Embed Size (px)
Citation preview

Angiogenesis, Metastasis, and the Cellular Microenvironment
New Insight into the SDF-1/CXCR4 Axis in a BreastCarcinoma Model: Hypoxia-Induced Endothelial SDF-1 andTumor Cell CXCR4 Are Required for Tumor Cell Intravasation
Fengyan Jin1, Ulf Brockmeier2, Friedrich Otterbach3, and Eric Metzen2
AbstractThe SDF-1/CXCR4 axis has been implicated in breast cancermetastasis. In contrast to its well-established role in
organ-specific homing and colonization of tumor cells, the involvement in intravasation, especially in a hypoxicenvironment, is still poorly understood. Initially, we detected both, the chemokine SDF-1 and its receptor CXCR4in microvessels in invasive ductal cancer samples. To elucidate the role of the SDF-1/CXCR4 axis in vascularendothelium for tumor intravasation, we evaluated the effects of CXCR4 activation in human umbilical vein anddermal microvascular endothelial cells (HUVEC andHDMEC) and in cultured mammary carcinoma cells (MDAMB231, and MCF7). We observed an upregulation of SDF-1 and CXCR4 in HUVECs in hypoxia, which led toproliferation, migration, and tube formation. Hypoxia induced adhesion of tumor cells to endothelial cells andstimulated transendothelial migration. The effects of hypoxia were dependent on the activity of the transcriptionfactor hypoxia-inducible factor. Adhesion to and migration through a HUVEC monolayer were significantlyreduced by lentiviral inhibition of CXCR4 in breast carcinoma cells or treatment of endothelial cells with an anti-SDF-1 neutralizing antibody. These data show that the interaction of SDF-1 secreted by ECs with tumor cellCXCR4 is sufficient to stimulate transendothelial migration of the tumor cells. Our results suggest that the SDF-1/CXCR4 axis is important in angiogenesis and tumor cell intravasation. Because both proteins were readilyidentifiable in a significant fraction of human breast cancer samples by immunohistochemistry, CXCR4 mayconstitute a molecular target for therapy when both, SDF-1, and CXCR4 are expressed. Mol Cancer Res; 10(8);1021–31. �2012 AACR.
IntroductionIn the United States, breast cancer is the most frequent
type of cancer with nearly 210,000 new cases diagnosed andalmost 40,000 deaths in 2010 (1). Metastasis, that istransport of tumor cells to and growth at secondary sites,is a major cause for morbidity and mortality (2, 3). Themetastatic process involves multiple steps including tumorcell mobilization, angiogenesis, intravasation, survival in thecirculation, extravasation, and proliferation in a new tissueenvironment. Despite increasing knowledge, all these stepsare incompletely understood. For example, at the primary
site, the communication of the tumor cells with their cellularmicroenvironment which is composed of fibroblasts, neu-trophils, mast cells, and endothelial cells, not only promotestumor growth, but also facilitates emigration from theoriginal location (4, 5). These studies strikingly suggest afunctionally important interaction between host-stromaand tumor. However, the mechanisms of intravasation aremostly elusive.In a hypoxic microenvironment, the function of endo-
thelial cells is modified toward an angiogenic phenotype.This response to hypoxia is indeed physiological, for exampleblood vessel development in the mammalian embryo is alsostimulated by hypoxia (6). In tumor biology, this pathway ishijacked by the tumor cells. When a tumor grows beyond adiameter of 2 mm, the tumor cells commonly turn hypoxic.As a response, the transcription factor complex hypoxia-inducible factor (HIF) is activated. HIF stimulates secretionof growth factors includingVEGF,which acts on endothelialcells to initiate angiogenesis. The developing capillary net-work in the tumor not only delivers nutrients and oxygen,but also provides the escape route by which detached cancercells can enter into the circulation.Stromal cell-derived factor-1 (SDF-1) is a highly con-
served chemokine that acts through its cognate receptorCXCR4. Chemokines are low molecular weight signaling
Authors' Affiliations: 1The First Hospital of Jilin University, Changchun,China; and 2Institute of Physiology, and 3Institute of Pathology and Neu-ropathology, University of Duisburg-Essen, Essen, Germany
Note: Supplementary data for this article are available at Molecular CancerResearch Online (http://mcr.aacrjournals.org/).
Current address for F. Jin: Department of Neurology, University of Duis-burg-Essen, Essen, Germany.
Corresponding Author: Eric Metzen, Institute of Physiology, University ofDuisburg-Essen, Hufelandstr. 55, Essen D45147, Germany. Phone: 49-201-7234626; Fax: 49-201-7235661; E-mail: [email protected]
doi: 10.1158/1541-7786.MCR-11-0498
�2012 American Association for Cancer Research.
MolecularCancer
Research
www.aacrjournals.org 1021
on April 13, 2017. © 2012 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst July 5, 2012; DOI: 10.1158/1541-7786.MCR-11-0498

peptides frequently involved in leucocyte mobilization andhoming. Specifically, the SDF-1/CXCR4 axis plays a criticalrole in B-cell development (7), angiogenesis (8), HIVinfection (9), as well as stem cell mobilization and homing(10). Both SDF-1 and CXCR4 have been reported to behypoxia-inducible (11, 12). Several lines of evidence suggestthat SDF-1 and CXCR4 are involved in tumor progressionand the development of distant metastases: SDF-1 secretedby tumor-associated fibroblasts has been reported to beproangiogenic (5). In a landmark study it was showed thatexpression of CXCR4 facilitates orthotopic tumor growthand metastatic potential in SCID mice although, notably,tumor cells were injected i.v., which indicates that theexperiments reflect late steps of metastasis (13). In anotherreport CXCR4 was identified in a human breast cancer cellline as 1 of 5 proteins which gives rise to bonemetastases afterintraventricular injection in mice (14). Correlation ofCXCR4 expression to HER2, a well-established prognosticfactor in mammary carcinoma was also observed in humanbreast tumors (15).However, neither SDF-1 nor CXCR4 has been identified
as a prognostic marker in large microarray DNA screenspublished by several independent groups (16, 17). A lowmolecular weight CXCR4 antagonist did not inhibit growthin amurine tumormodel (18). In one study it was found thatexpression of SDF-1 reduced the metastatic potential ofbreast carcinoma cell lines (19) and, furthermore, an immu-nohistochemical analysis of SDF-1 expression in humanbreast cancer patients came to the result that SDF-1 expres-sion was a marker of longer disease-free and overall survival(20). Collectively, these reports show that the role of SDF-1and CXCR4 in human tumors is still controversial, and inparticular, that the effect of SDF-1/CXCR4 activation intumor–host interaction at the primary site is elusive.It is still an open question whether tumor cells enter into
the vasculature passively or whether they migrate throughblood vessels actively. Evidence for active metastasis isprovided by the observation that the tumor microenviron-ment can induce migration-related pathways (21, 22) orcollaboration of stromal and tumor cells inmetastasis (4, 23).However, currently there is no study available in which thewhole sequence of events is followed in a single coculturemodel. Therefore, it was the aim of our study to providemechanistic insight into the role of the SDF-1/CXCR4 axisin the early steps of metastasis with particular emphasis onthe effects of hypoxia in a coculture system of breast cancerand endothelial cells. We observed that endothelial SDF-1stimulated active adhesion of CXCR4 expressing breastcarcinoma cells to endothelial cells and transendothelialmigration. Importantly, we confirmed endothelial SDF-1and tumor cell CXCR4 expression in a significant fraction ofsamples from mammary tumors which pinpoints the rele-vance of our data in a clinical setting.
Materials and MethodsImmunohistochemistry and immunofluorescenceStaining of SDF-1 and CXCR4 was carried out in 140
samples of primary breast carcinomas chosen from a collec-
tion of our Institute of Pathology. The patients presentedbetween 1989 and 1993 were operated with curative intentat the Department of Obstetrics and Gynaecology, Univer-sity Hospital Essen. The study was carried out according tothe Declaration of Helsinki and approved by the local EthicsCommittee. Immunohistochemistry was conducted asdescribed previously (24). Following individually optimizedheat-based antigen retrieval for each antibody, SDF-1 wasstained with a monoclonal anti- CXCL12/SDF1 [clone79018 (R&DSystems, Inc.) dilution 1:1000, antigen retriev-al: 20 minutes 95�C water bath, citrate buffer, pH 6.0].CXCR4 immunohistochemistry was done with a rabbitmonoclonal antibody generously provided by Prof. StefanSchulz, Jena, Germany, dilution 1:30, antigen retrieval as forSDF-1. CXCR7 was stained with a monoclonal antibody(MAB42273; R&D Systems). Immunofluorescence was car-ried out with the sameCXCL12/SDF-1 antibody and humanvon-Willebrand-Factor (vWF) antibody (dilution 1:100,rabbit polyclonal; Dako). SDF-1 and vWF were detectedwith an Alexa Fluor488 goat antimouse antibody and AlexaFluor568 goat antirabbit antibody (Invitrogen), respectively.
Cell cultureHuman umbilical vein endothelial cells (HUVEC) and
human dermal microvascular endothelial cells (HDMEC),both from PromoCell were grown in endothelial cell growthmedium (ECGM; PromoCell) containing the recom-mended additives. ECs were used for experiments frompassage 3 to 6. HEK-293T, MDA-MB 231, and MCF7cells were obtained from ATCC. MDA-MB 231 weremaintained in DMEM (Invitrogen), MCF-7 cells weregrown in RPMI1640 (Invitrogen), supplemented with10% fetal calf serum, penicillin, and streptomycin. Hypoxicculture conditions were achieved in a hypoxic chamber(Toepffer Lab Systems) by inflation of N2 and CO2 intothe air-filled chamber until 1% O2 and 5% CO2 werereached.
Chemokines, antibodies, and reagentsRecombinant CXCL12/SDF-1a, CXCL12/SDF-1
monoclonal antibody (MAb) (Clone 79018, Mouse IgG1),CXCR4 (Fusin) MAb (Clones 12G5 and 44716), andaffinity-purified HIF-2a antibody were purchased fromR&D Systems. CXCR4 polyclonal antibodies were fromAbcam. HIF-1a MAb was from BD Transduction Labo-ratories. Mouse Ig G2a (Clone G155-178) was purchasedfrom BD Biosciences. Human a-tubulin MAb was fromSanta Cruz Biotechnology. Human actin antibody was fromSigma. Dimethyloxalylglycine (DMOG) was purchasedfrom Alexis Biochemicals.
Bacterial strains, plasmids, lentiviral transduction, andsiRNA transfectionEscherichia coli XL1-blue served as the general host for
plasmid preparation, E. coli Stbl3 (Invitrogen) were used forproduction of pLKO.1 (8453, Addgene), pLKO.1-shRNA-HIF-1a-1 (Mission TRC shRNA TRCN0000003810;Sigma), pLKO.1-shRNA-HIF-1a-2 (Mission TRC shRNA
Jin et al.
Mol Cancer Res; 10(8) August 2012 Molecular Cancer Research1022
on April 13, 2017. © 2012 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst July 5, 2012; DOI: 10.1158/1541-7786.MCR-11-0498

TRCN0000003811; Sigma) pLKO.1-shRNA-HIF-2a-1(Mission TRC shRNA TRCN0000003803; Sigma),pLKO.1-shRNA-HIF-2a-2 (Mission TRC shRNATRCN0000003806; Sigma), and pLKO.1-shRNA-CXCR4 (Mission TRC shRNA TRCN0000256864; Sig-ma). Generation of recombinant lentivirus was carried out asdescribed previously (25). Lentivirus-infected MDA-MB231 and MCF7 cells were selected by treatment with 1 and3 mg/mL puromycin, respectively. CXCR4 siRNA transfec-tion of MCF7 cells was carried out according to the Dhar-maFECT siRNA transfection protocol (Thermo FisherScientific). A total of 2 � 105 MCF7 cells were transfected,the final concentration of the siRNA was 25 nmol/L. Thecells were incubated with the transfection solution for 72hours. Gene silencing was monitored by Western blotting.
Western blotWhole-cell lysates from HUVECs, MDA-MB 231, and
MCF-7 were prepared andWestern blotting was carried outas reported (25).
Fluorescent labeling and flow cytometryThe fluorescent labeling of HUVECs, MDA-MB 231,
and MCF7 cells with calcein-AM (Invitrogen) was con-ducted according to the manufacturer's protocol. Fluores-cence was detected using a Zeiss Axiovert 200 M confocalmicroscope (Carl Zeiss). For FACS analysis, HUVECs orMDA-MB231 cells were detachedwith 2mmol/L EDTA inPBS. For CXCR4 determination and membrane SDF-1expression, the unstained cells were fixed by incubationwith 1% paraformaldehyde (PFA) for 5 minutes. The cellswere incubated with 20 mL PE-conjugated human CXCR4MAb (Clone 12G5) for 30 minutes at 4�C. For intracellularSDF-1 detection, the cells were permeabilized with Cytofix/Cytoperm (BD Biosciences) and incubated with 25 mg/mLCXCL12/SDF-1 MAb (Clone 79018) for 30 minutes at4�C. SDF-1 was detected with a PE-conjugated goat anti-mouse antibody (BDPharmingen). Cells were analyzed witha Beckman Coulter FACSCalibur. A total of 10,000 eventswere assessed. Data were analyzed by CELLQuest software(BD Biosciences).
Tube formation assayHUVECs (2� 104/well) were transduced with pLKO.1-
shRNA-HIF-1a, pLKO.1-shRNA-HIF-2a or pLKO.1-shRNA-CXCR4 and seeded into Matrigel-coated wells ofa 96-well plate. Twenty-four hours later, photographs weretaken with a Leica DFC 290 camera (Leica Microsystems).Only perfectly continuous tubes between 2 branching pointswere regarded as a tube.
Proliferation assayTo assess HUVEC proliferation, 1� 104 HUVECs were
seeded into 24-well plates in the absence or presence ofrecombinant human SDF-1 (1–100 ng/mL). CXCR4 neu-tralizing antibodies (10 mg/mL) or control mouse IgG (BDBiosciences) were added 12 hours before treatment withSDF-1. The viable cells were counted at day 3 and 6. When
the effects of endogenous SDF-1were to be blocked, 1� 104
HUVECs were seeded into a 24-well plate and cultured inserum-free ECGM in the absence or presence of an SDF-1neutralizing antibody for 3 days. For proliferation assays afterHIF-1a, HIF-2a, or CXCR4 knockdown, 8 � 104
HUVECs were transduced with the appropriate vectors andcultured for 3 days. To quantify tumor cell proliferation, 1�105 MDA-MB 231 were plated and grown for 4 days.
Tumor cell-to-endothelium adhesion assayA total of 5 � 104 HUVECs transduced with pLKO.1-
shRNA-CXCR4, or pLKO.1 were exposed to normoxia orhypoxia for 12 hours after themonolayer had formed. A totalof 1 � 104 calcein-labeled MDA-MB 231 cells were added,adhesion was allowed to occur for 2 hours at 37�C. Non-adherent cells were washed off, adherentMDA-MB231 cellswere quantified.
Adhesion of tumor cells to endothelial tubesA total of 1 � 104 calcein-labeled MDA-MB 231 or
MCF7 cells were cocultured with 2 � 104 HUVEC orHDMEC on a Matrigel-coated 96-well pate. Ten mg/mLCXCR4 neutralizing antibody or 6 to 12 mg/mL SDF-1antibody (both from R&D Systems) was added to blockCXCR4 or SDF-1, respectively. After 24 hours, the adherenttumor cells were quantified.
Cell migration assaysAll cell migration assays were conducted in a modified
Boyden chamber, that is, with 24-well plates in whichtranswell inserts (polycarbonate membrane insert with6.5-mm diameter and 8.0 mm pores; Corning) were placed.To assess endothelial cell migration, 2 to 5� 104 HUVECswere seeded into the upper compartment and incubated for24 hours. The nonmigrated cells on the top side of thesurface were removed. Themigrated cells on the bottom sideof the membrane were counted in 12 standardized fields. Totest endothelial effects on tumor cell migration, 1 � 105
HUVECs or HDMECs were seeded in the lower compart-ment of the chamber. When the cells had formed a mono-layer, they were treated with SDF-1 neutralizing antibodies(R&D Systems) before calcein-labeled MDA-MB 231 cellswere seeded into the upper compartment. Exposure tohypoxia was for 24 hours. Migrated cells in the lowercompartment were counted and photographs were taken.For transendothelial migration assays, HUVECs were cul-tured in the upper compartment which was precoated with0.1% gelatine. Leakiness of the endothelial monolayer wastested with FITC dextran before addition of tumor cells. Atotal of 2 � 104 calcein-labeled MDA-MB 231 cells wereadded to the upper compartment and incubated for 24hours. The migrated cells were quantified. Because only thetumor cells were labeled, this assay allows discrimination oftumor cells from migrated endothelial cells.
Statistical analysisData are expressed as the mean � SD Studies involving
more than 2 groups were analyzed by 1-way ANOVA
SDF-1 and CXCR4 Are Required for Tumor Cell Intravasation
www.aacrjournals.org Mol Cancer Res; 10(8) August 2012 1023
on April 13, 2017. © 2012 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst July 5, 2012; DOI: 10.1158/1541-7786.MCR-11-0498

coupled to post hoc comparison tests for multiple pairwisecomparisons. All other experiments were analyzed withan unpaired 2-tailed Student t test. In all figures � indicatesP < 0.05, whereas �� indicates P < 0.001.
ResultsSDF-1 and CXCR4 are expressed in microvessels inhuman breast carcinoma tissuesSDF-1 and CXCR4 are expressed in a number of tissues
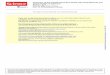
including bone marrow, lung, and lymph nodes. Becausethese factors were implicated in breast cancer progressionand metastasis, we regarded it as a prerequisite that they areindeed expressed in breast cancer samples. Immunohis-tochemistry showed that SDF-1 was expressed in the tumorcells in approximately 50% of 140 samples chosen from acollection of mammary tumor samples. We carried outdouble immunofluorescence by staining SDF-1 and vWFin 10 of these samples. A significant fraction of the cellsstained positive for SDF-1 but negative for vWF (Fig. 1A,case 1). These cells are most likely tumor cells. Interestingly,we detected SDF-1 also in the endothelium of microvessels.In 3 of 10 samples, SDF-1 was predominantly expressed inendothelial cells (Fig. 1A, case 2). Within the 140 samples,
CXCR4was expressed in 13 cases in the tumor cells (Fig. 1B,top). Interestingly, 23 samples showed necrotic areas, in 7 ofthese cases CXCR4 was apparently induced in the proximityof the necroses (Fig. 1B, center). Necroses are likely to behypoxic, which is in line with the idea that CXCR4 ishypoxia-inducible. From the 140 samples, a subgroup of24 samples showed CXCR4 expression in blood vessels(Fig. 1B, bottom). Which molecular events or pathwaysgovern CXCR4 expression in tumor cells as opposed toendothelial cells is unknown. There was no apparent cor-relation between CXCR4 and SDF-1 expression. In 13 of130 cases, tumor cells stained positive for CXCR7. Again,expression of CXCR7 did not correlate to SDF-1 or CXCR4expression. Tumor cells stained positive for CXCR4 andCXCR7 simultaneously in only 2 cases.
Autocrine SDF-1/CXCR4 signaling in hypoxia isrequired for ECs angiogenesisIn initial experiments, we observed that hypoxia induced
proliferation, tube formation, and migration in HUVECs(data not shown). To analyze the regulation of CXCR4under hypoxic conditions, we assessed the expression ofCXCR4 by FACS (Supplementary Fig. S1A). As expected,
Case 1 Case 2A B
H&E staining
VWF
SDF-1
DAPI
Merge
Figure 1. Vascular SDF-1 andCXCR4 immunoreactivity in humanbreast carcinoma tissues.A, immunofluorescent SDF-1staining in examples of humanprimary breast carcinoma tissue.Case 1 is a ductal carcinoma in situ(DCIS). Case 2 is an invasive ductalcarcinoma. Endothelial cells weredetected with an anti-vWFantibody (red), SDF-1 is stainedgreen. Cell nuclei are stained withDAPI (blue). B, CXCR4immunohistochemistry. Invasiveductal mammary carcinomasamples were stained for CXCR4,the signal was detected with asecondary antibody andDAB. Top,diffuse CXCR4 staining of thetumor cells; center, perinecroticinduction; bottom, endothelialstaining.
Jin et al.
Mol Cancer Res; 10(8) August 2012 Molecular Cancer Research1024
on April 13, 2017. © 2012 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst July 5, 2012; DOI: 10.1158/1541-7786.MCR-11-0498

CXCR4 increased markedly under hypoxia as comparedwith normoxia. Hypoxic induction was confirmed byWest-ern blotting (Supplementary Fig. S1B). As shown in Sup-plementary Fig. S1C, an intracellular SDF-1 upregulation byhypoxia was detected by FACS, which probably reflectsSDF-1 being processed before secretion because storage ofchemokines is uncommon. Importantly, secretion was alsoelevated as shown by ELISA in samples of the conditionedmedia (Supplementary Fig. S1D). In both, HUVEC andHDMEC, CXCR7 was neither detectable by Westernblotting nor by immunofluorescence in our hands. Next,we analyzed whether SDF-1 induced proliferation andangiogenesis. To do so, HUVECs were cultured in theabsence or presence of increasing concentrations of SDF-1 (1–100 ng/mL), CXCR4 neutralizing antibody (10 mg/mL), or Ig G control antibody. SDF-1 stimulated HUVECproliferation (Supplementary Fig. S2A) and tube formation(Supplementary Fig. S2B) in a dose-dependent manner.Anti-CXCR4 neutralizing antibodies inhibited proliferationand tube formation significantly.To assess the importance of CXCR4 in the endothelial
cells, HUVECs were transduced with pLKO.1-shRNA-CXCR4. Expression of CXCR4 was significantly reducedas compared with empty vector (Fig. 2A). Strikingly, reduc-tion of CXCR4 retarded HUVEC proliferation (Fig. 2A).We next tested whether CXCR4 also plays a role in migra-tion. As shown in Fig. 2B, HUVECs expressing shCXCR4exhibited attenuatedmigration under normoxia and hypoxiaas compared with controls. To show the involvement ofCXCR4 in capillary morphogenesis, we analyzed tube for-mation in HUVEC. After CXCR4 knockdown the tubenumbers were greatly reduced (Fig. 2C andD).We observedthat serum-deprived HUVECs died at day 3 in normoxia.However, many of the HUVECs survived when kept inhypoxia. An SDF-1 neutralizing antibody partly abrogatedthis effect in a dose-dependent manner (Fig. 2E and F).Furthermore, we carried out tube formation assays with theSDF-1 neutralizing antibody. We observed that blocking ofthe SDF-1 signal reduced tube numbers under normoxiaand under hypoxia (Fig. 2G and H).
HIF-1a and HIF-2a are essential for angiogenesisIn general, hypoxia significantly stimulates angiogenesis.
As in HUVEC, both HIF-a subunits were detectable andinduced by hypoxia in HDMEC. We also confirmed thatCXCR4 is hypoxia-inducible in HDMEC (SupplementaryFig. S4A). The relative contribution of HIF-1a as opposedtoHIF-2a to the transcriptional response of endothelial cellsin hypoxia is controversial (26, 27). To elucidate the role ofHIF-1a or HIF-2a in endothelial CXCR4 expression andangiogenesis, HUVECs were transduced with pLKO.1-shRNA-HIF-1a or pLKO.1-shRNA-HIF-2a alone or incombination. We then induced HIF activity chemicallyusing the prolyl hydroxylase inhibitor DMOG. AnalyzingCXCR4 expression by Western blotting, we showed thatboth HIF-1a and HIF-2a regulated CXCR4 expression inHUVECs (Fig. 3A). Apparently, both HIF-a subunitsaffected CXCR4 expression, although the precise contribu-
tion of each subunit is difficult to quantify because knock-down of 1 subunit affected also the other. HIF-1a and HIF-2a knockdown reduced proliferation (Fig. 3B). HIF-2aappeared to have a more pronounced effect. Apparently,there was some degree of synergy between the 2 factorsbecause double knockdown of HIF-1a andHIF-2a had themost striking effect. Similarly, migration of HUVECs afterHIF-1a or HIF-2a knockdown was reduced markedly,particularly after HIF-1a/2a double knockdown (Fig.3C). In addition, transduction of HIF-1a shRNA orHIF-2a shRNA abrogated hypoxic induction of tubeformation (Fig. 3D and E). Again, the most significantsuppression was achieved by combined HIF-1a/2aknockdown.
Hypoxia induces CXCR4 expression in MDA-MB 231and MCF7 and stimulates transendothelial migrationtoward an SDF-1 gradientHypoxia was shown previously to enhance CXCR4
expression in cultured tumor cells. Although CXCR4expression was readily detectable, we were unable to showexpression of CXCR7 in normoxia or hypoxia by Westernblotting or cell surface expression by immunofluorescence orFACS. To test regulation of CXCR4 by hypoxia, we exposed2 breast carcinoma cell lines, MDA-MB 231 (aggressivetype) and MCF7 (moderately aggressive) to hypoxia orDMOG treatment. In the case of MDA-MB 231, 24 hoursof hypoxia or DMOG led to a comparable increase inCXCR4 expression (Fig. 4A and B). However, 4 hours inhypoxia were insufficient to increase CXCR4 levels (Fig.4B). Similarly, CXCR4 expression of MCF7 showed amarked increase after treatment with DMOG or hypoxia(Supplementary Fig. S3). Knockdown of HIF-1a or HIF-2a led to decreased CXCR4 expression in MDA-MB 231(Fig. 4C) and MCF7 (Supplementary Fig. S3). Most effi-cient attenuation of CXCR4 expression was observed bycombined knockdown of HIF-1a/2a (Fig. 4C). Next, weobserved that hypoxia facilitated transendothelial migrationof MDA-MB 231 cells, in particular when an SDF-1gradient was present (Fig. 4D).
Collaboration of endothelial and breast carcinoma cellsmediates active adhesion and migration through theSDF-1/CXCR4 axisTo analyze whether hypoxia, CXCR4 and SDF-1 affect
the interaction of tumor cells with endothelial cells, a tumor-to-endothelium adhesion assay was developed. Hypoxiapromoted the adhesion ofMDA-MB 231 cells to HUVECs.This interaction was disrupted by selectively blockingCXCR4 in MDA-MB 231 cells, not by blocking CXCR4in HUVECs (Fig. 5A and B). This result indicated thatCXCR4 expression by tumor cells is important for theadhesive function. Next, calcein-labeled MDA-MB 231were cocultured with HUVECs in Matrigel for 24 hours.We observed that MDA-MB 231 were closely associatedwith the tubes formed by HUVECs. Again, blockingCXCR4 in MDA-MB 231 cells, but not in HUVECsinhibited this association (Fig. 5C and D). We carried out
SDF-1 and CXCR4 Are Required for Tumor Cell Intravasation
www.aacrjournals.org Mol Cancer Res; 10(8) August 2012 1025
on April 13, 2017. © 2012 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst July 5, 2012; DOI: 10.1158/1541-7786.MCR-11-0498

transendothelial migration assays to investigate whetherCXCR4 influences transendothelial migration.We observedthat CXCR4 knockdown inMDA-MB 231 cells, but not inHUVECs reduced transendothelial migration under nor-moxia and hypoxia (Fig. 5E and F). CXCR4 knockdownusing siRNA inMCF7 significantly reduced migration (Fig.5G and H). Because CXCR4 in HUVECs did not regulatethese processes we asked whether HUVEC-derived SDF-1regulates these steps. The presence of an SDF-1 neutralizingantibody attenuated the association of MDA-MB 231 aswell as MCF7 with HUVECs remarkably (Fig. 6A–D).
Importantly, adhesion of MDA-MB 231 to HDMECtubes was also inhibited by addition of an SDF-1 antibody(Supplementary Fig. S4B). When we blocked the activityof the SDF-1 secreted by HUVECs or HDMECs with anSDF-1 neutralizing antibody for 12 hours, migration ofMDA-MB 231 or MCF7 was substantially reduced inhypoxia (Fig. 6E and F, Supplementary Fig. S4C). Takentogether, these findings strongly indicate that SDF-1expressed in HUVECs or HDMECs promotes tumor-to-endothelium adhesion and transendothelial migrationof tumor cells.
CXCR4A C
B D
E F
G H
Actin
Control
Normoxia4
3
2
1
0
125
100
75
50
25
0
80
60
40
20
0
600
500
400
300
200
100
0
100
80
60
40
20
0
Cell
num
ber
(10
5)
Tube n
um
bers
Tube n
um
bers
Cell
num
bers
Mig
ration
(% o
f hypoxic
contr
ol)
Hypoxia
Normoxia
1% O2
Hypoxia
Normoxia Hypoxia
Normoxia
Hypoxia
3 d
d 0
NOX
HOX
NOX
HOX
d 4
shCXCR4
Control shCXCR4 Control
Control
Control
Control 3 6 12
Control 6 12
3 μg/mL SDF-1 Ab
SDF-1–neutralizing Ab (μg/mL)
SDF-1–neutralizing Ab (μg/mL)
Control 6 12
SDF-1–neutralizing Ab (μg/mL)
6 μg/mL SDF-1 Ab 12 μg/mL SDF-1 Ab
shCXCR4
shCXCR4ControlshCXCR4
Control shCXCR4
Control shCXCR4
Figure 2. Effects of hypoxia onHUVEC are dependent on SDF-1and CXCR4. A (top), CXCR4knockdown detected in whole-celllysates by Western blot; (Bottom)proliferation assay in HUVECs aftershRNA CXCR4 knockdown.Control cells were transduced withpLKO.l. HUVECs were cultured for4 days, viable cells were counted.The result gives means of 4independent experiments (� vs.normoxic control, P < 0.05; † vs.hypoxic control,P<0.05). B, in vitrotranswell migration assay:HUVECswereplated into the uppercompartment and incubated for 24hours. The migrated cells on thebottom of the surface werecounted in 12 standardized fields.Data are from 4 independentexperiments. C, tube formationassay after CXCR4 knockdown:HUVECs were seeded intoMatrigel-coated wells andincubated for 24 hours undernormoxia or hypoxia.Representative photographs oftube-like structures were taken. D,quantification of tube numbers inthe whole field, data representmeans of 3 independentexperiments. E, hypoxia survivalassay: 1 � 104 of serum-deprivedHUVECs were incubated inhypoxia in the presenceor absenceofSDF-1neutralizing antibodies for3 days. Normoxic serum-freeHUVEC served as additionalcontrol. Representativephotographs come from 4independent experiments. F, viableHUVECs were labeled with calceinand counted at day 3. The data givethe mean of 3 independentexperiments. G, tube formationassays in the presence or absenceof SDF-1 neutralizing antibodies.The results are from 4 independentexperiments. H, quantification oftube numbers in the whole fieldfrom 3 independent experiments.
Jin et al.
Mol Cancer Res; 10(8) August 2012 Molecular Cancer Research1026
on April 13, 2017. © 2012 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst July 5, 2012; DOI: 10.1158/1541-7786.MCR-11-0498

DiscussionIn this study, we show that the chemokine SDF-1 and its
cognate receptor CXCR4 are expressed in breast cancer invivo and analyze the effects of manipulating these proteins ina coculture model of mammary carcinoma and vascularendothelial cells. The evidence presented suggests thatactivation of the SDF-1/CXCR4 axis in a growing tumorcontributes to angiogenesis and the initial steps ofmetastasis.CXCR4 activation had surprisingly similar effects on tumorand endothelial cells: it induced proliferation and migration.In addition, SDF-1 binding to CXCR4 led to specific effectson endothelial and tumor cells: in endothelial cells, CXCR4stimulated tube formation which may point to proangio-genic activity in vivo, whereas in the tumor cells activation ofthis receptor resulted in adhesion to endothelial cells andtransendothelial migration.In addition to their hematopoietic activities, SDF-1 and
CXCR4 have been implicated in the regulation of angio-genesis in a number of (patho)physiological situations:the retina, the gastrointestinal tract, and the myocardiumafter infarction have been shown to rely on SDF-1 andCXCR4 for angiogenesis (28–30). Tumors also depend onangiogenesis because diffusion of oxygen and nutrients isinsufficient to sustain tumor growth beyond a size ofapproximately 2 mm. Therefore, the antiangiogenic anti-
body bevacizumab (Avastin) has been developed and is usedto antagonize the proangiogenic molecule VEGF. Withrespect to breast cancer, however, the use of this antibodyis discussed controversially (31). It is important to note thatour data do not simply suggest that the SDF-1/CXCR4 axisis proangiogenic. This is only one component which we haveobserved in vitro, as we have clearly shown that SDF-1stimulates proliferation, migration, and tube formation ofendothelial cells. In addition, and importantly, CXCR4signaling also fostered growth of cultured tumor cells (datanot shown) and the interaction between tumor and endo-thelial cells, which resulted in transendothelial migration.Transposed to the in vivo situation, these processes mayresult not only in angiogenesis but also in intravasation andmetastasis.The in vivo sequence of events which we propose and
which we have followed meticulously in vitro is as follows:rapid proliferation of tumor cells leads to areas of hypoxiawithin the primary tumor. Hypoxia then activates theubiquitous transcription factor HIF, which results in acti-vation of an array of HIF target genes including SDF-1 andCXCR4 which contribute to tumor cell migration andadhesion to endothelial cells. Eventually, activation of theSDF-1/CXCR4 axis results in transendothelial migration.Within this scenario, angiogenesis provides the escape route
Figure 3. HIF-1a and HIF-2acontribute to the effects of hypoxiaon HUVECs. Endothelial cells weretransduced with pLKO.1-shRNA-HIF-1a or pLKO.1-shRNA-HIF-2a.A, CXCR4 expression in HUVECsafter HIF-1a or HIF-2a knockdown.HUVECs were treated with 1 mmol/LDMOG for 4 hours. Data represent atleast 3 independent experiments. B,hypoxia survival assay: HUVEC werecultured in serum-free ECGM underhypoxia for 3 days. Representativephotographs from 2 independentexperiments are shown. C, HUVECmigration assay after HIF-1a or HIF-2a knockdown. Columns giveaverage of 3 independentexperiments. D, tube formationassay: HUVECs were exposed tohypoxia for 24 hours. Representativedata from 3 independentexperiments are shown. E,quantification of tube numbersshows means of 3 independentexperiments.
HIF-1α
HIF-2α
α-Tubulin
A
C
B
D E
140
120
100
80
60
40
20
0
80
60
40
20
0
Tube n
um
bers
Mig
ratio
n (
% o
f co
ntr
ol)
1% O2 (serum-free)
1% O2
ControlControl
shHIF-1αshHIF-1α
shHIF-1αshHIF-2α
shHIF-1αshHIF-2α
shHIF-2α
shHIF-2α
Control shHIF-1α
shHIF-1α+
shHIF-2αshHIF-2α
Control shHIF-1α shHIF-1αshHIF-2α
shHIF-2α
Control shHIF-1α shHIF-1αshHIF-2α
shHIF-2α
CXCR4
SDF-1 and CXCR4 Are Required for Tumor Cell Intravasation
www.aacrjournals.org Mol Cancer Res; 10(8) August 2012 1027
on April 13, 2017. © 2012 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst July 5, 2012; DOI: 10.1158/1541-7786.MCR-11-0498

for tumor cells. The secretion of SDF-1 by endothelial cells(potentially also by tumor and stroma cells) and CXCR4expression by the tumor cells results in entering of the tumorcells into the blood vessels. In principle, CXCR4 expressionof tumor cells also favors extravasation in tissues where SDF-1 is expressed. Indeed, this effect has been reportedpreviously (14, 32) and is regarded as a reason for theorgan-specific pattern of metastasis seen in many cases ofbreast cancer. Because our experiments focused on endo-thelial SDF-1 secretion and also showed effects on tumor cellmigration, our data suggest that in particular in hypoxicregions of the tumor an SDF-1 gradient develops, whichattracts tumor cells to the blood vessels and results in tumorcell intravasation at the site of primary tumor growth.Hypoxia is an important feature in many solid tumors.
Effects of hypoxia are associatedwith several aspects of tumorbiology including angiogenesis, metastasis as well as resis-tance to radiation and chemotherapy (33–35). In a hypoxicenvironment, angiogenesis is induced by a number of genesincluding vascular endothelial growth factor-A (VEGF-A),VEGFR1, Angiopoietin-2 (Ang-2), and others (36, 37).Hypoxia also stimulates expression of CXCR4 in differentcell types. We observed that both SDF-1 and CXCR4 inHUVECs are hypoxia-inducible. The key regulators of thecellular responses to low oxygen concentrations are twodistinct a-subunits termed HIF-1a and HIF-2a. Theindividual contributions of HIF-1a and HIF-2a to angio-
genesis are under debate. We observed that HUVECsexpressed both HIF-1a and HIF-2a when cultured underhypoxic conditions. In our hands, both HIF-1 and HIF-2contributed to migration and tube formation in response tohypoxia because inhibition of either HIF-1a or HIF-2aprovoked partial loss of induction of CXCR4 in response tohypoxia. Previously a reciprocal interaction between the twoa-subunits has been reported in tumor cells of renal origin(38). We could not observe this effect in our cells butfrequently noticed that inhibition of 1 a-subunit led to aparallel decrease of the other. The molecular basis of thisdiscrepancy is unclear. Because the effects of HIF-1a andHIF-2a knockdown were additive, we believe that bothsubunits contribute to CXCR4 induction by hypoxia.Importantly, we have showed by immunohistochemistry
that SDF-1 and CXCR4 are expressed in a significantfraction of mammary tumors. We observed that SDF-1 isexpressed in approximately 50% of our samples. Withinthese samples SDF-1wasmainly expressed by the tumor cellsbut frequently also in endothelial cells as showed by colo-calization with vWF. SDF-1 has also been detected inendothelial cells of human breast cancer samples in aprevious study (5, 20). Interestingly, none of our tumorcell lines (MDA MB231 and MCF-7) expressed SDF-1.However, CXCR4 expression in breast cancer samples wasmore difficult to assess than anticipated. Several commercialantibodies led to nuclear staining of almost all tumor cells,
HIF-1α
α-Tubulin
A
C
B
D
200
150
100
50
0
250
200
150
100
50
0
35.42% 70.49%
4 h 4 h 0.1 0.5 1 mmol/L
DMOG
DMOG 1 mmol/L
−SDF-1 +SDF-1 −SDF-1 +SDF-1
101 102 103
PE-A104 105 101 102 103
PE-A104 105
Ce
lls
Ce
lls
O2 (% )
21% O2 1% O2
21 21 21 211
Control shHIF-1α1 2 1 2
shHIF-1αshHIF-2α
shHIF-2α
CXCR4
HIF-1α
HIF-2α
α-Tubulin
CXCR4
CXCR4
Figure 4. CXCR4 is induced byhypoxia in breast carcinoma cells.A, CXCR4 expression on the cellsurface in MDA-MB 231 cells after24 hours of exposure to hypoxia orto DMOG as detected by flowcytometry. Representative data areshown from at least 3 independentexperiments. B, DMOG dose–response curve, incubations werefor 4 hours. C, MDA-MB 231 cellsexpressing shHIF-1a, shHIF-2a, orboth. The cells were treated with1 mmol/L DMOG for 4 hours. D,transendothelial migration assay:MDA-MB231 cells were added to aHUVEC monolayer in the uppercompartment in the presence orabsence of SDF-1 and incubatedfor 24 hours. Representativephotographs of MDA-MB 231 inthe lower chamber from 3independent experiments areshown.
Jin et al.
Mol Cancer Res; 10(8) August 2012 Molecular Cancer Research1028
on April 13, 2017. © 2012 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst July 5, 2012; DOI: 10.1158/1541-7786.MCR-11-0498

which we regarded as unreliable. Using another antibodywhich has been characterized extensively very recently (39),we observed that nearly 10% of the tumors we analyzed werepositive for CXCR4. From our data it is indeed conceivablethat these patients would benefit from antagonizing CXCR4signaling. Of note, very recently an in vitro study waspublished which implies that the effects of CXCR4 andCXCR7 inhibition may act synergistically (40). Important-ly, CXCR4 andCXCR7positive tumors can be identified ona routine basis by immunohistochemistry. For some tumorentities it was reported that the chemokine receptor termedCXCR7 can also bind SDF-1 and thus modulate CXCR4signaling. Because expression of both receptors was onlydetectable in 2 out of approximately 130 cases in our cohort,functional interaction is of course possible but seems to be arare event. However, CXCR7 could also transmit prolifer-
ative or angiogenic signals. Overall, our data suggest thatinhibition of CXCR4 may reduce growth and metastasis inbreast carcinoma. Interestingly, a low molecular weightcompound termed AMD3100, which antagonizes CXCR4function is already in clinical use to mobilize hematopoieticstem cells and has been reported to have a tolerable side effectprofile (41).Collectively, we discovered a crosstalk mechanism
between breast carcinoma and endothelial cells. Hypoxia-dependent induction of the SDF-1/CXCR4 pathway linksmetabolism to tumor angiogenesis and active transendothe-lial migration. Inhibition of the activated SDF-1/CXCR4signaling pathway in a primary tumor environment maytherefore attenuate angiogenesis and carcinoma intravasa-tion. Our data are primarily applicable to breast cancerbecause we have analyzed cultured breast carcinoma cell
Figure 5. CXCR4 in breastcarcinoma cells is responsible fortumor-to-endothelium adhesion andtransendothelial migration. MDA-MB231 and HUVEC were transduced bypLKO.1-shRNA-CXCR4 or pLKO.l.A, the adhesion of MDA-MB 231(control or shCXCR4) cells to aHUVEC monolayer (control orshCXCR4). HUVECs were exposedto normoxia or hypoxia for 12 hoursbefore the plating of calcein-labeledMDA-MB 231 cells. Nonadherentcells were thoroughly washed offafter 2 hours. Representativephotographs from 4 independentexperiments. B, quantification ofadherentMDA-MB231counted in 12continuous fields. C, adhesion ofcalcein labeled MDA-MB 231(control or shCXCR4) to HUVEC(control or treated with a CXCR4neutralizing antibody before theexperiment). D, quantification ofMDA-MB 231 associated withendothelial tubes. E, transendothelialmigration assay: MDA-MB 231(control or shCXCR4) cells wereadded to HUVECs (control orshCXCR4) on a transwell insert, andexposed to normoxia or hypoxia for24 hours. F, quantification ofmigrated MDA-MB 231. G, H,transendothelial migration assaywith MCF-7 cells (control or siRNA)carried out as with MDA-MB 231 inE and F.
MDA-MB 231
MDA-MB 231
MCF7
A
C
B
D
E F
G H
1,000
800
600
400
200
0
500
400
300
200
100
0
240
160
80
0
80
60
40
20
0
Normoxia
Hypoxia
Normoxia
Mig
rato
ry c
ells
Mig
rato
ry c
ells
Hypoxia
NOX
Adhere
nt
cells
Tum
or
cells
ssocia
ted w
ith tube (
Nr)
HOX
NOX
HOX
MDA-MB 231 Control shCXCR4 Control shCXCR4
HUVEC
MDA-MB 231
HUVEC
MDA-MB 231
HUVEC
MDA-MB 231
HUVEC
Control
Control
Control
shCXCR4
Control
CXCR4 Ab
shCXCR4
CXCR4Ab
Control
shCXCR4
Control
shCXCR4
Control shCXCR4 Control shCXCR4
Control shCXCR4Control shCXCR4
Control
Control
shCXCR4
CXCR4 siRNAControl CXCR4 siRNA
Control shCXCR4
Control shCXCR4Control shCXCR4
Control shCXCR4 Control shCXCR4
Control CXCR4AbControl CXCR4Ab
Control shCXCR4 Control shCXCR4
Control shCXCR4Control shCXCR4
SDF-1 and CXCR4 Are Required for Tumor Cell Intravasation
www.aacrjournals.org Mol Cancer Res; 10(8) August 2012 1029
on April 13, 2017. © 2012 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst July 5, 2012; DOI: 10.1158/1541-7786.MCR-11-0498

lines and confirmed SDF-1 and CXCR4 expression insamples of mammary carcinoma. Because importance ofCXCR4 and SDF-1 has already been proposed for tumors ofother origin, our data may also be relevant for other tumortypes.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: F. Jin, F. Otterbach, E. MetzenDevelopment of methodology: F. Jin, U. BrockmeierAcquisition of data (provided animals, acquired and managed patients, providedfacilities, etc.): F. Jin, F. OtterbachAnalysis and interpretation of data (e.g., statistical analysis, biostatistics, compu-tational analysis): F. Jin, F. OtterbachWriting, review, and/or revision of the manuscript: F. Jin, U. Brockmeier, F.Otterbach, E. Metzen
Administrative, technical, or material support (i.e., reporting or organizing data,constructing databases): U. Brockmeier, F. OtterbachStudy supervision: F. Otterbach, E. Metzen
AcknowledgmentsThe authors thank Kirsten Goepelt, Gabriele Ladwig, and Frank Splettstoesser for
excellent technical assistance and Petra Liebig for help with the manuscript. Theauthors are also grateful to Professor Dirk Hermann, Department of Neurology, forcontinuous support and many helpful discussions. Professor Stefan Schulz, Depart-ment of Pharmacology, Jena, Germany, provided us with the invaluable CXCR4antibody which is greatly appreciated.
Grant SupportF. Jin was supported financially by the Deutscher Akademischer Austauschdienst
(DAAD) and the Dr. Werner Jackst€adt-Stiftung.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be herebymarked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
Received October 14, 2011; revised June 4, 2012; accepted June 25, 2012;published OnlineFirst July 9, 2012.
References1. Jemal A, Siegel R, Xu J, Ward E. Cancer statistics, 2010. CA Cancer
J Clin 2010;60:277–300.2. Christofori G. Cancer: division of labour. Nature 2007;446:735–6.3. Podsypanina K, Du YC, Jechlinger M, Beverly LJ, Hambardzumyan D,
Varmus H. Seeding and propagation of untransformed mouse mam-mary cells in the lung. Science 2008;321:1841–4.
4. NakagawaH, Liyanarachchi S, Davuluri RV, Auer H,Martin EWJr, de laChapelle A, et al. Role of cancer-associated stromal fibroblasts in
metastatic colon cancer to the liver and their expression profiles.Oncogene 2004;23:7366–77.
5. Orimo A, Gupta PB, Sgroi DC, Arenzana-Seisdedos F, Delaunay T,Naeem R, et al. Stromal fibroblasts present in invasive human breastcarcinomas promote tumor growth and angiogenesis through elevat-ed SDF-1/CXCL12 secretion. Cell 2005;121:335–48.
6. Dunwoodie SL. The role of hypoxia in development of the mammalianembryo. Dev Cell 2009;17:755–73.
MDA-MB 231
MDA-MB 231
MCF7
A
C
B
D
E F
150
100
50
0
120
80
40
0
160
120
80
40
0
Mig
rato
ry 2
31 c
ells
231 a
sssocia
ted w
ith t
ube
MC
f7 a
sssocia
ted w
ith t
ube
Control
SDF-1–neutralizing Ab
6 μg/mL 12 μg/mL
Control
Control
SDF-1–neutralizing Ab
6 μg/mL 12 μg/mL
Control
SDF-1–neutralizing Ab
6 μg/mL 12 μg/mL
Control
SDF-1–neutralizing Ab
6 μg/mL 12 μg/mL
12 μg/mL SDF-1 AbHUVECsControl 12 μg/mL SDF-1 AbHUVECs
Figure 6. Adhesion andtransendothelial migration ofbreast carcinoma cells in thepresence or absence ofneutralizing SDF-1 antibodies. A,B, adhesion of calcein-labeledMDA-MB 231 to endothelial tubesin the presence or absence of SDF-1 neutralizing antibody. A,representative photographs; B,quantification of MDA-MB 231;C,D, adhesion of calcein-labeledMCF7 to endothelial tubes in thepresence or absence of SDF-1neutralizing antibody after 24 hoursof coculture; C, representativephotographs; D, quantification ofMCF7; E, F, migration assay:calcein-labeled MDA-MB 231migrated toward a HUVECmonolayer in the presence orabsence of neutralizing SDF-1antibodies in the bottom chamber;E, representative photographs;and F, quantification of migratedMDA-MB 231.
Jin et al.
Mol Cancer Res; 10(8) August 2012 Molecular Cancer Research1030
on April 13, 2017. © 2012 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst July 5, 2012; DOI: 10.1158/1541-7786.MCR-11-0498

7. Egawa T, Kawabata K, Kawamoto H, Amada K, Okamoto R, Fujii N,et al. The earliest stages of B cell development require a chemokinestromal cell-derived factor/pre-Bcell growth-stimulating factor. Immu-nity 2001;15:323–34.
8. Salvucci O, Yao L, Villalba S, Sajewicz A, Pittaluga S, Tosato G.Regulation of endothelial cell branching morphogenesis by endoge-nous chemokine stromal-derived factor-1. Blood 2002;99:2703–11.
9. Feng Y, Broder CC, Kennedy PE, Berger EA. HIV-1 entry cofactor:functional cDNA cloning of a seven-transmembrane, G protein-cou-pled receptor. Science 1996;272:872–7.
10. Broxmeyer HE, Orschell CM, Clapp DW, Hangoc G, Cooper S, PlettPA, et al. Rapidmobilization of murine and human hematopoietic stemand progenitor cells with AMD3100, a CXCR4 antagonist. J Exp Med2005;201:1307–18.
11. Zagzag D, Krishnamachary B, Yee H, OkuyamaH, Chiriboga L, Ali MA,et al. Stromal cell-derived factor-1alpha and CXCR4 expression inhemangioblastoma and clear cell-renal cell carcinoma: von hippel-lindau loss-of-function induces expression of a ligand and its receptor.Cancer Res 2005;65:6178–88.
12. Staller P, Sulitkova J, Lisztwan J, Moch H, Oakeley EJ, Krek W.Chemokine receptor CXCR4 downregulated by von hippel-lindautumour suppressor pVHL. Nature 2003;425:307–11.
13. Muller A, Homey B, Soto H, Ge N, Catron D, Buchanan ME, et al.Involvement of chemokine receptors in breast cancer metastasis.Nature 2001;410:50–6.
14. Kang Y, Siegel PM, Shu W, Drobnjak M, Kakonen SM, Cordon-CardoC, et al. A multigenic program mediating breast cancer metastasis tobone. Cancer Cell 2003;3:537–49.
15. Li YM, Pan Y, Wei Y, Cheng X, Zhou BP, Tan M, et al. Upregulation ofCXCR4 is essential for HER2-mediated tumor metastasis. Cancer Cell2004;6:459–69.
16. vandeVijverMJ,HeYD, van't Veer LJ,DaiH,Hart AA, Voskuil DW, et al.A gene-expression signature as apredictor of survival in breast cancer.N Engl J Med 2002;347:1999–2009.
17. Zhao X, Rodland EA, Sorlie T, Naume B, Langerod A, Frigessi A, et al.Combining gene signatures improves prediction of breast cancersurvival. PLoS One 2011;6:e17845.
18. Welford AF, Biziato D, Coffelt SB, Nucera S, Fisher M, Pucci F, et al.TIE2-expressing macrophages limit the therapeutic efficacy of thevascular-disrupting agent combretastatin A4 phosphate in mice.J Clin Invest 2011;121:1969–73.
19. Wendt MK, Cooper AN, Dwinell MB. Epigenetic silencing of CXCL12increases the metastatic potential of mammary carcinoma cells.Oncogene 2008;27:1461–71.
20. Mirisola V, Zuccarino A, Bachmeier BE, Sormani MP, Falter J, NerlichA, et al.CXCL12/SDF1expressionbybreast cancers is an independentprognostic marker of disease-free and overall survival. Eur J Cancer2009;45:2579–87.
21. Kopfstein L, Christofori G. Metastasis: cell-autonomous mechanismsversus contributions by the tumor microenvironment. Cell Mol Life Sci2006;63:449–68.
22. Ribatti D, Nico B, Vacca A. Importance of the bone marrow microen-vironment in inducing the angiogenic response in multiple myeloma.Oncogene 2006;25:4257–66.
23. Kitadai Y, Sasaki T, Kuwai T, Nakamura T, Bucana CD, Hamilton SR,et al. Expression of activated platelet-derived growth factor receptor instromal cells of human colon carcinomas is associatedwithmetastaticpotential. Int J Cancer 2006;119:2567–74.
24. Otterbach F, Callies R, Adamzik M, Kimmig R, Siffert W, Schmid KW,et al. Aquaporin 1 (AQP1) expression is a novel characteristic feature ofa particularly aggressive subgroup of basal-like breast carcinomas.Breast Cancer Res Treat 2010;120:67–76.
25. Brockmeier U, Platzek C, Schneider K, Patak P, Bernardini A, FandreyJ, et al. The function of hypoxia-inducible factor (HIF) is independent ofthe endoplasmic reticulum protein OS-9. PLoS One 2011;6:e19151.
26. Skuli N, Liu L, Runge A, Wang T, Yuan L, Patel S, et al. Endothelialdeletion of hypoxia-inducible factor-2alpha (HIF-2alpha) alters vascu-lar function and tumor angiogenesis. Blood 2009;114:469–77.
27. Vaupel P, Schlenger K, Knoop C, Hockel M. Oxygenation of humantumors: evaluation of tissue oxygen distribution in breast cancers bycomputerized O2 tension measurements. Cancer Res 1991;51:3316–22.
28. Strasser GA, Kaminker JS, Tessier-Lavigne M. Microarray analysis ofretinal endothelial tip cells identifies CXCR4 as a mediator of tip cellmorphology and branching. Blood 2010;115:5102–10.
29. Tachibana K, Hirota S, Iizasa H, Yoshida H, Kawabata K, Kataoka Y,et al. The chemokine receptor CXCR4 is essential for vascularization ofthe gastrointestinal tract. Nature 1998;393:591–4.
30. Dai S, Yuan F, Mu J, Li C, Chen N, Guo S, et al. Chronic AMD3100antagonism of SDF-1alpha-CXCR4 exacerbates cardiac dysfunctionand remodeling aftermyocardial infarction. JMol Cell Cardiol 2010;49:587–97.
31. Lenzer J. FDA committee votes to withdraw bevacizumab for breastcancer. BMJ 2011;343:d4244.
32. Smith MC, Luker KE, Garbow JR, Prior JL, Jackson E, Piwnica-WormsD, et al. CXCR4 regulates growthof both primary andmetastatic breastcancer. Cancer Res 2004;64:8604–12.
33. Pugh CW, Ratcliffe PJ. The von hippel-lindau tumor suppressor,hypoxia-inducible factor-1 (HIF-1) degradation, and cancer pathogen-esis. Semin Cancer Biol 2003;13:83–9.
34. Yotnda P, Wu D, Swanson AM. Hypoxic tumors and their effect onimmune cells and cancer therapy. Methods Mol Biol 2010;651:1–29.
35. Dewhirst MW, Cao Y, Li CY, Moeller B. Exploring the role of HIF-1 inearly angiogenesis and response to radiotherapy. Radiother Oncol2007;83:249–55.
36. Schioppa T, Uranchimeg B, Saccani A, Biswas SK, Doni A, RapisardaA, et al. Regulation of the chemokine receptor CXCR4 by hypoxia.J Exp Med 2003;198:1391–402.
37. Rey S, Semenza GL. Hypoxia-inducible factor-1-dependent mechan-isms of vascularization and vascular remodelling. Cardiovasc Res2010;86:236–42.
38. Raval RR, Lau KW, Tran MG, Sowter HM, Mandriota SJ, Li JL, et al.Contrasting properties of hypoxia-inducible factor 1 (HIF-1) and HIF-2in von hippel-lindau-associated renal cell carcinoma. Mol Cell Biol2005;25:5675–86.
39. Fischer T, Nagel F, Jacobs S, Stumm R, Schulz S. Reassessment ofCXCR4 chemokine receptor expression in human normal and neo-plastic tissues using the novel rabbit monoclonal antibody UMB-2.PLoS One 2008;3:e4069.
40. Hernandez L, MagalhaesM, Coniglio S, Condeelis J, Segall J. Oppos-ing roles of CXCR4 and CXCR7 in breast cancer metastasis. BreastCancer Res 2011;13:R128.
41. Devine SM, Vij R, Rettig M, Todt L, McGlauchlen K, Fisher N, et al.Rapid mobilization of functional donor hematopoietic cells without G-CSF using AMD3100, an antagonist of the CXCR4/SDF-1 interaction.Blood 2008;112:990–8.
SDF-1 and CXCR4 Are Required for Tumor Cell Intravasation
www.aacrjournals.org Mol Cancer Res; 10(8) August 2012 1031
on April 13, 2017. © 2012 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst July 5, 2012; DOI: 10.1158/1541-7786.MCR-11-0498

2012;10:1021-1031. Published OnlineFirst July 5, 2012.Mol Cancer Res Fengyan Jin, Ulf Brockmeier, Friedrich Otterbach, et al. Are Required for Tumor Cell IntravasationModel: Hypoxia-Induced Endothelial SDF-1 and Tumor Cell CXCR4 New Insight into the SDF-1/CXCR4 Axis in a Breast Carcinoma
Updated version
10.1158/1541-7786.MCR-11-0498doi:
Access the most recent version of this article at:
Material
Supplementary
http://mcr.aacrjournals.org/content/suppl/2012/07/05/1541-7786.MCR-11-0498.DC1
Access the most recent supplemental material at:
Cited articles
http://mcr.aacrjournals.org/content/10/8/1021.full.html#ref-list-1
This article cites 41 articles, 14 of which you can access for free at:
Citing articles
/content/10/8/1021.full.html#related-urls
This article has been cited by 1 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications Department at
on April 13, 2017. © 2012 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst July 5, 2012; DOI: 10.1158/1541-7786.MCR-11-0498