Embed Size (px)
Citation preview

Anno Accademico 2013/2014
Sintesi di sequenze peptidiche di enterovirus Coxsackie e hGAD per lo studio di una possibile correlazione tra agenti infettivi e diabete di tipo 1 Synthesis of peptide sequences of enterovirus Coxsackie and hGAD for the study of a possible correlation between infectious agents and type 1 diabetes
Relatore Prof. Anna Maria Papini Correlatore Prof. Chiara Azzari Candidato Lorenzo Altamore
Scuola di Scienze Matematiche
Fisiche e Naturali
Corso di Laurea in Scienze Chimiche

2
Abstract
Candidato: Lorenzo Altamore ([email protected])
Relatore: Anna Maria Papini ([email protected])
Correlatore: Chiara Azzari ([email protected])
Controrelatore: Antonella Capperucci ([email protected])
Il diabete mellito di tipo 1, sia nella sua forma giovanile (Insulin-Dependent
Diabetes Mellitus, IDDM) che negli adulti (Latent Autoimmune Diabetes in Adults,
LADA) è una patologia sulla quale sono stati effettuati, negli ultimi decenni
numerosi studi, al fine di comprenderne i meccanismi molecolari e soprattutto le
cause scatenanti, che accettate essere di natura genetica e/o ambientale. Fra gli
agenti ambientali più probabili, legati allo scatenamento e allo sviluppo di questa
patologia è sempre più accettato il ruolo di un’infezione virale precoce causata
dall’enterovirus Coxsackie B4. Il meccanismo che lega i due eventi rimane però
ancora ignoto. Una delle ipotesi più probabili al riguardo suggerisce un meccanismo
di mimicry verso una porzione, denominata P2C, di una proteina genomica del virus
rispetto a frammenti delle due isoforme della Glutamic Acid Decarboxylase (GAD65
e GAD67), un enzima coinvolto nella regolazione del glucagone. La P2C condivide
infatti con GAD65 e GAD67 un possibile epitopo, il frammento peptidico PEVKEK (J.-
C. Tong et al. Ann.N.Y.Acad.Sci. 2002). Questo mimetismo potrebbe generare una
cross-reattività degli anticorpi anti-Coxsackie verso le due isoforme della GAD e
portare al riconoscimento di neo-epitopi nella GAD stessa. Ne conseguirebbe un
danno all’enzima da parte del sistema immunitario e una conseguente
disregolazione dei livelli di glucagone, con possibile scatenamento della patologia.
La GAD è infatti riconosciuta ad oggi come uno dei più significativi autoantigeni
legati al diabete di tipo I e gli anticorpi anti-GAD sono accettati a livello clinico
come uno dei biomarker più caratteristici di questa patologia non solo a livello
diagnostico, ma anche predittivo.
Lo scopo di questo lavoro di tesi è stato quello di porre le basi per un
approfondimento mirato ad avvalorare questa ipotesi. Sono stati perciò selezionati
e sintetizzati tramite strategia di sintesi di peptidi su fase solida Fmoc/tBu assistita
da microonde, i tre frammenti peptidici:

3
hGAD65: 250AMMIARFKMFPEVKEKGMAALPRL273, hGAD67:
258SIMAARYKYFPEVKTKGMAAVPKL281 e CVB4 P2C: 28FIEWLKVKILPEVKEKHEFLSRL50.
I peptidi sono stati sintetizzati sia in forma non acetilata all’estremità N-terminale
(I-III), che acetilata (IV-VI). Il motivo di tale scelta è legato ad introdurre un
ulteriore legame ammidico in posizione N-terminale, mimando un frammento
interno alla putativa proteina antigenica e favorire l’interazione con anticorpi
specifici eventualmente presenti nel siero dei pazienti. I peptidi I, II e III sono stati
utilizzati per studi di immunoaffinità mediante Risonanza Plasmonica di Superficie
(BIACORE), mentre i peptidi IV, V e VI sono stati utilizzati in test immunoenzimatici
su fase solida SP-ELISA. In entrambi i casi sono stati utilizzati sieri di pazienti
diabetici e controlli provenienti sia dall’Azienda Ospedaliera Universitaria Meyer di
Firenze (sieri IDDM) che dall’Università di Padova (sieri LADA).
Nei sieri si pazienti IDDM è stato possibile rivelare, tramite SP-ELISA, anticorpi solo
di classe IgM. In particolare nel caso dei sieri 15 e 43 è stata osservata una netta
positività al test contro tutti e tre i peptidi IV, V e VI, mentre i sieri 1 e 37A hanno
dato risultati positivi nei confronti dei peptidi hGAD67 (258-281) e CVB4 P2C (28-
50) e dei peptidi hGAD65 (250-273) e CVB4 P2C (28-50) rispettivamente.
Inoltre sono stati ricercati gli stessi anticorpi anche in una coorte ridotta di sieri
appartenenti a pazienti LADA, al fine di verificare se questi fossero presenti anche
in soggetti adulti. Anche in questo caso non sono stati ritrovati anticorpi IgG, ma
solo IgM con valori di assorbanza elevati verso le tre sonde peptidiche IV, V e VI.
Essendo le IgM la prima classe di anticorpi prodotta in seguito a infezione, la loro
presenza può essere un indice di reminiscenza dell’infezione stessa. Possiamo
perciò affermare che anche se in un numero esiguo di pazienti, abbiamo una prima
dimostrazione dell’effetto di mimicry ipotizzato tra hGAD 65, hGAD 67 e la porzione
P2C della sequenza della proteina genomica virale del Coxsackievirus B4..
Gli studi di binding effettuati con i peptidi I, II e III mediante BIACORE T100,
ottimizzati ad oggi per il riconoscimento di anticorpi IgG, hanno dato come atteso
risultati negativi sia nei sieri dei pazienti diabetici che nei controlli. Misure
d’interazione tra le IgM identificate per la prima volta in questo lavoro di tesi e le
nuove sonde peptidiche I, II e III sono attualmente in corso.

4
INDICE
1. INTRODUZIONE 7
1.1. LA RISPOSTA IMMUNITARIA 7
1.1.1. Regio
1.1.2. ni variabili: caratteristiche strutturali e
legame con l’antigene 11
1.1.3. Regioni costanti: caratteristiche strutturali e
funzioni effettrici 12
1.1.4. Legame anticorpo-antigene 16
1.2. PATOLOGIE AUTOIMMUNI 17
1.2.1. Diabete mellito di tipo 1 (T1D) 18
1.2.1.1. Ruolo metabolico della GAD 20
1.2.1.2. Coxsackie Virus 23
2. PRESUPPOSTI DELLA RICERCA 25
2.1. CORRELAZIONE FRA DIABETE MELLITO DI TIPO
1 ED INFEZIONE DA COXSACKIE VIRUS:
MIMETISMO 25
2.1.1. Danneggiamento diretto e distruzione delle
β-cellule delle isole pancreatiche 25
2.1.2. Reazione autoimmune: danneggiamento del
procedimento chimico metabolico di
regolazione del glucosio nel sangue 26
2.1.2.1. Anticorpi anti-GAD 31
3. SVILUPPO DELLA RICERCA 32

5
3.1. SPPS 33
3.1.1. Teoria e applicazioni delle microonde in SPPS 37
3.1.2. Microonde in sintesi peptidica 40
3.1.2.1. Reazioni collaterali 41
3.1.2.2. Liberty BlueTM CEM 44
3.2. SINTESI DEI PEPTIDI hGAD65 (250-273) (I),
hGAD67 (258-281) (II) e CVB4 P2C (28-50) (III) 46
3.3. SINTESI DEI PEPTIDI hGAD65 Ac-(250-273) (I),
hGAD67 Ac-(258-281) (II) e
CVB4 P2C Ac-(28-50) (III) 46
3.3.1. Reazione di Acetilazione 47
3.4. PURIFICAZIONE E CARATTERIZZAZIONE DEI
PEPTIDI 47
3.5. RILEVAZIONE E QUANTIFICAZIONE DI
ANTICORPI TRAMITE TECNICA BIACORE
E TECNICA IMMUNOENZIMATICA ELISA 49
3.5.1. Biacore 49
3.5.1.1. Risonanza Plasmonica di Superficie 52
3.5.1.2. La superficie del biosensore 54
3.5.1.3. Procedura di pH-scouting 56
3.5.1.4. Immobilizzazione del ligando sul chip 60
3.5.1.5. Risultati dei test al BIACORE 64
3.5.2. Tecnica immunoenzimatica ELISA 68
3.5.2.1. Risultati dei test SP – ELISA 71

6
4. CONCLUSIONI 77
5. PARTE SPERIMENTALE 80
5.1. SPPS – MATERIALI E METODI 80
5.1.1. Sintesi automatica di peptidi su fase solida 81
5.1.2. Cleavage dalla resina 82
5.1.3. Acetilazione dell’estremità N-terminale 83
5.1.4. Purificazione dei peptidi 84
5.1.4.1. Pre-purificazione 84
5.1.4.2. HPLC semi-praparativa 84
5.1.4.3. Caratterizzazione dei peptidi 85
5.2. TEST DI IMMUNOAFFINITA’ BIACORE –
MATERIALI E METODI 86
5.2.1. Immobilizzazione dei peptidi sul chip 86
5.2.2. Studi di binding 87
5.3. TEST IMMUNOENZIMATICI IN FASE
SOLIDA (SP-ELISA) – MATERIALI E METODI 88
5.3.1. Test SP-ELISA – Fasi sperimentali 89
6. BIBLIOGRAFIA 91
7. RINGRAZIAMENTI 94

7
1 INTRODUZIONE
1.1 LA RISPOSTA IMMUNITARIA
Si definisce antigene (Ag) una qualunque sostanza in grado di essere riconosciuta
dal sistema immunitario, avente la caratteristica di combinarsi con i prodotti di una
reazione immunitaria (anticorpi o cellule T). Possono essere suddivisi in due
macrocategorie: “self”, generalmente tollerati dal sistema immunitario e spesso di
natura endogena, e “non-self”, in genere identificati come invasori e attaccati dal
sistema immunitario (spesso di natura esogena). Spesso si fa riferimento,
erroneamente, agli antigeni come “generatori di anticorpi”[1].
Si definisce invece immunogeno una sostanza che provoca in un organismo
suscettibile una risposta immunitaria (e quindi la produzione di molecole di
anticorpi specifici). Un immunogeno è un tipo specifico di antigene. Gli immunogeni
possono essere composti semplici (monomerici) o complessi (polimerici).
Generalmente però sono caratterizzati da un peso molecolare relativamente alto e
sono di natura proteica o polisaccaridica.
Molecole di piccole dimensioni non sono in grado, di per sé, di attivare i linfociti B,
ossia non funzionano da immunogeni. Per generare anticorpi diretti verso queste
piccole molecole è necessario coniugarle a macromolecola prima di procedere
all’immunizzazione: in questo caso la molecola piccola viene definita aptene,
mentre la macromolecola è chiamata carrier. Il complesso aptene-carrier,
diversamente dall’aptene da solo, è in grado di comportarsi da immunogeno.
Riassumendo, si definisce quindi immunogenicità la capacità di una sostanza di
indurre in un organismo una risposta umorale e/o cellulo-mediata di tipo immune;
l'antigenicità è invece la capacità di combinarsi specificamente con i prodotti finali
della risposta immunitaria (cioè gli anticorpi secreti e/o i recettori di superficie
presenti sulle cellule T). Anche se tutte le molecole che hanno proprietà
immunogene hanno anche proprietà antigeniche, non è vero il contrario[2].
Un anticorpo (o Ab, anche conosciuto come immunoglobulina) è invece un
substrato di natura proteica con una peculiare struttura quaternaria a forma di “Y”.
Gli anticorpi sono presenti nel sangue e in tutti gli altri fluidi degli organismi

8
vertebrati. Sono prodotti dal sistema immunitario in seguito al contatto con un
determinato antigene e la loro funzione è quella di identificarlo e neutralizzarlo[3].
Gli anticorpi hanno la capacità di legarsi in maniera specifica agli antigeni
(microorganismi infettivi come batteri, tossine o qualunque macromolecola
estranea che provochi la formazione di anticorpi). Negli organismi a sangue caldo
vengono prodotti dai linfociti B, trasformati per adempiere a questo compito, in
seguito a stimoli specifici, in plasmacellule. Le immunoglobuline, insieme ai
recettori dei linfociti T, TCR (T Cell Receptors), e alle molecole del complesso
maggiore di istocompatibilità, MHC (Major Histocompatibility Complex), sono le
uniche molecole capaci di legare l'antigene. La porzione strutturale dell’antigene
che viene riconosciuta e legata dall’anticorpo viene definita epitopo o
determinante (Fig. 1).
Figura 1 – Interazione Antigene-Anticorpo
Le macromolecole solitamente contengono molteplici epitopi, alcuni dei quali
possono essere ripetuti: ognuno di essi può ovviamente essere legato da un
anticorpo. Quando in uno stesso antigene sono presenti epitopi multipli identici si
parla di polivalenza o multivalenza. La maggior parte delle proteine globulari non
Epitopo

9
contengono epitopi identici ripetuti, e non sono quindi polivalenti, a meno che la
proteina non sia in forma aggregata.
Esistono tre diverse tipologie di epitopi:
• Epitopi lineari: sono definiti così quegli epitopi formati da una semplice
sequenza di residui amminoacidici. Di solito, il sito di legame per una
molecola anticorpale può alloggiare un determinante antigenico composto
da circa 6 amminoacidi. Gli epitopi lineari possono essere accessibili agli
anticorpi se sono esposti sulla superficie esterna dell’antigene, o in una
regione conformazionale estesa nella proteina nativa ripiegata; più spesso
invece gli epitopi lineari sono inaccessibili nella conformazione nativa della
proteina, e compaiono solo quando essa viene denaturata.
• Epitopi conformazionali: sono costituiti da amminoacidi non in sequenza e
si creano per avvicinamento spaziale, dovuto al ripiegamento della proteina,
di regioni distanti tra loro.
• Epitopi neo-antigenici: vengono generati in seguito a modifiche post-
traduzionali aberranti, quali fosforilazione o proteolisi, che, alterandone la
struttura covalente, possono generare nuovi epitopi.
In Figura 2 sono raffigurati i diversi tipi di epitopi.
Figura 2 – Tipologie di determinanti o epitopi antigenici

10
Tutte le interazioni di legame, generalmente non covalenti quali i legami a idrogeno,
le interazioni di Van der Waals ecc., avvengono quindi fra i siti di legame specifici
presenti sull’anticorpo/recettore e questa porzione del peptide antigenico
(epitopo).
La restante parte della sequenza amminoacidica può quindi eventualmente giocare
un ruolo prettamente strutturale, ovvero permettere l’esistenza stessa dell’epitopo
(nel caso di epitopi conformazionali) o coadiuvare l’avvicinamento dell’antigene
all’anticorpo.
L’approccio sintetico generalmente utilizzato per lo studio delle interazioni
antigene-anticorpo prevede quindi la sintesi non dell’intera proteina antigenica, ma
esclusivamente della sequenza che teoricamente rappresenta l’epitopo.
Come evidenziato in Figura 1, tutte le molecole anticorpali posseggono identiche
caratteristiche strutturali (regione costante), ma mostrano una variabilità
notevolissima nelle regioni che legano l’antigene (regione variabile). Questa
variabilità spiega la capacità delle diverse molecole anticorpali di legarsi ad un
numero enorme di antigeni strutturalmente differenti; ogni specie anticorpale è
dotata nel sito di combinazione con l’antigene di una sequenza amminoacidica
assolutamente unica, diversa da quella di tutti gli altri tipi di anticorpo. Le
caratteristiche chimico-fisiche e le funzioni effettrici delle Ig sono legate alla
porzione anticorpale che non lega l’antigene, che mostra una variabilità
relativamente bassa tra i diversi anticorpi. Ogni anticorpo è costituito da un nucleo
strutturale simmetrico, composto da due catene leggere identiche tra loro (ciascuna
del peso di circa 24 KD) e due catene pesanti anch’esse identiche tra loro (ciascuna
del peso di circa 55 o 70 KD). Ognuna delle due catene leggere è legata attraverso
un legame covalente ad una delle due catene pesanti da un ponte disolfuro; le due
catene pesanti sono a loro volta legate da ponti disolfuro. Sia le catene leggere che
quelle pesanti contengono una serie di unità omologhe, della lunghezza di circa 110
amminoacidi ciascuna, che prendono il nome di dominio immunoglobulinico. Un
dominio Ig contiene due strati di foglietto planare, ciascuno dei quali composto da
3-5 “nastri” polipeptidici ad andamento antiparallelo (Fig. 3)

11
Figura 3 – Domini immunoglobulinici della catena leggera degli anticorpi
Sia le catene pesanti che quelle leggere sono composte da una regione variabile
(rispettivamente VH e VL) N-terminale e da una regione costante (rispettivamente CH
e CL) C-terminale. Le regioni V sono definite tali in quanto contengono regioni di
variabilità della sequenza amminoacidica che differenziano gli anticorpi prodotti da
un clone di linfociti B da quelli prodotti da un clone diverso.
Dal momento che il nucleo strutturale di ogni molecola di anticorpo è costituito da
due catene pesanti e due catene leggere, ogni molecola Ig avrà due siti di legame
per l’antigene. I domini della regione C sono spazialmente separati dal sito di
legame per l’antigene e non partecipano al riconoscimento di quest’ultimo; tali
domini possono tuttavia interagire con molecole e cellule effettrici del sistema
immunitario, mediando così gran parte degli effetti biologici svolti dagli anticorpi.
L’estremità C-terminale delle catene pesanti serve, inoltre, ad ancorare gli anticorpi
sulla membrana dei linfociti B, dove svolgono le funzioni di recettore antigenico.
D’altra parte la regione C delle catene leggere non svolge alcuna funzione effettrice,
né funge da ancora per la membrana cellulare.
1.1.1 Regioni variabili: caratteristiche strutturali e legame con
l’antigene
La maggior parte delle differenze strutturali tra le diverse molecole di anticorpo si
concentra in tre brevi tratti situati nella regione V delle catene leggere e delle
catene pesanti, denominati segmenti ipervariabili. Ciascuna di queste regioni

12
altamente diversificate ha una lunghezza di circa 10 amminoacidi; i tre segmenti
ipervariabili sono tenuti assieme da regioni dette “cornice”.
Dal momento che queste sequenze possiedono una conformazione complementare
alla struttura tridimensionale dell’antigene che ad esse si lega, le regioni
ipervariabili sono spesso denominate anche CDR (Complementary-Determining
Regions) (Fig. 4)
Figura 4 – Regioni ipervariabili delle molecole di immunoglobuline
Queste regioni sono chiamate rispettivamente CDR1, CDR2 e CDR3. Sia nella
regione VH che in quella VL, il segmento più variabile delle tre sequenze è CDR3.
L’analisi cristallografica degli anticorpi ha rivelato che le CDR formano delle anse
esposte sulla superficie degli anticorpi e quindi disponibili ad interagire con
l’antigene.
1.1.2 Regioni costanti: caratteristiche strutturali e funzioni
effettrici
Le molecole anticorpali possono essere divise in classi in base a differenze nella
struttura delle regioni C delle catene pesanti. Queste classi sono chiamate anche
isotipi, e sono denominate IgA, IgD, IgE, IgG ed IgM (Tab. 1).

13
Tabella 1 – Isotipi delle immunoglobuline
Ognuna di queste classi possiede una specifica sottoclasse di catene pesanti, che
sono denominate rispettivamente α, δ, ε, γ, µ; sono invece noti due diversi tipi di
catena corta, denominati k e l.
Le IgG, o gammaglobuline, rappresentano la frazione più abbondante (70-75%)
delle immunoglobuline presenti nel sangue, svolgono un ruolo importante nella
difesa dalle infezioni: si legano ai macrofagi e ai leucociti permettendo loro di
individuare efficacemente il bersaglio da fagocitare. Le IgG sono anche capaci di
scatenare una serie di reazioni biochimiche, detta cascata del complemento, che si
conclude con l’eliminazione del microorganismo. Inoltre, le IgG sono l’unica classe
anticorpale efficace contro le tossine batteriche. Sono gli anticorpi maggiormente
impiegati durante la risposta immunitaria secondaria, cioè sono prodotte
tardivamente e in maniera massiva dai linfociti B differenziatisi in plasmacellule. In
questo esse si contrappongono alle IgM, che sono prodotte invece nelle fasi più
precoci dell'infiammazione, ma che hanno un'efficacia decisamente minore.
Le gammaglobuline hanno un'importantissima funzione nel proteggere il neonato
durante i primi mesi di vita, quando ancora non è in grado di produrre anticorpi da
solo. Le IgG sono infatti in grado di passare la barriera placentare, immettendosi nel
sangue del feto: questi possiede dunque lo stesso repertorio anticorpale della
madre per circa sei mesi, finché gli anticorpi ormai vecchi vengono degradati, ed è
quindi in grado di difendersi nei confronti di tutti quei patogeni con cui la madre è

14
venuta a contatto (anche solo per vaccinazione). Dopo la nascita, la madre continua
comunque a passare immunoglobuline al neonato mediante l'allattamento: il latte
materno contiene infatti grandi quantità di IgG ed IgA, che passano nell'apparato
digerente del neonato. Mentre le IgA si fermano nel canale digerente, le IgG
vengono assorbite dall'epitelio intestinale del lattante mediante uno specifico
recettore neonatale per le Fc, ed immesse nel torrente circolatorio.
Le IgM rappresentano circa il 10% delle immunoglobuline del sangue e sono
costituite dall’aggregazione di cinque, più raramente sei, monomeri di
immunoglobulina. La loro produzione è caratteristica delle risposte primarie agli
organismi infettivi. È sufficiente una sola molecola di IgM legata all’antigene per
attivare la cascata del complemento. L'emivita delle IgM nel siero è di 5-6 giorni,
circa tre volte inferiore a quella delle IgG.
Possono essere sintetizzate dai linfociti B in due forme: monomerica e pentamerica.
Nel primo caso si ritrovano (insieme alle IgD) sulla membrana del linfocita B
immaturo e la loro funzione principale è quella di captare gli antigeni circolanti per i
quali sono specifiche e di trasmettere all'interno del linfocita il segnale di
attivazione, che lo porterà a differenziarsi in una plasmacellula secernente
anticorpi. Le IgM vengono prodotte già in uno stadio molto precoce della
maturazione dei linfociti B: catene μ possono essere ritrovate già nel citoplasma dei
linfociti pre-B, dove vengono assemblate con una catena leggera sostitutiva e
permettono la formazione del recettore pre-B, utilissimo ai fini della selezione
positiva. Dopo l'attivazione del linfocita per il contatto con l'antigene ed il suo
differenziamento in cellula secernente anticorpi, le IgM subiscono un processo di
maturazione dell'affinità per l'antigene, per cui vengono prodotte IgM ad alta
affinità non più in forma di membrana ma in forma secretoria.
Nel secondo caso la principale funzione è l'opsonizzazione (potenziamento del
processo di fagocitosi dei batteri): essendo pentameriche, esse dispongono infatti
del quintuplo di siti di legame rispetto a tutte le altre immunoglobuline, e perciò
sono opsonine molto efficaci. Contrariamente alla maggior parte delle opsonine, la
funzione principale delle IgM non è però quella di favorire la fagocitosi della
particella opsonizzata da parte delle cellule immunitarie: pochissimi fagociti
possiedono, infatti, un recettore specifico per la porzione Fc delle IgM (Fig. 5).

15
Figura 5 – Le due porzioni derivanti dal clivaggio di un anticorpo
La loro funzione principale è invece quella di attivare il complemento: le IgM sono
infatti i migliori iniziatori della via classica dell'attivazione del complemento, meglio
ancora delle IgG.
Il sistema del complemento, insieme agli anticorpi, è un elemento essenziale del
sistema immunitario nei meccanismi di difesa umorale contro gli agenti infettivi.
Esso è costituito da una ventina di proteine circolanti e di membrana, capaci di
interagire reciprocamente e con le membrane cellulari. L'attivazione a cascata delle
sue proteine solubili, che convenzionalmente vengono chiamate componenti, è alla
base di attività biologiche varie come la lisi cellulare, batterica o virale. Queste si
introducono nelle membrane degli agenti patogeni provocando su di esse pori che
portano alla lisi. Durante l'attivazione del complemento si ha inoltre il reclutamento
di varie cellule immunocompetenti, quali le cellule fagocitarie (monociti, macrofagi,
polinucleati), i linfociti B e i linfociti T.
Le IgA sono gli anticorpi più rappresentati nelle secrezioni: saliva, latte, lacrime e
secrezioni respiratorie, digerenti e genitourinarie. Possono esistere sia in singola
unità (monomeri), sia aggregate in coppia (dimeri). Costituiscono un vero e proprio
sistema difensivo nelle mucose, rappresentando la prima barriera specifica che si
oppone alla penetrazione del materiale antigenico nell’organismo.
Le IgD rappresentano meno dell’1% delle immunoglobuline totali del plasma, ma
sono presenti in grande quantità sulla membrana di molti linfociti B circolanti. Esse
sembrerebbero svolgere un importante ruolo di recettori per l’antigene dei linfociti
B.
Frammenti “Fab” o “frammenti leganti l’antigene”
Frammento “Fc” o “frammento cristallizzabile”

16
Le IgE, pur trovandosi in piccolissime quantità nel siero plasmatico, sono presenti
sulla superficie dei basofili e dei mastociti di tutti gli individui, agendo come
recettori per gli antigeni. L’avvenuto legame stimola la degranulazione di queste
cellule che, liberando istamina e altri mediatori, provocano le manifestazioni
cliniche tipiche delle reazioni allergiche (asma, orticaria, febbre da fieno). Le IgE
svolgono inoltre un importante ruolo nella difesa dai parassiti, in particolar modo
dai vermi[4].
1.1.3 Legame antigene-anticorpo
Il legame tra l’antigene e l’anticorpo coinvolge diversi tipi di interazioni non
covalenti, tra cui forze elettrostatiche, ponti a idrogeno, interazioni di van der Waals
ed interazioni idrofobiche. La forza di legame tra un singolo sito combinatorio di un
anticorpo ed un epitopo antigenico, viene definita affinità dell’anticorpo. L’affinità è
solitamente espressa come costante di dissociazione (Kd), che definisce la
concentrazione di antigene necessaria per occupare i siti combinatori di metà delle
molecole di Ig presenti in una data soluzione di anticorpi. Una Kd bassa indica
un’affinità di interazione elevata, poiché significa che è necessaria una
concentrazione minore di antigeni per saturare la metà dei siti combinatori.
Per gli anticorpi rivolti verso antigeni d’interesse biologico, la Kd di solito oscilla tra
10-7 M e 10-11 M. Dal momento che la regione cerniera conferisce agli anticorpi una
certa flessibilità, una molecola di anticorpo può legarsi ad un antigene multivalente
con più di un sito combinatorio. Per le IgG e per le IgE questo legame può
coinvolgere al massimo due siti combinatori, situata su ognuno dei due Fab (cioè la
coppia di domini VL e VH, ognuno dei quali mantiene la capacità di legare
l’antigene); per le Ig pentameriche come le IgM, una singola molecola di anticorpo
può legare fino a 10 molecole di epitopi. Sebbene l’affinità di un sito combinatorio
sia la stessa nei confronti di ogni epitopo di un antigene polivalente, la forza di
legame dell’antigene all’anticorpo deve tener in conto il legame di tutti i siti
combinatori a tutti gli epitopi disponibili: questa forza complessiva di legame tra
antigene ed anticorpo è chiamata avidità, ed è ovviamente molto maggiore
dell’affinità di ogni singolo sito combinatorio. Il valore dell’avidità cresce in modo
quasi geometrico per ogni sito combinatorio occupato: pertanto, una molecola di

17
IgM a bassa affinità può legarsi in maniera molto forte ad un antigene polivalente,
poiché molte interazioni a bassa affinità possono produrre un’interazione
complessiva ad avidità elevata.
1.2 PATOLOGIE AUTOIMMUNI
Con il termine malattia autoimmune, in medicina, si indica l'alterazione del sistema
immunitario che dà origine a risposte immuni anomale o autoimmuni, cioè dirette
contro componenti dell'organismo umano in grado di determinare un'alterazione
funzionale o anatomica del distretto colpito[5].
L’autoimmunità consiste quindi in una risposta immunitaria contro antigeni self
(autologhi) e rappresenta un’importante causa di malattia.
È importante sottolineare che non sempre il verificarsi di una risposta
autoimmunitaria implica il successivo sviluppo di una patologia autoimmune. Sono
noti molti casi, legati soprattutto ad infezioni, in cui è presente una risposta
autoimmunitaria (transiente) che non sfocia in nessuna malattia autoimmune.
La malattia autoimmune è quindi una condizione patologica in cui è avvenuta una
rottura della tolleranza immunologica verso uno o più antigeni self. Il risultato della
scomparsa di questa tolleranza è la formazione di autoanticorpi e/o linfociti T
autoreattivi che determinano/inducono un danno tissutale e la malattia.
Si stima che l’1-2% degli individui soffra di malattie autoimmuni, sebbene si debba
ammettere che molte malattie associate a risposte immunitarie dirette contro
antigeni ignoti (e quindi non necessariamente self) vengano semplicisticamente
incluse tra le malattie autoimmuni.
Le malattie autoimmuni vengono convenzionalmente suddivise in due categorie:
organo-specifiche e non-organo-specifiche (o sistemiche). Nelle prime la risposta
autoimmunitaria è diretta verso antigeni multipli di un organo. Tipicamente
coinvolgono le ghiandole endocrine e gli autoantigeni sono spesso recettori
ormonali o enzimi intracellulari tipici dell’organo interessato (es. diabete di tipo 1).
Le seconde colpiscono invece più organi e sono associate ad una risposta
immunitaria diretta contro molecole self largamente distribuite nell’organismo, in
particolare molecole intracellulari coinvolte nella trascrizione del DNA (es. artrite
reumatoide).

18
I principali fattori che contribuiscono allo sviluppo dell’autoimmunità sono di
origine genetica (ereditarietà di geni predisponenti che contribuiscono alla mancata
tolleranza verso antigeni self) o di origine ambientale (come le infezioni che
possono attivare in modo patologico linfociti reattivi verso antigeni self) (Fig. 6).
Figura 6 – Meccanismi ipotizzati per lo sviluppo dell’autoimmunità
1.2.1 Diabete mellito di tipo 1 (T1D)
Il diabete mellito di tipo 1 è una forma di diabete che si configura come malattia
autoimmune caratterizzata dalla distruzione delle cellule β pancreatiche e
conseguente insulino-deficienza[6].
La forma più comune di T1D è conosciuta come Insulin-Dependent Diabetes
Mellitus (IDDM) o diabete giovanile.
La velocità con cui le cellule β vengono distrutte è variabile; è stata infatti osservata
una maggiore velocità di distruzione cellulare nei bambini, che risulta invece più
lenta negli adulti. In quest’ultimo caso si parla di Latent Autoimmune Diabetes in
Adults (LADA) [7].

19
Alcuni pazienti, specialmente bambini ed adolescenti, possono presentare una
rapida chetoacidosi come prima manifestazione della patologia. Altri mostrano
invece una più lenta iperglicemia, che assume rapidamente importanza in presenza
di infezioni o stress. Negli adulti, in alcuni casi, si è osservata una conservazione di
un’attività residua delle β cellule, sufficiente a prevenire la chetoacidosi [8].
Le cause scatenanti sono un insieme di fattori che riguardano la genetica,
l'ambiente e l'immunologia. Ad una predisposizione genetica di base si unisce uno
stimolo immunologico che, con il passare del tempo, porta alla distruzione delle
cellule β. L'insorgenza ha variazione stagionale e può seguire, tra l'altro, quella di
diverse patologie virali quali il morbillo, l'epatite o infezioni da Coxsackie Virus. Si
teorizza che tali infezioni realizzino una risposta autoimmunitaria, portando infine
alla comparsa di linfociti T citotossici che completino la distruzione delle cellule β
del pancreas, producenti insulina.
Per quanto riguarda i fattori ambientali, sembra che, come altre malattie
autoimmuni, il T1D sia correlato a particolari combinazioni di alleli HLA (Human
Leukocyte Antigen). La presenza di HLA del cromosoma 6 fu il primo locus ad essere
sospettato di una correlazione col diabete di tipo 1 mentre i B8 o B15 aumentano di
circa tre volte il rischio di sviluppare T1D[9].
L'eziologia genica è però ancora incerta. Inoltre non c'è prevalenza nella
trasmissione verticale.
Non esistono invece ancora prove certe riguardo al coinvolgimento dei fattori
ambientali nello sviluppo del T1D. Una recente ipotesi prende in considerazione
fenomeni di mimetismo con antigeni del Mycobacterium avium subspecie
paratuberculosis, già chiamato in causa per la patogenesi della malattia di Crohn[10].
Infezioni croniche subcliniche di tale micobatterio, contratto durante l'infanzia,
comporterebbero, in individui suscettibili, interazioni crociate di stampo
autoimmunitario, come rilevato in alcuni studi[11].
Inerentemente invece alle infezioni da Coxackie, il sierotipo B4 è stato correlato
all'insorgenza di diabete insulino-dipendente (vedi paragrafo “Correlazione fra T1D
ed infezione da Coxsackie Virus B4: mimetismo” nella sezione “Presupposti della
Ricerca”).

20
1.2.1.1 Ruolo metabolico della GAD
La GAD (Glutamic Acid Decarboxylase) è un enzima il cui ruolo primario è la
decarbossilazione del glutammato (1) (Glu, E) a GABA (2) e CO2 (Schema 1).
Schema 1 – Sintesi del GABA
Esiste nell’organismo umano in due isoforme denominate hGAD65 (65 kDa) e
hGAD67 (67 kDa) (Fig. 7).
Figura 7 – GAD65 e GAD67
Queste sono espresse da due geni diversi, denominati rispettivamente GAD2 e
GAD1, prevalentemente a livello cerebrale e pancreatico.
La GAD67 è caratterizzata da una distribuzione uniforme all’interno della cellula,
mentre la GAD65 è localizzata esclusivamente nelle terminazioni nervose e a livello
della membrana; questa caratteristica riflette una differenza funzionale delle due
GAD
+ CO2
(1) (2)
GAD65 (88-584) GAD67 (93 – 593)

21
isoforme. La GAD67 infatti ha il compito di sintetizzare il GABA per l’attività
neuronale non legata alla neurotrasmissione, some la sinaptogenesi e la protezione
da danni neurali, funzione che richiede una distribuzione ubiquitaria del GABA; la
GAD65 invece sintetizza il GABA specificatamente per la neurotrasmissione, che di
conseguenza è necessario solo a livello delle terminazioni nervose. La GAD67 viene
di conseguenza sviluppata per prima dall’organismo durante la fase iniziale di
sviluppo, mentre la GAD65 viene trascritta solo più tardi[12].
Ancora una volta questo riflette la differente funzione delle due forme
nell’organismo, la prima necessaria durante lo sviluppo per il normale
funzionamento della cellula, la secondo necessaria solo quando l’inibizione
sinaptica è più diffusa.
Entrambe le isoforme sono regolate tramite reazioni di fosforilazione[13], anche se in
modo diametralmente opposto; mentre la GAD65 è attivata dalla fosforilazione, la
GAD67 viene inibita da essa. La GAD67 viene fosforilata sulla treonina in posizione
91 ad opera della protein kinasi A (PKA); la GAD65 invece viene fosforilata dalla
protein kinasi C (PKC).
Esiste poi un ulteriore regolatore comune, denominato piridossal-5-fosfato (PLP). La
GAD risulta attiva solo se legata al PLP[14]; mentre, per gli stessi motivi già elencati
precedentemente, la GAD67 risulta sempre legata al PLP, la GAD65 si lega ad esso,
attivandosi, solo in caso di necessità.
Come già accennato, la GAD, nelle sue 2 isoforme 65 e 67, e di conseguenza il
GABA, sono espressi anche nelle cellule β delle isole di Langerhans, in quantità
paragonabili a quelle incontrate nel SNC.
Il GABA è un γ-amminoacido, principale neurotrasmettitore inibitorio nei
mammiferi del sistema nervoso centrale, responsabile della regolazione
dell'eccitabilità neuronale in tutto il sistema nervoso. Negli esseri umani GABA è
anche direttamente responsabile della regolazione del tono muscolare. Viene
rilasciato da neuroni dei circuiti locali presenti nel cervello (neuroni gabaergici). La
loro funzione fisiologica però rimane ad oggi ancora poco chiara. Alcune ipotesi
suggeriscono che il GABA prodotto nelle β-cellule possa servire da regolatore del
rilascio degli ormoni pancreatici oppure come molecola di segnalazione paracrina

22
per la comunicazione tra le suddette β-cellule e le altre cellule endocrine delle isole
di Langerhans.
La comunicazione paracrina si verifica quando la molecola segnale agisce a
brevissima distanza su cellule bersaglio diverse poste presso la cellula di
segnalazione. La velocità di questo segnale dipende dalla distanza delle cellule
bersaglio e dalla risposta generata che può avvenire in frazioni di secondo, qualora
sia finalizzata a cambiare la conformazione di una proteina, ma può richiedere
anche minuti o ore quando modifica l'espressione genica.
Ci sono inoltre prove convincenti a supporto del fatto che il GABA potrebbe avere
un effetto inibitorio sul rilascio del glucagone (in vitro), anche se non è ancora
chiaro come il GABA agisca da molecola segnale nell’inibizione della secrezione del
glucagone[15]. Il glucagone (Fig. 8) è un ormone peptidico secreto dal pancreas, più
precisamente dalle cellule α delle isole di Langerhans, che ha come bersaglio
principale alcune cellule del fegato; esso permette il controllo dei livelli di glucosio
nel sangue, affinché rimangano entro certi limiti: se il livello ematico di glucosio
scende sotto una soglia di circa 80 - 100 mg/dl (= 0,8 - 1 g/l), le cellule α cominciano
a secernere glucagone.
Figura 8 – Glucagone
Di seguito è riportata la sequenza completa del glucagone umano
(http://www.uniprot.org).
10 20 30 40 50 MKSIYFVAGL FVMLVQGSWQ RSLQDTEEKS RSFSASQADP LSDPDQMNED 60 70 80 90 100 KRHSQGTFTS DYSKYLDSRR AQDFVQWLMN TKRNRNNIAK RHDEFERHAE 110 120 130 140 150 GTFTSDVSSY LEGQAAKEFI AWLVKGRGRR DFPEEVAIVE ELGRRHADGS 160 170 180 FSDEMNTILD NLAARDFINW LIQTKITDRK

23
Questo si lega immediatamente ai suoi recettori presenti principalmente sugli
epatociti, attivando la degradazione del glicogeno (glicogenolisi) ed un conseguente
rilascio di glucosio nel sangue. Sebbene venga in genere considerato antagonista
dell'insulina per il suo compito di contrastare l'ipoglicemia, insulina e glucagone
intervengono in sinergia in seguito all'introduzione di proteine/amminoacidi, poiché
il primo determina lo stivaggio degli amminoacidi (proteosintesi) nei tessuti, mentre
il secondo previene l'ipoglicemia causata dall'insulina[16]. In Figura 9 è
rappresentato schematicamente il ciclo dell’insulina e del glucagone, a seconda che
l’organismo si trovi in condizioni di alta o bassa glicemia.
Figura 9 – Schema del ciclo dell’insulina e del glucagone
1.2.1.2 Coxsackie Virus
I Coxsackie sono virus citolitici della famiglia dei Picornaviridae, degli Enterovirus. Ci
sono 61 Enterovirus non poliomielitici che possono causare malattie umane, di
questi 23 sono Coxsackie A e 6 sono Coxsackie B. Gli Enterovirus sono il secondo più
comune agente infettante virale nella specie umana (dopo i Rhinovirus).
La classificazione moderna non prevede più che il genere Enterovirus abbia come
uno dei suoi gruppi i Coxsackievirus, ma che sia diviso, più rigorosamente, in varie
specie fra cui i Poliovirus, gli Enterovirus umani A (con vari Coxsackie A), gli
Enterovirus umani B (con i Coxsackie B), gli Enterovirus umani C (con altri Coxsackie

24
A), ecc. Inoltre due Coxsakie (A4 e A6) non sono ancora stati assegnati a nessuna
delle precedenti specie.
Tra le malattie causate dai sierotipi Coxsackie A, la più famosa è la malattia mano-
piede-bocca, ma anche congiuntivite emorragica, herpangina, e meningite asettica
(per i sierotipi sia A che B). Il Coxsackie B causa anche miocardite e pericardite
infettive e pleurodinia (malattia di Bornholm). Secondo recenti ipotesi, i virus
Coxsackie produrrebbero alterazioni a carico delle cellule β del pancreas, fenomeno
che quindi li chiama in causa come possibile fattore eziologico del T1D insulino-
dipendente. Tuttavia la sua virulenza è strettamente correlata alla funzionalità del
sistema immunitario: tale agente infettivo ha maggior facilità a manifestare
determinate patologie in un soggetto molto giovane o molto anziano o comunque
immunocompromesso piuttosto che in individui adulti e sani.
In Figura 10 è riportato a titolo esemplificativo il sottogruppo Coxsackie B3.
Figura 10 – Coxsackie Virus B3

25
2 PRESUPPOSTI DELLA RICERCA
2.1 CORRELAZIONE FRA T1D ED INFEZIONE DA
COXSACKIE VIRUS: IPOTESI DEL MIMETISMO
Negli ultimi anni è risultato sempre più evidente il coinvolgimento dei fattori
ambientali, in particolar modo le infezioni di tipo virale, nello sviluppo del T1D. Fra
gli agenti patogeni più comunemente associati a questa patologia si ritrovano
soprattutto virus appartenenti al genere degli enterovirus, e nello specifico il
sottogruppo B4 del Coxsackie Virus, appartenente alla specie degli enterovirus
umani B.
A partire dalla fine degli anni ’90, numerosi studi hanno cercato di comprendere
quale fosse la correlazione fra l’infezione e l’insorgere della patologia.
I danni evidenti riscontrati nei soggetti affetti da T1D e risultati positivi ad infezione
da Coxsackie sono fondamentalmente due:
• danneggiamento diretto e distruzione delle β-cellule delle isole
pancreatiche[17]
• reazione autoimmune con danneggiamento del processo
chimico/metabolico di regolazione del glucosio nel sangue[18]
2.1.1 Danneggiamento diretto e distruzione delle β-cellule
delle isole pancreatiche
In caso di infezione particolarmente aggressiva, le β-cellule delle isole pancreatiche
infette possono essere direttamente distrutte dalla citolisi, ovvero dalla rottura
della membrana cellulare, indotta dal virus. La distruzione diretta delle isole implica
ovviamente una cessazione della produzione di insulina ed un conseguente
aumento dei livelli di glucosio nel sangue, che sfocia poi nelle complicanze già
descritte precedentemente.
Un’infezione meno aggressiva potrebbe invece causare una reazione infiammatoria
nelle isole, generando una distruzione delle cellule β a livello subclinico ed un
conseguente rilascio di antigeni normalmente sequestrati, che quindi attivano la
risposta di cellule T, autoreattive patogene[17][19].

26
2.1.2 Reazione autoimmune: danneggiamento del processo
chimico/metabolico di regolazione del glucosio nel
sangue
Tutte le ipotesi gravitano intorno ad un evidente fattore comune, ovvero la possibile
omologia (Fig. 12) tra una porzione della sequenza amminoacidica della proteina
virale genomica del Coxsackie Virus B4, denominata protein 2C o P2C (1099 – 1427)
(Fig. 11), e due porzioni delle due isoforme, 65 (250 – 273) e 67 (259 – 282), della
glutammico decarbossilasi umana, o GAD, espressa nelle cellule β del pancreas da
un unico gene.
Figura 11 – CVB4 genome polyprotein (http://www.uniprot.org)

27
Figura 12 – Omologia nelle sequenze di GAD e P2C virale[18]
Di seguito sono riportate le sequenze complete delle due isoforme della GAD e
della P2C, nelle quali sono stati evidenziati i frammenti ipotizzati quali mimetici e
sintetizzati in questo lavoro di tesi (http://www.uniprot.org).
GAD67
10 20 30 40 50 MASSTPSSSA TSSNAGADPN TTNLRPTTYD TWCGVAHGCT RKLGLKICGF 60 70 80 90 100 LQRTNSLEEK SRLVSAFKER QSSKNLLSCE NSDRDARFRR TETDFSNLFA 110 120 130 140 150 RDLLPAKNGE EQTVQFLLEV VDILLNYVRK TFDRSTKVLD FHHPHQLLEG 160 170 180 190 200 MEGFNLELSD HPESLEQILV DCRDTLKYGV RTGHPRFFNQ LSTGLDIIGL 210 220 230 240 250 AGEWLTSTAN TNMFTYEIAP VFVLMEQITL KKMREIVGWS SKDGDGIFSP 260 270 280 290 300 GGAISNMYSI MAARYKYFPE VKTKGMAAVP KLVLFTSEQS HYSIKKAGAA 310 320 330 340 350 LGFGTDNVIL IKCNERGKII PADFEAKILE AKQKGYVPFY VNATAGTTVY 360 370 380 390 400 GAFDPIQEIA DICEKYNLWL HVDAAWGGGL LMSRKHRHKL NGIERANSVT 410 420 430 440 450 WNPHKMMGVL LQCSAILVKE KGILQGCNQM CAGYLFQPDK QYDVSYDTGD 460 470 480 490 500 KAIQCGRHVD IFKFWLMWKA KGTVGFENQI NKCLELAEYL YAKIKNREEF 510 520 530 540 550 EMVFNGEPEH TNVCFWYIPQ SLRGVPDSPQ RREKLHKVAP KIKALMMESG 560 570 580 590 TTMVGYQPQG DKANFFRMVI SNPAATQSDI DFLIEEIERL GQDL
Il frammento evidenziato (259 – 282) è caratterizzato strutturalmente da:
• Helix (252 – 267)
• Helix (270 – 274)
• Helix ( 276 – 278)
• Beta strand (282 – 287)

28
GAD65
10 20 30 40 50 MASPGSGFWS FGSEDGSGDS ENPGTARAWC QVAQKFTGGI GNKLCALLYG 60 70 80 90 100 DAEKPAESGG SQPPRAAARK AACACDQKPC SCSKVDVNYA FLHATDLLPA 110 120 130 140 150 CDGERPTLAF LQDVMNILLQ YVVKSFDRST KVIDFHYPNE LLQEYNWELA 160 170 180 190 200 DQPQNLEEIL MHCQTTLKYA IKTGHPRYFN QLSTGLDMVG LAADWLTSTA 210 220 230 240 250 NTNMFTYEIA PVFVLLEYVT LKKMREIIGW PGGSGDGIFS PGGAISNMYA 260 270 280 290 300 MMIARFKMFP EVKEKGMAAL PRLIAFTSEH SHFSLKKGAA ALGIGTDSVI 310 320 330 340 350 LIKCDERGKM IPSDLERRIL EAKQKGFVPF LVSATAGTTV YGAFDPLLAV 360 370 380 390 400 ADICKKYKIW MHVDAAWGGG LLMSRKHKWK LSGVERANSV TWNPHKMMGV 410 420 430 440 450 PLQCSALLVR EEGLMQNCNQ MHASYLFQQD KHYDLSYDTG DKALQCGRHV 460 470 480 490 500 DVFKLWLMWR AKGTTGFEAH VDKCLELAEY LYNIIKNREG YEMVFDGKPQ 510 520 530 540 550 HTNVCFWYIP PSLRTLEDNE ERMSRLSKVA PVIKARMMEY GTTMVSYQPL 560 570 580 GDKVNFFRMV ISNPAATHQD IDFLIEEIER LGQDL
Il frammento evidenziato (250 – 273) è caratterizzato strutturalmente da:
• Helix (243 – 258)
• Helix (262 – 265)
• Helix (267 – 269)
• Beta strand (273 – 278)
Recentemente sono state individuate due subunità della GAD65 denominate α e β,
aventi differente mobilità in elettroforesi su gel di SDS-poliacrilammide ed aventi
differente comportamento nei confronti di reazioni come la fosforilazione (solo la
subunità α viene fosforilata, sia in vivo che in vitro), fondamentale al fine di regolare
la funzione della proteina delle membrane vescicolari sinaptiche[20].

29
CVB4 P2C
1100 NN 1110 1120 1130 1140 1150 NWLKKFTEMT NACKGMEWIA VKIQKFIEWL KVKILPEVKE KHEFLNRLKQ 1160 1170 1180 1190 1200 LPLLESQIAT IEQSAPSQSD QEQLFSNVQY FAHYCRKYAP LYAAEAKRVF 1210 1220 1230 1240 1250 SLEKKMSNYI QFKSKCRIEP VCLLLHGSPG AGKSVATNLI GRSLAEKLNS 1260 1270 1280 1290 1300 SVYSLPPDPD HFDGYKQQAV VIMDDLCQNP DGKDVSLFCQ MVSSVDFVPP 1310 1320 1330 1340 1350 MAALEEKGIL FTSPFVLAST NAGSINAPTV SDSRALARRF HFDMNIEVIS 1360 1370 1380 1390 1400 MYSQNGKINM PMSVKTCDEE CCPVNFKKCC PLVCGKAIQF IDRRTQVRYS 1410 1420 1430 LDMLVTEMFR EYNHRHSVGA TLEALFQ
La P2C ha diversi ruoli, fra cui la morfogenesi del virione e l’incapsidazione dell’RNA
virale per interazione con la proteina VP3 del capside (per similarità[21]).
La similarità di due sequenze è una quantità misurabile che può essere espressa
come, ad esempio, percentuale di identità. Confrontando infatti le tre sequenze a
coppie tramite algoritmi di BLAST, si ottiene:
• hGAD65 (250-273) / hGAD67 (258-281) = 91% di sovrapposizione, 16
amminoacidi identici
• hGAD65 (250-273) / CVB4 P2C (28-50) = 37% di sovrapposizione, 7
amminoacidi identici, rispettivamente tutti nella regione (261-266) / (39-44)
• hGAD67 (258-281) / CVB4 P2C (28-50) = 26% di sovrapposizione, 5
amminoacidi identici, rispettivamente tutti nella regione (269-274) / (39-44)
Si evidenzia quindi in particolare la presenza di un frammento identico nelle
sequenze di hGAD65 e CVB4 P2C, ovvero PEVKEK, leggermente diverso invece nella
hGAD67 (PEVKTK). L’importanza di questo frammento come epitopo è stata
dimostrata in alcuni recenti studi, nei quali viene mostrato come mutazioni indotte
in questa regione riducono di circa l’80% la reattività di anticorpi monoclonali
specifici contro le due isoforme della GAD, rispetto alle forme intatte[22].
Questa omologia nei suddetti frammenti fra la GAD e la P2C del Coxsackie Virus,
teoricamente, può condurre ad una cross-reattività con esito di autoimmunità, che

30
inizialmente si manifesta come blocco della prima fase di secrezione dell’insulina,
quindi degenera in un vero e proprio danneggiamento delle isole di Langerhans[17].
In seguito all’infezione da Coxsackie, il sistema immunitario produce naturalmente
degli anticorpi specifici (principalmente IgM prima ed IgG dopo), contro le proteine
virali, come la P2C in questo caso. È stato quindi ipotizzato che, a causa della
similitudine di alcune porzioni delle suddette sequenze, gli anticorpi prodotti contro
le proteine virali non riescano a differenziare queste ultime dalle due isoforme della
GAD, attaccando indiscriminatamente sia l’una che le altre. In altre parole, si può
verificare una cross-reattività anticorpo-antigeni quando antigeni virali e antigeni
self condividono gli stessi determinanti antigenici.
Come già accennato precedentemente, la GAD ha la funzione biologica di produrre
il GABA per decarbossilazione dell’acido glutammico. Ci sono prove convincenti a
supporto del fatto che il GABA potrebbe avere un effetto inibitorio sul rilascio del
glucagone (in vitro), anche se non è ancora chiaro come il GABA agisca da molecola
segnale nell’inibizione della secrezione del glucagone[15]. L’attacco quindi da parte
degli anticorpi anti-Coxsackie sulle isoforme della GAD potrebbe conseguentemente
portare ad uno squilibrio nel sistema di gestione e regolazione dei livelli di glucosio
nel sangue, degenerando poi nella patologia oggetto del nostro studio.
A supporto di tale ipotesi, è stato dimostrato che topi NOD giovani trattati per via
endovenosa con GAD65 o GAD67 ricombinante sono stati significativamente
protetti da T1D. L’effetto antidiabetogenico è stato riscontrato anche
somministrando GAD65 per altre vie (intraperitoneale, intranasale, orale)[23].
Da segnalare è anche la presenza di specifici anticorpi anti-GAD nella quasi totalità
dei sieri provenienti da pazienti diabetici in caso di diabete giovanile, a conferma
comunque del ruolo, per quanto ancora ignoto, di questo enzima nello sviluppo e
nel meccanismo di questa patologia.
La maggior parte della letteratura disponibile a riguardo, risalente all’ultimo
decennio, concentra i propri sforzi nel cercare di evidenziare un’eventuale
correlazione o riconoscimento degli antigeni self (GAD) e di quelli virali (P2C) da
parte di cellule T. Solo recentemente l’attenzione della ricerca è stata posta anche
su molecole anticorpali di tipo IgM ed IgG, purtroppo con risultati spesso
discordanti o deludenti.

31
2.1.2.1 Anticorpi anti-GAD
La letteratura relativa gli studi effettuati finora in merito all’insorgenza ed allo
sviluppo del T1D riporta la proteina GAD come uno dei più importanti autoantigeni
legati a questa patologia[18][24][25][26]. A livello clinico il test per la rilevazione di tali
anticorpi è commercialmente disponibile ed effettuato di routine sui pazienti
diabetici. Viene considerato un marker predittivo importante, associato ad un alto
rischio di sviluppo della patologia[26]. Gli anticorpi anti-GAD rilevati attraverso
questo test appartengono alla classe delle IgG. Si ritrovano nel 65-85 % dei pazienti
affetti da IDDM e nel 70-90 % dei pazienti affetti da LADA, generalmente in
concentrazioni inferiori a 100 U/mL. I valori standard per uomini donne o bambini
stanno in un range fra 0 e 1 U/mL. Sotto le 10 U/ml il test viene comunque
dichiarato negativo.
Il test in questione è un test SP-ELISA quantitativo in vitro (Fig. 13). Come antigene
viene utilizzato hGAD65 ricombinante.
Figura 13 – Test SP-ELISA per la rilevazione di IgG anti-GAD.

32
3 SVILUPPO DELLA RICERCA
Il diabete mellito di tipo 1 (T1D) viene classificato all’interno delle patologie
autoimmuni e può presentarsi in due forme: giovanile (Insulin-Dependent Diabetes
Mellitus o IDDM) nei primi anni di vita dell’individuo, o adulto (Latent Autoimmune
Diabetes in Adult o LADA). Fra i più importanti autoantigeni legati a questa
patologia viene riportata la proteina acido glutammico decarbossilasi o GAD.
Questa esiste in due isoforme, GAD65 e GAD67. Negli ultimi anni è stata
evidenziata un’omologia fra le sequenze già molto simili delle due isoforme della
GAD con quella di una porzione, denominata P2C, della poliproteina genomica di un
virus appartenente al genere degli enterovirus, il Coxsackie B4 (CVB4). Il legame
esistente fra l’infezione da CVB4 e lo sviluppo dell’IDDM è sempre più evidente,
nonostante i meccanismi che determinano tale legame siano ancora ignoti. In
particolare, è stato individuato un sequone (PEVKEK) identico in tutte e tre le
sequenze. Sono state avanzate quindi numerose ipotesi, finora mai del tutto
verificate, riguardanti un possibile effetto di mimetismo nei confronti delle
sequenze in esame. L’esistenza di questo fenomeno porterebbe quindi ad una
mancata differenziazione, da parte degli anticorpi sviluppati per contrastare
l’infezione, della sequenza virale rispetto a quelle autoantigeniche. Avendo la GAD
un ruolo di regolazione nei confronti del glucagone, il suo danneggiamento e la sua
distruzione da parte di questi anticorpi porterebbero ad uno squilibrio nell’intero
sistema di regolazione del metabolismo del glucosio che sfocerebbe infine nella
patologia.
In base a questi presupposti, l’obiettivo di questo lavoro di tesi è quello di porre le
basi per cominciare a verificare l’esistenza di questo fenomeno di mimetismo, così
da poter cominciare a fare chiarezza sul reale conivolgimento del CVB4 nello
sviluppo del T1D giovanile.
A tal proposito sono quindi stati identificati e sintetizzati i tre frammenti peptidici
contenenti il sequone comune:
• hGAD65 (250-273) – 250AMMIARFKMFPEVKEKGMAALPRL273
• hGAD67 (258-281) – 258SIMAARYKYFPEVKTKGMAAVPKL281
• CVB4 P2C (28-50) – 28FIEWLKVKILPEVKEKHEFLSRL50

33
Le tre molecole sono state realizzate, sfruttando la strategia di sintesi di peptidi su
fase solida (SPPS) assistita da microonde tramite lo strumento Liberty BlueTM CEM,
sia in forma N-terminale libera (peptidi I, II, III) che acetilata (peptidi IV, V, VI). È
stato scelto di fare questo per valutare l’eventuale importanza di un ulteriore
legame ammidico in posizione N-terminale in termini di mimetismo da parte del
frammento rispetto alla proteina completa. Con i suddetti due gruppi di peptidi
sono stati quindi effettuati dei test immunoenzimatici e di immunoaffinità tramite
tecniche ELISA (per i peptidi IV, V, VI acetilati) e Biacore (per i peptidi I, II, III non
acetilati) su sieri, forniti dal reparto di immunologia pediatrica dall’azienda
ospedaliera Meyer di Firenze (IDDM) e dall’università di Padova (LADA), provenienti
da pazienti diabetici in età infantile o adolescenziale nel primo caso ed adulta nel
secondo. I sieri dei pazienti sono stati selezionati dalla Prof. Chiara Azzari (IDDM) e
dalla Prof. Annunziata Lapolla (LADA) ed appartengono a pazienti affetti da diabete I
ed a donatori sani. I pazienti sono stai selezionati in maniera casuale e non
precedentemente sottoposti a test per la reattività immune tramite test ELISA o
BIACORE. Questo studio, i metodi e le procedure di consenso sono stati approvati
dal Comitato Etico dell’Azienda Ospedaliera Universitaria Meyer di Firenze e
dell’Azienda Ospedaliera Universitaria Padovana. Tutti i dati utilizzati in questo
studio sono stati riportati in forma anonima.
Tramite questi test si è voluto studiare una eventuale cross-reattività fra i suddetti
anticorpi ed antigeni self, quali le due isoforme della GAD, e non-self, quali la
proteina virale, al fine di compiere una prima indagine preliminare per far luce
sull’ormai evidente ruolo dell’infezione virale nello sviluppo dell’autoimmunità
legata alla patologia del T1D e sul meccanismo che lega questi due eventi.
3.1 SPPS
La sintesi di peptidi su fase solida (SPPS) consente di far accrescere la catena
peptidica nella direzione C-terminale → N-terminale, a partire dal primo
amminoacido ancorato ad una matrice solida insolubile (resina polistirenica
funzionalizzata). Il supporto solido deve essere meccanicamente stabile ed inerte
nei confronti dei reagenti utilizzati per la sintesi. Per la sintesi dei tre peptidi

34
oggetto di questo lavoro di tesi si è scelto di utilizzare una resina di tipo Fmoc-Rink
Amide (Fig. 14).
Figura 14 – Resina Fmoc-Rink Amide
Tutte le operazioni di allungamento avvengono in fase eterogenea, con la possibilità
di rimuovere i prodotti secondari tramite lavaggi senza dover isolare ogni
intermedio di reazione. Il legame tra l’ultimo amminoacido della sequenza peptidica
e il supporto solido avviene tramite una molecola bifunzionale (linker). Questo
conferisce in genere una buona stabilità nei confronti di tutte le operazioni di sintesi
e di crescita del peptide e garantisce un distacco sicuro del prodotto dal supporto
una volta terminata la sintesi, evitando il più possibile danni alla struttura (per
quanto sensibile possa essere). La scelta del linker deve essere valutata
attentamente anche in base al tipo di peptide che si vuole ottenere.
Gli amminoacidi introdotti nella catena devono avere una protezione temporanea
sulla funzione α-amminica ed eventualmente una protezione semipermanente in
catena laterale (eliminata durante il distacco del peptide dalla resina). Nella
strategia Fmoc/tBu (introdotta da Sheppard e Atherton negli anni ’70) la protezione
temporanea sulla funzione amminica è rappresentata dal gruppo Fmoc (labile in
ambiente basico), mentre le eventuali protezioni semipermanenti presenti sulle
catene laterali degli amminoacidi sono rappresentate dai gruppi Pbf, tBu, Boc e Trt
(labili in ambiente acido) (Fig. 15). Questi gruppi protettori sono caratterizzati
quindi da un alto grado di selettività (ortogonalità) che consente di rimuoverli
indipendentemente (in condizioni diverse) l’uno dall’altro.

35
CH2
O
C O
Fmoc: Fluorenylmethoxycarbonyl
S OO
CH3
CH3
H3C
CH3
H3C
Pbf: 2,2,4,6,7-Pentamethyl-dihydrobenzofurane-5-sulfonyl
C
CH3
CH3H3C
tBu: t-Butyl
C
Trt: Trityl
C O
O
C CH3
CH3
H3C
Boc: t-Butoxycarbonyl
Figura 15 – Principali gruppi protettori
In questo lavoro di tesi, oltre al gruppo Fmoc per la protezione di tutte le estremità
N-terminali, sono stati utilizzati i seguenti gruppi protettori per la protezione
temporanea dei seguenti amminoacidi: Pbf per Arg, Trt per Hys, Boc per Lys e Trp e
tBu per Ser, Thr e Tyr.
Ciascun ciclo di accoppiamento per ogni amminoacido della sequenza prevede
quattro passaggi:
• deprotezione del gruppo amminico della funzione N-terminale dell’ultimo
amminoacido inserito nella catena che si accresce sulla resina;
• lavaggio dopo la deprotezione;
• accoppiamento con l’amminoacido protetto sulla funzione amminica in alfa
come Fmoc;
• lavaggio dopo l’accoppiamento.
La deprotezione del gruppo amminico della resina per l’ancoraggio del primo
amminoacido, e del gruppo α-amminico dell’ultimo amminoacido legato, è stata
effettuata trattando la resina con una soluzione di piperidina al 20% in DMF, seguito
da una serie di lavaggi con DMF per eliminare i residui di base.
36
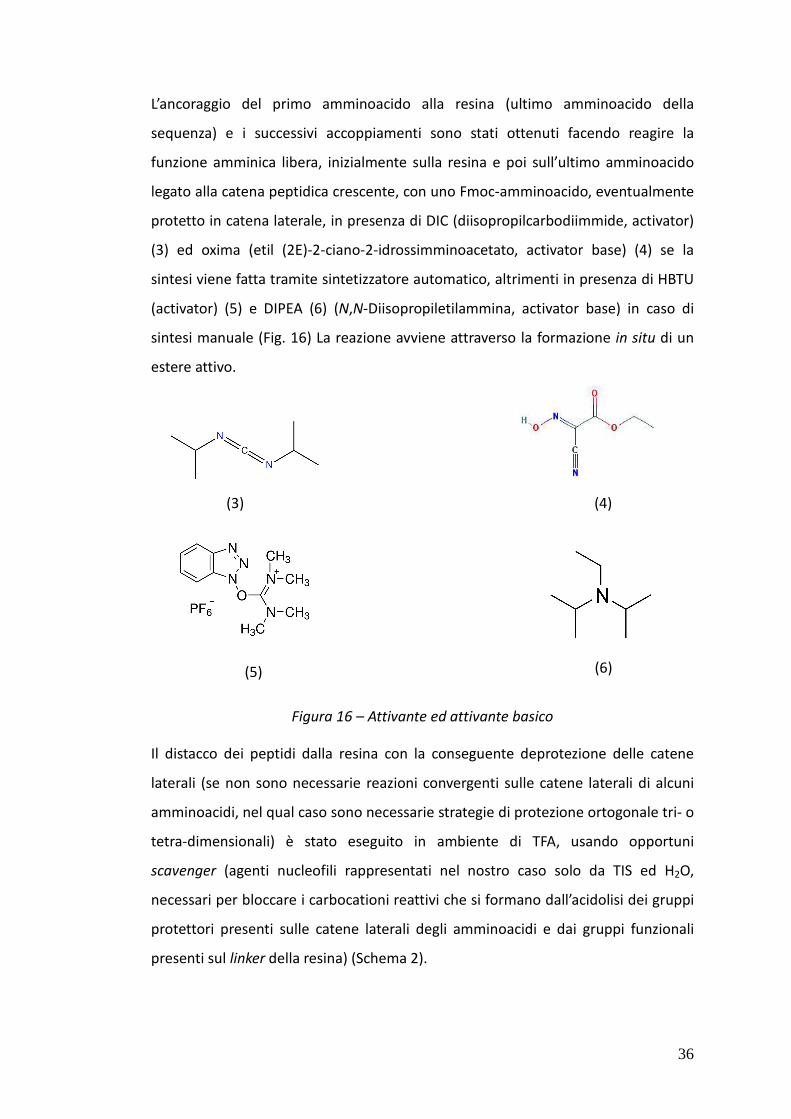
L’ancoraggio del primo amminoacido alla resina (ultimo amminoacido della
sequenza) e i successivi accoppiamenti sono stati ottenuti facendo reagire la
funzione amminica libera, inizialmente sulla resina e poi sull’ultimo amminoacido
legato alla catena peptidica crescente, con uno Fmoc-amminoacido, eventualmente
protetto in catena laterale, in presenza di DIC (diisopropilcarbodiimmide, activator)
(3) ed oxima (etil (2E)-2-ciano-2-idrossimminoacetato, activator base) (4) se la
sintesi viene fatta tramite sintetizzatore automatico, altrimenti in presenza di HBTU
(activator) (5) e DIPEA (6) (N,N-Diisopropiletilammina, activator base) in caso di
sintesi manuale (Fig. 16) La reazione avviene attraverso la formazione in situ di un
estere attivo.
Figura 16 – Attivante ed attivante basico
Il distacco dei peptidi dalla resina con la conseguente deprotezione delle catene
laterali (se non sono necessarie reazioni convergenti sulle catene laterali di alcuni
amminoacidi, nel qual caso sono necessarie strategie di protezione ortogonale tri- o
tetra-dimensionali) è stato eseguito in ambiente di TFA, usando opportuni
scavenger (agenti nucleofili rappresentati nel nostro caso solo da TIS ed H2O,
necessari per bloccare i carbocationi reattivi che si formano dall’acidolisi dei gruppi
protettori presenti sulle catene laterali degli amminoacidi e dai gruppi funzionali
presenti sul linker della resina) (Schema 2).
(3) (4)
(5) (6)

37
CH3C
O
ResinaLinkerHNC
O
CH
Rm
HNC
O
CH
Rn
Gruppo protettorein catena laterale
Gruppo protettorein catena laterale
n-1
HN
CH3C
O
NH2C
O
CH
Rm
HNC
O
CH
Rn
n-1
HN
TFA
TFA
TFA/H2O/TISt. a., 4 h
Schema 2 – Reazione di distacco del peptide dalla resina e contemporanea
deprotezione delle catene laterali
3.1.1 Teoria e applicazioni delle microonde alla SPPS
Sin dalla sua introduzione, è stato dimostrato che l’energia delle microonde è in
grado di aumentare la velocità di reazione, ridurre la presenza di reazioni
secondarie e permettere una maggiore scelta e risparmio di solventi.
Nella sintesi peptidica su fase solida, in particolare, l’uso delle microonde ha
ricoperto un ruolo di notevole importanza negli ultimi anni.
Le microonde sono una forma di energia elettromagnetica situata nello spettro fra
la radiazione infrarossa e le onde radio con frequenze nella scala tra 0.3-300 GHz
(ovvero lunghezze d'onda di 1 mm-1 m) (Fig. 17). Soltanto la frequenza di 2.45 GHz,
corrispondente ad una lunghezza d'onda di 12.24 cm, molto vicina alla frequenza di
rotazione molecolare, è utilizzata negli strumenti industriali e domestici a
microonde, per evitare interferenze con i radar e le attività di telecomunicazioni che
funzionano all'interno di questa scala.

38
Figura 17 – Lo spettro elettromagnetico
Quando una molecola è irraggiata da una frequenza esattamente pari alla sua
frequenza di rotazione, inizia a ruotare, mentre i dipoli o gli ioni molecolari provano
ad allinearsi al campo elettrico alternato delle microonde e ad assorbire energia,
innescando un processo di rotazione dipolare e conduzione ionica.
I vantaggi principali dell’utilizzo delle microonde sono essenzialmente legati ad un
trasferimento di energia dalla radiazione al composto più veloce e allo stesso tempo
più omogeneo rispetto alle tecniche di riscaldamento convenzionali.
Le microonde sono costituite da un campo elettrico e da un campo magnetico, tra
loro perpendicolari, in cui solo il primo interagisce con le molecole aumentando la
velocità delle reazioni tramite la conduttività dipolare o ionica (Figura 18).
Figura 18 – L’onda elettromagnetica

39
Il riscaldamento con le microonde sfrutta la capacità di alcune molecole (liquide o
solide), sulla base delle loro proprietà dielettriche, di trasformare l'energia
elettromagnetica in calore[27]. Questo riscaldamento dielettrico può avvenire
attraverso due meccanismi: polarizzazione dipolare e conduzione ionica (Fig. 19).
Figura 19 – Riscaldamento tramite microonde
Dato che solo il campo elettrico trasferisce energia per riscaldare una sostanza, la
sua azione induce i dipoli e gli ioni ad allinearsi al campo elettrico generato dalle
microonde. Poiché l'allineamento tuttavia non è mai completo, una parte di questa
energia è persa come calore e causa l’aumento improvviso della temperatura
molecolare e, quindi, dell’intera miscela di reazione.
L'irradiazione delle microonde è veloce e volumetrica, cioè la soluzione si riscalda
uniformemente, contrariamente al riscaldamento convenzionale dove il calore
diffonde dalla superficie del materiale e lentamente viene trasferito al bulk.
Non a caso l’utilizzo a volte di opportuni solventi, che possono assorbire la
radiazione, può incrementare la temperatura dell’ambiente di reazione e favorirne
lo svolgimento[26]. Gli idrocarburi per esempio non possono assorbire direttamente
le microonde, mentre solventi polari a basso peso molecolare ed alta costante
dielettrica, sono invece in grado di aumentare molto velocemente la loro
temperatura (es. acqua, metanolo, cloroformio, acetone, DMF, solvente tipico della
SPPS).

40
Un fenomeno indesiderato, tuttavia, di questa tecnica è l’incremento imprevisto
della temperatura della soluzione, che può portare ad un acceleramento anomalo
della reazione. In genere è causato dalla presenza di ioni o gruppi ionizzati
nell’ambiente di reazione. I vantaggi derivati comunque dall’utilizzo delle
microonde sono tali da consentirne l’impiego in molti ambiti, ad esempio per
migliorare i processi, modificare la selettività e persino per effettuare reazioni che
con riscaldamenti convenzionali non avverrebbero.
3.1.2 Microonde in sintesi peptidica
La sintesi in soluzione assistita dalle microonde ha trovato un grande successo nella
chimica organica, come confermato dalla vasta letteratura in questo campo,
migliorando fortemente la resa ed il completamento di molte reazioni[28]. Il grande
successo ottenuto in campo organico ne ha permesso anche l’applicazione nel
campo della chimica dei peptidi (in particolare della SPPS), determinando nel 1992
la pubblicazione del primo lavoro sull’uso di questa energia impiegando un forno a
microonde multimodale domestico. In questo studio tale energia era stata sfruttata
per accelerare la reazione di coupling con un’anidride asimmetrica e i metodi di
attivazione con l’estere attivo dell’N-idrossibenzotriazolo preformato in situ.
Dal momento che il successo di una sintesi peptidica necessita del completamento
di due reazioni per ciclo (deprotezione e accoppiamento), l’energia delle microonde
rappresenta una via efficiente per portare a termine queste reazioni.
Un problema frequente in sintesi peptidica, in particolare in sequenze peptidiche
lunghe (30 o più amminoacidi) o idrofobiche è l’aggregazione della catena. Si ritiene
che l’energia derivata dalle microonde sia in grado di rompere l’aggregazione
permettendo al peptide di assumere la conformazione più favorevole a minore
energia. L’uso delle microonde infine può talvolta migliorare, in termini di resa e di
purezza, la sintesi in fase solida convenzionale, che invece avviene a temperatura
ambiente. Entrambi i gruppi di peptidi sintetizzati in questo lavoro di tesi sono state
ottenute attraverso sintesi automatica coadiuvata da microonde (Liberty BlueTM
CEM).

41
3.1.2.1 Reazioni secondarie
Durante la sintesi peptidica possono avvenire alcune reazioni secondarie , come la
racemizzazione, la formazione dell’aspartimmide, la formazione del lattame
dell’arginina, il capping della guanidina e la formazione della dichetopiperazina.
Queste potenziali reazioni secondarie sono in generale ben documentate ed
esistono una serie di protocolli e precauzioni in grado di limitarle.
Racemizzazione
Le proprietà dei peptidi e delle proteine dipendono criticamente dalla
configurazione dei loro centri chirali. L’alterazione di un singolo centro chirale può
avere un effetto drastico sull’attività biologica: a parte la Gly, gli altri 20
amminoacidi naturali contengono un centro chirale sull’atomo di carbonio α e in più
l’Ile e la Thr ne contengono un altro in catena laterale. Nella SPPS la racemizzazione
è ben documentata durante la reazione di coupling, tuttavia alcuni casi sono stati
accertati anche durante le fasi di deprotezione catalizzate da basi. Durante la
reazione di coupling usando amminoacidi come His e Cys, sono frequenti
l’epimerizzazione, attraverso la formazione di intermedi ossazolone, o la diretta
enolizzazione. Infatti, durante il coupling, la conversione di tali amminoacidi nel
corrispondente estere attivo aumenta notevolmente l’acidità del carbonio α e
quindi la sua tendenza ad enolizzare e al riarrangiamento interno. Anche reazioni di
coupling ad elevate temperature tramite tecniche convenzionali o con le
microonde, possono determinare un aumento dell’epimerizzazione. Il fenomeno
dell’epimerizzazione viene di solito ridotto utilizzando un metodo di coupling
modificato, in cui le microonde vengono applicate nell’ultima parte
dell’accoppiamento. Una volta inseriti nel peptide, tali amminoacidi sono protetti
dall’epimerizzazione durante i successivi step di deprotezione e di coupling fino a
temperature di massimo 80°C.
Formazione dell’aspartimmide
Una delle più comuni e problematiche reazioni secondarie nella SPPS è la
formazione dell’aspartimmide (Schema 3), che di solito avviene in sequenze
contenenti il frammento “Asp X” (dove X=Gly, Asn, Ser,Thr). Questo fenomeno
coinvolge l’attacco dell’atomo di azoto, legato al carbonio α, di un acido aspartico o
di una asparagina sulla catena laterale di un gruppo estereo o di un gruppo

42
ammidico, rispettivamente. Il successivo attacco nucleofilo causa poi l’apertura
successiva dell’anello, determinando la formazione di una miscela di α- e β-aspartil
peptidi.
Schema 3 – Formazione dell’aspartimmide
Ogni ciclo di deprotezione dopo la sequenza “Asp-X” incrementa la possibilità di
formazione dell’aspartimmide, determinando così numerosi problemi durante la
sintesi di peptidi a lunga catena e con residui multipli di Asp. Il frammento “Asp-X”
in particolare è molto suscettibile alla formazione dell’aspartimmide, a causa della
mancanza di ingombro sterico della Gly in grado di inibire tale reazione.
L’uso della piperazina al posto della piperidina ha portato a notevoli diminuzioni dei
livelli di aspartimmide sia nella sintesi convenzionale che con le microonde. Tuttavia
la piperazina, avendo una pKa di 9.8 rispetto alla piperidina che invece presenta una
pKa di 11.1, risulta essere un reagente di deprotezione ad azione più lenta e poco
adatto per la sintesi convenzionale di sequenze peptidiche idrofobiche,
determinando una scarsa rimozione del gruppo Fmoc.
Formazione del lattame dell’Arg
Durante la reazione di coupling, la catena laterale nucleofila dell’Arg risulta
suscettibile alla formazione del δ-lattame (Schema 4). L’attivazione dell’acido
carbossilico promuove facilmente l’attacco da parte del gruppo δ-guanidinico
altamente basico (pKa=12.5) dell’Arg attraverso una ciclizzazione intramolecolare.

43
Questa reazione collaterale irreversibile rende inattivo il derivato argininico attivato,
durante il coupling.
Schema 4 – Ciclizzazione intramolecolare dell’Arg
Durante le normali sintesi peptidiche questo fenomeno può non essere riscontrato,
dal momento che la reazione di coupling avverrà sicuramente prima che si formi
una considerevole quantità di δ-lattame. Tuttavia, nella reazioni di coupling più
complesse, la formazione del δ-lattame può risultare favorita tanto quanto quella di
coupling: in tal caso persino un prolungamento della durata del tempo di coupling
non sarà in grado di portare a completamento la reazione. Per migliorare e rendere
più facili i coupling con l’Arg, sono stati progettati dei metodi che impiegano l’uso di
pseudo proline, di resine a base di PEG o delle microonde.
Formazione di dichetopiperazine
La formazione di dichetopiperazine può avvenire durante la deprotezione dello
Fmoc del secondo amminoacido sulla catena peptidica. Il gruppo amminico libero
può a questo punto staccare il legame estereo del peptide alla resina e formare il
piperizinedione: nella maggior parte dei casi questo comporta un distacco
quantitativo del dipeptide dalla resina. Questo fenomeno viene particolarmente
promosso in presenza di Gly o di Pro come primo o secondo amminoacido, a causa
dello scarso ingombro sterico nel caso della Gly e del legame cis-ammidico nel caso
della Pro. L’uso della resina 2-clorotritile è risultato essere ottimale per prevenire
tale reazione, a causa dell’ingombro sterico dell’atomo di cloro, in grado di
proteggere il legame estereo dall’attacco nucleofilo. Altrimenti occorre fare la
seconda deprotezione in tempi molto brevi (in genere due deprotezioni da 5 minuti
ciascuna).

44
3.1.2.2 Liberty BlueTM
CEM
Dai paragrafi precedenti è risultato chiaro come la SPPS convenzionale rimanga
sempre la strategia principale per la sintesi peptidica, ma anche come l’uso delle
microonde possa talvolta migliorare, in termini di resa, di purezza e di tempo, la
sintesi su fase solida convenzionale.
Ovviamente occorre ricordare che la sintesi peptidica su fase solida mediata da
microonde (MW-SPPS) non è priva di reazioni secondarie e che la maggior parte di
esse sono legate all’aumento di temperatura dei reagenti. Attualmente uno degli
strumenti più all’avanguardia nel campo della sintesi di peptidi mediata da
microonde (MW-SPPS) è il sintetizzatore di peptidi automatizzato Liberty BlueTM
CEM (Fig. 20). Costruito sulla piattaforma a microonde del Discover flessibile della
CEM Corporation, il Liberty Blue è in grado di sintetizzare peptidi su scale che vanno
da 0.05 a 5 mmol in modo più veloce ed efficace dei sintetizzatori manuali e delle
versioni automatiche senza microonde precedenti (tipo Aaptec), grazie alla cavità
circolare a microonde del sistema.
Figura 20 – Liberty BlueTM
CEM
Il Liberty Blue gestisce il trasferimento di tutti i solventi e soluzioni all’interno e
all’esterno del reattore, interfacciandosi con un sistema computerizzato in grado di
monitorare ogni step della sintesi peptidica. All’interno del reattore è presente un
termometro a fibra ottica per monitorare la temperatura di esercizio, in modo tale

45
da evitare surriscaldamenti anomali che potrebbero compromettere la sintesi
peptidica (es. reazioni secondarie).
Ogni ciclo della sintesi peptidica è caratterizzato da:
• Swelling: rigonfiamento della resina
• Deprotection Cycle: ciclo di deprotezione della resina con 20% piperidina in
DMF
• Coupling Cycle: cicli di accoppiamento diversi a seconda del tipo di
amminoacido da inserire; i reagenti attivante ed attivante basico utilizzati
sono stati comunque, rispettivamente, DIC e Oxima (Fig. 16).
Nel caso del Coupling Cycle dello strumento sono presenti programmi appositi per
l’inserimento di amminoacidi soggetti a reazioni secondarie (His e Arg): qui per
praticità ho riportato i due casi che ho applicato durante la sintesi peptidica.
Nel primo caso, quello dell’inserimento dell’His, il ciclo di coupling procede in modo
tale che nei primi due minuti venga svolto il coupling convenzionale, mentre negli
ultimi quattro minuti il coupling venga portato a termine con l’impiego delle
microonde a 50°C, limitando così la racemizzazione. In questo modo il primo step
permette ad una porzione significativa di tali amminoacidi di accoppiarsi, mentre il
secondo passaggio accelera la reazione anche nei siti più difficili rimasti.
Nel caso dell’accoppiamento dell’Arg, il metodo prevede l’applicazione delle
microonde alla fine del ciclo di coupling convenzionale, in modo tale che il derivato
attivato dell’Arg risulti protetto dall’energia delle microonde, che potrebbe
accelerare la reazione collaterale di formazione del δ-lattame dell’Arg. In questo
modo rimangono solo i siti non accoppiati più difficili da far reagire. La presenza di
un secondo ciclo di coupling infine permette che siano minimizzate le sequenze di
delezione, risultanti dalla formazione del δ-lattame.
Un ciclo completo di accoppiamento per ogni amminoacido avviene in circa 20
minuti.
Questo strumento rispetto alla prima generazione di Liberty CEM è in grado di
effettuare accoppiamenti arrivando ad una temperatura di 90°C, mentre per la
prima generazione la temperatura di accoppiamento era di 75°C. Questo ha
permesso di ridurre i tempi di accoppiamento e di deprotezione, riducendo ancora
di più i tempi totali di sintesi di un peptide.

46
Inoltre lo strumento è stato programmato per ridurre anche i volumi dei solventi
utilizzati: questo è un altro grande vantaggio visto che il maggior costo nella sintesi
peptidica sono i solventi.
3.2 SINTESI DEI PEPTIDI hGAD65 (250-273) (I),
hGAD67 (258-281) (II) e CVB4 P2C (28-50) (III)
I peptidi I,II,III sono hGAD65 (250-273) (I), hGAD67 (258-281) (II) e CVB4 P2C (28-
50) (III), in versione non acetilata nella posizione N-terminale. La sintesi delle tre
sequenze, non richiedendo alcuna modifica delle stesse o l’introduzione di building
block o amminoacidi modificati, è stata effettuata per intero attraverso lo
strumento Liberty BlueTM CEM, che sfrutta la tecnica SPPS mediata da microonde, i
cui vantaggi e caratteristiche sono già state descritte nel paragrafo 3.1.2.
3.3 SINTESI DEI PEPTIDI hGAD65 Ac-(250-273) (I),
hGAD67 Ac-(258-281) (II) e CVB4 P2C Ac-(28-50)
(III)
I peptidi IV, V, VI sono hGAD65 (250-273) (IV), hGAD67 (258-281) (V) e CVB4 P2C
(28-50) (VI), in versione acetilata nella posizione N-terminale. La sintesi delle tre
sequenze, esattamente come per le precedenti, è stata effettuata per intero
attraverso lo strumento Liberty BlueTM CEM.

47
3.3.1 Reazione di Acetilazione
Sui peptidi IV, V, VI è stata effettuata, prima del distacco dalla resina e della
deprotezione finale, una reazione di acetilazione dell’estremità N-terminale
utilizzando Ac2O.
Questa reazione è stata fatta al fine di valutare l’eventuale importanza di un
ulteriore legame ammidico in posizione N-terminale in termini di mimetismo da
parte del frammento rispetto alla proteina completa.
• Ac-hGAD65 (250-273) – Ac-250AMMIARFKMFPEVKEKGMAALPRL273 (IV)
• Ac-hGAD67 (258-281) – Ac-258SIMAARYKYFPEVKTKGMAAVPKL281 (V)
• Ac-CVB4 P2C (28-50) – Ac-28FIEWLKVKILPEVKEKHEFLSRL50 (VI)
3.4 PURIFICAZIONE E CARATTERIZZAZIONE DEI
PEPTIDI
Per poter effettuare i test ELISA sui peptidi sintetizzati I-VI, questi devono essere
caratterizzati da una purezza ≥ 95%. A questo scopo è stata necessaria una
purificazione preliminare su fase inversa, per allontanare l’elevata quantità di sali e
gli eventuali sottoprodotti formatisi durante la fase di cleavage, ed una successiva
purificazione tramite RP-HPLC semi preparativa. Per quanto riguarda la pre-
purificazione, come fase stazionaria è stata impiegata una colonna di silice
funzionalizzata con catene alchiliche idrofobiche C18 (la lunghezza delle catene
alchiliche idrofobiche impiegate dipende dalla polarità dei peptidi trattati), mentre
come fase mobile è stata usata una miscela di H2O e CH3CN in proporzioni variabili,
dipendenti dal metodo di pre-purificazione scelto. In questo lavoro di tesi i peptidi
sono stati pre-purificati tutti con lo stesso metodo, come riportato in Tabella 2.
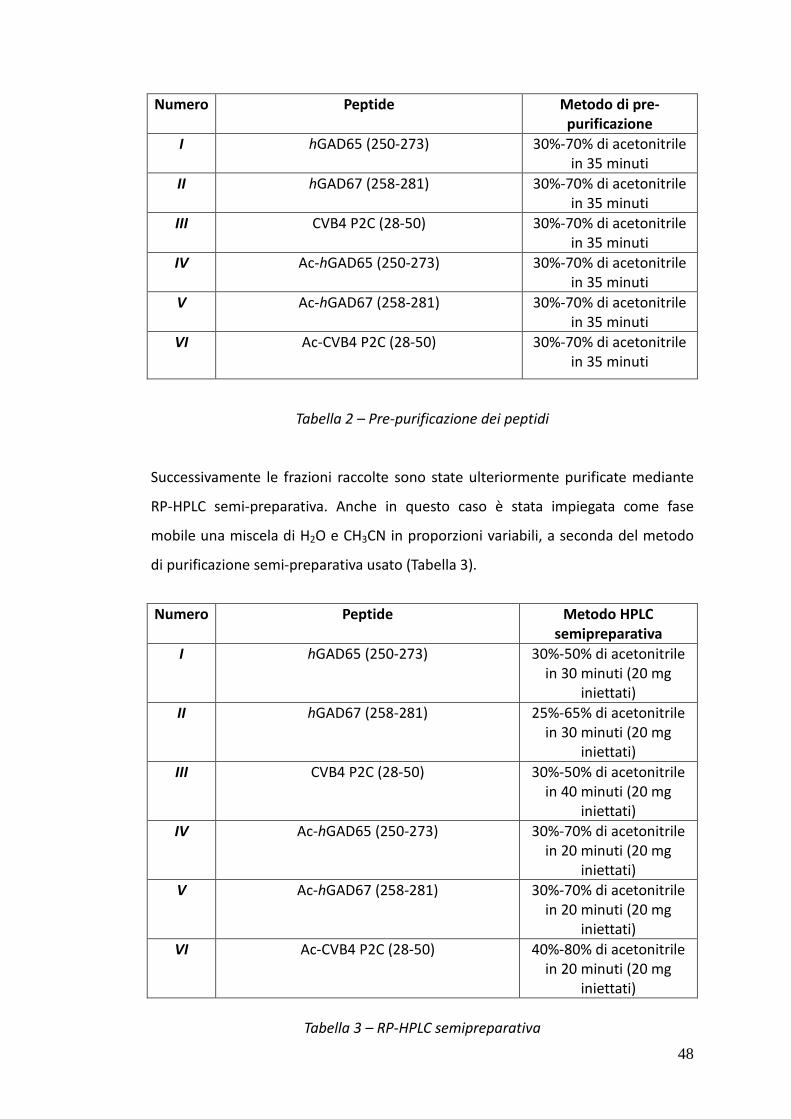
48
Numero Peptide Metodo di pre-
purificazione I hGAD65 (250-273) 30%-70% di acetonitrile
in 35 minuti II hGAD67 (258-281) 30%-70% di acetonitrile
in 35 minuti III CVB4 P2C (28-50) 30%-70% di acetonitrile
in 35 minuti IV Ac-hGAD65 (250-273) 30%-70% di acetonitrile
in 35 minuti V Ac-hGAD67 (258-281) 30%-70% di acetonitrile
in 35 minuti VI Ac-CVB4 P2C (28-50) 30%-70% di acetonitrile
in 35 minuti
Tabella 2 – Pre-purificazione dei peptidi
Successivamente le frazioni raccolte sono state ulteriormente purificate mediante
RP-HPLC semi-preparativa. Anche in questo caso è stata impiegata come fase
mobile una miscela di H2O e CH3CN in proporzioni variabili, a seconda del metodo
di purificazione semi-preparativa usato (Tabella 3).
Numero Peptide Metodo HPLC
semipreparativa I hGAD65 (250-273) 30%-50% di acetonitrile
in 30 minuti (20 mg iniettati)
II hGAD67 (258-281) 25%-65% di acetonitrile in 30 minuti (20 mg
iniettati) III CVB4 P2C (28-50) 30%-50% di acetonitrile
in 40 minuti (20 mg iniettati)
IV Ac-hGAD65 (250-273) 30%-70% di acetonitrile in 20 minuti (20 mg
iniettati) V Ac-hGAD67 (258-281) 30%-70% di acetonitrile
in 20 minuti (20 mg iniettati)
VI Ac-CVB4 P2C (28-50) 40%-80% di acetonitrile in 20 minuti (20 mg
iniettati)
Tabella 3 – RP-HPLC semipreparativa
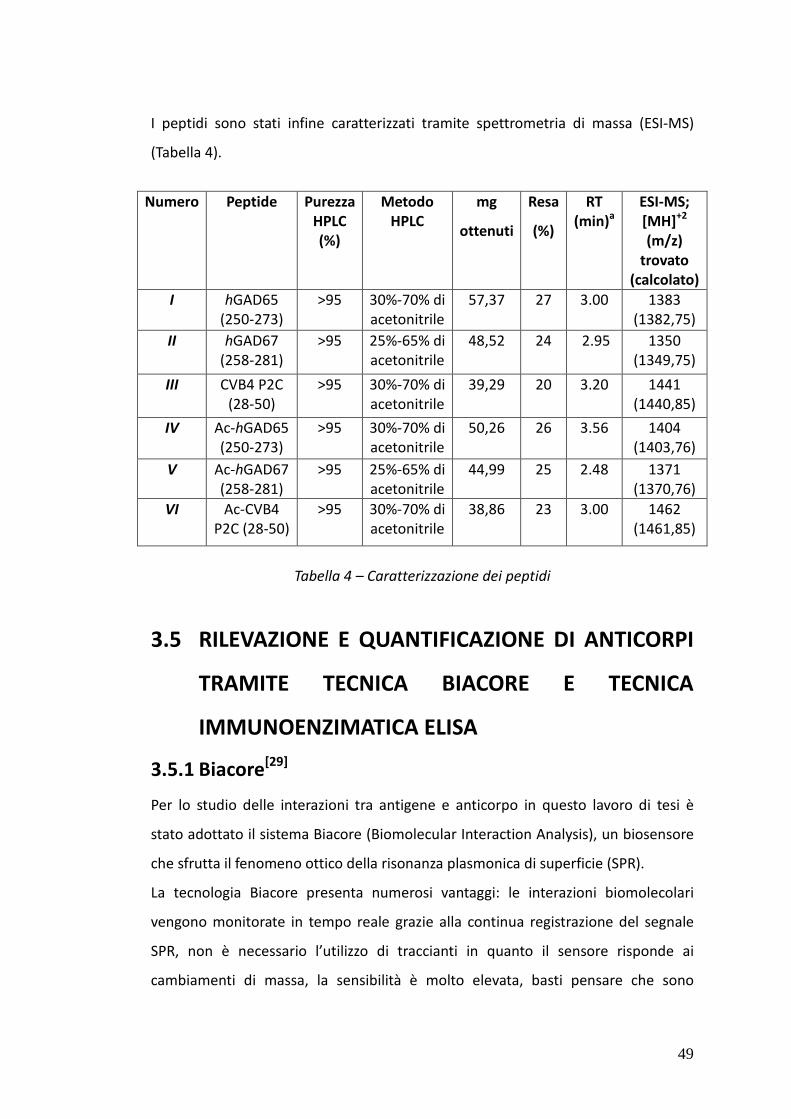
49
I peptidi sono stati infine caratterizzati tramite spettrometria di massa (ESI-MS)
(Tabella 4).
Numero Peptide Purezza
HPLC
(%)
Metodo
HPLC
mg
ottenuti
Resa
(%)
RT
(min)a
ESI-MS;
[MH]+2
(m/z)
trovato
(calcolato) I hGAD65
(250-273) >95 30%-70% di
acetonitrile 57,37 27 3.00 1383
(1382,75)
II hGAD67 (258-281)
>95 25%-65% di acetonitrile
48,52 24 2.95 1350 (1349,75)
III CVB4 P2C (28-50)
>95 30%-70% di acetonitrile
39,29 20 3.20 1441 (1440,85)
IV Ac-hGAD65 (250-273)
>95 30%-70% di acetonitrile
50,26 26 3.56 1404 (1403,76)
V Ac-hGAD67 (258-281)
>95 25%-65% di acetonitrile
44,99 25 2.48 1371 (1370,76)
VI Ac-CVB4 P2C (28-50)
>95 30%-70% di acetonitrile
38,86 23 3.00 1462 (1461,85)
Tabella 4 – Caratterizzazione dei peptidi
3.5 RILEVAZIONE E QUANTIFICAZIONE DI ANTICORPI
TRAMITE TECNICA BIACORE E TECNICA
IMMUNOENZIMATICA ELISA
3.5.1 Biacore[29]
Per lo studio delle interazioni tra antigene e anticorpo in questo lavoro di tesi è
stato adottato il sistema Biacore (Biomolecular Interaction Analysis), un biosensore
che sfrutta il fenomeno ottico della risonanza plasmonica di superficie (SPR).
La tecnologia Biacore presenta numerosi vantaggi: le interazioni biomolecolari
vengono monitorate in tempo reale grazie alla continua registrazione del segnale
SPR, non è necessario l’utilizzo di traccianti in quanto il sensore risponde ai
cambiamenti di massa, la sensibilità è molto elevata, basti pensare che sono

50
sufficienti cambiamenti di massa di pochi pg per mm2 di superficie per generare un
segnale. Le applicazioni di questa nuova tecnologia riguardano:
• la specificità dell’interazione: la semplice registrazione di una risposta SPR è
indice di una specificità tra l’analita in soluzione e il ligando immobilizzato;
• le costanti cinetiche: l’aumento del segnale che si verifica quando l’analita
passa sulla superficie riflette direttamente la velocità di formazione del
complesso, che sarà proporzionale alla concentrazione dell'analita; mentre
la diminuzione della risposta che si verifica quando il campione smette di
fluire sul chip indica la velocità di dissociazione di tale complesso. Attraverso
una serie di algoritmi di calcolo il software è in grado di fornire le rispettive
costanti di associazione e dissociazione del complesso;
• l’affinità dell’interazione: calcolata a partire dalle costanti cinetiche o in base
alla massima capacità di legame dell’analita allo stato stazionario;
• la concentrazione di molecole attive: dopo aver costruito un’apposita curva
di taratura si possono ottenere informazioni sulla concentrazione di una
specifica sostanza in una soluzione pura o in una miscela complessa.
Tutti questi studi possono essere applicati a interazioni tra molecole biologiche di
qualsiasi tipo: proteina-proteina, lipidi-proteina, acidi nucleici-proteina, o proteina
con molecole di basso peso molecolare quali farmaci, substrati e cofattori.
La strategia analitica Biacore grazie all’estrema versatilità che la caratterizza trova
applicazione in diversi settori:
• in campo biomedico può essere utilizzata per quantificare l’espressione di
specifiche proteine in diverse condizioni di salute del paziente, per
selezionare i migliori anticorpi da impiegare come strumento di ricerca o in
ambito terapeutico, per identificare e valutare l’efficacia di inibitori e
antagonisti, individuare marcatori di malattia in campioni biologici;
• nel Drug Discovery and Development per l’individuazione di candidati
farmaci, per ottimizzare il lead compound tramite studi cinetici, per definire
alcuni parametri farmacocinetici come ad esempio il legame alle proteine
plasmatiche.

51
La strategia Biacore è una tecnica ottica che usa il fenomeno dell’onda evanescente
per misurare cambiamenti dell’indice di rifrazione nella regione di spazio adiacente
alla superficie di un chip su cui è stata immobilizzata una delle due molecole
interagenti che prende il nome di ligando, mentre il campione contenente l’analita
libero in soluzione viene fatto passare continuamente sulla superficie grazie al
flusso di un tampone chiamato Runnig Buffer (Fig. 21)
Figura 21 – Ligando associato alla superficie del sensore e analita libero in soluzione
Nel presente lavoro di tesi è stato utilizzato uno strumento Biacore T 100 (Fig. 22)
interfacciato a un computer che permette di regolare lo strumento tramite il
software Biacore T 100 Control e di analizzare e rielaborare i sensogrammi tramite il
Biacore T 100 Evaluation.
Figura 22 – Biacore T100

52
Il funzionamento di uno strumento Biacore è basato su due tecnologie
fondamentali:
• il fenomeno ottico della SPR;
• le proprietà della superficie del sensor chip dove viene immobilizzato il
ligando;
3.5.1.1 Risonanza Plasmonica di Superficie
La SPR è un fenomeno ottico che si verifica all’interfaccia tra due mezzi con diverso
indice di rifrazione, quando la luce polarizzata linearmente colpisce un film
metallico in condizioni di riflessione interna totale (TIR). Una radiazione
elettromagnetica nel passaggio da un mezzo più denso a uno meno denso (con
diverso indice di rifrazione) in parte viene riflessa e in parte rifratta proseguendo il
suo cammino al di là della superficie di separazione dopo che ha subito una
deviazione. All’interfaccia tra i due mezzi, al di sopra di un angolo critico di
incidenza (proporzionale al rapporto tra gli indici di rifrazione), non ci sarà rifrazione
ma si verificherà la TIR. In questa condizione la luce incidente viene completamente
riflessa e una componente del campo elettromagnetico penetra a breve distanza
nel mezzo con indice di rifrazione più basso; questa componente è chiamata onda
evanescente, la sua ampiezza decresce esponenzialmente allontanandosi dalla
superficie e decade a una distanza di circa una lunghezza d’onda dall’interfaccia
(Fig. 23).
Figura 23 – Riflessione Interna Totale (TIR)

53
Se l’interfaccia tra i due mezzi è ricoperta da un sottile strato di metallo e la luce è
monocromatica e polarizzata linearmente, per uno specifico angolo di incidenza si
verificherà l’eccitazione dei plasmoni nel film metallico che comporterà un
caratteristico assorbimento di energia attraverso l’onda evanescente e il segnale
SPR sarà visto come una caduta dell’intensità della luce riflessa (Fig. 24).
Figura 24 – Risonanza Plasmonica di Superficie (SPR)
Questo fenomeno ottico è altamente sensibile all’indice di rifrazione della soluzione
presente all’interfaccia della superficie del biosensore. Come mostrato nella figura
6, inizialmente la diminuzione dell’intensità riflessa si registrerà a un certo angolo di
risonanza (angolo I), quando l’analita in soluzione andrà a legare la molecola
immobilizzata si verificherà un cambiamento di massa e quindi di indice di
rifrazione, che provocherà un cambiamento dell’angolo di risonanza (angolo II); nel
sensogramma è riportata la variazione di tale angolo di risonanza in funzione del
tempo di analisi.
L’angolo SPR è misurato in Unità di Risonanza (RU) e il suo cambiamento è
direttamente collegato alla concentrazione di biomolecole sulla superficie del chip:
in particolare una variazione dell’angolo SPR di 0.1° determina una risposta
strumentale pari a 1000 RU che corrispondono a una variazione di massa sulla
superficie del chip di 1 ng/mm2.
Le molecole rilevabili con questa tecnica devono quindi avere un peso molecolare
superiore a 100 Da in modo da indurre un sufficiente cambiamento dell’indice di

54
rifrazione; considerando inoltre che la profondità di penetrazione dell’onda
evanescente è di 300-400 nm, si spiega perché particelle di dimensioni superiori a
400 nm non possano essere misurate completamente in quanto il segnale che
generano non è lineare e quindi non può essere eseguita un’analisi cinetica o
quantitativa, ma è comunque possibile uno studio di tipo qualitativo.
Nel sistema Biacore la lunghezza d’onda e l’indice di rifrazione del mezzo più denso
sono tenute costanti e il fenomeno della SPR è usato per monitorare l’indice di
rifrazione nello strato acquoso immediatamente adiacente alla superficie d’oro del
chip. Oltre al biosensore sono quindi necessari una sorgente di luce polarizzata, un
detector bidimensionale e un sistema di flow cell che veicoli il mezzo in cui è
contenuto l’analita. I segnali misurati dal detector verranno poi processati da
algoritmi di interpolazione che calcolano l’angolo SPR con elevato grado di
accuratezza.
3.5.1.2 La superficie del biosensore
Il sensor chip è la chiave della tecnologia Biacore, indispensabile perché si verifichi
un segnale SPR, tutte le interazioni monitorate dipendono dalla capacità di
immobilizzare il ligando sulla superficie del chip. Le caratteristiche fondamentali di
un sensor chip sono due:
• una superficie di vetro ricoperta da un sottile film d’oro (50 nm) che fornisce
le condizioni fisiche richieste per generare un segnale SPR e che è quindi
una caratteristica comune a tutti i tipi di chip. Queste condizioni fisiche in
temini di angolo di risonanza e lunghezza d’onda della radiazione incidente
variano a seconda dello spessore e della natura del materiale conduttore
utilizzato e l’oro è l’elemento ideale in quanto consente di utilizzare
radiazioni vicine all’IR facilmente riproducibili ed è inerte nelle condizioni
fisiologiche dei tamponi utilizzati per le analisi;
• un rivestimento di qualsiasi genere al di sopra dello strato d’oro che varia a
seconda dei diversi tipi di chip e che fornisce il mezzo per ancorare il ligando
e rappresenta l’ambiente in cui avvengono le interazioni molecolari. Questo
rivestimento non interferisce col segnale SPR.

55
Lo strato d’oro e il rivestimento sono stabili a una varietà di condizioni che
includono valori di pH estremi e medie concentrazioni di solventi organici. Una volta
che il ligando è stato immobilizzato, la resistenza della superficie del biosensore
sarà determinata dalle proprietà di tale ligando; il rivestimento è fondamentale
perché un adsorbimento diretto del ligando su una superficie d’oro avrebbe un
effetto denaturante e non creerebbe un ambiente favorevole alle interazioni
biomolecolari.
In commercio sono disponbili sensor chip caratterizzati da matrici di rivestimento di
varia natura che permettono l’immobilizzazione di una vasta gamma di composti.
Ogni sensor chip è inserito in un supporto di plastica per renderlo più maneggevole
dall’operatore (Fig. 25).
Figura 25 – Sensor Chip
Il rivestimento più comune dei sensor chip è costituito da una matrice di destrano
carbossimetilato, si tratta di un polissacaride ramificato estremamente flessibile
che, nel caso del chip della serie CM5, utilizzato in questo lavoro di tesi, ha uno
spessore di 100 nm e consente l’immobilizzazione di diversi tipi di ligandi, sia
piccole molecole organiche che macromolecole come proteine, acidi nucleici e
carboidrati. La matrice di carbossidestrano presenta numerosi vantaggi: costituisce
un ambiente idrofilo favorevole alla maggior parte delle interazioni in soluzione,
evita la denaturazione delle molecole, può essere funzionalizzata in modo da
possedere le strutture chimiche necessarie per un’immobilizzazione covalente del
ligando, instaura interazioni elettrostatiche con molecole cariche positivamente
facilitando quindi il processo di immobilizzazione, è inerte e la flessibilità delle sue
catene permette all’analita in soluzione di muoversi liberamente (Fig. 26).

56
Figura 26 – Rappresentazione schematica di un chip della serie CM5
3.5.1.3 Procedura di pH-scouting
Al fine di ottenere un sufficiente livello di immobilizzazione di ligando sulla
superficie del chip, è indispensabile ottimizzare ogni fase del processo e fare in
modo che la maggior quantità possibile di ligando iniettato si avvicini alla matrice di
destrano. Questa situazione ideale può essere raggiunta grazie a una corretta
preconcentrazione elettrostatica del ligando che viene valutata con la procedura del
pH scouting.
Lo scopo del pH scouting è quindi quello di individuare le migliori condizioni in
termini di pH e di forza ionica che massimizzino l’interazione elettrostatica tra
ligando e chip; a questo punto sarà possibile attivare la matrice del chip e
immobilizzare covalentemente il ligando.
La matrice di destrano carbossimetilato dei chip CM5 ha una pKa di 3.5, quindi per
valori di pH superiori a 3.5 i gruppi carbossilici presentano cariche negative e sono
in grado di interagire elettrostaticamente con molecole cariche positivamente come
peptidi o proteine. Il requisito fondamentale per ottenere questa
preconcentrazione elettrostatica è che il pH della soluzione in cui è presente il
ligando sia compreso tra 3.5 e il Punto Isolelettrico del peptide in modo che ligando
e superficie del chip abbiano cariche nette opposte. A pH superiore a 3.5 si perdono
infatti le cariche negative della matrice per protonazione dei suoi gruppi carbossilici
mentre a un pH superiore al punto isoelettrico il peptide assume carica negativa e
non può interagire col chip (Fig. 27).

57
Figura 27 - B) Intervallo di pH ideale per favorire la preconcentrazione elettrostatica
del ligando sul chip; A) e C) Valori di pH rispettivamente troppo bassi o troppo alti
non consentono l’interazione elettrostatica tra le due entità.
Generalmente le interazioni in questione sono favorite da un basso valore di forza
ionica del tampone in cui è solubilizzato il ligando, quindi per l’immobilizzazione di
peptidi e proteine è comune utilizzare tamponi 10 mM di acetato di sodio a un pH
compreso tra 4 e 5.5; comunque le migliori condizioni devono essere identificate
volta per volta.
In una tipica analisi di pH scouting, per individuare il miglior tampone di
immobilizzazione senza alterare la superficie del biosensore, si procede con
l’iniezione del ligando sul chip non preventivamente attivato e alla fine di ogni
iniezione si effettuano brevi rigenerazioni con una soluzione di idrossido di sodio
0,1 M per eliminare il ligando residuo che ha interagito elettrostaticamente col chip.
La preconcentrazione elettrostatica del ligando sul chip sarà indicata da un aumento
del segnale SPR proporzionale all’entità dell’interazione stabilita e tale segnale
decadrà subito dopo la fine dell’iniezione (Fig. 28).
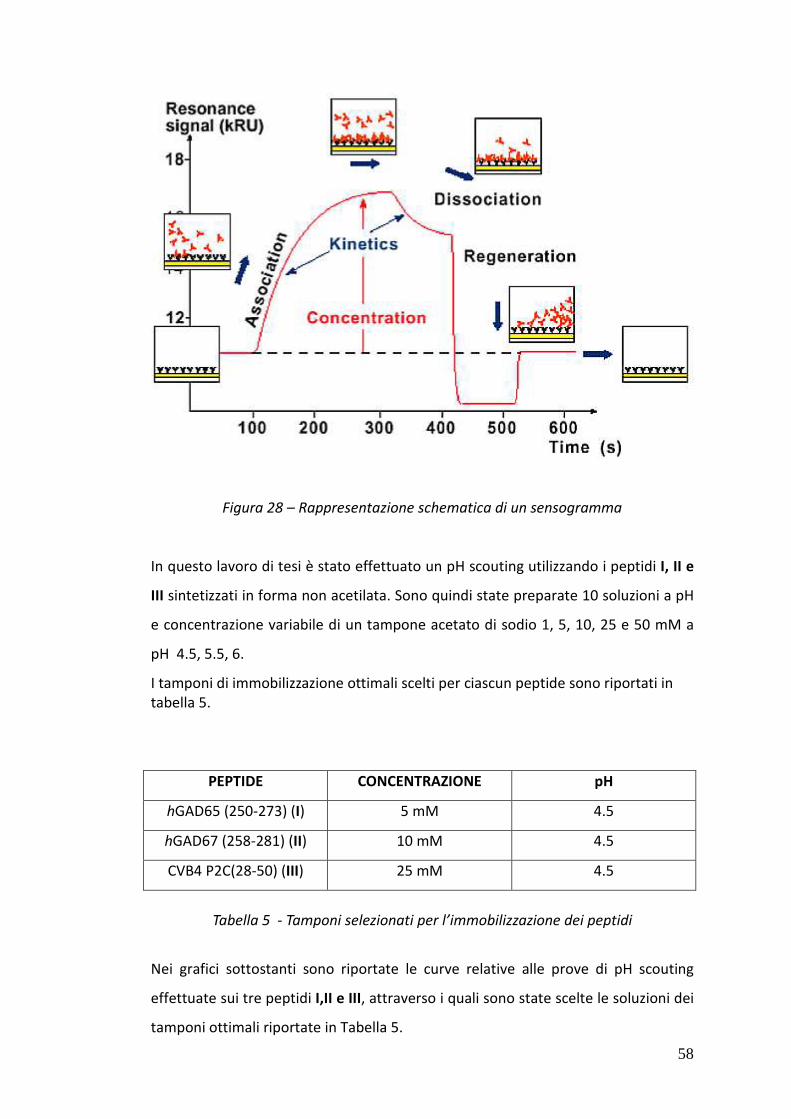
58
Figura 28 – Rappresentazione schematica di un sensogramma
In questo lavoro di tesi è stato effettuato un pH scouting utilizzando i peptidi I, II e
III sintetizzati in forma non acetilata. Sono quindi state preparate 10 soluzioni a pH
e concentrazione variabile di un tampone acetato di sodio 1, 5, 10, 25 e 50 mM a
pH 4.5, 5.5, 6.
I tamponi di immobilizzazione ottimali scelti per ciascun peptide sono riportati in tabella 5.
PEPTIDE CONCENTRAZIONE pH
hGAD65 (250-273) (I) 5 mM 4.5
hGAD67 (258-281) (II) 10 mM 4.5
CVB4 P2C(28-50) (III) 25 mM 4.5
Tabella 5 - Tamponi selezionati per l’immobilizzazione dei peptidi
Nei grafici sottostanti sono riportate le curve relative alle prove di pH scouting
effettuate sui tre peptidi I,II e III, attraverso i quali sono state scelte le soluzioni dei
tamponi ottimali riportate in Tabella 5.
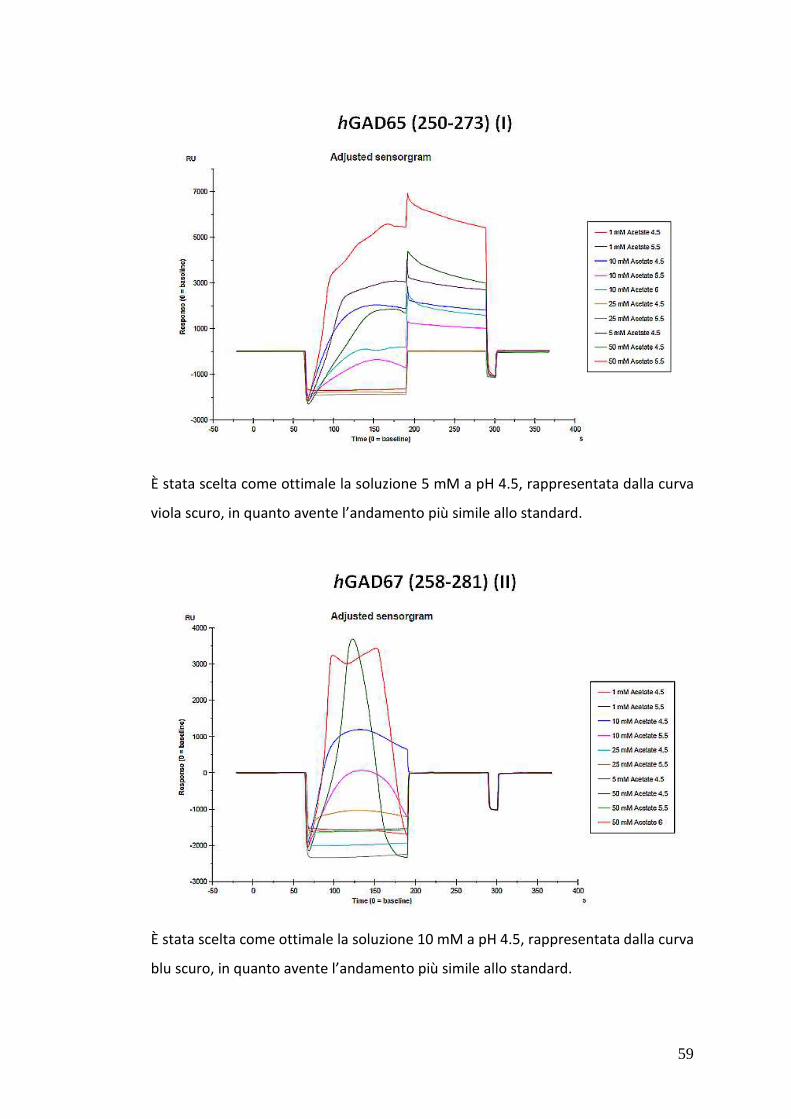
59
È stata scelta come ottimale la soluzione 5 mM a pH 4.5, rappresentata dalla curva
viola scuro, in quanto avente l’andamento più simile allo standard.
È stata scelta come ottimale la soluzione 10 mM a pH 4.5, rappresentata dalla curva
blu scuro, in quanto avente l’andamento più simile allo standard.
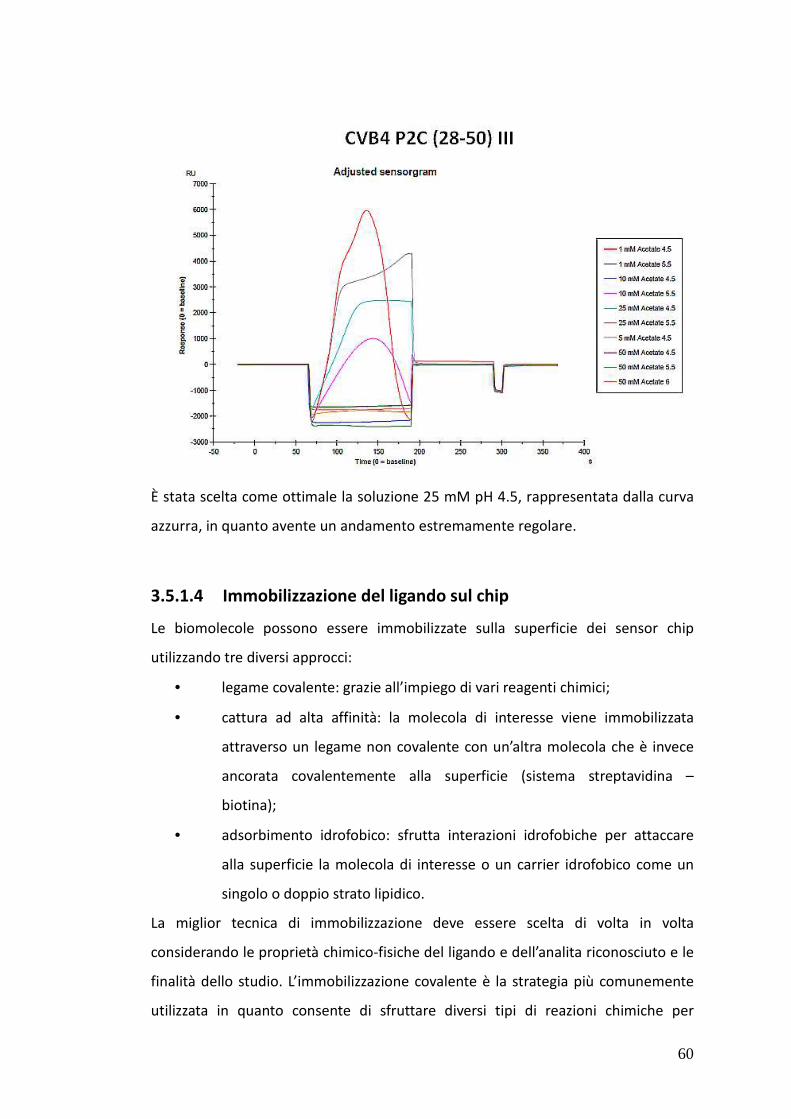
60
È stata scelta come ottimale la soluzione 25 mM pH 4.5, rappresentata dalla curva
azzurra, in quanto avente un andamento estremamente regolare.
3.5.1.4 Immobilizzazione del ligando sul chip
Le biomolecole possono essere immobilizzate sulla superficie dei sensor chip
utilizzando tre diversi approcci:
• legame covalente: grazie all’impiego di vari reagenti chimici;
• cattura ad alta affinità: la molecola di interesse viene immobilizzata
attraverso un legame non covalente con un’altra molecola che è invece
ancorata covalentemente alla superficie (sistema streptavidina –
biotina);
• adsorbimento idrofobico: sfrutta interazioni idrofobiche per attaccare
alla superficie la molecola di interesse o un carrier idrofobico come un
singolo o doppio strato lipidico.
La miglior tecnica di immobilizzazione deve essere scelta di volta in volta
considerando le proprietà chimico-fisiche del ligando e dell’analita riconosciuto e le
finalità dello studio. L’immobilizzazione covalente è la strategia più comunemente
utilizzata in quanto consente di sfruttare diversi tipi di reazioni chimiche per

61
ottenere un legame covalente tra i gruppi carbossilici del chip e i gruppi reattivi
presenti sulla molecola. Il legame che si forma tra chip e ligando con questo
approccio è sufficientemente stabile da rimanere inalterato durante i normali
processi di rigenerazione a cui è sottoposta la superficie per rimuovere l’analita che
ha interagito e rendere quindi disponibile il chip a un nuovo ciclo di analisi. I
principali svantaggi di questo approccio riguardano le modifiche chimiche e quindi
strutturali che vengono introdotte sul ligando e l’orientazione finale di quest’ultimo
che in alcuni casi può risultare non prevedibile: tutto ciò potrebbe limitare la
capacità di legame alla molecola target.
Qualsiasi metodo di immobilizzazione covalente prevede:
1. l’individuazione delle condizioni di pH e di concentrazione ionica più
adeguate applicando la metodologia del pH scouting;
2. l’attivazione della superficie del sensor chip tramite l’iniezione di appositi
reagenti (solitamente EDC/NHS);
3. l’iniezione della soluzione di ligando;
4. l’iniezione di reagenti che disattivano i rimanenti gruppi reattivi sulla
superficie e rimuovono il ligando non legato covalentemente.
Le modalità di accoppiamento covalente più comuni sono:
• Accoppiamento con la funzione amminica: porta alla formazione di un
legame amidico tra una funzione amminica primaria del ligando e un
carbossile della matrice di destrano preventivamente attivato;
• Accoppiamento con la funzione tiolica: sfrutta lo scambio tra gruppi tiolici e
ponti disolfuro attivi introdotti sul ligando o sulla matrice;
• Accoppiamento con la funzione aldeidica: si basa sulla reazione tra un
gruppo aldeidico del ligando e la superficie del chip opportunamente
attivata (Fig. 29).
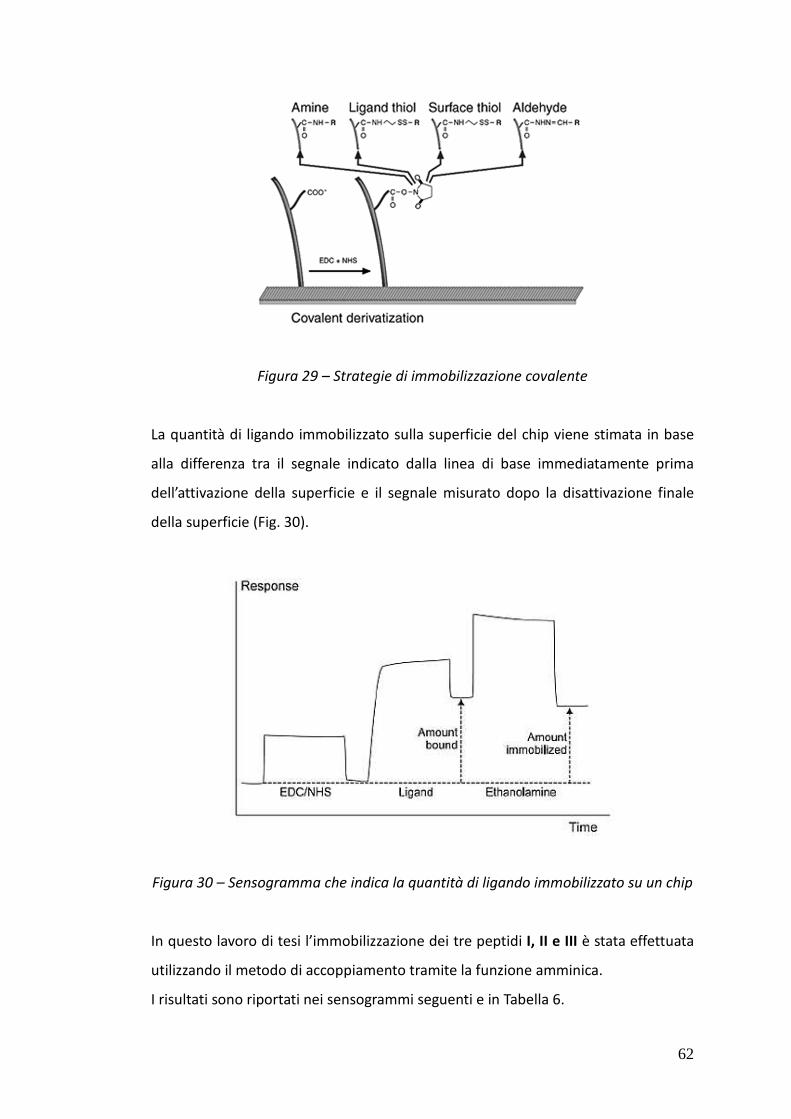
62
Figura 29 – Strategie di immobilizzazione covalente
La quantità di ligando immobilizzato sulla superficie del chip viene stimata in base
alla differenza tra il segnale indicato dalla linea di base immediatamente prima
dell’attivazione della superficie e il segnale misurato dopo la disattivazione finale
della superficie (Fig. 30).
Figura 30 – Sensogramma che indica la quantità di ligando immobilizzato su un chip
In questo lavoro di tesi l’immobilizzazione dei tre peptidi I, II e III è stata effettuata
utilizzando il metodo di accoppiamento tramite la funzione amminica.
I risultati sono riportati nei sensogrammi seguenti e in Tabella 6.
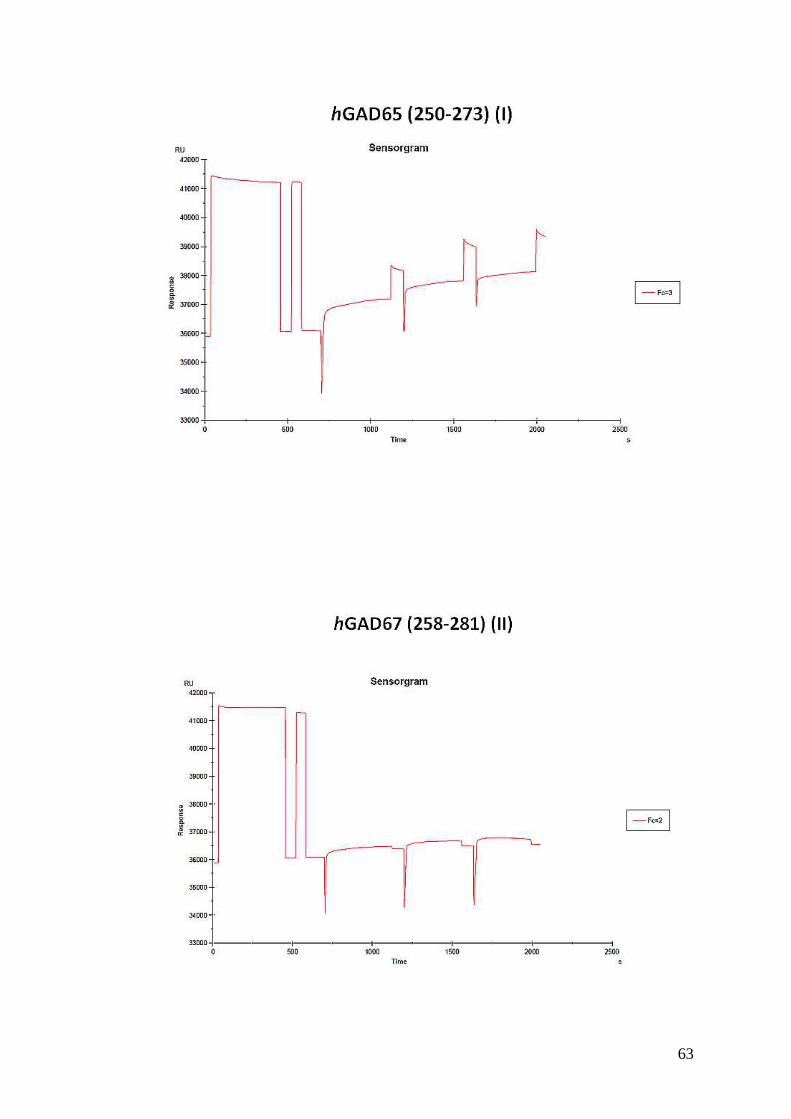
63
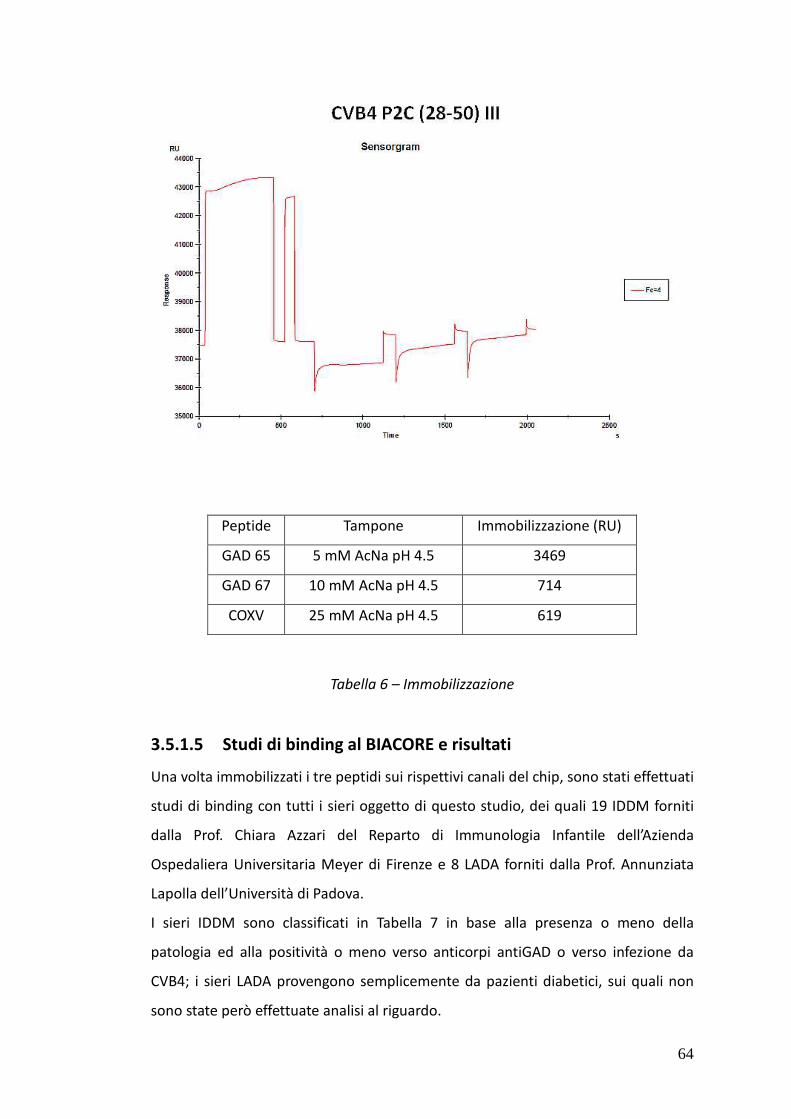
64
Peptide Tampone Immobilizzazione (RU)
GAD 65 5 mM AcNa pH 4.5 3469
GAD 67 10 mM AcNa pH 4.5 714
COXV 25 mM AcNa pH 4.5 619
Tabella 6 – Immobilizzazione
3.5.1.5 Studi di binding al BIACORE e risultati
Una volta immobilizzati i tre peptidi sui rispettivi canali del chip, sono stati effettuati
studi di binding con tutti i sieri oggetto di questo studio, dei quali 19 IDDM forniti
dalla Prof. Chiara Azzari del Reparto di Immunologia Infantile dell’Azienda
Ospedaliera Universitaria Meyer di Firenze e 8 LADA forniti dalla Prof. Annunziata
Lapolla dell’Università di Padova.
I sieri IDDM sono classificati in Tabella 7 in base alla presenza o meno della
patologia ed alla positività o meno verso anticorpi antiGAD o verso infezione da
CVB4; i sieri LADA provengono semplicemente da pazienti diabetici, sui quali non
sono state però effettuate analisi al riguardo.
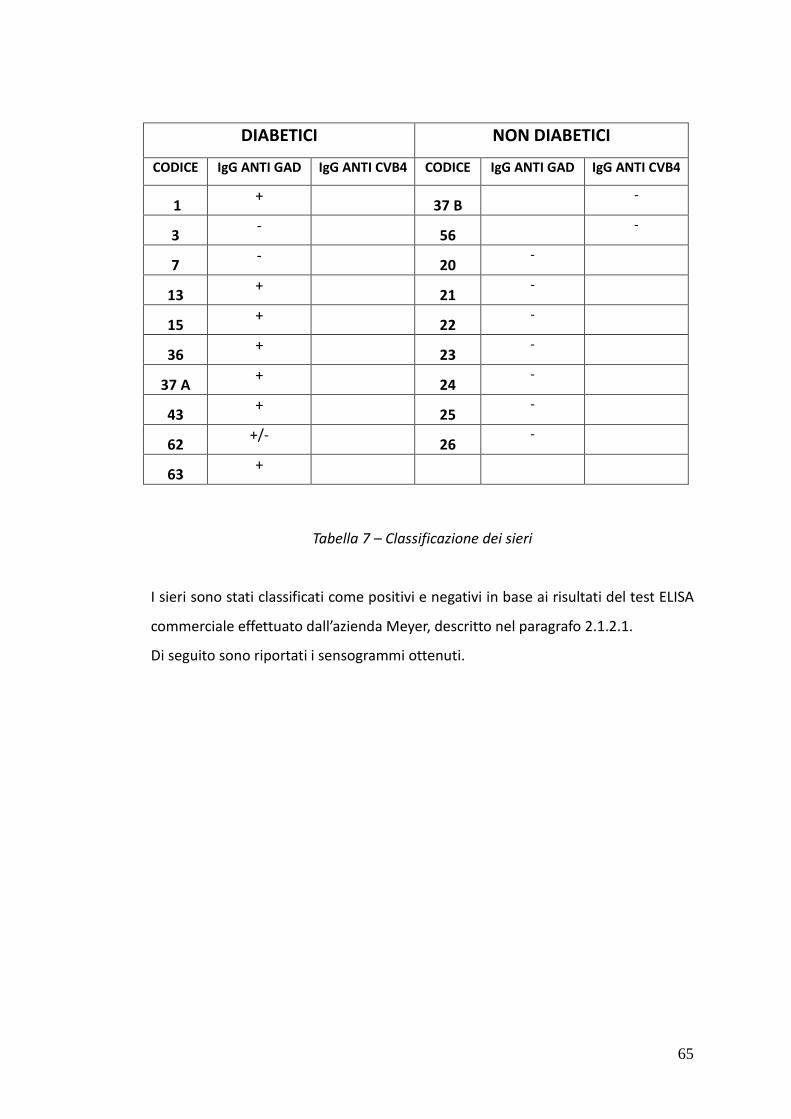
65
DIABETICI NON DIABETICI
CODICE IgG ANTI GAD IgG ANTI CVB4 CODICE IgG ANTI GAD IgG ANTI CVB4
1 +
37 B -
3 -
56 -
7 -
20 -
13 +
21 -
15 +
22 -
36 +
23 -
37 A +
24 -
43 +
25 -
62 +/-
26 -
63 +
Tabella 7 – Classificazione dei sieri
I sieri sono stati classificati come positivi e negativi in base ai risultati del test ELISA
commerciale effettuato dall’azienda Meyer, descritto nel paragrafo 2.1.2.1.
Di seguito sono riportati i sensogrammi ottenuti.
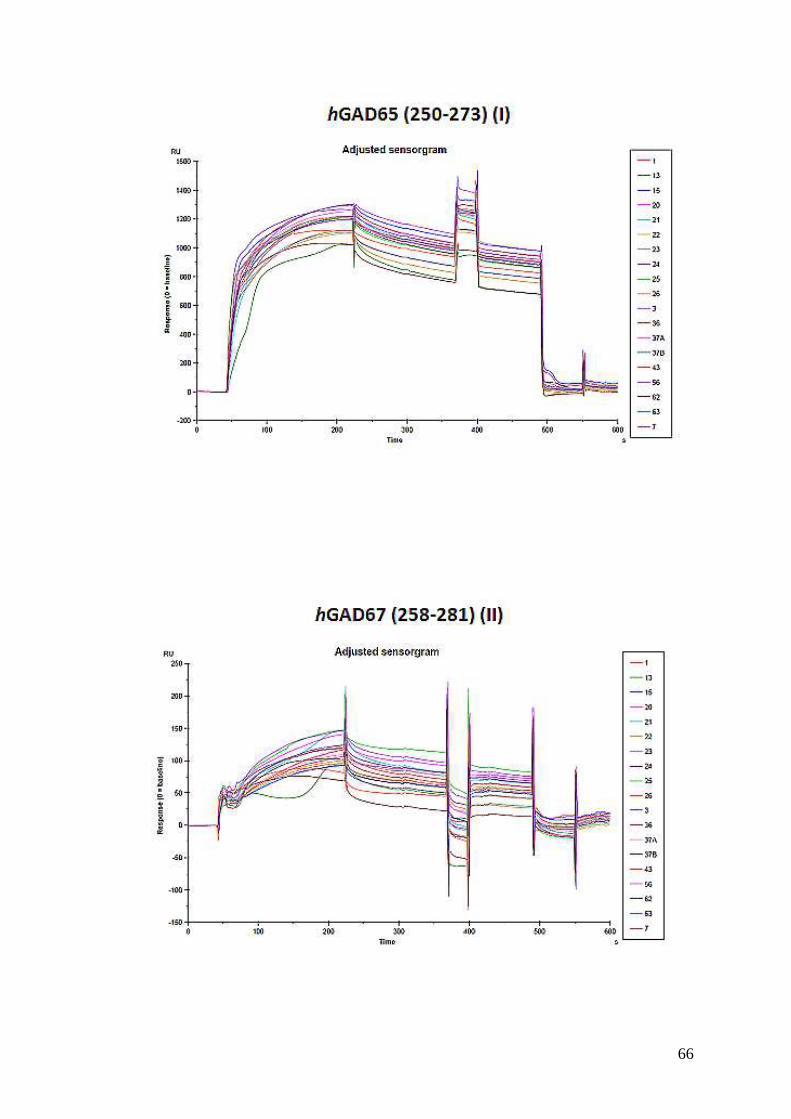
66
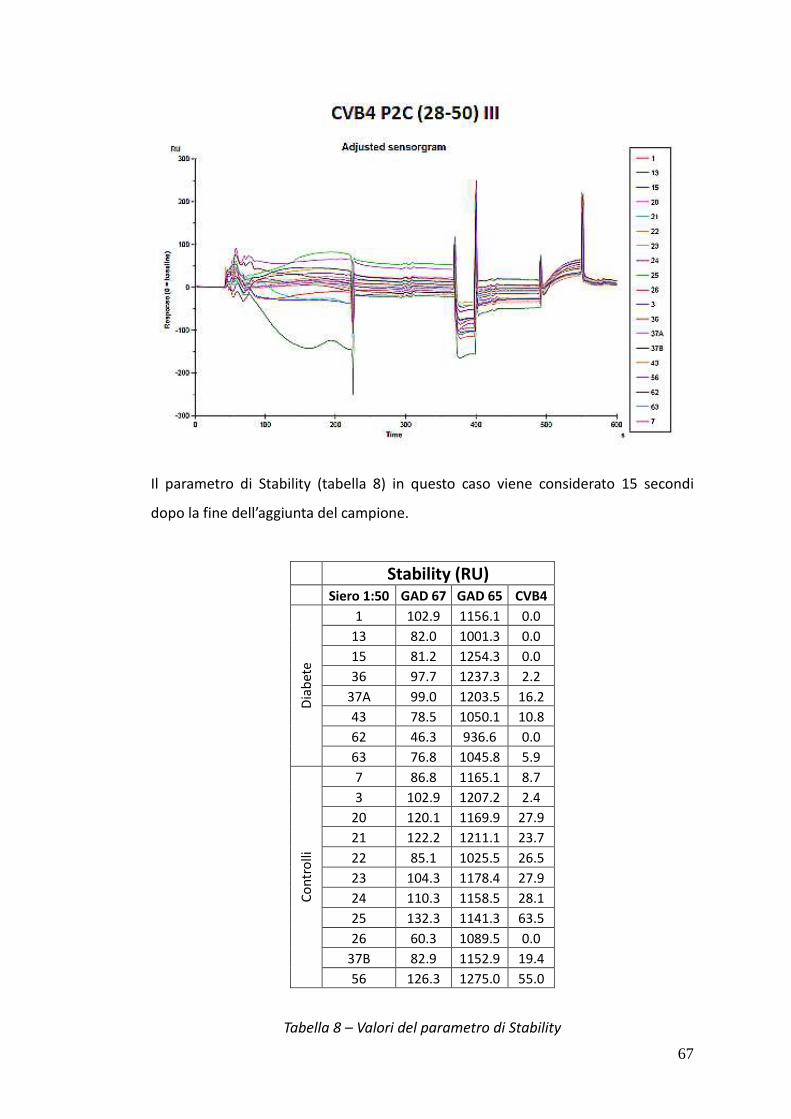
67
Il parametro di Stability (tabella 8) in questo caso viene considerato 15 secondi
dopo la fine dell’aggiunta del campione.
Stability (RU)
Siero 1:50 GAD 67 GAD 65 CVB4
Dia
bet
e
1 102.9 1156.1 0.0
13 82.0 1001.3 0.0
15 81.2 1254.3 0.0
36 97.7 1237.3 2.2
37A 99.0 1203.5 16.2
43 78.5 1050.1 10.8
62 46.3 936.6 0.0
63 76.8 1045.8 5.9
Co
ntro
lli
7 86.8 1165.1 8.7
3 102.9 1207.2 2.4
20 120.1 1169.9 27.9
21 122.2 1211.1 23.7
22 85.1 1025.5 26.5
23 104.3 1178.4 27.9
24 110.3 1158.5 28.1
25 132.3 1141.3 63.5
26 60.3 1089.5 0.0
37B 82.9 1152.9 19.4
56 126.3 1275.0 55.0
Tabella 8 – Valori del parametro di Stability
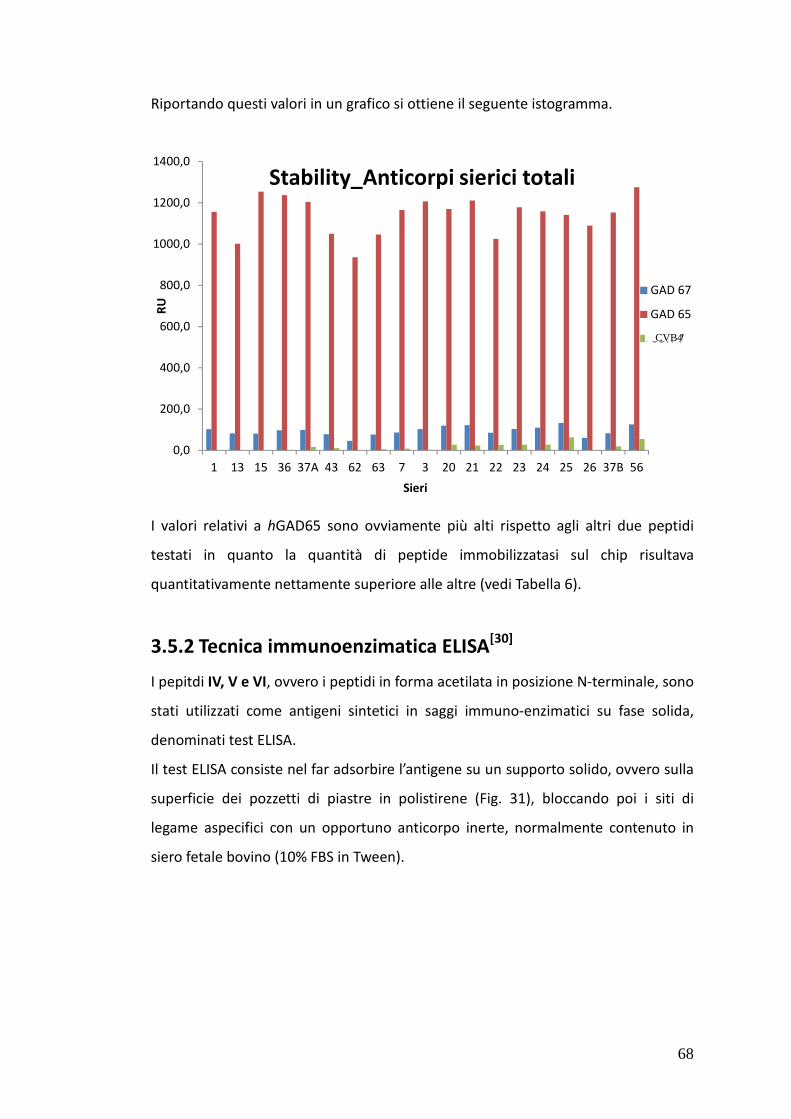
68
Riportando questi valori in un grafico si ottiene il seguente istogramma.
I valori relativi a hGAD65 sono ovviamente più alti rispetto agli altri due peptidi
testati in quanto la quantità di peptide immobilizzatasi sul chip risultava
quantitativamente nettamente superiore alle altre (vedi Tabella 6).
3.5.2 Tecnica immunoenzimatica ELISA[30]
I pepitdi IV, V e VI, ovvero i peptidi in forma acetilata in posizione N-terminale, sono
stati utilizzati come antigeni sintetici in saggi immuno-enzimatici su fase solida,
denominati test ELISA.
Il test ELISA consiste nel far adsorbire l’antigene su un supporto solido, ovvero sulla
superficie dei pozzetti di piastre in polistirene (Fig. 31), bloccando poi i siti di
legame aspecifici con un opportuno anticorpo inerte, normalmente contenuto in
siero fetale bovino (10% FBS in Tween).
0,0
200,0
400,0
600,0
800,0
1000,0
1200,0
1400,0
1 13 15 36 37A 43 62 63 7 3 20 21 22 23 24 25 26 37B 56
RU
Stability_Anticorpi sierici totali
GAD 67
GAD 65
COX V
Sieri
CVB4

69
Figura 31 – Piastra ELISA
Successivamente nei pozzetti sono stati aggiunti i sieri contenenti gli anticorpi,
ovvero le immunoglobuline IgG e IgM. Ogni siero è stato aggiunto in triplicato sia
sulla piastra relativa al test per le IgM che su quella per le IgG, in modo da poter
esprimere il risultato come media dei tre valori di assorbanza ottenuti, ed entrambi
i test sono stati effettuati due volte. I sieri trattati nei test, fornitici dalla Prof. Chiara
Azzari e dalla Prof. Annunziata Lapolla, appartengono a pazienti affetti da diabete I
(IDDM nel primo caso e LADA nel secondo) e/o da infezione da Coxsackie B4 e a
donatori sani. Le immunoglobuline se presenti dovranno legarsi quindi ai
determinanti antigenici specifici, che nel nostro caso sono i peptidi sintetizzati. Il
legame antigene-anticorpo è stato successivamente rivelato aggiungendo ai
pozzetti un anti-siero contenente anti-immunoglobuline, marcate con un enzima
(nel caso specifico una fosfatasi alcalina), capaci di legarsi alle immunoglobuline già
impegnate nel complesso con l’antigene. La struttura di un anticorpo infatti è
costituita da quattro catene polipeptidiche (due sequenze costituite da circa 450
amminoacidi cosiddette “catene pesanti” e due di circa 200 amminoacidi “catene
leggere” variabili in ogni anticorpo), unite da legami non covalenti e ponti disolfuro.
La regione N-terminale (Fab) riconosce l’epitopo dell’antigene legandosi in modo
specifico, mentre la porzione C-terminale (Fc) è la responsabile dell’interazione con
l’anti-immunoglobulina marcata con la fosfatasi alcalina, accennata prima.
Successivamente è stato aggiunto un adeguato substrato, il p-NPP (para-

70
nitrofenilfosfato), capace di legarsi all’anticorpo mediante interazioni idrofobiche e
di essere trasformato dall’enzima dando luogo ad una reazione colorimetrica (Fig.
32). Ovviamente non deve essere presente nei fluidi biologici utilizzati nel test, per
evitare possibili interferenze.
Figura 32 – Reazione colorimetrica per la rivelazione del test ELISA
Il complesso finale è stato quindi determinato quantitativamente, tramite l’uso di
uno spettrofotometro per la misurazione dell’assorbanza a determinate lunghezze
d’onda: in questo caso è stata effettuata la lettura a 405 nm con lo strumento
Tecan-Sunrise (Fig. 33).
Figura 33 – Tecan Sunrise
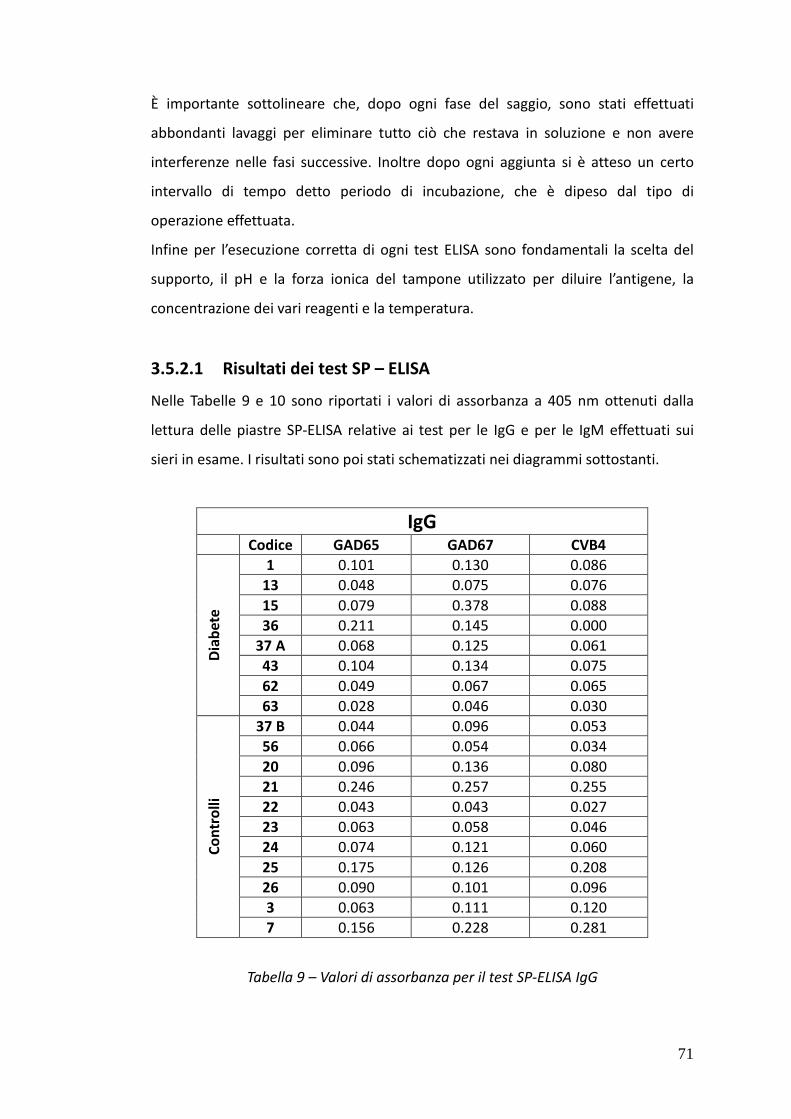
71
È importante sottolineare che, dopo ogni fase del saggio, sono stati effettuati
abbondanti lavaggi per eliminare tutto ciò che restava in soluzione e non avere
interferenze nelle fasi successive. Inoltre dopo ogni aggiunta si è atteso un certo
intervallo di tempo detto periodo di incubazione, che è dipeso dal tipo di
operazione effettuata.
Infine per l’esecuzione corretta di ogni test ELISA sono fondamentali la scelta del
supporto, il pH e la forza ionica del tampone utilizzato per diluire l’antigene, la
concentrazione dei vari reagenti e la temperatura.
3.5.2.1 Risultati dei test SP – ELISA
Nelle Tabelle 9 e 10 sono riportati i valori di assorbanza a 405 nm ottenuti dalla
lettura delle piastre SP-ELISA relative ai test per le IgG e per le IgM effettuati sui
sieri in esame. I risultati sono poi stati schematizzati nei diagrammi sottostanti.
IgG
Codice GAD65 GAD67 CVB4
Dia
be
te
1 0.101 0.130 0.086 13 0.048 0.075 0.076 15 0.079 0.378 0.088 36 0.211 0.145 0.000
37 A 0.068 0.125 0.061 43 0.104 0.134 0.075 62 0.049 0.067 0.065 63 0.028 0.046 0.030
Co
ntr
oll
i
37 B 0.044 0.096 0.053 56 0.066 0.054 0.034 20 0.096 0.136 0.080 21 0.246 0.257 0.255 22 0.043 0.043 0.027 23 0.063 0.058 0.046 24 0.074 0.121 0.060 25 0.175 0.126 0.208 26 0.090 0.101 0.096 3 0.063 0.111 0.120 7 0.156 0.228 0.281
Tabella 9 – Valori di assorbanza per il test SP-ELISA IgG
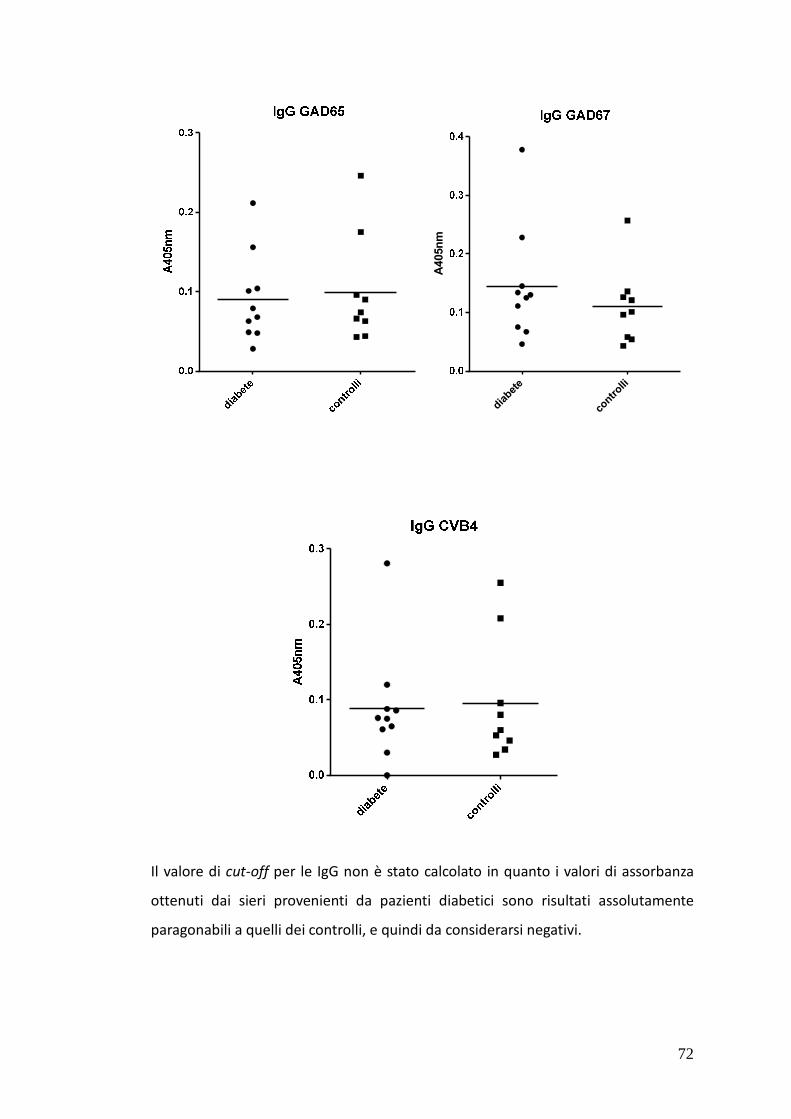
72
A405nm
diabete
controlli
Il valore di cut-off per le IgG non è stato calcolato in quanto i valori di assorbanza
ottenuti dai sieri provenienti da pazienti diabetici sono risultati assolutamente
paragonabili a quelli dei controlli, e quindi da considerarsi negativi.
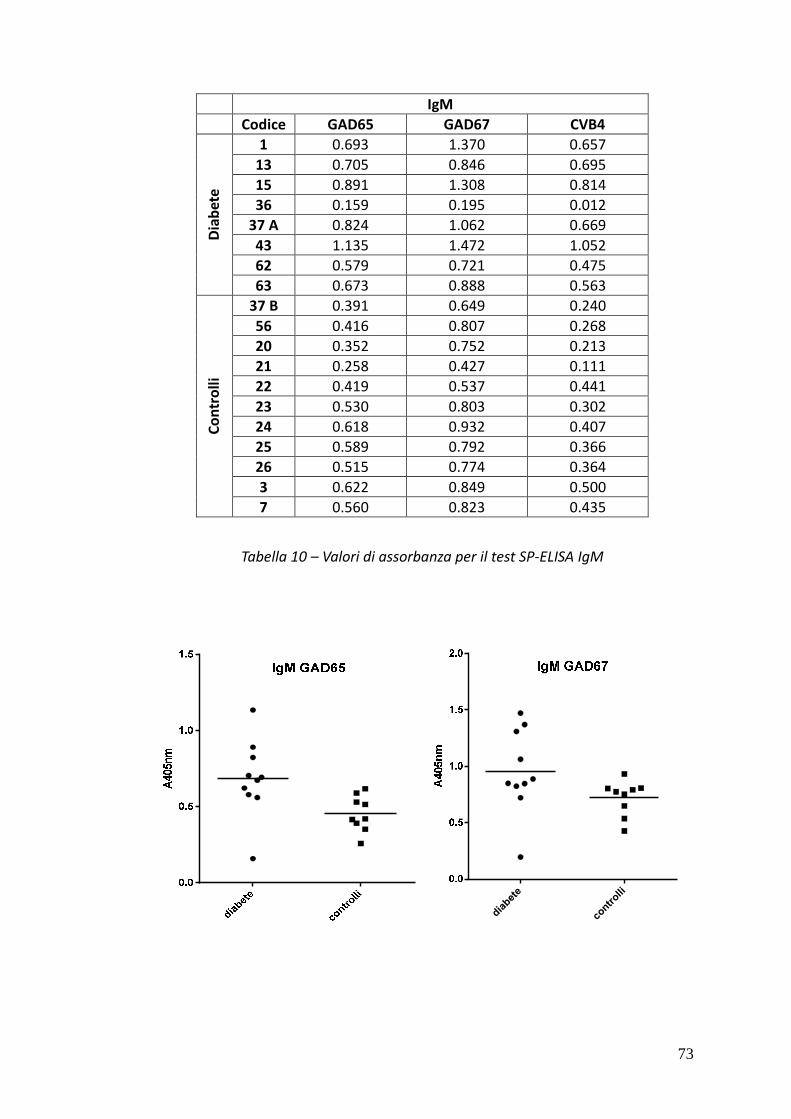
73
IgM
Codice GAD65 GAD67 CVB4
Dia
be
te
1 0.693 1.370 0.657 13 0.705 0.846 0.695 15 0.891 1.308 0.814 36 0.159 0.195 0.012
37 A 0.824 1.062 0.669 43 1.135 1.472 1.052 62 0.579 0.721 0.475 63 0.673 0.888 0.563
Co
ntr
oll
i
37 B 0.391 0.649 0.240 56 0.416 0.807 0.268 20 0.352 0.752 0.213 21 0.258 0.427 0.111 22 0.419 0.537 0.441 23 0.530 0.803 0.302 24 0.618 0.932 0.407 25 0.589 0.792 0.366 26 0.515 0.774 0.364 3 0.622 0.849 0.500 7 0.560 0.823 0.435
Tabella 10 – Valori di assorbanza per il test SP-ELISA IgM
diabete
controlli
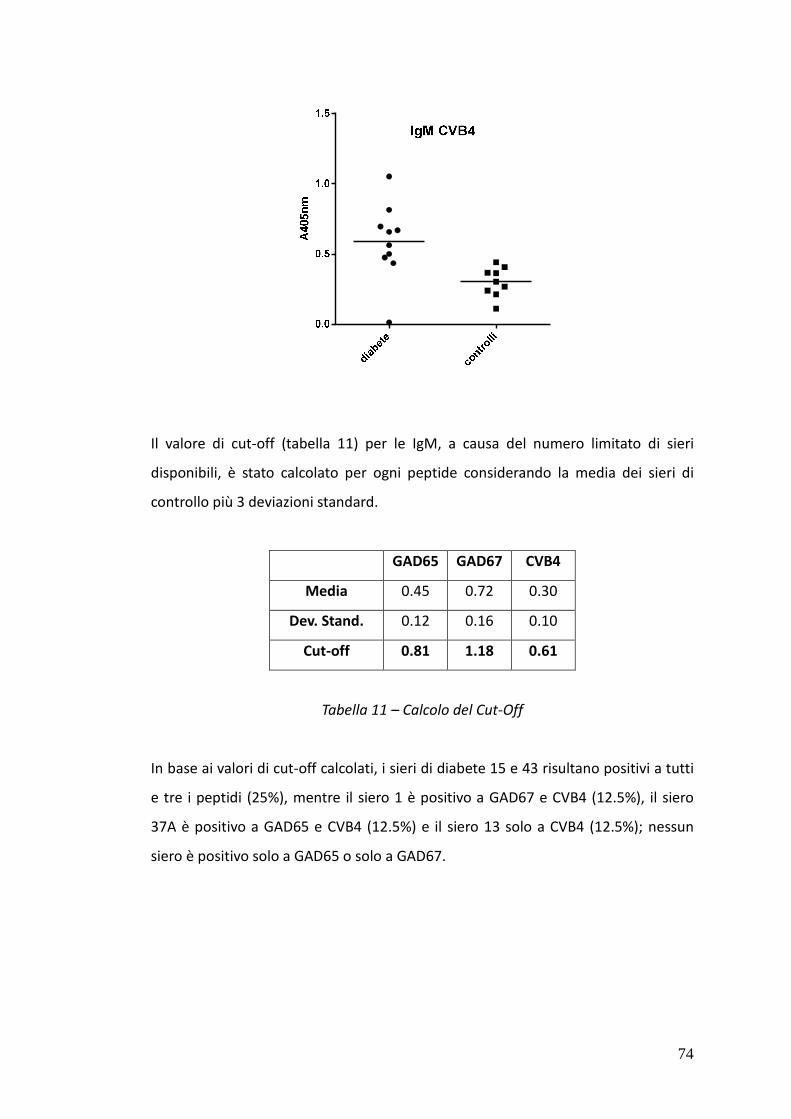
74
Il valore di cut-off (tabella 11) per le IgM, a causa del numero limitato di sieri
disponibili, è stato calcolato per ogni peptide considerando la media dei sieri di
controllo più 3 deviazioni standard.
GAD65 GAD67 CVB4
Media 0.45 0.72 0.30
Dev. Stand. 0.12 0.16 0.10
Cut-off 0.81 1.18 0.61
Tabella 11 – Calcolo del Cut-Off
In base ai valori di cut-off calcolati, i sieri di diabete 15 e 43 risultano positivi a tutti
e tre i peptidi (25%), mentre il siero 1 è positivo a GAD67 e CVB4 (12.5%), il siero
37A è positivo a GAD65 e CVB4 (12.5%) e il siero 13 solo a CVB4 (12.5%); nessun
siero è positivo solo a GAD65 o solo a GAD67.
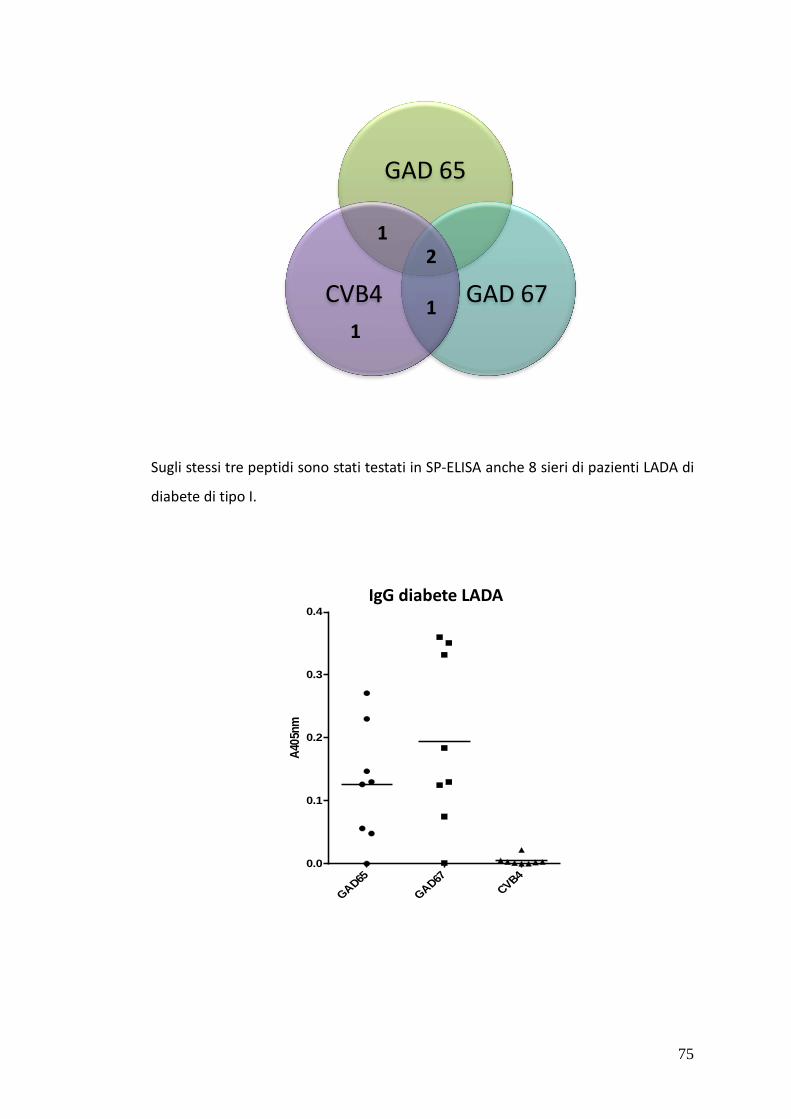
75
Sugli stessi tre peptidi sono stati testati in SP-ELISA anche 8 sieri di pazienti LADA di
diabete di tipo I.
IgG diabete Lapolla
A40
5nm
GAD65
GAD67CVB4
0.0
0.1
0.2
0.3
0.4
GAD 65
GAD 67CVB4
2
1
1
1
IgG diabete LADA
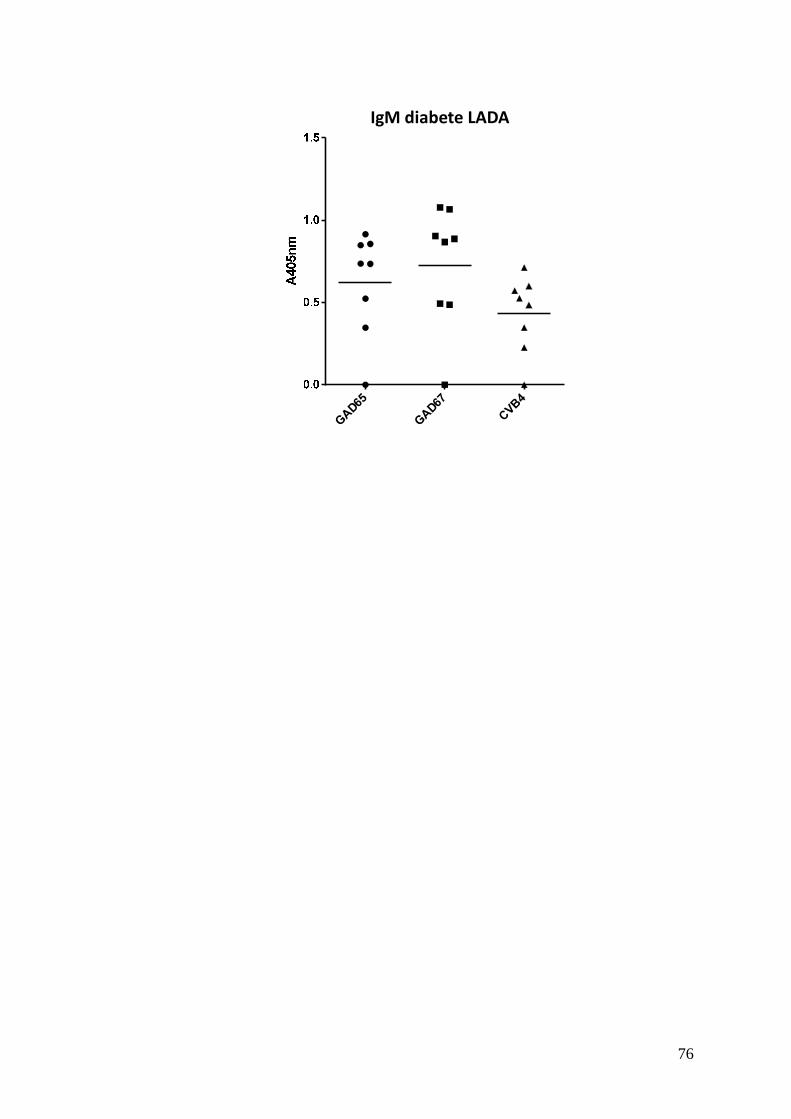
76
GAD65
GAD67
CVB4
LADA IgM diabete LADA

77
4 CONCLUSIONI E SVILUPPI FUTURI
Il diabete mellito di tipo 1, sia nella sua forma giovanile (Insulin-Dependent
Diabetes Mellitus, IDDM) che negli adulti (Latent Autoimmune Diabetes in Adults,
LADA) è una patologia sulla quale sono stati effettuati, negli ultimi decenni
numerosi studi, al fine di comprenderne i meccanismi molecolari e soprattutto le
cause scatenanti, che sono accettate essere di natura genetica e/o ambientale. Fra
gli agenti ambientali più probabili, legati allo scatenamento e allo sviluppo di questa
patologia, è sempre più accettato il ruolo di un’infezione virale precoce causata
dall’enterovirus Coxsackie B4. Il meccanismo che lega i due eventi rimane però
ancora ignoto. Una delle ipotesi più probabili al riguardo suggerisce un meccanismo
di mimicry verso una porzione, denominata P2C, di una proteina genomica del virus
rispetto a frammenti delle due isoforme della Glutamic Acid Decarboxylase (GAD65
e GAD67), un enzima coinvolto nella regolazione del glucagone. La P2C condivide
infatti con GAD65 e GAD67 un possibile epitopo, il frammento peptidico PEVKEK.
Questo mimetismo potrebbe generare una cross-reattività degli anticorpi anti-
Coxsackie verso le due isoforme della GAD e portare al riconoscimento di neo-
epitopi nella GAD stessa. Ne conseguirebbe un danno all’enzima da parte del
sistema immunitario e una conseguente disregolazione dei livelli di glucagone, con
possibile scatenamento della patologia. La GAD è infatti riconosciuta ad oggi come
uno dei più significativi autoantigeni legati al diabete di tipo I e gli anticorpi anti-
GAD sono accettati a livello clinico come uno dei biomarker più caratteristici di
questa patologia, non solo a livello diagnostico, ma anche predittivo.
Lo scopo di questo lavoro di tesi è stato quello di porre le basi per un
approfondimento mirato ad avvalorare questa ipotesi. Sono stati perciò selezionati
e sintetizzati tramite strategia di sintesi di peptidi su fase solida Fmoc/tBu assistita
da microonde, i tre frammenti peptidici:
• hGAD65 (250-273): 250AMMIARFKMFPEVKEKGMAALPRL273
• hGAD67(258-281): 258SIMAARYKYFPEVKTKGMAAVPKL281
• CVB4 P2C (28-50): 28FIEWLKVKILPEVKEKHEFLSRL50.
I peptidi sono stati sintetizzati sia in forma non acetilata all’estremità N-terminale (I-
III), che acetilata (IV-VI). Il motivo di tale scelta è introdurre un ulteriore legame

78
ammidico in posizione N-terminale, mimando un frammento interno alla proteina
putativamente antigenica e favorire così l’interazione con anticorpi specifici
eventualmente presenti nel siero dei pazienti. I peptidi sintetizzati sono stati
ottenuti con rese grezze valutate tramite HPLC rispettivamente dell’ 80% (I), 78% (II)
e 71% (III). I peptidi grezzi sono stati quindi pre-purificati tramite SPE ed infine
sottoposti a purificazione tramite HPLC semi-preparativa. Sono stati quindi ottenuti
tutti i peptidi con una purezza >95% (rese dal 20% al 30%). In tabella 16 paragrafo
5.1.4.3 sono riportati i dati di caratterizzazione tramite HPLC analitica ed ESI-MS.
I peptidi I, II e III sono stati utilizzati per studi di immunoaffinità mediante
Risonanza Plasmonica di Superficie (BIACORE T100), mentre i peptidi IV, V e VI sono
stati utilizzati in test immunoenzimatici su fase solida SP-ELISA. In entrambi i casi
sono stati utilizzati sieri di pazienti diabetici e controlli provenienti sia dall’Azienda
Ospedaliera Universitaria Meyer di Firenze (sieri IDDM) che dall’UOC di
Diabetologia e Dietetica dell’Università di Padova (sieri LADA).
In particolare, i sieri provenienti dal Meyer sono classificati, oltre che come diabete
di tipo 1 o controlli, anche in base alla presenza o meno di anticorpi anti-GAD,
identificati tramite un test commerciale utilizzato dalla AOU Meyer al momento del
prelievo. Nei sieri di pazienti IDDM è stato possibile rivelare, tramite SP-ELISA,
anticorpi solo di classe IgM. In particolare nel caso dei sieri 15 e 43 è stata osservata
una netta positività al test SP-ELISA da noi messo a punto contro tutti e tre i peptidi
IV, V e VI, mentre i sieri 1 e 37A hanno dato risultati positivi verso i peptidi hGAD67
(258-281) e CVB4 P2C (28-50) e verso i peptidi hGAD65 (250-273) e CVB4 P2C (28-
50) rispettivamente. I risultati sono riportati in tabella 10, paragrafo 3.5.2.1.
Inoltre sono stati ricercati anticorpi anche in una coorte ridotta di sieri appartenenti
a pazienti LADA, al fine di verificare se fossero presenti anche in soggetti adulti.
Anche in questo caso non sono stati ritrovati anticorpi IgG, ma solo IgM con valori
di assorbanza elevati verso le tre sonde peptidiche IV, V e VI. I risultati sono
riportati nei grafici del paragrafo 3.5.2.1. Dall’analisi dei dati emerge che nei sieri
IDDM e LADA sono presenti solo IgM, dirette contro le tre nuove sonde peptidiche
oggetto di questo lavoro di tesi ed in particolare hGAD65 (250-273), hGAD67 (258-
281) e CVB4 P2C (28-50). Essendo le IgM la prima classe di anticorpi prodotta in
caso di infezione nonché la prima a manifestarsi prima dello shift a IgG, è evidente

79
che la loro presenza può essere indice di reminiscenza di un’infezione mai
riconosciuta. Una possibile spiegazione dei nostri risultati potrebbe ritrovarsi in una
persistente e prolungata infezione, e quindi in una produzione latente di anticorpi
che, sulla base della nostra ipotesi di cross-reattività tra CVB4 P2C (28-50), hGAD65
(250-273) e hGAD67 (258-281), porterebbe nei sieri di pazienti diabetici, sia IDDM
che LADA, IgM dirette contro i peptidi della GAD. Avendo inoltre le tre sequenze in
comune tra loro solo il sequone PEVKEK, l’ipotesi di un fenomeno di mimicry
confermerebbe anche la natura del neo-epitopo scatenante la risposta aberrante
perché presente anche nei peptidi GAD da noi selezionati. Inoltre, dall’analisi del
Diagramma di Venn, riportato a pag. 73, si evince che, per quanto riguarda i
campioni provenienti da pazienti diabetici IDDM, i sieri 15 e 43 risultano positivi a
tutti e tre i peptidi (25%), mentre il siero 1 è positivo a hGAD67 (258-281) e CVB4
P2C (28-50) (12.5%), il siero 37A è positivo a hGAD65 (250-273) e CVB4 P2C (28-50)
(12.5%), mentre il siero 13 è positivo solo a CVB4 P2C (28-50) (12.5%). Nessun siero
è positivo solo a a hGAD65 (250-273) o a hGAD67 (258-281). Infine i peptidi I, II, e
III sono stati utilizzati per studi di immunoaffinità tramite risonanza plasmonica di
superficie (BIACORE T100). Il metodo utilizzato per questi test è ad oggi ottimizzato
per la rivelazione di soli anticorpi IgG, come mostrato in tabella 8, sono negativi..
Questi risultati confermano i dati negativi per le IgG ottenuti in SP-ELISA con i tre
peptidi testati. Misure d’interazione via SPR tra le IgM identificate per la prima volta
in questo lavoro di tesi e le nuove sonde peptidiche I, II e III sono attualmente in
corso. L’esiguo numero di sieri analizzati, nonostante abbia permesso di identificare
una cross-reattività tra le frammenti della sequenza virale e dell’enzima, non ha
permesso di ottenere un dato statisticamente significativo. L’analisi di un maggior
numero di sieri e il confronto dei nostri risultati con quelli ottenuti utilizzando il test
commercialmente disponibile, potranno portare ad una migliore comprensione del
fenomeno e ad una eventuale conferma dell’esistenza di cross-reattività con
l’infezione da Coxsackievirus. Inoltre l’analisi di un numero maggiore di sieri
provenienti da pazienti LADA potrebbe avvalorare ulteriormente la nostra ipotesi,
permettendo di estendere la nostra ipotesi anche a soggetti adulti precocemente
infettati.

80
5 PARTE SPERIMENTALE
5.1 SPPS – MATERIALI E METODI
L’acetonitrile (CH3CN) per HPLC, la dimetilformammide (DMF) e il diclorometano
anidro (DCM) per sintesi peptidica sono stati utilizzati senza ulteriori trattamenti.
Le sintesi peptidiche in fase solida sono state effettuate con il sintetizzatore Liberty
Blue (CEM Corporation), utilizzando una resina Fmoc-Rink Amide 0,48 mmol/g.
Il distacco dei peptidi dalla resina è stato effettuato con una miscela di TFA/H2O/TIS
(triisopropilsilano) (98:1:1). La reazione di acetilazione è stata effettuata usando
una miscela di anidride acetica/NMM (10 eq) in DCM. I prodotti sono stati analizzati
mediante HPLC analitica in fase inversa; si è utilizzato un apparecchio Waters
Alliance (modello 2695) equipaggiato con rivelatore a schiera di diodi, colonna
Jupiter Phenomenex C18 (5 μm, 300 Å, 250 × 4.6 mm di diametro interno), ad un
flusso di 1 ml/min di una miscela di eluenti a composizione variabile nel tempo: (A)
0.1% TFA in H2O (MilliQ) e (B) 0.1% TFA in CH3CN (84%). Per la purificazione dei
peptidi sono state utilizzate due tecniche:
1. Pre-purificazione mediante Solid-Phase Extraction (SPE) usando come
fase stazionaria gel di silice Merck LiChroprep RP-8 (40-63 m) e come
eluente una miscela a composizione variabile di H2O (MilliQ) e CH3CN ;
2. HPLC semipreparativa in fase inversa con strumento Waters modello 600
su colonne Jupiter C18 (10 μm, 250 × 10 mm) e colonne tipo Varian
Polaris di tipo Aqua (5 μm, C18, 200Å, 250 × 10 mm), a flussi di 4 ml/min.
con miscela H2O (MilliQ)/ CH3CN come eluente.
Gli spettri di massa sono stati registrati mediante spettrometro Waters micromass
ZQ, accoppiato ad un sistema UPLC analitico Acquity ultra performance LC Waters
dotato di una colonna C18 con flusso di 450 μL/min, usando come eluenti miscele a
composizione variabile nel tempo di: (A) 0.1% TFA in H2O (MilliQ) e (B) 0.1% TFA in
CH3CN 84% in H2O. Per la liofilizzazione è stato utilizzato un liofilizzatore Edwards
mod. Modulyo.
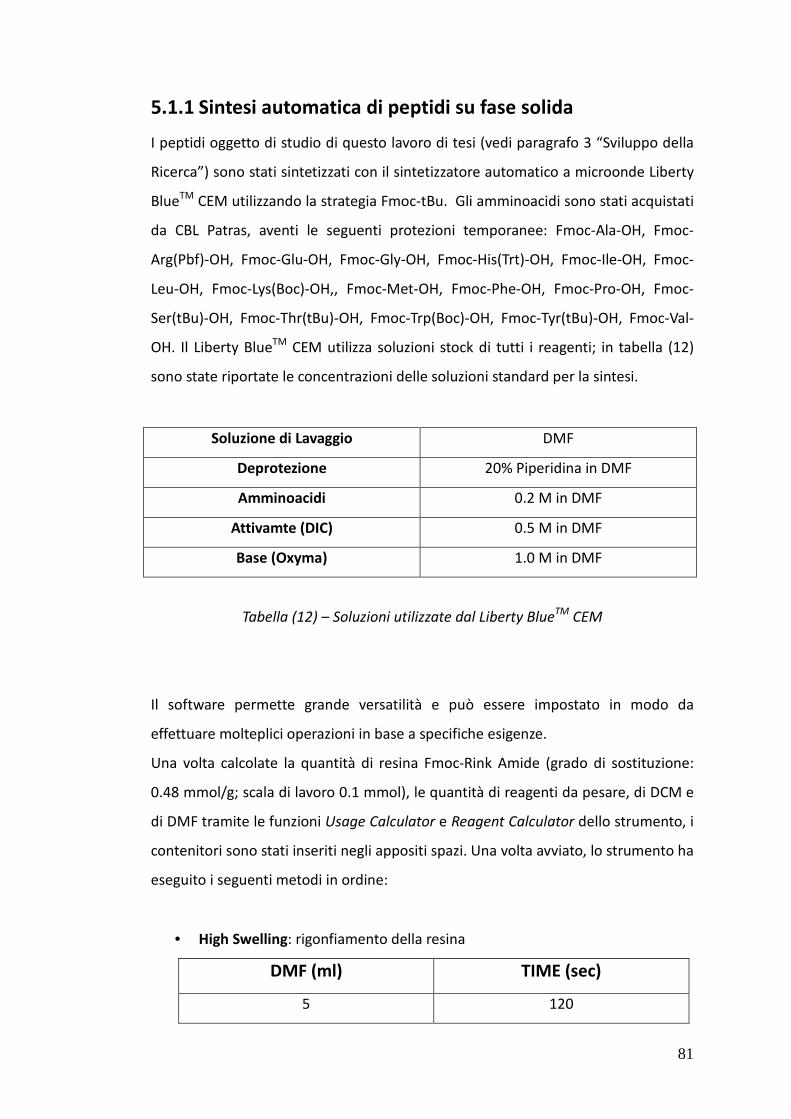
81
5.1.1 Sintesi automatica di peptidi su fase solida
I peptidi oggetto di studio di questo lavoro di tesi (vedi paragrafo 3 “Sviluppo della
Ricerca”) sono stati sintetizzati con il sintetizzatore automatico a microonde Liberty
BlueTM CEM utilizzando la strategia Fmoc-tBu. Gli amminoacidi sono stati acquistati
da CBL Patras, aventi le seguenti protezioni temporanee: Fmoc-Ala-OH, Fmoc-
Arg(Pbf)-OH, Fmoc-Glu-OH, Fmoc-Gly-OH, Fmoc-His(Trt)-OH, Fmoc-Ile-OH, Fmoc-
Leu-OH, Fmoc-Lys(Boc)-OH,, Fmoc-Met-OH, Fmoc-Phe-OH, Fmoc-Pro-OH, Fmoc-
Ser(tBu)-OH, Fmoc-Thr(tBu)-OH, Fmoc-Trp(Boc)-OH, Fmoc-Tyr(tBu)-OH, Fmoc-Val-
OH. Il Liberty BlueTM CEM utilizza soluzioni stock di tutti i reagenti; in tabella (12)
sono state riportate le concentrazioni delle soluzioni standard per la sintesi.
Tabella (12) – Soluzioni utilizzate dal Liberty BlueTM
CEM
Il software permette grande versatilità e può essere impostato in modo da
effettuare molteplici operazioni in base a specifiche esigenze.
Una volta calcolate la quantità di resina Fmoc-Rink Amide (grado di sostituzione:
0.48 mmol/g; scala di lavoro 0.1 mmol), le quantità di reagenti da pesare, di DCM e
di DMF tramite le funzioni Usage Calculator e Reagent Calculator dello strumento, i
contenitori sono stati inseriti negli appositi spazi. Una volta avviato, lo strumento ha
eseguito i seguenti metodi in ordine:
• High Swelling: rigonfiamento della resina
DMF (ml) TIME (sec)
5 120
Soluzione di Lavaggio DMF
Deprotezione 20% Piperidina in DMF
Amminoacidi 0.2 M in DMF
Attivamte (DIC) 0.5 M in DMF
Base (Oxyma) 1.0 M in DMF

82
• Deprotection Cycle: metodo di deprotezione della resina
T (°C) POWER (W) TIME (sec)
75 155 15
90 30 30
• Coupling Cycle: metodi di accoppiamento diversi a seconda che si tratti di
un comune amminoacido (Standard Coupling), di un’Arg (Double Arg
Coupling: viene ripetuto due volte un Arg Coupling) o di un’His (50 °C
Coupling)
STANDARD COUPLING
T(°C) POWER (W) TIME (sec)
90 170 15
90 30 110
DOUBLE Arg COUPLING
T(°C) POWER (W) TIME (sec)
25 0 1500
75 30 300
50°C COUPLING
T(°C) POWER (W) TIME (sec)
25 0 120
50 35 140
5.1.2 Cleavage dalla resina
Per il distacco del peptide dalla resina e la contemporanea deprotezione delle
catene laterali, è stato effettuato il cleavage con una miscela di TFA/H2O/TIS
(95:2.5:2.5) (1 ml ogni 100 mg di resina), mantenendo il tutto sotto continua
agitazione a temperatura ambiente per circa 3 ore. Successivamente è stata filtrata
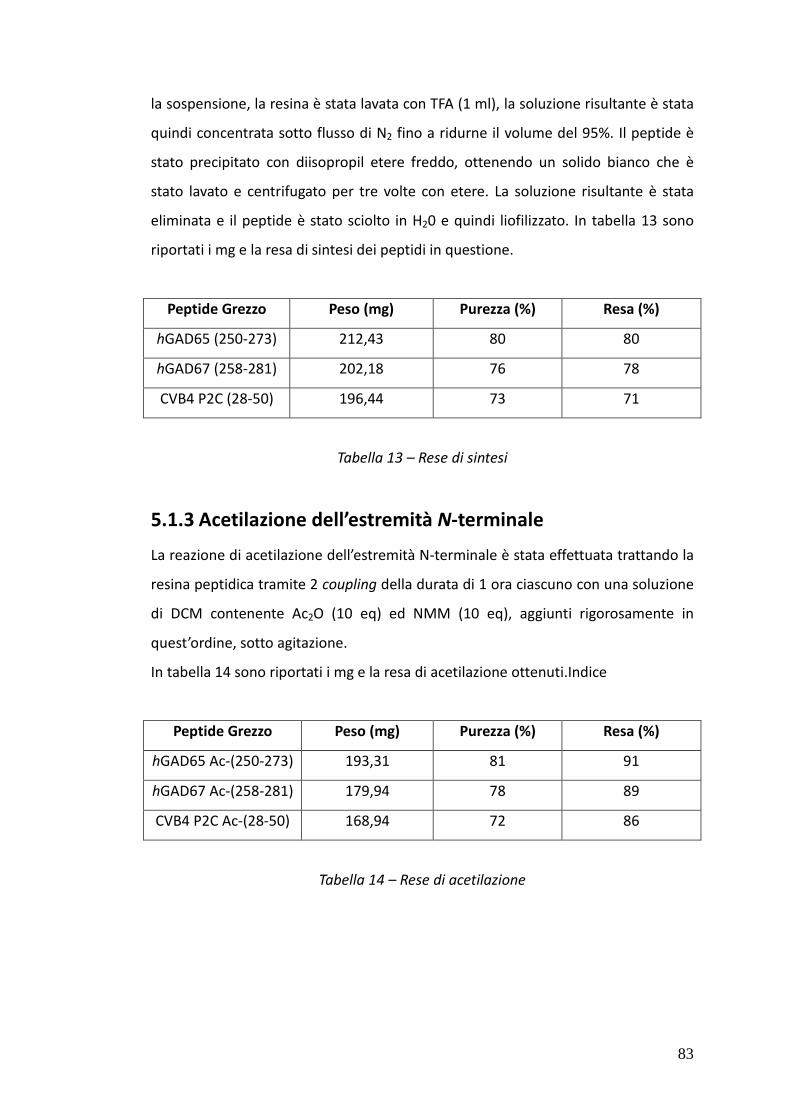
83
la sospensione, la resina è stata lavata con TFA (1 ml), la soluzione risultante è stata
quindi concentrata sotto flusso di N2 fino a ridurne il volume del 95%. Il peptide è
stato precipitato con diisopropil etere freddo, ottenendo un solido bianco che è
stato lavato e centrifugato per tre volte con etere. La soluzione risultante è stata
eliminata e il peptide è stato sciolto in H20 e quindi liofilizzato. In tabella 13 sono
riportati i mg e la resa di sintesi dei peptidi in questione.
Peptide Grezzo Peso (mg) Purezza (%) Resa (%)
hGAD65 (250-273) 212,43 80 80
hGAD67 (258-281) 202,18 76 78
CVB4 P2C (28-50) 196,44 73 71
Tabella 13 – Rese di sintesi
5.1.3 Acetilazione dell’estremità N-terminale
La reazione di acetilazione dell’estremità N-terminale è stata effettuata trattando la
resina peptidica tramite 2 coupling della durata di 1 ora ciascuno con una soluzione
di DCM contenente Ac2O (10 eq) ed NMM (10 eq), aggiunti rigorosamente in
quest’ordine, sotto agitazione.
In tabella 14 sono riportati i mg e la resa di acetilazione ottenuti.Indice
Peptide Grezzo Peso (mg) Purezza (%) Resa (%)
hGAD65 Ac-(250-273) 193,31 81 91
hGAD67 Ac-(258-281) 179,94 78 89
CVB4 P2C Ac-(28-50) 168,94 72 86
Tabella 14 – Rese di acetilazione

84
5.1.4 Purificazione dei peptidi
5.1.4.1 Pre-purificazione
I peptidi ottenuti sono stati sottoposti ad una prima fase di purificazione (pre-
purificazione) mediante lo strumento Armen Instrument. A questo scopo è stata
utilizzata una colonna impaccata con silice C18 (fase stazionaria, capacità di carico
5% in peso), e come eluente un gradiente crescente di H2O/CH3CN. L’operazione
prevede alcuni step principali:
• condizionamento della colonna con H2O;
• solubilizzazione del peptide in H2O (massimo 7 ml) ed eventuale
centrifugazione se rimangono dei residui solidi;
• inserimento della soluzione acquosa del peptide nel loop apposito dello
strumento
• eluizione con un gradiente H2O/CH3CN variabile a seconda del metodo
utilizzato. Le frazioni sono state controllate tramite HPLC analitica, riunite e
successivamente liofilizzate.
5.1.4.2 HPLC semipreparativa
I peptidi pre-purificati sono stati ulteriormente purificati mediante HPLC
semipreparativa, dopo aver selezionato il metodo più adatto tramite HPLC analitica.
Le frazioni raccolte sono state quindi caratterizzate sempre tramite HPLC analitica,
andando poi a separare le frazioni più pure da quelle più impure e liofilizzandole.
In tabella 15 sono riportate le rese di purificazione.
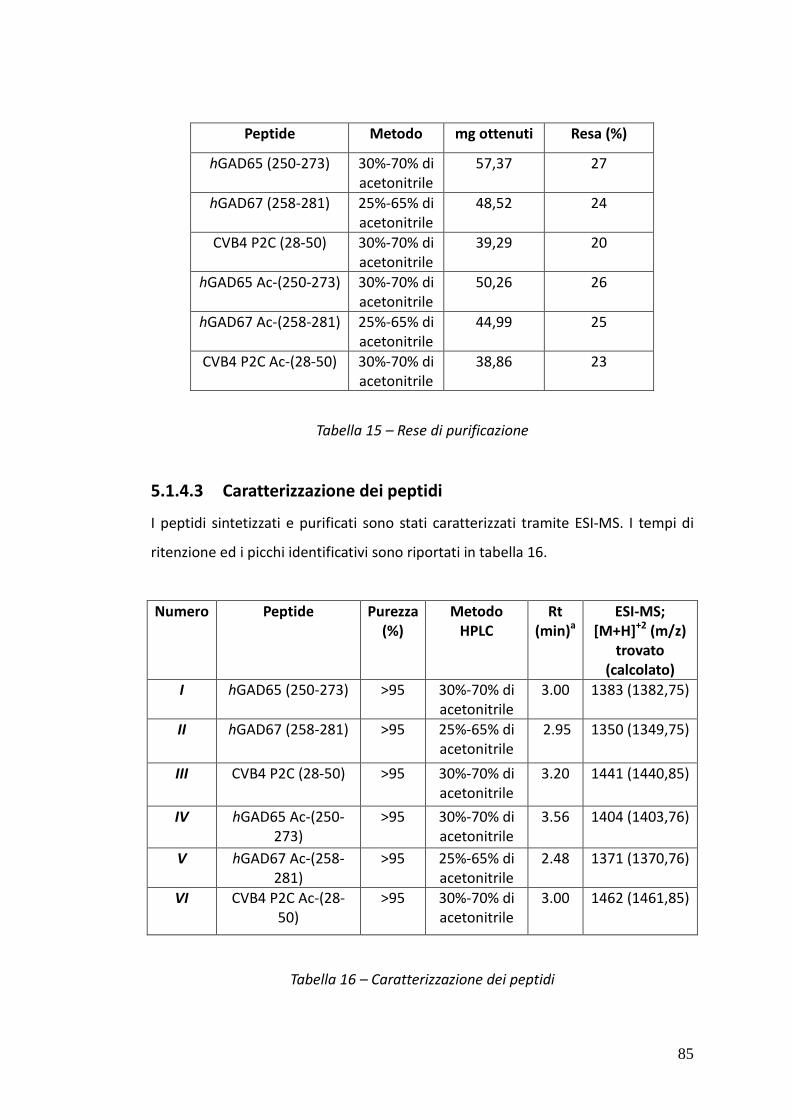
85
Peptide Metodo mg ottenuti Resa (%)
hGAD65 (250-273) 30%-70% di acetonitrile
57,37 27
hGAD67 (258-281) 25%-65% di acetonitrile
48,52 24
CVB4 P2C (28-50) 30%-70% di acetonitrile
39,29 20
hGAD65 Ac-(250-273) 30%-70% di acetonitrile
50,26 26
hGAD67 Ac-(258-281) 25%-65% di acetonitrile
44,99 25
CVB4 P2C Ac-(28-50) 30%-70% di acetonitrile
38,86 23
Tabella 15 – Rese di purificazione
5.1.4.3 Caratterizzazione dei peptidi
I peptidi sintetizzati e purificati sono stati caratterizzati tramite ESI-MS. I tempi di
ritenzione ed i picchi identificativi sono riportati in tabella 16.
Numero Peptide Purezza
(%)
Metodo
HPLC
Rt
(min)a
ESI-MS;
[M+H]+2
(m/z)
trovato
(calcolato) I hGAD65 (250-273) >95 30%-70% di
acetonitrile 3.00 1383 (1382,75)
II hGAD67 (258-281) >95 25%-65% di acetonitrile
2.95 1350 (1349,75)
III CVB4 P2C (28-50) >95 30%-70% di acetonitrile
3.20 1441 (1440,85)
IV hGAD65 Ac-(250-273)
>95 30%-70% di acetonitrile
3.56 1404 (1403,76)
V hGAD67 Ac-(258-281)
>95 25%-65% di acetonitrile
2.48 1371 (1370,76)
VI CVB4 P2C Ac-(28-50)
>95 30%-70% di acetonitrile
3.00 1462 (1461,85)
Tabella 16 – Caratterizzazione dei peptidi

86
5.2 TEST DI IMMUNOAFFINITA’ BIACORE – MATERIALI
E METODI
I test di immunoaffinità sui peptidi I,II,III sono stati effettuati tramite lo strumento
Biacore® T100 (GE Healthcare).
Tutte le soluzioni standard e i buffers sono stati preparati con acqua Milli-Q
ottenuta con il sistema Sartorius (arium® 611 VF). I sensor chips della serie CM5, N-
idrossisuccinimmide (NHS 0.1 M), sono stati forniti dalla GE Healthcare. Il running
buffer con cui si eseguono tutte le analisi di binding/cientica è il buffer HBS-EP+
(10x) così costituito: 0.1M HEPES, 1.5M NaCl, 30mM EDTA, 0.5% v/v Tween ed è
stato fornita dalla GE Healthcare e diluito 10 volte con acqua Milli-Q a pH 7.4. Il
sodio acetato e l’idrossido di sodio per i tamponi di immobilizzazione e
rigenerazione sono forniti dalla Carlo Erba (Milano). I buffers vengono filtrati con un
sistema MILLIPORE Express™ PLUS 0,22 µm.
5.2.1 Immobilizzazione dei peptidi sul chip
Il tampone di immobilizzazione migliore per ogni peptide è stato scelto eseguendo
la procedura del pH scouting. A tale scopo sono stati preparati i tamponi alle
concentrazioni ioniche e ai valori di pH desiderati:
- tampone acetato di sodio 1, 5, 10, 25 e 50 mM a pH 4.5, 5.5, 6
(quest’ultimo solo nel caso del 50 mM).
I tre peptidi sono stati solubilizzati nei tamponi elencati partendo da una soluzione
stock acquosa 1mg/ml ed eseguendo una diluizione 1:100 per arrivare a una
concentrazione finale di peptide 10 μg/ml.
La procedura del pH scouting è stata eseguita ripetendo cicli costituiti da:
- iniezione del campione per 120 secondi a flusso 10 μl/min;
- rigenerazione della superficie del chip dopo ogni iniezione per 60 secondi
con NaOH 0.1 M a flusso 10 μl/min.
I tamponi di immobilizzazione ottimali scelti per ciascun peptide sono riportati in
tabella 17:

87
PEPTIDE CONCENTRAZIONE pH
hGAD65 (250-273) (I) 5 mM 4.5
hGAD67 (258-281) (II) 10 mM 4.5
CVB4 P2C (28-50) (III) 25 mM 4.5
Tabella 17 - Buffer selezionati per l’immobilizzazione dei peptidi
L’immobilizzazione dei tre peptidi I, II, III in esame, rispettivamente sui canali 3, 2 e
4 del chip, è stata eseguita ripetendo i cicli riportati in seguito per ognuno dei tre
antigeni:
1) Due iniezioni di una soluzione di EDC/NHS 0.4 M / 0.1 M rispettivamente
per 420 e 60 s al flusso di 10 µl/min
2) Tre iniezioni del peptide in una soluzione 10 mM del tampone scelto tramite
pH scouting rispettivamente per 420, 360 e 360 s al flusso di 10 µl/min
5.2.2 Studi di Binding
Gli studi di binding sono stati effettuati iniettando contemporaneamente ogni siero
testato sui 4 canali del chip (rispettivamente BIANCO, I, II, III), attraverso il seguente
ciclo:
1) Iniezione del campione per 180 s al flusso di 30 µl/min
2) Dissociazione (iniezione di running buffer) per 60 s al flusso di 30 µl/min
3) Rigenerazione (1) attraverso l’iniezione di una soluzione di Gly 50 mM pH
2.5 per 30 s al flusso di 30 µl/min
4) Rigenerazione (2) attraverso l’iniezione di una soluzione di NaOH 0.1 M per
60 s al flusso di 30 µl/min
I risultati (parametro di stability) vengono considerati dopo 15 secondi dalla fine
dell’iniezione del campione, ovvero dopo 15 secondi dall’iniezione del running
buffer (punto 2).

88
5.3 TEST IMMUNOENZIMATICI IN FASE SOLIDA (SP-
ELISA) – MATERIALI E METODI
In tutti i test ELISA sono state utilizzate le piastre di polistirene da 96 pozzetti (ICN
Flow, Linbro/Titertek) e il coating è stato effettuato con i peptidi IV, V, VI. Le
soluzioni per eseguire il test ELISA sono le seguenti:
1. il coating buffer, che è un tampone carbonato a pH 9.6 costituito da
carbonato di sodio (Na2CO3) e bicarbonato di sodio (NaHCO3) 0.9%, con
aggiunta di sodio azide (NaN3) 0.02%
2. il lavaggio è stato eseguito con una soluzione Washing Buffer di NaCl 0.9% e
0.05% di tensioattivo Tween (poliossietilene sorbitan monolaurato)
3. il tampone FBS (siero bovino fetale), che è una soluzione che si deteriora
molto facilmente per cui è indispensabile farla fresca ogni qualvolta si
esegua l’esperimento. Il siero bovino fetale è diluito 1:10 con soluzione
Washing Buffer e Tween
4. il coniugato IgG e IgM va diluito in tampone FBS e una volta diluito può
essere mantenuto in frigorifero per non più di una settimana
5. I due coniugati sono:
• anticorpo IgM [A-1067, (γ-chain specific), F(ab’)2 fragment of goat
antibody SIGMA] marcato con l’enzima fosfatasi alcalina e diluito
1:200 in SBF/Tween
• anticorpo IgG [A-3312, (µ-chain specific), F(ab’)2 fragment of goat
antibody SIGMA], marcato con l’enzima fosfatasi alcalina e
diluito1:8000 in FBS/Tween
6. il Substrate Buffer, che è una soluzione costituita da 10% dietanolammina,
0.1% MgCl2 e portato a pH 9.8 con piccole aggiunte di HCl 37% e sodio azide
(NaN3) 0.02%.
7. la soluzione di bloccaggio, che è una soluzione 1 M di idrossido di sodio
(NaOH).
8. Le letture delle assorbanze sono state effettuate a 405 nm mediante
spettrofotometro Tecan-Sunrise.

89
I sieri dei pazienti sono stati selezionati dalla Prof. Chiara Azzari (IDDM) e dalla Prof.
Annunziata Lapolla (LADA) ed appartengono a pazienti affetti da diabete I ed a
donatori sani. I pazienti sono stai selezionati in maniera casuale e non
precedentemente sottoposti a test per la reattività immune tramite test ELISA o
BIACORE. Questo studio, i metodi e le procedure di consenso sono stati approvati
dal Comitato Etico dell’Azienda Ospedaliera Universitaria Meyer di Firenze e
dell’Azienda Ospedaliera Universitaria Padovana. Tutti i dati utilizzati in questo
studio sono stati riportati in forma anonima.
5.3.1 Test SP-ELISA – Fasi sperimentali
1 Coating: i pozzetti delle piastre sono stati funzionalizzati con la soluzione di
peptide sciolto in tampone carbonato 0.05 M (pH 9.6) alla concentrazione di
1µg/µL. Ciascun pozzetto è stato quindi riempito con 1 µL di soluzione e la
piastra incubata a 4°C per tutta la notte. Per ogni campione di peptide sono
state preparate due piastre in modo da testare l’interazione dell’antigene con
due anticorpi diversi;
2 Lavaggio: le piastre sono state lavate 5 volte con la soluzione di lavaggio;
3 Blocking: i siti di legame non specifico sono stati bloccati con 110 µL di una
soluzione al 10% di siero di bovino fetale (FBS), 0.9 % NaCl e Tween allo 0,05%
a temperatura ambiente per 1 h;
4 Applicazione dei sieri: i sieri sono stati aggiunti in triplicato alla diluizione di
1:100 in FBS–Tween. Le soluzioni sono state applicate in quantità di 100 µL per
pozzetto ed incubate a 4°C per tutte la notte;
5 Lavaggio: le piastre sono state lavate 5 volte con soluzione di lavaggio;
6 Aggiunta dell’antisiero: per ogni campione di peptide sono stati aggiunti 100
µl per ogni pozzetto di anticorpo secondario anti IgG AP umano, diluito 1:8000
in FBS-Tween o l’anticorpo secondario anti IgM AP umano con una diluizione
1:1200. Gli anticorpi si legano al frammento costante (Fc) delle Ig che sono
legate all’antigene. Le piastre sono state incubate per 3 h a temperatura
ambiente;
7 Lavaggio: le piastre sono state lavate 5 volte con la soluzione di lavaggio;

90
8 Applicazione del substrato: sono stati applicati 100 µL per pozzetto di
substrato (p-NPP, para-nitrofenilfosfato alla concentrazione di 1 mg/mL) in
tampone substrato. Si tratta di un cromogeno, cioè un composto non colorato
sul quale agisce la fosfatasi alcalina coniugata all’antisiero, un enzima che
reagisce in modo specifico con il substrato convertendolo in un prodotto di
colore giallo. L’intensità della colorazione di ogni singolo pozzetto dipende
dalla quantità di enzima presente e quindi dalla concentrazione di IgG e IgM
nel campione da testare. (Il cut-off per le IgG è Abs ≥ , mentre per le IgM è
Abs >)
9 Blocco della reazione: la reazione colorimetrica del substrato catalizzata dalla
fosfatasi alcalina, è stata bloccata dopo 30 min aggiungendo una soluzione di
NaOH 1 M (50 μL per pozzetto);
10 Lettura: l’assorbanza è stata letta mediante spettrofotometro Tecan-Sunrise
alla lunghezza d’onda di 405 nm.

91
6. Bibliografia
1. Guyton and Hall, Textbook of Medical Physiology, 2006, 11th edition., Elsevier
2. Richard A. Goldsby, Thomas J. Kindt, Barbara A. Osborne, Kuby Immunology,
2006 Aug 15, 6th edition, Macmillan
3. IUPAC, Glossary for chemists of terms used in biotechnology, 1992
4. Abul K. Abbas, Andrew W. Lichtman, Jordan S. Pober, Meccanismi Effettori
dell'Immunità Umorale in Immunologia cellulare e molecolare, 2002, 4ª ed., Piccin
5. Abul K. Abbas, Andrew W. Lichtman, Le basi dell’immunologia: fisiopatologia
del sistema immunitario, 2006, 2ª ed., Masson
6. Research Laboratories Merck, The Merck Manual, 2008, 5ª ed., Springer-Verlag
7. K.G. Alberti, Zimmet P.Z., Definition, diagnosis and classification of diabetes
mellitus and its complications. Part 1: diagnosis and classification of
diabetes mellitus provisional report of a WHO consultation, Diabetic Med 1998, Jul; 15(7):539-53
8. Tuomi T, Groop LC, Zimmet PZ, Rowley MJ, Knowles W, Mackay IR, Antibodies to glutamic acid decarboxylase reveal latent autoimmune
diabetes mellitus in adults with a non-insulin-dependent onset of disease, Diabetes 1993 Feb; 42(2):359–62
9. Cudworth AG, Woodrow JC., HLA system and diabetes mellitus, Diabetes 1975 Apr; 24(4):345-9
10. Naser SA, Ghobrial G, Romero C, Valentine JF, Culture of Mycobacterium
avium subspecies paratuberculosis from the blood of patients with Crohn's
disease, Lancet 2004 Sep; 364(9439):1039-44
11. van Halteren AG, Roep BO, Gregori S, Cooke A, van Eden W, Kraal G, Wauben MH, Cross-reactive mycobacterial and self hsp60 epitope recognition in I-
A(g7) expressing NOD, NOD-asp and Biozzi AB/H mice, J Autoimmun 2002 Mar; 18(2):139-47
12. Pinal C, Tobin A, Uniqueness and redundancy in GABA production, Perspectives on developmental neurobiology 1998; 5(2-3):109-18.
13. Wei J, Davis K, Wu H, Wu J, Protein Phosphorylation of Human Brain
Glutamic Acid Decarboxylase GAD65 and GAD67 and Its Physiological
Implications, Biochemistry 2004 May; 43(20):6182-9.
14. Battaglioli G, Liu H, Martin D (2003). "Kinetic differences between the
isoforms of glutamate decarboxylase: Implications for the regulation of
GABA synthesis". J. Neurochem 2003 Aug; 86(4):879-87
15. Shi Y, Kanaani J, Menard-Rose V, Hui Ma Y, Chang P, Hanahan D, Tobin A, Grodsky G, Baekkeskov S. Increased expression of GAD65 and GABA in

92
pancreatic β-cells impairs first-phase insulin secretion, Am J Physiol Endocrinol Metab. 2000 Sep; 279(3):E684-94.
16. Perea A, Clemente F, Martinell J, Villanueva-Peñacarrillo ML, Valverde I. Physiological effect of glucagon in human isolated adipocytes. Horm Metab Res. 1995 Aug; 27(8):372-5
17. Bason C., Lorini R., Lunardi C., In T1D a subset of anti-coxsackievirus B4
antibodies recognize autoantigens and induce apoptosis of pancreatic beta
cells, PLoS One. 2013; 8(2):e57729. doi: 10.1371/journal.pone.0057729
18. D L Kaufman, M G Erlander, M Clare-Salzler, M A Atkinson, N K Maclaren, and A J Tobin, Autoimmunity to two forms of glutamate decarboxylase in
insulin-dependent diabetes mellitus, J Clin Invest. 1992 Jan; 89(1):283-92.
19. Marc S. Horwitz1, Linda M. Bradley1, Judith Harbertson1, Troy Krahl1, Jae Lee1 & Nora Sarvennick, Diabetes induced by Coxsackie virus: Initiation by
bystander damage and not molecular mimicry, Nat Med. 1998 Jul; 4(7):781-5
20. Mark Namchuk, LeAnn Lindsay, Christoph W. Turck, Jamil Kanaani, and Steinunn Baekkeskov, Phosphorylation of Serine Residues 3, 6, 10, and 13
Distinguishes Membrane Anchored from Soluble Glutamic Acid
Decarboxylase 65 and Is Restricted to Glutamic Acid Decarboxylase 65α, J Biol Chem. 1997 Jan 17;272(3):1548-57
21. http://www.uniprot.org; Accessed 12-02-15
22. Tong J.C., Myers M.A., Mackay I.R., Zimmet P.Z., Rowley M.J. The PEVKEK
region of the pyridoxal phosphate binding domain of GAD65 expresses a
dominant B cell epitope for type 1 diabetes sera Ann N Y Acad Sci 2002 Apr; 958:182-9
23. Vreugdenhil GR, Batstra MR, Aanstoot HJ, Melchers WJ, Galama JM., Analysis of antibody responses against coxsackie virus B4 protein 2C and the
diabetes autoantigen GAD(65), J Med Virol. 1999 Oct; 59(2):256-61.
24. Wild T, Scherbaum WA, Gleichmann H, Landt M, Santiago J, Endl J, Landgraf R, Cavallo MG, Ganz M, Pozzilli P., Comparison of a new anti-glutamic acid
decarboxylase (GAD) enzyme-linked immunosorbent assay (ELISA) with
radioimmunoassay methods: a multicenter study, Horm Metab Res. 1997 Aug; 29(8):403-6.
25. Wasserfall CH, Atkinson MA, Autoantibody markers for the diagnosis and
prediction of type 1 diabetes, Autoimmun Rev. 2006 Jul; 5(6):424-8.
26. Tuomilehto J, Zimmet P, Mackay IR, Koskela P, Vidgren G, Toivanen L, Tuomilehto-Wolf E, Kohtamäki K, Stengård J, Rowley MJ., Antibodies to
glutamic acid decarboxylase as predictors of insulin-dependent diabetes
mellitus before clinical onset of disease, Lancet 1994 Jun 4; 343(8910):1383-5.
27. Gabriel, C.; Gabriel, S.; Grant, E. H.; Grant, E. H.; Halstead, B. S. J.; Mingos, D. M. P., Chem. Soc. Rev. 1998; 27(3): 213-224
28. A. Fini, A. Breccia, Pure appl. Chem., 1999; 71(4):573–579, IUPAC

93
29. Giada Rossi, Biosensor analysis of anti-citrullinated protein/peptide antibody
affinity, PhD Thesis, The University of Florence, 2014,
30. Wells A. Exploring the development of the independent, electronic, scholarly journal. M.Sc. Thesis, The University of Sheffield. 1999.
31. D. Kennett., J. Immunol. Methods, 1998; 106, 203-209.

94
7. RINGRAZIAMENTI
Sono tante le persone che voglio ringraziare; alcune perché hanno reso possibile la
realizzazione di questo progetto di tesi, che per me significa la degna conclusione di
un percorso durato tanti anni, fondamentale nella mia crescita come persona prima
di tutto e come scienziato in secondo luogo, altre perché non solo mi hanno
permesso di arrivare in fondo a questo viaggio, ma sono al mio fianco ogni giorno,
in tutte le esperienze, buone o cattive, che la vita mi ha riservato e mi riserverà
negli anni a venire.
Voglio cominciare quindi ringraziando i miei genitori, Mario e Patrizia, senza i quali,
ovviamente, non sarei qui e senza il supporto dei quali forse non sarei riuscito
nell’impresa. Mi hanno cresciuto secondo i più sani princìpi, mi hanno dato
coscienza, razionalità e buon senso, mi hanno lasciato la libertà di decidere e capire
da solo cosa è giusto e cosa è sbagliato, semplicemente mostrandomi quello che
secondo loro, fra i tanti, era il cammino corretto, ma permettendomi sempre di
scegliere, alla fine, quale imboccare.
Voglio ringraziare la mia principessa, la mia luna splendente, il mio bocciolo il cui
profumo è per me profumo di casa, Marika, che è stata ed è per me l’altra metà di
qualunque cosa io fossi stato nella mia interezza. È in lei che trovo ogni giorno la
forza di fare sempre una cosa in più del necessario, rendendo la vita traboccante di
esperienze, di emozioni e di gioie, ed impedendomi così di sprecare attimi
irripetibili di un’esistenza che, alla fine dei conti, non è poi tanto lunga.
Voglio ringraziare mio fratello, Mattia, che con i suoi eloquenti silenzi sa sempre
cosa non dirmi al momento giusto, ma che, quando infine apre bocca, non dice mai
qualcosa di superfluo o impulsivo, rendendo impossibile qualsivoglia litigio e
risultando di fatto il miglior fratello di sempre.
Voglio ringraziare mia sorella, Costanza, che è mia sorella pur non essendolo, con la
quale ho condiviso molto di più che con chiunque altro, l’unica persona al mondo
che, probabilmente, conosce la vera essenza di me stesso, perché quell’essenza ha
contribuito a plasmarla in 26 anni di meraviglioso, puro e genuino rapporto.

95
Voglio ringraziare i miei amici, che per fortuna sono tanti e che, chi in un modo chi
in un altro, hanno portato un pezzo indelebile di gioia nella mia esistenza: Dario, un
altro fratello non fratello, la cui spontaneità, genuinità e lealtà sono sempre state
per me fonte di ispirazione ed ammirazione; Matteo, con cui ho condiviso una parte
enorme della mia infanzia e della mia non indifferente capigliatura; Bernardo e
Benedetta, due dei miei punti di riferimento da bambino, ai quali mi sono ispirato e
che hanno avuto un’influenza non indifferente nella creazione in me un’ideologia
ed un modo di essere che tutt’ora sono cardine della mia personalità; Lorenzo, con
cui ho condiviso tanto, troppo per poterlo raccontare, ma che è per me sinonimo di
certezza, di presenza e di stabilità, perché so che non mancherà mai nella mia vita;
Francesco, perché è l’unica persona con cui ho condiviso anche tanto dolore, dolore
che però ci ha avvicinato in un modo che non percepisco con nessun altro; Vittoria,
altrettanto sorella, che è amore e bontà allo stato puro, uno di quei rapporti che
rimangono indissolubili negli anni; Edoardo, il mio modello per tutto il periodo delle
scuole medie e adesso un amico capace di rendere esilarante ogni momento,
sagace, intelligente e divertente come nessun altro abbia mai conosciuto; Giulia,
mia cugina, l’unica persona insieme alla quale posso dire di aver vissuto esperienze
letteralmente mortali, esperienze che però hanno creato un legame a cui non potrei
rinunciare per niente al mondo; il gruppo di Scandicci, Marco, Vanessa, Barbara,
Simone, Sara, Simone, Linda, Andrea, Dario e Giulia, la mia compagnia, alcuni fra i
miei amici più cari, che sono parte fondamentale della mia vita e che sono ciò che
mi aiuta a rimanere in contatto col mondo, impedendomi di perdermi nella mia
innegabile sociopatia; i Nature Mistakes, Stefano, Maria, Simone, Lorenzo e
Arianna, con i quali mi sono avvicinato ad una delle più grandi passioni della mia
vita, la musica, con i quali ho condiviso le prime esperienze su un palco, con i quali
ho sudato, sofferto e fatto sacrifici, ma grazie ai quali ho potuto vivere alcune fra le
emozioni più grandi di tutta la mia vita, suonando la nostra musica davanti alle
persone; David e Francisco, grazie ai quali questa passione continua ad ardere
sempre più forte; il mio maestro di chitarra, Iacopo, che, nonostante tutto, ha
l’innegabile merito di avermi trasmesso l’amore per la musica e per la chitarra; il
mio maestro, di vita e di capoeira, CM Paahppi, che mi ha raccolto come un filo
d’erba in mezzo alla bufera del mondo e che mi ha insegnato, negli anni, come

96
affrontare qualunque avversità, insegnandomi a camminare a testa alta,
insegnandomi il valore della mia individualità ed il rispetto per me stesso,
introducendomi alla cultura Bahiana e brasiliana, permettendomi di conoscere un
mondo ed una disciplina che ora sono colonna portante del mio io, del mio modo di
essere, di interpretare la mia vita e del mio modo di rapportarmi con le persone e
con il mondo; sua moglie Veronica, che mi ha insegnato il valore imprescindibile
della convinzione, che mi ha insegnato l’importanza fondamentale della sicurezza
ed a credere nelle mie affermazioni, e la cui razionalità e spontaneità è per me un
modello da seguire; i loro figli, Sirio ed Aynarah, che sono due angeli sulla terra;
tutti i ragazzi del gruppo Capoeira Angola Palmares, dai più vecchi, Lapo, Gnocco,
Pitù, Pititinga, Shrek, Jolly, Mami, Chiara, ai più nuovi, Poeta, Piu Piu, Minimè,
Ahmed, Michele, Andrea, Lisa, Zeppola, Cosimo, Irene, Mummia, Pulga, Cupido,
Marco, Bambu, Queixo, Acquatico, Giulia, Matilde e tutti gli altri, nei quali ho
trovato il gruppo di compagni di avventure che cercavo da sempre; tutti i capoeristi
che ho incontrato nella mia vita, fra cui CM Malicia e sua moglie Valentina, con la
loro stupenda figlia Isabella, grazie ai quali cresco ogni volta di più; I miei “allievi”,
Edoardo e Isabella, con i quali ho condiviso questa passione e che adesso sono due
amici importanti; gli amici incontrati all’università, Flavia, Fabiana, Ottavia, Matteo,
Gherardo, Giulia, Francesca, Martina, Samuele, Marco, Elena, e tanti altri, insieme
ai quali ho condiviso gioie, dolori e paure, ed insieme ai quali ho vissuto questa
magica avventura universitaria; i miei cuccioli, Liuba, Luna Darko, Nymeria e
Shamrock, che sono le mie gioie quotidiane; lo Zoo di 105, senza il quale non avrei
avuto la forza di affrontate con determinazione e ridendoci sopra tutte le difficoltà e
le prove degli ultimi anni.
Voglio ringraziare tutti i miei parenti, che sono tanti, tantissimi, ma che sono la
famiglia più bella che potessi chiedere di avere: Lidia e Daniela, le zie pazzerelle,
Stefano, il mio zio Fanano, il mio idolo, Rosalba, Mariangela, Aldo, Paolo, Giuseppe,
Leila, Massimo e Donatella, gli zii dei pranzi e delle cene, gli zii delle partite a carte,
gli zii della grande famiglia, Vincenzo e Bruna, i nonni della forza e della bontà, Tina
e Franco, i nonni della mia infanzia e del divertimento, tutti i miei mille cugini ed i
loro rispettivi mariti/mogli e figli, Andrea, Simone, Alessio, Carolina, Alessandra,
Ilaria, Alessandra, Chiara, che non mancano mai; mia zia Patrizia, che è zia pur non

97
essendolo, che mi ha insegnato il valore dell’arte, che mi ha insegnato a non
sprecare la mia vita, che mi ha insegnato a viverla al massimo col sorriso sulle
labbra, e che ora, da qualche parte nell’universo, controlla che io non faccia
ca****e, e ride della stupidità umana.
Voglio ringraziare tutte le persone che hanno fatto, in un modo o in un altro, parte
della mia vita; Pietro, Margherita, Federica e Michele, la cui pazienza (e, in
quest’ultimo caso, la cui palpebra) ha fatto si che, nella mia infanzia, avessi intorno
una famiglia anche fuori casa; Emilio, Rosalba e Guido, che si sono impegnati
affinché nella mia infanzia non mancasse mai quel pizzico di follia che rende la vita
interessante; Donatella e Giuseppe, che, come gli altri alle Croci, ci trattavano come
se fossimo tutti una grande famiglia allargata; Elisabetta e Gianluca, da cui mi sono
sempre sentito a casa; Mauro e Giovanna, con la loro giovinezza, ed i loro figli,
Corinna e Mattia, gli animatori della mia infanzia; Massimo, Gloria, Donatella e
Mauro, ancora una volta famiglie fuori casa; Alessandro e Cristina, divertenti,
frizzanti, spensierati, con cui ho vissuto alcune delle estati più belle; Jean e Ian, che
mi hanno fatto conoscere ed amare la Spagna; Lorenzo, il mio primo vero datore di
lavoro, che mi ha insegnato un mestiere, che mi ha raccontato mille storie e grazie
al quale ho provato per la prima volta l’ebrezza dell’indipendenza.
Voglio ringraziare tutti i ragazzi del PeptLab, Chiara, Francesca, Raffaello, Simona,
Alessandro, Mario e Giuseppina, per la pazienza, per gli insegnamenti, per i pranzi,
per aver reso il luogo di lavoro piacevole e privo di stress; la professoressa Anna
Maria Papini, per aver creduto in me e per avermi spronato quando ne avevo
bisogno. Voglio ringraziare tutte le persone che hanno contribuito alla mia
istruzione, che è una delle cose più importanti che si possano ricevere nella vita.
Ci sarebbero ancora tante, tantissime persone da ringraziare, comprese quelle che
in futuro entreranno a far parte della mia vita; purtroppo lo spazio scarseggia.
Quindi chiedo perdono a tutti coloro che mi sono dimenticato di citare, questo non
è altro che un debole tentativo di esprimere la mia gratitudine a tutti coloro che mi
hanno condotto fin qui; sia io che voi sappiamo che siete stati parte della mia vita,
indipendentemente dal fatto che siate o meno scritti in queste pagine.
Grazie, di cuore Lorenzo