Embed Size (px)
Citation preview

ORIGINAL PAPER
The biological behavior of SDF-1/CXCR4 in patientswith myelodysplastic syndrome
Rui Yang • Jie Pu • Juan Guo • Feng Xu •
Zheng Zhang • Youshan Zhao • Xi Zhang •
ShuCheng Gu • ChunKang Chang • Xiao Li
Received: 28 March 2011 / Accepted: 1 April 2011 / Published online: 10 May 2011
� Springer Science+Business Media, LLC 2011
Abstract The purpose of this investigation is to evaluate
the biological behavior of stromal cell-derived factor-l
(SDF-1) in migration, adhesion, and apoptosis as well as
the related signaling transduction pathways in patients with
myelodysplastic syndrome (MDS) and acute myeloid leu-
kemia (AML). We chose 22 patients with MDS, 7 patients
with de novo AML, and 8 patients with non-clonal cyto-
penia diseases. We performed flow cytometric analysis of
CD34? cells apoptosis using annexinV-FITC, which binds
to exposed phosphatidylserine on apoptotic cells. The cell
adhesion capability was detected by CCK-8 assay. The
migration ability of the cell was checked by transwell
assay. Furthermore, we measured SDF-1 levels in BM
plasma from patients by enzyme-linked immunosorbent
assay (ELISA). Our results indicated that the apoptosis of
CD34? cell was significantly increased in the Low-grade
MDS (IPSS score B 1.0) patients compared with the high-
grade MDS (IPSS score C 1.5) (21.33% vs. 7.27%,
P \ 0.001) and patients with de novo AML (21.33% vs.
7.53%, P \ 0.001). SDF-1 promoted CXCR4 high
expression cells adhesion to the stroma cells (MSC) and
induced these cells migration. SDF-1 could trigger the
occurrence of polarized morphology of the cells that
expressed CXCR4 high. After addition of wortmannin or
PTX, the ability of adhesion and migration of the cells that
expressed CXCR4 high decreased. But in the patient’s cells
that expressed CXCR4 low, there was no above-mentioned
phenomenon. So we can suppose that the signaling path-
way of SDF-1/CXCR4 axis is PI3K pathway, and we
should do more things about this pathway and may find out
the target treatment of MDS.
Keywords SDF-1 � Migration � Adhesion �Myelodysplastic syndromes
Introduction
Myelodysplastic syndromes (MDS) comprised a hetero-
geneous group of hematopoietic stem cell malignancies
characterized by ineffective bone marrow (BM) hemato-
poiesis, peripheral blood (PB) cytopenias, and substantial
risk for progression to acute myeloid leukemia (AML).
Either bone marrow failure or progression to AML can
cause patients’ death.
Chemokines are a superfamily of small molecule
chemoattractive cytokines that mediate several cellular
functions. Stromal cell-derived factor-1 (SDF-1) is member
of the CXC subfamily of chemokines, is produced by
stromal and other cell types, and interacts with the seven-
transmembrane G-protein-coupled receptor CXCR4 [1].
CXCR4 is expressed constitutively in a wide variety of
normal tissue, including lymphatic tissues, thymus, brain,
spleen, stomach, and small intestine [2]. SDF-1 and its
receptor CXCR4 were involved in the mobilizing and
trafficking of hematopoietic progenitor cells [3]. SDF-1
also enhanced survival/anti-apoptosis of hematopoietic
progenitor cells, in vitro as well as in vivo [4], suggesting
R. Yang � J. Guo � F. Xu � Z. Zhang � Y. Zhao � X. Zhang �S. Gu � C. Chang (&) � X. Li
Department of Hematology, Shanghai Sixth People’s Hospital
Affiliated of Shanghai Jiaotong University, 200233 Shanghai,
China
e-mail: [email protected]
R. Yang � Y. Zhao
Medical college, Su Zhou University, 215123 Suzhou, China
J. Pu
Department of Hematology, Hospital Affiliated of Guilin
Medical College, 541001 Guilin, China
123
Med Oncol (2012) 29:1202–1208
DOI 10.1007/s12032-011-9943-7

that the SDF-1/CXCR4 system played an important role in
hematopoietic cell regulation.
In this study, we detected the expression of SDF-1/
CXCR4 axis, and its role in the migration, adhesion and
apoptosis of patients’ cells with MDS was evaluated either.
We may understand the mechanism of MDS and then find
out new pathways of diagnosis and treatment.
Materials and methods
Study population
After informed consent was given, bone marrow (BM)
mononuclear cells were obtained from 22 patients with
newly diagnosed MDS (15 patients’ IPSS score were not
more than 1 and 7 patients’ were not less than 1.5), from 7
de novo AML patients and from 8 patients with non-clonal
cytopenia diseases.
Material
All the monoclonal antibodies and the flow cytometry were
purchased from Becton–Dickinson. ELISA Kits were
purchased from R&D Systems. MicroplateReader was
Labsystems Dragon Wellscan MK-3. Cell Counting Kit-8
(CCK-8) was purchased from DOJINDO Laboratories.
AMD3100, pertussis toxin (PTX), and wortmannin were
purchased from Sigma. 5 lm membrane diameter of the
Transwell inserts cell culture plates were purchased from
Corning. Cell incubator was purchased from SANYO.
Fluorescence microscope was purchased from Olympus.
Mesenchymal stem cells (MSC) were the gift of hematol-
ogy laboratory of Shanghai sixth people’ hospital.
Flow cytometry assay
Bone marrow aspirates of 22 consecutive patients diag-
nosed with MDS between 2009 and 2010 at Shanghai Sixth
People’s Hospital (Shanghai, China) were obtained after
informed consent at the time of diagnosis and examined for
CXCR4 expression during routine flow cytometry work-up,
generally within 2–4 h after the sample was drawn. The
diagnosis of MDS was established using standard mor-
phology, cytochemistry, immunophenotypic criteria, and
cytogenetic evaluation according to the WHO classifica-
tion. For immunodetection of CXCR4 on MDS cells, a
whole-blood lysis method was used with directly labeled
primary antibodies following the manufacturer’s instruc-
tions (Becton–Dickinson, Franklin Lakes, NJ, USA).
Briefly, mononuclear cells were isolated from 2 mL bone
marrow with heparin sodium as the anticoagulant. After
adjusted to 106/mL by the phosphate buffered solution
(PBS), 100 lL of the cell suspension were incubated for
15 min with appropriately diluted monoclonal antibody
conjugates at room temperature protected from the light.
For CXCR4 detection, 10 lL of APC-conjugated anti-
CXCR4 (12G5) was used in combination with 10 lL of
anti-CD34-PerCP. A control sample was incubated with
the appropriate isotype control antibodies. For apoptosis
detection, 5 lL of FITC-conjugated AnnexinV was used in
combination with 2 lL PI. After incubation, cells were
washed once and analyzed within 2 h on a FACS Calibur
(Becton–Dickinson). All antibodies were purchased from
Becton–Dickinson. Flow cytometry data were analyzed
using the FlowJo software (version 2.7.4; Tree Star,
Ashland, OR).
ELISA assay
Plasma obtained from a heparin sodium-anticoagulated
bone marrow sample was kept at -80�C until required for
ELISA determinations. SDF-1 ELISA was developed with
appropriate pairs of coating and catcher anti-SDF-1 anti-
bodies, as well as with standard recombinant human
SDF-1a (R&D Systems, USA). The ELISA conditions for
antibody coating, blocking, reagent for plasma dilution,
and enzyme-developing reaction were as recommended by
the manufacturer (R&D Systems, USA) with ELISA high-
binding capacity plates (R&D Systems, USA). All deter-
minations were done in duplicate, and in most cases, they
were confirmed in an independent experiment.
CCK-8 assay
MSC were seeded in 96-well cell culture plates at a density
of 1 9 105 cells/well and allowed to grow overnight, and
then we planted patients’ bone marrow mononuclear cell
into the plates for 5 9 105 cells/well. The inhibitor groups’
bone marrow mononuclear cells were pretreated with the
corresponding drugs for 2 h. The final concentration of
each drug was as follows: 100 ng/mL of SDF-1, 1 lg/mL
of AMD3100, 200 ng/mL of PTX, 400 ng/mL of wort-
mannin. SDF-1 was added to each well, either. The control
group used PBS instead of SDF-1. Then incubated them
under the environment of 37�C, 5% CO2 for 6 h. 10 lL of
CCK-8 was added into each well, then incubated them
under the environment of 37�C, 5% CO2 for 4 h. Detected
the OD at 450 nm within 30 min.
Transwell assay
A concentration of 5 9 105/mL bone marrow mononuclear
cell was incubated with 10 lL CXCR4-APC protected
from light for 15 min. After washed twice using PBS, cells
were suspended by 100 lL DMEM. The inhibitor groups’
Med Oncol (2012) 29:1202–1208 1203
123
bone marrow mononuclear cell were pretreated with the
corresponding drugs for 2 h. The final concentration of
each drug was as follows: 100 ng/mL of SDF-1, 1 lg/mL
of AMD3100, 200 ng/mL of PTX, and 400 ng/mL of
wortmannin. Then we added 600 lL DMEM (contained
60 ng SDF-1) into the bottom room, the control group
using 600 lL DMEM not contained SDF-1. Then we
planted cells into the upper room. Then incubated them
under the environment of 37�C, 5% CO2 for 4 h.
Results
Apoptosis assay
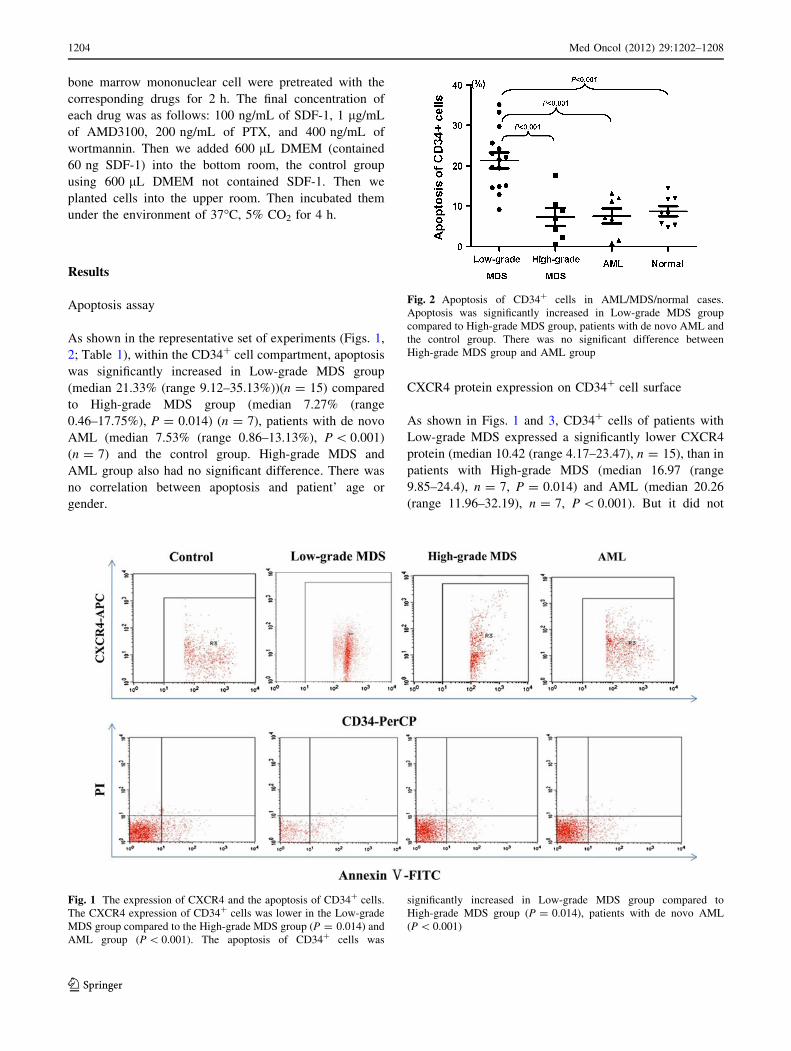
As shown in the representative set of experiments (Figs. 1,
2; Table 1), within the CD34? cell compartment, apoptosis
was significantly increased in Low-grade MDS group
(median 21.33% (range 9.12–35.13%))(n = 15) compared
to High-grade MDS group (median 7.27% (range
0.46–17.75%), P = 0.014) (n = 7), patients with de novo
AML (median 7.53% (range 0.86–13.13%), P \ 0.001)
(n = 7) and the control group. High-grade MDS and
AML group also had no significant difference. There was
no correlation between apoptosis and patient’ age or
gender.
CXCR4 protein expression on CD34? cell surface
As shown in Figs. 1 and 3, CD34? cells of patients with
Low-grade MDS expressed a significantly lower CXCR4
protein (median 10.42 (range 4.17–23.47), n = 15), than in
patients with High-grade MDS (median 16.97 (range
9.85–24.4), n = 7, P = 0.014) and AML (median 20.26
(range 11.96–32.19), n = 7, P \ 0.001). But it did not
Fig. 1 The expression of CXCR4 and the apoptosis of CD34? cells.
The CXCR4 expression of CD34? cells was lower in the Low-grade
MDS group compared to the High-grade MDS group (P = 0.014) and
AML group (P \ 0.001). The apoptosis of CD34? cells was
significantly increased in Low-grade MDS group compared to
High-grade MDS group (P = 0.014), patients with de novo AML
(P \ 0.001)
Fig. 2 Apoptosis of CD34? cells in AML/MDS/normal cases.
Apoptosis was significantly increased in Low-grade MDS group
compared to High-grade MDS group, patients with de novo AML and
the control group. There was no significant difference between
High-grade MDS group and AML group
1204 Med Oncol (2012) 29:1202–1208
123
reach statistical significance between Low-grade MDS and
the control group (P = 0.496). We were also unable to
find significant difference between High-grade MDS and
AML.
SDF-1 levels in BM plasma
Furthermore, we measured SDF-1 levels in BM plasma
from patients by enzyme-linked immunosorbent assay
(ELISA) using Quantikines, human SDF-1a immunoassay
(R&D systems, Minneapolis, MN, USA). But there were
no significant difference among the four groups as shown
in Fig. 4.
The adhesion rate of bone marrow mononuclear cells
with MSC
In the high-grade MDS group, the adhesion rate of SDF-1
group was increased significantly compared with other
groups (P \ 0.001, Table 2). And this increase could be
blocked by drugs. But in the low-grade MDS group, there
was no significant difference among groups.
The migration rate of bone marrow mononuclear cells
In the high-grade MDS group, the migration rate of SDF-1
group was increased significantly compared with other
groups (P \ 0.001, Table 2). And this increase could be
blocked by drugs. But in the low-grade MDS group, there
was no significant difference among groups.
Polarized morphology of the cells
After the migration, cells showed a polarized morphology
in the SDF-1 group of the high-grade MDS group.
This phenomenon did not appear in other groups (Fig. 5).
Discussion
SDF-1 is a CXC chemokine, secreted constitutively from
several cell types. There are 2 isoforms of SDF-1, a and b,
which are generated by differential splicing from a single
gene. CXCR4 is a 7-transmembrane surface structure
linked to G proteins. The chemotactic effect of SDF-1 on
hematopoietic progenitor cells has been shown to be
mediated by the CXCR4 receptor. CXCR4 plays a central
role in both progenitor cell homing and mobilization
[5–10].
Phosphoinositide 3-kinase family members are proto-
oncogenes. Generation of 3-phosphoinositides has been
documented to influence diverse cellular pathways and
hence alter a spectrum of fundamental cellular activities. It
can generate 4 forms of the phosphoinositide: PI(3)PI,
PI(3,4)P2, PI(3,5)P2, PI(3,4,5)P3. The upregulation of PI3K
can cause Akt decrease by phosphorylation [11, 12]. Liang
et al. [13] had shown that CXCR4 upregulates the phos-
phorylation of Akt and increases VEGF expression at both
mRNA and protein levels. CXCL12 increases phosphory-
lation of Akt in a dose-dependent manner, indicating a
Fig. 3 CXCR4 expression of CD34? cells in AML/MDS/normal
cases. CD34? cells of patients with Low-grade MDS expressed a
significantly lower CXCR4 protein than in patients with High-grade
MDS and AML. High-grade MDS group expressed a significantly
higher CXCR4 protein than the control group. There were no
significant difference between Low-grade MDS and the control group
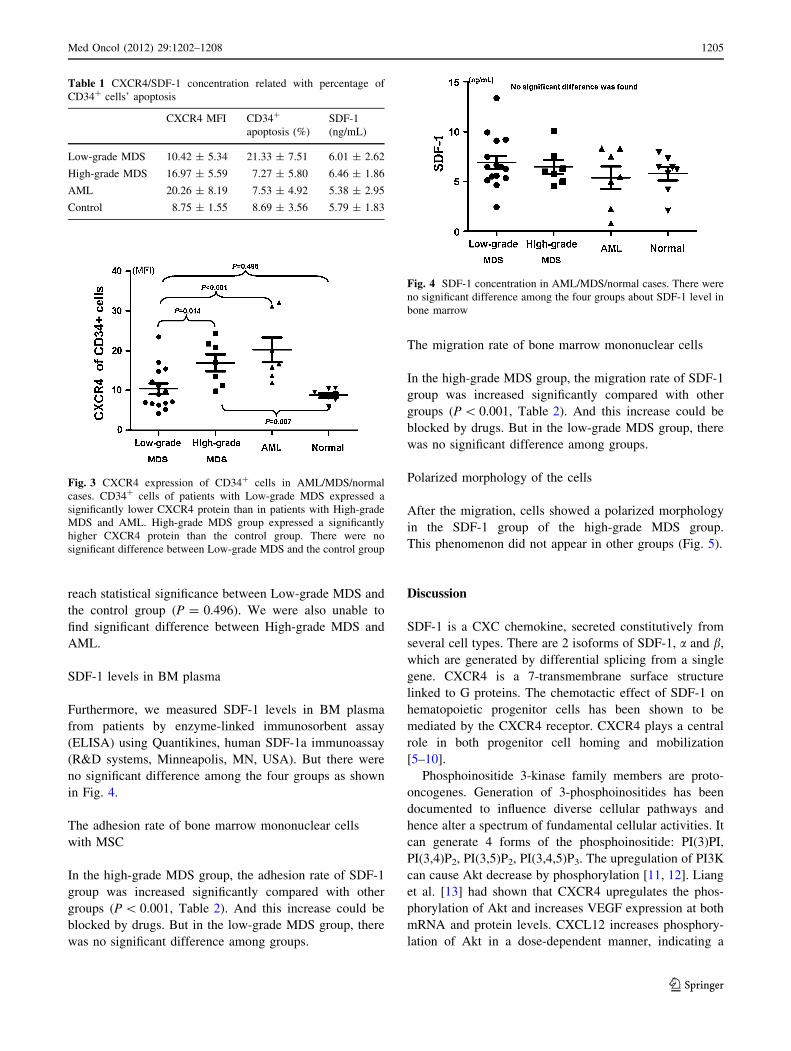
Fig. 4 SDF-1 concentration in AML/MDS/normal cases. There were
no significant difference among the four groups about SDF-1 level in
bone marrow
Table 1 CXCR4/SDF-1 concentration related with percentage of
CD34? cells’ apoptosis
CXCR4 MFI CD34?
apoptosis (%)
SDF-1
(ng/mL)
Low-grade MDS 10.42 ± 5.34 21.33 ± 7.51 6.01 ± 2.62
High-grade MDS 16.97 ± 5.59 7.27 ± 5.80 6.46 ± 1.86
AML 20.26 ± 8.19 7.53 ± 4.92 5.38 ± 2.95
Control 8.75 ± 1.55 8.69 ± 3.56 5.79 ± 1.83
Med Oncol (2012) 29:1202–1208 1205
123

direct relationship between CXCR4/CXCL12 axis and Akt.
Inversely, blocking the activation of Akt signaling led to a
decrease in VEGF protein levels; blocking CXCR4/
CXCL12 interaction with a CXCR4 antagonist suppressed
tumor angiogenesis and growth in vivo.
There was significant difference on the development and
termination between Low-grade MDS and High-grade
MDS. In our study, we divided MDS patients into two
groups according to the International Prognostic Scoring
System (IPSS) [14]: Low-grade MDS (IPSS score B 1.0)
and High-grade MDS (IPSS score C 1.5).
There was no significant difference on CXCR4 expres-
sion between Low-grade MDS and the control group. But
the apoptosis of CD34? cells was higher in the Low-grade
MDS than it in controls. Considering that SDF-1 was the
only ligand for CXCR4, we furthermore detected the SDF-
1 level in the BM plasma by using ELISA assays. But there
was no significant difference between two groups (Fig. 3).
Our results were different from Matsuda’s report [15].
Broxmeyer et al. [4, 16, 17] reported that in the study of
SDF-1 transgene mice, SDF-1 played anti-apoptisis role on
the hemopoietic progenitor cells. Yin et al. [18] verified
that SDF-1 inhibited hypoxia/serum deprivation induced
bone marrow-derived mesenchymal stem cells apoptosis
through PI3K/Akt signaling pathways. We supposed that in
the Low-grade MDS, the SDF-1/CXCR4 signal pathway
might be injured. Liang [13] reported that SDF-1/CXCR4
signaling axis react though the activation of PI3K/Akt
pathway. Fuhler [19, 20] found that PI3K pathway was
abnormal in MDS patients.
In the High-grade MDS group, the CXCR4 expression
and apoptosis of CD34? cells was similar to AML group.
Clinically, High-grade MDS patients had higher risk for
progression to acute myeloid leukemia compared with
Low-grade MDS. CXCR4 expression is higher in High-
grade group than the control group (P = 0.007) (Fig. 2),
while the apoptosis between two groups being no signifi-
cant difference. It needed higher CXCR4 level to remain
low apoptosis compared with controls. This also can
demonstrate that the SDF-1/CXCR4 signal pathway was
injured.
In the adhesion assays, because of the SDF-1, cells of
high-expression CXCR4 can adhere to MSC, but there was
no this phenomenon on cells of low-expression CXCR4.
Furthermore, we found that this phenomenon can be
blocked by AMD3100, PTX, and wortmannin. The
malignant cells can avoid the cytotoxicity of drugs by
adhering to the stroma cells. So, our study indicated that
SDF-1/CXCR4 axis can promote tumor cells adhering to
MSC. Zhang et al. [21] found that the adhesion capability
of AML cells was enhanced through changing the
expression of the adhesion molecules (ICAM-1 and
LFA-1). So, we thought that SDF-1/CXCR4 axis played an
important role in the adhesion of the MDS clone cells.
Table 2 The adhesion rate and migration rate in each group
Groups The adhesion rate (%) The migration rate (%)
Low grade-MDS High-grade MDS Low grade-MDS High-grade MDS
Control 19.78 ± 2.35 27.00 ± 4.58 16.71 ± 3.49 14.07 ± 2.87
SDF-1 23.24 ± 1.72 52.33 ± 7.51* 22.59 ± 2.41 70.18 ± 3.09**
SDF-1 ? AMD3100 17.48 ± 2.84 21.97 ± 5.01 18.35 ± 3.20 11.38 ± 2.37
SDF-1 ? PTX 20.33 ± 3.75 22.33 ± 2.52 16.27 ± 3.13 13.72 ± 2.61
SDF-1 ? wortmannin 22.59 ± 4.11 20.86 ± 4.04 17.94 ± 2.74 13.29 ± 4.56
* The adhesion rate of SDF-1 group was increased significantly compared with other groups (P \ 0.001)
** The migration rate of SDF-1 group was increased significantly compared with other groups (P \ 0.001)
Fig. 5 Polarized morphology of the migrated cells. The SDF-1
group, the number of cells migrated from upper-room to lower-room
was higher than others. And we can found a polarized morphology on
the migrated cells in the SDF-1 group. But in the other groups, there
was no this phenomenon
1206 Med Oncol (2012) 29:1202–1208
123

Furthermore, we found that in High-grade MDS group,
the number of cells that migrate from upper-room to lower-
room in the SDF-1 group increased significantly compared
with in other groups. And, it can be neutralized by
AMD3100, PTX, and wortmannin. In the High-grade MDS
group, morphous polarization was detected in the migrated
cells. Kimura et al. [22] reported that SDF-1/CXCR4 axis
can enhance the migration of bone marrow CD34? cells. Li
et al. [23] demonstrated that the PI3K/Akt signaling path-
way activated enhanced migration of neural-like cells
toward SDF-1. Some researchers reported that chemotaxis
of SDF-1 can be intercepted by PTX [24–26]. In the
AMD3100 group, the number of migrated cells decreased
significantly compared with the SDF-1 group. So, SDF-1
produced a marked effect through binding CXCR4. After
adding PTX and wortmannin, the migration disappeared.
This demonstrated that SDF-1/CXCR4 axis delivered
signals through PI3K pathways. In the High-grade MDS
group, cells were high-expression CXCR4; so, they had
high capability of adhesion and migration; So, High-grade
MDS patients usually developed into AML. In the Low-
grade MDS group, cells were low-expression CXCR4; so,
they had low capability of adhesion and migration; So,
Low-grade MDS patients usually developed into marrow
failure.
Totally, SDF-1/CXCR4 axis can promote the ability of
survival, adhesion to MSC and migration of high-expres-
sion CXCR4 cells. And, this promotion can be disrupted by
AMD3100, PTX, and wortmannin. The faculty for adhe-
sion and migration of tumor cells plays an important role in
the development of disease. We presume that SDF-1/
CXCR4 axis plays an important role in the mechanism of
MDS. Detecting the expression of SDF-1/CXCR4 and the
PI3K pathways can be the new target of diagnosis and
therapy of MDS.
References
1. Federsppiel B, Melhado IG, Duncan AM, et al. Molecular cloning
of the cDNA and chromosomalclocalization of the gene for a
putative seven-transmembrane segment (7-TMS) receptor iso-
lated from human spleen. Genomics. 1993;16(3):707–12.
2. Nagasawa T, Kikutani H, Kishimoto T. Molecular cloning and
structure of a pre-B-cell growth-stimulating factor. Proc Natl
Acad Sci USA. 1994;91(6):2305–9.
3. Aiuti A, Webb IJ, Bleul C, et al. The chemokine SDF-1 is a
chemoattractant for human hematopoietic progenitor cells and
provides a new mechanism to explain the mobilization of CD34?
progenitors to peripheral blood. J Exp Med. 1997;185(1):111–20.
4. Broxmeyer HE, Cooper S, Kohli L, et al. Transgenic expression
of stromal cell-derived factor-1/CXC chemokine ligand 12
enhances myeloid progenitor cell survival/antiapoptosis in vitro
in response to growth factor withdrawal and enhances myelo-
poiesis in vivo. J Immunol. 2003;170(1):421–9.
5. Peled A, Petit I, Kollet O, et al. Dependence of human stem cell
engraftment and repopulation of NOD/SCID mice on CXCR4.
Science. 1999;283(5403):845–8.
6. Lapidot T. Mechanism of human stem cell migration and
repopulation of NOD/SCID and B2mnull NOD/SCID mice. Ann
N Y Acad Sci. 2001;938:83–95.
7. Mohle R, Bautz F, Rafii S, Moore MAS, Brugger W, Kanz L.
The chemokine receptor CXCR4 is expressed on CD34?
hematopoietic progenitors and leukemic cells and mediates
transendothelial migration induced by stromal cell-derived factor-
1. Blood. 1998;91(12):4523–30.
8. Lapidot T, Petit I. Current understanding of stem cell mobilization:
the roles of chemokines, proteolytic enzymes, adhesion mole-
cules, cytokines, and stromal cells. Exp Hematol. 2002;30(9):
973–81.
9. Petit I, Szyper-Kravitz M, Nagler A, et al. G-CSF induces stem
cell mobilization by decreasing bone marrow SDF-1 and
up-regulating CXCR4. Nat Immunol. 2002;3(7):687–94.
10. Shen H, Cheng T, Olszak I, et al. CXCR-4 desensitization is
associated with tissue localization of hemopoietic progenitor
cells. J Immunol. 2001;166(8):5027–33.
11. Hanada M, Feng J, Hemmings BA. Structure, regulation and
function of PKB/AKT—a major therapeutic target. Biochim
Biophys Acta. 2004;1697(1–2):3–16.
12. Vanhaesebroeck B, Leevers SJ, Ahmadi K, et al. Synthesis and
function of 3-phosphorylated inositol lipids. Annu Rev Biochem.
2001;70:535–602.
13. Liang Z, Brooks J, Willard M, et al. CXCR4/CXCL12 axis
promotes VEGF-mediated tumor angiogenesis through Akt sig-
naling pathway. Biochem Biophys Res Commun. 2007;359(3):
716–22.
14. Greenberg P, Cox C, LeBeau MM, et al. International scoring
system for evaluating prognosis in myelodysplastic syndromes.
Blood. 1997;89(6):2079–88.
15. Matsuda M, Morita Y, Hanamoto H, et al. CD34? progenitors
from MDS patients are unresponsive to SDF-1, despite high
levels of SDF-1 in bone marrow plasma. Leukemia. 2004;18(5):
1038–40.
16. Broxmeyer HE, Cooper S, Hangoc G, et al. Stromal cell-derived
factor-1/CXCL12 selectively counteracts inhibitory effects of
myelosuppressive chemokines on hematopoietic progenitor cell
proliferation in vitro. Stem Cells Dev. 2005;14(2):199–203.
17. Broxmeyer HE, Kohli L, Kim CH, et al. Stromal cell-derived
factor-1/CXCL12 directly enhances survival/antiapoptosis of
myeloid progenitor cells through CXCR4 and G(alpha)i proteins
and enhances engraftment of competitive, repopulating stem
cells. J Leukoc Biol. 2003;73(5):630–8.
18. Yin Q, Jin P, Liu X, et al. SDF-1a inhibits hypoxia and serum
deprivation-induced apoptosis in mesenchymal stem cells
through PI3K/Akt and ERK1/2 signaling pathways. Mol Biol
Rep. 2011;38(1):9–16.
19. Fuhler GM, Drayer AL, Vellenga E. Decreased phosphorylation
of protein kinase B and extracellular signal-regulated kinase in
neutrophils from patients with myelodysplasia. Blood. 2003;
101(3):1172–80.
20. Fuhler GM, Drayer AL, Olthof SG, et al. Reduced activation of
protein kinase B, Rac, and F-actin polymerization contributes to
an impairment of stromal cell–derived factor-1–induced migra-
tion of CD34?cells from patients with myelodysplasia. Blood.
2008;111(1):359–68.
21. Zhang W, Zhung X, Fan X, et al. Effect of ICAM-1 and LFA-1 in
hyperleukocytic acute myeloid leukaemia. Clin Lab Haematol.
2006;28(3):177–82.
22. Kimura T, Boehmler AM, Seitz G, et al. The sphingosine 1-phos-
phate receptor agonist FTY720 supports CXCR4-dependent
Med Oncol (2012) 29:1202–1208 1207
123

migration and bone marrow homing of human CD34? progenitor
cells. Blood. 2004;103(12):4478–86.
23. Li S, Deng L, Gong L, et al. Upregulation of CXCR4 favoring
neural-like cells migration via AKT activation. Neurosci Res.
2010;67(4):293–9.
24. Bleul CC, Fuhlbrigge RC, Casasnovas JM, et al. A highly effi-
cacious lymphocyte chemoattractant, stromal cell-derived factor
1 (SDF-1). J Exp Med. 1996;184(3):1101–9.
25. Dutt P, Wang JF, Groopman JE. Stromal cell-derived factor-1
alpha and stem cell factor/kit ligand share signaling pathways in
hemopoietic progenitors: a potential mechanism for cooperative
induction of chemotaxis. J Immunol. 1998;161(7):3652–8.
26. Kim CH, Broxmeyer HE. In vitro behavior of hematopoietic
progenitor cells under the influence of chemoattractants: stromal
cell-derived factor-1, steel factor, and the bone marrow envi-
ronment. Blood. 1998;91(1):100–10.
1208 Med Oncol (2012) 29:1202–1208
123





























