Embed Size (px)
Citation preview

Acc
epte
d A
rtic
le
This article has been accepted for publication and undergone full peer review but has not been through the copyediting, typesetting, pagination and proofreading process which may lead to differences between this version and the Version of Record. Please cite this article as an 'Accepted Article', doi: 10.1111/tpj.12663 This article is protected by copyright. All rights reserved.
Received Date : 04-Jun-2014 Revised Date : 15-Aug-2014 Accepted Date : 29-Aug-2014 Article type : Original Article The cell-wall targeted purple acid phosphatase AtPAP25 is critical for
acclimation of Arabidopsis thaliana to nutritional phosphorus-deprivation
Hernan A. Del Vecchio1, Sheng Ying1, Joonho Park2, Vicki L. Knowles1, Satomi
Kanno3, Keitaro Tanoi3, Yi-Min She4 and William C. Plaxton1,5*
1Department of Biology and 5Department of Biomedical and Molecular Sciences, Queen’s
University, Kingston, Ontario K7L 3N6, Canada
2Department of Fine Chemistry, Seoul National University of Science and Technology, 138
Gongreung-Gil, Nowon-Gu, Seoul 139-743, Korea
3Graduate School of Agricultural and Life Sciences, University of Tokyo, Bunkyo-ku, Tokyo
113-8657, Japan
4Shanghai Center for Plant Stress Biology, Chinese Academy of Sciences, Songjiang,
Shanghai 201602, China
Corresponding author: William C. Plaxton, Department of Biology, Queen’s University,
Kingston, Ontario K7L 3N6, Canada. Phone: 1-613-533-6150; FAX: 1-613-533-6617; E-
mail: [email protected]
Running title: Cell wall PAPs of Pi-deprived Arabidopsis
Key words: Arabidopsis thaliana (thale cress), functional genomics, cell wall proteome,
phosphorus nutrition, purple acid phosphatase.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
SUMMARY
Plant purple acid phosphatases (PAPs) belong to a relatively large gene family whose
individual functions are poorly understood. Three PAP isozymes upregulated in cell walls of
phosphate (Pi) starved (–Pi) Arabidopsis thaliana suspension cells were purified and
identified by MS as AtPAP12 (At2g27190), AtPAP25 (At4g36350), and AtPAP26
(At5g34850). AtPAP12 and AtPAP26 were previously isolated from the culture media of the
–Pi cell cultures and shown to be secreted by roots of Arabidopsis seedlings to facilitate Pi
scavenging from soil-localized organophosphates. AtPAP25 exists as a 55-kDa monomer
containing complex NX(S/T) glycosylation motifs at Asn172, Asn367, and Asn424. Transcript
profiling and immunoblotting with anti-AtPAP25 immune serum indicated that AtPAP25 is
exclusively synthesized under –Pi conditions. Coupled with AtPAP25’s mixed-type inhibition
by Pi (I50 = 50 µM), this indicates a tight feedback control by Pi that would prevent AtPAP25
from being synthesized or functioning as a phosphatase except when Pi levels are quite low.
Promoter:β-glucuronidase reporter assays revealed AtPAP25 expression in shoot vascular
tissue of –Pi plants. Development of an atpap25 T-DNA insertion mutant was arrested
during cultivation on a soluble Pi-deficient soil, but rescued upon Pi fertilization or
complementation with AtPAP25. Transcript profiling by qPCR indicated that Pi starvation
signaling was attenuated in the atpap25 mutant. AtPAP25 exhibited near-optimal
phosphatase activity with several phosphoproteins and phosphoamino acids as substrates.
We hypothesize that AtPAP25 plays a key signaling role during Pi-deprivation by functioning
as a phosphoprotein phosphatase, rather than as a non-specific scavenger of Pi from
extracellular organic-P esters.
INTRODUCTION
Acid phosphatases (APases; EC 3.1.3.2) catalyze the release of orthophosphate (Pi) from a
wide variety of P-monoesters and anhydrides with an acidic pH optimum. They function in
the production and recycling of Pi, a crucial but environmentally limiting macronutrient for
cellular metabolism and bioenergetics (Tran et al., 2010a). Purple APases (PAPs) are the

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
largest class of non-specific plant APase, and have a characteristic pink or purple color in
solution which is caused by the occurrence of a binuclear metal ion complex at their active
site. Upregulation of PAPs is a ubiquitous plant response to Pi deficiency, a frequent abiotic
stress that restricts plant growth in natural ecosystems (Tran et al., 2010a). Intracellular
PAPs recycle Pi from dispensable P-monoesters and anhydrides during Pi stress.
Extracellular PAPs belong to an assortment of Pi starvation inducible (PSI)
phosphohydrolases that cleave Pi from external organic-P molecules, the main form of P in
soil solutions which can account for up to 80% of the soil’s total P content (Tran et al.,
2010a).
Genomic and proteomic methodologies have helped to identify PAP genes that
mediate efficient Pi acquisition and use by vascular plants. Genome annotation identified 29
putative PAP genes in Arabidopsis thaliana, several of which are induced during Pi
deprivation (Tran et al., 2010a). The variation and complexity of AtPAP1-29 expression has
been well documented (Robinson et al., 2012a; Tran et al., 2010a, 2010b; Veljanovski et al.,
2006; Wang et al., 2011; Wang et al., 2014; Zamani et al., 2014; Zhang et al., 2008; Zhu et
al., 2005). To assess the function of specific AtPAP genes in the metabolic adaptations of Pi-
starved (–Pi) Arabidopsis, we have purified, characterized, and identified native intracellular
and secreted AtPAP isozymes upregulated during Pi stress. Cell suspension cultures have
been instrumental in this respect since a large quantity of –Pi cells and the corresponding
liquid media containing secreted enzymes can be readily acquired (Tran et al., 2010b; Tran
and Plaxton, 2008; Veljanovski et al., 2006). Thus, the primary intracellular (vacuolar) PAP
upregulated by –Pi Arabidopsis was isolated and identified as AtPAP26 (At5g34850)
(Veljanovski et al., 2006). Subsequent analyses of an atpap26 loss-of-function mutant
confirmed vacuolar AtPAP26 as the predominant contributor to intracellular APase activity,
and that it plays essential roles to scavenge and remobilize Pi during nutritional Pi
deprivation or leaf senescence (Hurley et al., 2010; Robinson et al., 2012a; 2012b; Shane et
al., 2014). Additional studies revealed that AtPAP26 is also targeted to the cell wall (CW)

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
and secretome of –Pi Arabidopsis (Robinson et al., 2012a). For example, AtPAP26 and the
closely related AtPAP12 (At2g27190) account for about 75% of the total APase activity
secreted by roots of Arabidopsis seedlings cultivated under –Pi conditions, as well as on
various exogenous organic-P sources such as glycerol-3-P or salmon sperm DNA (Robinson
et al., 2012a). AtPAP12 and AtPAP26 were also upregulated on root surfaces and in shoot
CW proteomes of –Pi seedlings (Robinson et al., 2012a; Wang et al., 2014). CW-associated
AtPAP12 and AtPAP26 were hypothesized to facilitate P-status maintenance by recycling Pi
from cytoplasmic P-monoesters that had leaked into CWs during Pi deprivation (Robinson et
al., 2012a). Furthermore, AtPAP10 is a PSI PAP predominantly localized to the surface of
root epidermal cells, and that also functions in the acclimation of Arabidopsis to Pi
deprivation (Wang et al., 2011; 2014).
The CW is a dynamic structure intimately involved with plant growth and
development, responses to abiotic stress, and interactions with pathogens and symbionts.
The complexity and importance of the CW is reflected by the large number of genes
suspected to play a role in its biogenesis, assembly, and modifications (Albenne et al.,
2013). CW proteins represent about 10% of the CW mass, and encompass hundreds of
different proteins that function as CW modifying enzymes, structural proteins, or stress-
inducible defense proteins (Albenne et al., 2013). The aim of the current study was to target
the CW proteome for purification, identification, and eventual biochemical, molecular, and
functional characterization of CW PAPs upregulated by –Pi Arabidopsis suspension cells.
CW-localized AtPAP12 and AtPAP26 were shown to be strongly upregulated, corroborating
previous results obtained with –Pi Arabidopsis seedlings (Robinson et al., 2012a). However,
the previously uncharacterized AtPAP25 was discovered to be exclusively expressed and
targeted to CWs of –Pi Arabidopsis while exhibiting kinetic features consistent with its
possible function as a phosphoprotein phosphatase. Analysis of an atpap25 mutant
indicated that AtPAP25 makes a pivotal contribution to the ability of Arabidopsis to acclimate

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
to Pi deficient soils, and that Pi-starvation signaling appears to have been attenuated in the
mutant.
RESULTS AND DISCUSSION
Influence of Pi-deprivation on cell wall acid phosphatase activity and immunoreactive
PAP polypeptides of Arabidopsis suspension cell cultures
When Arabidopsis suspension cells were subcultured into media containing 5 mM Pi and
harvested 7-d later, the cell culture filtrates (CCFs) contained 1.8 mM Pi, demonstrating that
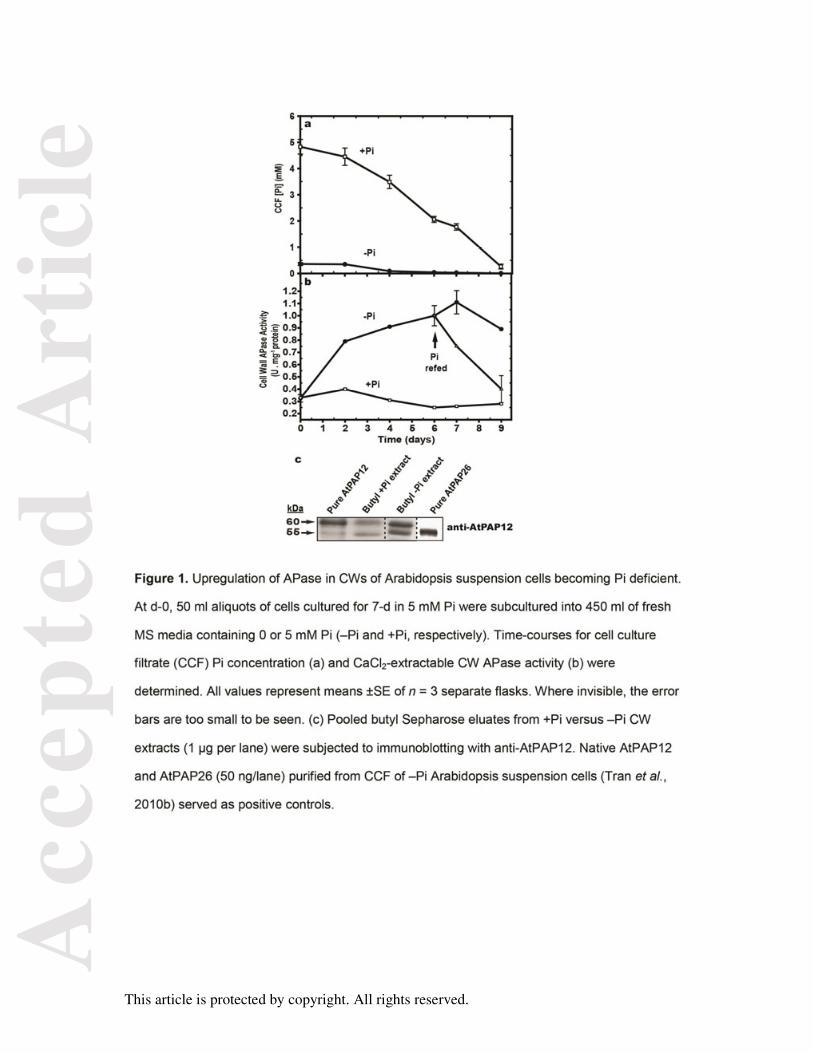
the cells were still Pi-sufficient (+Pi) (Figure 1a). However, extracellular Pi became
undetectable when parallel –Pi cells were cultivated for more than 4-d in media containing 0
mM Pi; this was paralleled by a 4-fold increase in CW APase activity (Figure 1b). Resupply
of the –Pi cells with 2.5 mM Pi at day-6 triggered a steadily decrease in CW APase activity to
+Pi levels over the next several days (Figure 1b). Hydrophobic interaction chromatography
of the CW extracts was performed using butyl Sepharose to enrich CW APase activity. The
resulting samples from the –Pi cells showed increased specific APase (5.11 ±0.09 units mg-
1) relative to those from the +Pi cells (1.26 ±0.03 units mg-1). This was correlated with the
upregulation of 60 and 55 kDa immunoreactive polypeptides on immunoblots of butyl
Sepharose-enriched –Pi CW extracts probed with anti-AtPAP12 immune serum (anti-
PAP12), which respectively co-migrated with AtPAP12 and AtPAP26 isolated from the CCF
of the –Pi cell cultures (Figure 1c). These results agree with earlier work demonstrating that
AtPAP12 and AtPAP26 are also strongly upregulated in shoot CW proteomes and on root
surfaces of –Pi Arabidopsis seedlings (Robinson et al., 2012a).
Previous research established that secreted CCF proteins from the +Pi or –Pi
Arabidopsis cell cultures were not contaminated by cytoplasmic marker enzymes (Tran et
al., 2010b; Tran and Plaxton, 2008). Likewise: (i) immunoreactive cytosolic
phosphoenolpyruvate (PEP) carboxylase and aldolase polypeptides were absent on
immunoblots of the CW extracts, but detected on immunoblots of corresponding clarified

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
extracts prepared from cells that had previously been treated with CaCl2 for CW protein
extraction, and (ii) PEP carboxylase and aldolase activities were detected in the clarified cell
extracts, but not in the corresponding CW proteomes (Figure S1).
Purification and identification of cell wall PAPs of Pi-deprived Arabidopsis
suspension cells
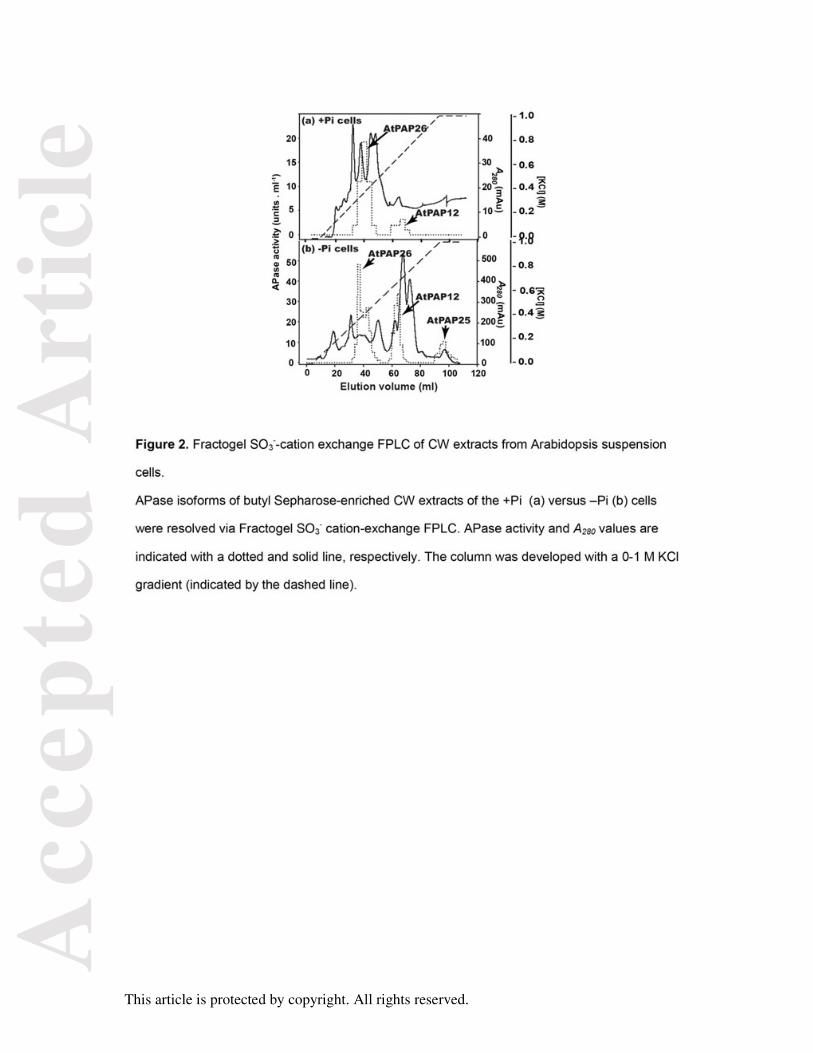
Three peaks of APase activity (corresponding to AtPAP12, AtPAP25, and AtPAP26; see
below) were resolved during SO3--cation exchange fast protein liquid chromatography
(FPLC) of the concentrated butyl Sepharose fraction obtained from the CW proteome of the
–Pi cells (Figure 2b). As reported during AtPAP26 purification from the CCF of the –Pi cells
(Tran et al., 2010b), the pooled AtPAP26 activity fractions from the butyl Sepharose column
resolved as a pair of putative glycoforms (AtPAP26-CW1 and AtPAP26-CW2) during lectin-
affinity chromatography on conA. AtPAP26-CW1 did not bind to the conA column, whereas
AtPAP26-CW2 (and AtPAP12 and AtPAP25) was bound and subsequently eluted by a
methyl-α-D-mannopyranoside gradient (denotes existence of terminal mannose residues).
The proportion of AtPAP26-CW1 to AtPAP26-CW2 activities eluting during conA
chromatography was about 2:1. AtPAP26-CW1’s failure to bind to the conA matrix was not
due to column overloading since immediate application of this sample on a second, freshly
prepared conA column led to quantitative recovery of AtPAP26-CW1 in the resultant
unbound fractions. Additional research is needed to determine glycan structures and
linkages at each N-linked glycosylation site of AtPAP26 glycoforms secreted into the CW
and apoplast by –Pi Arabidopsis. However, our earlier study of a pair of AtPAP26 glycoforms
purified from the CCF of –Pi Arabidopsis cells: (i) employed a glycan differentiation kit that
provided independent evidence that are diffentially glycosylated, and (ii) reported that
differential glycosylation appears to modulate their respective substrate specificities (Tran et
al., 2010b).

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
AtPAP12, AtPAP25, AtPAP26-CW1, and AtPAP26-CW2 were purified to final PEP
hydrolyzing specific activities of 66, 5.5, 252, and 108 units mg-1, respectively (Table S1).
The concentrated final preparations of AtPAP12, AtPAP26-CW1, and AtPAP26-CW2
showed a pink color in solution, while AtPAP25 was purple. The purified PAPs were
subjected to N-terminal sequencing and peptide mass fingerprinting via matrix-assisted laser
desorption/ionization time-of-flight (MALDI-TOF) MS. Comparison with databank sequences
identified them as AtPAP12 (At2g27190), AtPAP25 (At4g36350), and two glycoforms of
AtPAP26 (At5g34850) (Figure 3 and Table S2). AtPAP25 was previously detected in the CW
proteome of Arabidopsis suspension cells (Borderies et al., 2003; Chivasa et al., 2002).
These studies were likely conducted with Pi-limited cells, as the cells were subcultured for
≥7-d in conventional MS media containing 1.25 mM Pi, which is suboptimal for maintaining
Arabidopsis suspension cells fully +Pi during one week in batch culture (Tran and Plaxton,
2008; Veljanovski et al., 2006).
Physical and immunological properties
When the purified CW PAPs were analyzed by SDS-PAGE, major protein-staining
polypeptides of approximately 60 (AtPAP12) and 55 kDa (AtPAP25 and AtPAP26-CW1/-
CW2) were observed, which cross-reacted with anti-AtPAP12 (Figure 4a and 4b). By
contrast, anti-AtPAP25 and anti-AtPAP26 were relatively monospecific for their respective
antigens (Figure 4b). SDS-PAGE followed by protein staining indicated that AtPAP12 and
AtPAP26-CW2 were highly purified, whereas AtPAP25 and AtPAP26-CW1 were purified to
apparent and near homogeneity, respectively (Figure 4a). The native Mr of AtPAP12 was
estimated by analytical gel filtration to be 120 ±10 kDa, whereas that of AtPAP25, AtPAP26-
CW1, and AtPAP26-CW2 were each 55 ±5 kDa (means ± SEM, n = 3 determinations). The
combined results indicate that purified AtPAP12 exists as a homodimer, while AtPAP25 and
AtPAP26-CW1/-CW2 are monomeric. Identical results were obtained with AtPAP12 and a
pair of AtPAP26 glycoforms isolated from the CCF of the –Pi suspension cells (Tran et al.,
2010b).

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Bioinformatic analysis of AtPAP25
The AtPAP family is encoded by 29 genes that were categorized into three discrete groups
following bioinformatics analysis of their deduced amino acid sequences (Tran et al., 2010).
Group Ia-1 is represented by AtPAP5, AtPAP6, AtPAP11, AtPAP19, and AtPAP25, whereas
Group Ia-2 is composed of AtPAP10, AtPAP12, and AtPAP26. AtPAP25 shares 51-71% and
55-60% sequence identity with Group Ia-1 and 1a-2 members of the AtPAP family,
respectively, while its sequence identity with AtPAP25 orthologs from other plants ranges
from 56-68% (Table S3). In silico analysis of the deduced AtPAP25 sequence indicated that
the mature protein has molecular mass of 53 kDa (Table S3). The 2 kDa discrepancy with
the subunit Mr of the purified native AtPAP25 as estimated by SDS–PAGE (Figure 4) is
explained by the post-translational (covalent) incorporation of glycan groups (Table S4).
Signal P incorrectly predicted that the N-terminus of the deduced AtPAP25 polypeptide
contains a 21 amino acid signal peptide. AtPAP25’s actual signal peptide of 29 amino acids
was revealed by comparing the N-terminal sequence obtained with the purified AtPAP25
with its deduced full-length amino acid sequence (Figure 3). It is notable that the signal
peptides of AtPAP12, AtPAP25, and AtPAP26 were all cleaved at an identical site, beginning
after a conserved arginine residue (Figure 3).
AtPAP25 glycosylation motifs
Glycosylation is an essential post-translational modification that can modulate enzyme
stability, localization, solubility, and kinetic properties (Varki, 1993). Secreted plant proteins
usually contain highly heterogeneous glycan pools. Although all characterized plant PAPs
are glycosylated (Olczak et al., 2003; Olczak and Olczak, 2007; Tran et al., 2010a), few
details of their N-linked glycans are available. The majority of the glycan residues of plant
PAPs studied to date consist of complex oligosaccharides containing mannosylated,
fucosylated and/or xylosylated residues (Olczak et al., 2003; Stahl et al., 1994). Plant PAPs
having 55-kDa subunit sizes usually have three to five N-glycosylation sites and
approximately 10% sugar content (Olczak et al., 2003; Olczak and Olczak, 2007; Stahl et al.,

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
1994). N-linked oligosaccharides of purified AtPAP25 were characterized by high-resolution
ultra-performance liquid chromatography (UPLC) separation coupled with linear ion-trap
Fourier transform ion cyclotron resonance tandem MS (LTQ-FT MS/MS) analysis of
glycopeptides generated by either specific trypsin or nonspecific proteinase K digestion. The
three putative N-glycosylation sites of AtPAP25 (Asn172, Asn367, and Asn424; Figure 3)
possess the consensus NX(S/T) glycosylation motif (where X is any amino acid except
proline), were each determined to contain two compositions of xylomannosides;
GlcNAc(Fuc)GlcNAc(Xyl)Man3 and GlcNAc(Fuc)GlcNAc(Xyl)Man3GlcNAc (Supplemental
Table S4), but no high-mannose-type glycan. The Asn424 glycosylation site was further
modified by the addition of a terminal galactose residue to form an extended side chain of
GlcNAc(Fuc)GlcNAc(Xyl)Man3GlcNAcGal (Table S4). A site-directed mutagenesis study of
Phaseolus vulgaris (common bean) and Lupin luteus (European white lupin) PAP isozymes
(orthologous to AtPAP12 and AtPAP26, respectively) was employed to generate various
PAP mutants lacking N-glycosylation at specific asparagine residues (Olczak and Olczak,
2007). The results established that glycosylation, particularly at the conserved glycosylation
site located near the C-terminus of all known plant PAPs (e.g., corresponding to Asn424 of
AtPAP25) is an essential prerequisite for PAP stability and secretion.”
A study of the phosphotyrosylated proteome of Arabidopsis cell cultures reported that
AtPAP25 is phosphorylated at Tyr423 (Sugiyama et al., 2008). However, MS/MS analysis of
AtPAP25 failed to detect evidence for AtPAP25 phosphorylation at any site, including Tyr423
(Figure S2). Phosphorylation of AtPAP25 at Tyr423 is improbable given the bulky N-linked
oligosaccharide that is covalently bonded to the adjacent Asn424 residue (Figure 3, Table
S4).
AtPAP25 kinetic properties
Similar to other plant PAPs, AtPAP25 displayed a wide pH-activity profile with a maximum at
pH 5.6 (Figure S3), and hyperbolic PEP saturation kinetics (Km(PEP) = 0.42 mM). AtPAP25

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
showed a low Vmax with PEP (5.5 units mg-1) as well as other in vitro substrates that were
identified, relative to values ranging from about 60 - 1700 units mg-1 obtained with AtPAP12
or AtPAP26 isoforms isolated from CW or CCF of the –Pi Arabidopsis suspension cells
(Tables 1 and S1) (Tran et al., 2010b). Although AtPAP25 hydrolyzed Pi from a broad variety
of P-esters and anhydrides, it exhibited a substrate selectivity consistent with its possible
function as a phosphoprotein phosphatase (Table 1). Comparable activities were obtained
with saturating levels of the phosphoamino acids P-Thr, P-Ser, and P-Tyr, as well as with the
storage phosphoproteins ovalbumin and phosvitin (Table 1), unlike AtPAP12 or AtPAP26
(Tran et al., 2010b; Veljanovski et al., 2006). However, AtPAP25 failed to use 5 mM phytate
as a substrate which demonstrates that in contrast to AtPAP15 (Zhang et al., 2008), it is not
a phytase.
AtPAP25 activity was unaffected by the addition of MgCl2, CoCl2, MnCl2, or EDTA (5
mM each). As with other PAPs, the APase activity of AtPAP25 was insensitive to 5 mM L-
tartrate, but potently inhibited by vanadate, arsenate, molybdate, Zn2+, Fe2+ and Cu2+ (Table
S5). Apart from its low Vmax, another unusual kinetic feature of AtPAP25 was its strong
feedback inhibition by Pi as reflected by a low I50(Pi) value of 50 ±6 µM (determined with 0.5
mM PEP). Interestingly, Pi functions as a mixed-type inhibitor since increasing
concentrations of Pi significantly decreased and increased AtPAP25’s Vmax and Km(PEP)
values, respectively (Figure S4). The enzyme’s competitive and uncompetitive dissociation
constants for Pi were determined to be 0.43 ±0.04 and 0.74 ±0.05 mM, respectively. By
contrast, Pi typically serves as a competitive inhibitor of plant PAPs (i.e., increases Km,
without influencing Vmax) (Tran et al., 2010a).
Influence of P-status on AtPAP25 gene and protein expression
Transcripts of AtPAP25 were detected by RT-PCR in –Pi, but not +Pi, Arabidopsis
suspension cells (Figure 5a). This effect was quantified by qPCR, which revealed striking
induction of AtPAP25 transcripts in roots and particularly shoots of –Pi seedlings (Figure 5b).

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Similarly: (i) comparison of APase activity elution profiles during cation-exchange FPLC of
CW extracts from the +Pi versus –Pi cells indicated that AtPAP25 was de novo synthesized
in response to Pi deprivation, whereas AtPAP12 and AtPAP26 were markedly upregulated
(Figure 2), and (ii) immunoblot analysis using anti-AtPAP25 indicated that unlike AtPAP12
and AtPAP26 (Figure 1C and Figure 2), AtPAP25 polypeptides were exclusively expressed
in CWs of the –Pi cells (Figure 5c). In contrast, immunoreactive AtPAP25 polypeptides were
absent on immunoblots of CW or total protein extracts of shoots or roots of –Pi Col-0
seedlings (using chromogenic or chemiluminscent detection). Likewise, immunoreactive
AtPAP25 polypeptides were not apparent on immunoblots of flower extracts from the Col-0
seedlings. This suggests that AtPAP25 polypeptide expression is very low in planta, relative
to other PSI secreted PAPs such as AtPAP12 and AtPAP26 (Robinson et al., 2012a; Tran et
al., 2010b).
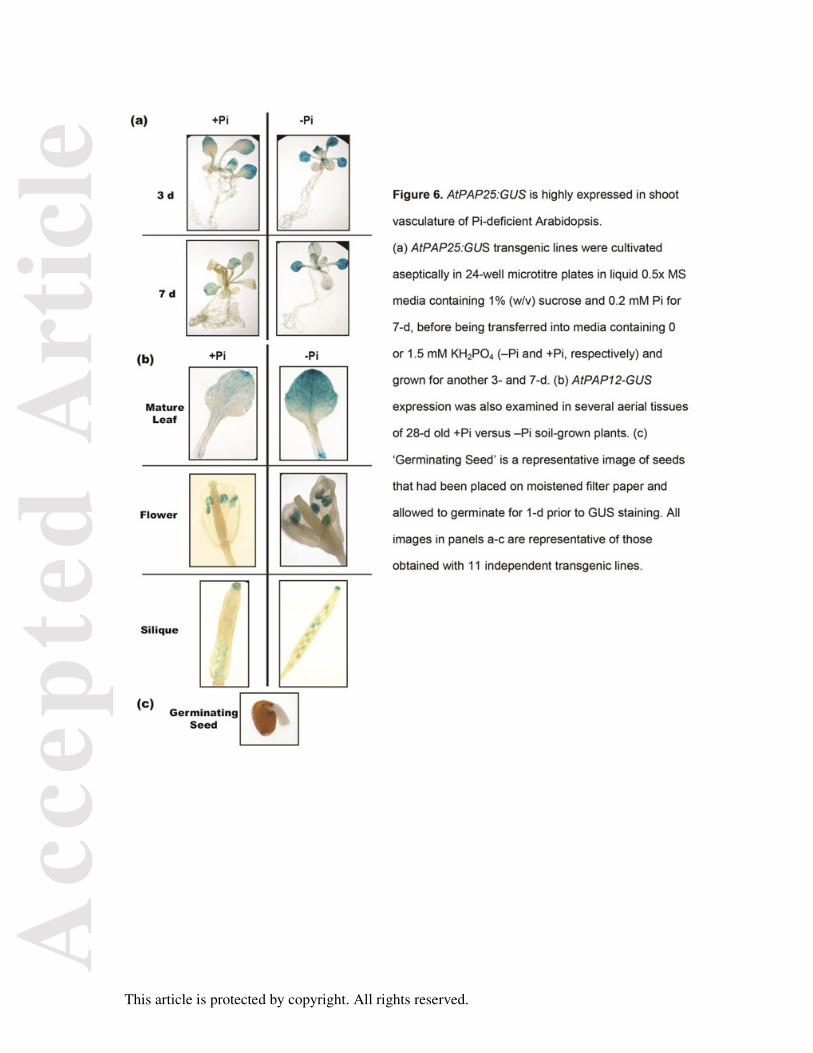
To assess the tissue specificity of AtPAP25 expression, an AtPAP25 promoter: β-
glucuronidase (GUS) reporter gene fusion was generated. The expression of GUS activity
was examined in eleven independent AtPAP25:GUS transgenic lines, all of which indicated
strong expression of the AtPAP25 promoter in shoot vascular tissue during vegetative
growth (particularly during –Pi growth), as well as in anthers and developing embryos of
flowering seedlings (Figure 6). Flower specific-expression of AtPAP25 was previously
demonstrated in Arabidopsis, with AtPAP25 transcripts being particularly abundant in
stamens (Zhu et al., 2005).
Characterization of an atpap25 loss-of-function mutant
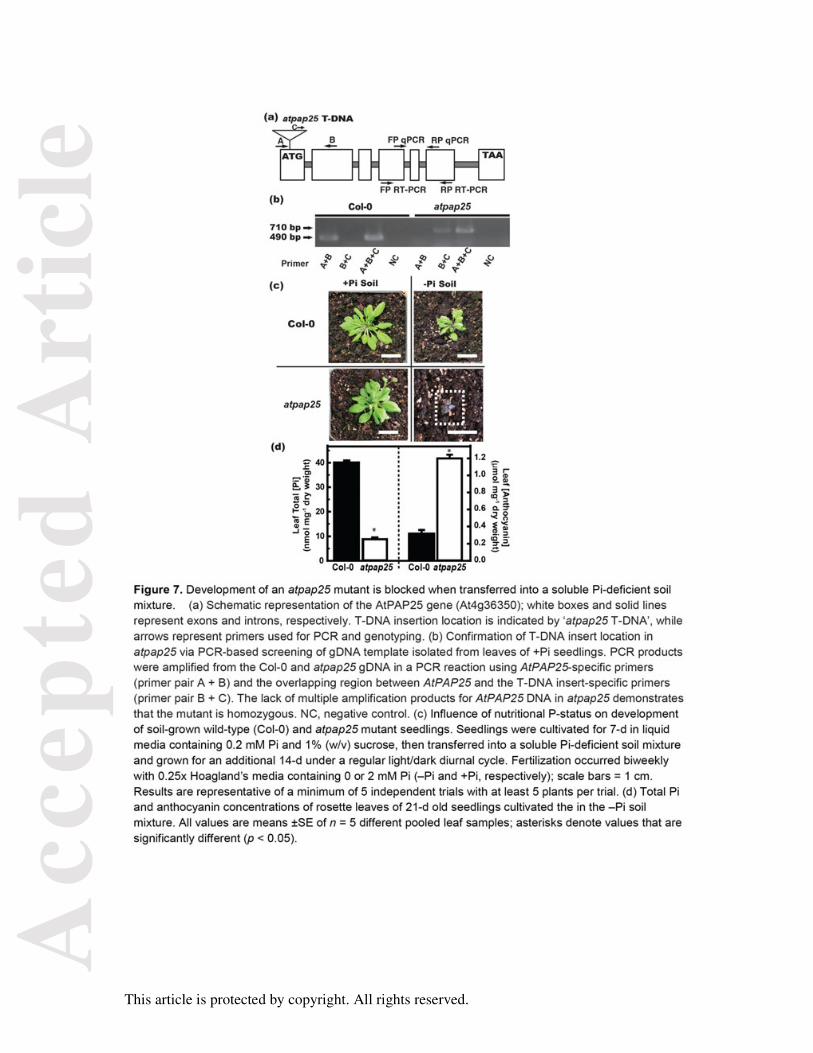
An atpap25 mutant was obtained in which a T-DNA insertion was predicted to occur in the
first exon of the AtPAP25 gene (locus At4g36350). This was confirmed by PCR screening of
gDNA using an AtPAP25-specific primer and a T-DNA right border primer (Figure 7a and
7b). Mutant homozygosity was established by PCR of gDNA using AtPAP25-specific primers
(Figure 7b). Elimination of AtPAP25 expression was confirmed by RT-PCR of cDNA

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
prepared from shoots of –Pi atpap25 plants (Figure 8b). Total shoot CW APase activity of –
Pi atpap25 mutant seedlings was not significantly different from that of the Col-0 controls
(0.27 ±0.05 and 0.30 ±0.04 units mg-1, respectively). Histochemical staining of APase activity
on root surfaces of –Pi atpap25 and Col-0 seedlings using 5-Br-4-Cl-3-indoyl-P (which
serves as an effective substrate for purified AtPAP25; Table 2) was also indistinguishable
(Figure S5b). These results further indicate that AtPAP25 polypeptide levels are very low in
–Pi Col-0 plants.
No obvious differences were noted relative to Col-0 controls in the development or
appearance of atpap25 plants when 7-d hydroponically-cultivated +Pi seedlings were
transferred to sterile –Pi liquid or agar-solidified MS media, or a Pi-fertilized soil mixture
(Figures S5, 7c, 8a, and 8c). Although Pi deprivation stimulated 33Pi uptake capacity of
hydroponically-cultivated Col-0 and atpap25 seedlings, no differences in the capacity to
translocate 33Pi from roots to the shoots were apparent (Figure S6). Similarly, no phenotypic
differences relative to Col-0 were revealed when +Pi atpap25 seedlings were cultivated on
nitrogen, sulfur, or potassium deficient sois. However, atpap25 development was blocked
when 7-d old +Pi seedlings were transplanted into an organic-P containing soil mix lacking
soluble Pi and cultivated for an additional 14-d under a regular light:dark regime (Figure 7c).
Their leaves turned dark purple due to marked anthocyanin accumulation, and total leaf Pi
concentration was over 4-fold lower than that of –Pi Col-0 controls (Figure 7d). The arrested
development of the 21-d-old soil-cultivated –Pi atpap25 plants was rapidly reversed when
they were fertilized with 2 mM Pi and cultivated for an additional 14-d (Figure 8A). Likewise,
complementation with AtPAP25 resulted in normal growth of the resulting transgenic plants
on the +Pi or –Pi soil mix (Figure 8b and 8c). qPCR also revealed that AtPAP25 expression
was undetectable in leaves of –Pi atpap25 plants, but was massively induced (by 840 ±60%)
in leaves of soil-cultivated –Pi Col-0 plants, relative to +Pi controls. This corroborates qPCR
results obtained with Col-0 seedlings cultured in sterile MS liquid media (Figure 5B).

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
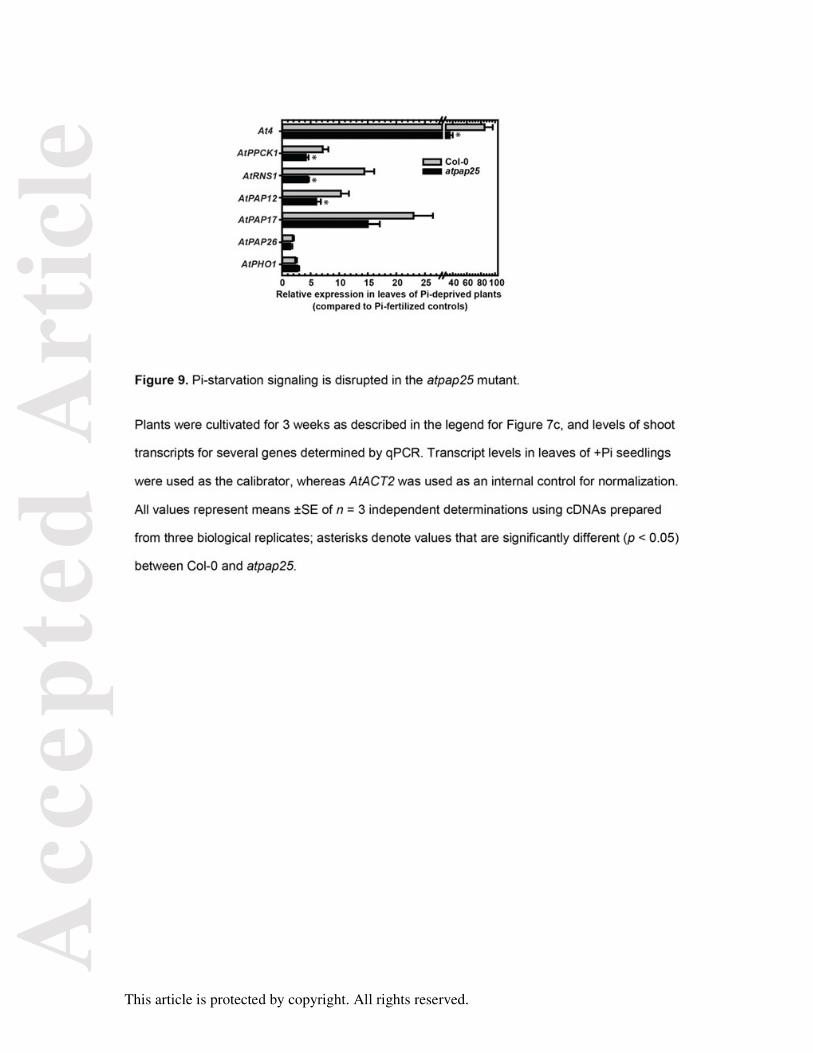
Pi-starvation signaling is attenuated in the atpap25 mutant
Expression of AtPAP12 and AtPAP17 as well as several additional, well-documented PSI
genes were also evaluated by qPCR to determine if Pi-starvation signaling was affected in
the soil-cultivated atpap25 mutant. At4 is a strongly PSI gene in vascular tissue of
Arabidopsis root and shoots that is believed to function in Pi distribution between shoots and
roots during Pi stress (Ha and Tran, 2014; Shin et al., 2006). AtPPCK1 encodes a cytosolic
protein kinase that phosphorylates and activates phosphoenolpyruvate carboxylase in –Pi
Arabidopsis (Gregory et al., 2009), whereas AtRNS1 encodes a vacuolar and secreted (cell
wall, secretome) ribonuclease involved in Pi-recycling from expendable RNA pools (Ha and
Tran, 2014; Tran and Plaxton, 2008). Expression of At4, AtPPCK1, AtRNS1, AtPAP12, and
AtPAP17 was significantly reduced (by 35-70%) in atpap25 shoots relative to Col-0 controls,
whereas expression of AtPAP26 and AtPHO1 was not significantly affected (Figure 9).
AtPAP26 and AtPHO1 represent genes known to play essential roles in Arabidopsis Pi-
starvation responses, but whose transcripts are weakly responsive to Pi deprivation stress
(Ha and Tran, 2014; Tran et al., 2010b; Veljanovski et al., 2006).
CONCLUDING REMARKS
The current results demonstrate that AtPAP25, AtPAP12, and AtPAP26 are the predominant
AtPAP isozymes upregulated and targeted to the extracellular matrix of –Pi Arabidopsis
suspension cells. In contrast to AtPAP12 and AtPAP26 (Hurley et al., 2010; Robinson et al.,
2012a; Robinson et al., 2012b; Tran et al., 2010b; Veljanovski et al., 2006; Wang et al.,
2014), we are unaware of earlier reports describing the biochemical or functional properties
of AtPAP25, nor its involvement in the Arabidopsis Pi-starvation response. It is remarkable
that the transit peptides of AtPAP25, as well as AtPAP12 and its soybean (Glycine max)
ortholog, secreted and vacuolar AtPAP26 glycoforms, and AtPAP26 orthologs purified from
onion (Allium cepa) bulbs and –Pi tomato (Lycopersicon esculentum) cell cultures are all
cleaved at the same site, beginning after an invariant arginine residue (Figure 3) (Tran et al.,
2010a). Future studies are needed to identify the signal peptidase that hydrolyzes the transit

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
peptide from precursors of vacuolar and secreted PAP isoforms during their subcellular
sorting in –Pi plant cells. AtPAP25’s restricted CW localization may be the result of
glycosylation-driven targeting, and its putative signaling role might also rely on this, adding
more complexity to its in vivo function. LTQ-FT MS/MS analysis of AtPAP25 revealed a
complex array of glycans, N-linked to three Asn residues possessing the conserved NX(S/T)
glycosylation motif. The importance of glycosylation in controlling enzyme localization,
turnover, activity, and association with binding partners may transcend PAPs and Pi
starvation. The involvement of both the individual protein and its cellular environment in
determining glycosylation site occupancy and glycan processing allows cells to generate an
assortment of glycoforms, thus endowing a single glycosylated enzyme with an assortment
of functional and dynamic activities (Varki, 1993).
GUS reporter assays indicated that AtPAP25 is specifically and strongly expressed in
shoot vascular tissue during vegetative growth of –Pi plants, as well as in stamens and
developing embryos of the corresponding flowering plants. Similar results were obtained with
AtPAP10:GUS seedlings, although the APase activity of AtPAP10 appears to be
predominantly associated with root surfaces of –Pi Arabidopsis (Wang et al., 2011). By
contrast, –Pi AtPAP12:GUS and AtPAP26:GUS plants showed widespread GUS activity
(Robinson et al., 2012a). Another uncommon feature of AtPAP25 was that it appeared to be
exclusively expressed and synthesized in response to Pi deprivation, but at low levels
compared to other PSI PAPs such AtPAP12 and AtPAP26 (Robinson et al., 2012a; Tran et
al., 2010b). Coupled with AtPAP25’s mixed-inhibition by Pi, this indicates a tight feedback
control by Pi that would prevent AtPAP25 from being synthesized or functioning as a
phosphatase except when Pi levels are very low.
Unusual kinetic properties of AtPAP25 also included its relatively low Vmax, and a
substrate selectivity suggesting that it might function as a phosphoprotein phosphatase
rather than as a non-specific scavenger of Pi from extracellular organic-P compounds, as

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
demonstrated for AtPAP10, AtPAP12, and AtPAP26 (Robinson et al., 2012a; Tran et al.,
2010b; Wang et al., 2011; Shane et al., 2014; Wang et al., 2014). Similarly, NtPAP12 is a
CW PAP of +Pi tobacco cells orthologous to AtPAP10 and that appears to function as a
phosphoprotein phosphatase that participates in the control of cellulose metabolism (Kaida
et al., 2009, 2010). Extracellular protein phosphorylation is widespread in eukaryotic cells. It
will thus be important to determine the in vivo substrate(s) for AtPAP25, and whether
AtPAP25 contributes to reprogramming of the CW phosphoproteome upon nutritional Pi-
deficiency.
AtPAP25 clearly plays an important role in the Arabidopsis Pi-starvation response as
the atpap25 mutant demonstrated a dramatic arrested growth phenotype when 7-d old +Pi
seedlings were cultivated for two weeks in a peat-vermiculite soil mixture lacking soluble Pi
(Figures 7 and 8). Quantification of transcripts encoding five PSI genes indicated that Pi
starvation signaling was attenuated in shoots of the soil-cultivated atpap25 mutant (Figure
9). It is notable, however, that AtPAP26 and AtPHO1 transcript levels were unaffected in the
mutant. Although AtPAP26 and AtPHO1 play essential roles in Arabidopsis Pi starvation
responses, levels of their respective transcripts are largely unresponsive to Pi deprivation
(Ha and Tran, 2014; Hurley et al., 2010; Robinson et al., 2010a; Tran et al., 2010b;
Veljanovski et al., 2006). This suggests that the reduced expression of At4, AtPPCK1,
AtRNS1, AtPAP12, and AtPAP17 observed in the –Pi atpap25 mutant was not due to a
general restriction in cellular function, as opposed to a specific disruption of the Pi starvation
response. Additional studies are needed to test our hypothesis that AtPAP25 functions as a
phosphoprotein phosphatase in CWs of shoot vascular tissue of –Pi plants, and that this
somehow participates in the overall signaling pathway by which Arabidopsis responds to
nutritional P deprivation at the molecular level. In this regard, microarray transcriptome and
(phospho)proteomic profiling to assess global gene and (phospho)protein expression
changes in atpap25 plants would help to further establish the degree to which Pi-starvation
signaling has been impaired in the mutant. Overall, these studies are relevant to current

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
applied efforts to bioengineer P-efficient crops, urgently required to reduce the overuse of
non-renewable and polluting Pi-containing fertilizers in agriculture.
EXPERIMENTAL PROCEDURES
Plant material and growth conditions
Heterotrophic Arabidopsis (Arabidopsis thaliana, cv. Landsberg erecta) suspension cells
were maintained at 21 oC in the dark as previously described (Tran et al., 2010b; Veljanovski
et al., 2006). For large-scale –Pi subculture, 50 ml aliquots of 7-d Pi-replete (+Pi; 5 mM
K2HPO4) cultures were used to inoculate 2.6 l Fernbach flasks containing 450 ml of fresh MS
media lacking Pi. Cells were collected after 7-d by filtration, frozen in liquid N2, and stored at
-80 °C.
For routine plant growth and mutant isolation, Arabidopsis (Col-0 ecotype) seeds
were sown in a standard soil mixture (Sunshine Aggregate Plus Mix 1; SunGro, Vancouver,
Canada) and stratified at 4 oC for 3-d. Plants were cultivated in growth chambers at 23 °C
(16/8 h photoperiod at 100 µmol m-2 s-1 PAR) and fertilized biweekly by subirrigation with
0.25x Hoagland’s media. To assess the influence of Pi deprivation on soil-grown plants,
seedlings were established for 7-d in a 24-well microtitre plate (1 seedling per well)
containing 0.5 ml of 0.5x MS media supplemented with 1% (w/v) sucrose and 0.2 mM Pi,
then transplanted into a 75-85% sphagnum peat moss/perlite soil mix lacking all nutrients
(Sunshine Mix 2; SunGro). Plants were grown for an additional 14-d and fertilized biweekly
with 0.25x Hoagland’s media (pH 6.0) containing either 0 or 2 mM KH2PO4. Whenever Pi
was eliminated, it was replaced by 2 mM KH2SO4 and 0.5 mM MES. For liquid cultures, 5 mg
of seeds were surface sterilized, stratified, placed in 250 ml Magenta boxes containing 50 ml
of 0.5x MS media, pH 5.7, 1% (w/v) sucrose and 0.2 mM KH2PO4, and placed on an orbital
shaker (80 rpm) at 24 oC under continuous illumination (100 µmol m-2 s-1). After 7-d the
medium was replaced with fresh media containing 0 or 1.5 mM filter-sterilized KH2PO4. The
14-d seedlings were blotted dry, snap frozen in liquid N2, and stored at -80 oC. Stratified

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
seeds were also placed on vertically oriented 1% (w/v) agar (Micropropagation Type I Agar
from Caisson Labs, Cat. #A038) plates containing 0.5x MS media and 1% (w/v) sucrose
supplemented with 50 µM or 1.5 mM KH2PO4, or 1.5 mM DL-glycerol-3-P (Sigma Chemical
Co.) and cultivated for up to 21-d as described for the liquid grown seedlings.
Enzyme activity assays and kinetic studies
All enzyme assays were linear with respect to time and concentration of enzyme assayed,
and optimized with respect to pH and substrate concentration. One unit of activity is defined
as the amount resulting in the use of 1 µmol min-1 of substrate at 25 °C. Aldolase and PEP
carboxylase activities were determined using coupled spectrophotometric assays (Tran and
Plaxton, 2008). For routine measurements of APase activity, the hydrolysis of PEP to
pyruvate was coupled to the lactate dehydrogenase reaction and assayed at 25 °C by
monitoring the oxidation of NADH at 340 nm using a Molecular Devices Spectromax Plus
Microplate spectrophotometer. Standard APase assay conditions were: 50 mM Na-acetate
(pH 5.6) containing 5 mM PEP, 0.2 mM NADH, and 3 units of rabbit muscle lactate
dehydrogenase (final volume = 0.2 ml). Assays were corrected for any background NADH
oxidation by omitting PEP from the assay mixture. Substrate selectivity studies were
performed by quantifying the Pi released by the APase reaction as previously described
(Veljanovski et al., 2006). AtPAP25’s: (i) apparent Km(PEP) and I50(Pi) values were
calculated using a non-linear regression enzyme kinetics program (Brooks, 1994), and (ii)
competitive and uncompetitive inhibition constants for Pi were determined using the
graphical method described by Cornish-Bowden (1974). All kinetic parameters are the
means of at least three independent experiments and are reproducible to within ±10% of the
mean value.
Protein electrophoresis and immunoblotting
SDS-PAGE, subunit Mr estimation via SDS–PAGE, immunoblotting onto poly(vinylidene)
difluoride (PVDF) membranes, imaging of antigenic polypeptides using an alkaline

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
phosphatase-tagged secondary antibody, and chromogenic detection were carried out as
previously described (Veljanovski et al., 2006). All immunoblot results were replicated at
least three times with representative results shown in the figures.
RT-PCR and qPCR analysis
Total RNA was extracted and purified as described previously (Gregory et al., 2009;
Robinson et al., 2012a; Tran et al., 2010b). RNA samples were evaluated for purity via their
A260/A280 ratio and integrity by resolving 1 µg of total RNA on a 1% (w/v) denaturing agarose
gel. RNA (5 µg) was reverse transcribed with Superscript III (Invitrogen, Carlsbad, CA,
USA), and noncompetitive RT-PCR accomplished using gene specific primers listed in Table
S6 or as previously described (Gregory et al., 2009; Hurley et al., 2010). Conditions were
optimized for all RT-PCR reactions to ensure linearity of response for comparison between
samples. All PCR products were verified by sequencing.
Quantitative real-time PCR (qPCR) was performed as previously described
(Robinson et al., 2012b) using gene-specific primers listed in Table S6. Relative expression
was determined by normalizing transcript abundance with AtActin2 (NM_112764) transcripts.
Relative expression (fold-change) was calculated using the 2–∆∆Ct method as previously
described (Robinson et al., 2012b). All qPCR experiments were repeated at least three times
using cDNA prepared from three independent samples.
GUS analysis
The AtPAP25 promoter (982 bp sequence upstream of the start codon of the AtPAP25 gene)
was amplified from genomic DNA using primers listed in Table S6. Amplified promoter region
of AtPAP25 was mixed in a 3:1 molar ratio with pBI101 N1 linearized by HindIII and XbaI,
incubated with In-Fusion reaction mix, and transformed according to manufacturer’s
(Clontech) protocol to yield AtPAP25:GUS. The construct was transferred into
Agrobacterium tumefaciens strain LBA4404 and transformed into Arabidopsis plants via the

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
floral dip method as previously described (Robinson et al., 2012a). Transformed plants were
selected on 0.8% (w/v) agar plates containing 0.5x MS media, 1% (w/v) sucrose, and 30 µg
ml-1 kanamycin, and transferred to soil for self-pollination and propagation. For analysis of
mature plants, seeds were planted in soil and grown for 28-d while being fertilized bi-weekly
with 0.25x Hoagland’s media containing 0 or 2 mM Pi. Histochemical staining of GUS
activity was performed as previously described (Robinson et al., 2012a). Tissues were
incubated at 37 °C overnight in GUS staining buffer (100 mM NaPi, pH 7.0, containing 1 mM
X-Gluc, 0.5 mM EDTA, 0.1% (v/v) Triton X-100, 2 mM K3Fe(CN)6, and 2 mM K4Fe (CN)6).
Stained tissues were cleared with 70% ethanol before imaging with a dissecting microscope.
Mutant isolation and complementation
A potential atpap25 mutant line (GK-004D10-014797) was obtained from The European
Arabidopsis Stock Centre (NASC). Initial screening was performed by growing plants on
0.8% (w/v) agar plates containing 0.5x MS media, 1% (w/v) sucrose, and 5 µg ml-1
sulfadiazine. Resistant plants were subjected to a second round of selection with
sulfadiazine and then transferred to soil for self-pollination and propagation. gDNA was
extracted from leaves and PCR-screened for homozygous mutants using T-DNA left-border
and gene specific primers listed in Table S6.
For complementing the atpap25 mutant phenotype, AtPAP25 was amplified by using
primers (Table S6) designed to introduce BglII and PmlI restriction enzyme sites at the 5’
and 3’ ends of its coding region, respectively. AtPAP25 was cloned into the pCAMBIA3301
(CAMBIA, Canberra, Australia) vector, thus allowing the gene to be driven by the CaMV 35S
promoter. The constructs were introduced into Agrobacterium strain GV3101 and
transformed into the atpap25 knock-out mutant using the floral dip method as previously
described (Robinson et al., 2012a). Seeds of the T0 generation were harvested and sown in
soil, and two-week-old seedlings of T1 plants screened by spraying with 0.5% (v/v)
phosphinothricin (PhytoTechnology Laboratories, USA). The surviving transformants (T1)

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
were confirmed by PCR to amplify AtPAP25 and Bar genes (primers listed in Table S6). T2
seeds were placed on agar-solidified 0.5x MS media containing 7 mg l-1 phosphinothricin
and transgenic lines with a 3:1 (resistant:sensitive) segregation ratio were selected to
produce T3 seeds. The T3 lines displaying 100% phosphinothricin resistance (due to
expression of Bar gene) were considered homozygous and used for further experiments.
ADDITIONAL EXPERIMENTAL PROCEDURES
Additional experimental procedures are described in Methods S1.
ACKNOWLEDGEMENTS
We are grateful to Prof. Tom McKnight (Texas A & M University) for the gift of anti-
(recombinant AtPAP12) immune serum. Financial support was generously provided by
grants from The Natural Sciences and Engineering Research Council of Canada (NSERC)
and Queen’s Research Chairs program to WCP.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the online version of this article.
Figure S1. Immunological detection of PEP carboxylase and cytosolic aldolase in clarified
extracts from Arabidopsis suspension cells.
Figure S2. AtPAP25 is not phosphorylated at Tyr423.
Figure S3. AtPAP25 pH-activity profile.
Figure S4. Influence of Pi on PEP saturation kinetics of AtPAP25.
Figure S5. Comparison of development of Col-0 and atpap25 seedlings cultivated in liquid
or agar-solidified MS media.
Figure S6. Imaging of 33P-accumulation by shoots of Col-0 and atpap25 seedlings.
Table S1. Purification of native cell wall PAPs from 400 g of 7-d-old –Pi Arabidopsis
suspension cells

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Table S2. MALDI-TOF MS analysis of tryptic peptides derived from native AtPAP isoforms
isolated from cell wall extracts of –Pi Arabidopsis.
Table S3. Comparison of the amino acid sequence identity of AtPAP25 with AtPAP12 and
AtPAP26, as well as with AtPAP25 orthologs from other vascular plant sources.
Table S4. Identification of AtPAP25 glycopeptides by UPLC LTQ-FT LC MS/MS.
Table S5. Effect of various substances on the activity of purified AtPAP25.
Table S6. Primers used for cloning and PCR analysis.
Methods S1. Additional experimental procedures.
This material is available as part of the online article from http://www.blackwell-synergy.com
REFERENCES
Albenne, C., Canut, H. and Jamet, E. (2013) Plant cell wall proteomics: the leadership of
Arabidopsis thaliana. Front. Plant Sci. 4, 111.
Borderies, G., Jamet, E., Lafitte, C., Rossignol, M., Jauneau, A., Boudart, G.,
Monsarrat, B., Esquerre-Tugaye, M.T., Boudet, A. and Pont-Lezica, R. (2003)
Proteomics of loosely bound cell wall proteins of Arabidopsis thaliana cell suspension
cultures: a critical analysis. Electrophoresis 24, 3421-3432.
Brooks, S.P.G. (1992) A simple computer program with statistical tests for the analysis of
enzyme kinetics. Biotechniques 13, 906-911.
Chivasa, S., Ndimba, B.K., Simon, W.J., Robertson, D., Yu, X.L., Knox, J.P., Bolwell, P.
and Slabas, A.R. (2002) Proteomic analysis of the Arabidopsis thaliana cell wall.
Electrophoresis 23, 1754-1765.
Cornish-Bowden, A. (1974) A simple graphical method for determining the inhibition
constants of mixed, uncompetitive and non-competitive inhibitors. Biochem. J. 137, 143-
144.
Gregory, A.L., Hurley, B.A., Tran, H.T., Valentine, A.J., She, Y.M., Knowles, V.L. and
Plaxton, W.C. (2009) In vivo regulatory phosphorylation of the phosphoenolpyruvate

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
carboxylase AtPPC1 in phosphate-starved Arabidopsis thaliana. Biochem. J. 420, 57-
65.
Ha, S. and Tran, L. (2014) Understanding plant responses to phosphorus starvation for
improvement of plant tolerance to phosphorus deficiency by biotechnological
approaches. Crit. Rev. Biotechnol. 34, 16-30.
Hurley, B.A., Tran, H.T., Marty, N.J., Park, J., Snedden, W.A., Mullen, R.T. and Plaxton,
W.C. (2010) The dual-targeted purple acid phosphatase AtPAP26 is essential for
efficient acclimation of Arabidopsis thaliana to nutritional phosphate deprivation. Plant
Physiol. 153, 1112-1122.
Kaida, R., Satoh, Y., Bulone, V., Yamada, Y., Kaku, T., Hayashi, T. and Kaneko, T.S.
(2009) Activation of β-glucan synthases by wall-bound purple acid phosphatase in
tobacco cells. Plant Physiol. 150, 1822-1830.
Kaida, R., Serada, S., Norioka, N., Norioka, S., Neumetzler, L., Pauly, M., Sampedro, J.,
Zarra, I., Hayashi, T. and Kaneko, T.S. (2010) Potential role for purple acid
phosphatase in the dephosphorylation of wall proteins in tobacco cells. Plant Physiol.
153, 603-610.
Olczak, M., Morawiecka, B. and Watorek, W. (2003) Plant purple acid phosphatases -
genes, structures and biological function. Acta Biochim. Pol. 50, 1245-1256.
Olczak, M. and Olczak, T. (2007) N-glycosylation sites of plant purple acid phosphatases
important for protein expression and secretion in insect cells. Arch. Biochem. Biophys.
461, 247-254.
Robinson, W.D., Park, J., Tran, H.T., Del Vecchio, H.A., Ying, S., Zins, J.L., Patel, K.,
McKnight, T.D. and Plaxton, W.C. (2012a) The secreted purple acid phosphatase
isozymes AtPAP12 and AtPAP26 play a pivotal role in extracellular phosphate-
scavenging by Arabidopsis thaliana. J. Exp. Bot. 63, 6531-6542.
Robinson, W.D., Carson, I., Ying, S., Ellis, K. and Plaxton, W.C. (2012b) Eliminating the
purple acid phosphatase AtPAP26 in Arabidopsis thaliana delays leaf senescence and
impairs phosphorus remobilization. New Phytol. 196, 1024-1029.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Shane, M.W., Stigter, K., Fedosejevs, E.T. and Plaxton, W.C. (2014) Senescence-
inducible cell-wall and intracellular purple acid phosphatases: implications for
phosphorus remobilization in Hakea prostrata (Proteaceae) and Arabidopsis thaliana
(Brassicaceae). J. Exp. Bot. (in press).
Shin, H., Shin, H.S., Chen, R. and Harrison, M.J. (2006) Loss of At4 function impacts
phosphate distribution between the roots and the shoots during phosphate starvation.
Plant J. 45, 712-726.
Stahl, B., Klabunde, T., Witzel, H., Krebs, B., Steup, M., Karas, M. and Hillenkamp, F.
(1994) The oligosaccharides of the Fe(III)-Zn(II) purple acid phosphatase of the red
kidney bean. Determination of the structure by a combination of matrix-assisted laser
desorption/ionization mass spectrometry and selective enzymic degradation. Eur. J.
Biochem. 220, 321-330.
Sugiyama, N., Nakagami, H., Mochida, K., Daudi, A., Tomita, M., Shirasu, K. and
Ishihama, Y. (2008) Large-scale phosphorylation mapping reveals the extent of
tyrosine phosphorylation in Arabidopsis. Mol. Syst. Biol. 4, 193.
Tran, H.T. and Plaxton, W.C. (2008) Proteomic analysis of alterations in the secretome of
Arabidopsis thaliana suspension cells subjected to nutritional phosphate deficiency.
Proteomics 8, 4317-4326.
Tran, H.T., Hurley, B.A. and Plaxton, W.C. (2010a) Feeding hungry plants: The role of
purple acid phosphatases in phosphate nutrition. Plant Sci. 179, 14-27.
Tran, H.T., Qian, W., Hurley, B.A., She, Y., Wang, D. and Plaxton, W.C. (2010b)
Biochemical and molecular characterization of AtPAP12 and AtPAP26: The
predominant purple acid phosphatase isozymes secreted by phosphate-starved
Arabidopsis thaliana. Plant Cell Environ. 33, 1789-1803.
Varki, A. (1993) Biological roles of oligosaccharides - all of the theories are correct.
Glycobiology 3, 97-130.
Veljanovski, V., Vanderbeld, B., Knowles, V.L., Snedden, W.A. and Plaxton, W.C.
(2006) Biochemical and molecular characterization of AtPAP26, a vacuolar purple acid

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
phosphatase up-regulated in phosphate-deprived Arabidopsis suspension cells and
seedlings. Plant Physiol. 142, 1282-1293.
Wang, L., Lu, S., Zhang, Y., Li, Z., Du, X. and Liu, D. (2014) Comparative genetic analysis
of Arabidopsis purple acid phosphatases AtPAP10, AtPAP12, and AtPAP26 provides
new insights into their roles in plant adaptation to phosphate deprivation. J. Integr. Plant
Biol. 56, 299-314.
Wang, L., Li, Z., Qian, W., Guo, W., Gao, X., Huang, L., Wang, H., Zhu, H., Wu, J., Wang,
D. and Liu, D. (2011) The Arabidopsis purple acid phosphatase AtPAP10 is
predominantly associated with the root surface and plays an important role in plant
tolerance to phosphate limitation. Plant Physiol. 157, 1283-1299.
Zamani, K., Lohrasebi, T., Sabet, M.S., Malboobi, M.A. and Mousavi, A. (2014)
Expression pattern and subcellular localization of Arabidopsis purple acid phosphatase
AtPAP9. Gene Expr. Patt. 14, 9-18.
Zhang, W., Gruszewski, H.A., Chevone, B.I. and Nessler, C.L. (2008) An Arabidopsis
purple acid phosphatase with phytase activity increases foliar ascorbate. Plant Physiol.
146, 431-440.
Zhu, H.F., Qian, W.Q., Lu, X.Z., Li, D.P., Liu, X., Liu, K.F. and Wang, D.W. (2005)
Expression patterns of purple acid phosphatase genes in Arabidopsis organs and
functional analysis of AtPAP23 predominantly transcribed in flower. Plant Mol. Biol. 59,
581-594.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Table 1 Substrate specificity of purified AtPAP25
Substrate Relative Activity
β-naphthyl-P
PEP
Ovalbumin
5-Br-4-Cl-3-indoyl-P
Phosphothreonine
para-nitrophenyl-P
Phosphoserine
Phosvitin
ATP
Phenyl-P
Phosphotyrosine
PPi
AMP
Glucose-6-P
6-P-Gluconate
Glucose-1-P
α-naphthyl-P
111 ±9
100 ±6
94 ±7
91 ±8
81 ±4
82 ±6
77 ±6
71 ±5
64 ±3
56 ±4
50 ±5
48 ±3
41 ±2
32 ±3
30 ±3
14 ±1
9 ±1
APase activity was determined with saturating concentrations of the storage
phosphoproteins ovalbumin and phosvitin (5 mg ml-1 each), or the indicated phosphorylated
compounds (5 mM each) using the spectrophotometric Pi assay described in the Methods.
Activity is expressed relative to the rate of Pi hydrolysis obtained with 5 mM PEP, set at
100%. All values represent means ±SE of n = 3 separate determinations.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Figure legends
Figure 1. Upregulation of APase in CWs of Arabidopsis suspension cells becoming Pi
deficient.
At d-0, 50 ml aliquots of cells cultured for 7-d in 5 mM Pi were subcultured into 450 ml of
fresh MS media containing 0 or 5 mM Pi (–Pi and +Pi, respectively). Time-courses for cell
culture filtrate (CCF) Pi concentration (a) and CaCl2-extractable CW APase activity (b) were
determined. All values represent means ±SE of n = 3 separate flasks. Where invisible, the
error bars are too small to be seen. (c) Pooled butyl Sepharose eluates from +Pi versus –Pi
CW extracts (1 µg per lane) were subjected to immunoblotting with anti-AtPAP12. Native
AtPAP12 and AtPAP26 (50 ng/lane) purified from CCF of –Pi Arabidopsis suspension cells
(Tran et al., 2010b) served as positive controls.
Figure 2. Fractogel SO3--cation exchange FPLC of CW extracts from Arabidopsis
suspension cells.
APase isoforms of butyl Sepharose-enriched CW extracts of the +Pi (a) versus –Pi (b) cells
were resolved via Fractogel SO3- cation-exchange FPLC. APase activity and A280 values are
indicated with a dotted and solid line, respectively. The column was developed with a 0-1 M
KCl gradient (indicated by the dashed line).
Figure 3. Deduced amino acid sequence alignment of CW-targeted AtPAP isozymes
upregulated by Pi-deficient Arabidopsis.
Identical amino acids are denoted by an asterisk; the arrow indicates their predicted transit
peptide cleavage site. N-terminal sequences obtained by automated Edman degradation of
AtPAP12, AtPAP25, and AtPAP26-CW1 purified from CWs of –Pi cells are indicated in bold
font above the deduced sequences. Amino acid sequences obtained during MS/MS of tryptic
and proteinase K peptides derived from purified AtPAP25 are underlined. The boxes denote
the positions of AtPAP25’s Asn172, Asn367, and Asn424 N-linked glycosylation sites.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Figure 4. SDS-PAGE and immunoblot analysis of CW PAPs purified from Pi-starved
Arabidopsis suspension cells.
(a) Purified PAPs (5 µg each) were subjected to 10% SDS-PAGE and stained with
Coomassie Blue R-250. ‘MW’ denotes several molecular mass standards. (b) Immunoblot
analysis was performed following SDS-PAGE as indicated. For anti-AtPAP12 and AtPAP26
immunoblots, 250 ng of each purified PAP was loaded, while for anti-AtPAP25 blots 100 ng
of AtPAP12, 200 ng of AtPAP26-CW1, 300 ng of AtPAP26-CW2, and 55 ng of AtPAP25
were loaded.
Figure 5. AtPAP25 is strongly induced during nutritional Pi deprivation.
Analysis of gene expression in suspension cell cultures and shoots and roots of
hydroponically-cultivated seedlings was performed as described in the Experimental
Procedures. (a) Levels of mRNA were analyzed by semi-quantitative RT-PCR. AtPAP12 and
AtPPCK1 served as positive controls for PSI genes (Gregory et al., 2009; Tran et al.,
2010b), whereas AtACT2 was used as a reference to ensure equal template loading. All
PCR products were taken at cycle numbers determined to be non-saturating. Control RT-
PCR reactions lacking reverse transcriptase did not show any bands. The AtPAP12,
AtPAP25, AtPAP26, AtPPCK1, and AtACT2 transcripts amplified as 244, 610, 627, 483 and
220 bp fragments of the 5′ region of their respective cDNAs, as expected. (b) Levels of
AtPAP25 transcripts were quantified by qPCR. AtPAP25 transcript levels in shoots and roots
of +Pi seedlings were used as the calibrator, whereas AtACT2 was used as an internal
control for normalization. All values represent means ±SE of n = 3 independent
determinations using cDNAs prepared from three biological replicates. (c) Pooled butyl
Sepharose APase activity fractions from CW extracts of +Pi and –Pi suspension cells were
subjected to SDS-PAGE and immunoblotted with anti-AtPAP25 (1 µg protein per lane).
AtPAP25 purified from CWs of the –Pi suspension cells (50 ng) served as a positive control.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Figure 6. AtPAP25:GUS is highly expressed in shoot vasculature of Pi-deficient Arabidopsis.
(a) AtPAP25:GUS transgenic lines were cultivated aseptically in 24-well microtitre plates in
liquid 0.5x MS media containing 1% (w/v) sucrose and 0.2 mM Pi for 7-d, before being
transferred into media containing 0 or 1.5 mM KH2PO4 (–Pi and +Pi, respectively) and grown
for another 3- and 7-d. (b) AtPAP12-GUS expression was also examined in several aerial
tissues of 28-d old +Pi versus –Pi soil-grown plants. (c) ‘Germinating Seed’ is a
representative image of seeds that had been placed on moistened filter paper and allowed to
germinate for 1-d prior to GUS staining. All images in panels a-c are representative of those
obtained with 11 independent transgenic lines.
Figure 7. Development of an atpap25 mutant is blocked when transferred into a soluble Pi-
deficient soil mixture.
(a) Schematic representation of the AtPAP25 gene (At4g36350); white boxes and solid lines
represent exons and introns, respectively. T-DNA insertion location is indicated by ‘atpap25
T-DNA’, while arrows represent primers used for PCR and genotyping. (b) Confirmation of T-
DNA insert location in atpap25 via PCR-based screening of gDNA template isolated from
leaves of +Pi seedlings. PCR products were amplified from the Col-0 and atpap25 gDNA in
a PCR reaction using AtPAP25-specific primers (primer pair A + B) and the overlapping
region between AtPAP25 and the T-DNA insert-specific primers (primer pair B + C). The lack
of multiple amplification products for AtPAP25 DNA in atpap25 demonstrates that the mutant
is homozygous. NC, negative control. (c) Influence of nutritional P-status on development of
soil-grown wild-type (Col-0) and atpap25 mutant seedlings. Seedlings were cultivated for 7-d
in liquid media containing 0.2 mM Pi and 1% (w/v) sucrose, then transferred into a soluble
Pi-deficient soil mixture and grown for an additional 14-d under a regular light/dark diurnal
cycle. Fertilization occurred biweekly with 0.25x Hoagland’s media containing 0 or 2 mM Pi
(–Pi and +Pi, respectively); scale bars = 1 cm. Results are representative of a minimum of 5
independent trials with at least 5 plants per trial. (d) Total Pi and anthocyanin concentrations
of rosette leaves of 21-d old seedlings cultivated the in the –Pi soil mixture. All values are

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
means ±SE of n = 5 different pooled leaf samples; asterisks denote values that are
significantly different (p < 0.05).
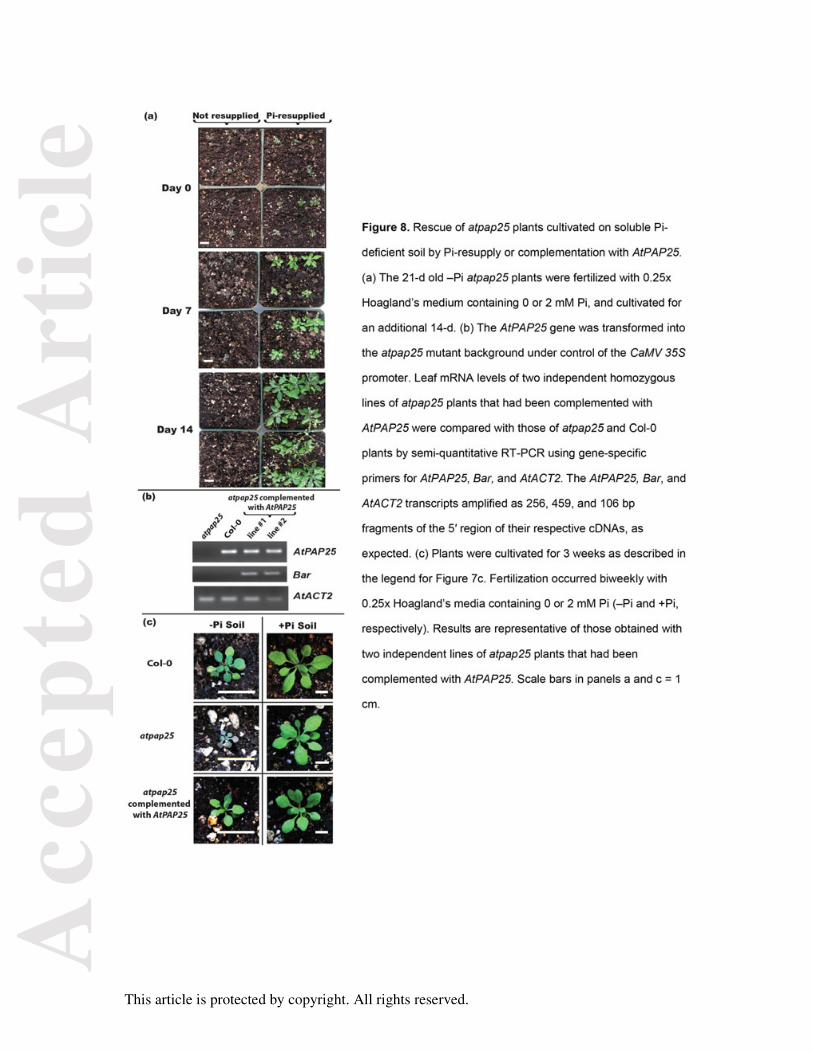
Figure 8. Rescue of atpap25 plants cultivated on soluble Pi-deficient soil by Pi-resupply or
complementation with AtPAP25.
(a) The 21-d old –Pi atpap25 plants were fertilized with 0.25x Hoagland’s medium containing
0 or 2 mM Pi, and cultivated for an additional 14-d. (b) The AtPAP25 gene was transformed
into the atpap25 mutant background under control of the CaMV 35S promoter. Leaf mRNA
levels of two independent homozygous lines of atpap25 plants that had been complemented
with AtPAP25 were compared with those of atpap25 and Col-0 plants by semi-quantitative
RT-PCR using gene-specific primers for AtPAP25, Bar, and AtACT2. The AtPAP25, Bar,
and AtACT2 transcripts amplified as 256, 459, and 106 bp fragments of the 5′ region of their
respective cDNAs, as expected. (c) Plants were cultivated for 3 weeks as described in the
legend for Figure 7c. Fertilization occurred biweekly with 0.25x Hoagland’s media containing
0 or 2 mM Pi (–Pi and +Pi, respectively). Results are representative of those obtained with
two independent lines of atpap25 plants that had been complemented with AtPAP25. Scale
bars in panels a and c = 1 cm.
Figure 9. Pi-starvation signaling is disrupted in the atpap25 mutant.
Plants were cultivated for 3 weeks as described in the legend for Figure 7c, and levels of
shoot transcripts for several genes determined by qPCR. Transcript levels in leaves of +Pi
seedlings were used as the calibrator, whereas AtACT2 was used as an internal control for
normalization. All values represent means ±SE of n = 3 independent determinations using
cDNAs prepared from three biological replicates; asterisks denote values that are
significantly different (p < 0.05) between Col-0 and atpap25.
Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.
Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved.





























