Embed Size (px)
Citation preview

Teaching cells new tricksPhilippe Collas1,2 and Anne-Mari Hakelien1
1Institute of Medical Biochemistry, University of Oslo, PO Box 1112 Blindern, 0317 Oslo, Norway2Nucleotech, 33 Riverside Avenue, Westport, CT 06880, USA
The direct conversion of one differentiated cell type into
another – a process referred to as transdifferentiation –
would be beneficial for producing isogenic (patient’s
own) cells to replace sick or damaged cells or tissue.
Adult stem cells display a broader differentiation poten-
tial than anticipated and might contribute to tissues
other than those in which they reside. As such, they
could be worthy therapeutic agents. Recent advances in
transdifferentiation involve nuclear transplantation,
manipulation of cell culture conditions, induction of
ectopic gene expression and uptake of molecules from
cellular extracts. These approaches open the doors to
new avenues for engineering isogenic replacement
cells. To avoid unpredictable tissue transformation,
nuclear reprogramming requires controlled and herita-
ble epigenetic modifications. Considerable efforts
remain to unravel the molecular processes underlying
nuclear reprogramming and evaluate stable of the
changes in reprogrammed cells.
The human body has some ability to replace old, diseasedor damaged tissue throughout life. Skin and blood arecontinuously replaced; blood vessels, liver, bone andmuscle have a limited capacity for self-repair. However,age- or accident-related conditions, such as Parkinson’s orAlzheimer’s disease, osteoporosis, diabetes, cardiovascu-lar disease or spinal cord injury, cause damage that thebody’s regenerative capacity cannot cope with. Scarssometimes form that functionally impair tissues. Drugscan alleviate symptoms and relieve pain but they cannotpermanently repair diseased or damaged tissues.
Ideally, damaged organs have to be repaired orreplaced. Organ transplantation from immunologicallycompatible donors is the obvious method but the desperateshortage of donors imposes limitations on the number oftransplantations and the list of patients awaiting organtransplant augments each year. Artificial donor organsor devices might be an option; however, despiteencouraging advancements, their functional propertiesand life-span are often limiting. A third alternativecould be xenotransplantation – the transfer of cells ororgans from another species – but xenotransplantationremains controversial primarily because of risks ofdisease (e.g. retroviral) transmission from the graftedtissue. Additionally, organ transplants from human donorsand xenografting involve the use of immunosuppressants,which often have serious side effects when taken overa lifetime.
Imagine one could take healthy cells from your body, forexample in the form of a skin biopsy, convert them into thecell type that you need to cure your disease and implantthe modified healthy cells back into your body. Skin cells orbone marrow cells could become neurons, pancreas, heart,liver, gut or muscle. Imagine one could even teach cellsnew tricks inside your body: part of the liver could becomepancreas and help overcome diabetes. This might soundlike realms of science fiction but advances in methods forreprogramming cells open doors to the promised land ofautologous cell-based therapy.
Therapeutic potential of adult stem cells
Stem cells undergo self-renewing divisions, whereby thedaughter cells retain the exact same characteristics as theparent but can also give rise to more committed progenitorcells that differentiate into specific cell types (Fig. 1).Tissue-specific stem cells are found in adult tissuesincluding blood, skin, central nervous system, liver,gastrointestinal tract, fat and skeletal muscle [1,2]. Stemcells are responsible for regenerating damaged tissue andmaintaining tissue homeostasis. In contrast to thepluripotent embryonic stem (ES) cells, the differentiationpotential of adult stem cells has been thought to berestricted to the tissues in which they reside. Adult stemcells, however, can differentiate into a variety of tissues(Table 1). Bone marrow-derived cells not only replenishblood but can also contribute to heart, brain, liver, muscle,
Fig. 1. Stem cells, progenitor cells and differentiated cells. Pluripotent stem cells
proliferate into self-renewing stem cells. Stem cells can also divide asymmetrically
to produce more committed progenitors, which in turn are destined to give rise to
differentiated cells.
Stem cell
Progenitorcells
Differentiated cells
TRENDS in Biotechnology
Corresponding author: Philippe Collas ([email protected]).
Review TRENDS in Biotechnology Vol.21 No.8 August 2003354
http://tibtec.trends.com 0167-7799/03/$ - see front matter q 2003 Elsevier Science Ltd. All rights reserved. doi:10.1016/S0167-7799(03)00147-1
skin, and vascular endothelium. Neural stem cells can giverise to blood or skeletal muscle; hepatic stem cells can turninto pancreatic islet cells or intestine; skin stem cells canproduce smooth muscle, neurons, glia or adipocytes;adipose tissue-derived stem cells can differentiate intoneuronal, chondrogenic, myogenic or osteogenic cells.Thus, at least in some situations, adult stem cells appearto show sufficient plasticity to differentiate into cell types
outside their predicted developmental lineage and evencross germ layer boundaries.
The therapeutic potential of adult stem cells has drawncohorts of biotechnology companies to test the differen-tiation and engraftment possibilities of precursors ofblood, bone, cartilage, muscle or skin (Table 2). Effortsare being put towards defining clinically suitable culturesystems for ex vivo culture of stem cells. This would allownot only their expansion but also enrichment of specificstem cell subpopulations (which might have differentproliferation and differentiation properties), repair ofdefective genes or elimination of tumour cells fromstem cells. Predictions of tissue repair or replacementpossibilities should not, however, undermine how little weknow about the reliability of the changes induced ondifferentiation of adult stem cells. Demonstration of cellphenotype alteration does not necessarily provide anyindication as to whether the cell is functional or will be oftherapeutic use.
Slippery road?
Despite the hype resulting from the differentiationpotential of adult stem cells, the extent of their plasticityremains debated [2]. First, alterations in stem cell fate arerare occurrences [3]. Second, one must be certain that thedifferentiated cells are derived from the stem cells ofthe donor tissue and not from contaminating cells. Third,the ploidy of differentiated stem cells should be assessed.Co-culture of somatic cells or adult stem cells with ES cellscan lead to rare cell fusion events (one in 105 cells),resulting in hyperdiploid hybrids that display an ES cell-like phenotype [4,5]. However, it seems unlikely that cellfusion would account for all instances of reporteddifferentiation plasticity of stem cells.
Table 1. Differentiation potential of adult stem cells
Precursor cell Derivative Refs
Neural Skeletal muscle [43]
Epidermis [43]
Intestine & liver epithelium [43]
Heart tissue [43]
All germ layers [43]
Erythroid & myeloid lineage [44]
Oligodendrocyte Neurons [45]
Bone marrow Neurons [46–48]
Hepatocytes [49–52]
Skeletal muscle [49,53]
Cardiomyocytes [54,55]
[56]
All germ layers [57]
Endothelial cells [55,58–60]
Muscle Bone marrow [61–63]
Blood [44,53]
Liver Islet cells [64]
Pancreatic duct, acini [65]
Hepatocytes [64]
Bile duct [65]
Intestine epithelium [65]
Skin (dermis) Smooth muscle [66]
Neurons, glia [66]
Adipocytes [66]
Erythroid & myeloid lineage [67]
Fat Adipocytes, cartilage, bone [68,69]
Muscle [68–70]
Neurons [69,71,72]
Table 2. Biotechnology companies with focus on cell-based therapeutics
Company Focus
Stem cells
(allogeneic)
Stem cells
(autologous)
Cell reprogramming
(autologous)
Xenotransplantation
Aastrom Biosciences: http://www.aastrom.com X X
Aegera Therapeutics: http://www.aegera.com X X
Alexion Pharmaceuticals: http://www.alexionpharm.com X
Artecel Sciences: http://www.artecel.com X X
BioTransplant: http://www.biotransplant.com X X
BresaGen Ltd: http://www.bresagen.com.au X X X
Circe Biomedical: http://www.circebio.com X
Curis: http://www.curis.com X
Geron: http://www.geron.com X X X
Incara Pharmaceuticals: http://www.incara.com X X
Intercytex: http://www.intercytex.net X X
Ixion Biotechnology: http://www.ixion-biotech.com X
Layton Bioscience: http://www.laytonbio.com X
MicroIslet: http://www.microislet.com X
Novocell: http://www.novocell.com X
Nucleotech: http://www.nucleotech.com X
Oncosis: http://www.oncosis.com X X
Osiris Therapeutics: http://www.osiristx.com X
PPL Therapeutics: http://www.ppl-therapeutics.com X
StemCells: http://www.stemcellsinc.com X X
TheraTec: http://www.theratec.com X X
Viacell: http://www.viacellinc.com X
Review TRENDS in Biotechnology Vol.21 No.8 August 2003 355
http://tibtec.trends.com

Transdifferentiation
Transdifferentiation can be defined as a direct switch of analready differentiated cell to another type of differentiatedcell (Fig. 2a). Accordingly, transdifferentiation does notinvolve dedifferentiation – a resetting of the develop-mental clock. For a cell to reach a transdifferentiated state,several criteria should be fulfilled. Imperatively, a set ofgenes must be turned on while another set of genes isturned off. If this transition takes place rapidly, then theproducts of both sets of genes could be transiently co-expressed in the cell [6]. Markers of both donor and targetcell types might also co-exist if the reprogramming processleading to transdifferentiation is incomplete. This might
result in cells with a hybrid phenotype. If transdifferentia-tion is slow, there might be a stage at which the originalgenes are off before the new set of genes is activated [6].However, this does not necessarily mean that the cell hasreached a dedifferentiated state. Transdifferentiated cellsshould acquire a new, characterized phenotype that isstably expressed. If used in transplantation studies, thecells should be genetically labeled to ensure their trace-ability in the organism. Furthermore, the transplantedcells should be able to be withdrawn, eliciting as a resultthe re-occurrence of symptoms. This would demonstratethat rescue of a phenotype is a consequence of engraftmentof the transdifferentiated cells.
The microenvironment, such as contact with surround-ing cells, extracellular matrix, growth and differentiationfactors, has a key role in redirecting cell fate. Environ-mentally induced changes in cell fate occur naturally ininsects, amphibians, avians and mammals [7–9]. Strat-egies for reprogramming cells through nuclear transplan-tation (‘therapeutic cloning’; Box 1), inducing ectopic geneexpression or promoting the introduction of molecular‘soups’ into cells are being investigated. All raise hopes fora future application of a novel cell-replacement therapy.
Turning myoblasts into adipocytes
A common tissue culture model for myogenesis relies onmouse G8 myoblasts, which can differentiate into myo-tubes in vitro. Myoblasts are geared to muscle differen-tiation. However, ectopic expression of PPARg andC/EBPa, two transcription factors controlling adipogen-esis, can inhibit myogenesis and enable the conversion ofG8 myoblasts to an adipocyte phenotype on hormonalstimulation [10]. Wnts are a family of paracrine andautocrine factors that regulate cell growth and cell fate.Wnt signaling acts as an adipogenic switch that repressesadipogenesis when turned on and stimulates adipogenesiswhen switched off [11]. Disruption of Wnt signalingtransdifferentiates C2C12 myoblasts into adipocytesaccumulating lipids [11].
Transdifferentiation of osteoblasts into adipocytes
Osteoblasts and adipocytes are phenotypically distinct butare believed to differentiate from a common stromalprogenitor [12]. Their respective phenotype can be fosteredor maintained in vitro. Manipulating culture conditionscan induce transdifferentiation of mature human osteo-blasts into adipocytes [13]. Single human adipocytes mightalso dedifferentiate into fibroblast-like cells, which in turncan convert into osteoblasts or adipocytes depending onculture conditions [14]. Communication through gapjunctions modulates the expression of genes regulatingthe osteoblastic phenotype, and gap junctions are downregulated during adipogenesis. Inhibition of gap junctionsconverts human osteoblast cell lines and bone-derivedprimary osteoblasts cultured under osteogenic conditionsinto cells with an adipocyte phenotype [15] (Fig. 2b). To ourknowledge, converting fat tissue to bone has not yet beenachieved. Nevertheless, Schiller and colleagues [15] raisethe possibility of modulating gap junction communicationas a therapeutic approach to favour bone, rather thanadipose tissue, formation in patients with osteoporosis.
Fig. 2. Examples of transdifferentiation. (a) Differentiation and transdifferentiation.
(b) Lipophilic staining (Sudan IV labeling) of MC3T3-E1 preosteoblastic cells
treated with the gap junction inhibitor 18-a-glycyrrhetinic acid or a control non-
inhibitory analogue. The inhibitor induces transdifferentiation of the preosteo-
blasts into adipocytes containing Sudan IV-labeled lipid droplets (brown label).
Taken from [15] with permission. (c) Differentiation of endothelial cells into cardiac
cells in vitro. Co-culture of primary endothelial cells isolated from a LacZ-expres-
sing mouse embryo with cardiomyocytes induces expression of the cardiac sarco-
meric-specific marker troponin 1 (red label). Left panel, b-gal staining of the
differentiated cell. Arrows point to the two nuclei of a binucleated cell. Taken from
[16] with permission. (d) Electron micrograph of a differentiated cardiomyocyte
showing sarcomeres (arrow) and positive b-gal staining (arrowhead). Taken from
[16] with permission. (e) Induction of ectopic pancreatic tissue (EP) in the liver of a
Xenopus laevis tadpole expressing a modified form of the pancreas-specific tran-
scription factor PDX-1 exclusively in the liver. P refers to the endogenous pancreas.
Green-fluorescent protein label (green) marks both endogenous and ectopic
pancreatic tissues. Reproduced, with permission, from [23].
(a)
(b)
(c)
(e)
(d)
Stemcell
Progenitorcell
Mature cell
Mature cell
Transdifferentiation
Differentiation
GJinhibitorControl
TRENDS in Biotechnology
Review TRENDS in Biotechnology Vol.21 No.8 August 2003356
http://tibtec.trends.com
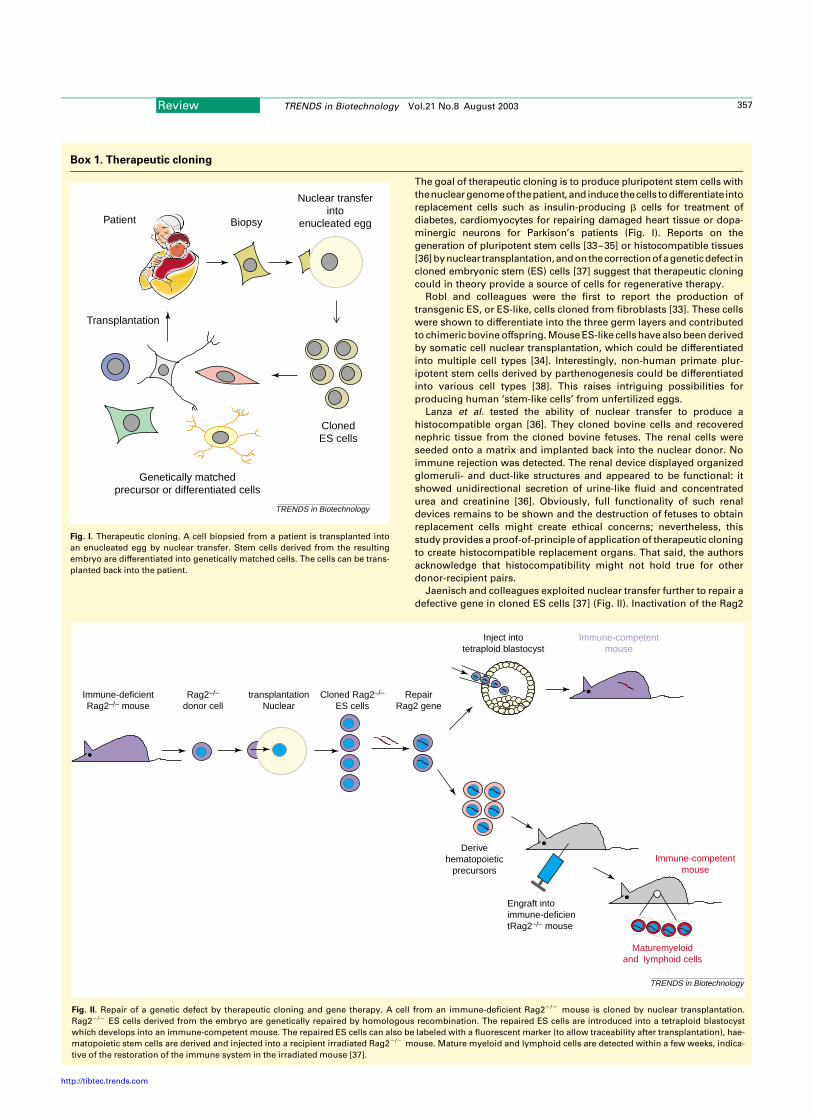
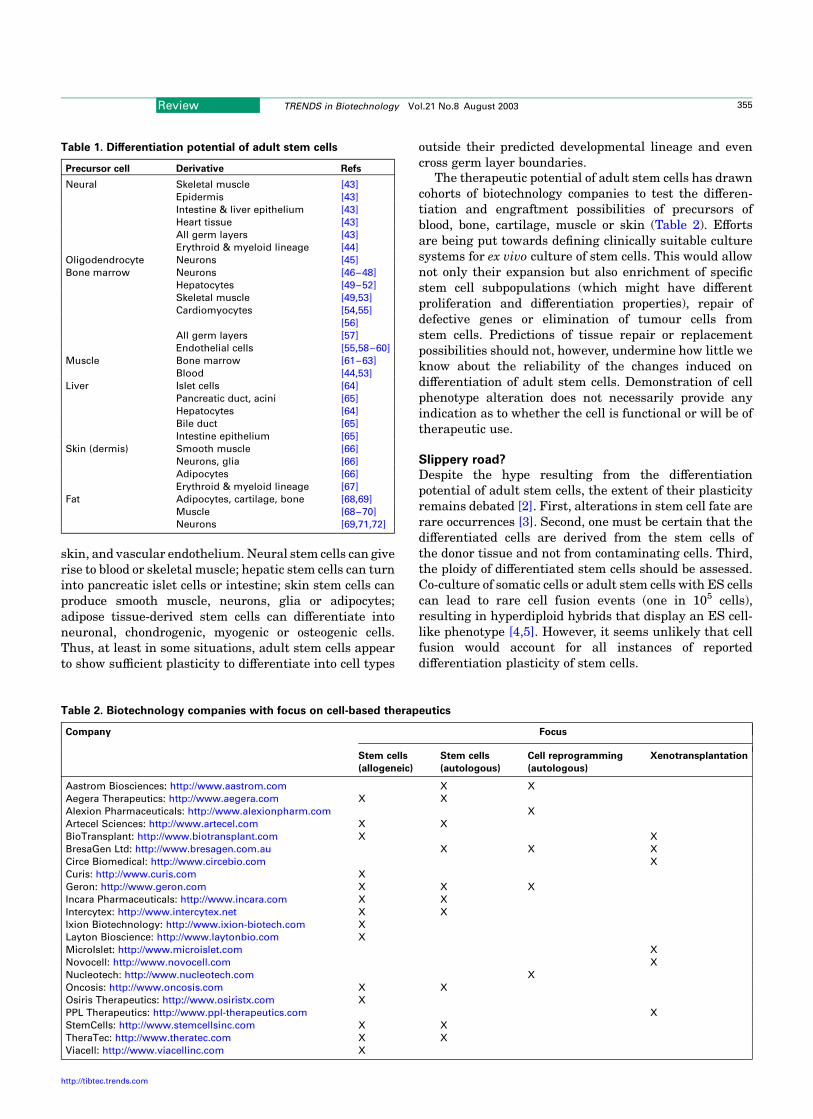
Box 1. Therapeutic cloning
The goal of therapeutic cloning is to produce pluripotent stem cells with
thenucleargenomeof thepatient,and inducethecells todifferentiate into
replacement cells such as insulin-producing b cells for treatment of
diabetes, cardiomyocytes for repairing damaged heart tissue or dopa-
minergic neurons for Parkison’s patients (Fig. I). Reports on the
generation of pluripotent stem cells [33–35] or histocompatible tissues
[36]bynuclear transplantation,andonthecorrectionofageneticdefect in
cloned embryonic stem (ES) cells [37] suggest that therapeutic cloning
could in theory provide a source of cells for regenerative therapy.
Robl and colleagues were the first to report the production of
transgenic ES, or ES-like, cells cloned from fibroblasts [33]. These cells
were shown to differentiate into the three germ layers and contributed
to chimeric bovine offspring. Mouse ES-like cells have also been derived
by somatic cell nuclear transplantation, which could be differentiated
into multiple cell types [34]. Interestingly, non-human primate plur-
ipotent stem cells derived by parthenogenesis could be differentiated
into various cell types [38]. This raises intriguing possibilities for
producing human ‘stem-like cells’ from unfertilized eggs.
Lanza et al. tested the ability of nuclear transfer to produce a
histocompatible organ [36]. They cloned bovine cells and recovered
nephric tissue from the cloned bovine fetuses. The renal cells were
seeded onto a matrix and implanted back into the nuclear donor. No
immune rejection was detected. The renal device displayed organized
glomeruli- and duct-like structures and appeared to be functional: it
showed unidirectional secretion of urine-like fluid and concentrated
urea and creatinine [36]. Obviously, full functionality of such renal
devices remains to be shown and the destruction of fetuses to obtain
replacement cells might create ethical concerns; nevertheless, this
study provides a proof-of-principle of application of therapeutic cloning
to create histocompatible replacement organs. That said, the authors
acknowledge that histocompatibility might not hold true for other
donor-recipient pairs.
Jaenisch and colleagues exploited nuclear transfer further to repair a
defective gene in cloned ES cells [37] (Fig. II). Inactivation of the Rag2
Fig. I. Therapeutic cloning. A cell biopsied from a patient is transplanted into
an enucleated egg by nuclear transfer. Stem cells derived from the resulting
embryo are differentiated into genetically matched cells. The cells can be trans-
planted back into the patient.
ClonedES cells
Genetically matchedprecursor or differentiated cells
Transplantation
Nuclear transferinto
enucleated eggPatient Biopsy
TRENDS in Biotechnology
Fig. II. Repair of a genetic defect by therapeutic cloning and gene therapy. A cell from an immune-deficient Rag22/2 mouse is cloned by nuclear transplantation.
Rag22/2 ES cells derived from the embryo are genetically repaired by homologous recombination. The repaired ES cells are introduced into a tetraploid blastocyst
which develops into an immune-competent mouse. The repaired ES cells can also be labeled with a fluorescent marker (to allow traceability after transplantation), hae-
matopoietic stem cells are derived and injected into a recipient irradiated Rag22/2 mouse. Mature myeloid and lymphoid cells are detected within a few weeks, indica-
tive of the restoration of the immune system in the irradiated mouse [37].
TRENDS in Biotechnology
Immune-competentmouse
Maturemyeloidand lymphoid cells
Derivehematopoietic
precursors
Engraft intoimmune-deficientRag2–/– mouse
Inject intotetraploid blastocyst
Immune-competentmouse
Immune-deficientRag2–/– mouse
Rag2–/–
donor celltransplantation
NuclearCloned Rag2–/–
ES cellsRepair
Rag2 gene
Review TRENDS in Biotechnology Vol.21 No.8 August 2003 357
http://tibtec.trends.com

Converting endothelial cells to cardiomyocytes
Endothelial cells isolated from mouse embryonic vessels,the umbilical vein or established cultures can transdiffer-entiate into beating cardiomyocytes on co-culture withneonatal cardiomyocytes [16]. Differentiation was evi-denced by expression of troponin I, a cardiac-specificsarcomeric protein (Fig. 2c), formation of sarcomeres(Fig. 2d) and electrical coupling of the cardiac cells throughgap junctions. Further indication of transdifferentiationwas provided by injection of genetically labeled endothelialprogenitors near the damaged area of a heart afterocclusion of a coronary vessel. The labeled cells differ-entiated into myosin-positive cells with a typical cardio-myocytes phenotype, suggesting the regeneration of amyocardium [16]. The plasticity of endothelial cells mightcreate possibilities for autologous cell replacement.
From pancreas to liver…
The appearance of hepatic foci in the pancreas undervarious experimental conditions is well-documented [17].Early studies have shown that a copper-depleted dietinduces a loss of pancreatic acinar cells in adult rats [18].Development of hepatic foci occurs when the animals arereturned to a normal diet, presumably from pancreaticduct cells. These cells resemble progenitor cells of the liverand grafting these cells into the liver can elicit transdiffer-entiation into hepatocytes [19]. Overexpression of thekeratinocyte growth factor KGF7 under the control of a ratinsulin promoter in the pancreas of transgenic mice alsopromotes the appearance of hepatocytes within islets ofLangerhans [20]. These studies show the potential ofpancreatic duct, acinar and endocrine cells to transdiffer-entiate into hepatocytes.
The development of controlled systems for investigatingtransdifferentiation of pancreas to liver shed light on theunderstanding of this process. Tosh and colleaguesrecently demonstrated that a rat pancreatic cell line andmouse fetal pancreatic tissue could take on characteristicsof hepatocytes after treatment with dexamethasone orafter transfection with C/EBPb [21]. A proportion of thehepatocytes converted directly from differentiated exo-crine cells without cell division. Although the paper did not
address whether these hepatocytes could effectivelyrepopulate the liver, it outlines a pathway of transdiffer-entiation of pancreas to liver and shows that C/EBPb is akey factor distinguishing between hepatic and pancreaticdifferentiation pathways. The study also demonstratesplasticity within the endodermal lineage [21]. Reprogram-ming somatic cells into hepatocytes would be useful totransiently create a population of hepatocytes to replenishthe liver after lobectomy. It would also be useful forresearch purposes because primary human hepatocytesare not readily accessible.
… back to pancreas
In addition of the ability of hepatic stem cells todifferentiate into pancreatic endocrine cells (Table 1),transdifferentiation of mature hepatocytes to pancreas isalso possible. Expression of the insulin gene is restricted tothe islet b cells of the pancreas. The transcription factorPDX-1 is essential for pancreatic development, islet cellfunction and insulin gene activation. Recombinant adeno-virus-mediated transfer of the Pdx1 gene to the liver ofmice induced expression of the insulin 1 and 2 genes andprohormone convertase 1/3, the enzyme that convertsproinsulin to insulin [22]. Plasma insulin levels increasedthreefold compared with those of control mice and insulinreduced hyperglycemia in a diabetic mouse treated withstreptozotocin [22]. Ectopic expression of PDX-1 mightconstitute a valuable approach for producing autologousinsulin-producing cells for treatment of type-1 diabetes.
Another appealing development of a PDX-1 expression-based approach for autologous cell replacement therapywas reported earlier this year [23]. Expression in theliver of Xenopus laevis tadpoles of a constitutively activeform of PDX-1 induces conversion of part of the liver topancreas, with both endocrine and exocrine cells(Fig. 2e). Similar effects were seen in culturedhuman hepatoma cells [23]. Loss of liver functionswas detected, indicative of transdifferentiation. Butperhaps the most remarkable observation in this studywas that the ectopic pancreas persisted even after thePdx1 transgene was turned off. It seems therefore thatonce the orchestra of pancreatic genes has been assembled
recombinase in mice creates a severe combined immune deficiency
caused by the complete absence of mature B and T lymphocytes. Cells
from immune-deficient Rag22/2 mice were cloned by nuclear trans-
plantation and an ES cell line was derived. Homologous recombination
allowed repair of one of the mutant alleles in the Rag22/2 ES cells
to restore normal Rag2 structure and function. Immune-competent
mice were produced from the repaired ES cells by tetraploid
complementation1; these mice were successfully used as donors of
bone marrow donors for grafting. Haematopoietic precursors were also
derived from the repaired ES cells and injected into Rag22/2 mice to
replenish lymphoid and myeloid tissues. This study provides an elegant
paradigm for the treatment of a genetic disorder by therapeutic cloning
in combination with gene therapy.
Therapeutic cloning has the advantage that a somatic cell can in theory
bereprogrammedintoanycell typethroughtheproductionof (cloned)ES
cells. These ES cells can be suitable for gene therapy [37]. However, low
efficiency of nuclear transfer is a concern: to date, hundreds of attempts
are typically required to produce a clinically suited ES cell line. This raises
the issue of human oocyte supply for therapeutic cloning. This limitation
might be alleviated with oocytes from other species [39] but mitochon-
drial genome differences between species are likely to pose a problem.
DifferentiatinganEScell line intoasufficientnumberofspecializedcells is
also a challenge. But perhaps the biggest current challenge of
therapeutic cloning is to overcome abnormalities encountered in
cloned animals [40] as these may reflect defects in cloned ES cells. In
spite of the production of cloned animals and ES-like cells by nuclear
transplantation, reports of unstable or abnormal gene expression
patterns in cloned embryos and fetuses [41,42] suggest incomplete
reprogramming. Nevertheless, proponents of therapeutic cloning may
argue that this inconvenience could be minimized because therapeutic
cloning does not require recapitulation of the entire development.
Another issue faced by therapeutic cloning is legal and ethical concerns
resulting from the production and manipulation of human embryos.
1 A technique whereby ES cells are injected into a tetraploid
blastocyst. The blastocyst strictly provides the placenta (tetraploid
mammalian embryos do not develop to term) while the ES cells develop
into an embryo and viable offspring.
Review TRENDS in Biotechnology Vol.21 No.8 August 2003358
http://tibtec.trends.com
by the transgene, transdifferentiation is irreversible.Because the liver is largely redundant, redirecting partof this organ to pancreas might present therapeuticbenefits. These findings [23] suggest that one mightsome day be able to induce transient expression of aspecific protein to elicit permanent phenotypic changes.An implication of this strategy is that knowledge of theregulatory factors required to achieve the desired cell typeis necessary.
Epigenetic reprogramming by cell hybridization
Epigenetic changes are modifications of DNA or chromatinthat do not involve DNA sequence alteration or deletion[24,25]. They allow differentiated cells to perpetuate themolecular memory needed to retain their identity [24,26].These modifications are heritable but they can beexperimentally reversed. A differentiated nucleus can bereprogrammed to a pluripotent state by fusion of a somaticcell with a stem or germ cell. Surani and colleagues haveshown that when mouse thymic lymphocytes are fusedwith embryonic germ (EG) cells, demethylation of severalimprinted and non-imprinted genes in the thymocytenucleus takes place [27]. The epigenetic modificationscorrelate with expression of a silent maternal allele of thePeg1/Mest imprinted gene in the thymocyte nucleus.Furthermore, the EG-thymocyte heterokaryons displaypluripotency because they contribute to various tissues inchimeric embryos [27]. Reversion of lymphocyte nuclei alsooccurs after fusion with ES cells, as shown by reactivationof the inactive X chromosome from female thymocytes,induction of a normally repressed Oct4-GFP transgene inthe thymocyte nuclei and contribution of the ES cell-thymocyte hybrids to the three primary germ layers inchimeras [28]. Nevertheless, the methylation pattern ofthe imprinted Igf2r and H19 genes is maintained [28]. EScells fused with mouse brain adult stem cells alsoreactivate a silent ES cell-specific gene in the neurongenome and confer pluripotentiality of the hybrids [29].These studies clearly demonstrate that epigenetic modi-fications can be induced in a differentiated somatic cellgenome and that such alterations are heritable, at least aslong as the ES or EG cell reprogramming components aremaintained in the hybrid cell. It will obviously beimportant to characterize the molecules eliciting epige-netic reprogramming and to determine to what extentepigenetic modifications occur in the transdifferentiationsituations illustrated earlier.
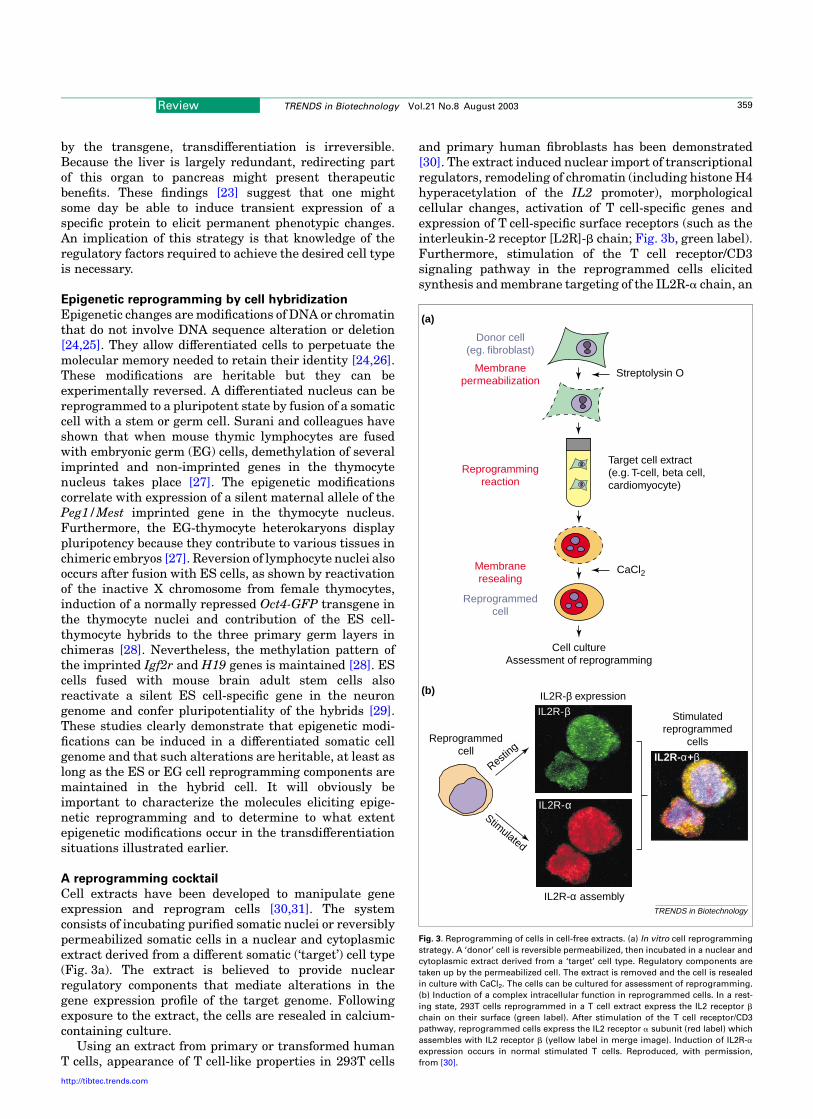
A reprogramming cocktail
Cell extracts have been developed to manipulate geneexpression and reprogram cells [30,31]. The systemconsists of incubating purified somatic nuclei or reversiblypermeabilized somatic cells in a nuclear and cytoplasmicextract derived from a different somatic (‘target’) cell type(Fig. 3a). The extract is believed to provide nuclearregulatory components that mediate alterations in thegene expression profile of the target genome. Followingexposure to the extract, the cells are resealed in calcium-containing culture.
Using an extract from primary or transformed humanT cells, appearance of T cell-like properties in 293T cells
and primary human fibroblasts has been demonstrated[30]. The extract induced nuclear import of transcriptionalregulators, remodeling of chromatin (including histone H4hyperacetylation of the IL2 promoter), morphologicalcellular changes, activation of T cell-specific genes andexpression of T cell-specific surface receptors (such as theinterleukin-2 receptor [L2R]-b chain; Fig. 3b, green label).Furthermore, stimulation of the T cell receptor/CD3signaling pathway in the reprogrammed cells elicitedsynthesis and membrane targeting of the IL2R-a chain, an
Fig. 3. Reprogramming of cells in cell-free extracts. (a) In vitro cell reprogramming
strategy. A ‘donor’ cell is reversible permeabilized, then incubated in a nuclear and
cytoplasmic extract derived from a ‘target’ cell type. Regulatory components are
taken up by the permeabilized cell. The extract is removed and the cell is resealed
in culture with CaCl2. The cells can be cultured for assessment of reprogramming.
(b) Induction of a complex intracellular function in reprogrammed cells. In a rest-
ing state, 293T cells reprogrammed in a T cell extract express the IL2 receptor b
chain on their surface (green label). After stimulation of the T cell receptor/CD3
pathway, reprogrammed cells express the IL2 receptor a subunit (red label) which
assembles with IL2 receptor b (yellow label in merge image). Induction of IL2R-a
expression occurs in normal stimulated T cells. Reproduced, with permission,
from [30].
Target cell extract(e.g. T-cell, beta cell,cardiomyocyte)
Streptolysin O
CaCl2
Cell cultureAssessment of reprogramming
IL2R-α
IL2R-β
IL2R- +α βResti
ng
Stimulated
Stimulatedreprogrammed
cells
IL2R-β expression
IL2R-α assembly
Reprogrammedcell
(a)
(b)
Membranepermeabilization
Donor cell(eg. fibroblast)
Reprogrammedcell
Membraneresealing
Reprogrammingreaction
TRENDS in Biotechnology
Review TRENDS in Biotechnology Vol.21 No.8 August 2003 359
http://tibtec.trends.com

indication that a complex T cell-specific regulatory func-tion took place (Fig. 3b). This suggests that it might bepossible to elicit other complex (e.g. secretory) functions infibroblasts reprogrammed in extract from other cell types.This functional aspect will be of crucial importance forpotential therapeutic applications of this technology.
It is clearly too premature to evaluate whether in vitrocell reprogramming might be useful in therapeuticcontext. Nevertheless, the use of cell extracts mightprove useful to investigate the molecular processes ofnuclear reprogramming, at least as they take place in vitro.Studies with cell extracts are also anticipated to extendcell fusion studies of nuclear reprogramming [27,28]because conditions of exposure of nuclei or cells toreprogramming factors can easily be manipulated.
The future of cell-based therapy
The immune system poses a barrier to allografts, whichmakes autologous cell-based therapies attractive alterna-tives. Isogenic transplantation eliminates problems ofimmune rejection but raises the economic and regulatorychallenges of producing a custom therapy for each patient.These issues are equally relevant for designing cell-baseddrug delivery systems.
In autologous cell therapy the patient would supplyher/his own donor cells for transplantation. These cellsmight be stem cells that can be easily accessed – such ashaematopoietic stem cells, stem cells of adipose tissueor skin precursor cells – and potentially multiplied(self-renewed) to obtain clinically relevant quantities.Stem cells can also be modified to repair genetic defectsbefore engraftment (see Box 1). Alternatively, the patient’sdonor cells might be healthy differentiated cells, such asskin fibroblasts, which can be readily obtained in largeamounts. One might envision that these cells can bedirectly turned into a cell type of choice ex vivo before goingback into the patient. Nevertheless, additional data arerequired to assess whether transdifferentiation of maturecells can be applied into a cell replacement therapy.Another emerging possibility is that organ conversion canbe achieved in situ using transient exogenous geneexpression [23]. This would be an advantage because itwould eliminate the need for biopsy and in vitro handlingof cells (associated with risks of contamination). Finally, acombination of both transgenesis with direct cell repro-gramming might provide optimum conditions to drive onecell type towards another.
Many fundamental issues need to be addressed,however, before a therapeutic application of transdiffer-entiation can be envisaged. For example, which donor celltype is most amenable for reprogramming; that is, which isthe most readily accessible and exhibits a chromatinorganization suitable for epigenetic modification? Howcomplete is the conversion of epigenetic regulation in thereprogrammed cells? Which role has the tissue environ-ment (including extracellular matrix, cell-to-cell contact)in reprogramming gene expression (see [32])? What is thestability of function of the reprogrammed cells? Why can asubset of cells transdifferentiate within an organ whereasthe majority of the tissue is not capable of transformation(see, e.g. the regional conversion of hepatocytes into
pancreatic cells [23])? Will the reprogrammed cells willbe of any therapeutic value in animal models?
One might also speculate on whether for therapeuticapplication, full transdifferentiation is necessary orwhether only partial reprogramming suffices. This coulddepend on the condition to be treated. For instance, fortreating type 1 diabetes, it might be sufficient to engineer acell that secretes regulated levels of insulin in response toglucose stimulation, very much like a pancreatic b cell;however, the replacement cell might not need to be a b cellstricto sensu. This hypothesis, however, is speculative andwill require extensive investigation. Nevertheless, wetentatively propose that a partially reprogrammed cellthat displays the necessary therapeutic properties, or‘therapeutic cell’, may be what it takes to treat certaintraumas or diseases. More work will tell.
AcknowledgementsThe authors are grateful to Dr. Maureen O’Leary for research oncompanies, Dr. Thomas Kuntziger for critical reading of the manuscriptand referees for comments. Research by the authors is supported by theResearch Council of Norway, the Norwegian Cancer Society, the HumanFrontiers Science Programme, the Juvenile Diabetes Foundation andNucleotech, LLC.
References
1 Tosh, D. and Slack, J.M. (2002) How cells change their phenotype. Nat.Rev. Mol. Cell Biol. 3, 187–194
2 Verfaillie, C.M. (2002) Adult stem cells: assessing the case forpluripotency. Trends Cell Biol. 12, 502–508
3 Wagers, A.J. et al. (2002) Little evidence for developmental plasticity ofadult hematopoietic stem cells. Science 297, 2256–2259
4 Terada, N. et al. (2002) Bone marrow cells adopt the phenotype of othercells by spontaneous cell fusion. Nature 416, 542–545
5 Ying, Q.L. et al. (2002) Changing potency by spontaneous fusion.Nature 416, 545–548
6 Slack, J.M. and Tosh, D. (2001) Transdifferentiation and metaplasia -switching cell types. Curr. Opin. Genet. Dev. 11, 581–586
7 DeRuiter, M.C. et al. (1997) Embryonic endothelial cells transdiffer-entiate into mesenchymal cells expressing smooth muscle actins invivo and in vitro. Circ. Res. 80, 444–451
8 Mochii, M. et al. (1998) Role of Mitf in differentiation and transdiffer-entiation of chicken pigmented epithelial cell. Dev. Biol. 193, 47–62
9 Wei, G. et al. (2000) Stem cell plasticity in mammals and transde-termination in Drosophila: common themes? Stem Cells 18, 409–414
10 Hu, E. et al. (1995) Transdifferentiation of myoblasts by the adipogenictranscription factors PPAR gamma and C/EBP alpha. Proc. Natl. Acad.Sci. U. S. A. 92, 9856–9860
11 Ross, S.E. et al. (2000) Inhibition of adipogenesis by Wnt signaling.Science 289, 950–953
12 Owen, M. (1988) Marrow stromal stem cells. J. Cell Sci. Suppl. 10,63–76
13 Nuttall, M.E. et al. (1998) Human trabecular bone cells are able toexpress both osteoblastic and adipocytic phenotype: implications forosteopenic disorders. J. Bone Miner. Res. 13, 371–382
14 Park, S.R. et al. (1999) Interconversion potential of cloned humanmarrow adipocytes in vitro. Bone 24, 549–554
15 Schiller, P.C. et al. (2001) Inhibition of gap-junctional communicationinduces the trans-differentiation of osteoblasts to an adipocyticphenotype in vitro. J. Biol. Chem. 276, 14133–14138
16 Condorelli, G. et al. (2001) Cardiomyocytes induce endothelial cells totrans-differentiate into cardiac muscle: implications for myocardiumregeneration. Proc. Natl. Acad. Sci. U. S. A. 98, 10733–10738
17 Shen, C.N. et al. (2003) Transdifferentiation of pancreas to liver. Mech.Dev. 120, 107–116
18 Rao, M.S. et al. (1988) Almost total conversion of pancreas to liver inthe adult rat: a reliable model to study transdifferentiation. Biochem.Biophys. Res. Commun. 156, 131–136
19 Dabeva, M.D. et al. (1997) Differentiation of pancreatic epithelial
Review TRENDS in Biotechnology Vol.21 No.8 August 2003360
http://tibtec.trends.com

progenitor cells into hepatocytes following transplantation into ratliver. Proc. Natl. Acad. Sci. U. S. A. 94, 7356–7361
20 Krakowski, M.L. et al. (1999) Pancreatic expression of keratinocytegrowth factor leads to differentiation of islet hepatocytes andproliferation of duct cells. Am. J. Pathol. 154, 683–691
21 Shen, C.N. et al. (2000) Molecular basis of transdifferentiation ofpancreas to liver. Nat. Cell Biol. 2, 879–887
22 Ferber, S. et al. (2000) Pancreatic and duodenal homeobox gene 1induces expression of insulin genes in liver and amelioratesstreptozotocin-induced hyperglycemia. Nat. Med. 6, 568–572
23 Horb, M.E. et al. (2003) Experimental conversion of liver to pancreas.Curr. Biol. 13, 105–115
24 Jones, P.A. and Takai, D. (2001) The role of DNA methylation inmammalian epigenetics. Science 293, 1068–1070
25 Jenuwein, T. and Allis, C.D. (2001) Translating the histone code.Science 293, 1074–1080
26 Turner, B.M. (2002) Cellular memory and the histone code. Cell 111,285–291
27 Tada, M. et al. (1997) Embryonic germ cells induce epigeneticreprogramming of somatic nucleus in hybrid cells. EMBO J. 16,6510–6520
28 Tada, M. et al. (2001) Nuclear reprogramming of somatic cells by invitro hybridization with ES cells. Curr. Biol. 11, 1553–1558
29 Pells, S. et al. (2002) Multipotentiality of neuronal cells afterspontaneous fusion with embryonic stem cells and nuclear reprogram-ming in vitro. Cloning Stem Cells 4, 331–338
30 Hakelien, A.M. et al. (2002) Reprogramming fibroblasts to expressT-cell functions using cell extracts. Nat. Biotechnol. 20, 460–466
31 Landsverk, H.B. et al. (2002) Reprogrammed gene expression in asomatic cell-free extract. EMBO Rep. 3, 384–389
32 Jaenisch, R. and Bird, A. (2003) Epigenetic regulation of geneexpression: how the genome integrates intrinsic and environmentalsignals. Nat. Genet. 33 (Suppl.), 245–254
33 Cibelli, J.B. et al. (1998) Transgenic bovine chimeric offspringproduced from somatic cell-derived stem-like cells. Nat. Biotechnol.16, 642–646
34 Munsie, M.J. et al. (2000) Isolation of pluripotent embryonic stem cellsfrom reprogrammed adult mouse somatic cell nuclei. Curr. Biol. 10,989–992
35 Wakayama, T. et al. (2001) Differentiation of embryonic stem cell linesgenerated from adult somatic cells by nuclear transfer. Science 292,740–743
36 Lanza, R.P. et al. (2002) Generation of histocompatible tissues usingnuclear transplantation. Nat. Biotechnol. 20, 689–696
37 Rideout, W.M. III et al. (2002) Correction of a genetic defect by nucleartransplantation and combined cell and gene therapy. Cell 109, 17–27
38 Cibelli, J.B. et al. (2002) Parthenogenetic stem cells in nonhumanprimates. Science 295, 819
39 Lanza, R.P. et al. (1999) Prospects for the use of nuclear transfer inhuman transplantation. Nat. Biotechnol. 17, 1171–1174
40 Cibelli, J.B. et al. (2002) The health profile of cloned animals. Nat.Biotechnol. 20, 13–14
41 Humpherys, D. et al. (2002) Abnormal gene expression in cloned micederived from embryonic stem cell and cumulus cell nuclei. Proc. Natl.Acad. Sci. U. S. A. 99, 12889–12894
42 Bortvin, A. et al. (2003) Incomplete reactivation of Oct4-related genesin mouse embryos cloned from somatic nuclei. Development 130,1673–1680
43 Clarke, D.L. et al. (2000) Generalized potential of adult neural stemcells. Science 288, 1660–1663
44 Bjornson, C.R. et al. (1999) Turning brain into blood: a hematopoieticfate adopted by adult neural stem cells in vivo. Science 283, 534–537
45 Kondo, T. and Raff, M. (2000) Oligodendrocyte precursor cells
reprogrammed to become multipotential CNS stem cells. Science289, 1754–1757
46 Brazelton, T.R. et al. (2000) From marrow to brain: expression ofneuronal phenotypes in adult mice. Science 290, 1775–1779
47 Mezey, E. et al. (2000) Turning blood into brain: cells bearing neuronalantigens generated in vivo from bone marrow. Science 290, 1779–1782
48 Zhao, L.R. et al. (2002) Human bone marrow stem cells exhibit neuralphenotypes and ameliorate neurological deficits after grafting into theischemic brain of rats. Exp. Neurol. 174, 11–20
49 Ferrari, G. et al. (1998) Muscle regeneration by bone marrow-derivedmyogenic progenitors. Science 279, 1528–1530
50 Petersen, B.E. et al. (1999) Bone marrow as a potential source ofhepatic oval cells. Science 284, 1168–1170
51 Krause, D.S. et al. (2001) Multi-organ, multi-lineage engraftment by asingle bone marrow-derived stem cell. Cell 105, 369–377
52 Lagasse, E. et al. (2000) Purified hematopoietic stem cells candifferentiate into hepatocytes in vivo. Nat. Med. 6, 1229–1234
53 Gussoni, E. et al. (1999) Dystrophin expression in the mdx mouserestored by stem cell transplantation. Nature 401, 390–394
54 Bittner, R.E. et al. (1999) Recruitment of bone-marrow-derived cells byskeletal and cardiac muscle in adult dystrophic mdx mice. Anat.Embryol. 199, 391–396
55 Jackson, K.A. et al. (2001) Regeneration of ischemic cardiac muscleand vascular endothelium by adult stem cells. J. Clin. Invest. 107,1395–1402
56 Orlic, D. et al. (2001) Bone marrow cells regenerate infarctedmyocardium. Nature 410, 701–705
57 Jiang, Y. et al. (2002) Pluripotency of mesenchymal stem cells derivedfrom adult marrow. Nature 418, 41–49
58 Reyes, M. et al. (2001) Purification and ex vivo expansion of postnatalhuman marrow mesodermal progenitor cells. Blood 98, 2615–2625
59 Reyes, M. et al. (2002) Origin of endothelial progenitors in humanpostnatal bone marrow. J. Clin. Invest. 109, 337–346
60 Shi, Q. et al. (1998) Evidence for circulating bone marrow-derivedendothelial cells. Blood 92, 362–367
61 Alison, M.R. et al. (2000) Hepatocytes from non-hepatic adult stemcells. Nature 406, 257
62 Theise, N.D. et al. (2000) Liver from bone marrow in humans.Hepatology 32, 11–16
63 Theise, N.D. et al. (2000) Derivation of hepatocytes from bone marrowcells in mice after radiation-induced myeloablation. Hepatology 31,235–240
64 Yang, L. et al. (2002) In vitro trans-differentiation of adult hepatic stemcells into pancreatic endocrine hormone-producing cells. Proc. Natl.Acad. Sci. U. S. A. 99, 8078–8083
65 Suzuki, A. et al. (2002) Clonal identification and characterization ofself-renewing pluripotent stem cells in the developing liver. J. CellBiol. 156, 173–184
66 Toma, J.G. et al. (2001) Isolation of multipotent adult stem cells fromthe dermis of mammalian skin. Nat. Cell Biol. 3, 778–784
67 Lako, M. et al. (2002) Hair follicle dermal cells repopulate the mousehaematopoietic system. J. Cell Sci. 115, 3967–3974
68 Zuk, P.A. et al. (2001) Multilineage cells from human adipose tissue:implications for cell-based therapies. Tissue Eng. 7, 211–228
69 Zuk, P.A. et al. (2002) Human adipose tissue is a source of multipotentstem cells. Mol. Biol. Cell 13, 4279–4295
70 Mizuno, H. et al. (2002) Myogenic differentiation by human processedlipoaspirate cells. Plast. Reconstr. Surg. 109, 199–209
71 Safford, K.M. et al. (2002) Neurogenic differentiation of murine andhuman adipose-derived stromal cells. Biochem. Biophys. Res. Com-mun. 294, 371–379
72 Ashjian, P.H. et al. (2003) In vitro differentiation of human processedlipoaspirate cells into early neural progenitors. Plast. Reconstr. Surg.111, 1922–1931
Review TRENDS in Biotechnology Vol.21 No.8 August 2003 361
http://tibtec.trends.com





























