Embed Size (px)
Citation preview

INTERNATIONAL UNION OF GEOLOGICAS SCIENCES SUBCOMISSION ON NEOGENE STRATIGRAPHY
FOURTH CONGRESS Buenos Aires (Argentina), December, 2008
THE ATLANTIC NEOGENE IN THE INTERNATIONAL YEAR OF THE EARTH PLANET
ABSTRACTS RESÚMENES
J.A. GONZÁLEZ DELGADO & J. CIVIS (eds.)

Cover photo: c) J.A. González Delgado y J. Civis (eds.), Facultad de Ciencias, Universidad de Salamanca (España) Depósito Legal: S.1498_2008

PRESENTACIÓN
El grupo R.C.A.N.S. (Regional Committee on Atlantic Neogene Stratigrpahy), dependiente de la IUGS (Internacional Union of Geological Sciences - Subcomission on Neogene Stratigraphy) tiene por objeto congregar a científicos que desarrollan su investigación en el Neógeno del dominio Atlántico, y ya lleva una andadura de 16 años. En Octubre de 1992, celebró su primer Congreso en la ciudad de Lisboa (Portugal); en 1997 tuvo lugar, en Salamanca (España), el segundo encuentro, y en año 2003, en Tetouan (Marruecos), el tercero. Se han celebrado, además, diversos interim-colloquium en Tours (Francia, 1995) y en Salamanca (2002). La celebración del IV Congreso en Buenos Aires (Argentina) supone un importante hito ya que por primera vez el congreso de los neogenistas atlánticos tiene lugar en la parte occidental del océano.
Este Congreso se ha organizado conjuntamente por instituciones argentinas, uruguayas
y españolas, en el marco del Año Internacional del Planeta Tierra. Por parte argentina participan el Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, la Universidad de Buenos Aires, la Universidad Nacional del Sur y la Universidad Nacional de La Plata; por parte de Uruguay participa la Facultad de Ciencias de la Universidad de la República y por parte española, la Universidad de Salamanca y la Junta de Castilla y León. Además de las sesiones científicas, comunicaciones orales, paneles, debates y coordinación de proyectos comunes, se llevará a cabo una excursión geológica al extraordinario registro paleontológico del Mioceno marino de Península Valdés.
En este Libro se recogen los resúmenes de las comunicaciones orales y de los paneles
que serán objeto de exposición y debate, así como las conferencias invitadas. Desde estas líneas expresamos nuestro agradecimiento a todas las instituciones colaboradoras, al Comité organizador del Congreso y al Comité científico que ha revisado los trabajos presentados.

COMITÉ ORGANIZADOR:
Presidente
Jorge Civis (Universidad de Salamanca, España)
Vicepresidente Ester Farinati (Universidad Nacional del Sur, Bahía Blanca, Argentina)
Secretario General José Angel González Delgado (Universidad de Salamanca, España)
Coordinadores
Viviana Barrera (Museo Nacional de Ciencias Naturales “Bernardino Rivadavia” Buenos Aires, Argentina)
Claudia J. del Río (Museo Nacional de Ciencias Naturales “Bernardino Rivadavia” Buenos Aires, Argentina)
Roberto Scasso (Universidad de Buenos Aires, Argentina) Marina Aguirre (Universidad Nacional de La Plata, Argentina)
Sergio Martínez (Universidad de la República, Uruguay) Salvador Mulero (Junta de Castilla y León, España)
COMITÉ CIENTÍFICO:

Aguirre, Julio Dpto. Estratigrafía y Paleontología. Universidad de Granada (España) Alonso Gavilán, Gaspar Dpto. Geología (Estratigrafía). Universidad de Salamanca (España) Antunes, Miguel T. Academia de las Ciencias. Lisboa (Portugal) Armenteros, Ildefonso Dpto. Geología (Estratigrafía). Universidad de Salamanca (España) Barreda, Viviana Museo Argentino de Ciencias Naturales Bernardino Rivadavia, Buenos Aires, (Argentina) Civis, Jorge Dpto. Geología (Paleontología). Universidad de Salamanca (España) Dabrio, Cristino J. Dpto. Estratigrafía, Facultad de Geología, Universidad Complutense, Madrid (España) Del Río, Claudia J. Museo Argentino de Ciencias Naturales Bernardino Rivadavia, Buenos Aires (Argentina) Farinati, Ester Dpto. Geología, Universidad Nacional del Sur, Bahía Blanca (Argentina) Flores, José Abel Dpto. Geología (Paleontología). Universidad de Salamanca (España) Fucks, Enrique Fac. de Ciencias Nat. y Museo. UNLP, La Plata (Argentina) González Delgado, José Angel Dpto. Geología (Paleontología). Universidad de Salamanca (España) Goy Goy, José Luis Dpto. Geología (Geodinámica Externa). Universidad de Salamanca (España) Jiménez Fuentes, Emiliano Dpto. Geología. Universidad de Salamanca (España) Legoinha, Paulo Centro de Estratigrafía y Paleobiologia, Universidade Nova de Lisboa (Portugal) Martínez, Sergio Universidad de la República, Montevideo (Uruguay) Pais, Joao Centro de Estratigrafía y Paleobiologia, Universidade Nova de Lisboa (Portugal) Rodríguez Lázaro, Julio Dpto. Geología, Universidad del País Vasco (España)

Ruiz, Francisco Dpto. Geodinámica y Paleontología. Universidad de Huelva (España) Sierro, F. Javier Dpto. Geología (Paleontología). Universidad de Salamanca (España) Tonni, Eduardo P. División Paleontología Vertebrados, Museo de La Plata, 1900-La Plata (Argentina) Valle, María Dpto. Geología (Paleontología). Universidad de Salamanca (España)

INDICE CONFERENCIAS: página ZÁRATE, M. El registro estratigráfico continental del Neógeno del centro de Argentina………. 1 CIVIS, J. Ciencias de la Tierra para la Sociedad: hacia un Planeta sostenible………………. 3 RESÚMENES: AGUIRRE, M.L. y FARINATI, E. Moluscos del Cuaternario marino de Argentina: herramientas útiles para interpretaciones paleoambientales y paleoclimáticas………………………… 6 AGUIRRE, M.L., FARINATI, E., PÉREZ, S.I. y PISANO, F. Problemática de la variabilidad morfológica en moluscos del Cuaternario marino de Argentina: implicancias paleoambientales………………… 8 AGUIRRE, M.L., RICHIANO, S., ÁLVAREZ, M.F., FUCKS, E., FARINATI, E. y EASTOE, CH. Gastropoda y Bivalvia del Cuaternario del sector costero Bahía Lángara - Puerto Deseado (Santa Cruz, Patagonia, Argentina): interpretaciones paleoambientales…..... 10 ALIOTTA, S., FARINATI, E. y SPAGNUOLO, J.O. Caracteres distintivos de los depósitos marinos holocenos del estuario de Bahía Blanca……………………………………………………………………… 11 ANGELOZZI, G.N., RONCHI, D.I. y PÉREZ PANERA, J.P. Asociaciones de nanofósiles calcáreos y foraminíferos del Mioceno temprano del Atlántico sur occidental…………………………………………………………. 12 ARAMAYO, S.A. Registros paleoicnológícos de la Formación Río Negro, facies continental (Mioceno tardío – Plioceno temprano), costa atlántica de la Provincia de Río Negro, Argentina………………………………………………………………… 14

ARMENTEROS, I., GONZÁLEZ DELGADO, J.A., CIVIS, J., ALONSO GAVILÁN, G. & DABRIO, C.J. Pliocene shoreline, high-energy, non-tropical coarse biocalcarenites in the Atlantic central region of Maroc………………………………………………… 16 CIVIS, J., ALONSO GAVILÁN, G., GONZÁLEZ DELGADO, J.A., RUIZ MUÑOZ, F. y GONZÁLEZ REGALADO, M.L. Tránsito El Mio-Plioceno en el sondeo de Montemayor (Neógeno de Huelva, Oeste de la Cuenca del Guadalquivir, España). Diagrafías y Foraminíferos bentónicos…… 18 CRAWFORD, R.S., CASADÍO, S., FELDMANN, R., GRIFFIN, M. & SCWEITZER, C.E. The ecologic and taphonomic effects of a Miocene volcanic ashfall on Atlantic inner-shelf communities………………………………………………………………… 20 ERRA, G., FERRERO, L., FUCKS, E., AGUIRRE, M.L y SCHNACK, E. Evidencias paleoambientales del Pleistoceno Marino tardío de la Formación Pascua basadas en moluscos y microfósiles……………………………………………………… 21 FARINATI, E.A. y FERNÁNDEZ, A.L. Estudios biométricos en ostras del Neógeno marino de Patagonia, Argentina……………… 22 GARCÍA, E.X.M., ANTUNES, M.T., BALBINO, A.C., CIVIS, J., RUIZ, F. y RICO GARCÍA, A. Las familias Scyliorhinidae, Triakidae, Hemigaleidae y Carcharhinidae (Neoselachii, Galeomorphii) del Plioceno inferior de la Fm. Arenas de Huelva, España…… 23 GARCÍA CHAPORI, N. & LAPRIDA, C. Plio-Pleistocene SST reconstruction: essay for the Southwest South Atlantic……………… 25 GONZÁLEZ, C.A., AGUIRRE, M.L., MIQUEL, S.E., KRÖHLING, D., ZUKOL, A. y BREA, M. Malacofauna continental holocena y paleoambientes en Villa Valle María (Departamento Diamante, Entre Ríos, Argentina)……………………………………. 27 GONZÁLEZ-DELGADO, J.A., BEN MOUSSA, A., CIVIS, J. & RICO GARCÍA, A. Malacology of the Pliocene Atlantic basins of Northern Morocco………………………….. 28 GORDILLO, S. & ISLA, F.I. Quaternary mollusk assemblages from the northern Atlantic coast of Tierra del Fuego (southern South America): taphonomy and paleoecology……………………….. 29

GUTIÉRREZ TELLEZ, B., SCHILLIZZI, R., LUNA, L., CABALLERO, N. y NASS, Y. Cambios paleoambientales en el Cuaternario de la costa Norte del Río Negro (paraje Playa Chica), provincia de Buenos Aires, Argentina………………………………………………. 30 LEGOINHA, P. The Serravallian-Tortonian boundary in the Lower Tagus Basin (Portugal) and the new GSSP of the Tortonian Stage……………………………………………………. 31 LUENGO, J., ARMENTEROS, I., MONTES, M., NOZAL, F., CORROCHANO, A. LÓPEZ OLMEDO, F. y RODRÍGUEZ, J. Síntesis del Neógeno de la Cuenca del Duero, Noroeste de España…………………………. 32 MAESTRO, M.V., GARCÍA, E.X.M., BAJO, I., CÁRDENAS CARRETERO, J. RICO GARCÍA, A. y CIVIS, J. Resultados preliminares del contenido paleoictiológico de la sección “Hacienda Clarevot” (Cuenca del Guadalquivir, Sevilla, SO España)………………………………… 34 PAIS, J., CUNHA, P.P. & LEGOINHA, P. The Palaeogene and Neogene of the Lower Tejo Basin (western Iberia, Portugal)………… 35 PISANO, M.F., AGUIRRE, M.L., FARINATI, E. y FUCKS, E. Gastropoda y Bivalvia característicos del Pleistoceno de Bahía San Blas: estudios preliminares……………………………………………………………….. 36 RICO GARCÍA, A., CÁRDENAS CARRETERO, J., BAJO, I., MAESTRE, V. y CIVIS, J. La sección “Hacienda Clarevot” (Sevilla, SO España) y su registro paleontológico (Neógeno Superior, Cuenca del Guadalquivir)………………………………………………... 37 RICHIANO, S.M., AGUIRRE, M.L., FARINATI, E.A., FUCKS, E. y PISANO, F. Tafonomía comparativa de bivalvos dominantes en el Cuaternario marino de Argentina……. 24 ROMERO, M.V., RODRÍGUEZ, V., CASADÍO, S., MARTÍN, M.C. y PASCUAL, M. Las esponjas perforantes: una clave para conocer aspectos de la vida de una ostra Miocena del Atlántico Sur…………………………………………………………………….. 38 RUIZ, F., ABAD, M., PENDÓN, J.G., GONZÁLEZ REGALADO, M.L., TOSQUELLA, J., PRUDENCIO, M.I. y DIAS, M.I. Análisis microfaunístico del nivel condensado Tortoniense en el margen pasivo de la Depresión del Guadalquivir (España)…………………………………………………………. 39

RUIZ, F., GONZÁLEZ REGALADO, M.L., ABAD, M. y CIVIS, J. Ostrácodos Neógenos marinos del Suroeste de Europa………………………………………. 40 RUIZ, F., GONZÁLEZ REGALADO, M.L., ABAD, M., CIVIS, J. GONZÁLEZ DELGADO, J.A., MUÑOZ, J.M. y PENDÓN, J.G. Efecto de las tormentas en las asociaciones de Foraminíferos bentónicos del Plioceno de Huelva (Suroeste de España)………………………………………………………………. 41 SPRECHMANN, P., CHEMALE, F. Jr. Y GAUCHER, C. Presencia del Burdigaliense en las secuencias basales en la Perforación IGU Chuy Nº 364 del Uruguay datadas con 87Sr/86Sr en tecas de foraminíferos Cibicides…………….. 42 TOMASSINI, R.L. y MONTALVO, C.I. Evidencias de depredación en restos de vertebrados asignados a la edad Montehermosense en Farola Monte Hermoso, Buenos Aires, Argentina…………………….. 44 TOSQUELLA, J., ABAD, M., PENDÓN, J.G., RUIZ, F. y GONZÁLEZ REGALADO, M.L. Niveles de concentración de Heterostegina (Nummulitidae, Foraminifera) en los sedimentos del Tortoniense (Mioceno Superior) en el margen pasivo del sector Occidental de la Cuenca del Guadalquivir (SW de España)……………………….. 45

CONFERENCE 4th CONGRESS, Buenos Aires (Argentina), December 2008
El registro estratigráfico continental del Néogeno del centro de Argentina
The continental Neogene stratigraphic record of central Argentina.
Marcelo Zárate INCITAP (CONICET-UNLPAM) Avenida Uruguay 151, 6300 Santa Rosa La Pampa e-mail: [email protected] El registro estratigráfico continental del Neógeno exhibe una amplia distribución geográfica en el centro de Argentina. La región analizada se extiende desde los 34º S hasta los 40º S; abarca principalmente la llanura pampeana y el extremo norte de la Patagonia.. Los depósitos conforman una extensa cubierta sedimentaria de espesor relativamente reducido, variable entre 100 y 200 metros en promedio según el sector analizado. Están integrados por depósitos limo-arenosos a areno-limosos de composición volcaniclástica dominante, procedentes del área andina, con aportes secundarios de las sierras circundantes. Entre otros rasgos, se caracteriza por la presencia de paleosuelos y acumulaciones calcáreas de morfología variable. La acción eólica y el retrabajamiento por agentes ácueos son los principales agentes de transporte y acumulación vinculados con la génesis de los depósitos. Debido a la homogeneidad litológica y la discontinuidad de los afloramientos, la estratigrafia de la sucesión sedimentaria neógena se ha basado fundamentalmente en criterios paleontológicos. De esta manera, se identificaron diversas entidades (unidades litoestratigráficas) de edades relativas distintas, inferidas a partir de las asociaciones de vertebrados fósiles exhumados de los depósitos (edades mamífero en el sentido de Pascual et al., 1965). La aparente monotonía de la llanura, cuando es analizada en escala regional, enmascara variaciones geológicas significativas que definen áreas o sectores con características propias de la sucesión sedimentaria continental neógena. Por ello, el vasto ámbito del territorio considerado, se ha subdividido en distintas unidades de paisaje individualizables por sus características hidrográficas, topográficas, geomorfológicas y estratigráficas. En cierta manera, estas unidades de paisaje reflejan la compleja composición litológico-estructural del sustrato de la sucesión sedimentaria neógena, . agrupable en bloques o áreas positivas y cuencas tectónicas con rellenos sedimentarios de varios miles de metros cuyo origen se remonta a la apertura del océano Atlántico sur en el Mesozoico.

La acumulación de los depósitos continentales neógenos comienza hace alrededor de unos 12 Ma con posterioridad al retiro del Mar Paranense, extensa ingresión marina que cubrió las áreas de las cuencas tectónicas (Cuenca del Colorado, Cuenca del Salado, Cuenca de Macachin, entre otras). Paralelamente, los sectores de las áreas positivas o bloques (Positivo Bonaerense, Bloque de la Pampa central, Bloque de Chadileuvú, Sierras Pampeanas) permanecieron emergidos. La cubierta sedimentaria continental neógena se acumuló tanto en las áreas de bloques como de cuencas tectónicas. En el Bloque de la Pampa central y el Positivo Bonaerense, así como en la cuenca de Macachín y el sector norte de la cuenca del Colorado, la secuencia se caracteriza por un prolongado hiato estratigráfico que se extiende desde el Mioceno tardío-Plioceno hasta el Pleistoceno tardío cuando se reinicia la sedimentación. Hacia el norte, en el ámbito de la cuenca tectónica del Salado, el registro sedimentario continúa durante el Cuaternario. Las cuencas sedimentarias del Mioceno tardío y Plioceno respectivamente, exhiben distintas extensiones y están parcialmente superpuestas. La del Mioceno tardío es la de mayores dimensiones abarcando gran parte de la región considerada. En tanto, la cuenca de sedimentación pliocena se ubicaría básicamente dentro de la cuenca tectónica del Colorado y en el margen sur del Positivo Bonaerense. Actualmente, para comprender la naturaleza de la distribución y variaciones regionales del registro estratigráfico se explora el papel que pueden haber desempeñado entre otros, los cambios climáticos, las variaciones del nivel del mar, así como el papel potencial que puede haber desempeñado la tectónica de los Andes y de las cuencas sedimentarias (Salado, Colorado, Macachin, entre otras). Referencia Pascual, R., Ortega Hinojosa, J., Gondar, D., Tonni, E.P. 1965. Las edades del cenozoico mamalífero de la Argentina, con especial atención a aquellas del territorio bonaerense. Anales Comisión de Investigaciones Científicas de la Provincia de Buenos Aires, 6: 165-193.

CONFERENCE 4th CONGRESS, Buenos Aires (Argentina), December 2008
Ciencias de la Tierra para la Sociedad: hacia un Planeta sostenible
Jorge Civis Dpto. Geología, Facultad de Ciencias. Universidad de Salamanca (España); [email protected] El pasado día 12 de Febrero de 2008, tuvo lugar en la Sede de la UNESCO en París,
la inauguración oficial del Año Internacional del Planeta Tierra, proclamado por la Asamblea General de las Naciones Unidas, el 22 de Diciembre del año 2005 y como una contribución en el marco del “Decenio para la Educación del Desarrollo Sostenible”(1).
En la declaración de la ONU se señala, en uno de los párrafos, “que la abundante información científica disponible sobre el Planeta Tierra no se aprovecha y es poco conocida del público y los responsables de políticas y decisiones” y se declara convencida de que la enseñanza de las ciencias de la Tierra proporciona a la humanidad instrumentos para el uso sostenible de los recursos naturales y para construir la infraestructura científica esencial para el desarrollo sostenible.
Esta declaración ha sido promovida por la División de Ciencias de la Tierra de la UNESCO y la IUGS (Unión Internacional de Ciencias Geológicas), porque la comunidad de geocientíficos es consciente de que sus conocimientos pueden contribuir muy significativamente en el bienestar social y puso en marcha esta iniciativa durante la celebración del 32 Congreso Geológico Internacional (IGC) celebrado en Florencia (Italia), en 2004, con 140 paises representados y 7000 participantes. Así conjuntamente la IUGS y el IGC proclamaron las siguientes declaraciones:
“Las geociencias pueden contribuir de forma significativa a un mundo más seguro, más saludable y más próspero”
“Esta potencial contribución está siendo seriamente subutilizada por la sociedad y debería ser sustancialmente incrementada”
“La proclamación de un año internacional bajo el auspicio de los estados miembros de las Naciones Unidas ayudaría a las Ciencias de la Tierra a realizar su plena contribución al desarrollo sostenible de planeta”
La vida ha ido evolucionando y adaptándose a los cambios sufridos por la Tierra; una evolución simultánea y conjunta: la Tierra condiciona la vida y la vida transforma el Planeta. Vivimos por cortesía del Planeta.
Sin embargo, la sociedad en general tiene poco conocimiento del escenario en el cual se desarrolla nuestra vida y nuestro progreso y es poco consciente de la necesidad de su buen uso y de la fragilidad del mismo, y debe plantearse un reto y un compromiso: perseguir que las generaciones futuras alcancen un mundo más seguro y más próspero y utilizar el conocimiento que los científicos del Sistema Tierra han generado y generan.

En la resolución de las Naciones Unidas puede leerse “teniendo en cuenta el papel fundamental que podría desempeñar el Año, entre otras cosas, en la sensibilización del público sobre la importancia para el desarrollo sostenible de los procesos y los recursos de al Tierra, la prevención, reducción y mitigación de los desastres, la creación de capacidad para la gestión sostenible de los recursos, y su importante contribución al Decenio de las Naciones Unidas de la Educación para el Desarrollo Sostenible. DECIDE proclamar 2008 Año Internacional del Planeta Tierra”.
Las actividades, con motivo del Año Internacional, se llevan acabo durante un Trienio
2007-2009, siendo el año 2008 el año central de las actividades del programa diseñado para: .- Fomentar actividades de aprendizaje, difusión e investigación del campo de las
Ciencias de la Tierra .- Incrementar la conciencia del gran público y de la clase política sobre el potencial
de la Ciencias de la Tierra para mejorar la calidad de vida de los ciudadanos y la salvaguarda del planeta
Esta concienciación debe abarcar a los sistemas educativos, legislación y deben afrontarse tomas de decisión y medidas que contemplen la importancia que el conocimientos de las ciencias de la Tierra y su aplicación tienen para el futuro
En el logotipo de esta conmemoración están reflejados los componentes del Sistema: la Tierra sólida (Litosfera) es indicada en rojo, la Hidrosfera en azul oscuro, en verde la Biosfera y en azul claro la Atmósfera.
Muchos son los países que se han sumando a esta iniciativa y están llevando a cabo
diversas actividades, tanto en el marco de Proyectos de Investigación como mediante actividades divulgativas, y se está consiguiendo que la sociedad se implique en el proyecto, y que en muchas ciudades el logo del Año Internacional del Plantea Tierra se haga visible en exposiciones, ciclos de conferencias y eventos científicos entorno a una serie de temas que cubren los objetivos (2):
.- Aguas subterráeas: un recurso que debe ser explotado con responsabilidad
.- Riesgos naturales: investigación e intervención para prevenir, evitar y minimizar los efectos de desastres naturales de un planeta vivo
.- Tierra y salud: para asegurar unas condiciones ambientales saludables, conocer las propiedades beneficiosas o nocivas de muchas sustancias naturales, su distribución y mecanismos en el planeta
.- Cambio climático: el registro geológico nos permite un conocimiento de los cambios, por procesos naturales, del clima a lo largo del tiempo y este conocimiento nos permite entender el clima del futuro
.- Recursos naturales: tanto el uso racional de los recursos naturales, necesarios para nuestro desarrollo, como la investigación sobre nuevos recursos son fundamentales para alcanzar un futuro equilibrado y sostenible
.- La Tierra profunda: la Tierra, como planeta activo, se mueve, evoluciona y cambia, desde la parte más superficial a las capas profundas. El conocimiento del interior de la Tierra es fundamental para entender el efecto en la superficie: procesos, recursos etc.
.-Océanos: conocer e investigar las zonas frágiles de las áreas litorales de la Tierra, conocer la profundidad de los océanos, su dinámica, recursos que encierra, conocer la salud de la parte líquida del Sistema

.- Suelos: conocer y cuidar la piel del planeta sobre la que se asienta la vida. Su deterioro puede conducir a la destrucción de la base del desarrollo de la vida en la superficie de la tierra; la deforestación, sobreexplotación del suelo etc.
.-Tierra y vida: porque la vida hace al planeta Tierra único en el Universo y porque tierra y vida se desarrollan conjuntamente; la Tierra condiciona la vida y la vida transforma al planeta.
.-Megaciudades: El desarrollo de grandes núcleos urbanos requiere un exhaustivo conocimiento de su asentamiento; construcción de infraestructuras, utilización de recursos, generación de residuos y consecuencias
En España, la inauguración oficial de las actividades, tuvo lugar el día 13 de Febrero
del 2008. Los promotores de la celebración de este Año Internacional han sido el Instituto Geológico y Minero de España (IGME, dependiente del Ministerio de Ciencia e Innovación, a través de la Comisión Nacional de Geología y la Comisión Española de la UNESCO, y se ha constituido un Comité Español del AIPT para desarrollar las actividades programadas a las que se han sumado Universidades e Instituciones públicas y privadas, unidas con los objetivos de este Trienio.
Junto con los proyectos de investigación, congresos y reuniones científicas que se llevan a cabo en el marco del AIPT, y con convocatoria específica por parte del Gobierno español, merecen atención las actividades de divulgación a diferentes niveles, estudiantes, profesores, sociedad en general, como edición de folletos, vídeos, excursiones geológicas etc.
Destaca una Exposición Itinerante que está recorriendo diversas capitales de España, con el objetivo de sensibilizar a la sociedad en general, sobre los temas del AIPT. Se divide en varias partes en las que se pretende concienciar al visitante, mediante una presentación audiovisual, sobre los “peligros” que puede generar el Sistema Tierra, así como los beneficios que nos proporciona nuestro planeta., abordando las diez áreas temáticas del programa; conocer efectos de fenómenos naturales como circulación de las corrientes oceánicas o movimiento de las placas tectónicas o sentir la sensación de los movimientos sísmicos, la humedad de grutas subterráneas y recorrer una zona didáctica que permite ejercitar al visitante sobre conocimientos básicos en el campo de las ciencias de la Tierra.
Muchas instituciones han programado ciclos de conferencias, así como itinerarios geológicos para dar a conocer la variedad geológica de España, nuestros recursos, riesgos etc., con una aceptación muy alta. Una de ellas, ha sido llevada a cabo recientemente en Alicante, por iniciativa de su Universidad y del Parque Natural de Serra Gelada, con la participación , en esta jornada geológica, de más de 600 personas.
En el marco de este Trienio se celebra, en Buenos Aires, el 4º Congreso Internacional del R.C.A.N.S. (Regional Committee on Atlantic Neogene Stratigrpahy) dependiente de la IUGS, que tiene por objeto congregar a científicos que desarrollan su investigación en el Neógeno del dominio Atlántico. Este Congreso se ha organizando conjuntamente por instituciones argentinas, uruguayas y españolas.
(1).- Cumbre Mundial sobre el desarrollo Sostenible .Johannesburgo (Sudáfrica) 2002 (2) Resumen del folleto divulgativo editado por el Instituto Geológico y Minero de España, dependiente del Ministerio de Ciencia e Innovación

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Moluscos del Cuaternario marino de Argentina: herramientas útiles para interpretaciones paleoambientales y paleoclimáticas
Aguirre, M.L 1 y Farinati, E.A. 2
1. FCNYM, UNLP, calle 64 No.3, 1900 La Plata; CONICET; [email protected] 2. Departamento de Geología, Universidad Nacional del Sur. San Juan 670, 8000 Bahía Blanca, [email protected] Los cambios climáticos durante los episodios marinos del Neógeno atlántico, reflejados en numerosos grupos de organismos bentónicos y planctónicos, están bien documentados a nivel global. La malacofauna del Cuaternario marino de Argentina, aunque aún poco conocida en sectores costeros patagónicos, brinda información sobre variaciones ambientales y climáticas de distinto grado ocurridas durante el Pleistoceno medio y superior (desde el MIS9/7?) y Holoceno (MIS1) hasta la actualidad, y del mar posterior al LGM según estudios preliminares de testigos del nearshore y plataforma interna bonaerense (ca. 33-35 ka A.P.). Existe mayor información y control cronológico para el Holoceno, con malacofauna en conjunto indicativa de comunidades bentónicas litorales principalmente del infralitoral superior hasta 100 m de profundidad, controladas por corrientes oceánicas superficiales (Brasil, Malvinas).
En el área costera bonaerense para el lapso 7- 4,5 ka A.P. sugiere temperatura oceánica superficial (SST) ligeramente más altas y gradientes de salinidad diferentes en comparación con el litoral vecino moderno. Los taxones holocenos característicos (Littoridina australis, Mactra isabelleana, Corbula patagonica, Brachidontes rodriguezi, Tagelus plebeius) son típicamente eurihalinos y estuariales, del intermareal-infralitoral, sustratos lodosos o areno-lodosos; aproximadamente 5% son euritérmicos, 35% cálidos o templado-cálidos (vs. 14% actual), 55% templado-fríos, 5% exclusivamente fríos. Posteriormente a ca. 4,5 ka A.P. se habría definido el patrón biogeográfico actual con extinciones o retracciones de algunos taxones (i.e., Conus iheringi Frenguelli, Thais haemastoma (Linné), Triphora nigrocincta (C.B. Adams), Tegula atra (Lesson), Anachis avara (Say), Noetia bisulcata (Lamarck), Crassostrea rhizophorae (Guilding) y Anomalocardia brasiliana (Gmelin). En el nearshore la malacofauna es típica de fondos arenosos y aguas superficiales más cálidas, mientras que en la plataforma interna es menos diversa y abundante, típica de fondos lodosos y aguas más frías. Para Patagonia no se registran extinciones (excepto Tegula atra (Lesson), Ostrea tehuelcha Feruglio, Chama del Pleistoceno tardío, MIS5) y se reconocen desplazamientos

geográficos holocenos (Mactra spp., Corbula patagonica d´Orbigny, Diplodonta vilardeboana (d´Orbigny), Clausinella gayi Hupé). Durante el lapso ca. 9-6 ka A.P. habría existido mayor porcentaje de elementos cálidos o templado cálidos y menor abundancia de taxones estenotermos fríos hasta ca. 45-47° S. Desplazamientos geográficos y extinciones responderían a cambios en la circulación oceánica y atmosférica, ya que la respuesta malacológica a cambios climáticos y eustáticos es consecuencia del desplazamiento austral en los límites de las corrientes marinas, masas de agua oceánica superficial y provincias malacológicas. Estas evidencias requieren revisiones y abundantes análisis geocronológicos, geoquímicos y microestructurales para acotar inferencias paleoclimáticas del Holoceno e indagar sobre el Pleistoceno marino.

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Problemática de la variabilidad morfológica en moluscos del Cuaternario marino de Argentina: implicancias paleoambientales
Aguirre, M.L.1, Farinati, E.A.2, Pérez, S.I. 3 y Pisano, F.1, 4
1. Facultad de Ciencias Naturales y Museo, UNLP, calle 64 No.3, 1900 La Plata; CONICET; [email protected] 2. Departamento de Geología, Universidad Nacional del Sur. San Juan 670, 8000 Bahía Blanca, [email protected] 3. División Antropología, Facultad de Ciencias Naturales y Museo, [email protected] 4. Becaria Doctoral ANPCYT La problemática sobre la identificación taxonómica objetiva, genérica y/o específica, de moluscos del Cuaternario marino argentino ha comenzado a analizarse en estudios complexivos incluyendo material fósil y moderno. Continúa vigente la polémica sobre la significación de los conceptos tipológico y morfológico de especie y el criterio apropiado en determinaciones de material fósil-subfósil para taxones con representantes vivientes, con amplia abundancia y variabilidad morfológica (con morfos intermedios asignables a distintas especies descriptas). El estudio de la “forma” ha cobrado revitalizado interés y su importancia en términos evolutivos ha comenzado a revalorizarse, inclusive en el campo de la biología, desde donde frecuentemente se cuestiona el concepto morfológico (morfoespecie) priorizando la “especie biológica”, aunque proponiendo nuevos enfoques morfológicos (endosomáticos). Esta situación está bien ilustrada por taxones representativos del Cuaternario marino por su constancia y dominancia, difíciles de discriminar objetivamente en muestras abundantes tomadas al azar. Entre otros: Littoridina Souleyet (Hydrobiidae, Gastropoda) y Mactra Linné (Mactridae, Bivalvia) en el litoral de la provincia de Buenos Aires, Brachidontes Swainson (Mytilidae, Bivalvia) del litoral argentino. Herramientas como análisis biométricos tradicionales, junto con técnicas modernas de morfometría geométrica (Relative Warps sobre coordenadas de landmarks, puntos morfológicos anatómica o geométricamente homólogos, y semilandmarks, puntos espaciados regularmente a lo largo de un contorno) aplicados a conchillas con alto número de individuos completos, excelente preservación, distintos estadíos ontogenéticos (poblaciones), diferentes grados de variación morfológica inter y/o intraespecífica, reconocibles como componentes autóctonos de las comunidades originales, pueden aportar una valiosa información. Su utilidad radica en facilitar la discriminación taxonómica objetiva, establecer patrones de paleobiodiversidad y evaluar su posible valor paleoambiental. Enfoques multivariados (partes duras y blandas), junto con el análisis de la distribución geográfica de linajes génicos, son fundamentales para determinar si las diferencias entre “especies” son significativas, si impedirían el cruzamiento (diferentes especies biológicas), si representan diferencias reales o si se repite la problemática de la alta

plasticidad fenotípica que exhiben las conchillas cuaternarias. Hasta el momento se carece de esta información (fundamento “biológico” para discriminar especies sin utilizar criterios tipológicos-morfológicos exclusivamente). Frecuentemente se comete el error de considerar que los indicadores paleoambientales deben ser necesariamente especies biológicas. Sin embargo, la variabilidad morfológica de moluscos cuaternarios, aún cuando no fuera posible alcanzar acuerdo unánime sobre la nominación de “especies”, puede brindar información paleoambiental relevante, documentando variaciones fenotípicas en espacio y tiempo, útiles para interpretar la historia evolutiva de los taxones ligada a cambios ambientales y climáticos durante el Cuaternario tardío.

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Gastropoda y Bivalvia del Cuaternario del sector costero Bahía Lángara-Puerto Deseado (Santa Cruz, Patagonia, Argentina): interpretaciones
paleoambientales
Aguirre, M.L. 1,2, Richiano, S. 1, 3, Álvarez, M.F. 1, 4, Fucks, E.5, Farinati, E.A. 6 y Eastoe, Ch. 7
1, CONICET; 2, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, INGEA, calle 64 No. 3, 1900 La Plata, Argentina; 3, CIG, Centro de Investigaciones Geológicas, 1 nº 644, 1900 La Plata, Argentina; 4, Laboratorio de Ecología (FCEYN) Universidad Nacional de Mar del Plata, Funes 3350,7600; Argentina; 5, Latyr, Facultad de Ciencias Naturales y Museo de La Plata; 6, Departamento de Geología, Universidad Nacional del Sur; 7, Environmental Isotope Laboratory, Department of Geoscience, University of Arizona, USA. Tucson, Arizona 85721-0077, USA.
En terrazas marinas (TM) del Cuaternario del área costera entre Bahía Lángara y Puerto Deseado (norte de Santa Cruz) se identificaron 49 moluscos (33 Gastropoda y 16 Bivalvia) y los primeros registros cuaternarios de Ataxocerithium pullum, Crepidula protea, C. aculeata, Adelomelon ferrusacii, Brachidontes rodriguezi, Aequipecten tehuelchus, Ostrea tehuelcha, Solen tehuelchus. Las asociaciones malacológicas sugieren condiciones marinas típicas y paleoambientes del nearshore similares al litoral moderno vecino (Atlántico Sudoccidental, Provincia Magallánica), de sustratos principalmente duros, y aguas poco profundas y frías. La TMVI (+ 5-15m s.n.m, ca. 8-1,2 ka A.P.) se caracteriza fundamentalmente por la dominancia de Brachidontes spp., Nacella spp. y Mytilus edulis, asociados a Siphonaria lessoni. No existen evidencias para la TMVI de respuestas bióticas o efectos paleobiogeográficos marcados durante el lapso del Optimo Climático (“Hypsithermal”, Holoceno medio), documentadas más al norte en otras áreas costeras de Argentina, hasta ca. 44-45° S. Pocas diferencias distinguen la TMV (+ 15-30 m s.n.m., ca. 108-137 ka A.P. MIS5 ?) y TMIV (+20-30 m s.n.m; ca. 242 ka A.P.) del Holoceno y litoral moderno. En la TMV son exclusivos Clausinella, Eurhomalea, Solen tehuelchus y Brachidontes cf. darwinianus (típica de aguas más cálidas); en la TMIV son característicos Trophon, Buccinanops, Brachidontes; Pectinidae indet. El predominio de Trophon spp. (típicos de aguas frías magallánicas) en muestras modernas y su escasez o ausencia en las TM sugiere temperatura oceánica superficial ligeramente menor actualmente. Algunas similitudes taxonómicas con la costa pacífica sudamericana (i.e., Fissurella, Nacella, Tegula, Crepidula, Trochita, Acanthina) responderían a corrientes frías que posteriormente a la apertura del Pasaje de Drake facilitaron dispersiones de elementos fríos al Atlántico sudamericano.

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Caracteres distintivos de los depósitos marinos holocenos del estuario de Bahía Blanca.
Aliotta S.1,2, Farinati E. A1. y Spagnuolo J. O.1,2
1. Instituto Argentino de Oceanografía (CONICET-UNS). 2. Departamento de Geología, Universidad Nacional del Sur. [email protected]; [email protected]; [email protected] Sobre la costa norte del estuario de Bahía Blanca se presentan depósitos elongados, aproximadamente paralelos al litoral, atribuibles al Holoceno marino y de los cuales se analizan sus características morfosedimentológicas, paleontológicas, tafonómicas y genéticas. Estos cordones areno conchilíferos, se distribuyen desde General Cerri (sector más interno del estuario) hasta aproximadamente 20 km al este de la ciudad de Punta Alta y, generalmente, se apoyan sobre material fango arenoso. La cota de sus bases se ubica entre 5 y 6 m sobre el nivel medio del mar y alcanzan alturas de 0,70 m a 2 m, con anchuras inferiores a 70 m. El perfil transversal de dichas geoformas normalmente es asimétrico, con su mayor pendiente (9º) hacia el continente. La disposición de sus estratos es paralela con intercalaciones de arena mediana a gruesa. Asimismo, presentan niveles con abundantes restos de moluscos (fábrica bioclasto soporte) y rodados de cuarzo, tosca, pumicita y limolita, éstos últimos con signos de bioerosión. La composición taxonómica de estos depósitos marinos señala la presencia dominante de moluscos (38 especies de bivalvos y 32 de gastrópodos), entre los que se destacan por su abundancia Brachidontes rodriguezi y Littoridina australis respectivamente, más una fauna asociada compuesta por poliplacóforos, escafópodos, cnidarios (pennatuláceos y antozoos), crustáceos (cirripedios y decápodos) y briozoos. Entre los microfósiles calcáreos se reconocen foraminíferos y ostrácodos. Varias dataciones radimétricas por carbono 14 realizadas sobre el material conchilífero indican edades que van desde los 5200 a los 6650 años A.P. para los cordones más antiguos, hasta edades de 1430 a 1740 correspondientes a los últimos depósitos marinos holocenos. El origen de estas geoformas se atribuye a eventos multiepisódicos de tormentas asociados al máximo transgresivo holoceno y al período regresivo subsecuente. La historia tafonómica de los restos conchilíferos comprende una breve etapa bioestratinómica, donde es escasa la bioerosión e incrustación. También, la abrasión y fragmentación valvar resultan poco significativas, consecuencia de un rápido transporte por olas de tormenta de energía intermedia desde la zona de producción hasta la zona de depositación. Durante la etapa fosildiagenética se hace notorio un importante proceso de disolución de las valvas por efecto del intemperismo (acción de aguas meteóricas y freáticas, calentamiento solar, fluctuaciones diarias de temperatura e influencia de la vegetación).

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Asociaciones de nanofósiles calcáreos y foraminíferos del Mioceno temprano
del Atlántico sur occidental
Angelozzi, G.N.1, Ronchi, D.I.1 y Pérez Panera, J.P.2
1GEMA SRL. Servicios Bioestratigráficos, Av. Calchaquí Km 23,5 (1888) Florencio Varela, Argentina. [email protected] 2 CONICET-División Paleontología Invertebrados – Museo de Ciencias Naturales de La Plata. Paseo del Bosque s/n, La Plata. CP 1900. Buenos Aires, Argentina. [email protected] Se dan a conocer las asociaciones de nanofósiles calcáreos y foraminíferos del Mioceno temprano ( biozonas NN2 y NN1 de Martini, 1971) registradas en muestras de recorte de perforación de siete pozos, en el sector comprendido entre la Isla Grande de Tierra del Fuego y las islas Malvinas, en el área sudoccidental del atlántico, plataforma continental Argentina. Las asociaciones registradas presentan rasgos particulares que las diferencian con los sectores de menor latitud y similitudes con las asociaciones recuperadas en el Plateau de Malvinas (Boltovskoy, 1978; Wise et al., 1982; Basov y Krasheninnikov, 1980) aunque en ese sector el Mioceno temprano aparece con retrabajo del Oligoceno y en numerosos sondeos se lo consideró ausente (Wise, 1983; Wise y Wind, 1977). Las asociaciones microfosilíferas para el Mioceno temprano en la región que aquí se estudia presentan una alta diversidad y número de ejemplares permitiendo reconstruir las condiciones ambientales en las que se desarrolló esta paleocomunidad. De acuerdo a los nanofósiles se interpreta un ambiente de aguas superficiales templado-frías, aunque se reconoce la influencia de una zona de mezcla que aporta elementos de aguas relativamente cálidas. A través del análisis de índices de plancton/bentos referidos a los foraminíferos se interpretaron una serie de pulsos regresivos que aumentan hacia el tope de la secuencia, pero siempre manteniéndose en el orden de un ciclo mayor transgresivo en un ambiente que se interpreta como de plataforma abierta media-externa (80-200 mbnm). Así mismo se infieren procesos de up welling en varios niveles, evidenciados por la ocurrencia simultánea de elementos calcáreos y silíceos (diatomeas). La asociación de nanofósiles calcáreos está caracterizada por la dominancia de la especie indicadora de aguas frías Coccolithus pelagicus (Wallich) Schiller. También son abundantes las helicosphaeras, reticulofenestras, especialmente Reticuofenestra pseudoumbilicus (Gartner) Gartner y Cyclicargolithus abisectus (Müller) Wise. Pontosphaera multipora (Kamptner) Roth, Cyclicargolithus floridanus (Roth y Hay en Hay et al.) Bukry y Braarudosphaera bigelowii (Gran y Braarud) Deflandre son elementos comunes de la asociación. Como elementos raros, pero persistentes, se registraron especies características de aguas templado - cálidas de los

géneros Sphenolithus, y Discoaster deflandrei Bramlette y Riedel. La asociación de foraminíferos está caracterizada por las especies planctónicas Globigerina woodi woodi Jenkins, G. woodi conecta Jenkins, G. brazieri Jenkins y Globorotalia (T) praescitula Blow. Entre las especies bentónicas se destacan Uvigerina proboscidea Schwager, U. spinocostata Cushman, Siphonina lamarckiana Cushman, Martinottiella antarctica Parker y Jones, Laticarinina pauperata (Parker y Jones) y Textularia leuzingeri Cushman. Referencias.
Basov, I.A. y Krasheninnikov, V.A. 1983. Benthic foraminifers in Mesozoic and Cenozoic sediments of the
southwestern Atlantic as an indicator of paleoenvironment, DSDP, leg 71. En Initial Reports of the Deep Sea Drilling Project 71,1: 445-460.
Boltovskoy, E. 1978. Estudio bioestratigráfico y paleontológico (foraminíferos bentónicos) del Cenozóico
superior al este de las Islas Malvinas (DSPD, crucero 36, sitios 327 y 329) Revista del Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Geología. Tomo 8, 2: 1-95.
Martini, E., 1971: Standard Tertiary and Quaternary calcareous nannoplankton zonation. En A. Farinacci
(Ed.), Proceedings 2nd Planktonic Conference, Roma, 1970. 2: 739-785. Wise, S.W.Jr. 1983. Mesozoic and Cenozoic calcareous nannofossils recovered by Deep Sea Drilling Project
Leg 71 in the Falkland Plateau Region, Southwest Atlantic Ocean. En Initial Reports of the Deep Sea Drilling Project 71: 481-550.
Wise, S.W.Jr. y Wind, F.H. 1977. Mesozoic and Cenozoic calcareous nannofossils recovered by DSDP Leg
36 drilling on the Falkland Plateau, SW Atlantic sector of the Southern Ocean. En Initial Reports of the Deep Sea Drilling Project 36: 296-309.ç
Wise, S.W., Ciesielski, P.E, MacKenzie, D.T., Wind, F.H., Busen, K.E., Gombos, A.M., Haq, B.U.,
Lohmann, G.P, Tjalsma, R.C., Harris, W.K., Hedlund, R.W, Beju, D.N., Jones, D.L., Plafker, G., and Slitter, W.V., 1982, Paleontologic and paleoenvironmental synthesis for the southwest Atlantic Ocean based on Jurassic to Holocene faunas and floras from the Falkland Plateau: En Campbell Craddock (ed.), Antarctic Geoscience: Third symposium on Antarctic Geology and Geophysics, 4: 155-163.

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Registros paleoicnológícos de la Formación Río Negro, facies continental (Mioceno tardío – Plioceno temprano), costa atlántica de la Provincia de Río
Negro, Argentina
Silvia A. Aramayo
Departamento de Geología, Universidad Nacional del Sur, calle San Juan 670, B8000ICN Bahía Blanca, Argentina. E-mail: [email protected]
La costa atlántica de la Provincia de Río Negro, entre las localidades de Balneario el Cóndor y Las Lobería- 30 km de extensión- está caracterizada por acantilados de gran altura ( 30 a 40 m) de los cuales se desprenden bloques debido a la acción del mar. En los planos de dichos bloques, se registran huellas de mamíferos y aves que han sido objeto de estudio. Angulo y Casamiquela (1982) y recientemente Aramayo (2005) se refieren a la icnología del área. El estudio geológico de las barrancas ha sido actualizado por Zavala y Freije (2001); se distinguen tres miembros: 1) Miembro basal eólico continental 2º ) Miembro marino, no considerado en este trabajo, 3º) Miembro eólico continental superior donde se registra el mayor número de paleoicnitas. Estas corresponden a mamíferos terrestres, destacándose las de perezosos terrestres, (Megatherichnum isp.) del Orden Xenarthra, mamíferos autóctonos sudamericanos que adoptaban generalmente una locomoción bípeda; la morfología de manos y pies está bien diferenciada, característica observada también en yacimientos cuaternarios como el de Pehuen Co, provincia de Buenos Aires. Numerosas icnitas de ungulados, de contorno subcircular manifiestan una incipiente tridactilia, asignándolas a notoungulados o litopternos, órdenes autóctonos sudamericanos. Huellas tetradáctilas y tridáctilas se asignan a manos y pies respectivamente de carpinchos extinguidos (Rodentia, Caviomorpha) ( cf. Porcellusignum isp.) y huellas de marsupiales carnívoros, cuya característica es presentar un diámetro latero medial mayor que la longitud del pie, cinco dedos terminados en garras, y una almohadilla palmar o plantar muy desarrollada. Entre las aves se observan huellas de palmípedas, tridáctilas con dedos delgados, otras pequeñas y numerosas formando un pisadero (charadriformes) y una rastrillada de cuatro icnitas tridáctilas de gran porte, asignadas a Phorusrhacidae. El registro en arcillitas realizado hasta el presente indica un ambiente de lagunas donde las icnitas quedaban impresas en sus orillas; ellas representan el ámbito de encuentro de la fauna de mamíferos y aves en búsqueda de agua y alimento; muchas huellas están impresas también en las areniscas eólicas, por ejemplo en un bloque de 16 m2 donde se entrecruzan una rastrillada de xenartro, otra de ungulado y la rastrillada de Phorusrhacidae. Dada la fauna autóctona de herbívoros representada por perezosos terrestres y ungulados, los marsupiales carnívoros y las aves Phorusrhacidae cumplían el rol carnicero o tal vez carroñero durante el período Terciario, cuando el continente sudamericano estuvo aislado y los verdaderos carnívoros no habían ingresado aún procedentes de América del Norte.

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Pliocene non-tropical platform coarse biocalcarenites in the Atlantic central region of Marocco
Armenteros, I.1,a, González Delgado, J.A.1,b, Civis, J.1,c, Alonso Gavilán, G. 1,d & Dabrio, C.J. 2,e 1: Dpto. Geología, Facultad de Ciencias, c/Parque s/n, 37008 Salamanca (España) a: [email protected]; b: [email protected]; c: [email protected]; d: [email protected] 2: Dpto. Estratigrafía, Facultad de Geología, Univ. Complutense e Instituto de Geología Económica, CSIC, Madrid. Ciudad Universitaria, e: [email protected] Carbonate pliocene successions in the region of Safi (southern part of the the “domaine mésetien” extending to the west of the moroccan Atlas Mountains between Rabat and Agadir) consist of grainstone and rudstone facies dominated by a temperate shallow water association very rich in mollusks, barnacles, echinoids and benthic foraminifera that has been grouped as being part of the barnamol association (sensu Hayton et al. (1995). Their study is worthy of attention since the situation around latitude 32º N would allows including it within the tropical domain, above all if the Pliocene age of the succession is taken into account. However, the composition of the association of bioclasts indicates that some different circumstances must concur to produce a non-tropical carbonate sediments. Preliminary studies (Gonzalez Delgado et al. 2007) have recognized this singularity and this paper tries to characterise in depth the carbonate sedimentation in the Pliocene shallow carbonate platforms of the Safi region.
The succession is around 20 m thick. Superimposed nodular calcrete profiles commonly crown the succesion. The best outcrop to the south of Safi is Jorf al Yhoudi section. To the north, Pliocene carbonates are well represented in Borj Nador and Sidi Khelil sections.
The Jorf Al Yhoudi section consists of clast-supported biocalcirudites and grain-supported, very coarse biocalcarenites, with a fine–grained carbonate matrix with variable amounts (4 to 30 %) of silty to fine sand-sized quartz. It shows massive to crudely stratified beds ranging from 0.4 m to 2 metres in thickness and is characterized by the presence of decimetre-thick levels of paucispecifical shell concentrations dominated by the bivalves Ostrea lamellosa or Pecten benedictus, the echinoid Rotuloidea fimbriata, or the barnacle Balanus. Benthic foraminifera are poorly preserved, being Elphidium (E. crispum, E. advenum), Lobatula lobatula and Pararotalia sp. the most abondant.
The outcrops to the north of Safi consist of well-sorted, moderately cemented, coarse, grain-supported biocalcarenites, devoid of micrite and silty particles. These facies occur well stratified in tabular to cuneiform beds some centimetres to 0.8 metres thick, with internal cross lamination. Some of them are arranged in cosets forming large-scale cross stratification and upward-thickenning sequences. Well-preserved molluscs are scarce, but benthic foraminifers

have higher diversity, being the most common Elphidium, Ammonia beccarii, Lobatula lobatula and Cibicides refulgens. The ostracod Callistocythere is also abundant.
C and O isotopes from Pecten and Ostrea range from 1,49 to -6,19%o δ13CPDB, and from 0,08 to -4,87 δ18OPDB respectively, although most data have intermediate values. Data for the whole rock range from -7,48 to -4,31 for δ13CPDB, and -5,56 to -4,45 for δ18OPDB. These values suggest a different preservation of the original signature between specimens and also between stratigraphic horizons, even in samples where the original microstructure of shells is well preserved. The coarse textures, fossil associations, taphonomic signatures, sedimentary structures and shallowing character of the regional succession point to deposition in platform interior between the lower shoreface facies and beach facies in a ramp context (sensu Tucker and Wright, 1990). Northern sections show facies that indicate high energy regime, whereas southern sections point to a more restricted area of the inner platform probably related to coastal embayments. These shoreline sequences developed during still-stands in shorelines of moderate to low gradient. References: González Delgado, J.A., Civis, J., Alonso Gavilán, G., El Hajjaji, K., Ben Moussa, A., Rivas Carballo, A. & Valle, M.F., 2007.- New paleontological data in the neogene of Safi (Morocco). Abstracts First MAPG Congress, Marrakech: 185-186 Hayton, S., Nelson, C.S. and Hood, S.D., 1995.- A skeletal classification system for non-
tropical carbonate deposits based on New Zealand Cenozoic limestone. Sedimentary Geology 100, 123-141.
Tucker, M.E. and Wright, V.P., 1990.- Carbonate Sedimentology. Blackwell Scientific
Publications, Oxford. 482 p.
ACKNOWLEDGMENTS: financial support of the European Union and the Spanish Ministerio
de Ciencia y Tecnología, Project CGL2006-05473BTE

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Tránsito Mio-Plioceno en el sondeo “Montemayor” (Neógeno de Huelva, Cuenca del Guadalquivir, España). Diagrafías y Foraminíferos bentónicos
Civis, J.*, Alonso Gavilán, G.*, González Delgado, J.A.*, Ruiz Muñoz, F.** y González Regalado, M.L.**
*Dpto. de Geología; Universidad de Salamanca (España) ** Dpto. de Geodinámica y Paleontología; Universidad de Huelva (España)
El Neógeno marino del borde occidental de la Cuenca del Guadalquivir (provincia de Huelva, España) registra sedimentos del Tortoniense superior, Messiniense, Plioceno inferior y Plio-Pleistoceno, habiéndose definido cuatro Formaciones (Civis et al., 1987; Mayoral y Pendón, 1987).
Con el fin de obtener un registro completo de dichas Formaciones, se ha realizado un sondeo de testigo contínuo (sondeo Montemayor) situado cerca de Moguer (Huelva). El sondeo se realizó mediante un convenio con el Instituto Geológico y Minero de España y en el marco de un proyecto de investigación del Ministerio de Ciencia y Tecnología. Abarca 271 metros, agrupado en cuatro unidades litosísmicas caracterizadas por asociaciones de electrofacies diferentes. Las edades han sido calibradas mediante eventoestratigrafía y paleomagnetismo (González Delgado et al, 2002; Larrasaoña et al, 2008). En este trabajo, se estudian los 85 metros superiores que incluyen el límite Messiniense – Plioceno.
El objetivo de este trabajo es definir e interpretar los cambios de las asociaciones de foraminíferos bentónicos en el tránsito Mio/Plioceno y en el Plioceno inferior, así como evaluar su significado en la evolución de la cuenca una vez interrumpida la comunicación Atlántico / Mediterráneo durante la crisis de salinidad Messiniense, cuando esta parte del estrecho Nord-Bético, se conformó como un golfo abierto al océano Atlántico. También se interpretan los resultados de las diagrafías de rayos gamma, potencial espontáneo, resistividad y neutrón.
Se diferencian cuatro asociaciones de foraminíferos bentónicos denominadas A, entre los 86 y 59 metros, B (59-41m), C (41–27m) y D (27-16 m), que corresponde al último registro marino. Las 4 asociaciones coinciden parcialmente con diferentes electrofacies. La asociación A se define por la presencia de formas que son prácticamente exclusivas: Sphaeroidina bulloides, Cibicidoides floridanus y Hoeglundina elegans; la asociación B se caracteriza por el predominio de Nonion boueanum y Fursenkoina schreibersiana; la C por N. boueanum, Ammonia becccarii, Nonionella turgida y la D por el predominio en Elphidium spp. Bulimínidos, Bolivinidos, y Cibicides/Heterolepa están presentes a lo largo de todo el testigo estudiado, si bien las especies dominantes varían. La abundancia de indivíduos se mantiene aproximadamente constante a lo largo del testigo a excepción del tramo con la asociación B, que coincide con niveles glauconíticos de la base de la Fm. Arenas de Huelva, y se caracteriza por un

brusco incremento en la abundancia junto a una caída en la diversidad. El número de especies es mayor en las arcillas que en las arenas.
Los cambios en las asociaciones se han interpretado como el resultado de cambios en la sedimentología, batimetría, temperatura y materia orgánica relacionados con la propia dinámica del borde occidental de la Cuenca.
Referencias Civis, J., Sierro, F.J., González Delgado, J.A., Flores, J.A., Andrés, I., Porta J. y Valle, M.F. (1987). In Paleont. Neog. Huelva, Dpto Paleont.Univ.Salamanca (ed): 9-21 González Delgado, J.A., Civis, J., Larrasoaña, J.C., Sierro, F.J., García Cortés, A. y Alonso Gavilán, G. (2002).In XVIII Jornadas de la S.E.P. –II Congr.Ibérico de Paleont., Civis y González Delgado (Ed):194-195 Larrasoaña, J.C., González Delgado, J.A., Civis, J., Sierro, F.J., Alonso Gavilán, G. y Pais, J. (2008). Geotemas 10: 1175-1178 Mayoral, E. y Pendón, J.G. (1987). Acta Geológica Hispánica 21-22:507-513 Agradecimientos .- IGME ( Instituto Geológico y Minero de España) por la ejecución del sondeo Montemayor I .- Proyecto CGL2006-05473 BTE (Ministerio de Educación y Ciencia, España / Unión Europea, Programa FEDER)

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
The ecologic and taphonomic effects of a Miocene volcanic ashfall on
Atlantic inner-shelf communities
Crawford, R. S.1, Casadío, S.2, Feldmann, R.1, Griffin, M.2, and Schweitzer, C. E.3
1 Department of Geology, Kent State University, Kent, OH 44242, USA 2 Facultad de Ciencias Exactas y Naturales, Universidad Nacional de La Pampa, Av. Uruguay 151, L6300 CLB Santa Rosa, La Pampa, Argentina and CONICET 3 Department of Geology, Kent State University Stark Campus, 6000 Frank Ave. NW, North Canton, Ohio 44720 Explosive eruptions were frequent throughout the Cenozoic in Patagonia. South of 45ºS this volcanism showed a maximum activity during the Miocene. This is evidenced in the thick tuffaceous deposits that make up the continental and marine rocks of that age. However, the effects that this volcanism had on Patagonian Miocene ecosystems remain poorly understood. Recognition within the early Miocene Monte León Formation of four paleosurfaces containing different invertebrate assemblages and in close geographic and stratigraphic proximity to each other allows assessment of the ecological and taphonomic impacts of volcanic ashfall on Miocene inner-shelf communities. The paleosurfaces are exposed within the boundaries of Monte León National Park (Paleosurface I, lat. 50°19´22.5 S, long. 68°51´31.6 W; Paleosurface II, lat. 50°19´22.6 S, long. 68°51´32.2 W; Paleosurface III, lat. 50°19´26.8 S, long. 68°51´43.1 W; and Paleosurface IV, lat. 50°19´32.4 S, long. 68°51´55.1 W) and are accessible during low tide on the wide abrasion platform along the beach there. The paleosurfaces were measured using a 1.5 m2 grid system, and described and mapped in plan view recording the precise location of all fossil specimens. Sedimentologic, stratigraphic, and taphonomic features suggest a shallow water habitat, above storm wave base. Two different invertebrate assemblages were recorded: 1) a relatively diverse, normal marine benthic fauna consisting of bivalves, gastropods, bryozoans, echinoids, and decapods; and 2) a unique assemblage consisting solely of numerous, exceptionally well-preserved, fully-articulated crabs. The crab-rich assemblage was stratigraphically below the surfaces containing the normal marine assemblage. The crabs are contained within a slightly compacted, argillaceous volcanic tuff. Biogenic fragments other than the crabs are noticeably absent from the deposit, unlike samples collected from the stratigraphically higher surfaces. Most of the crabs were preserved with their third maxillipeds in an open, gaping posture. This is consistent with postures observed in crabs suffering from respiratory distress. From the above evidence it is inferred that the crabs were suffocated, killed, and rapidly buried. The apparent faunal disparity observed on the four paleosurfaces within the Monte León Formation is interpreted as representing the initial decimation and eventual re-establishment of the benthic marine fauna following a catastrophic volcanic event.

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Evidencias paleoambientales del Pleistoceno Marino Tardío de la Formación Pascua basadas en moluscos y microfósiles
Erra, G. 1,2, Ferrero, L.3, Fucks, E. 4, 5, Aguirre. M.2, 5, y Schnack, E.5, 6
1, Museo de La Plata, Paseo del Bosque s/n, 1900 La Plata 2, CONICET 3, Centro de Geología de Costas y del Cuaternario, Universidad de Mar del Plata 4, Facultad de Ciencias Agrarias y Forestales y Latyr, Facultad de Ciencias Naturales y Museo, UNLP 5, Facultad de Ciencias Naturales y Museo, UNLP 6, Comisión de Investigaciones Científicas de la Provincia de Buenos Aires La Formación Pascua (Pleistoceno marino; +6-8m s.n.m., 120 ka?, MIS 5e ?) en su localidad tipo (Puente de Pascua) presenta los afloramientos más completos del NE bonaerense, compuestos por secuencias arenosas, arcillosas y limosas con una importante concentración de conchillas de moluscos, esqueletos de otros invertebrados, microfósiles (foraminíferos y ostrácodos) y en la parte superior restos de mamíferos. La revisión de aspectos paleontológicos, estratigráficos y geomorfológicos, en su conjunto, aporta características inusuales para el área Pampeana. Esta localidad se encuentra ubicada a 35º 55’ 37” S y 57º 43’ 7,6” W en el sector costero de la Bahía Samborombón, sobre un sustrato de limos loessoides de la Fm. Pampeano (continental) constituida por limos arcillosos a arcillo-arenoso de color castaño a castaño oscuro con importantes contenidos de tosca. La secuencia marina, que indica el límite más interno de la ingresión pleistocena en toda el área de la Cuenca del Salado, está representada por sedimentos arenosos a arcillosos. El contenido fosilífero de invertebrados es típico de ambientes litorales estuariales y consiste mayoritariamente de conchillas de moluscos, en su mayoría sueltas y enteras (sin cementación, fragmentación ni recristalización): valvas completas y en posición de vida de Tagelus plebeius asociadas con Mactra isabelleana, Pitar rostrata, Ostrea sp. y Corbula patagonica (Bivalvia) y Adelomelon brasiliana, Buccinanops cochlidium, B. globulosus, Olivancillaria spp., Dorsanum moniliferum, Terebra sp. y Biomphalaria peregrina (continental) (Gastropoda). La microfauna está compuesta por al menos 90 taxones y presenta variado estado de preservación, parte del material está recristalizado y otra parte presenta signos de disolución. Entre los foraminíferos predominan Buccella peruviana campsi, Ammonia beccarii y Discorbis williamsoni y Elphidium discoidale. Entre los ostrácodos, mucho más escasos, no existen especies dominantes. La mayoría de los taxones son marinos litorales (e.g., Frenguellicythere argentinensis, Oculocytheropteron spp., Cytheretta punctata) pero aparecen otros continentales frecuentes en ambientes estuariales (Cyprideis salebrosa, C. multidentata, Limnocythere staplini y L. solum). Los datos aportados por la microfauna presente sumados a los obtenidos a partir de la malacofauna, indicarían paleoambientes marginales marinos y la ocurrencia de cambios ambientales, principalmente de salinidad, con respecto al litoral moderno vecino. Éstos están directamente relacionados con cambios climáticos y eustáticos globales que causaron el desplazamiento del gradiente de salinidad horizontal del Río de La Plata tal como se observa en la actualidad.

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Estudios biométricos en ostras del Neógeno marino de Patagonia, Argentina
Farinati, E.A. y Fernández, A.L.
Departamento de Geología, Universidad Nacional del Sur. [email protected] CONICET. Departamento de Geología. ana.ferná[email protected] Los acantilados de la Formación Río Negro, aflorantes en la zona costera atlántica del noreste de la provincia de Río Negro, Argentina, se caracterizan por grandes concentraciones de Ostrea patagonica d´Orbigny procedentes de una secuencia depositacional que incluye el cortejo transgresivo y el cortejo de nivel alto y ofrecen una excelente oportunidad para estudiar las condiciones paleoambientales durante el intervalo Mioceno superior-Plioceno inferior. Las ostras son organismos sésiles y filtradores que segregan sus valvas carbonatadas en equilibrio con el agua marina registrando su relación con el medio y los posibles eventos estresantes que sufrieron a lo largo de su vida. Así, en ambientes contaminados pueden incorporar elementos tóxicos causantes de malformaciones (engrosamiento de la valva derecha, cámaras, etc) como se ve en muchos estuarios europeos donde los cultivos de ostras presentan malformaciones debido a la presencia en el medio del TBT (biocida de la pintura de los barcos). En base al Principio del Actualismo, la intención fue analizar si en las grandes concentraciones de ostras del Neógeno atlántico pudo ocurrir algo similar, ya sea en respuesta a la manifiesta actividad bioerosiva o a la actividad volcánica. Se analizaron 30 ejemplares de O. patagonica de ambos niveles cuyas valvas derechas tienen en promedio 130 mm de largo, 93 mm de ancho y 24 mm de espesor. Se midieron los índices biométricos: engrosamiento, peso y volumen con el objeto de detectar valores anómalos que pudieran indicar estrés ambiental o contaminación También se obtuvieron datos de la especie actual alóctona (Crassostrea gigas) de la localidad de Pocitos distante 90 km de los afloramientos neógenos a los efectos de una comparación. El Indice de Engrosamiento (normal: mayor a 10) analizado para la valva derecha arrojó valores anómalos (promedio:5,77) mientras que los índices de peso y de volumen dieron valores ambiguos en comparación con los ejemplares actuales. Se realizaron cortes en sentido longitudinal según el eje de crecimiento de valvas derechas y secciones delgadas de las mismas para un futuro análisis microestructural. En ambos niveles se observó que las valvas están afectadas por la actividad bioerosiva de organismos perforantes (bivalvos, esponjas, anélidos) la que pudo haber sido la causa del estrés desencadenante de un engrosamiento anormal de naturaleza defensiva, siempre y cuando la bioerosión haya sido en vida del organismo. Otra causa de estos valores anómalos podría ser la presencia en la secuencia de materiales volcánicos que pudieran contaminar la composición del agua-sedimento donde se desarrollaron las ostras.

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Las familias Scyliorhinidae, Triakidae, Hemigaleidae y Carcharhinidae (Neoselachii, Galeomorphii) del Plioceno inferior de la Formación Arenas de
Huelva, España
García, E.X.M.1, Antunes, M. T. 2,3, Balbino, A.C. 4, Civis, J.5, Ruiz, F.6 y Rico-García, A.5
1 Departamento de Botánica y Zoología, Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara, Km 15.5 carretera Guadalajara-Nogales, Las Agujas, Zapopan, c.p. 45110. Apdo. postal 1-139, Jalisco, México; [email protected] 2 Centro de Investigação em Ciência e em Engenharia Geológica da Universidade Nova de Lisboa. 3 Academia das Ciências de Lisboa, Rua da Academia das Ciências, 19, 1249-122 Lisboa, Portugal; [email protected] 4 Departamento de Geociências, Universidade de Évora, Apartado 94, 7002-554 Évora, Portugal; [email protected] 5 Departamento de Geología, Universidad de Salamanca, Plaza de la Merced, s/n, 37008-Salamanca, España; [email protected] 6 Departamento de Geodinámica y Paleontología, Facultad de Ciencias Experimentales, Universidad de Huelva, Avda. de las Fuerzas Armadas, s/n, 21071-Huelva, España; [email protected] Se estudiaron dientes de seláceos provenientes del Plioceno inferior de la Formación Arenas de Huelva, como parte del estudio de Tesis Doctoral realizada por García (2008). La Formación Arenas de Huelva, descrita por Civis et al. (1987), se inicia con un nivel de limos glauconíticos, rico en dientes de seláceos y es considerado como el marcador Mioceno-Plioceno (5,33 Ma) para el suroeste de la cuenca del Guadalquivir (González-Delgado et al., 2004). Por encima aparecen una serie de niveles arenosos con frecuentes intercalaciones de lumaquelas de moluscos, en donde se han llegado a citar más de 300 especies de moluscos (González-Delgado, 1983). 31 taxones de seláceos fueron determinados a partir del estudio de 8 secciones en las localidades de Niebla, Bonares, Moguer y El Rompido (Fig. 1).
Fig. 1.- Localidades de estudio en el sur de la provincia de Huelva.
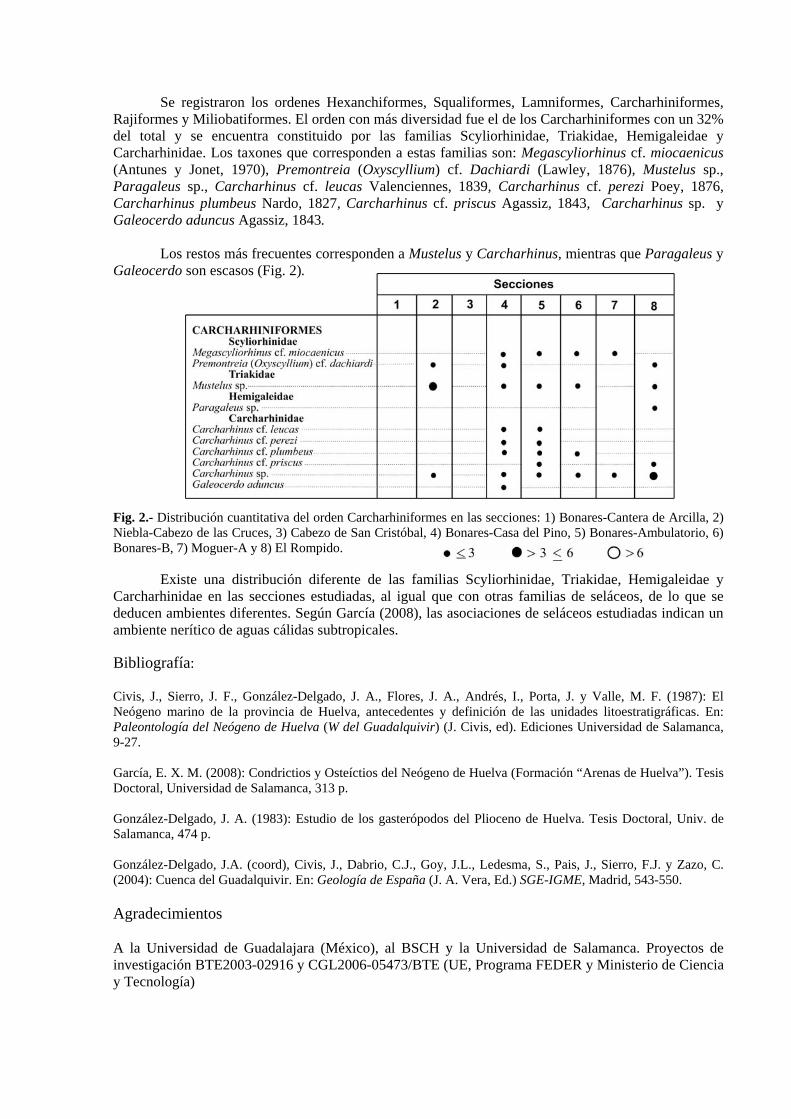
Se registraron los ordenes Hexanchiformes, Squaliformes, Lamniformes, Carcharhiniformes, Rajiformes y Miliobatiformes. El orden con más diversidad fue el de los Carcharhiniformes con un 32% del total y se encuentra constituido por las familias Scyliorhinidae, Triakidae, Hemigaleidae y Carcharhinidae. Los taxones que corresponden a estas familias son: Megascyliorhinus cf. miocaenicus (Antunes y Jonet, 1970), Premontreia (Oxyscyllium) cf. Dachiardi (Lawley, 1876), Mustelus sp., Paragaleus sp., Carcharhinus cf. leucas Valenciennes, 1839, Carcharhinus cf. perezi Poey, 1876, Carcharhinus plumbeus Nardo, 1827, Carcharhinus cf. priscus Agassiz, 1843, Carcharhinus sp. y Galeocerdo aduncus Agassiz, 1843. Los restos más frecuentes corresponden a Mustelus y Carcharhinus, mientras que Paragaleus y Galeocerdo son escasos (Fig. 2). Fig. 2.- Distribución cuantitativa del orden Carcharhiniformes en las secciones: 1) Bonares-Cantera de Arcilla, 2) Niebla-Cabezo de las Cruces, 3) Cabezo de San Cristóbal, 4) Bonares-Casa del Pino, 5) Bonares-Ambulatorio, 6) Bonares-B, 7) Moguer-A y 8) El Rompido.
Existe una distribución diferente de las familias Scyliorhinidae, Triakidae, Hemigaleidae y Carcharhinidae en las secciones estudiadas, al igual que con otras familias de seláceos, de lo que se deducen ambientes diferentes. Según García (2008), las asociaciones de seláceos estudiadas indican un ambiente nerítico de aguas cálidas subtropicales. Bibliografía: Civis, J., Sierro, J. F., González-Delgado, J. A., Flores, J. A., Andrés, I., Porta, J. y Valle, M. F. (1987): El Neógeno marino de la provincia de Huelva, antecedentes y definición de las unidades litoestratigráficas. En: Paleontología del Neógeno de Huelva (W del Guadalquivir) (J. Civis, ed). Ediciones Universidad de Salamanca, 9-27. García, E. X. M. (2008): Condrictios y Osteíctios del Neógeno de Huelva (Formación “Arenas de Huelva”). Tesis Doctoral, Universidad de Salamanca, 313 p. González-Delgado, J. A. (1983): Estudio de los gasterópodos del Plioceno de Huelva. Tesis Doctoral, Univ. de Salamanca, 474 p. González-Delgado, J.A. (coord), Civis, J., Dabrio, C.J., Goy, J.L., Ledesma, S., Pais, J., Sierro, F.J. y Zazo, C. (2004): Cuenca del Guadalquivir. En: Geología de España (J. A. Vera, Ed.) SGE-IGME, Madrid, 543-550. Agradecimientos A la Universidad de Guadalajara (México), al BSCH y la Universidad de Salamanca. Proyectos de investigación BTE2003-02916 y CGL2006-05473/BTE (UE, Programa FEDER y Ministerio de Ciencia y Tecnología)

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Plio-Pleistocene SST reconstruction: essay for the Southwest South Atlantic
García Chapori, N. & Laprida, C. Departamento de Ciencias Geológicas, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Consejo Nacional de Investigaciones Científicas y Técnicas. Ciudad Autónoma de Buenos Aires, Argentina. [email protected]
In the oceans sea surface temperature (SST) and salinity variations are ultimately driven by climate so that paleoceanographic reconstructions allow to recognize climate variations.
The reconstruction of ocean history has employed a large variety of methods. Planktic foraminifera have been extensible used because of their widespread geographical and geological occurrence (Hale and Plaufmann, 1999). Their spatial distribution is controlled by physiological requirements related to salinity, productivity, feeding preferences and specially temperature, the dominant factor (Bé and Hamilton, 1967; Bé and Tolderlund, 1971). Planktic foraminifera are excellent proxies of Quaternary climate because Recent and Quaternary faunas are practically the same and have similar ecologic requirements, so one is able to draw conclusions with respect to Quaternary environments by comparing Quaternary faunas with Recent ones. This is the base of the Modern Analog Techniques.
The SP1251 core, 877 cm long, is located at 38°29.7’S-53°40.7’W at 3400m depth in the southwestern South Atlantic, in the Convergence Zone. This is an excellent place to perform paleoceanographic reconstructions because this zone is sensitive to climatic change.
Nineteen samples were recovered and their foraminifers analyzed. The age estimation of the core is based on the presence of G. tosaensis at 60 cm which, even in low proportion, indicates Upper Pliocene-Lower Pleistocene (Boltovskoy, 1973; Watanabe et al., 2005). Assemblages were compared with a data set of modern samples (SPECMAP archive #1) to determine the paleotemperatures based on the most similar modern samples (MAT) using PaleoAnalogs 3.0 software (Theron, 2004).
Southern Ocean cores are characterized by alternations of cold-water faunas composed almost entirely of left-coiling G. pachyderma, with warmer water faunas which include several other species (Kennett, 1970), and this core is not an exception.
The core reflects a prominent cold period with summer SSTs between 5-11°C and winter SSTs between 2.6-6.4°C. Nowadays, the summer SST in the area is 20-22°C (Martinez Avellaneda, 2005; Pathfinder, NOAA) and the winter SST is 8-12°C (Pathfinder, NOAA). A moment of warmer waters was observed at 60-74 cm depth, with summer SSTs of 12-14°C and winter SSTs of 7-8°C.
This reconstruction fits well with the Plio-Pleistocene boundary climatostratigraphy event proposed by Shackleton (1997) (around 2.5 Ma), and ratified by Mauz (1998) as a level that marks the onset of the prominent cold period around the marine isotope stages 100, 98 and 96.

References: Bé, A.W.H. and Hamilton, W.H. 1967. Ecology of Recent planktonic foraminifera. Micropaleontology
13, 87–106. Bé, A.W.H. and Tolderlund, D.S. 1971. Distribution and ecology of living planktonic foraminifera in
surface waters of the Atlantic and Indian Oceans. In: Funnell, B.M. and Riedel, R.R. (eds.). The Micropaleontology of the Oceans. Cambridge University Press, New York, pp 105-149.
Boltovskoy, E. 1973. Estudios de testigos submarinos del Atlántico Sudoccidental. Revista del Museo Argentino de Ciencias Naturales, Geología 7 (4), 215-340.
Hale, W. and Pflaumann, U. 1999. Sea-Surface Temperature Estimations Using a modern Analog Technique with Foraminiferal Assemblages from Western Atlantic Quaternary Sediments. In: Fischer, G. and Wefer, G. (eds.). Use of Proxies in Paleoceanography: Examples from the South Atlantic. Springer, Berlin Heidelberg, pp 69-90.
Kennett, J.P. 1970. Pleistocene paleoclimates and foraminiferal biostratigraphy in subantartic deep-sea cores. Deep-Sea Research 17, 125-140.
Martinez Avellaneda, N. 2005. El Ciclo Anual y Variabilidad de baja frecuencia de la Temperatura Superficial del Mar en el Océano Atlántico Sudoccidental, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, 139pp.
Mauz, B. 1998.The onset of the Quaternary: a review of new findings the Pliocene-Pleistocene chronostratigraphy. Quaternary Science Reviews 17, 357-364.
Pathfinder Proyect. National Environmental Satellite, Data, and Information Service (NESDIS; NOAA). National Oceanographic Data Center (NODC; NOAA), University of Miami's Rosenstiel School of Marine and Atmospheric Sciences (RSMAS), Physical Oceanography Distributed Active Archive Center (PO.DAAC; NASA).
Shackleton, N.J. 1997. The deep-sea sediment record and the Pliocene-Pleistocene boundary. Quaternary International 40, 33-35.
SPECMAP Archive #1. IGBP PAGES/World Data Center-A for Paleoclimatology Data Contribution Series # 90-001. NOAA/NGDC Paleoclimatology Program, Boulder CO, USA.
Theron, R. 2004. Visual Analytics of Paleoceanographic Conditions. Prepare for Return of el Nio. In: Lean, G. Environment Editor, The Independent.
Watanabe, S., Totah, V., Laprida, C., García, N., Violante, R. y Parker, G. 2005. Estudio preliminar de los foraminíferos planctónicos y bentónicos de un testigo del talud bonaerense, Atlántico sudoccidental. 16º Congreso Geológico Argentino, Actas 3, 357-364.

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Malacofauna continental holocena y paleoambientes en Villa Valle María (Departamento Diamante, Entre Ríos, Argentina)
González, C.A.1, Aguirre, M.L.2,5, Miquel, S.E.3,5, Kröhling, D.4,5, F. Zucol, A.1,5 y Brea, M.1,5
1Laboratorio de Paleobotánica, Centro de Investigaciones Científicas, Diamante. CICYTTP-Diamante, Dr. Materi y España SN, (E3105BWA) Diamante, Entre Ríos, Argentina. [email protected]; [email protected], [email protected] 2Facultad de Ciencias Naturales y Museo, INGEA, Lab. 6 (Malacofauna Cuaternaria), calle 64 N°3, (1900) La Plata. Buenos Aires, Argentina. [email protected] 3Museo de Ciencias Naturales “Bernardino Rivadavia”, Av. Ángel Gallardo 470, (1405) Buenos Aires, Argentina. [email protected] 4Facultad de Ingeniería y Ciencias Hídricas, Universidad Nacional Del Litoral, C.C. 217 (3000) Santa Fe, Argentina. [email protected] 5Consejo Nacional de Investigaciones Científicas y Técnicas. La malacofauna continental preservada en sedimentos loéssicos continentales del Holoceno de Villa Valle María (Departamento Diamante, Entre Ríos, Argentina) corresponde a un depósito sedimentario datado en 2490 ± 45 años 14 C A.P. sobre conchillas de Bulimulus sp. En el perfil descripto en la localidad fosilífera se efectuó la identificación y revisión taxonómica de los moluscos recuperados, el análisis cuantitativo de los distintos taxones reconocidos en los niveles muestreados, así como su análisis paleoecológico y de distribución. Los ejemplares encontrados corresponden únicamente a gastrópodos, predominantemente terrestres asociados a formas palustres. Los ejemplares pertenecen a cinco familias (Pupillidae, Succineidae, Ferussaciidae, Orthalicidae y Scolodontidae) y a 10 especies: Gastrocopta nodosaria (d’Orbigny, 1835), Succinea meridionalis d´Orbigny, 1846, Cecilioides consobrina (d’Orbigny, 1837), Bulimulus apodemetes (d’Orbigny, 1835), B. bonariensis bonariensis (Rafinesque, 1833), Naesiotus pollonerae (Ancey, 1897), N. deletangi (Parodiz, 1946), Miradiscops brasiliensis (Thiele, 1927) y Scolodonta semperi Doering, 1875. Se da a conocer el primer registro de Miradiscops para la provincia de Entre Ríos. La asociación de gastrópodos (típica de hábitats con cobertura vegetal y hojas y troncos como sustrato) es equivalente a la que se encuentra actualmente en el sur de Entre Ríos (Distrito Mesopotámico del Dominio Subtropical) caracterizado por un escenario continental de estepas templado-cálidas. No obstante, el registro de S. meridionalies, especie típica de ambientes con contenido de humedad más elevados o pantanos, sugiere condiciones ambientales locales. Los datos aportados en este trabajo brindan evidencia adicional respecto de la existencia de un episodio eólico que habría producido el retrabajo y subsiguiente acumulación de sedimentos superficiales deflacionados sobre las llanuras centrales de la Argentina durante un período seco entre 3.500 y 1.400 años A.P.

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Malacology of the Pliocene Atlantic basins of Northern Morocco
González Delgado, J.A.1,a, Ben Moussa, A.2, Civis, J.1,b & Rico García, A.1,c 1: Dpto. Geología, Facultad de Ciencias, c/Parque s/n, 37008 Salamanca (España) a: [email protected]; b: [email protected]; c: [email protected] 2: Dpt. Géologie, Faculté des Sciences, Univ. A. Essaadi, B.P. 2121, Tétouan (Morocco) ; [email protected] In the Atlantic coast of Morocco, from the Gibraltar strait to Essaouira (35 to 30º N), three marine Neogene areas are characterized by their high malacological richness: the Asilah basin, the western edge of the Bou Regreg basin, and the Safi area. The Asilah basin is a small siliciclastic basin (100 square km and up to 25 m thickness) of Upper Zanclean age (N-19). Sediments consist of a succession of yelow very fine-grained sands, highly bioturbated, and grey silts. Paucispecifical event-type shell concentrations of Pinna, Corbula, Ostrea, Acanthocardia, Modiolus, Pecten and Palliolum excisum, alternate with other composite multispecifical shell concentrations of more complex taphonomic signature, and with levels of scarce malacofauna. Molluscan diversity reach the 110 taxa (73 of them corresponding to bivalves, wich are also the most abundant molluscs). Inicial transgressive and posterior onshore environments are interpreted on the basis of the palaeoecological requirements of the malacofauna. The oxygen isotopic signal of P. excisum records changes of until 4ºC on the sea surface temperature where the bivalve grew. The pliocene sediments (Upper Zanclean) of the Bou Regreg basin (known as South Rifian Corridor which connected the Atlantic and the Mediterranean realms in Upper Miocene) outcrop in a non-continuous mode from the coastline to 100 Km eastward, and 20 m of thickness. They consist in grey silts with scarce malacofauna and yellow fine-sands very rich in molluscs, arranged in composite multispecifical shell concentrations, being the Oued Arjat and Oued Kell sections the most representatives. Diversity reach the 230 taxa, and is higher in Gastropods (124 taxa), but the most abundant are the Bivalves. The malacofauna records circalittoral settings affected by distal storms westward, and progresively more littoral environments eastward. In the Safi area, the pliocene sediments are calcareous. They are located in scarce isolated outcrops arranged 15 km in a NE-SW direction, and up to 21 m of thickness. They consist in grainstones and rudstones dominated by bioclasts of molluscs, barnacles, foraminifers, algae, echinoids and bryozoans, arranged in fining-upward sequences highly bioturbated. Paucispecifical event-type shell concentrations are common, very rich in Ostrea, or Pecten, and alternate with levels of articulated moulds of infaunal bivalves in life-position like Panopea and Lutraria. The molluscan diversity is low (30 taxa), due to intense dissolution of the aragonitic shells. Taphonomic signatures suggest a latitudinal variety of marine nearshore environments. Acknowledgments: financial support of the European Union and the Spanish Ministerio de Ciencia y Tecnología, Project CGL2006-05473BTE

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Quaternary mollusk assemblages from the northern Atlantic coast of Tierra del Fuego (southern South America): taphonomy and paleoecology.
Gordillo, S.1 & Isla, F.I.2
1CICTERRA, CONICET. Centro de Investigaciones Paleobiológicas (CIPAL), Universidad Nacional de Córdoba. Córdoba, Argentina. [email protected]
2CONICET. Centro de Geología de Costas y Cuaternario, Universidad Nacional de Mar del Plata. Mar del Plata, Argentina. [email protected]
Pleistocene and Holocene marine deposits along the northern Atlantic coast of Tierra del Fuego represent different transgressive-regressive phases throughout the Quaternary. Previous works in this region provided data which permitted chronological reconstruction of these sequences. They were deposited during different sea level highstands corresponding to oxygen isotopic stages (OIS) from the Middle and Late Pleistocene to the Holocene.
The objective of this work is to characterize the typical mollusk assemblages developed along the northern Atlantic coast of Tierra del Fuego and, on this basis, to analyze their relationships with environmental changes during the Quaternary. Fossil bulk samples were taken from 5 localities (Laguna Arcillosa, Laguna Las Vueltas, La Sara, Río Chico y San Pablo) previously dated in age, and studied from geological and geomorphological viewpoints. Different taphonomic attributes (articulation, fragmentation, surface alteration, bioerosion and shell coloration) were recorded for each specimen from each sample, and then were averaged over the entire sample. These signatures were observed in the field and/or in the laboratory. Results show that the faunal composition of Quaternary marine terraces consists mostly of bivalve and gastropod mollusk shells. All the species recovered are extant taxa, living in the Magellan Province. Although gastropods (15 species) outnumber bivalves (5 species), bivalves are more abundant. The highest richness of gastropods and the dominance of bivalves appear to be the result of natural ecological conditions of the fauna in the region. Taphonomic alteration of these shells is related to different energies and periods of exposure within each environment. When comparing the Pleistocene assemblages with the Holocene and modern ones, some differences arise: the first ones exhibit lower diversity and are mostly represented by infaunal bivalves (venerids), while the second ones are more diverse, with more presence of gastropods, and dominated by epifaunal taxa (mytilids). They represent typical shallow benthic communities which developed within the Atlantic fuegian marine realm during the Quaternary. The fact that they are similar to the benthic communities living today in the region reinforces the theory that the climatic conditions have been maintained without significant changes since at least the Middle Pleistocene. However, the local disappearance of Retrotapes in postglacial benthic communities from the studied area is interpreted on the basis of local environmental conditions affecting wave dynamics and sediment deposition, and not to any significant climatic change.

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Cambios paleoambientales en el Cuaternario de la costa Norte del Río Negro (paraje Playa Chica), provincia de Buenos Aires, Argentina.
Gutiérrez Téllez, B.1, Schillizzi, R. 2, Luna, L. 1, Caballero, N.3 y Nass, Y.3
1Departamento de Geología, Universidad Nacional del Sur, Bahía Blanca. E-mail: [email protected] 2 IADO, CONICET, 3Departamento de Biología, Universidad Nacional del Sur, Bahía Blanca
El área estudiada corresponde al paraje denominado Playa Chica (S 40º 59´ 17”, W 62º 47´ 43”) cercano a la desembocadura del Río Negro, en la provincia de Buenos Aires. El objetivo planteado es la caracterización sedimentológica, mineralógica y micropaleontológica a partir del análisis de asociaciones de diatomeas. Se estudió un perfil de 7 m de potencia ubicado en una barranca a orillas del río, del cual se extrajeron 19 muestras representativas de los distintos estratos. Los sedimentos, de colores pardo a pardo rojizos, moderadamente consolidados, varían desde limo arcillosos hasta areno limosos en menor proporción. La mineralogía es similar en todos los niveles y esta representada por cuarzo, plagioclasa, feldespato potásico, mafitos, trizas de vidrio volcánico, carbonatos y óxidos de Fe. Se analizaron los porcentajes de carbonato de calcio y se observó una variación ascendente en los diferentes niveles desde 0,5 % hasta 1,7 %. La materia orgánica, cuyos valores promedio están entre 0,7 y 1,5 %, está representada por restos vegetales, espículas de esponjas, cistos de crisostomatáceas y frústulos de diatomeas. En la base de la barranca se observan restos caulinares en sección transversal y longitudinal, los que se presentan en forma de nódulos oxidados y carbonosos. Los frústulos de diatomeas se observaron con lupa binocular en sedimentos sin tratamiento químico y con microscopio óptico en muestras procesadas según técnicas diatomológicas apropiadas. Se identificaron especies como Hyalodiscus subtilis Bailey y Surirella striatula Turpin, bentónicas y mesohalobias junto con Actinoptychus splendens (Shadbolt) Ralfs, Paralia sulcata (Ehremberg) Cleve, Hyalodiscus laevis Ehrenberg, Coscinodiscus radiatus Ehrenberg, Rhaphoneis amphiceros (Ehrenberg) Ehrenberg y Rhaphoneis surirella (Ehrenberg) Grunow, planctónicas y polihalobias/mesohalobias. Todas las asociaciones de diatomeas determinadas para el perfil muestran una depositación rítmica. El nivel inferior corresponde a una planicie de marea, caracterizada por sedimentos finos y vegetación asociada. En el nivel superior los depósitos presentan estratificación poco marcada. Ambos ambientes exhiben variaciones cíclicas en la abundancia de frústulos, contenido de carbonato de calcio y materia orgánica total. Por lo tanto, estas características permiten determinar un paleoambiente de transición, entre el dominio marino y continental, relacionado con el mar de manera permanente o episódica. La presente contribución forma parte del proyecto de investigación PICTO 905 de la Agencia Nacional de Promoción Científica y Tecnológica, en convenio con al Universidad Nacional del Sur de la ciudad de Bahía Blanca, provincia de Buenos Aires.

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
The Serravallian-Tortonian boundary in the Lower Tagus Basin (Portugal) and the new GSSP of the Tortonian Stage
Legoinha, P.
Centro de Investigação em Ciência e Engenharia Geológica (CICEGe), Faculdade de Ciências e Tecnologia, Universidade Nova de Lisboa, Quinta da Torre, P-2829-516 Caparica, Portugal; [email protected]
A comprehensive chronostratigraphical study of the Miocene from the distal sector of the Lower Tagus Basin has been carried out by researchers of the CICEGe. The author has studied the planktonic foraminifera.
The succession, of mostly marine with some interfingering continental beds, ranges from Early (Aquitanian) to Late Miocene (Tortonian). The stratigraphic and environmental framework is supported on data concerning foraminifera, ostracoda, vertebrates, palinomorphs. K-Ar glauconite ages, 87Sr/86Sr dating, and paleomagnetism. A fairly accurate local time scale was obtained and ten depositional sequences were characterized.
The lower boundary (regional disconformity D9) of the depositional sequence T1 was dated at 11.6 Ma because of the last occurrences of Globigerinoides subquadratus under the disconformity (LAD: 11,8 Ma; Berggren et al., 1995) and of Globorotalia mayeri above the disconformity (LAD: 11,4 Ma; Berggren et al., 1995). Furthermore, a Sr isotopic age of a pectinid shell from the lowermost part of this depositional sequence yielded 11.6 (+2, - 0.6) Ma. Meanwhile, the new Tortonian Stage GSSP was astronomically dated at 11,608 Ma Hilgen et al. (2005). It can thus be correlated with the beginning of the T1 depositional sequence. Biostratigraphic and chronostratigraphic data of two geological sections related to this boundary are presented and compared with the Tortonian Stage GSSP. Main conclusions are (Legoinha, 2008):
a) depositional sequence T1 can be correlated with the 3rd order eustatic cycle 3.1 of Haq et al.; b) disconformity D8 represents a hiatus between 11,6 Ma and 12,7 Ma which was not recognized
in previous works; c) the Serravallian-Tortonian boundary is shown to be present between the last occurence of
Globigerinoides subquadratus and the last occurence of Globorotalia mayeri both in the Mediterranean and in the Iberian Atlantic coast;
d) first regular occurrence of Globigerina apertura is above the last occurence of Globorotalia mayeri, in the Lower Tortonian. The same situation is reported in the Mediterranean;
e) Globigerinopsis aguasayensis, an index species of the Mediterranean middle Miocene, attains the basal Tortonian (N15).
References Hilgen, F., Aziz, H.A., Bice, D., Iaccarino, S., Krijgsman, W., Kuiper, K., Montanari, A., Raffi,I., Turco, E. & Zachariasse, W.-J. (2005) – The Global boundary Stratotype Section and Point (GSSP) of the Tortonian Stage (Upper Miocene) at Monte Dei Corvi. Episodes, v. 28, no. 1, 6-17. Legoinha, P. (2008) - The Serravallian-Tortonian boundary in the Lower Tagus Basin (Portugal) and the new GSSP of the Tortonian Stage. e-Terra, vol. 6, n. 1. (http://metododirecto.pt/ojs/index.php/e-terra).

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Síntesis del Neógeno de la cuenca del Duero, noroeste de España
Luengo, J.1, Armenteros, I.2, Montes. M.1, Nozal, F.1, Corrochano, A.2,
López Olmedo, F.1 y Rodriguez, J.1
1 Departamento de Infraestructura Geocientífica. Área de Cartografía Geocientífica. IGME. C/ Calera 1. 28760-Tres Cantos. Madrid. [email protected]; [email protected]; [email protected]; fa.lopez@igme. 2 Departamento de Geología, Facultad de Ciencias, Universidad de Salamanca, 37071 Salamanca, [email protected]
La cuenca del Duero es una de las mayores cuencas cenozoicas de la península Ibérica, con una extensión de unos 50.000 km2. Representa una depresión intraplaca rellena de sedimentos continentales de edad cenozoica, depositados en un régimen endorreico. La cuenca tuvo una evolución compleja y se originó por los esfuerzos que afectaron a la placa Ibérica durante la orogenia Alpina, como consecuencia de la convergencia de las placas Eurásica y Africana. Durante buena parte del ciclo paleógeno y hasta el Mioceno inferior se generaron varios depocentros en el margen norte (junto a la Cordillera Vasco-Cantábrica), en la franja oriental (junto a la Cordillera Ibérica) y en la zona sur (junto al Sistema Central, sobre todo en su mitad oriental), que se constituyeron como cuencas de antepaís respecto a estas cordilleras. Junto al margen occidental, constituido por el basamento varísco del Macizo Ibérico, se produjo durante todo el terciario una sedimentación ligada al basculamiento hacia el este y a la fracturación por fallas de dicho zócalo.
El relleno de la cuenca se ha dividido en tres secuencias principales: 1) finicretácica-paleocena; 2) eocena-oligocena; y 3) neógena (Corrochano y Armenteros, 1989). La extensa representación cronoestratigráfica de la sucesión neógena, junto a su escasa deformación y su rico repertorio de facies hacen que sea una de las cuencas endorreicas neógenas de la península más interesantes para estudios cronoestratigráficos, de estratigrafía secuencial y de sistemas sedimentarios fluvio-lacustres. Los estudios previos de carácter regional (Portero et al., 1982; Armenteros et al., 2002; Alonso Gavilán et al., 2004; Nozal et al., 2006), los trabajos de cartografía regional (Armenteros y Mediavilla, 1997; Pineda et al., 2006), la correlación de unidades litoestratigráficas significativas y la correlación de discontinuidades mayores con distribución a escala de cuenca revelan dentro de la secuencia principal neógena, 4 secuencias menores. El hallazgo de nuevos yacimientos fósiles, el refinamiento de la posición estratigráfica de los ya existentes y las aportaciones magnetoestratigráficas (Montes et al., 2006) permiten una división cronoestratigráfica más detallada que hasta ahora.

La primera secuencia neógena está comprendida entre el Mioceno inferior (Rambliense) y el Mioceno medio (Aragoniense medio). En ella quedan agrupadas un conjunto de litofacies carbonatadas, y equivalentes detríticos laterales (U. Roja de Toro, U. de Villalba de Adaja, U. de Turégano). La segunda secuencia abarca desde el Aragoniense medio hasta el superior. En la zona central está representada por las facies Dueñas y Tierra de Campos, entre otras unidades, y en el sector centro-oriental la Unidad Detrítica de Aranda. Estas aparecen culminadas en esta última región por un nivel bastante continuo y de gran expresión morfológica (calizas del Páramo Inferior) presente en el sector oriental. En los bordes, se encuentran sus equivalentes laterales, constituidos por litofacies detríticas gruesas depositadas por abanicos aluviales (conglomerados de Montejo de la Vega, facies de Grijalva-Villadiego…).
La tercera secuencia neógena comprende el Aragoniense superior y el Vallesiense. En el centro de cuenca está constituida por la facies Cuestas y, por encima, las calizas y dolomías que soportan el páramo culminante en el centro de cuenca. Hacia el sector oriental, éste constituye un litosoma intermedio (“Páramo Intermedio” de Nozal et al., 2006) de la sucesión terrígeno-carbonatada entre las calizas del Páramo Inferior y Calizas del Páramo Superior que culminan generalmente la sucesión miocena en dicho sector. Por último la cuarta secuencia, de escasa representación y difícil correlación por presentarse en zonas aisladas, está datada en el tránsito Mioceno-Plioceno. Está constituido por un conjunto de facies detríticas que suelen culminar con facies calcáreas, lacustres y palustres (i.e., conglomerados de Tariego en el centro de cuenca y otras secuencias detritico-carbonatadas que aparecen ligadas a los bordes en el sector oriental y septentrional).
Agradecimientos: El presente trabajo ha contado con el soporte económico del proyecto CGL 2008-05584-CO2-01/BTE y del IGME.
Bibliografía Alonso Gavilán, G., Armenteros, I., Carballeira, J., Corrochano, A., Huerta, P. y Rodríguez, J.M. 2004. Cuenca
del Duero. En: Geología de España (J.A. Vera, Ed.), SGE-IGME, Madrid, pp. 550-556. Armenteros. I. y Mediavilla, B., 1997. Terciario y Cuaternario. En: Mapa Geológico y Minero de Castilla y León.
SIEMCALSA, Valladolid, .pp.. 54-82. Armenteros, I., Corrochano, A., Alonso Gavilán, G., Carballeira, J. y Rodríguez, J.M., 2002. Duero basin
(northern Spain). In: Gibbons, W. and Moreno, M.T. (eds) The Geology of Spain. Geological Society, London,. pp. 309-315.
Corrochano, A., y Armenteros, I., 1989. Los sistemas lacustres de la Cuenca del Duero. En: Sistemas Lacustres Cenozoicos de España, Acta Geol. Hispánica, 24, (3/4), 259-279.
Nozal, F., Montes, M., López Olmedo, F., y Luengo, J., 2006. Los “tres páramos” del SE de la Cuenca del Duero. Geo-Temas, 9, 189-192.
Montes, M., Beamud B., Garcés, M., Nozal F., López-Olmedo, F. y Luengo, J. (2006): Datos preliminares de la magnetoestratigrafía del sureste de la Cuenca del Duero. Geo-Temas, 9: 171-175.
Pineda, A., Nozal, F., Montes, M., López Olmedo, F., y Martín-Serrano, A., J, 2006. Mapa Geológico de la Cuenca del Duero. Geo-Temas, 9, 193-196.
Portero García, J. M., Del Olmo, P., Olivé, A. (1983): El Neógeno de la transversal norte-sur de la Cuenca del Duero. In: Libro Hom. J.M: Ríos, Geología de España. (Comba, J.A., Ed.). IGME. II: 494-502.

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Resultados preliminares del contenido paleoictiológico de la sección “Hacienda Clarevot” (Cuenca del Guadalquivir, Sevilla, España)
Maestre, M.V.1, García, E.X.M.2, Bajo, I.3, Cárdenas Carretero, J.4, Rico-García, A.5 y Civis, J. 5
1 C/ Nuestra Sra. del Aguila 51. 41500 Alcalá de Guadaíra. Sevilla). ([email protected])
2 Departamento de Botánica y Zoología, Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara, Km 15.5 carretera Guadalajara-Nogales, Las Agujas, Zapopan, c.p. 45110. Apdo. postal 1-139, Jalisco, México; [email protected]
3 C/ Fuerteventura 2. 41700 Dos Hermanas, Sevilla. ([email protected]) 4 Plaza España, 10 1ºD, 41700, Dos Hermanas (Sevilla). ([email protected]) 5 Departamento de Geología, Facultad de Ciencias, Universidad de Salamanca, Pz. Merced s.n, 37008, Salamanca, España; [email protected]; [email protected]
La sección “Hacienda de Clarevot”, situada en el término municipal de Alcalá de Guadaíra se ha dividido en cuatro tramos ó unidades, denominadas U1 a U4. La unidad U2, que presenta una elevada y diversa representación de ictiofauna, ha sido datada, por su contenido en foraminíferos planctónicos, como Messiniense inferior, cercana a los 6,43 Ma. Esta unidad consta de 1 m de arenas limo-arcillosas ocres-marronáceas con granos de glauconita. Junto a la ictiofauna, conservada con muy buena impronta tafonómica, son frecuentes los moldes internos de moluscos, trazas fósiles y fragmentos óseos de cetáceos. En las unidades restantes tan sólo se ha localizado en U3 un fragmento de Megaselachus megalodon (Agassiz in Charlesworth, 1837).
Se han recogido hasta la fecha 245 dientes, de los que 191 corresponden a condrictios y 54 a osteíctios. Los condrictios son especialmente abundantes, aportando 14 taxones cuyas familias dominantes son Odontaspididae (35,2%), Carcharhinidae (33,5%) y Lamnidae (26,2%), destacando respectivamente: Carcharias acutissima (Agassiz, 1843), Carcharhinus cf. plumbeus Nardo, 1827 y Cosmopolitodus hastalis (Agassiz, 1843). En menor proporción, se localizan taxones de las familias Squatinidae (2%), Otodontidae (1%), y Hemigaleidae (0,5%), así como del orden Myliobatiformes (1,6%). Todos los dientes de osteíctios pertenecen a la familia Sparidae, destacando la abundancia de Sparus cinctus (Agassiz, 1843) y Sparus neogenus Arambourg, 1927.
Desde el punto de vista paleoecológico se localizan, aunque escasamente, formas estenotérmicas de aguas tropicales/subtropicales como Hemipristis serra Agassiz, 1843 y Galeocerdo aduncus Agassiz, 1843, aunque son predominantes las especies nectónicas de aguas cálidas a templadas. Las especies más abundantes, especialmente Carcharias acutissima (Agassiz, 1843), así como datos de fauna y trazas fósiles asociadas, parecen indicar un rango batimétrico litoral/nerítico. El estudio y comparación del contenido paleoictiológico en otras secciones messinienses cercanas, que incluyen las unidades U1-U3, podría señalar un progresivo enfriamiento de las aguas y una disminución de la profundidad relativa durante el Messiniense, marcando un proceso regresivo. Cronoestratigráficamente, la edad dada por la asociación, aunque con un rango amplio, es compatible con la edad Messiniense inferior dada por los foraminíferos planctónicos.

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
The Palaeogene and Neogene of the Lower Tejo Basin (western Iberia, Portugal)
João Pais 1a, Pedro P. Cunha 2 & Paulo Legoinha 1b 1 Centro de Investigação em Ciência e Engenharia Geológica, Departamento de Ciências da Terra, Faculdade de Ciências e Tecnologia, Quinta da Torre, 2829-516 Caparica, Portugal. http://www.cicege.fct.unl.pt; a - [email protected]; b - [email protected] 2 Dep. Ciências da Terra e Instituto do Mar – CIC, da Universidade de Coimbra; proj. PPCDT/CTE-GEX/58120/2004; [email protected]
The importance of the Cenozoic basins of western Iberia is underlined by its Atlantic location, which allows the use of detailed biostratigraphic and sedimentary information in the study of the major tectonic, climatic and eustatic events affecting the North Atlantic margin.
Since the late Cretaceous, Iberia evolved located between the Eurasian and African plates while it was displaced towards east by the opening of the North Atlantic Ocean. Mesozoic extension was replaced by a compressive regime, leading to the development of NE-SW trending basins in western Iberia. In the Cenozoic basins of western Iberia, erosional unconformities can be related to major tectonic events generated within distinct phases of compression between Iberia, Europe and Africa. The main Portuguese Cenozoic basins (Mondego and Lower Tejo) are located in the western border of the Iberian Massif and separated by the Central System, Estremenho Massif and Arrabida chain, southwest-trending relieves uplifted since the Miocene. Their depositional records include onshore continental units grading westwards into offshore marine sediments that document palaeogeographic, tectonic, climatic and eustatic events. Allostratigraphic units were established in the western Iberian margin and high-resolution sequence stratigraphy is currently applied to the Neogene. The specific distribution of depositional facies - alluvial deposits interfingering with marginal marine siliciclastics - and the abundance of fossil macro- and microfaunas demonstrate the exceptional importance of the record. Good biostratigraphic correlations are based on foraminifera, ostracoda, dinoflagellates, pollen, spores and mammals, coupled with isotopic ages (K/Ar in glauconites; 87Sr/86Sr), providing a chronostratigraphic framework and the definition of 10 main depositional sequences, in part agreeing with the Haq’s 3d order eustatic cycles. Inland, continental successions clearly reflect the influence of tectonics and climate on the evolution of the basin.
The LTB was endorreic during the Paleogene, with alluvial fan sedimentation. The first marine connection, from the South, occurred at the early Aquitanian, originating a gulf with a N-S coral reef that attained the Lisboa area; a West link to the sea was established in the early Burdigalian. Untill the middle Tortonian, a sandy braided depositional system drained NE-SW, to the Atlantic. During the late Tortonian to Zanclean, endorheic alluvial fan sedimentation was present along tectonic scarps. An exorheic drainage system developed at the transition to the more humid conditions of the Piacenzian, when the ancestral Tejo river became a gravelly braided river with abundant bed load after the capture of the drainage network of the Madrid Cenozoic Basin. During the following incision stage, deep valleys were excavated, and in tectonic depressions terrace staircases have developed.

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Gastropoda y Bivalvia característicos del Pleistoceno de Bahía San Blas (provincia de Buenos Aires, Argentina): estudios preliminares
Pisano M. F.1, Aguirre M. L. 2, 3, Farinati E. 4 & Fucks E. 3,5
1. Becaria Doctoral ANPCYT, Facultad de Ciencias Naturales y Museo, UNLP [email protected]; 2, CONICET [email protected]; 3, Facultad de Ciencias Naturales y Museo, INGEA, UNLP; 4. Departamento de Geología, Universidad Nacional del Sur. San Juan 670, 8000 Bahía Blanca [email protected] 5. LATYR, FCNYM, UNLP [email protected]
El Cuaternario marino del sector costero en los alrededores de Bahía San Blas (SE de la provincia de Buenos Aires, Argentina) está representado por geoformas litorales pleistocenas y holocenas con alto contenido malacológico (30 gastrópodos y 30 bivalvos) y microfauna asociada (foraminíferos y ostrácodos). En el área se reconocen tres niveles de terrazas marinas: I y II del Pleistoceno tardío (edades mínimas de 28.700 + 1400 a 43.000 14C A.P.; Último Interglacial ?, MIS5 ?) y el nivel III del Holoceno (2.170 + 110, 4.100, 5.370 + 110 años 14C A.P). Los moluscos pleistocenos exhiben una excelente preservación y poca pérdida de información postmortem durante su depositación final. Esta situación, muy poco común para el Pleistoceno marino bonaerense, caracterizado por los escasos registros ricos en malacofauna, brindan una excelente oportunidad para iniciar estudios complexivos tafonómicos, paleoecológicos, paleobiogeográficos y paleoambientales, con el fin de completar la caracterización malacológica en un área poco analizada del litoral bonaerense. Especialmente los ejemplares fósiles (15 gastrópodos y 13 bivalvos) provenientes de cordones conchiles pleistocenos cercanos al canal de Villalonga (Bahía Anegada) aportan información paleoecológica útil como complemento de importantes estudios geológicos y geocronológicos efectuados previamente en la región. En conjunto esta malacofauna es indicativa de ambientes litorales, principalmente del intermareal a infralitoral superior, de fondos mayormente blandos, y en su mayoría típicos de masas de agua oceánica superficial cálidas y templado-cálidas de la Provincia Malacológica Argentina (sur de Brasil Golfo San Matías). Todos los taxones cuentan con representantes vivientes en el Mar Argentino vecino, con excepción de Anachis avara (Say), actualmente desplazada al norte, hasta las costas de Uruguay y Brasil. Este desplazamiento, junto con el predominio de elementos de aguas cálidas o templado-cálidas, son indicativos de un episodio marino caracterizado por temperatura oceánica superficial ligeramente superior a la moderna en el área, en respuesta al aumento global de temperatura y del nivel del mar documentado para el Pleistoceno tardío. Sin embargo, el escaso control cronológico disponible no permite por el momento corroborar con precisión si estos niveles son correlacionables con el MI5e (máximo del Último Interglacial, ca. 125 ka A.P.). Nuevas dataciones por distintos métodos modernos, análisis isotópicos y un exhaustivo estudio de la malacofauna (en curso) permitirán arribar a conclusiones más precisas sobre los niveles pleistocenos y su comparación con la transgresión holocena y el litoral moderno.

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
La sección “Hacienda Clarevot” (Sevilla, SO España) y su registro paleontológico (Neógeno superior, Cuenca del Guadalquivir)
Rico-García, A.1,Cárdenas Carretero, J.2, Bajo, I. 3, Maestre, V.4 y J. Civis, J.1
1 Departamento de Geología, Facultad de Ciencias, Universidad de Salamanca, Pz. Merced s.n, 37008, Salamanca, España. [email protected]; [email protected] 2 Plaza España, 10 1ºD, 41700, Dos Hermanas (Sevilla). ([email protected]) 3 C/ Fuerteventura 2. 41700 Dos Hermanas, Sevilla. ([email protected]) 4 C/ Nuestra Sra. del Aguila 51. 41500 Alcalá de Guadaíra. Sevilla). ([email protected])
La sección Hacienda Clarevot, situada en el término municipal de Alcalá de Guadaíra
(Sevilla), consta de 34 metros y se divide en cuatro tramos, que de muro a techo son: U1) 18 metros de arcillas gris azuladas masivas, U2) 1 metro de arenas limo-arcillosas ocres-marronáceas con glauconita, U3) 11 metros de calcarenitas de grano medio-grueso de color amarillento cuyo techo está alterado y erosionado, y U4) 4 metros de arcillas verdosas y arenas blanquecinas.
El alto contenido en foraminíferos planctónicos de las unidades inferiores U1 y U2 ha permitido determinar su edad. La abundancia de individuos de Globorotalia miotumida y la presencia muy escasa de G. margaritae indica una edad Messiniense inferior, cercana a los 6,34 Ma y al Evento PF5. La ausencia de biomarcadores en U3 y U4 no hace posible su datación, aunque regionalmente por su posición estratigráfica permite asignar una edad Messiniense inferior-superior para U3 y Plioceno para U4.
Se han determinado en la sección un total de 147 especies: 21 de vertebrados (condríctios, osteíctios y cetáceos) y 126 de invertebrados (moluscos, corales, briozoos, anélidos, cirrípedos, equinodermos y crustáceos), de los que el grupo más importante con106 especies es el de los moluscos (5 escafópodos, 52 bivalvos y 49 gasterópodos). La U1 presenta la mayor diversidad y contenido en fósiles, con un 61% de las especies totales de la sección, destacando su gran riqueza en moluscos. El grado de conservación es excepcional, con conchas completas, articuladas, nulos efectos de desgaste y conservando trazas originales de color y estructuras ornamentales delicadas. En la U2 se han determinado 46 especies, destacando los dientes de condrictios y osteíctios y moldes de moluscos. La U3 presenta 36 especies con un mayor desgaste tafonómico que U2, destacando la abundancia de pectínidos y trazas fósiles de Gastrochaenolites en la base. Por último, la U4 presenta esterilidad en restos de macrofauna.
Este afloramiento supone el mejor registro del Messiniense de moluscos, por su diversidad y conservación, en la cuenca del Guadalquivir e incluso en la Península Ibérica. Este registro malacológico apoya la datación microfaunística de Messiniense, ya que ciertas especies sólo han sido citadas en ese rango temporal. Los cambios en las asociaciones de macrofauna y su impronta tafonómica, reflejan un importante y progresivo descenso del nivel del mar durante el Messiniense, pasando de un ambiente batial-circalitoral inferior hasta el infralitoral.

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Tafonomía comparativa de bivalvos dominantes en el Cuaternario marino de Argentina
Richiano, S.M. 1,2, Aguirre, M.L. 1,3, Farinati, E.A. 4, Fucks, E. 3 y Pisano, F. 5
1. CONICET 2. CIG, Centro de investigaciones Geológicas, calle 1 No 644, 1900 La Plata 3. Facultad de Ciencias Naturales y Museo, INGEA, calle 64 No 3, 1900 La Plata 4. Departamento de Geología, Universidad Nacional del Sur. San Juan 670, 8000 Bahía Blanca 5. Becaria Agencia Nacional de Promoción Científica y Tecnológica La tafonomía comparativa en ambientes sedimentarios marinos permite analizar las preservaciones diferenciales que presentan los restos esqueléticos de diversos grupos de moluscos y otros organismos bentónicos litorales, al estar sujetos a procesos bioestratinómicos y diagenéticos tempranos. El análisis del estado de conservación de dos bivalvos dominantes en el Cuaternario marino de Argentina, Mactra isabelleana d´Orbigny y Brachidontes rodriguezi (d´Orbigny) procedentes de depósitos del Pleistoceno y Holoceno medio de distintos sectores del litoral atlántico bonaerense, brindan la oportunidad de establecer comparaciones semicuantitativas objetivas y evaluar evidencias paleoambientales. La elección de estas especies se debe a su abundancia (superior al 80 %). en las concentraciones muestreadas. Para el área de Bahía Samborombón (Noreste de la provincia de Buenos Aires) se recolectaron muestras de las Formaciones Pascua (Pleistoceno, ambiente de playa), Destacamento Río Salado (Holoceno, planicie de mareas) y Las Escobas (Holoceno; Mb Canal 18, laguna costera y Mb Cerro de la Gloria, barra litoral) en las cuales se analizó un total de 2200 ejemplares de Mactra isabelleana. Por otro lado, en el estuario de Bahía Blanca (Sur de la provincia de Buenos Aires) se estudió el evento transgresivo-regresivo holoceno (cordones litorales) y se obtuvieron 1800 ejemplares de Brachidontes rodriguezi. Primeramente se compararon ambos taxones desde el punto de vista morfológico (arquitectura de la conchilla) y paleoecológico (modo de vida y batimetría, entre otros). Luego se analizaron distintos parámetros tafonómicos como articulación, fragmentación, abrasión, color, bioerosión, incrustación y disolución clasificando a los mismos en tres grados de alteración (0, bueno; 1, regular; 2, pobre). Los datos fueron cuantificados y ploteados sobre tafogramas ternarios. Los resultados muestran que la articulación, fragmentación, bioerosión, incrustación y disolución, no señalan diferencias entre las diferentes facies sedimentarias, y en general presentan valores comparables. En cambio, abrasión y color, son los atributos que permiten hacer una distinción evidente entre las facies de menor y mayor energía. Estos resultados tienen el valor agregado de ser independientes de los tan diferentes hábitos y hábitat de los taxones. En ambas áreas analizadas la cuantificación obtenida es coherente con el análisis sedimentario de las unidades. Así, las facies de playa y cordón habrían sido originadas por eventos multiepisódicos de tormenta que transportaron y acumularon los bioclastos desde su zona de origen hasta el área de depositación. Mientras que en la planicie costera y lagoon predominaría la baja energía con escasa abrasión y conservación del color original.

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Las esponjas perforantes: una clave para conocer aspectos de la vida de una
ostra miocena del Atlántico Sur
Romero, M. V.1, Rodríguez2, V., Casadío, S.1,2, Martín, M. C. 1 y Pascual, M.3 1Facultad de Ciencias Exactas y Naturales, Universidad Nacional de La Pampa, Uruguay 151, (6300) Santa Rosa, La Pampa. [email protected] 2CONICET. 3Instituto de Biología Marina y Pesquera “Alte. Storni”, 8520 San Antonio Oeste, Río Negro, Argentina.
Una de las ostras más frecuentes del Mioceno tardío de la Patagonia es Ostrea alvarezii d’Orbigny, 1842 que muestra afinidades importantes con la especie actual O. puelchana d’Orbigny, 1842. Los estudios preliminares señalan que ambas especies compartirían características morfológicas, estrategias reproductivas y requerimientos ambientales similares. Tanto O. alvarezii como O. puelchana registran en sus valvas una comunidad de epibiontes muy diversa, entre los que se destacan las esponjas perforantes. En las ostras actuales, la posición que adoptan los ejemplares con respecto al substrato es uno de los factores que controla la distribución y abundancia de las esponjas, por lo que esta información es de utilidad para inferir la posición de vida en las ostras fósiles. En este trabajo se compara la distribución de las perforaciones de esponjas en valvas de O. alvarezii (n=60) recolectadas en facies de canales de marea de la Formación Puerto Madryn, expuestas en los acantilados de Puerto Pirámide (provincia de Chubut) con la observada en ejemplares de O. puelchana (n=52) coleccionados en el banco de ostras “Las Grutas”, Golfo San Matías. La presencia de las perforaciones se registró según un esquema que divide a las valvas de las dos especies de ostras en distintas regiones. Estas divisiones reflejan diferencias en la morfología de la conchilla que podrían influir en el asentamiento y/o supervivencia de las esponjas. La valva izquierda se dividió en 6 áreas y la derecha en 5. En la especie fósil el 76,67% de las valvas izquierdas y el 30 % de las derechas presentaron perforaciones, mientras que en la especie actual el 92 % de las izquierdas y el 44 % de las derechas registraron esponjas. En las valvas derechas de ambas especies se observó que, preferentemente, las esponjas se encuentran en el ápice y en menor medida en el margen ventral (p<0,03 para O. puelchana y p<0,0001 para O. alvarezii). Mientras que en las valvas izquierdas de O. puelchana se observó una ubicación preferencial de las perforaciones en el ápice y en el zócalo (p<0,01) y, en las de O. alvarezii en el margen ventral y el zócalo (p<0,0000004). Esta diferencia en la valva izquierda se debería a que en la ostra fósil el área de fijación es, con frecuencia, más grande y abarca el ápice. Los resultados sugieren que la posición de O. alvarezii con respecto al substrato habría sido similar a la observada para O. puelchana (i. e. con la valva derecha hacia abajo).

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Análisis microfaunístico del nivel condensado Tortoniense en el margen pasivo de la Depresión del Guadalquivir (SO de España)
Ruiz, F.1, Abad, M.1, Pendón, J.G.2, González-Regalado, M.L.1, Tosquella, J.1, Prudencio, M.I.3 y Dias, M.I.3
1Departamento de Geodinámica y Paleontología, Universidad de Huelva, 21071-Huelva, España. 2Departamento de Geología, Universidad de Huelva, 21071-Huelva, España. 3Instituto Tecnologico e Nuclear, EN 10, 2686-953 Sacavem, Portugal.
En este trabajo, se analiza el contenido microfaunístico de un nivel condensado de limos glauconíticos presente en tres secciones situadas en los sectores central y occidental de la Depresión del Guadalquivir (SO España), de edad Tortoniense Superior. Pueden definirse dos tramos:
a) Tramo basal, con abundantes foraminíferos bentónicos (Cibicides floridanus, Cibicidoides pseudoungerianus, Heterolepa bellincionii, Marginulina costata), mezcla de ostrácodos típicos de medios infralitorales y circalitorales y un bajo índice de icnofábrica. Esta coexistencia podría ser indicativa de bajas tasas de sedimentación en un intervalo temporal donde hay una transición desde condiciones someras a medios oceánicos más profundos;
b) Tramo superior, con mayores porcentajes de foraminíferos bentónicos típicos de medios
de plataforma media-externa, raros ostrácodos (Costa tricostata, Henryhowella sarsi profunda) y un índice de icnofábrica superior.
La razón P/B oscila entre las diversas secciones, con máximos del 47 % en el tramo
superior de las secciones más occidentales. Entre los foraminíferos planctónicos, se observa la coexistencia de taxones típicos de aguas frías y cálidas, lo que podría deberse a la entrada episódica de aguas atlánticas frías o a oscilaciones climáticas de alta frecuencia.

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Ostrácodos Neógenos marinos del Suroeste de Europa
F. Ruiz1, M. L. González-Regalado1, M. Abad1 y J. Civis2
1Departamento de Geodinámica y Paleontología, Universidad de Huelva, 21071-Huelva, España. 2Departamento de Geología, Universidad de Salamanca, 37008-Salamanca, España
En este trabajo, se estudian los ostrácodos presentes en 17 secciones del suroeste de la Depresión del Guadalquivir (Sur de España), con una edad comprendida entre el Tortoniense Superior al Plioceno Inferior. Su comparación con los resultados obtenidos en el Neógeno Superior de Portugal y Francia ha permitido diferenciar las siguientes asociaciones: Tortoniense Superior. En la zona infralitoral, abundaban Cytheretta simplex, Cytheretta orthezensis, Mutilus labiatus y Urocythereis favosa. Los medios circalitorales se reconocen por la presencia conjunta de Bosquetina carinella, Costa batei, Echinocythereis scabra y Paracypris polita. Las especies más representativas del talud superior serían Henryhowella partenopaea, Krithe soustaensis y Costa tricostata. Messiniense. Los medios litorales más someros estaban poblados básicamente por diversas especies de los géneros Urocythereis, Cistacythereis y Carinocythereis. En la plataforma media, eran frecuentes los especimenes de Acanthocythereis hystrix, Ruggieria tetraptera, Xestoleberis prognata y Pterigocythereis spp.. En las zonas epibatiales, la diversidad era baja, con una frecuente presencia de Buntonia multicostata y diversas especies de Krithe, Parakrithe y Cytherella. Plioceno Inferior. Pueden diferenciarse 5 asociaciones: a) Infralitoral, con Pontocythere elongata, Flexus triebeli, Quadracythere prava y Leptocythere faveolata; b) Circalitoral somera, representada por Costa edwardsii, Buntonia sublatissima y Celtia quadridentata; c) Circalitoral profunda, con abundantes ejemplares de Ruggieria tetraptera, Henryhowella asperrima y Pterigocythereis jonesii; d) Batial superior, con escasos individuos de Henryhowella asperrima, Monoceratina mediterranea, Krithe spp. y Parakrithe spp.; y e) Batial inferior, con raros ejemplares de Krithe, Agrenocythere y Abyssocythere sulcatoperforata.

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Efecto de las tormentas en las asociaciones de Foraminíferos bentónicos
del Plioceno de Huelva (Suroeste de España)
Ruiz, F.1, González-Regalado, M.L.1, Abad, M.1, Civis, J.2, González-Delgado, J.A.2, Muñoz, J.M.3, y Pendón, J.G.4
1Departamento de Geodinámica y Paleontología, Universidad de Huelva, 21071-Huelva, España. 2Departamento de Geología, Universidad de Salamanca, 37008-Salamanca, España 3Departamento de Estadística e Investigación Operativa, Universidad de Sevilla, 41071-Sevilla, España. 4Departamento de Geología, Universidad de Huelva, 21071-Huelva, España.
El análisis litoestratigráfico y faunístico (foraminíferos bentónicos, moluscos) de
cinco secciones pliocénicas localizadas en el suroeste de la Depresión del Guadalquivir permite definir tres facies sedimentarias principales, que se habrían depositado en un medio marino somero bajo condiciones de calma energética (facies FA-1), debido a la acción de las tormentas (facies FA-2) o en areas fluvio-litorales (facies FA-3).
El estudio estadístico de las poblaciones de foraminíferos bentónicos revela la
presencia de 6 asociaciones, representativas de zonas circalitorales (asociaciones de Fursenkoina shreibersiana y Globobulimina auriculata), infralitorales (asociaciones de Ammonia inflata y Ammonia beccarii) o bien ubicuas de zonas de plataforma interna (asociaciones de Florilus boueanum y Reusella spinulosa). Las variaciones verticales y horizontales de estas asociaciones, los cambios en diversos indices faunísticos, así como la distribución de los moluscos, sugiere la presencia de un medio marino somero, con una zona litoral que recibía aportes de aguas dulces (Lucena, Bonares y Casa del Pino) y una profundidad creciente hacia el suroeste (Moguer y El Rompido).
Los efectos de las tormentas sobre las poblaciones de foraminíferos bentónicos se
limitaron a una ligera disminución del número de individuos y especies de foraminíferos bentónicos, una importante disminución en el número de foraminíferos planctónicos y muy leves diferencias en otros índices faunísticos. Estos datos y la presencia de poblaciones relativamente bien conservadas de moluscos serían indicativas de un transporte limitado desde las paleobiocenosis originales. Las condiciones de post-tormenta se caracterizarían por: a) un aumento en el número de individuos; b) un progresivo aumento en el cociente P/B; y c) un ligero incremento de diversos índices faunísticos.

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Presencia del Burdigaliense en las secuencias basales en la Perforación IGU Chuy Nº 364 del Uruguay datadas con 87Sr/86Sr en tecas de
foraminíferos Cibicides.
Sprechmann, P.1; Chemale, F. Jr.2 y Gaucher, C.1.
1Instituto de Ciencias Geológicas, Facultad de Ciencias, Iguá 4225, Montevideo 11400, Uruguay. 2Laboratório de Geologia Isotópica, IG-UFRGS, Cx. P. 15001, Porto Alegre – RS, Brasil. En el territorio continental del Uruguay se han descrito 4 depocentros con sedimentos de la Formación Camacho desarrollados durante la transgresión “Paranense” y/o “Entrerriense”, siendo la única unidad con asociaciones fosilíferas marinas del Neógeno. Existen desavenencias sobre su edad, postulándose una depositación en el Mioceno Medio o Mioceno Superior. Unos afirman que la depositación fue isócrona y otros diacrónica. Se aportan nuevos datos sobre secuencias del Mioceno halladas en la Perforación IGU N364 (33º42´S, 53º26´W) ubicada en el borde occidental de la Cuenca de Pelotas. Las muestras corresponden al intervalo de 133,00-113,00 m. Fueron obtenidas por percusión. Contienen, en lo esencial, asociaciones de foraminíferos bentónicos, moluscos, braquiópodos, ostrácodos, briozoos, otolitos y dientes de condrictios. Las asociaciones son normal-marinas: las razones isotópicas del Sr reflejan equilibrio con el agua de mar. Estas asociaciones han sido asignadas bioestratigráficamente a la Formación Camacho. Para dilucidar algunas de los incógnitas citadas se procedió, en el Laboratorio de Geologia Isotópica IG-UFRGS, Porto Alegre, al análisis de las razones 87Sr/86Sr en tecas del foraminíferos Cibicides aknerianus y Cibicides "pseudoungerianus”. Los resultados obtenidos son:
Muestra 87Sr/86Sr Error (%)
CHUYM40: 122.10-124.00 0,70819 0,0019
CHUYM41: 124.00-125.40 0,70856 0,0024
CHUYM42: 125.40-128.00 0,70874 0,0018 Los análisis de las muestras 42 y 41 (125,40-122.10 m) presentan resultados muy similares con razones isotópicas de 0.7087 y 0.7086, respectivamente. Según Mc Artur et al. (2001) comprenden a edades absolutas de 17 y 18 Ma, perteneciendo al Burdigaliense (Mioceno Inferior). La muestra 40 indica una relación 0.7082, es decir Oligoceno Superior. Su ubicación estratigráfica por encima de las anteriores, permite descartarla y suponer que se trata de alteración diagenética de los valores primarios. Las edades mencionadas indican que la

depositación de los foraminíferos se verificó entre los dos importantes hiatos hallados por Anjos-Zerfass et al. (2006) en el Mioceno Inferior y Mioceno Inferior-Medio de las Cuenca de Pelotas del offshore del sur del Brasil. Las secuencias datadas de la Perf. Chuy 364 corresponden a eventos acaecidos en la plataforma continental adyacente a dicha cuenca oceánica, y no pertenecen a la Formación Camacho. La presencia del Burdigaliense en la Perforación Chuy 364 se confirma por la presencia de los foraminíferos bentónicos Amphistegina lessonii, Cancris sagra y Poroeponides lateralis. Referencias Anjos-Zerfass, G.S., Carreño, A.L. y Coimbra, J.C. 2006. Neogene foraminifera from Pelotas Basin, southern Brazil: Biostratigraphy and paleoceanography. Anuário do Instituto de Geociencias – UFRJ 29: 319-320. McArthur, J.M., Howarth, R.J. y Bailey, T.R. 2001. Strontium Isotope Stratigraphy: LOWESS Version 3: Best Fit to the Marine Sr-Isotope Curve for 0-509 Ma and Accompanying Look-up Table for Deriving Numerical Age. The Journal of Geology 109: 155-170.

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Evidencias de depredación en restos de vertebrados asignados a la edad Montehermosense en Farola Monte Hermoso, Buenos Aires, Argentina
Tomassini, R.L.1,2 y Montalvo, C. I.2 1. CONICET; 2. Facultad de Ciencias Exactas y Naturales, Universidad Nacional de La Pampa, Argentina La Formación Monte Hermoso en su localidad tipo, Farola Monte Hermoso (Buenos Aires), ha aportado mamíferos y otros vertebrados asignados a las edades Montehermosense y Chapadmalalense temprano. Se trata de un sistema fluvial meandriforme en el que se describieron varias asociaciones de facies. Los materiales que se analizan en esta oportunidad provienen de limos arcillosos correspondientes a la facies Fl, interpretada como un ambiente de llanura de inundación. Se trata de dos pequeñas concentraciones constituidas por material fósil muy fragmentario de mamíferos y peces. Si bien no se encontraron próximas entre sí, por sus características y las similitudes que presentan se las estudió juntas. La primera, de 7,7 y 4,5 cm de diámetros máximo y mínimo respectivamente, y la otra dividida en cuatro sectores de acumulaciones, con diámetros máximos de entre 1 y 5 cm. En los dos casos el espesor de las acumulaciones no superaba los 50 mm. En ambos, el sedimento aglutinante se diferenciaba del que las contenía por su color, más amarillento, y su alto grado de cementación. Se analizaron tafonómicamente todos los elementos esqueléticos y fragmentos incluidos en estas concentraciones. El 87,55% de los restos correspondían a astillas indeterminables anatómicamente cuyo tamaño no superó los 90 mm de largo y 40 mm de ancho. Entre los mamíferos fueron identificados Actenomys priscus y Paedotherium sp.; estos taxa avalarían la coetaneidad de estas asociaciones con la fauna del nivel portador, que clásicamente se ha asignado al Montehermosense. En cuanto a la representación anatómica los elementos esqueléticos más abundantes fueron las vértebras, molares y metápodos. Todos los materiales mostraron signos de redondeamiento, en algunos casos había pérdida de material óseo compacto y en aquellos en los que fue posible evaluar grados de digestión (fémur proximal, molares e incisivos) ésta era de leve a fuerte. Por el grado de fragmentación y las evidencias de digestión estas asociaciones se interpretan como acumulaciones de restos digeridos por un depredador. La sumatoria de los atributos tafonómicos analizados señalan que se trataría de un depredador que produce modificaciones de moderadas a fuertes en los huesos de sus presas. En este contexto las asociaciones se podrían interpretar como egagrópilas fósiles, producidas por un ave diurna o coprolitos colapsados, producidos por un mamífero carnívoro. Ya sea que se trate de un ave o de un mamífero, este registro constituye la primera evidencia de depredación en la Formación Monte Hermoso.

ABSTRACTS 4th CONGRESS, Buenos Aires (Argentina), December 2008
Niveles de concentración de Heterostegina (Nummulitidae, Foraminifera) en los sedimentos del Tortoniense (Mioceno Superior) en el margen pasivo
del sector Occidental de la Cuenca del Guadalquivir (SW de España
Tosquella, J., Abad, M., Pendón, J.G., Ruiz, F. y González-Regalado, M.L.
Departamento de Geodinámica y Paleontología, Facultad de Ciencias Experimentales, Universidad de Huelva Avda. 3 de Marzo s/n, Campus Universitario de “El Carmen”, HUELVA-21071. E-mail: [email protected] La base de la sedimentación neógena en el margen pasivo septentrional del sector occidental de la Cuenca del Guadalquivir (Fm. Niebla o Complejo Basal Transgresivo) se caracteriza por una alternancia de eventos transgresivos y regresivos bien reflejados por las variaciones en el modelo de procesos sedimentarios, de arquitectura deposicional, y en especial por el contenido fósil. De este modo, los eventos transgresivos se hallan desarrollados esencialmente en un medio de plataforma marina mixta en contexto de rampa, con una sedimentación principalmente calcarenítica caracterizada por la presencia de algas rodofíceas, briozoos, equinodermos y diversos tipos de moluscos. En numerosas ocasiones presentan asimismo niveles decimétricos a métricos de acumulación de macroforaminíferos del género Heterostegina, cuyas características de depósito son motivo de controversia. Heterostegina es un nummulítido de pared hialina que contiene algas unicelulares simbiontes en su citoplasma que contribuyen a su nutrición, lo que le hace ocupar habitualmente sectores de la plataforma marina con presencia de luz, en condiciones oligo- a mesotróficas. El carácter prácticamente monoespecífico de estos niveles (Heterostegina gomez-angulensis) y las observciones realizadas a través del análisis tafonómico de sus esqueletos permiten inferir numerosos datos sobre el contexto bio-deposicional de su formación. La morfología plana y ornamentación poco desarrollada de las conchas, la no evidencia de abrasión en las mismas, con fracturaciones angulosas por compactación diagenética del sedimento y por meteorización, así como el análisis de las proporciones numéricas entre formas microsféricas (con reproducción asexual) y megalosféricas (con reproducción sexual), de alrededor del 5%, las habituales en poblaciones actuales de este género, parece indicar que se trata de concentraciones autóctonas y démicas en medios infralitorales de aguas templadas a cálidas, con paleobatimetrías no superiores a los 30 m. Estos datos se hallan apoyados por el análisis del contenido fosilífero de microforaminíferos bentónicos y ostrácodos. Por otra parte, en este trabajo se pretende discutir el significado biológico de tal tipo de concentraciones en determinados momentos de la historia geológica y su relación con el contexto sedimentario y climático del sector occidental de la Cuenca del Guadalquivir durante el Tortoniense medio-superior, edad en la que se encuentran la mayor parte de estas concentraciones.