 31
•oc
io.oEo
100-1
soq
-4x10*
-3X104
-2X104
oIi6o
•o
co
.O
3-1x10*
0 20 40Fibronectin added (/jg)
Fig. 10. The competitive binding of ceruloplasmin (200^g per dish), in the presence offibronectin to untreated polystyrene ( •——•) and the effect on BHK cell adhesion(O O).
DISCUSSION
The main findings of this paper are as follows.(1) The hydroxyl-rich surfaces formed after chloric acid treatment of polystyrene
show reduced binding of those plasma proteins (i.e. alpha-1-antitrypsin, alpha-2-macroglobulin, albumin) that tend to inhibit cell attachment to hydrophobicpolystyrene, but increase binding of fibronectin.
(2) When fibronectin competes with one of these proteins for adsorption to suchsurfaces there is a reduction in adsorption of both proteins. Fibronectin binding isgreater to the hydroxylated surfaces than to the non-hydroxylated surfaces and thisextends to situations in which fibronectin is in competition with a protein that binds

32 A.S.G. Curtis andj. V. Forrester8OO-1 r-4xio4
-3x10*OIi
O
u
cOJ3
O- 1x104
Ceruloplasmin added (/<g)Fig. 11. The converse of Fig. 10. The competitive binding of fibronectin in the presenceof ceruloplasmin ( • • ) to untreated polystyrene and the effect on BHK cell adhesion(O O); 80 fig of fibronectin was present per dish during the adsorption.
Table 6. The binding of cells to proteins adsorbed from synthetic sera
Control, no serumAlbumin, transferrin, IgGAlbumin, transferrin, IgG,
alpha-1-antitrypsinAlbumin, transferrin, IgG,
alpha-2-macroglobulinAlbumin, transferrin, IgG,
alph-1 -antitrypsin,alpha-2-macroglobulin
BHK cells
27989 (43%)31430 (17%)24174 (18%)
665 (63%)
673 (101%)
Cells bound cm 2
A\
Leucocytes
55410 (17%)45146 (19%)24326 (54%)
N.D.
N.D.
Bacteriological grade polystyrene. Proteins preadsorbed to dish, no proteins added to culturemedium.
General conditions as for other experiments. Protein mixture solutions contained 3-5 mgml"' ofhuman serum albumin, 0-35 mgml"1 human transferrin, 017mgml~' immunoglobulin G (IgG)and for experiments with BHK cell 35^gml~' bovine fibronectin. Alpha-1-antitrypsin was at aconcentration of 0-15 mg ml"1. Alpha-2-macroglobulin was at a concentration of 0-3 mg ml"1. Theseconcentrations approximate to the levels to be found in a 10% serum medium (see Diem & Lentner(1970) for data on serum composition). Data are the means and standard deviations of 10 measure-ments on three different dishes per treatment. N.D., not determined.

Adsorbed proteins and cell adhesion 33
preferentially to a non-hydroxylated surface. The adhesion of BHK cells to suchsurfaces of two adsorbed proteins is reduced below the level that would be expectedfor the actual level of fibronectin bound. The effect is more marked on hydrophobicsurfaces than on hydrophilic surfaces.(3) The adsorption of alpha-2-macroglobulin or alpha-1-antitrypsin to plain(hydrophobic) polystyrene inhibits leucocyte adhesion but actually increases BHKcell adhesion at low levels of binding. The fall in BHK adhesion at higher levels ofbinding of these proteins might indicate a conformational change in the adsorbedlayer.
The results obtained by Grinnell & Feld (1981, 1982) and by Klebe et al. (1981)showed that fibronectin and bovine serum albumin compete in their binding to bothbacteriological and tissue culture grade polystyrene as well as to other plastics. Grin-nell & Feld (1982) found that other unidentified proteins from serum also competedin adsorption with fibronectin: we suggest that these proteins may be alpha-1-antitrypsin and alpha-2-macroglobulin. Grinnell & Feld (1982) suggested thatfibronectin binding may be aided by albumin adsorption at very low levels of the latterprotein. We did not test for this with albumin but found no evidence for such aphenomenon with ceruloplasmin, alpha-2-macroglobulin or alpha-1-antitrypsin.
Our results suggest that the failure of cell adhesion, or presumably of cell spreading,on untreated polystyrene surfaces (hydrophobic surfaces) is to be explained, not byan inadequate binding of plasma fibronectin from the serum in the medium, but bythe level of binding of the antiadhesive proteins. This interpretation is supported bythe experiment with artificial sera reported in Table 6, in which exogenous fibronectinwas absent. Addition of these proteins diminished adhesion appreciably. On the otherhand the close parallel between declining fibronectin binding in the presence of alpha-2-macroglobulin and cell adhesion (seen in Figs 7 and 8) argues to the contrary.However, since such very small quantities of alpha-2-macroglobulin (less than40 ngcm"2) would only occupy a small fraction of the area that might otherwise havebeen taken up by fibronectin, and since the fibronectin level remains high enough byitself to support a fairly high level of cell adhesion, it seems likely that the effects ofthe antiadhesive proteins are direct. This conclusion can be reached even more directlyby noting the reduction in adhesion once one of the proteins has been adsorbed to asurface (see for example, Figs 2 and 3).
We do not know the nature of the binding sites on polystyrene dishes for proteins,but it seems likely that proteins that bind preferentially to hydrophobic surfaces willdo so by various hydrophobic interactions whilst those proteins adhering tohydroxylated surfaces may do so by hydrogen bonding. In view of these considera-tions it seems unlikely that the proteins compete for the same sites even though theymay compete for space. It is worth noting that the mode of adsorption of proteins tohydrophobic and to hydroxylated surfaces may be vastly different. Binding of aprotein to a hydrophobic surface may result in its hydrophilic groupings facing out-wards into the medium whilst binding of a protein to a hydrophilic surface may resultin the protein displaying some of its hydrophobic groupings to the medium.
Grinnell & Feld (1982) found appreciable adhesion of cells to bacteriological grade

34 A. S. G. Curtis andjf. V. Forrester
polystyrene in the absence of fibronectin whereas we did not. The explanation of thisdifference almost certainly lies in the fact that the BHK cells they used had beenexposed to serum components for some hours after trypsinization and before their usein adhesion assays in serum-free media, while in the present work no serum com-ponents were allowed to come in contact with the cells after trypsinization. Subsidiaryexperiments carried out by us show that even brief exposure of BHK cells to serummedia allows sufficient adsorption of components for the cells to become markedlyadhesive to plain polystyrene.
Various authors (e.g. Van Oss, Gillman & Neumann, 1975) have proposed thatwettability of a surface is directly correlated with adhesiveness for cells. It is note-worthy that we observe that adsorption of alpha-1-antitrypsin, alpha-2-macroglobulinor ceruloplasmin to bacteriological dishes renders these surfaces wettable butdiminishes cell adhesion. Such observations would seem to destroy the wettabilityhypothesis, but we remark, in its defence, that the cells may be making local areas ofsuch surfaces unwettable so that they are unable to adhere to the nearby surfaces.
One of the questions that we have not resolved in this work is the extent of surfacecoverage required for an effect on cell adhesion. Grinnell & Feld (1981) suggest thatfibronectin coverage must approach 100% for maximal effects on cell adhesion toappear. One important question that needs an answer is whether suboptimal levels offibronectin (or other protein) adsorption lead to 100% coverage by a more unfoldedmolecule, or whether the configuration of the molecule is unchanged but appreciableareas of unmodified substrate 'show through' at various places. It is noticeable thatboth alpha-1-antitrypsin and alpha-2-macroglobulin reduce adhesion at fairly lowlevels of binding and that adhesion of cells decreases as the amount of protein increases,which suggests that the new surface afforded by the protein is inimical to cell adhesion.
In summary, the results suggest that the reduced adhesiveness of bacteriologicalpolystyrene in media containing sera is due, not only to the reduced fibronectinbinding to such surfaces, but also to the binding of relatively high levels of anti-adhesive proteins such as albumin, alpha-2-macroglobulin and alpha-1-antitrypsin.
We thank Michael McGrath for excellent technical assistance, and R.G.G., C.K. and R.D. fordonations of human blood. The work was supported from the General Funds of the University ofGlasgow.
REFERENCES
BROWN, A. F. & LACKIE, J. M. (1981). Fibronectin and collagen inhibit cell-substratum adhesionof neutrophil granulocytes. Expl Cell Res. 136, 225-231.
CURTIS, A. S. G., FORRESTER, J. V., MCINNES, C. & LAWRIE, F. (1983). Adhesion of cells topolystyrene surf aces. J . Cell Biol. 97, 1500-1506.
DIEM, K. & LENTNER, C. (1970). Scientific Tables. Basel: J. R. Geigy Ltd.EDWARDS, J. G. & CAMPBELL, J. A. (1971). The aggregation of trypsinized BHK cells. J . CellSci.
8, 53-72.ENGVALL, E. & RUOSLAHTI, E. (1977). Binding of soluble form of fibroblast surface protein,
fibronectin, to collagen. Int.J. Cancer 20, 1-5FORRESTER, J. V., LACKIE, J. M. & BROWN, A. F. (1983). Neutrophil behaviour in the presence
of protease inhibitors. J. Cell Sci. 59, 213-230.

Adsorbed proteins and cell adhesion 35
FORRESTER, J. V. & WILKINSON, P. C. (1981). Inhibition of leucocyte locomotion by hyaluronicacid.7. Cell Set. to, 315-331.
FORRESTER, J. V., WILKINSON, P. C. & LACKIE, J. M. (1983). Effect of modified macroglobulinon leucocyte locomotion and chemotaxis. Immunology 50, 251-259.
GRINNELL, F. (1978). Cellular adhesiveness and extracellular matrix. Int. Rev. Cytol. 53, 65-144.GRINNELL, F. & FELD, M. K. (1979). Initial adhesion of human fibroblasts in serum-free medium:
possible role of secreted fibronectin. Cell 17, 117-129.GRINNELL, F. & FELD, M. K. (1981). Adsorption characteristics of plasma fibronectin in relation-
ship to biological activity.^, biomed. Mater. Res. 15, 363-381.GRINNELL, F. & FELD, M. K. (1982). Fibronectin adsorption on hydrophilic and hydrophobic
surfaces detected by antibody binding and analysed during cell adhesion in serum-containingmedium. J . biol. Chem. 257, 4888-4893.
HUGHES, R. C , PENA, S. D. J., CLARK, J. & DOURMASHKIN, R. R. (1979). Molecular require-ments for the adhesion and spreading of hamster fibroblasts. Expl Cell Res. 121, 307-314.
HYNES, R. O. (1976). Cell surface protein and malignant transformation. Biochim. biophys. Ada458, 73-107.
KLEBE, R. J., BENTLEY, K. L. & SCHOEN, R. L. (1981). Adhesive substrates for fibronectin. J. cell.Physiol. 109, 481-488.
VAN OSS, C. J., GILLMAN, C. F. & NEUMANN, A. W. (1975). Phagocytic Engulfment and CellAdhesiveness as Cellular Surface Phenomena. New York: Dekker.
WESTON, J. A. & HENDRICKS, K . I . (1972). Reversible transformation by urea of contact-inhibitedfibroblasts. Proc. natn. Acad. Sci. U.SA. 69, 3727-3731.
YAMADA, K. M. & OLDEN, K. (1978). Fibronectin: adhesive glycoproteins of cell surface andblood. Nature, Lond. 275, 179-185.
(Received 2 April J984-Accepted 8 May 1984)